

Purification and characterisation of a phosphatidylcholine-binding protein from duck Biceps femoris muscle
D. Y. Wang A , M. H. Zhang A , F. Liu A , Y. Z. Zhu A and W. M. Xu A BA Institute of Agricultural Products Processing, Jiangsu Academy of Agricultural Sciences, Nanjing 210014, People’s Republic of China.
B Corresponding author. Email: weiminxu2002@yahoo.com.cn
Animal Production Science 54(2) 194-199 https://doi.org/10.1071/AN12321
Submitted: 22 May 2012 Accepted: 28 February 2013 Published: 9 April 2013
Abstract
The interaction between protein and phospholipids is a widespread phenomenon involving several physiological events in postmortem muscle. We hereby report a method for one-step purification of a phosphatidylcholine-binding (PC-binding) protein from duck Biceps femoris muscle with relatively high purity and yield using ion-exchange chromatography. This PC-binding protein has an inhibitory effect on the activity of phospholipase A2 (PLA2). A decrease (~62.3%) in PLA2 activity was observed. It had a strong affinity to bind PC at pH range of 6.2–6.8 with a peak at pH 6.6 (13.36 ± 0.48 g PC/g protein); in addition, raising ATP content from 1 to 5 μmol/mL enhanced the binding capacity. The PC-binding protein plays a potential role in the integrity of membrane and meat quality.
Additional keywords: characterisation, phospholipase A2, purification.
Introduction
Phospholipids (PL) are a class of lipids that are a major component of intramuscular fat. Phospholipids make up 50% of the total fat and 1% of the wet weight and are important to the quality of meat products (Gray et al. 1996). Phospholipids are very sensitive to hydrolysis or oxidation due to their high polyunsaturated fatty acids content (Hernández et al. 1999). Previous studies have demonstrated that PL were the main substrates of lipolysis in poultry and pig muscles postmortem (Sklan et al. 1983; Buscailhon et al. 1994). In cooked and dry-cured meat products, lipolysis of PL contributes to the generation of free fatty acids, which were considered main precursors to the flavour compounds (Chizzolini et al. 1998; Wang et al. 2009a). On the other hand, as the PL bilayer forms the cell membrane, the degradation of membrane PL would cause the water loss from meat products. As previously reported, the majority of water in muscle is held within the myofibrils, between the myofibrils and between the myofibrils and membrane, between muscle cells, and between bundles (Offer and Cousins 1992).
Phospholipid hydrolysis, as the first stage and main cause of phopholipid degradation, is generally attributed to phospholipases. Variables affecting activity of phospholipases in postmortem muscle, however, are poorly studied. Some identified membrane proteins (such as calpactins, lipocortins and Hsp70) are able to inhibit the activity of phospholipase A2 (PLA2) and maintain the integrity, fluidity and permeability of membrane. Nevertheless, their function on meat quality is unknown (Davidson et al. 1986; Jaattela 1993).
Since phosphatidylcholine (PC) is very abundant in animal tissues (almost 50% of total PL; Wang et al. 2009b), the objective of this study was to investigate the possible relationship between a PC-binding protein and PC lipolysis. In recent years, duck meat has gained significant popularity in Asia but very few studies were conducted on duck meat quality. For this reason we developed a simple and efficient method to purify this protein from duck Biceps femoris muscle and evaluated the effects of PC-binding protein on the activity of PLA2. As pH and ATP are two major changes during the transition to postmortem meat, we subsequently analysed their effects on the binding capacity of protein to PC, which may reflect the postmortem status of PC in duck muscle.
Materials and methods
Materials
Gaoyou ducks (fed for 10 months) from Jiangsu Waterfowl Research and Development Centre were slaughtered following commercial practices in a poultry processing plant (Jiangsu Furun Food Ltd, China). After being slaughtered, six ducks were packed separately and transported in ice to the laboratory immediately. Phospholipid standards, including PC (2-oleoyl-1-palmitoyl-sn-glycero-3-phosphocholine) and lysophosphatidylcholine (LPC, 1-palmitoyl-sn-glycero-3-phosphocholine) were purchased from Sigma-Aldrich Chemical Co. (St Louis, MO, USA); PLA2 was from Cayman Chemical Co. (USA); methanol, chloroform and acetonitrile were chromatographic pure grade; ATPNa2, CaCl2, Tris, HCl, NaCl, polyacrylamide, 2-mercaptoethanol, formic acid, NaAc, HAc, NaTDC, Na2HPO4, NaOH were analytic pure grade.
Preparation of crude protein
Biceps femoris muscles were removed from duck carcasses and trimmed of all visible subcutaneous fat and connective tissues. Thirty grams of muscle sample was homogenised immediately with 100 mL TRIS-HCl buffer (100 mM, pH 8.0) at 1200 rpm on ice using an Ultra Turrax (T25, IKA, Germany). The homogenate was then centrifuged for 20 min at 12 000g at 4°C (Allegra 64R, Beckman, USA), supernatant was collected and filtered through four-layer gauze. Afterwards, the filtrate was precipitated by ammonium sulfate at a saturation of 70% at 4°C for 12 h under gentle stirring, and centrifuged for 20 min at 12 000g at 4°C, the precipitate was dissolved in a minimal volume of TRIS-HCl buffer (100 mM, pH 8.0). The crude extract was dialysed for 24 h in TRIS-HCl buffer (100 mM, pH 8.0), then lyophilised and stored at −20°C.
Determination of binding capacity of extracted protein to PC
The PC microemulsions were prepared by dissolving 1.0 g PC in 5.0 mL ethyl ether, followed by mixing with 20 mL buffer or double distilled water (DDW). Buffer solutions of various pH values as indicated were prepared with 50 mM Na-HAc buffer (pH 5.2, 5.5 and 5.9), 50 mM sodium phosphate buffer (pH 6.2, 6.6 and 6.8) or 50 mM TRIS-HCl buffer (pH 7.2). The ethyl ether was removed by rotatory evaporator (RE-85C, Yarong, China) in a 44°C water bath. Then these solutions were dissolved in 20 mL buffer or DDW again and were emulsified (2 × 2 min) at a frequency of 80% with a 130-Watt autotune series high intensity ultrasonic sonicator. Then 0.55 g CaCl2, 2.07 g NaTDC and DDW or buffer was supplemented to make a constant volume of 100 mL, thus 10 mg/mL PC suspension was obtained. To determine binding capacity of extracted protein to PC, 40 µL protein (1.662 mg/mL) was incubated for 1.0 min at 15°C in the suspension of 200 µL PC. After incubation, 400 µL chloroform was added to extract PC, and centrifuged at 4000g at 4°C for 15 min. The substratum containing PC was analysed by high performance liquid chromatography (HPLC). Binding capacity of protein to PC was calculated by dividing the amount of PC decrease in the reaction mixture by quantity of protein.
Purification of PC-binding protein
Crude protein (10.0 mg) was dissolved in 1.0 mL TRIS-HCl buffer (100 mM, pH 8.0) and centrifuged at 12 000g at 4°C for 10 min, then filtered through a 0.45-μm membrane. The filtrate was applied to ion-exchange chromatography with source 15Q column (1.0 × 5.0 cm), which has been previously equilibrated with TRIS-HCl buffer (50 mM, pH 8.0) and eluted with the same buffer, until unbound protein passed through the column. The filtrate was eluted with a linear gradient of 0–1.0 M NaCl in equilibration buffer. A flow rate of 1.0 mL/min was used in the whole procedure and it was monitored spectrophotometrically by absorbance at 280 nm. The fraction containing the protein that had the optimal binding capacity to PC were pooled and concentrated by ultrafiltration using a 10-kDa cut-off membrane.
SDS-PAGE of PC-binding protein
Sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) was performed according to Sarkar (Sarkar et al. 2009) in 10% SDS-polyacrylamide mini gels (5% stacking, 1 mm thick) at a constant voltage of 140 V. Protein samples and molecular weight standards were heated to 95°C for 4 min in the presence of 5% 2-mercaptoethanol before electrophoresis. Protein bands were stained with Coomassie Blue R-250. The protein markers were resolved into clearly identifiable sharp bands from 10 to 200 kDa.
Identification and molecular mass analysis of PC-binding protein
Protein sample obtained by SDS-PAGE was identified using the Nano LC-ESI-MS/MS peptide sequencing technology. The mass spectrometric data acquired was used to search for the most recent non-redundant protein database with ProtTech’s proprietary software suite.
Effect of pH and ATP on the binding capacity of extracted protein to PC
The PC suspension was prepared at various pH values ranging from 5.2 to 7.2. To evaluate the effect of pH on the binding capacity of extracted protein to PC, 40 µL protein was added to 200 µL PC suspension with different pH values. At optimal pH, the effect of ATP on the binding capacity of extracted protein to PC was evaluated by mixing 40 µL protein with ATP at concentrations from 1 to 5 μmol/mL, respectively. Binding capacity of protein to PC was measured with HPLC.
Effect of PC-binding protein on the activation of PLA2
The reaction mixture of PLA2 untreated with extracted protein consisted of 200 µL PC suspension, 40 µL of TRIS-HCl buffer (50 mM, pH 8.0) and 10 µL PLA2, whereas the protein-treated reaction mixture consisted of 200 µL PC suspension, 40 µL protein and 10 µL PLA2. After a 30-min incubation at 37°C, the reaction products were analysed by HPLC.
Determination of PC-binding protein concentration
All protein quantifications were performed by Bradford method and monitored spectrophotometrically at 595 nm with bovine serum albumin as the standard.
HPLC analyses of PC and LPC
The samples were analysed in an Agilent 1100 HPLC system (Agilent, USA) using a Sinochrom SI 60 silica gel column (5 µm, 250 × 4.6 mm) operating at 30°C according to our previous methods (Wang et al. 2009b). Invariable elution methanol was used at a flow rate of 1.0 mL/min. Chromatographic peaks were detected with an ELSD (evaporative light scattering detector), running at 85°C with N2 at 2.5 L/min. PC and LPC peaks were identified by comparing their retention time and UV spectra with those of PC and LPC standards.
Statistical analyses
Results of the binding capacity of protein to PC were presented as mean ± s.d., which were from six duck samples and each sample was repeated 3 times. Significant difference was determined with Student’s t-test (Microsoft Office Excel 7.0). The significance level was settled at P < 0.05.
Results
Development of a new method for the purification of PC-binding protein from Biceps femoris muscle in duck
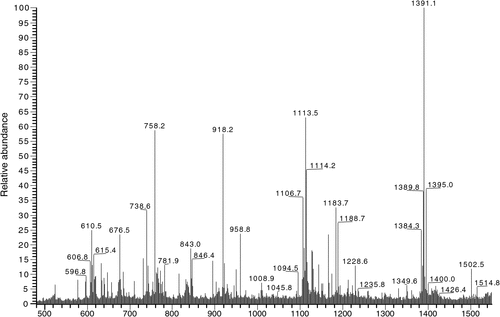
PC-binding protein was purified from crude protein through a one-step purification procedure, which yields roughly 1 mg PC-binding protein from 3 g muscle. During the ion-exchange chromatography, the column bound proteins were separated into several peaks. These fractions were analysed by binding capacity to PC. Spectrophotometric analysis revealed that PC-binding protein was collected predominantly in one fraction (Fig. 1a). Eluted fraction was applied to SDS-PAGE analysis. The analysis of the pooled fraction revealed the presence of a highly homogeneous, ~90-kDa-band protein (Fig. 1b). Identified by HPLC-ESI-MS/MS (Fig. 2) and BLAST in NCBInr database, it exhibited a high degree of homology with sequences of other biology ones namely heat shock protein 90-α (Hsp 90α) with molecular mass of 84 405.73.
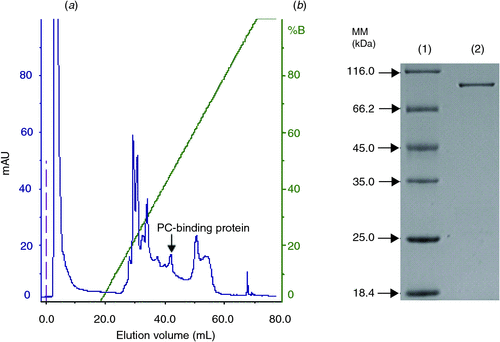
|
|
Binding capacity of protein to PC analyses
The PC suspension was prepared as 10 mg/mL and content of PC was measured to be relatively high by HPLC (Fig. 3a), whereas after PC-binding protein treatment, the PC content significantly declined (P < 0.05) (Fig. 3b), which indicated that the protein could bind to PL.
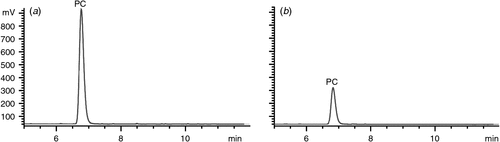
|
Effect of pH and ATP on the binding capacity of PC-binding protein to PC
The protein showed strong binding capacity at a pH range of 6.2–6.8, the maximum amount (13.36 ± 0.48 g PC/g protein) was achieved at pH 6.6 (Fig. 4a). When pH values were below 5.5 binding capacity decreased substantially. Each gram of protein could bind 23.94 ± 1.15 g PC at ATP level of 5 μmol/mL (Fig. 4b), which was almost double than that at 1 μmol/mL ATP level.
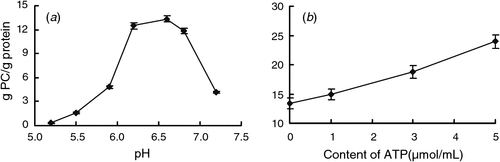
|
Effect of PC-binding protein on the activation of PLA2
PLA2 can lead to hydrolysis of the sn-2 position of PC, resulting in the production of free fatty acids and LPC. The reaction between PLA2 and PC in absence of PC-binding protein converts some amount of PC to LPC (Fig. 5a). After PC-binding protein treatment, an obvious rise in PC level and decline in LPC level were detected (Fig. 5b). The activity of PLA2 was reduced by 62.3%. This indicates that this protein binds PC more strongly than PLA2. As a result, PLA2 is now less effective in catalysing lipolysis in the sn-2 position.
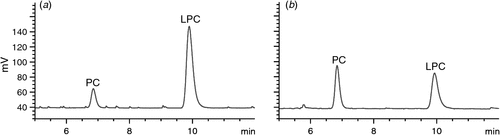
|
Discussion
2-oleoyl-1-palmitoyl-sn-glycero-3-phosphocholine is a major molecular species of PC in Gaoyou duck (Wang et al. 2009b), the protein that binds to it shows the status of PL and physiological reactions occurring in muscle. In our study, the PC-binding protein had strong binding capacity at pH range of 6.2–6.8 with peak at pH of 6.6. The binding capacity dropped as ATP decreased, which implied that binding capacity declined during the conversion from muscle to meat. The decrease of the PC-binding protein may release more unprotected PC, which is likely to undergo hydrolysis and oxidation processes. This may affect the ability of membrane to act as a semipermeable barrier and may contribute to exudative loss from fresh meat. In many studies, pH was one of the most important parameters to assess the meat quality, and water holding capacity was believed to decrease during the post-mortem period. It was found that the degradation of membrane proteins such as integrin has been associated with larger drip channel size and increased drip loss (Huff-Lonergan and Lonergan 2005). Bee et al. (2007) found early pH decline contributed to the degradation of desmin and talin and shrinkage of the muscle fibre. The present study may serve as another approach to explain the decreased water holding capacity during post-mortem in these studies.
It was found that PLA2 catalyses the hydrolysis and oxidation of membrane PL, resulting in the loss of cell integrity, a main factor contributing to drip formation (Lambert et al. 2001). Some researchers have indicated that PLA2 plays a fundamental role in the occurrence of PSE in pork and chicken meat (Lambert et al. 2001; Chen et al. 2010). The decline in PC hydrolysis after the PC-binding protein treatment suggests the protein could protect PC or partly inhibit the activity of PLA2. The PC-binding protein identified was Hsp 90, which is abundantly expressed in nearly all organisms, making up 1–2% of cytosolic proteins (Buchner 1996). It plays an essential role in protecting the cells and as ‘molecular chaperone’. Small Hsp and Hsp 70 have been reported to bind with PL and maintain the integrity and physical state of cell membranes under stress conditions (Tsvetkova et al. 2002; Chowdary et al. 2007; Harada et al. 2007). Hsp70 has also been shown to interfere with the signal transduction pathway to PLA2 activation (Jaattela 1993). Yu et al. (2009) found increases in plasma creatine kinase and lactate dehydrogenase, which indicate pig muscle damage and disruptions in cell membrane is associated with decrease in Hsp expression and higher drip loss after transportation stress. Similar results were obtained by Di Luca et al. (2011) who found a correlation between decreased Hsp content and a decline in water-holding capacity. However, the underlying mechanisms and processes of the Hsp and water-holding capacity changes were not elucidated in these studies. On the basis of these results, it can be inferred that the existence of PC-binding protein may protect PC against PLA2 hydrolysis and improve the water holding capacity, and therefore meat quality, by maintaining the membrane integrity.
Conclusions
We developed a simple and efficient method to purify a PC-binding protein from duck muscle, and demonstrated that PC-binding protein could restrain the activity of phospholipase on PC lipolysis. The binding capacity is largely influenced by pH and ATP. The results reflect the postmortem interactions of membrane protein and PL, and implied the potential function of this protein on PC lipolysis. Further studies are required to examine the PC-binding capacity of protein in vivo from postmortem meat and mechanism of its function.
Acknowledgements
This study was funded by the National Natural Science Foundation of China (31071569), Natural Science Foundation Program of Jiangsu Province (BK2012785) and Innovation of Agricultural Science and Technology of Jiangsu Province [CX(11)4028].
References
Bee G, Anderson AL, Lonergan SM, Huff-Lonergan E (2007) Rate and extent of pH decline affect proteolysis of cytoskeletetal proteins and water-holding capacity in pork. Meat Science 76, 359–365.| Rate and extent of pH decline affect proteolysis of cytoskeletetal proteins and water-holding capacity in pork.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2sXit12ltro%3D&md5=1ce4fa003a7f67dbf2794734ef944150CAS | 22064307PubMed |
Buchner J (1996) Supervising the fold: functional principles of molecular chaperones. The FASEB Journal 10, 10–19.
Buscailhon S, Gandemer G, Monin G (1994) Time-related changes in intramuscular lipids of French dry-cured ham. Meat Science 37, 245–255.
| Time-related changes in intramuscular lipids of French dry-cured ham.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaK2cXkvVWgtrg%3D&md5=1b9174bfc51bd15a1c6eee5ea2fb0ef2CAS | 22059498PubMed |
Chen T, Zhou GH, Xu XL (2010) Phospholipase A2 and antioxidant enzyme activities in normal and PSE pork. Meat Science 84, 143–146.
| Phospholipase A2 and antioxidant enzyme activities in normal and PSE pork.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD1MXht1GmsbfF&md5=fd64a03792f45e464348004d56927ee2CAS | 20374766PubMed |
Chizzolini R, Novwlli E, Zanardi IE (1998) Oxidation in traditional Mediterranean meat products. Meat Science 49, S87–S99.
| Oxidation in traditional Mediterranean meat products.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaK1cXmtFGltb8%3D&md5=ad87f6cda118c73bb21c57cbd67a3009CAS |
Chowdary TK, Raman B, Ramakrishna T, Rao CM (2007) Interaction of mammalian Hsp22 with lipid membranes. Biochemical Journal 401, 437–445.
| Interaction of mammalian Hsp22 with lipid membranes.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2sXhtVaiuw%3D%3D&md5=7c92bd83e5a132a20314eece43c3c5dfCAS | 17020537PubMed |
Davidson FF, Dennis EA, Powell M, Glenny JR (1986) Inhibition of phospholipase A2 by ‘lipocortins’ and calpactins. The Journal of Biological Chemistry 262, 1698–1705.
Di Luca A, Mullen AM, Elia G, Davey G, Hamill RM (2011) Centrifugal drip is an accessible source for protein indicators of pork ageing and water-holding capacity. Meat Science 88, 261–270.
| Centrifugal drip is an accessible source for protein indicators of pork ageing and water-holding capacity.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3MXisFGmt7w%3D&md5=f291cade5f30b3ff5d298233549c98feCAS | 21292408PubMed |
Gray JI, Gomaa EA, Buckley DJ (1996) Oxidative quality and shelf life of meats. Meat Science 43, 111–123.
| Oxidative quality and shelf life of meats.Crossref | GoogleScholarGoogle Scholar |
Harada Y, Sato C, Kitajima K (2007) Complex formation of 70-kDa heat shock protein with acidic glycolipids and phospholipids. Biochemical and Biophysical Research Communications 353, 655–660.
| Complex formation of 70-kDa heat shock protein with acidic glycolipids and phospholipids.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2sXmtVahsg%3D%3D&md5=f3008f8b2123069e3abae7f5c8c791bcCAS | 17204248PubMed |
Hernández P, Navarro JL, Toldra F (1999) Lipolytic and oxidative changes in two Spanish pork loin products: dry-cured loin and pickled-cured loin. Meat Science 51, 123–128.
| Lipolytic and oxidative changes in two Spanish pork loin products: dry-cured loin and pickled-cured loin.Crossref | GoogleScholarGoogle Scholar | 22061695PubMed |
Huff-Lonergan E, Lonergan SM (2005) Mechanisms of water-holding capacity of meat: the role of postmortem biochemical and structural changes. Meat Science 71, 194–204.
| Mechanisms of water-holding capacity of meat: the role of postmortem biochemical and structural changes.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2MXmt1Wiurc%3D&md5=4351e176749d38246cf323f41ba5362fCAS | 22064064PubMed |
Jaattela M (1993) Overexpression of major heat shock protein 70 inhibits tumor necrosis factor-induced activation of phospholipase A2. Journal of Immunology (Baltimore, MD.: 1950) 151, 4286–4294.
Lambert LH, Nielsen JH, Andersen HJ (2001) Cellular model for induction of drip loss in meat. Journal of Agricultural and Food Chemistry 49, 4876–4883.
| Cellular model for induction of drip loss in meat.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD3MXntVygtL0%3D&md5=21e444024503773565eee054289a4c03CAS |
Offer G, Cousins T (1992) The mechanism of drip production: formation of 2 compartments of extracellular-space in muscle postmortem. Journal of the Science of Food and Agriculture 58, 107–116.
| The mechanism of drip production: formation of 2 compartments of extracellular-space in muscle postmortem.Crossref | GoogleScholarGoogle Scholar |
Sarkar MK, Kinter M, Mazumder B (2009) Purification and characterisation of a novel antioxidant protein molecule from Phyllanthus niruri. Food Chemistry 114, 1405–1412.
| Purification and characterisation of a novel antioxidant protein molecule from Phyllanthus niruri.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD1MXitFCjtL4%3D&md5=bf5cd2b8ec8102219b044e0253feeaebCAS |
Sklan D, Tenne Z, Budowski P (1983) Simultaneous lipolytic and oxidative changes in turkey meat stored at different temperatures. Journal of the Science of Food and Agriculture 34, 93–99.
| Simultaneous lipolytic and oxidative changes in turkey meat stored at different temperatures.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaL3sXhsF2htbs%3D&md5=c95a6a3a0310410aa05b347be45997cfCAS |
Tsvetkova NM, Horvath I, Torok Z, Wolkers WF, Balogi Z, Shigapova N, Crowe LM, Tablin F, Vierling E, Crowe JH, Vígh L (2002) Small heat-shock proteins regulate membrane lipid polymorphism. Proceedings of the National Academy of Sciences of the United States of America 99, 13504–13509.
| Small heat-shock proteins regulate membrane lipid polymorphism.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD38XotVKms7g%3D&md5=8294e27aaf045e09336815f1c9384deaCAS | 12368478PubMed |
Wang DY, Zhu YZ, Xu WM (2009a) Composition of intramuscular phospholipids and free fatty acids in three kinds of traditional Chinese duck meat products. Poultry Science 88, 221–226.
| Composition of intramuscular phospholipids and free fatty acids in three kinds of traditional Chinese duck meat products.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD1MXhtlamtLg%3D&md5=66b8b103f04cc63087ddb7a2921d28b8CAS | 19096077PubMed |
Wang DY, Xu WM, Xu XL (2009b) Determination of intramuscular phospholipid classes and molecular species in Gaoyou duck. Food Chemistry 112, 150–155.
| Determination of intramuscular phospholipid classes and molecular species in Gaoyou duck.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD1cXovFyjt7o%3D&md5=eaa432c3695fee2ef60e16da6a417308CAS |
Yu J, Tang S, Bao E, Zhang M, Hao Q, Yue Z (2009) The effect of transportation on the expression of heat shock proteins and meat quality of M. longissimus dorsi in pigs. Meat Science 83, 474–478.
| The effect of transportation on the expression of heat shock proteins and meat quality of M. longissimus dorsi in pigs.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD1MXhtV2rt7vP&md5=6a2df2d5bc4fee8b850159cfe740a8d0CAS | 20416675PubMed |