

Identifying indicator traits for breech strike in Merino sheep in a Mediterranean environment
J. C. Greeff A B , L. J. E. Karlsson A and A. C. Schlink AA Department of Agriculture and Food Western Australia, 3 Baron Hay Court, South Perth, WA 6151, Australia.
B Corresponding author. Email: Johan.greeff@agric.wa.gov.au
Animal Production Science 54(2) 125-140 https://doi.org/10.1071/AN12233
Submitted: 4 July 2012 Accepted: 13 March 2013 Published: 3 May 2013
Abstract
Data from 2833 unmulesed Merino lambs from 49 sires and 1535 dams that were born from 2005 to 2009, were used to identify indicator traits for breech strike in a Mediterranean environment. Sheep were managed in a normal production environment where no preventative treatments were applied to protect them from being struck by flies, but were immediately treated when found to be fly struck. Seventy-seven skin, body conformation, wool, disease and production traits that may contribute to breech strike and that could qualify as potential indicator traits were recorded on progeny at birth, weaning, post-weaning, yearling, spring and at pre- and post-hogget age at ~18 months of age. In all, 31% of ewes and 22% of rams were fly struck in the breech area between birth and hogget shearing. The heritability of and the phenotypic and genetic correlations between the potential indicator traits and breech strike were estimated. The results showed that breech strike is a heritable trait (h2 = 0.51 ± 0.10), and that the most important indicator traits for breech strike were dags, scored at any time, especially at hogget age (rg = 0.81 ± 0.15), spring (rg = 0.77 ± 0.13) or at weaning (rg = 0.64 ± 0.00), urine stain scored at weaning (rg = 0.54 ± 0.13) and marking (rg = 0.51 ± 0.15), neck wrinkle at hogget age (rg = 0.46 ± 0.13) and breech cover (rg = 0.32 ± 0.15) scored at post-weaning shearing. These traits could be used to select indirectly for breech strike resistance in a Mediterranean environment.
Additional keywords: breech cover, dags, genetic correlation, heritability, selection, skin wrinkles, urine stain.
Introduction
Fly strike, cutaneous myiasis, is one of the major diseases affecting wool-producing sheep. It is generally caused by the blowfly, Lucilia cuprina, in Australia, and can result in dramatic losses if not treated in a timely fashion. Moisture is crucial for the development of fly strike and most fly strikes occur in the crutch and tail region of sheep.
Extensive genetic experiments were carried out in the late 1920s and 1930s, to find genetic solutions to breech strike (Seddon et al. 1931; James 2006). However, with the advent of effective chemicals and surgical mulesing, genetic solutions were abandoned. Today, breech strike is still largely being controlled by surgical mulesing, regular crutching, jetting with insecticides and by treating individually struck sheep with chemicals. The annual costs of applying these preventative techniques are approximately AU$4/sheep (J. Larsen, pers. comm.) but the collective losses due to indirect factors and breech strike are generally accepted to be huge, especially in unmulesed sheep.
The practice of mulesing has recently become increasingly unpopular and a renewed effort was initiated by the wool industry in 2005 to find genetic solutions to breech strike. Several genetic studies have been carried out during the past 70 years to identify the causal factors of fly strike (Seddon et al. 1931; Watts and Merritt 1981; Gherardi et al. 1983; Lewer et al. 1987; Raadsma and Rogan 1987; Raadsma 2000; James 2006; Scholtz et al. 2010). This has led to the identification of body wrinkle, wool colour, fleece rot, dermatophilosis and fibre variability traits as potential indicator traits to breed indirectly for body strike resistance (Raadsma and Rogan 1987). However, until recently, very little genetic research has been carried out to identify potential indicator traits for breech strike in Merino sheep. This was probably due to the effectiveness of the mulesing operation that was introduced in the early thirties, regular crutching and jetting sheep with chemicals. Several studies have indicated that faecal soiling (dags) in the breech, wrinkles and bare area in the breech (Scobie et al. 2008, 2002, 2007; Edwards et al. 2009; Greeff and Karlsson 2009; Smith et al. 2009; Scholtz et al. 2010) are the main candidates as potential indicator traits for breech strike resistance. The size of the bare area in the breech received strong focus of attention as a solution for surgical mulesing, as removal of wool around the anus through breeding reduces the potential to develop dags, as was shown by Scobie et al. (2008) in Romney sheep. However, apart from the preliminary studies by Greeff and Karlsson (2009), no study to elucidate the role of dags, skin wrinkle, body conformation and wool traits that may contribute to breech strike has been carried out on Merino sheep in a Mediterranean environment.
The present paper investigated a wide range of skin, body conformational, visual wool and the wool production traits in Merino sheep to elucidate factors contributing to breech strike. The sheep were managed without preventative treatments for breech strike in a Mediterranean environment, so as to identify indicator traits that could potentially be used to breed indirectly for resistance to breech strike.
Materials and methods
The trial was approved by the Animal Ethics Committee of the Department of Agriculture and Food Western Australia (AEC 4-05-30 and AEC 1-02-01) and was carried out according to the Code of Practice of using animals in experiments.
Design
The experimental flock was established in 2006 and consisted of three lines, namely a Select A line, Select B line and an unselected Control line, to demonstrate whether selecting for breech strike resistance will result in genetic changes in flocks. The Select A line represented a scenario of a ram breeding flock that was selected intensely against breech strike by using all breech-strike and the known indicator-trait information to determine what progress can be made in breech strike resistance, and the correlated responses in production traits. The Select B line represented a scenario of a commercial flock that buys rams from a ram breeding flock that selects intensively for breech strike resistance, and where replacement ewes are culled on breech strike. The unselected control line was established to compare the genetic changes in the incidence of breech strike in the Select A and Select B lines.
Modified design
To facilitate a larger database for more robust genetic parameter estimates of potential indicator traits for breech strike, the Rylington Merino (RM) flock was included in this experiment in 2008. This flock consists of 500 breeding ewes mated to 10 sires annually and has been a closed flock since its inception in 1987. Karlsson and Greeff (2006) described the establishment of the flock and the genetic changes that have occurred in this flock due to selection for low faecal worm egg count.
Animals
Sourcing mature ewes
Six hundred mature Merino ewes were sourced from the Department of Agriculture and Food Western Australia (DAFWA) experimental flocks. Previous research and other anecdotal information from industry has shown that moisture content, fleece rot, dermatophilosis, wool cover, quantity of dags, bare skin area around the anus and vulva, amount of wool wax, urine stain, wool colour to indicate suint content and wrinkles may play a role in body strike (Belschner 1953; Dun and Eastoe 1970; Watts and Merritt 1981; Raadsma and Rogan 1987). As the mature ewes had been mulesed, wrinkles in the breech area were not considered to be a reliable indicator of wrinkles; therefore, neck and body wrinkles were used to identify plain-bodied ewes using the visual-scoring charts published by AWI (2007).
The mature Merino ewes from four experimental flocks of the Department of Agriculture of Western Australia were classed before shearing in December 2005. These flocks were the Rylington Merino flock, the Mount Barker Research station general commercial flock, the Great Southern Agricultural Research Institute experimental flock at Katanning and the Badgingarra Research station general flock. The Rylington Merino flock and the breech strike flocks were managed together since 2006 and therefore received exactly the same treatment. However, with its inclusion in the breech strike experiment in 2008, seven of the best performing rams from the breech strike experiment on the 7% dual-purpose index of Sheep Genetics were also used in the Rylington Merino flock to increase the genetic links between these two flocks. The Rylington Merino flock contributed 20% of the ewes and 14 of the 49 sires to the breech strike flock. These 14 rams produced 496 progeny, which indicates that the RM and breech flocks were genetically well connected.
Ewes with low wrinkle, low breech cover, white wool colour, low dags, no fleece rot, no dermatophilosis and that had no history of fly strike, were selected for the Select A line, while the ewes for the control (C line) and Select B line were selected randomly among the remaining ewes in each flock. In total, 600 ewes were selected, consisting of 200 ewes per line. Equal numbers were selected for each line from each of the four flocks. The ewes were transferred to the Mount Barker research station in January 2006. The research station is situated at 34°34′39″S, 117°31′10″E.
Sourcing ewe lambs from industry flocks
Selection of sheep for each line was carried out as follows. Sheep were sourced from 10 industry flocks and these flocks had to consist of at least 300 mature breeding ewes before they could contribute ewe weaners to the experiment. Only ewe lambs that were born during June to August 2005 were considered for selection. The 2005-born ewes were initially identified on the lamb-marking cradle.
All ewe lambs in a particular flock were measured or scored for tail wrinkle, breech wrinkle, breech cover and wool colour by using the Australian Wool Innovation (AWI) visual-scoring system. Thirty ewe lambs in each flock were initially scored to obtain an estimate of the mean and variation between ewes for each trait. Then, from each flock, 22 ewe lambs were randomly identified for the control line, 22 average ewe lambs were then identified for the Select B line and the best 22 lambs with a score of one (1) for tail and breech wrinkle (no wrinkles) were selected for the Select A line among the remainder of the lambs.
Half of each group were mulesed and the other half were left unmulesed on an alternate basis as the lambs came through the lamb-marking cradle. However, for the present paper, only unmulesed animals were included in the analysis. Following selection of the experimental sheep at lamb marking, they remained on the property with their dams until weaning. The selected young ewe lambs were tagged with a unique identification tag at lamb marking. These animals were then collected from each participating farm after weaning and transported to the Manurup Annex, Mount Barker research station in Western Australia, on December 2005. Any of the Select A lambs that subsequently may have become fly struck were not included in the final selection for this line.
Sourcing rams for the different lines
All available information on potential indicator traits and breech strike were used to identify suitable rams for the Select A and Select B lines. Merino rams with low breech wrinkle, low breech cover, white wool colour, low dags, no fleece rot, no signs of dermatophilosis and that had no history of fly strike were selected for the Select A and Select B lines. Twenty-five different rams were used during the first 2 years, of which nine rams were sourced from industry flocks, eight from the Rylington Merino flock and eight rams from the Katanning flock. A large fully pedigreed database was available on dags, worm egg count, production traits and breech strike records from the Rylington Merino and Katanning flocks. The rest of the rams were sourced from industry flocks when it was considered that they could contribute to the breech strike flock by increasing the variation among animals. However, very limited objective information was available on most of the industry sires sourced.
The control rams were chosen from the Katanning experimental flock. The sheep in this flock were recorded extensively for all the economically important production traits as well as for wrinkles and dags, and it has been shown to be a very good representation of a typical Western Australian Merino flock (Greeff and Cox 2006). This information was used to select rams on bodyweight and wool production traits, to ensure that they were genetically representative of industry flocks.
Replacements
After the first year, the ewes from Select A line that were more susceptible to breech strike, and had high scores for urine stains, occurrence of dags and wrinkles were culled. Older ewes in Select B and control lines were culled randomly but within age groups to ensure that the age structure of the three lines stayed relatively similar. Sheep were scored at the annual selection before shearing and culled if found unfit for the experiment. The majority of the mature ewes were culled for age in 2007 and 2008 and replaced by the 2005-born ewes that were sourced from industry flocks and from ewes bred from within the flock.
In 2008, most of the replacement rams used were bred from within the flock but one outside sire with strong industry links was also used, as well as two rams from the CSIRO breech strike flock at Chiswick, Armidale (Smith et al. 2009), to generate genetic links between these two experimental flocks.
Management
Mating
Each line consisted of 200 ewes mated to four sires, with 50 ewes each. The mature ewes were randomly allocated to the sire groups within each line, as all the rams and ewes were unrelated. Mating was carried out over a 5-week period, starting in mid-February each year. Semen was obtained from selected industry rams in the first 2 years of the experiment to inseminate the mature ewes of the Select A and B lines, whereas the selected rams from the Rylington Merino flock and the control rams from the Katanning flocks were used through natural mating in the selection and control lines, respectively. All the inseminated ewes were pooled after insemination into two groups within selection line and mated in the following oestrus cycle to a ram from each line. This backup mating allowed for the fertilisation of ewes that did not conceive from the artificial insemination. This increased the number of single-sire groups from 12 to 14 in 2006 and 2007.
A rotational mating system was followed in which ewes stayed in the family group they were born in, while rams were replaced by their best performing son for each line, and used in the following family group other than the birth group that they were born in. To create genetic links across years, a minimum of two rams that were used previously, were used again in the following years but in different family groups according to the rotational mating system.
Lambing
Two weeks before lambing, the ewes were drafted into their single-sire mating groups as at mating, and placed on lambing plots for lambing. Lambs were weighed and recorded for sex, birth coat and birth type within 24 h after birth and given a unique identification tag. Each lamb was mothered-up with its dam; thus, full sire and dam pedigrees were available on all animals.
Management treatments
In this Mediterranean environment, breech strike generally occurs during a relatively small time frame from mid-October to end of December. However, a small number of strikes may occur in summer and late autumn. The fly challenge was monitored during the year by using fly traps for 1 day of the week from September to May inclusively. This information, combined with local weather station data, was used to predict the occurrence of fly strike in the flock.
For the 2006- and 2007-born progeny, half of the lambs of each sire within each of the three lines (Select A, Select B and Control) were mulesed and the other half were left unmulesed by only mulesing every second lamb of each sire. Mulesing was discontinued in 2008 to generate more information on unmulesed sheep. All ram lambs were left entire. After lamb-marking, the different sire lambing groups were combined and managed as one flock until lambs were weaned at about 4 months of age. At this age, the ram and ewe lambs were separated, and subsequently managed in two separate flocks.
The sheep in this experimental flock were exposed to two fly strike seasons, first from birth until weaner shearing at ~5 months of age in early December, and then again from weaner shearing until hogget shearing 12 months later.
No preventative crutching was carried out until hogget shearing as this would have had an impact on the incidence of breech strike. Hoggets were crutched immediately before shearing to remove stained and daggy wool to prevent fleece contamination at shearing. Faecal worm egg counts were regularly collected to monitor the sheep for worm infection. It is important to note that most of the scouring and development of dags in hoggets at Mount Barker is due to a hyper-sensitivity reaction to ingested immature worm larvae and not to an established high adult worm burden (Larsen et al. 1994; Williams et al. 2010).
No preventative chemical treatments against breech strike were applied during this experiment. The sheep were monitored every 1–3 days, depending on the predicted rate of blowfly challenge. Sheep that were positively identified as being struck were treated immediately. The wool around the strike area was clipped away and the area treated with the short-acting chemical Spinosad (Elanco Animal Health, Australia) according to the manufacturer’s recommendations. Treated animals were spray-marked for easy identification and then returned to the flock. Their subsequent health and recovery were continuously monitored and they were treated again if necessary.
Data
The data file consisted of 2833 records of unmulesed sheep from 49 sires and 1535 dams born from 2005 to 2009. After cleaning the dataset, 2581 records were kept, which resulted in an average of 41 progeny per sire that ranged from 2 to 106 animals.
All fly strikes were recorded continuously from birth up to post-hogget shearing but only breech strikes are reported in the present study. For the purposes of the Gibbs analysis (Misztal et al. 2002), the number of breech strikes from birth to hogget shearing were recorded as 1 for no strikes, 2 for 1 strike, 3 for 2 strikes, 4 for 3 strikes and 5 for 4 strikes. The visually scored wool, body conformation, skin traits and dags were scored from 1 to 5, using the visual sheep-scoring system (AWI 2007), where 1 was low or none and 5 was the highest for the trait. However, birth coat was scored on a 1–6 point scale. These traits were recorded on each sheep at different times from birth to post-hogget shearing and are shown in Table 1. Additional traits such as wrinkle on the rump, belly cover, dag moisture content, tail wrinkle and tail bareness at marking and urine stain, and traits such as hairy birth coat, white shoulder and a belly pluck score were also recorded. A rump and a tail wrinkle (1 = no wrinkles, … 5 = very wrinkly) score were also recorded in 2008 and 2009 because it appeared that some sheep may be very plain on the rump but display clear wrinkles on the tail. It was assumed that these tail wrinkles could predispose sheep to be struck. Dag moisture content (1 = dry dags, …5 = very wet dags) was scored separately from the quantity of dags, to determine whether wetness of the dags is more important than the quantity of dags.

|
Dermatophilosis was scored in spring on a 1 (no signs) to 5 (severe dermatophilosis) point scale. Fleece rot and wool colour of a midside were also scored in spring before weaner and hogget shearing, using the visual sheep-scoring system (AWI 2007). An additional trait that consists of a white patch that is sometimes found between the shoulder blades on sheep in this environment, was also scored to determine whether this characteristic could be correlated with breech strike. It was scored on a 1 (low) to 5 (very white) point scale. A belly pluck score was also given on all animals from 2009, to determine whether the looseness of the wool on the belly could be related to wool coverage across the body later in life. A bundle of wool was plucked from the belly and the ease and amount of wool plucked were scored from 1 (low) to 5 (high). In all cases, half scores were given to visually scored traits where appropriate.
Bare area around the anus (bareness across anus and length of bareness down from the root of the tail), tail length, tail width and the bare length and bare width on the inside of the tail were measured in centimetres with a measuring apparatus at marking. Faecal worm egg count (WEC) at weaning and in spring was measured as the number of eggs per gram faeces at weaning and at hogget age as this could be related to the presence of dags. WEC was measured as the number of worm eggs per gram faeces at weaning and at hogget age as this could be related to the presence of dags. FS was also scored on the faeces collected for the WEC on a 1 (hard pellets) to 5 (fluid faeces) point scale, to determine whether it is correlated with breech strike.
Lambs were shorn as weaners in late spring and fleece weights were recorded 12 months later at hogget age. A midside wool sample was collected on each animal and tested in a commercial laboratory for fibre diameter (FD), standard deviation of fibre diameter (FDSD), coefficient of variation of fibre diameter (FDCV), proportion of fibre below 15 micron (F <15), proportion of fibre above 30 micron (F >30), coarse edge (CE), comfort factor (CF), staple strength (SS), staple length (STPL), yield (Yld), fibre curvature (Cur) and standard deviation of fibre curvature (SDCur). The percentage yield was used to calculate clean fleece weight (CFW).
Statistical analyses
Summary statistics were calculated with Genstat (2011). The data were then analysed with ASREML (Gilmour et al. 2006) and a series of univariate analyses were carried out by fitting every trait in Table 1 as a dependent variable. The non-normally distributed traits were analysed using linear models with and without transforming the data, to identify the fixed factors that affected these traits significantly. The incidence of fleece rot and dermatophilosis was extremely low in this flock and therefore not included in the analysis. The following animal model was fitted for all traits.
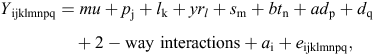
where Y = a vector of observations for one trait, mu = overall mean for that trait, pj = effect of property of origin of ewes (j = 1–10), lk = effect of selection lines Select A, B and C and RM flock (k = 1–4), yrl = effect of birth year 2005–09 (l = 1–5 for indicator traits and l = 1–4 for breech strike), sm = effect of sex: male or female (m = 1 or 2), btn = effect of birth type: single, twin or triplet (n = 1–3), adp = effect of age of dam: 7 age groups (P = 2–7+), dq = effect of qth day of birth: days from 1 January, ai = random effect of ith animal and eijklmnpq = residual effects.
Property was fitted as a fixed factor to accommodate the effect of origin of the animals. All two-way interactions were initially fitted, but factors were dropped from the model if found to be not significant.
Genetic maternal and permanent environmental maternal effects were initially fitted with ASREML, but had no significant effect on breech strike or on any of the visually scored traits. Genetic maternal effects were significant (P < 0.05) only for CFW and explained 3% of the total variation. These effects were therefore dropped from the model.
The genetic analyses were carried out by using the Gibbs sampling procedure of Misztal et al. (2002). This software is suitable for estimating the variance components for any combination of categorical and continuous traits (Lee et al. 2002; Cloete et al. 2009). Breech strike was treated as a categorical trait, as the Gibbs methodology accommodates categorical threshold traits and it assumes that the underlying genetic distribution of these traits is normally distributed.
All the potential indicator traits, including the non-normally distributed traits and the traits that were scored on a 1–5 point scale, were analysed with the Gibbs procedure without transforming the data. Traits such as urine stain and dag scores that were not normally distributed, and that had a low incidence at certain times of the year, were analysed as threshold and as continuous traits. The heritability estimates of these two methods were generally similar, except that the threshold approach resulted in unrealistically high variance components for these traits. As the Gibbs methodology accommodates non-normal distributed traits, all the potential indicator traits were analysed using linear models and assuming a continuous underlying distribution.
The environmental factors that were significant (P < 0.05) in the univariate ASREML analysis were fitted as fixed effects in this analysis. The Gibbs analysis was carried out by fitting a two-trait model between breech strike and the potential indicator traits.
The equation for the two-trait model was as follows:
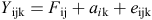
In this model, Yijk is a vector of observations where i indicates the trait (i = 1 or 2), where 1 equals total breech strike and 2 the potential indicator trait. Fij consists of the significant fixed effects j of trait i, aik is the additive genetic effect of the kth animal for the ith trait, and eijk is the vector of randomly distributed residual effects.
THRGIBBSF90 software (Misztal et al. 2002) was used to obtain the variance components to estimate the heritability of, and the genetic, environmental and phenotypic correlations between breech strike and the indicator traits. The POSTGIBBSF90 software was used for the post-Gibbs analysis (Misztal et al. 2002), which allows for the prediction of solutions for fixed and random effects.
A single chain of 200 000 cycles was run for each analysis, with the first 40 000 samples used as the burn-in period, as required by this methodology (Donoghue et al. 2004; Ntzoufras 2009). Every 10th sample was stored after 40 000 iterations, giving a total of 16 000 values for the computation of posterior means, posterior standard deviations (PSD) and the 95% highest posterior density confidence intervals. The success of the Gibbs sampling process depends on the variance components stabilising during iterations (Ntzoufras 2009). The sample values were then plotted against iterations and inspected to determine whether the values stabilised. Visual assessment of the sampled genetic variance and residual values against iteration shows that the sampling process has stabilised for all the traits reported in the present study.
Point estimates were calculated of the additive genetic and residual variance component parameters. Although breech strike and the visually scored traits had several thresholds in the present study, it was assumed that breech strike and the indicator traits followed a normal distribution on the underlying scale and consisted of a combination of additive genetic and environmental effects. The heritability of each trait, as well as the genetic and phenotypic correlations between breech strike and the potential indicator traits, were estimated according to methods from Falconer (1989).
Responses to selection were predicted using the breeder’s equations for the direct and indirect correlated responses (Falconer 1989). In this scenario, the direct response to selection depends on the incidence of the disease as it affects the selection differential. If the incidence of breech strike is 20%, with a heritability of 0.57, then it is predicted that the incidence of breech strike will be reduced by 14% in one generation of selection. The ratio of the correlated response (CR) to the direct response (D) can be used to determine which indirect traits would be the most effective indicator traits for breech strike in breeding programs. Where the selection intensities of the direct and correlated traits are the same, this ratio is given as:
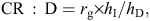
where rg = genetic correlation between breech strike and the indicator, hI = square root of heritability of indicator trait and hD = square root of heritability of breech strike (Falconer 1989).
The selection intensity of breech strike will, in most cases, be different from that of the indicator trait. However, for different incidences of breech strike, the ratio of the two selection intensities will be a constant and for the purposes of this exercise it was assumed that it is the same and will not affect the final conclusion. This method was therefore used to identify the most important indicator traits that will give at least 30% of the response by selecting directly for breech strike resistance.
Results
Tables 2–4 show the number of records, mean, minimum, maximum, standard deviation, skewness and kurtosis of breech strike and the subjectively scored body, skin, faecal, urine and wool traits. Table 5 shows the objectively measured bare area, tail traits, and the faecal consistency and faecal WECs. The number of records differs, because some traits were not recorded in the initial years but were recorded in the subsequent years as more information became available, which indicated that other potential traits may contribute in making animals more susceptible or resistant to breech strike.

|

|

|

|
The average scores for wrinkle traits in this population were relatively low, as indicated by the low average scores on the breech, rump, body and tail wrinkle at yearling and post-hogget shearing.
Skewness and kurtosis were estimated for all traits in the present study, and the results in Tables 2–5 show that most of the dag score, urine stain and wrinkle traits, as well as WECs had skewed distributions.
Table 2 shows that the breech cover traits were generally normally distributed (skewness < 1.0). Belly cover was skewed to the left at yearling but skewed to the right at post-hogget shearing. The skin wrinkle traits were generally skewed to the right (Table 3) while some had very high (>3) kurtosis values. Dag score (Table 4) recorded up to yearling age was generally right-skewed but slightly skewed to the left when recorded in spring and before hogget shearing. Urine stain scored on ewes only was generally skewed to the right with high kurtosis, except for urine stain at weaning, which was skewed to the left and with low kurtosis.
Table 5 shows that WECs at weaning and in spring were skewed to the right while all the other traits were normally distributed. The results of the posterior density distribution of the traits (not shown) showed that although breech strike on the normal scale is a categorical threshold trait, with thresholds at zero, one, two and three, the additive genetic and residual density distributions for total breech strike followed a normal distribution pattern. This pattern was also found for all the potential indicator traits, whether they were fitted as a threshold or continuous trait.
The flock in the present study produced a medium-type wool that is very representative of sheep in this region. Table 6 summarises the different wool traits. Most of the wool traits had a normal distribution, except F<15 and F>30, CE and CF, which had skewed distributions.

|
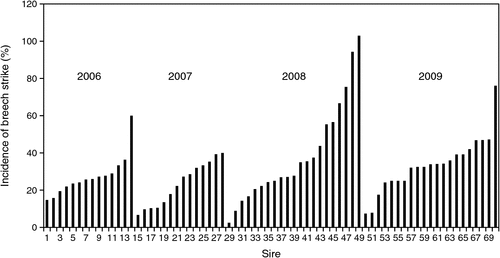
Incidence of breech strike
Figure 1 shows the raw means of the incidence of breech strike from birth to hogget shearing among progeny groups from different sires born from 2006 to 2009. On average, 31% of ewes and 22% of rams were struck in the breech from birth to hogget shearing from 2006 to 2010. Thus, the sheep received a reasonable challenge to identify genetic differences among sheep. Large differences were found among sire groups within each year. The most resistant and most susceptible groups were found in 2008. The most resistant sire progeny group had a strike rate of only 2.4%, while 103% of the progeny of the most susceptible sire group were struck in the breech. Some animals in the last mentioned progeny group were struck up to three times, which resulted in a more than 100% strike rate. It is clear that marked variation exists among progeny groups from different sires, which indicates that genetic differences are playing a significant part in differences in breech strike among sires.
|
Environmental factors
Year of birth and line affected all the traits in the present study significantly (P < 0.05). Sex affected most traits significantly (P < 0.01), except for Cur. A year × sex interaction was found for some traits that were recorded after weaning. This interaction was most likely caused by different management regimes imposed on the lambs because they were separated on sex at weaning and then subsequently run in different management groups. Property of origin also affected some wrinkle, breech cover and wool traits significantly (P < 0.05), which could be due to different selection protocols and breeding programs on these properties.
Age of the dam had no significant effect on any of the conformational and visually assessed wool traits, but had a significant (P < 0.05) effect on CFW and FD. Progeny of older dams produced more and broader wool.
Type of birth affected most wrinkle and breech cover traits significantly (P < 0.01). It was found that multiple-born lambs were generally plainer and had higher breech cover scores. Breech cover of twins were 3% lower than that of singletons at weaning and the difference increased to 13% and 15% at yearling and hogget classing, respectively. Interestingly, the effect was not significant for face and crutch cover, but triplets had significantly (P < 0.01) lower face and crutch cover scores. Singletons were also up to 17% more wrinkled in the breech than were twin-born animals at hogget classing.
Heritability estimates
Breech strike
Breech strike has a moderately high heritability on the underlying scale, as shown in Table 7. The heritability of breech strike up to weaner shearing, and from weaner shearing up to hogget shearing, was the same, namely 0.57 ± 0.13 and 0.57 ± 0.16, respectively, while the total heritability of breech strike from birth to hogget shearing was 0.51 ± 0.10.

|
Bare skin area and wool cover traits
The heritability estimates of the subjectively scored skin and wool cover traits are shown in Table 8. The belly pluck score had a very low heritability of 0.06 ± 0.03, which indicates that this trait may not be useful as an indicator trait. Face cover at weaning and yearling had the highest heritability estimates of 0.77 ± 0.07 and 0.73 ± 0.05, respectively. However, at post-hogget shearing, the heritability of face cover dropped to 0.31 ± 0.06. All the breech cover traits were heritable, variance ratios increasing from ~0.25 ± 0.06 at marking to 0.48 ± 0.07 at post-hogget shearing. Crutch cover was generally more heritable than breech cover at weaning, yearling and hogget age, while the heritability of the belly cover traits was very similar to that of the breech cover traits at the same stage. Heritability of belly and of breech cover was highest at hogget age, but the heritability of crutch cover declined with age.
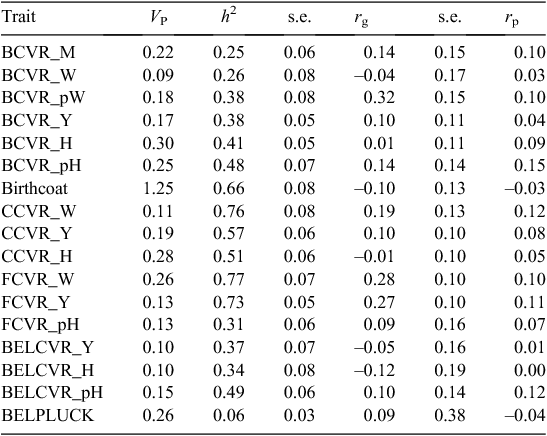
|
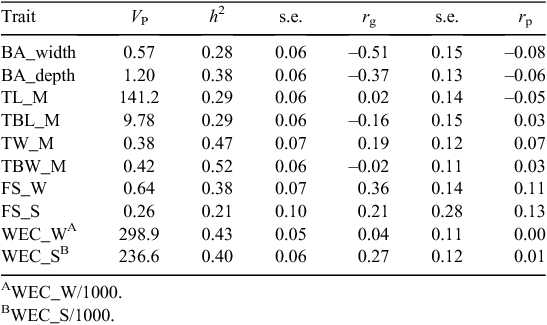
Bare perineum areas across and along the depth of the anus at marking were both heritable (BA_width: h2 = 0.26 ± 0.06; BA_depth: h2 = 0.38 ± 0.06), but BA_depth displayed more additive genetic variation than did BA_width (Table 11).
Tail bare width (TBW_M) and bare depth (TBL_M) on the inside of the tail at marking were also scored to determine whether the bare area under the tail contributed to increased resistance to breech strike. Both of these traits were heritable (h2 = 0.52 ± 0.06 and 0.29 ± 0.06, respectively, for TBW_M and TBL_M), which indicates that they will respond to selection (Table 11).
Faecal traits
Moisture content of the dags (DMDags) had a heritability of ~0.25 at weaning, spring and at hogget age, but the heritability was slightly lower at yearling age, i.e. 0.20 ± 0.11 (Table 9). However, the heritability of dags was ~0.35 at marking, post-weaning, spring and hogget age, but was also much higher (h2 = 0.63 ± 0.08) at yearling age. The dag score at yearling age was based on 1909 records collected over 4 years. Thus, it appears that dag score has a much higher heritability at yearling age than at other times.
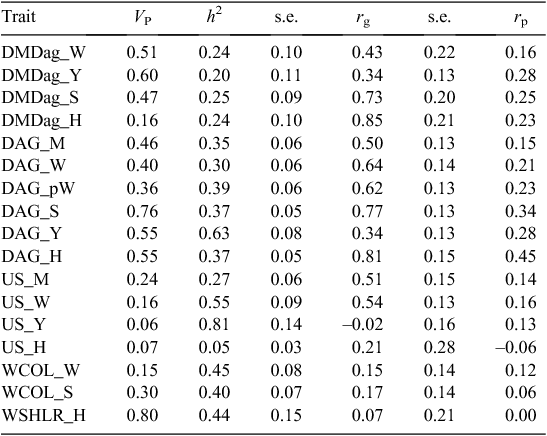
|
Faecal WECs at weaning and in spring (Table 11) were moderately heritable (h2 = 0.43 ± 0.05 and h2 = 0.40 ± 0.06, respectively). Faecal consistency score (Table 11) of the faecal samples that were collected for faecal WECs at weaning and in spring had a heritability of 0.38 ± 0.07 at weaning and 0.21 ± 0.10 in spring.
Urine traits
Urine stains were scored only on females and the average urine stain means were generally very low (Table 4). However, Table 9 shows that this trait was highly heritable at yearling (h2 = 0.81 ± 0.14) and at weaning (h2 = 0.55 ± 0.09), but much lower at marking (h2 = 0.27 ± 0.06) and very low (h2 = 0.05 ± 0.03) at hogget age. This low value at hogget age was most likely caused by the presence of dags as the animals were not crutched. This made it virtually impossible to score urine stain accurately in ewes with daggy breeches.
Wrinkle traits
Wrinkles were scored across the body (neck, body, breech, tail, rump) at marking, weaning, post-weaning and post-hogget shearing. Table 10 shows that the heritability of all wrinkle traits was moderately low to high.
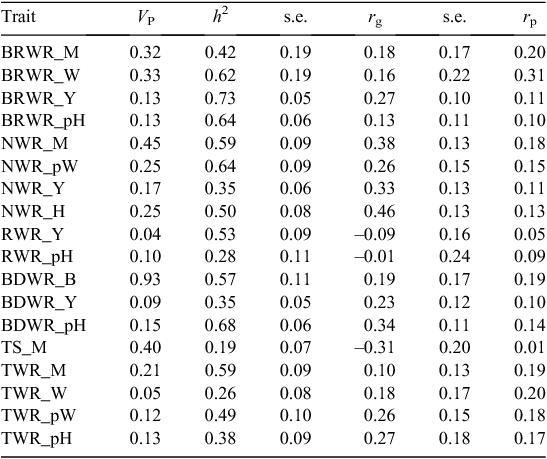
|
|
Breech wrinkle at yearling age had the highest heritability (h2 = 0.73 ± 0.05), followed by body wrinkle at post-hogget shearing (h2 = 0.68 ± 0.06), neck wrinkle post-weaning (h2 = 0.64 ± 0.06), breech wrinkle at weaning (h2 = 0.62 ± 0.19), and breech wrinkle post-hogget shearing (h2 = 0.62 ± 0.06). The wrinkle traits that had the lowest heritability estimates were tail wrinkle at weaning (h2 = 0.26 ± 0.08) and wrinkle on the rump at post-hogget shearing (h2 = 0.28 ± 0.11). Most wrinkle traits in the studied population had quite low mean averages (<1.5), except for body wrinkle at birth, neck wrinkle at marking and post-weaner shearing.
Wool traits
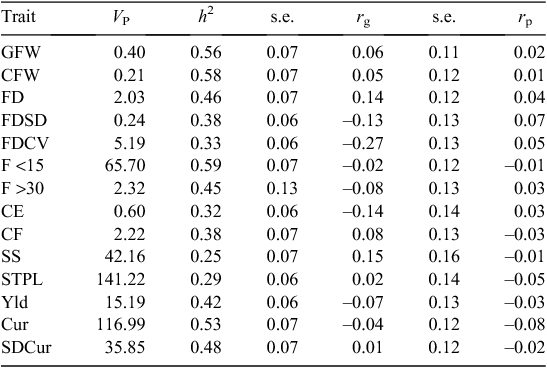
The heritability estimates of the wool and fibre traits are shown in Table 12. In general, the heritability estimates found in the present study agree well with estimates from Australian sheep flocks (Safari et al. 2005; Huisman et al. 2008), indicating that these traits will respond to selection. The heritability estimates for proportion of fibres below 15 micron and for proportion of fibres above 30 micron were 0.59 and 0.45, respectively. No comparative estimates could be found in the literature for these two traits.
|
Wool colour was a moderately heritable trait, with a heritability of 0.45 ± 0.08 and 0.40 ± 0.07 at weaning and hogget age, respectively (Table 9). The heritability estimates of the objectively measured wool traits are shown in Table 12.
Relationships between potential indicator traits and breech strike
Breech strike from birth to weaning was genetically moderately correlated with breech strike from post-weaning up to hogget shearing (rg = 0.44 ± 0.17). The environmental correlation was virtually zero, which resulted in the phenotypic correlation of 0.29.
The genetic and phenotypic relationships between total breech strike and the subjectively scored visual skin, body conformation and wool traits are shown in Tables 8–12.
Bare area and wool coverage traits
Table 8 shows that the phenotypic correlation between breech strike and breech cover was generally low. The highest phenotypic correlation was found between BCVR_pH and breech strike (rp = 0.15). The highest genetic correlation was found when the animals were scored post-weaning after weaner shearing (rg = 0.32 ± 0.15). The correlations between crutch cover scored at the different ages and breech strike were also found to be generally low. The same trend was found for belly cover at yearling, and at pre- and post-hogget shearing. Belly pluck score was not genetically or phenotypically correlated with breech strike.
Face cover (Table 8) at weaning and at yearling was genetically correlated with breech strike (rg = 0.27 and 0.28, respectively, with a standard error of 0.10). The highest genetic relationship between breech strike and the bare area traits was found for BA_depth and BA_width (Table 11). Both these traits were genetically moderately correlated with breech strike (–0.51 ± 0.15 and –0.37 ± 0.13, respectively). Bare area under the tail at marking was not significantly genetically correlated with breech strike.
Faecal traits
Table 9 shows that dag score and dag moisture content of the dag traits measured at any stage were on average the strongest correlated with total breech strike of all the traits scored. The genetic correlation between dag score and total breech strike ranged from 0.34 ± 0.13 at yearling age, to 0.81 ± 0.15 before hogget shearing. Dag moisture content scored in spring and before hogget shearing followed the same pattern. The phenotypic correlation between breech strike and dag score varied from 0.15 at marking to 0.34 in spring, and 0.45 before hogget shearing. All the correlations with dag moisture content scored at the same age were lower, 0.25 and 0.23, respectively.
Table 11 shows that WEC in spring had a moderately low genetic relationship with breech strike of 0.27 ± 0.12 in spring, but no relationship with WEC at weaning. However, FS and DMdags had moderate genetic correlations (0.36 ± 0.14 and 0.43 ± 0.22) with breech strike at weaning.
Urine traits
Urine stain was recorded only on ewes. Table 9 shows that it was genetically moderately strongly correlated with total breech strike when scored at marking (rg = 0.51 ± 0.15) and at weaning (rg = 0.54 ± 0.13). The phenotypic correlations at these times were generally positive but low, i.e. <0.16. Urine stain at yearling was not genetically correlated with breech strike, but a low genetic correlation was found again before hogget shearing. A moderately high environmental correlation was found at yearling age, although this was not significantly different from zero (re = 0.48 ± 0.27).
Wrinkle traits
The wrinkle traits measured at different stages on different sites on the body were generally phenotypically and genetically positively correlated with total breech strike and are shown in Table 10. The phenotypic correlations varied from 0.05 measured on the rump at yearling, to 0.31 measured in the breech at weaning. Apart from a non-significant negative genetic correlation between breech strike and wrinkle score measured on the rump at yearling and hogget age, all other genetic correlations between the wrinkle traits and breech strike were positive. The strongest genetic correlations were found between neck wrinkle measured at yearling (rg = 0.46 ± 0.13), marking (rg = 0.38 ± 0.13) and at hogget age (rg = 0.33 ± 0.13). Except for breech wrinkle at yearling (rg = 0.27 ± 0.10), all the other genetic correlations between breech wrinkle and breech strike were not significantly different from zero. The genetic correlation between breech strike and body wrinkle at birth was not significantly different from zero (rg = 0.19 ± 0.17), but it increased to 0.23 ± 0.12 at yearling and to 0.34 ± 0.11 at hogget age.
Wrinkle was also scored independently on the tail of the lambs at marking, at pre- and post-lamb shearing and again at post-hogget shearing. The phenotypic and genetic correlations between total breech strike and tail wrinkle scored after weaner and hogget shearing were moderately low (rp ~0.2 and rg ~0.26), which indicates that tail wrinkle was not correlated with breech strike in the present study.
Wool traits
The genetic and phenotypic correlations between breech strike and wool production traits are shown in Table 12. FDCV was the only trait that was genetically significantly and unfavourably correlated with breech strike (rg = –0.27 ± 0.13). None of the genetic correlations was significantly different from zero, which implies that breech strike resistance was not correlated with wool quantity or wool quality traits. Similarly, wool colour or white shoulder traits were not correlated with breech strike resistance, as shown in Table 9.
Potential indirect indicator traits for breech strike
Table 13 shows the indicator traits that offer the best opportunity to be used to select indirectly for breech strike resistance.

|
Dags scored before hogget shearing, or scored in spring, followed by urine stain scored before weaner shearing, were the most important traits in unmulesed sheep that received no preventative treatment from crutching or jetting in this Mediterranean environment. Selecting on these traits will result in a response of ~60% of that which can be achieved by selecting directly for breech strike if the selection intensities are the same. The next-best wrinkle indicator trait was neck wrinkle scored at marking and at hogget age, followed by dags scored post-weaning and at yearling age. Of the wool cover traits, only face cover at weaning and yearling could be considered as potential indicator traits.
Discussion
The present study investigated the underlying genetic differences among sheep that contribute to breech strike in a Mediterranean environment. The paper focuses on identifying individual traits that could be used as indicator traits to breed indirectly for breech strike resistance. Seventy-seven traits were scored or measured at different times and up to post-hogget shearing. The phenotypic and genetic relationships among potential indicator traits, or among the same indicator trait but measured at different times, will be addressed in a subsequent study.
In all, 22% of rams and 31% of ewes were struck from birth to hogget shearing. This indicates that the sheep in the present study received an acceptable blowfly challenge to identify the underlying causes of breech strike. Variation was found in the means of the same trait scored at the same site on the body but scored at different times of the year for virtually all the potential indicator traits investigated in the study. Due to the confounding nature of season and age, it was not possible to distinguish whether these large differences were caused by age or season, or both, but season certainly played a dominant role, especially for dags as this trait clearly followed the annual rainfall pattern (Karlsson et al. 2004).
Breech strike in this Mediterranean environment is a heritable trait, with a h2 estimate of higher than 0.5 in unmulesed sheep. This estimate is slightly higher than the value estimated by Scholtz et al. (2010) for absence of breech strike, but significantly higher than the heritability of 0.32 reported by Smith et al. (2009) for breech strike. These differences in heritability estimates could possibly have been caused by different fly strike frequencies and different models, and also because Smith et al. (2009) treated breech strike as a continuous trait, whereas Scholtz et al. (2010) treated breech strike as a threshold trait, as was done in the present study. As this flock was composed of sheep from different unrelated genetic sources, strain or origin of flock effects could also have contributed to the higher heritability of breech strike in spite of the fact that the property of origin was fitted in the model to accommodate genetic groups. However, this clearly indicates that breech strike is heritable and that it would respond to selection provided that adequate challenge conditions prevail.
Phenotypic correlations between breech strike and indicator traits
A positive phenotypic correlation of 0.29 was found between breech strike early in life (before weaner shearing) and breech strike between weaner and hogget shearing. This indicates that breech strike is a repeatable trait and that animals that are struck early in life should be culled as they will have a higher propensity to be struck later in life.
None of the standard wool production and wool quality traits was phenotypically correlated with breech strike. The highest phenotypic correlations between breech strike and any other potential indicator trait was found for dags scored before hogget shearing (rp = 0.45), dags scored in spring (rp = 0.34) and for breech wrinkle scored at weaning (rp = 0.31). The correlations with the remaining dag traits varied between 0.2 and 0.3, while the correlations with the wrinkle and breech cover traits were equal to or below 0.20. All the other phenotypic correlations with any of the remaining traits were very low. This confirms farmers’ observations that dags and wrinkle are important traits for breech strike susceptibility. However, it does not support the notion that breech cover is an important trait in unmulesed sheep in this environment. This may be due to the relative small size of the bare area in Merino sheep and the fact that a larger area is required in regions where dags are a problem before it will show any effect.
Genetic correlations between breech strike and indicator traits
Breech strike early in life (before weaner shearing) was genetically not the same trait as breech strike from weaner shearing up to hogget shearing. The genetic correlation was only 0.44 ± 0.17, which is significantly different from unity. However, this positive genetic correlation indicates that culling on early breech strike susceptibility should result in a correlated response later in life. Although breeders protect their sheep from being struck, a small proportion of animals can become struck under favourable conditions. Therefore, sheep breeders should be advised that struck sheep are probably the most susceptible sheep in the flock and therefore should not be kept for breeding purposes as this would increase the susceptibility of the flock.
The present study has identified several traits that are genetically significantly correlated with breech strike, and that could qualify as potential indicator traits that could be used to select indirectly for breech strike resistance on the genetic level.
FDCV was the only wool trait that was correlated with breech strike (–0.27 ± 0.13). This indicates that susceptibility or resistance to breech strike is not genetically associated with fleece weight. This is an important finding that should give sheep breeders confidence that breeding for resistance to breech strike will not lead to a reduction in wool production.
Dag score was the single most important indicator trait for breech strike in unmulesed and uncrutched sheep that did not receive any chemical preventative treatment against breech strike. It was genetically strongly correlated with breech strike, with a genetic correlation generally higher than 0.5, except for yearling dags. Dags at marking, post-weaning, spring and hogget age had a heritability of ~0.35, which resulted in dags scored pre-hogget shearing in spring to be the most effective indicator trait for breech strike. This is also the time when the flies are present in this environment. Selecting on this trait will result in ~60% of the gain that can be achieved by selecting directly for breech strike.
Interestingly, dags scored at yearling age had a much higher heritability estimate (0.63 ± 0.08) and a much lower genetic correlation (0.34 ± 0.13) with breech strike than did dags scored at other ages. The low correlation could be due to the fact that flies are generally not found at this time of the year in this environment. This measurement also coincided with the onset of winter in this environment, which could be influenced by a hypersensitivity response of certain sires’ progeny to pasture larvae after a long dry summer (Larsen et al. 1994), rather than dags caused by high worm burdens. This may indicate that, in this environment, dags scored at the onset of winter at yearling age could be a genetically different trait from dags scored at other ages. The development of dags at various ages thus warrants further investigation.
Urine stain scored at weaning was the second most important trait in unmulesed and uncrutched sheep. It was a highly heritable trait at weaning; however, this estimate should be treated with caution because of the relatively low proportion of animals affected by urine stain at this time; more research is needed on this relationship. This also coincided with the time when the flies are present in this environment. Urine stain before hogget shearing could not be recorded because the presence of dags made it very difficult or even impossible to score urine stain with confidence. Although the results suggested that selecting on this trait will result in a response of ~60% of that which can be achieved by selecting directly for breech strike, it is important to note that only ~26% of ewe lambs showed any sign of urine stain, which resulted in an average urine score of 1.15 at weaning. The incidence of urine stain was less than 20% at marking and at hogget age, and only 9% at yearling age. The confounding nature of dags and the low incidence of urine stain indicated that it will be difficult to identify genetically superior sheep for this trait.
Previous research (Seddon et al. 1931) has shown that skin wrinkle is strongly associated with breech strike, which was recently confirmed by Smith et al. (2009) in a fine wool flock. It is also a highly heritable trait (Turner and Young 1969; Greeff and Karlsson 2009; Smith et al. 2009; Brown et al. 2010). In the present study, all wrinkle traits were heritable, but contrary to expectation, wrinkle scored on the neck at marking (rg = 0.38 ± 0.13) and at post-hogget shearing (rg = 0.46 ± 0.13) had the genetically strongest correlation with breech strike. The only stage when breech wrinkle was genetically significantly correlated with breech strike was when it was scored at a yearling age (rg = 0.27 ± 0.10).
This flock was a relatively plain-bodied flock (Table 3). Anecdotal information indicates that the Western Australian sheep flock is plainer than the Merino flocks from eastern states. Therefore, highly wrinkly animals were not sourced when the flock was established, because it is well known that wrinkly animals are generally more susceptible to breech strike (Seddon et al. 1931). The focus of the present study was to source representative animals from participating flocks.
It was sometimes difficult to differentiate among animals using body, tail or breech wrinkle, especially on sheep with long wool. The highest wrinkle scores were recorded on the neck of the animals. Scholtz et al. (2010) showed that neck wrinkle is genetically strongly correlated with breech wrinkle in mulesed (rg = 0.80) and unmulesed (rg = 0.89) sheep. They also found a stronger genetic relationship between the absence of breech strike and neck wrinkle (rg = –0.57) in unmulesed sheep than that between the absence of breech strike and breech wrinkle (rg = –0.45), which strongly agrees with the results of the present study. This confirms that neck wrinkle is a good indirect indicator trait for breech strike in plainer-bodied sheep and in mulesed flocks. But it is unclear as to why neck wrinkle is a better indicator trait than breech or tail wrinkle in an uncrutched and unmulesed flock, as it is generally assumed that the best wrinkle traits would be found at the site with the highest risk of being struck. A plausible explanation could be that it is difficult to assess tail wrinkle at weaning on unshorn weaners because it is not possible to see the wrinkles under the wool on the tail at this stage. It is also unclear as to why the heritability of rump wrinkle after the animals have been shorn should be lower than that at other ages. This may be related to the changes in skin wrinkle after shearing, and also perhaps to weight loss that weaners normally experience in this environment following weaning and shearing. However, heritability of tail wrinkle at post-hogget shearing was moderate (0.38 ± 0.11), which indicates that it should respond to selection. Under a management system were animals are crutched to prevent formation of dags and urine stain, breech wrinkle may still be an important indicator trait, especially in more wrinkled flocks. The role of breech wrinkle in breech strike requires further investigation in Merino flocks that are not subject to dags and urine stain.
McGuirk and Atkins (1980) found that greasy wool colour scored on the midside was positively correlated with fleece rot, while Raadsma and Rogan (1987) showed that fleece rot was a key indicator trait for body strike. In the present study, greasy, midside wool colour, although moderately heritable, was not genetically correlated with breech strike. There was a tendency for sheep with higher average greasy wool colour (yellowness) to have a higher incidence of breech strike. This trend was, however, not significant. Raadsma and Rogan (1987) showed that greasy wool colour is positively correlated with body strike. The fact that no genetic relationship was found in the present study may be due to the overriding effect that dags and urine stain had on the incidence of breech strike in this environment, thereby perhaps masking or overshadowing the impact of wool colour. Thus, although greasy wool colour is a heritable trait, it was not genetically or phenotypically associated with breech strike and, therefore, does not seem to offer potential as an indicator trait for breech strike for the Mediterranean regions.
Scobie et al. (2008) showed that larger bare breech skin area was significantly correlated with a lower dag score in non-Merino crosses. The present study found that breech cover is a heritable trait at different ages, which supports the results of Edwards et al. (2009). Although breech cover before post-weaning shearing had the highest genetic correlation with breech strike (rg = 0.32 ± 0.15), it did not feature highly in the list of indicator traits on the basis of the CR : D ratio, in spite of having a heritability estimate of 0.38 ± 0.08. Edwards et al. (2009) found that breech cover was not highly repeatable in females (r = 0.42), which indicates that scoring breech cover at different ages could be problematic in ewes. It would imply that other environmental factors could affect the expression of this trait. It is important to note that in this environment, mulesed Merino sheep with a large artificially bare area, can still develop dags and must still be crutched before winter to protect them against breech strike. This may indicate that a much larger bare breech is required before sheep are protected against breech strikes where dags are a problem.
Face cover at weaning (rg = 0.28 ± 0.10) and yearling age (rg = 0.27 ± 0.10) had a lower genetic correlation with breech strike than did breech cover, but both traits were highly heritable traits, with heritability estimates of 0.77 ± 0.13 and 0.73 ± 0.13, respectively. This resulted in the CR : D ratio for face cover scored at weaning and at yearling to be equal to 0.44 and 0.39, respectively, which indicates that they are better indicator traits than is breech cover. However, the heritability of face cover dropped to 0.31 ± 0.06 at post-hogget shearing, which indicates that shearing may reduce the accuracy of scoring face cover, or that other environmental factors affect the expression of the trait. Edwards et al. (2009) found that breech cover had a low repeatability; thus, if face cover and breech cover are correlated, then a similar trend may be found for face cover. These results strongly suggest that face cover should be scored before shearing or wigging.
Faecal WEC in spring and faecal consistency score at weaning were genetically moderately correlated with breech strike (rg = 0.27 and 0.36, respectively), but their CR : D ratios were not high enough for them to qualify as potential indicator traits for breech strike resistance. However, as these traits are caused by the presence of worms, they contributed to the formation of dags, which has been shown to be the most important indicator trait for breech strike in this environment.
Greeff et al. (2010) showed that the indicator traits investigated in the present study explain only up to 20% of the total variation in breech strike. The fact that dags and urine stain are the best indicator traits for breech strike susceptibility again shows that moisture is a critical component for the development of breech strike. As breech wrinkle and breech cover were not consistently correlated with breech strike, it appears that there may be an interaction with the moisture content, and that other factors may be involved. A phenotypic comparison of extreme individuals for breech strike in the present study (Greeff et al. 2010) has shown no clear phenotypic differences in indicator traits between extremely susceptible and resistant sire progeny groups. This indicates that other factors are probably involved in making animals more or less susceptibility to breech strike. If breech wrinkle and breech cover traits were the primary traits, then one would expect them to explain more of the variation in breech strike; however, it appears that differences in these traits have only the effect of modifying the expression and severity of breech strike.
The present study also found that large differences exist between the heritability estimates for the same indicator trait scored at different ages. This variability could have been caused by the varying environmental conditions during the year. The long, dry, hot summers of the Mediterranean regions are generally followed by cold, wet winters with lush pastures in late winter and spring. These conditions will certainly affect both flies and sheep, as dags are most prevalent during late winter and spring when flies are more likely to be present and active. The present study has indicated that the heritability estimates are not constant. This requires further investigation to determine how different environment conditions affect the same indicator traits at different times of the year, to determine when would be the best time to score animals for these traits. The best time would be when the heritability estimate is the highest, provided it is genetically the same trait at different times.
Conclusions
The aim of the present study was to identify indicator traits that could be used to breed indirectly for resistance to breech strike, as it would be unpractical and unacceptable for industry to challenge their breeding animals with flies to select directly for breech strike resistance. The results showed that breech strike is a heritable trait and that early breech strike is phenotypically and genetically moderately correlated with breech strike later in life. Dags are genetically strongly correlated with breech strike and are the most important indicator trait for breech strike in this environment. Dags and urine stain scored at the time when flies are present, neck wrinkle at marking, and breech and face cover at weaning could be used as indirect indicator traits for breech strike resistance in a winter-rainfall environment such as in Western Australia. No significant genetic associations were found between breech strike and the wool production and wool quality traits. Only FDCV had a small unfavourable association with breech strike.
Sheep Genetics currently estimate breeding values for dags, wrinkle and breech cover that allow breeders to select indirectly for breech strike resistance. The present study has shown that a trait for urine stain should be added. Face cover and neck wrinkle also qualify as potential indicator traits, especially in mulesed flocks and in flocks that are already relatively plain, to breed indirectly for breech strike resistance.
Acknowledgements
We acknowledge our technical farm staff, especially Nicola Stanwyck and Noreen Underwood who managed the flock and who were responsible for the collection of data for this study. We also thank Australian Wool Innovations Ltd for funding and Mr Geoff Lindon of AWI for his continued interest and constructive comments.
References
AWI (2007) ‘Visual sheep scores.’ (Australian Wool Innovation and Meat and Livestock Australia: Sydney)Belschner HG (1953) ‘Sheep management and diseases.’ (Angus and Robertson: Sydney)
Brown DJ, Swan AA, Gill JS (2010) Within- and across-flock genetic relationships for breech flystrike indicator traits. Animal Production Science 50, 1060–1068.
| Within- and across-flock genetic relationships for breech flystrike indicator traits.Crossref | GoogleScholarGoogle Scholar |
Cloete SWP, Misztal I, Olivier JJ (2009) Genetic parameters and trends for lamb survival and birth weight in a Merino flock divergently selected for multiple rearing ability. Journal of Animal Science 87, 2196–2208.
| Genetic parameters and trends for lamb survival and birth weight in a Merino flock divergently selected for multiple rearing ability.Crossref | GoogleScholarGoogle Scholar |
Donoghue KA, Rekaya R, Bertrand JK, Misztal I (2004) Threshold-linear analysis of measures of fertility in artificial inseminations data and days to calving in beef cattle. Journal of Animal Science 82, 987–993.
Dun RB, Eastoe R (1970) ‘Science and the Merino breeder.’ New South Wales Department of Agriculture. (Government printer: Sydney)
Edwards NM, Hebart M, Hynd PI (2009) Phenotypic and genotypic analysis of bare breech trait in Merino sheep as a potential replacement for surgical mulesing. Animal Production Science 49, 56–64.
| Phenotypic and genotypic analysis of bare breech trait in Merino sheep as a potential replacement for surgical mulesing.Crossref | GoogleScholarGoogle Scholar |
Falconer DS (1989) ‘Introduction to Quantitative genetics.’ (Longman Scientific Technical: New York)
Genstat (2011) ‘Users manual. Version14.1.’ (VSN International: Hemel Hempstead, UK)
Gherardi SG, Sutherland SS, Monzu N, Johnson KG (1983) Field observations on body strike in sheep affected with dermatophilosis and fleece-rot. Australian Veterinary Journal 60, 27–28.
| Field observations on body strike in sheep affected with dermatophilosis and fleece-rot.Crossref | GoogleScholarGoogle Scholar | 6830546PubMed |
Gilmour AR, Gogel BJ, Cullis BR, Thompson R (2006) ‘ASReml user guide. Release 2.0.’ (VSN International: Hemel Hempstead, UK)
Greeff JC, Cox G (2006) Genetic changes generated within the Katanning Merino resource flocks. Australian Journal of Experimental Agriculture 46, 803–808.
| Genetic changes generated within the Katanning Merino resource flocks.Crossref | GoogleScholarGoogle Scholar |
Greeff JC, Karlsson LJE (2009) Opportunities to breed for resistance to breech strike in Merino sheep in a Mediterranean environment. In ‘Proceedings of the 18th conference of the Association for the Advancement of Animal Breeding and Genetics’. pp. 272–278.
Greeff JC, Karlsson LJE, Schlink AC (2010) Final report to Australia Wool Innovations Ltd on Breeding for breech strike resistance. Australian Wool Innovations Ltd final report of Project EC940, Sydney.
Huisman AE, Brown DJ, Ball AJ, Graser H-U (2008) Genetic parameters for body weight, wool, and disease resistance and reproduction traits in Merino sheep. 1. Description of traits, model comparison, variance components and their ratios. Australian Journal of Experimental Agriculture 48, 1177–1185.
| Genetic parameters for body weight, wool, and disease resistance and reproduction traits in Merino sheep. 1. Description of traits, model comparison, variance components and their ratios.Crossref | GoogleScholarGoogle Scholar |
James PJ (2006) Genetic alternatives to mulesing and tail docking in sheep: a review. Australian Journal of Experimental Agriculture 46, 1–18.
| Genetic alternatives to mulesing and tail docking in sheep: a review.Crossref | GoogleScholarGoogle Scholar |
Karlsson LJE, Greeff JC (2006) Selection response in fecal worm egg counts in the Rylington Merino parasite resistant flock. Australian Journal of Experimental Agriculture 46, 809–811.
| Selection response in fecal worm egg counts in the Rylington Merino parasite resistant flock.Crossref | GoogleScholarGoogle Scholar |
Karlsson LJE, Pollott GE, Eady SJ, Bell A, Greeff JC (2004) Relationship between faecal worm egg counts and scouring in Australian Merino sheep. Animal Production in Australia 25, 100–103.
Larsen JWA, Anderson N, Vizard AL, Anderson GA, Hoste H (1994) Diarrhoea in Merino ewes during winter: association with trichostrongylid larvae. Australian Veterinary Journal 71, 365–372.
| Diarrhoea in Merino ewes during winter: association with trichostrongylid larvae.Crossref | GoogleScholarGoogle Scholar |
Lee D, Misztal I, Bertrand K, Rekaya R (2002) National evaluation of calving ease, gestation length and birth weight by linear and threshold model methodologies. Journal of Applied Genetics 43, 209–216.
Lewer RP, Gherardi SG, Sutherland SS (1987) Realised heritability estimates for resistance to dermatophilosis in Merino sheep. In ‘Merino improvement programs in Australia. Proceedings of a national symposium, Leura, New South Wales’. (Ed. BJ McGuirk) pp. 347–350. (Australian Wool Corporation: Melbourne)
McGuirk B, Atkins KD (1980) Indirect selection for increased resistance to fleece rot and body strike. In ‘Proceedings of the 13th Australian Society for Animal Production’. pp. 92–95.
Misztal I, Tsuruta S, Strabel T, Auvray B, Druet T, Lee DH (2002) BLUPF90 and related programs (BGF90). In ‘Proceedings of the 7th world congress on genetics applied to livestock production’. pp. 743–744. (Montpellier, France)
Ntzoufras I (2009) ‘Bayesian modeling using Winbugs.’ (John Wiley and Sons: New York)
Raadsma HW (2000) Genetic aspects of resistance to ovine cutaneous myiasis. In ‘Breeding for disease resistance in farm animals’. (Eds RFE Axford, SC Bishop FW Nicholas, JB Owen) pp. 171–193. (CAB International: Wallingford, UK)
Raadsma HW, Rogan IM (1987) Genetic variation in resistance to blowfly strike. In ‘Merino improvement programs in Australia. Proceedings of a national symposium, Leura, New South Wales’. (Ed. BJ McGuirk) pp. 321–340. (Australian Wool Corporation: Melbourne)
Scholtz AJ, Cloete SWP, van Wyk JB, Misztal I, du Toit E, van der Linde TC de K (2010) Genetic covariances between wrinkle score and absence of breech strike in mulesed and unmulesed Merino sheep, using a threshold model. Animal Production Science 50, 201–218.
| Genetic covariances between wrinkle score and absence of breech strike in mulesed and unmulesed Merino sheep, using a threshold model.Crossref | GoogleScholarGoogle Scholar |
Scobie DR, O’Connell DO, Bray A, Cunningham P (2002) Breech strike can be reduced by increased area of naturally bare skin around the perineum of lambs. In ‘Proceedings of the 24th Animal Production Society of Australia’. pp. 201–204.
Scobie DR, O’Connell DO, Morris CA, Hickey SM (2007) A preliminary genetic analysis of breech and tail traits with the aim of improving the welfare of sheep. Australian Journal of Experimental Agriculture 58, 161–167.
Scobie DR, O’Connell DO, Morris CA, Hickey SM (2008) Dag score is negatively correlated with breech bareness score in sheep. Australian Journal of Experimental Agriculture 48, 999–1003.
| Dag score is negatively correlated with breech bareness score in sheep.Crossref | GoogleScholarGoogle Scholar |
Seddon HR, Belschner HG, Mulhearn CR (1931) Studies on cutaneous myiasis of sheep (sheep blowfly attack). Science Bulletin, No. 37. New South Wales Department of Agriculture.
Smith J, Brewer H, Dyall T (2009) Heritability and phentopyic correlations for breech strike and breech strike resistance indicators in Merino. In ‘Proceedings of the 18th conference of the Association for the Advancement of Animal Breeding and Genetics’. pp. 334–337.
Turner HN, Young SSY (1969) ‘Quantitative genetics in sheep breeding.’ (MacMillan of Sydney)
Watts JE, Merritt GC (1981) Body strike in sheep. In ‘Proceedings no. 58. Refresher course for veterinarians, 10–14 August 1981, the Stephen Roberts Lecture Theatre, The University of Sydney’. pp. 171–193.
Williams AR, Karlsson LJE, Palmer DG, Vercoe PE, Williams IH, Greeff JC, Emery DL (2010) Relationships between faecal dry matter, worm burdens and inflammatory mediators and cells in parasite-resistant Merino rams. Veterinary Parasitology 171, 263–272.
| Relationships between faecal dry matter, worm burdens and inflammatory mediators and cells in parasite-resistant Merino rams.Crossref | GoogleScholarGoogle Scholar | 20430531PubMed |