

Improving the nutrition of Merino ewes during pregnancy increases the fleece weight and reduces the fibre diameter of their progeny’s wool during their lifetime and these effects can be predicted from the ewe’s liveweight profile
A. N. Thompson A C D G , M. B. Ferguson A C D , D. J. Gordon A E , G. A. Kearney A F , C. M. Oldham B and B. L. Paganoni BA Department of Primary Industries Victoria, Private Bag 105, Hamilton, Vic. 3300, Australia.
B Department of Agriculture and Food Western Australia, 3 Baron-Hay Court, South Perth, WA 6151, Australia.
C Present address: Department of Agriculture and Food Western Australia, 3 Baron-Hay Court, South Perth, WA 6151, Australia.
D Present address: School of Veterinary and Biomedical Sciences, Murdoch University, 90 South Street, Murdoch, WA 6150, Australia.
E Present address: Rural Industries Skills Training, Private Bag 105, Hamilton, Vic. 3300, Australia.
F Present address: 36 Payne Street, Hamilton, Vic. 3300, Australia.
G Corresponding author. Email: andrew.thompson@agric.wa.gov.au
Animal Production Science 51(9) 794-804 https://doi.org/10.1071/AN10161
Submitted: 27 August 2010 Accepted: 4 August 2011 Published: 14 September 2011
Journal Compilation © CSIRO Publishing 2011 Open Access CC BY-NC-ND
Abstract
Nutrition of ewes during pregnancy can have permanent impacts on the production potential of their progeny. The hypothesis tested in the experiments reported in this paper was that improving the nutrition of Merino ewes during pregnancy and lactation increases the fleece weight and reduces the fibre diameter of their progeny’s wool during their lifetime. In addition, that these effects on the progeny’s wool production can be predicted from the ewe’s liveweight profile. At sites in Victoria and Western Australia in each of 2 years, a wide range in the liveweight and condition score profiles of Merino ewes was generated by varying the amount of supplements fed from joining to Day 100 of pregnancy and the amount of feed on offer grazed from Day 100 to weaning. The site in Victoria was based on perennial pastures and included both single- and twin-bearing ewes whereas the site in Western Australia was based on annual pastures and included single-bearing ewes only. The production and characteristics of wool from the progeny were measured until 51 months of age at the site in Victoria and 33 months of age at the site in Western Australia. The nutritional treatments and the resulting changes in ewe liveweight had significant impacts on the fleece weight and to a lesser extent the fibre diameter of wool produced by their progeny, but there were no consistent effects on other characteristics of progeny fleece wool. The fleece weight of the progeny was related to the liveweight change during pregnancy of their mothers (P < 0.05) and the relationships were similar for the two experiments at each site. At the site in Victoria, a loss of 10 kg in ewe liveweight between joining and Day 100 of pregnancy reduced fleece weight by ~0.2 kg at each shearing until 51 months of age whereas gaining 10 kg from Day 100 of pregnancy to lambing had the opposite effect. The effect of changes in ewe liveweight during late pregnancy on the fleece weight of their progeny at each shearing was of similar magnitude at the site in Western Australia. When evident, the effect of the ewe liveweight profile on the fibre diameter of progeny wool was opposite to the effect on clean fleece weight and the effect of poor nutrition in early to mid pregnancy could be completely overcome by improving nutrition during late pregnancy. Twin-born and reared progeny produced ~0.3 kg less clean wool at each shearing (P < 0.001) that was 0.3-μm broader (P < 0.001) than that from single-born progeny at the site in Victoria. However, the effects of varying ewe nutrition and ewe liveweight change during pregnancy on fleece weight and fibre diameter of progeny wool were similar (P > 0.05) for both single- and twin-born or reared progeny. Overall, these results supported our hypothesis and it is clear that the nutritional management of Merino ewes during pregnancy is important for optimal wool production from their progeny during their lifetime.
Introduction
The weight and diameter of wool produced by sheep is influenced by the ratio of secondary to primary wool follicles and the number of secondary follicles is genetically controlled (Jackson et al. 1975; Hocking Edwards et al. 1994). Secondary wool follicles are initiated between Day 80 of fetal life and just before birth and most secondary follicles commence producing a fibre 1–3 weeks after birth (Short 1955a; Hocking Edwards 1999). The development of the follicle population in the fetus is influenced by the nutrition of the ewe (Short 1955b; Schinckel and Short 1961; Everitt 1967). Progeny born to underfed ewes may grow less wool that is broader than progeny born to ewes that were better fed during pregnancy (Denney 1990; Kelly et al. 1996, 2006). Kelly et al. (1996) showed that in comparison to progeny from single-bearing ewes fed to maintain maternal liveweight during pregnancy, genetically identical progeny from ewes fed to lose 10 kg between Days 50 and 140 of pregnancy produced ~0.14 kg less clean wool that was also ~0.1-μm broader at their hogget shearing. These differences in clean fleece weight and fibre diameter between single-born progeny were permanent (Kelly et al. 2006), and this is consistent with the differences in wool production and quality that persist between single- and twin-born lambs until at least 2–3 years of age (Huisman et al. 2008).
The effects of maternal nutrition on fetal and wool follicle development, and therefore the wool production of the progeny as adults, will be influenced by the ‘timing’, duration and severity of the nutrient restrictions experienced by the ewe (Robinson et al. 1999). Most studies of the impacts of nutrition on fetal growth and development have tended to focus on late pregnancy or have only considered extreme nutritional regimes often outside the boundaries of commercial reality. The outstanding question is whether the variations of nutrient supply experienced during different stages of pregnancy and lactation under more typical grazing conditions are sufficient to induce permanent changes in the wool production of adults. The extent that nutrition can be restricted during different stages of the reproductive cycle before adverse impacts on the wool production potential of the progeny are observed, or if any adverse effects can be overcome by subsequent nutritional management, are not known. There are also no reports in the literature that have quantified the effects of different levels of pasture or different rates of ewe liveweight change during pregnancy and lactation on the production and quality of wool produced by single and twin progeny over their lifetime. The hypothesis tested in the experiments reported in this paper is that improving the nutrition of Merino ewes during pregnancy and lactation increases the fleece weight and reduces the fibre diameter of their progeny’s wool during their lifetime, and that these effects of nutrition can be reliably predicted from the liveweight profile of the ewe over pregnancy and lactation.
Materials and methods
All procedures reported in this paper were conducted according to the guidelines of the Australian Code of Practice for the Use of Animals for Scientific Purposes and received approval from the various State Department Animal Ethics Committees.
Experimental sites and design
Experiments were conducted in 2001 and 2002 at two sites located on commercial properties near Hamilton in south-west Victoria (Vic.; 141.7°E/41′25″, –37.6°S/36′1″) and Kendenup in Western Australia (WA; 117.6°E/37′25″, –34.5°S/29′13″). Both sites experience predominantly winter–spring rainfall, and dry, hot summers, with a long-term average annual rainfall of 590 and 540 mm for the Vic. and WA sites. Actual rainfall received in 2001 and 2002 was 717 and 548 mm at the Vic. site and 522 and 466 mm at the WA site. The pastures at the Vic. and WA sites were based on perennial grasses (Lolium perenne and Phalaris aquatica) and annual grasses (Lolium rigidum) respectively, with a much higher proportion of subterranean clover in the pastures at the WA than Vic. site over the 2 years (46 versus 11%). Other papers in this series provide additional details on the sites and experimental design, pasture and ewe management and measurements (Ferguson et al. 2011) and progeny management and measurements (Oldham et al. 2011; Thompson et al. 2011).
In brief, a factorial design was used with two (WA) or three (Vic.) replicates of the following 10 treatments: (i) two target ewe condition scores (2.0 and 3.0) at Day 100 of pregnancy, after being joined in condition score 2.5–3.0; and (ii) five target amounts of feed on offer (800, 1100, 1400, 2000, and >3000 kg green DM/ha) from Day 100 of pregnancy until lambs were weaned (Vic.) or when pasture growth could no longer maintain the feed on offer targets (WA). The lambs at the WA site were weaned ~30 days after removal from plots. The target feed on offer levels for each treatment group were maintained by varying the number of non-experimental sheep on each plot and or the area grazed by experimental sheep.
Ewe management and measurements
Adult Merino ewes at the Vic. site were 2.5 or 3.5 years old (n = 1600) and at the WA site between 2.5 and 5.5 years old (n = 1400) in each experiment, and different ewes were used in each experiment at both sites. The ewes were artificially inseminated to commence lambing in late August (Vic.) or late July (WA). Average dates of artificial insemination (Day 0) were 2 April and 29 March in 2001 and 2002 at the Vic. site, and 1 March in both years at the WA site. Semen was used from 20 sires each year selected from four fine-medium wool genotypes with at least 40% genetic linkage between sites and years.
After artificial insemination, ewes were randomly split into two flocks on the basis of liveweight, condition score and sire source. They were then managed to achieve a target condition score of 2 or 3 by Day 100 of pregnancy by altering the supplementary feeding regime and grazing pressure. Ewes were scanned at Day 60–70 of gestation to identify pregnancy status. At Day 100 of pregnancy, single- and twin-bearing ewes at the Vic. site or single-bearing ewes at the WA site from each condition score flock were allocated to pasture plots on the basis of liveweight, condition score and sire source. The plots were maintained at five target levels of feed on offer and treatments ceased at lamb weaning (Vic.) or 30 days prior to weaning (WA). At the Vic. site plots were grazed with 303 single- and 375 twin-bearing ewes in 2001 and 467 single- and 219 twin-bearing ewes in 2002. At the WA site 320 single-bearing ewes were used in each experiment.
Ewes were weighed and condition scored during pregnancy and lactation every 2–4 weeks at both sites. Ewe liveweight was corrected for cumulative fleece weight, estimated from five or six dye-bands spaced through the year (Wheeler et al. 1977), and for conceptus weight using the equations developed by Wheeler et al. (1971). Condition score was assessed using the method described by Jefferies (1961). The liveweight, condition score and characteristics of wool produced by the ewes are reported by Ferguson et al. (2011).
Progeny management and measurements
All lambs were weighed and double-tagged within 24 h of birth and their sex and dam recorded. Lambs were weighed 2–4 weekly until weaning at ~11 weeks of age at the Vic. site and ~16 weeks of age at the WA site. They were tailed, mulesed, and castrated (males) at lamb marking at 5–6 weeks and vaccinated at lamb marking and at weaning. After weaning, all progeny at each site were grazed together until they were at least 30 months of age. The ewe progeny at the Vic. site were not mated until 30 months of age (after their third shearing) whereas the WA progeny were mated at 19 months of age just before their second shearing. They were weighed every 1–2 months until 12 months of age and then up to three times per year until 63 or 33 months of age at the Vic. and WA sites, respectively. The birthweights, survival and liveweight of progeny to maturity are reported by Oldham et al. (2011) and Thompson et al. (2011).
Progeny were shorn at varying ages as shown in Table 1. A sample of wool (50 g) was taken from the mid-side region of all progeny before shearing and used to measure: (i) yield; (ii) mean and variation in fibre diameter; (iii) staple length; (iv) staple strength; and (v) position of break along the staple. The methods used for measuring wool characteristics have been described by Thompson et al. (1994). The total weight of greasy wool was recorded for individual animals at shearing.

|
Statistical analyses
All statistical analyses were performed using Genstat (Genstat Committee 2008). In the first analysis, the method of restricted maximum likelihood (REML) was used to fit progeny clean fleece weight, mean and variation in fibre diameter, staple length, staple strength and position of break with target ewe condition score at Day 100 of pregnancy, age at shearing (15–51 months) and average feed on offer per plot during late pregnancy and lactation as a linear or quadratic effect, and ewe age and progeny sex and rear type as fixed effects within year. Replicate, plot and sire were fitted as random effects. This method of analysis was also used for the first shearing at 5 months with the age at shearing fixed effect removed. Statistical significance was accepted at P < 0.05.
Data generated from the four experiments were then utilised to determine whether liveweight or change in liveweight of individual ewes or feed on offer during specific periods could predict progeny clean fleece weight, mean and variation in fibre diameter, staple length, staple strength and position of break. For prediction of the progeny wool characteristics REML was used with ewe liveweight at joining, ewe liveweight change from joining until Day 100 of pregnancy, ewe liveweight change from Day 100 of pregnancy until lambing, and ewe liveweight change from lambing until weaning (or when removed from plots at the WA site). Ewe age and rearing type and sex and pregnancy status of progeny were fitted as fixed effects where appropriate, and year, replicate, plot and sire were fitted as random effects. All possible models were examined to define statistical significance of effects and interactions accepted at P < 0.05.
Finally, lamb birthweight and growth rate to weaning from the four experiments were used to predict progeny clean fleece weight and fibre diameter. For this REML was used with lamb birthweight and growth rate to weaning fitted as variates, ewe age, rearing type and sex of progeny fitted as fixed effects where appropriate, and year, replicate, plot and sire fitted as random effects. All possible models were examined to define statistical significance of effects and interactions accepted at P < 0.05.
Results
Treatment effects on ewe liveweight and condition score profiles
The average (±s.e.m) liveweight and condition score of the ewes at or just before artificial insemination in 2001 and 2002 were 46 ± 0.2 kg and condition score 2.7 ± 0.01 and 45 ± 0.2 kg and condition score 3.0 ± 0.01 at the Vic. site, and 46 ± 0.3 kg and condition score 2.9 ± 0.02 and 47 ± 0.3 kg and condition score 2.5 ± 0.03 at the WA site. On average across all experiments, the nutritional treatments after artificial insemination generated differences in ewe liveweight and condition score by Day 100 of pregnancy of 7.1 kg (range 5.5–8.7 kg) and 0.7 of a condition score (range 0.6–0.8). Grazing different feed on offer levels from Day 100 of pregnancy amplified the spread in ewe liveweight and condition score between treatments, such that the average differences between extreme treatments at lambing were 9.7 kg (range 4.9–14.3 kg) and 1.1 of a condition score (range 0.5–1.6) and at weaning 15.5 kg (range 12.5–22.7 kg) and 1.4 of a condition score (range 0.9–2.3). The range in liveweight of progeny at weaning between the extreme treatments in 2001 and 2002 was 14.9–20.3 kg and 13.8–20.0 kg at the Vic. site and 15.4–28.3 kg and 22.4–27.7 kg at the WA site, respectively.
Treatment effects on progeny fleece weight and fibre diameter
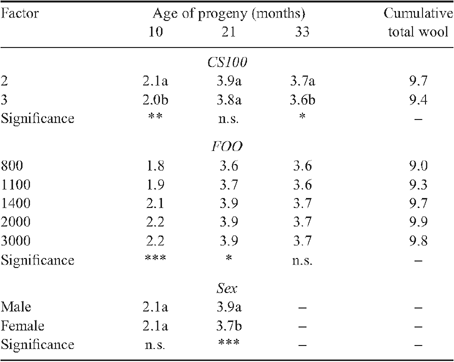
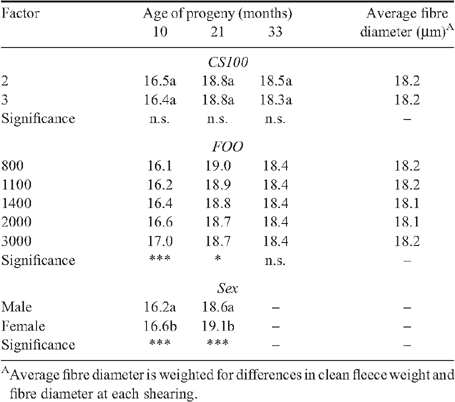
The effects of ewe condition score treatment at Day 100 of pregnancy and feed on offer grazed by ewes during late pregnancy and lactation on progeny clean fleece weight and fibre diameter are shown in Tables 2–5. At the Vic. site, progeny from ewes managed to target condition score 2 at Day 100 of pregnancy tended to produce ~0.1 kg less wool at each shearing, with the exception of the lamb shearing at 5 months of age, and their wool was broader than that of progeny from ewes managed to target condition score 3 at Day 100 of pregnancy. Over five shearings the progeny from ewes fed less during early to mid pregnancy produced ~0.4 kg less wool that on average was also 0.1-μm broader. There were no significant effects of ewe nutrition during early- and mid pregnancy on progeny fibre diameter at the WA site, but the treatment effects on progeny clean fleece weight appeared to be opposite to those observed at the Vic. site.

|

|
|
|
The effects of feed on offer treatments from Day 100 of pregnancy to weaning on progeny clean fleece weight and fibre diameter were greater than the effects of ewe nutrition to Day 100 of pregnancy at both sites. Progeny from ewes that grazed higher feed on offer levels from mid pregnancy until weaning produced more wool that was broader at their first shearing at 5 months of age at the Vic. site or 10 months of age at the WA site. At the hogget shearing at both sites treatment effects on progeny clean fleece weight and fibre diameter were significant (P < 0.05), and progeny from ewes that grazed higher levels of feed on offer from mid pregnancy to weaning produced more wool that was also significantly finer (P < 0.05).
There was a significant feed on offer by age at shearing interaction (P < 0.01), such that treatments effects on progeny clean fleece weight and fibre diameter decreased between hogget shearing and shearing at 51 months of age at the Vic. site or 33 months of age at the WA site. However, while treatment effects on progeny clean fleece weight and fibre diameter were not statistically significant beyond the hogget shearing, the trends were mostly in the same direction as that observed at their hogget shearing. The cumulative difference between the extreme feed on offer treatments was ~1.1 kg of clean wool and 0.2 μm over five shearings at the Vic. site and 0.8 kg of clean wool over three shearings at the WA site. There were no differences in cumulative fibre diameter of progeny wool at the WA site between extreme treatments largely because progeny from the higher feed on offer treatment produced broader wool at their first shearing at 10 months of age than progeny from the lower feed on offer treatments.
At the Vic. site there were consistent effects of birth and rear type of progeny on their clean fleece weight (Table 2). At all shearings between 5 and 51 months of age single-born progeny produced significantly (P < 0.001) more wool than twin-born and reared progeny, and the average fleece weight for twin-born single reared progeny was intermediate. Single-born progeny produced broader wool than twin-born progeny at their lamb shearing at 5 months of age but thereafter single-born progeny tended to produce finer wool than the twin-born progeny even though the difference was not significant at all shearings (Table 3). The effects of rear type on the fibre diameter of wool produced by twin-born progeny varied with age but was not significant at 27 months of age and older.
When progeny were grazed together until 27 months of age at the Vic. site and 21 months of age at the WA site, male progeny produced as much or more wool that was also significantly finer than wool from female progeny. At the Vic. site male progeny generally produced more and broader wool at the fourth and fifth shearing than ewe progeny, but sex was confounded with management group.
There was no significant effects of ewe age on progeny fleece weight or fibre diameter at either site, and no significant interactions between target condition score of ewes at Day 100 of pregnancy, target feed on offer during late pregnancy and lactation, ewe age, progeny sex and or progeny birth and rear type. There were also no consistent effects of these factors on yield, variation in fibre diameter, staple length, staple strength or position of break of progeny wool at different ages of shearing at either site.
Prediction of progeny fleece weight from ewe liveweight profile
Progeny clean fleece weight at each shearing from hogget age through to 51 months of age at the Vic. site or 33 months of age at the WA site could be predicted from the liveweight profile of the ewe (Tables 6 and 7). There were no significant (P > 0.05) interactions with year of experiment so data from the different experiments at each site was combined. Most importantly the coefficients in the models which predicted progeny clean fleece weight from ewe liveweight profile did not differ significantly with age of progeny at shearing.

|

|
At the Vic. site, ewes that were heavier at joining or lost less weight between joining and Day 100 of pregnancy or gained more weight from Day 100 of pregnancy to lambing produced progeny that had heavier fleece weights. These aspects of the ewe liveweight profile were significant when fitted together and each explained additional variance in progeny clean fleece weight. An additional 1 kg of ewe liveweight at joining increased the fleece weight from progeny by ~0.10 kg. A loss of 10 kg in liveweight of ewes between joining and Day 100 of pregnancy consistently reduced fleece weight by ~0.2 kg at all shearings whereas gaining 10 kg from Day 100 of pregnancy to lambing had the opposite effect (Table 6).
The fleece weight responses of the progeny to the liveweight profile of the ewe were consistent regardless of progeny birth type or rear type and sire genotype at the Vic. site as there were no interactions between these factors. However, the effect of progeny birth and rear type on their fleece weight remained significant after adjusting for the effects of birth and rear type on ewe liveweight profile. For the same ewe liveweight profile, on average single-born progeny produced 0.27 kg more wool than twin-born and reared progeny and 0.12 kg more wool than twin-born single reared progeny.
At the WA site, only liveweight change from Day 100 of pregnancy until lambing significantly influenced progeny clean fleece weight and the magnitude of the progeny wool response to ewe liveweight change during this period was very similar to that observed at the Vic. site (Table 7). Various combinations of liveweight change of the ewe over shorter periods (≥2 weeks WA, ≥4 weeks Vic.) failed to find other periods of change in the liveweight of individual ewes that was related to the clean fleece weight of their progeny. There was also no significant effect (P > 0.05) of changes in ewe liveweight during lactation on progeny fleece weight.
Prediction of progeny fibre diameter from ewe liveweight profile
Fibre diameter of wool from progeny at hogget age could be predicted from the liveweight profile of the ewe and the data from the different experiments at each site could be combined (Tables 8 and 9). At the Vic. site, ewes which lost less weight between joining and Day 100 of pregnancy or gained more weight from Day 100 of pregnancy to lambing reared progeny that produced finer wool. A loss of 10 kg in ewe liveweight between joining and Day 100 of pregnancy increased fibre diameter of progeny wool by ~0.3–0.4 μm whereas gaining 10 kg of liveweight from Day 100 of pregnancy to lambing had the opposite effect.

|

|
The progeny fibre diameter responses at hogget age to ewe liveweight profile were consistent regardless of progeny birth type, rear type and sire genotype. However, with the exception of the shearing at 51 months of age, the effects of progeny birth and rear type on their fibre diameter remained significant after adjusting for the effects of birth and rear type on ewe liveweight profile. For the same ewe liveweight profile, on average single-born progeny produced wool that was 0.48-μm finer than that from twin-born and reared progeny and 0.13-μm finer wool than twin-born single reared progeny at 15 months of age (Table 8). The effects of ewe liveweight profile and birth or rear type on fibre diameter of progeny wool were not significant beyond shearing at 15 months and 27 months of age, respectively, even though the birth type and rear type effects were significant when combines across all shearings.
At the WA site, ewe liveweight change from both joining to Day 100 of pregnancy and Day 100 of pregnancy until lambing, influenced the fibre diameter of wool produced by the progeny. While the effects were not quite statistically significant at 33 months of age (P < 0.1), the responses to ewe liveweight change during this period were similar to or greater than that observed at the Vic. site (Table 9). The fibre diameter responses were also similar to clean fleece weight in that at both sites there was no evidence that fibre diameter of wool from progeny was influenced by changes in ewe liveweight over periods of only a few weeks during pregnancy or during lactation.
Prediction of progeny wool production from their birthweight and growth to weaning
Low birthweight lambs produced less wool that was broader and these effects of birthweight persisted until 51 months at the Vic. site and 33 months at the WA site (Tables 10–13). The fleece weight and fibre diameter responses to birthweight were similar across experiments and sites, and on average lambs that were 1-kg lighter at birth produced ~0.11–0.13 kg less wool that was 0.18–0.24-μm broader over their lifetime.

|

|

|

|
Lambs that grew faster to weaning also produced significantly more wool at all shearings at both sites, and broader wool at all shearings at the Vic. site but not at the WA site. At the Vic. site, lambs that grew an extra 50 g/day from birth to weaning produced an extra 0.15 kg of clean wool across all shearings that was also 0.1-μm broader.
The difference in fleece weight between single- and twin-born or reared progeny was explained fully via the effect of birth type on birthweight and rear type on lamb growth rate to weaning. By contrast, significant effects of birth type and rear type on the fibre diameter of wool produced by progeny remained after adjustment for differences in their birthweight and growth to weaning.
Discussion
Improving the nutrition of Merino ewes during pregnancy and lactation increased the fleece weight and reduced the fibre diameter of wool produced by their progeny at their hogget shearing. This is consistent with the findings of Schinckel and Short (1961), Everitt (1967) and Kelly et al. (1996). The high repeatability and small errors about the coefficients across years and sites (Tables 6–9) confirms the goodness of fit and predictability of the relationships derived in this study. Further validation of these relationships is also provided by the finding that the equations derived from individual ewes and their progeny in the present study were similar to those derived for whole flocks from a series of paddock-scale experiments conducted on farms across southern Australia (Behrendt et al. 2011). In some cases the effects of the maternal environment on progeny clean fleece weight and fibre diameter decreased between hogget shearing and shearing at 51 months of age at the Vic. site or 33 months of age at the WA site, depending on whether the maternal environment was defined as the nutritional treatments, the liveweight profile of individual dams or the birth and rear type of the progeny themselves. Nevertheless, the cumulative difference between the extreme nutritional treatments that covered the range typical of those experienced by many Merino ewes during pregnancy and lactation on commercial farms across southern Australia (Kelly 1992; Kleemann and Walker 2005; Saul et al. 2011), was ~1.1 kg of clean wool and 0.2 μm over five shearings at the Vic. site and 0.8 kg of clean wool over three shearings at the WA site. These effects were comparable to the 1.2–1.4 kg and 0.1–0.3-μm differences in productivity of progeny measured over 4–6 shearings reported by Kelly et al. (2006). It is clear that development of optimum feeding strategies for breeding Merino ewes must take into account the impacts of ewe nutrition on the lifetime wool production of the progeny (Young et al. 2011).
The liveweight profile of ewes provided an effective tool for predicting the effects of maternal nutrition on progeny fleece weight. Restricting the level of nutrition to the ewe reduced progeny fleece weight and this effect was dependent on the timing and severity of the restriction and subsequent nutrition. At the Vic. site a loss of 10 kg in ewe liveweight between joining and Day 100 of pregnancy reduced fleece weight by ~0.2 kg at each shearing whereas a gain of 10 kg from Day 100 of pregnancy to lambing had the opposite effect. The responses in progeny clean fleece weight were consistent across both experiments at this site and the coefficients to predict clean fleece weight from ewe liveweight changes did not change significantly with increasing age of progeny. This result is the strongest evidence reported in the literature confirming that the effects of ewe liveweight change during pregnancy on progeny clean fleece weight were permanent and predictable.
The model coefficients derived to predict the clean fleece weight of progeny at the WA site from ewe liveweight change during late pregnancy were not significantly different to those for the Vic. site, and again they did not change significantly with increasing age of progeny. The impact of ewe liveweight change from joining to Day 100 of pregnancy on clean fleece weight showed a similar trend to the Vic. site, but was not significant. There was no obvious explanation for this result given ewe liveweight change to Day 100 of pregnancy was related to the fibre diameter of their progeny’s wool and birthweight (Oldham et al. 2011) and liveweight to 33 months of age (Thompson et al. 2011) at this site. While Kelly et al. (2006) did not attempt to quantify the relative effects of ewe liveweight change during different stages of pregnancy on progeny wool production, the prediction coefficients we derived from their work, where most of the differences in ewe liveweight were generated in mid and late pregnancy and the ewes lambed in autumn, were similar to the present study (average 0.26 kg fleece weight per 10-kg change in ewe liveweight). The results from the WA site, in conjunction with Kelly et al. (2006), further support our hypothesis that improving the nutrition of Merino ewes during pregnancy permanently increases the progeny’s fleece weight. Furthermore they suggest that the progeny wool production responses to ewe liveweight profile are likely to apply across environments and for different times of lambing.
There were consistent effects of the liveweight profile of ewes during pregnancy on fibre diameter of the wool of progeny as hoggets but in most cases the effects on fibre diameter declined as they got older (Tables 8 and 9). By contrast, Behrendt et al. (2011) reported that the effects of ewe liveweight profile during pregnancy on progeny fibre diameter were more apparent at the adult shearing than the hogget shearing for 13 different flocks across southern Australia. Kelly et al. (2006) also reported that the adverse impacts of poor ewe nutrition during pregnancy on the fibre diameter of wool produced by the progeny were permanent. Similarly, we found effects of birthweight of lambs, regardless of whether this was caused by poor ewe nutrition or increasing litter size, resulted in a permanent difference in their fibre diameter over their lifetime. Hence, when considered together with other literature our data supports the hypothesis that improving the nutrition of Merino ewes during pregnancy permanently decreases the progeny’s fibre diameter.
When progeny clean fleece weight and fibre diameter were related to changes in ewe liveweight between joining and Day 100 of pregnancy and Day 100 of pregnancy and lambing, the effects of ewe liveweight change during the different periods of pregnancy were additive. In other words, the effects of poor nutrition in early- and mid pregnancy on progeny wool production could be completely overcome by improving nutrition during late pregnancy. This is consistent with the effect of changes in ewe liveweight on lamb birthweight (Oldham et al. 2011), but is the first work to our knowledge to report significant effects of nutrition during early- and mid pregnancy on the wool production of single and twin progeny during adulthood. The overall effects of changes in nutrition during pregnancy on progeny fleece characteristics are consistent with permanent reductions in follicle density and the ratio of secondary to primary follicles (Short 1955b; Schinckel and Short 1961; Everitt 1967; Kelly et al. 2006). However, we expected that improving nutrition during late pregnancy, the period of maximum initiation and branching of secondary wool follicles, would have a greater impact of progeny wool production and quality than improving nutrition during early- and mid pregnancy, but this proposition was not supported. Hutchison and Mellor (1983) suggested that the ratio of secondary to primary follicles was most sensitive to underfeeding during the period from Days 112 to 132 of gestation, the period just before completion of secondary follicle initiation (Schinckel 1955). However, there was again no evidence in our work that progeny wool production could be significantly manipulated by managing ewe liveweight during more acute critical ‘windows’ of only a few weeks during pregnancy. This suggests that consideration of ewe nutrition throughout the whole pregnancy is important in optimising progeny wool production.
Birth type and rear type of the progeny had variable effects on their clean fleece weight and fibre diameter. Twin-born animals produced significantly less wool (–0.19 kg) with higher fibre diameter (+0.26 μm) than single-born animals across all shearings at the Vic. site. On average, twin-born progeny reared as twins also produced less wool (–0.15) than those reared as a single lamb, but fibre diameter was less affected by rear type. Several studies (Brown et al. 2002, 2006; Safari et al. 2005) reported this birth-type effect on clean fleece weight and fibre diameter but small or non-significant effects of rear type on these traits. In our work the differences in progeny fleece weight between birth and rear types were consistent from 15 months to at least 51 months of age. By contrast, while birth-type effects on fibre diameter of wool produced by progeny generally persisting to maturity rear-type effects were only evident at hogget age. Few studies have reported the effects of birth type or rear type on clean fleece weight and fibre diameter beyond 2–3 years (Huisman et al. 2008).
When evident, the progeny fleece weight and fibre diameter responses to ewe liveweight profile were consistent regardless of progeny birth or rear type. In other words, the effects of ewe liveweight change and birth-rear type on progeny wool production were additive and there was no evidence that the effects of varying ewe nutrition on progeny wool production was greater for twin- than single-born progeny. This was expected given Oldham et al. (2011) also reported that there was no interaction between ewe liveweight change and birth type for lamb birthweight, and lamb birthweight is related to progeny wool production. For the same ewe liveweight profile, on average single-born progeny produced 0.27 kg more wool than twin-born and reared progeny and 0.12 kg more wool than twin-born single reared progeny.
In practical terms, a comparison of the coefficients that predict lamb birthweight (Oldham et al. 2011) or progeny fleece weight and fibre diameter from ewe liveweight profile versus the relative effects of birth type and rear type on these traits, suggests that it might not be possible to preferentially feed ewes that conceive and or rear twins such that their progeny will perform at similar levels to single-born and reared progeny. However, to the contrary, Hocking Edwards et al. (2011) showed that it was possible to manage twin-bearing ewes to ensure that their surviving progeny produced a similar quantity and quality of wool to progeny from single-bearing ewes fed lower levels of nutrition in late pregnancy. Scanning for litter size and differential management in late pregnancy such that twin-bearing ewes are ~3 kg or 0.3 of a condition score fatter at lambing result in changes in lamb survival and wool production of progeny that improve farm profits (Young et al. 2008). These benefits from differential management of twin-bearing ewes are over and above those achieved from better nutritional management of the whole flock reported by Young et al. (2011).
The effects of birth type on progeny clean fleece weight and fibre diameter were largely explained by the differences in the birthweight between single- and twin-born lambs since birth type explained no additional variance in these traits in addition to that explained by birthweight. Differences in birthweights reflect the combined effects of the maternal environment and the genotype of the lamb. At the Vic. site, single-born lambs were 1.1-kg heavier at birth than twin-born lambs (Oldham et al. 2011), and this resulted in a 0.15-kg increase in clean fleece weight and a 0.20-μm decrease in fibre diameter in single-born lambs compared with twin-born lambs. The clean fleece weight and fibre diameter responses to changes in birthweight were similar for the WA and Vic. sites regardless of the age of the progeny. This is further evidence that nutrition of the developing fetus has permanent effects of the wool production potential of the progeny. As low birthweight had negative impacts on both progeny wool production and lamb survival (Oldham et al. 2011), the effects of nutritional treatments on progeny wool production would be even more evident if strategies were adopted to improve the survivability of low birthweight lambs.
Progeny that grew faster to weaning produced heavier fleeces at each shearing and this effect was evident in both experiments and at both sites. On average, an extra 50 g/day gain in lamb growth to weaning was related to an increase in progeny fleece weight of ~0.15 kg at the Vic. site and 0.14 kg at the WA site. This is consistent with Schinckel (1955) and Hocking Edwards et al. (1994) who showed that many follicles that initiate do not get to the stage of producing a fibre, especially in animals that have a relatively low birthweight and low growth rates in the first few months. Lambs born and raised as twins grew ~50 g/day slower to weaning than those born as twins and raised as a single, and these differences in growth rates were not related to ewe liveweight change during lactation (Thompson et al. 2011). The effects of rear type on progeny clean fleece weight could be explained by their differences in growth to weaning. By contrast, rear type had minimal effects on progeny fibre diameter and there was no consistent effect of progeny growth rate to weaning. Taken together, these results suggest that the maternal environment during gestation and during early post-natal life both influence fleece weight of the progeny, but fibre diameter of progeny wool was mostly influenced by prenatal nutrition.
To optimise whole-farm stocking rate and manage risk in situations of fluctuating and unpredictable nutrient supply, it is inevitable that Merino ewes will be managed to achieve less than maximum rates of production for both herself and her progeny. However, as progeny fleece weight and fibre diameter are significantly affected by maternal nutrition, it is essential that these responses are included in the models used to estimate the cost effectiveness of different ewe management strategies.
Acknowledgements
This work was funded by Australian Wool Innovation Limited, Department of Primary Industries, Victoria and Department of Agriculture and Food Western Australia. The authors thank David and Fiona Robertson and Bill and Kay and Geoff Sandilands for providing the research sites and experimental sheep. Mr Brian Hurley, Mr Tom Plaisted, Ms Kazue Tanaka and Ms Amber Walker are thanked for their invaluable technical assistance.
References
Behrendt R, van Burgel AJ, Bailey A, Barber P, Curnow M, Gordon DJ, Hocking Edwards JE, Oldham CM, Thompson AN (2011) On-farm paddock-scale comparisons across southern Australia confirm that increasing the nutrition of Merino ewes improves their production and the lifetime performance of their progeny. Animal Production Science 51, 805–812.| On-farm paddock-scale comparisons across southern Australia confirm that increasing the nutrition of Merino ewes improves their production and the lifetime performance of their progeny.Crossref | GoogleScholarGoogle Scholar |
Brown DJ, Ball AJ, Mortimer R, Oppenheimer M (2002) Incorporating SRS(R)/Elite wool traits into genetic evaluations for Merino sheep 1. Phenotypic variation and heritabilities. Wool Technology and Sheep Breeding 50, 373–377.
Brown DJ, Mortimer RL, Mortimer ML (2006) Genetic aspects of greasy wool colour assessments in Merino sheep. Proceedings of the Association of Animal Breeding and Genetics 14, 119–122.
Denney GD (1990) Effects of preweaning farm environment on adult wool production of Merino sheep. Australian Journal of Experimental Agriculture 30, 17–25.
| Effects of preweaning farm environment on adult wool production of Merino sheep.Crossref | GoogleScholarGoogle Scholar |
Everitt GC (1967) Residual effects of prenatal nutrition on the postnatal performance of Merino sheep. Proceedings of the New Zealand Society of Animal Production 27, 52–68.
Ferguson MB, Thompson AN, Gordon DJ, Hyder MW, Kearney GA, Oldham CM, Paganoni BL (2011) The wool production and reproduction of Merino ewes can be predicted from changes in liveweight during pregnancy and lactation. Animal Production Science 51, 763–775.
| The wool production and reproduction of Merino ewes can be predicted from changes in liveweight during pregnancy and lactation.Crossref | GoogleScholarGoogle Scholar |
Genstat Committee (2008) ‘Genstat for Windows.’ 11th edn. (VSN International: Hertfordshire, UK)
Hocking Edwards JE (1999) Reduction in wool follicles prior to birth in Merino sheep. Reproduction, Fertility and Development 11, 229–234.
| Reduction in wool follicles prior to birth in Merino sheep.Crossref | GoogleScholarGoogle Scholar | 1:STN:280:DC%2BD3czovFensg%3D%3D&md5=b6dec18252d4d2a098ae732963e8c3c4CAS |
Hocking Edwards JE, Birtles MJ, Harris PM, Parry A, Paterson E, Wickham GA, McCutcheon SN (1994) Prenatal follicle development in Romney, Merino and Merino-Romney cross sheep. Proceedings of the New Zealand Society of Animal Production 54, 131–134.
Hocking Edwards JE, Copping KJ, Thompson AN (2011) Managing the nutrition of twin-bearing ewes during pregnancy using Lifetimewool recommendations increases production of twin lambs. Animal Production Science 51, 813–820.
| Managing the nutrition of twin-bearing ewes during pregnancy using Lifetimewool recommendations increases production of twin lambs.Crossref | GoogleScholarGoogle Scholar |
Huisman AE, Brown DJ, Ball AJ, Graser HU (2008) Genetic parameters for bodyweight, wool, and disease resistance and reproduction traits in Merino sheep. 1. Description of traits, model comparison, variance components and their ratios. Australian Journal of Experimental Agriculture 48, 1177–1185.
| Genetic parameters for bodyweight, wool, and disease resistance and reproduction traits in Merino sheep. 1. Description of traits, model comparison, variance components and their ratios.Crossref | GoogleScholarGoogle Scholar |
Hutchison G, Mellor DJ (1983) Effects of maternal nutrition on the initiation of secondary wool follicles in foetal sheep. Journal of Comparative Pathology 93, 577–583.
Jackson N, Nay T, Turner HN (1975) Response to selection in Australian Merino sheep. VII. Phenotypic and genetic parameters for some wool follicle characteristics and their correlation with wool and body traits. Australian Journal of Agricultural Research 26, 937–957.
| Response to selection in Australian Merino sheep. VII. Phenotypic and genetic parameters for some wool follicle characteristics and their correlation with wool and body traits.Crossref | GoogleScholarGoogle Scholar |
Jefferies BC (1961) Body condition scoring and its use in management. Tasmanian Journal of Agriculture 32, 19–21.
Kelly RW (1992) Lamb mortality and growth to weaning in commercial Merino flocks in Western Australia. Australian Journal of Agricultural Research 43, 1399–1416.
| Lamb mortality and growth to weaning in commercial Merino flocks in Western Australia.Crossref | GoogleScholarGoogle Scholar |
Kelly RW, Macleod I, Hynd P, Greeff JC (1996) Nutrition during fetal life alters annual wool production and quality in young Merino sheep. Australian Journal of Experimental Agriculture 36, 259–267.
| Nutrition during fetal life alters annual wool production and quality in young Merino sheep.Crossref | GoogleScholarGoogle Scholar |
Kelly RW, Greeff JC, Macleod I (2006) Lifetime changes in wool production of Merino sheep following differential feeding in fetal and early life. Australian Journal of Agricultural Research 57, 867–876.
| Lifetime changes in wool production of Merino sheep following differential feeding in fetal and early life.Crossref | GoogleScholarGoogle Scholar |
Kleemann DO, Walker SK (2005) Fertility in South Australian commercial Merino flocks: relationships between reproductive traits and environmental cues. Theriogenology 63, 2416–2433.
Oldham CM, Thompson AN, Ferguson MB, Gordon DJ, Kearney GA, Paganoni BL (2011) The birthweight and survival of Merino lambs can be predicted from the profile of liveweight change of their mothers during pregnancy. Animal Production Science 51, 776–783.
| The birthweight and survival of Merino lambs can be predicted from the profile of liveweight change of their mothers during pregnancy.Crossref | GoogleScholarGoogle Scholar |
Robinson JJ, Sinclair KD, McEvoy TG (1999) Nutritional effects on foetal growth. Animal Science (Penicuik, Scotland) 68, 315–331.
Safari E, Fogarty NM, Gilmour AR (2005) A review of genetic parameter estimates for wool, growth, meat and reproduction traits in sheep. Livestock Production Science 92, 271–289.
| A review of genetic parameter estimates for wool, growth, meat and reproduction traits in sheep.Crossref | GoogleScholarGoogle Scholar |
Saul G, Kearney G, Borg D (2011) Pasture systems to improve productivity of sheep in south-western Victoria. 2. Animal production from ewes and lambs. Animal Production Science 51, in press.
Schinckel PG (1955) The postnatal development of the skin follicle population in a strain of Merino sheep. Australian Journal of Agricultural Research 6, 68–76.
| The postnatal development of the skin follicle population in a strain of Merino sheep.Crossref | GoogleScholarGoogle Scholar |
Schinckel PG, Short BF (1961) The influence of nutritional level during prenatal and early postnatal life on adult fleece and body characteristics. Australian Journal of Agricultural Research 12, 176–202.
| The influence of nutritional level during prenatal and early postnatal life on adult fleece and body characteristics.Crossref | GoogleScholarGoogle Scholar |
Short BF (1955a) Development of the secondary follicle population in sheep. Australian Journal of Agricultural Research 6, 62–67.
| Development of the secondary follicle population in sheep.Crossref | GoogleScholarGoogle Scholar |
Short BF (1955b) Developmental modification of fleece structure by adverse maternal nutrition. Australian Journal of Agricultural Research 6, 863–872.
| Developmental modification of fleece structure by adverse maternal nutrition.Crossref | GoogleScholarGoogle Scholar |
Thompson AN, Doyle PT, Grimm M (1994) Effects of differential grazing of annual pastures in spring on sheep and wool production. Australian Journal of Agricultural Research 45, 367–389.
| Effects of differential grazing of annual pastures in spring on sheep and wool production.Crossref | GoogleScholarGoogle Scholar |
Thompson AN, Ferguson MB, Campbell AJD, Gordon DJ, Kearney GA, Oldham CM, Paganoni BL (2011) Improving the nutrition of Merino ewes during pregnancy and lactation increases weaning weight and survival of progeny but does not affect their mature size. Animal Production Science 51, 784–793.
| Improving the nutrition of Merino ewes during pregnancy and lactation increases weaning weight and survival of progeny but does not affect their mature size.Crossref | GoogleScholarGoogle Scholar |
Wheeler JL, Reardon TF, Hedges DA, Rocks RL (1971) The contribution of the conceptus to weight change in pregnant Merino ewes at pasture. The Journal of Agricultural Science 76, 347–353.
| The contribution of the conceptus to weight change in pregnant Merino ewes at pasture.Crossref | GoogleScholarGoogle Scholar |
Wheeler JL, Hedges DA, Mulcahy C (1977) The use of dyebanding for measuring wool production and fleece tip wear in rugged and unrugged sheep. Australian Journal of Agricultural Research 28, 721–735.
| The use of dyebanding for measuring wool production and fleece tip wear in rugged and unrugged sheep.Crossref | GoogleScholarGoogle Scholar |
Young JM, Thompson AN, Oldham CM (2008) Increasing profitability by pregnancy scanning ewes. Livestock Updates, Department of Agriculture and Food, WA, 1–2 July 2008, Perth.
Young JM, Thompson AN, Curnow M, Oldham CM (2011) Whole-farm profit and the optimum maternal liveweight profile of Merino ewe flocks lambing in winter and spring are influenced by the effects of ewe nutrition on the progeny’s survival and lifetime wool production. Animal Production Science 51, 821–833.
| Whole-farm profit and the optimum maternal liveweight profile of Merino ewe flocks lambing in winter and spring are influenced by the effects of ewe nutrition on the progeny’s survival and lifetime wool production.Crossref | GoogleScholarGoogle Scholar |