

Proteolysis of meat and bone meal to increase utilisation
G. J. Piazza A B and R. A. Garcia AA US Department of Agriculture1, Agricultural Research Service, Eastern Regional Research Centre, Biobased and Other Animal Coproducts Research Unit, 600 East Mermaid Lane, Wyndmoor, PA 19038, USA.
B Corresponding author. Email: george.piazza@ars.usda.gov
Animal Production Science 54(2) 200-206 https://doi.org/10.1071/AN13041
Submitted: 1 February 2013 Accepted: 19 March 2013 Published: 7 May 2013
Abstract
Meat and bone meal (MBM), an important by-product of the meat industry, is the ground, rendered remainder of farm animals after removal of the hide and meat. Most protein in MBM is insoluble, which limits its usefulness. Defatted, milled porcine MBM was subjected to saturating amounts of trypsin, a selective protease, and subtilisin, a protease with broad selectivity. Samples were withdrawn over a 48-h time course of hydrolysis and filtered to remove insoluble material. The rate at which the MBM protein was converted to a soluble form was equivalent for both proteases. Over the time course, trypsin generated fewer free amino groups than did subtilisin, and at a specified time, the molecular weight (MW) of the soluble trypsin hydrolysate was higher than that of the subtilisin hydrolysate. Assay of amino group formation showed that the proteases were still active even after soluble protein generation had ceased. The hydrolysates are useful for a variety of food and non-food uses. The hydrolysates were tested for flocculation activity in an ongoing effort to find sources for renewable flocculant. Kaolin flocculant activity was observed with the soluble fraction obtained before hydrolysis of MBM and also observed with the relatively high MW hydrolysates from short treatment with trypsin and subtilisin. Low MW fractions obtained from by subtilisin treatment at 30–48 h also showed kaolin-settling ability, probably through a coagulation or charge neutralisation process.
Additional keywords: bridging, kaolin, patch.
Introduction
Approximately 10% of the gross income of the meat industry comes from by-products (Jayathilakan et al. 2012). One such by-product is meat and bone meal (MBM). MBM is the ground, rendered remainder of farm animals after removal of the hide and meat for leather and food use. MBM demand has been limited by governmental restrictions due to disease concerns (Arvanitoyannis and Ladas 2008) and by an increase in available protein. MBM is currently used as an animal feed ingredient in some countries and as a solid fuel. MBM has poor solubility, due to the nature of its protein component plus the high heat used its preparation (Garcia et al. 2006).
Recently there has been an influx into the market place of protein-rich materials, which are by-products of biofuel production. There has also been a surge of interest in the use of proteins in non-food applications due to a need for renewable materials. A partial list of such applications is wood adhesives (Park et al. 2000; Liu and Li 2007), media ingredients for industrial fermentation (Garcia et al. 2011; Liang et al. 2011; Solaiman et al. 2011), commodity chemicals from protein-derived amino acids (Lammens et al. 2012), bio-based plastics (Swain et al. 2004), and renewable flocculant (Piazza and Garcia 2010b).
It has been known for some time that protein hydrolysis of meat industry by-products gives soluble extracts with high protein content (Webster et al. 1982). This research area is still under active investigation (Bhaskar et al. 2007) and has been recently expanded to include the production of bioactive peptides by hydrolysis of edible meat by-products (Toldrá et al. 2012). Nevertheless, a review of the literature shows few studies of MBM hydrolysis. Acid hydrolysis was applied for the determination of amino acid content of MBM (Rayner 1985). Basic and enzymatic hydrolysis was applied to MBM to obtain soluble hydrolysates (Garcia et al. 2011). Basic hydrolysis gave a product with degraded amino acids and high ash content. Enzymatic hydrolysis employed non-specific proteases, which gave soluble hydrolysates with number average molar mass of only 500 Da, and the percent organic matter solubilised was only 40%.
An aqueous extract of (unhydrolysed) MBM was reported to have kaolin (clay) flocculant activity (Piazza and Garcia 2010a). This extract contained only a fraction of the MBM protein, since most MBM protein is insoluble. Base-catalysed hydrolysis of MBM gave hydrolysates, which showed intriguing differences in their flocculation ability depending upon the time they were taken from the hydrolysis mixture (Piazza and Garcia 2010b). The only data concerning flocculation by protease-derived MBM hydrolysates in the literature is a test of a single Versazyme hydrolysate. Thus a more rigorous investigation of flocculation by enzyme-produced MBM hydrolysates is warranted.
In the research results presented here, MBM was hydrolysed with two proteases, trypsin and subtilisin. Trypsin is a selective protease that preferentially cleaves the peptide bond on the carboxyl side of arginine and lysine (Yin et al. 2008). Subtilisin (protease from Bacillus licheniformis) is a protease with broad reactivity (Hill and Schmidt 1962; Guérard et al. 2001). The soluble hydrolysates were tested for their flocculant activity. Finding sources of renewable flocculants is important because currently used flocculants are prepared from materials manufactured from natural gas and petroleum (Salehizadeh and Shojaosadati 2001). Also renewable flocculants may be safer than synthetic flocculants, which are known to be toxic or to degrade to toxic products (Bolto and Gregory 2007).
Materials and methods
Materials
A fine kaolin (clay) with the trade name ‘Polygloss 90’ from Huber Engineered Materials was a gift from the M. F. Cachat Co. (Lakewood, OH, USA). Polygloss 90 had a surface area of 22.0 m2/g. Porcine MBM was a gift from Hormel (Austin, MN, USA). DL-dithiothreitol, MES [2-(morpholino)ethanesulfonic acid], o-phthaldialdehyde, sodium tetraborate decahydrate, protease from Bacillus licheniformis (subtilisin) [Type VIII, lyophilised powder, 7–15 units/mg (1 unit = 1.0 μmol trypsin/min), CAS number 9014–01–1, EC number 3.4.21.14], and trypsin from bovine pancreas [powder, ≥7500 BAEE units/mg solid, CAS number 9002–07–7, EC number 3.4.21.4] were obtained from Sigma-Aldrich (St Louis, MO, USA). Ammonium hydrogen carbonate 98% was from Alfa Aesar (Lancaster, UK). Titan 2, 30-mm Filter Red 0.45 μm, PVDF membrane filters were from Sun Sri (Rockwood, TN, USA). Water was purified to a resistance of 18 MΩ-cm using a Barnstead E-pure system from Thermo Fisher Scientific (Waltham, MA, USA).
Preparation of MBM fractions
Fat removal from MBM was conducted by mixing 3 mL hexane per 1 g MBM for 1 h at 22°C and centrifugation of the mixture at 7000g for 45 min at 20°C. The MBM was air-dried overnight. In 100-g portions, part of the MBM was processed in a cryogenic mill (6800 Freezer Mill, SPEX CertiPrep, Metuchen, NJ, USA) to reduce the particle size. Small portions of the defatted, milled MBM were vacuum dried to constant weight, which showed that MBM contained 2.8 ± 0.1% (s.e., n = 6) water, and relevant calculations were corrected for this water. The defatted, milled MBM contained 48.1 ± 1.1% protein and had an ash content of 34.4 ± 0.1%.
Extraction of MBM protein without protease
Milled, defatted MBM (625 mg; dry weight 606 mg) was extracted with 24 mL aqueous NH3HCO3 (50 mM, pH 9.0) at 30°C in a rotary shaker. At measured times, a portions of the mixture was filtered through 0.45-μm PVDF filters (17 mm, Titan, Rockwood, TN, USA). Duplicate 1-mL portions of the filtrates were transferred to preweighed vials. The buffer and water were removed by placing the vials in a nitrogen stream at 55°C for 12 h (Nitrogen Evaporator, Organomation Associates, Berlin, MA, USA), followed by placement in a vacuum oven at 55°C until constant weight was obtained.
Proteolysis and extraction of MBM protein
Milled, defatted MBM was subjected to hydrolysis by proteases. In a typical procedure 10 g (dry weight 9.7 g) of MBM was suspended in 400 mL aqueous NH3HCO3 (50 mM, pH 9.0), and the reaction vessel was placed in a rotary shaker at 30oC. After 20 min, a 10-mL sample was removed for the ‘zero’ point. The indicated amount of protease was added, and samples were subsequently withdrawn at the indicated times. The pH of the samples was lowered to 3.0 to terminate the reaction. The acidified samples were filtered through 0.45-μm PVDF filters, and dry weight of the filtrates was determined as described above.
Protein measurement of MBM from nitrogen analyses
Nitrogen content was determined using an automated analyser (model FP-2000, Leco Corp., St Joseph, MI, USA). A nitrogen to protein conversion factor of 5.37 is based on complete amino acid analysis of MBM (Sriperm et al. 2011).
Degree of peptide bond hydrolysis
The time course of MBM hydrolysis was followed by peptide bond cleavage using a spectrophotometric assay for amino group formation by derivation with o-phthalaldehyde (Nielsen et al. 2001) with the addition of dimethylsulfoxide to stabilise the chromophore, as recommended (Church et al. 1985). The ‘o-phthalaldehyde reagent’ contained 90 mg o-phthalaldehyde, 127 mg sodium dodecyl sulfate, 100 mg dithiothreitol. To this mixture, the following liquids were added sequentially with stirring: 2 mL ethanol, 80 mL sodium tetraborate decahydrate (50 mM), and 18 mL dimethyl sulfoxide. To assay amino groups, 1 mL of o-phthalaldehyde reagent was added to a 0.2-mL sample with mixing. The absorbance at 340 nm was measured after 3 min. Calibration curves were prepared using serine as an amino group standard. Total amino groups in completely hydrolysed MBM were obtained using the hydrolysis method described in Gehrke et al. (1985).
Flocculation testing
A published method was used for measuring the flocculation ability of the hydrolysates (Piazza and Garcia 2010a). Hydrolysate (volume adjusted to give 8 mg dry weight) was dispersed in aqueous buffer [2-(N-Morpholino)ethanesulfonic acid], and the pH was adjusted to 5.5. Settling of kaolin (clay) was determined at 1 and 5 h. All measurements were repeated 5 times, and the mean values ± s.e. were calculated.
Gel filtration chromatography
Molecular weight (MW) standards: dipeptide (Ser-Gly) and pentapeptide (Peptide 6A) were from Bachem Americas (Torrance, CA, USA). Aprotinin, cytochrome C, carbonic anhydrase, albumin, and bradykinin were from a kit for MW 6500–68 000 for gel filtration chromatography, Sigma-Aldrich. Thyroglobulin, gamma globulin, ovalbumin were components of a gel filtration standard from Biorad (Richmond, CA, USA). Alanine, glutamic acid, and phenylalanine were from Sigma.
HPLC gel filtration chromatography was conducted under isocratic conditions on a Waters 2695 Separation Module with Varian 380-LC evaporative light scattering detector using a Superdex Peptide column 10/300 GL (300 × 10 mm, GE Healthcare, Piscataway, NJ, USA). Each chromatography run was for 60 min. The eluting solvent was 50 mM ammonium hydrogen carbonate.
Results and discussion
Extraction of MBM without protease
Milled, defatted MBM was extracted with aqueous ammonium hydrogen carbonate buffer (pH 9.0) to measure its water-soluble content. The time course of extraction of soluble mass (15 min to 24 h) was followed, and the percent mass was calculated. After 15 min of extraction, the mass percent of extracted material was 28.3 ± 1.5%. The mass percent of extracted material at 24 h was identical within experimental error to that obtained after 15 min. Thus, the majority of MBM consists of insoluble material under these extraction conditions, and increasing extraction time does not increase MBM solubility.
Enzymatic extraction of MBM mass
Milled, defatted MBM was hydrolysed with the proteases trypsin and subtilisin. The amount of protease used was determined empirically to give the maximal rate of hydrolysis. The time course of hydrolysis was followed by measuring the dried mass of the water-soluble material and calculating percent mass. Fig. 1 shows composite data from three experiments for each protease.
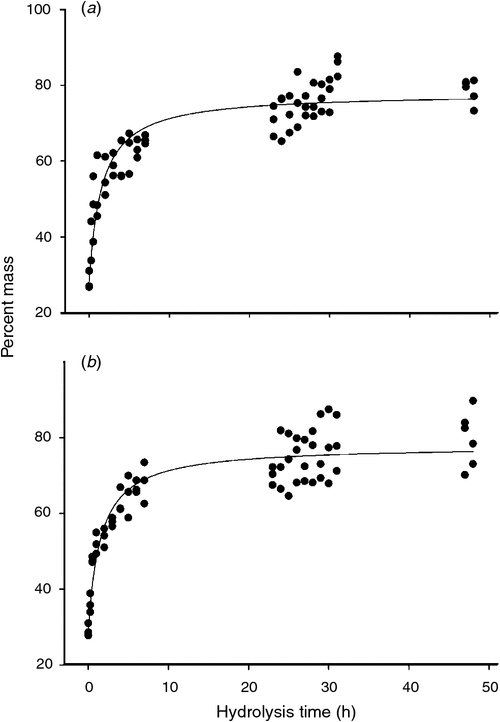
|
The percent of soluble mass increased at a rapid rate during the first 7 h of hydrolysis with trypsin (Fig. 1a). At times greater than 7 h, the percent of soluble mass increased only modestly. The curved line in Fig. 1a is the fit of the data by non-linear regression analysis to the following function: f(t) = [(Mpt)/(K + t)] + Mint where Mp is the percent of soluble mass attainable at infinite time by protease addition; K is a constant; t is the hydrolysis time; Mint is the initial percent of soluble mass obtained by mixing MBM and aqueous buffer before protease was added. For the data in Fig. 1a the values of Mp, K, and Mint are 49.7 ± 1.2%, 1.6 ± 0.2, and 28.2 ± 1.4%. The sum of Mint and Mp is the maximum percent of soluble MBM mass (Mmax) and is 77.9 ± 1.9%.
The data shown in Fig. 1b shows the percent dried mass of the water-soluble material obtained by treatment of MBM with the protease subtilisin. From non-linear regression analysis, the values of Mp, K, and Mint are 48.7 ± 1.1%, 1.6 ± 0.2, and 29.1 ± 1.9%, giving a value for Mmax as 77.8 ± 1.9%. Thus, the estimates of Mp, K, Mint, and Mmax obtained from trypsin or subtilisin hydrolysis were remarkably similar.
Protein measurement of extracts
Duplicate portions of the dried trypsin and subtilisin-treated soluble extracts obtained during the time course were analysed for nitrogen, and percent protein was calculated (Sriperm et al. 2011). The mean percent protein in the soluble extracts was 63.9 ± 1.2% after correction for the protein in trypsin or subtilisin. Since average Mmax is 77.85 ± 1.90% for trypsin and subtilisin-treated MBM, the percent of protein extracted from MBM is 49.7 ± 1.5% (0.7785 × 63.9). This product is, within experimental error, equal to the percent protein in dried MBM: 48.1 ± 1.1%. The equivalence of these values shows that all the protein contained in MBM was converted to a soluble form by proteolysis with trypsin or subtilisin.
A reviewer questioned whether the collagen present in MBM is hydrolysed by proteases. While it is true that the triple helix structure of collagen usually prevents or dramatically slows proteolysis, the collagen structure is disrupted by heat which allows protease digestion (Bruckner and Prockop 1981). Specific recent examples of proteolysis of collagen by trypsin are given in Yuanlong et al. (2012) and Zhang et al. (2006). An example of proteolysis of collagen by subtilisin is given in Vasileva-Tonkova et al. (2007). Additionally, it has been claimed that finely divided collagen powders are also digested by trypsin (Thomas and Seymour-Jones 1923), although the heat generated in the collagen sample by the grinding process may have been sufficient to denature the collagen. Normal procedures for cooking and drying MBM use temperatures in the range of 130−150°C, which are sufficiently high to denature the collagen in MBM and make it susceptible to proteolysis.
Peptide bond hydrolysis
Peptide bond hydrolysis was monitored over the hydrolysis time course by measuring the appearance of free amino groups. After an initial rapid increase in free amino groups, the rate of amino formation continued from 5 to 48 h at a reduced rate. At times after 30 min, the free amino groups generated by subtilisin were greater than those generated by trypsin. By 48 h, the free amino groups given by subtilisin were 1.15 ± 0.10 mmol/g MBM, while those generated by trypsin were 0.66 ± 0.04 mmol/g MBM. It is notable, that there was a significant increase in amino groups going from the second day of collection (23–31 h) to the third day of collection (47–48 h) for both subtilisin- and trypsin-promoted hydrolysis. This shows that subtilisin and trypsin were still active during the 23–48-h time period. The flattening of the soluble mass curves (Fig. 1) during this time period was not due to insufficient proteolytic activity, but was instead due to depletion of insoluble protein from MBM.
Fig. 2 shows the concentration of amino groups (μmol/mL). The amino group concentration data show that amino groups increased throughout the time course. Subtilisin generated a higher concentration of free amino groups than did trypsin. Fig. 2 also shows the ratio of amino groups to the soluble mass (μmol/mg). In contrast to the concentration data, the ratio of amino to soluble mass for trypsin was approximately constant up to 30 h. After 30 h, there was almost no additional soluble mass released from MBM, but amino groups continued increasing, and the ratio of amino to soluble mass increased. In contrast to the trypsin results, the ratio of amino to soluble mass for subtilisin increased throughout the time course. According to Figs 1a and b, subtilisin released soluble mass at nearly the same rate as trypsin. However, with subtilisin, the degree of proteolysis was in excess of what was needed to release soluble mass. Thus the data show that while proteolysis is important for mass extraction, there is not a correlation between the degree of proteolysis and the formation of soluble mass.
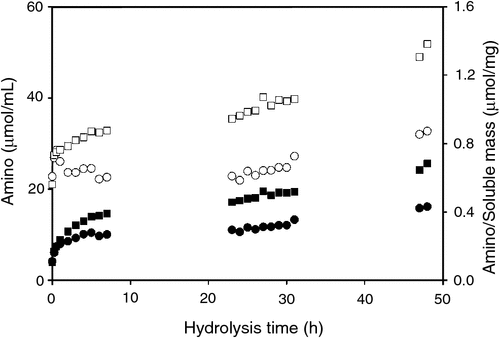
|
The equivalence in the rates of soluble mass generation by subtilisin and trypsin suggests that in addition to amide bond hydrolysis rate, there is a factor which is important in controlling the rate of soluble mass generation. One possibility is that that after the enzymes hydrolyse the protein in MBM, the process of solubilising the resulting protein fragments is not instantaneous, and the rate of this solubilising process limits the overall rate of formation of soluble mass. Also, the proteases may not be able to contact all of the MBM protein at once and may instead only hydrolyse that protein localised at or near the surface of the MBM particle. The proteases are able to increase their penetration into the MBM particle as the surface proteins are hydrolysed and solubilised. This overall process, penetration and hydrolysis, occurs at a rate which is equal for each protease.
Molecular weight distribution
The influence of time on the MW distribution of material in the soluble fraction of the trypsin hydrolysates is shown in Fig. 3. In the first 15 min (0.25 h) large decreases in >30 and 10–30 kDa materials were observed, with a corresponding increase in material with MW <10 kDa. With times greater than 15 min, there was a relatively gradual change in MW distribution with the lower MW materials gradually increasing. At 47–48 h, the material of greatest abundance was 1–5 kDa, and there was also some increase in material with MW <0.2 kDa (free amino acids).
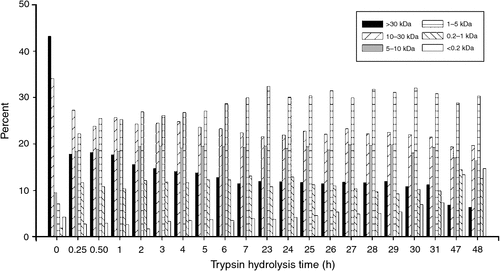
|
The MW distribution of soluble material in the subtilisin hydrolysates is shown in Fig. 4. A gradual decrease in the >30 kDa proteinaceous material was observed to 31 h. The 10–30 and 5–10 kDa materials also decreased to 23 h, and thereafter the amounts of these materials remain approximately constant until 47–48 h where there was a decrease. At 47–48 h, the >30 kDa materials were absent, and the 10–30, 5–10, and 1–5 kDa materials were at their lowest levels. At 47–48 h there was a gradual increase in the <0.2 kDa material up to 31 h, and at 47–48 h, there was a large increase in the <0.2 kDa material.
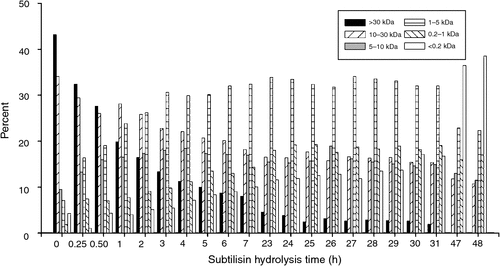
|
The MW distribution data in Figs 3 and 4 are consistent with the amino group formation data in Fig. 2, which show that more amide bond hydrolysis is catalysed by subtilisin than by trypsin. Trypsin is a highly selective proteinase, and this selectivity promotes the formation of higher MW fragments, although some lower MW fragments form also. In contrast, subtilisin has relatively broad selectivity, and its action gives a higher amide bond hydrolysis rate resulting in greater amounts of lower MW material.
Flocculation activity
An equal dry weight portion of the soluble filtrate from each sampled time was tested for its ability to flocculate kaolin. Flocculation testing was conducted at pH 5.5 because at this pH, the kaolin particles do not spontaneously flocculate, but proteins, peptides, and amino acids, which compose the MBM hydrolysates, have net positive charge that acts to neutralise the negative charge on the surface of the kaolin particles. The pooled results of the flocculation tests from trypsin hydrolysates are shown in Fig. 5. Kaolin settling activity was noted from the soluble portion of untreated MBM, which contained 28% of the starting MBM mass (zero time in figure). Kaolin settling activity from the soluble hydrolysates was retained to 7 h of hydrolysis, which rendered soluble 68% of the MBM. In interpreting the results, it is important to note that Fig. 5 shows the results from a fixed quantity (0.8 g dry weight) of the soluble material. The total kaolin settling activity of the 7-h hydrolysate is actually greater than indicated in the Figure because the 7-h hydrolysate contained 40% more soluble material than prehydrolysis MBM. Nevertheless, the kaolin settling activity in the soluble hydrolysates was no longer evident in those samples taken after 7 h, and this shows that larger MW polypeptides are necessary for flocculant activity. The need for larger MW polypeptides is consistent with the idea that the flocculant is operating through a bridging mechanism, such that the flocculant ‘bridges’ many individual particles to build a floc (Bratby 2006). The lack of kaolin settling activity from the longer time hydrolysates also shows that even a selective protease such as trypsin cannot release all the protein from MBM and maintain large enough protein fragments to give an active flocculant material.
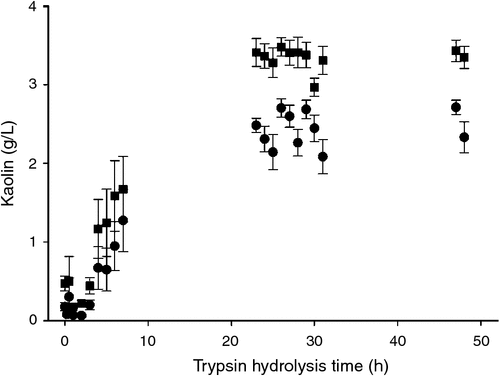
|
The soluble filtrates taken from the subtilisin hydrolysis time course trials were also tested for flocculation activity (Fig. 6). Subtilisin hydrolsates gave kaolin settling only from hydrolysates obtained from 1 h or less. Since the size data (Fig. 4) clearly shows that subtilisin gives small polypeptide fragments, the loss of kaolin settling activity after only 1 h is consistent with the idea that flocculant that functions by the bridging mechanism which requires larger MW polypeptides. The most interesting aspect of Fig. 6 was the unexpected kaolin settling activity exhibited by the 30–48-h soluble hydrolysates. Fig. 4 shows that fractions 30–48 h contained mostly lower MW material.
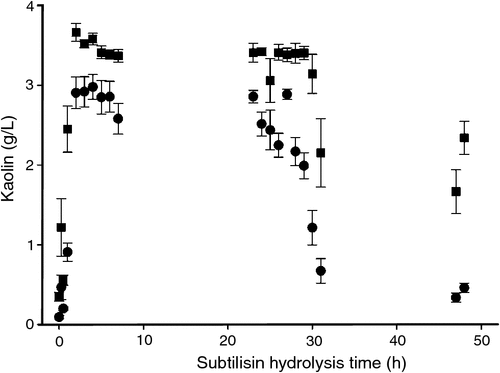
|
The predominant mechanism of kaolin settling activity by the low MW hydrolysates is likely to be the patch or neutralisation mechanism; the positively charged amino groups in the MBM hydrolysate bind to the surface of the kaolin particles and neutralise the negative surface charge of the kaolin (Bratby 2006). In order to effectively neutralise the negative charge of the kaolin particles, the amino groups must be closely spaced on the surface of the kaolin particles. To achieve such close spacing, the size of the protein remnants needs to be relatively small. Lower MW synthetic or inorganic materials that cause sedimentation are usually referred to as coagulants (Bratby 2006).
Flocculation testing of the hydrolysates of trypsin and subtilisin revealed that there are two kinds of kaolin settling activity formed by proteolysis, a high MW and a low MW material. With respect to the high MW hydrolysate, there was 10.1 μmol/mL amino group formation at 7 h of trypsin hydrolysis and 8.9 μmol/mL amino group formation with 1 h subtilisin hydrolysis. Thus, when amino group formation is less than or equal to ~9–10 μmol/mL, the hydrolysates exhibited kaolin flocculant activity.
For the low MW material from subtilisin hydrolysis that gave kaolin coagulant activity, amino group formation is greater than or equal to 19.2 μmol/mL at 30–48 h (Fig. 2). This is higher than the amino group concentration measured in any of the trypsin hydrolysates. The need for a high degree of proteolysis explains why no low MW kaolin coagulant activity was observed in the trypsin hydrolysates.
Surprisingly, there is not a great deal of difference in the mass of flocculant or coagulant materials that may be generated by treatment with either enzyme. With 7 h treatment with trypsin, 17 mg/mL of high MW flocculant was prepared. With 30–48 h treatment with subtilisin, 18 mg/mL of low MW coagulant was prepared.
Conclusions
As indicated by free amine formation, the rate of proteolysis with subtilisin was greater than that given by trypsin, but the rate of soluble mass generation was the same for both proteases, suggesting that structural characteristics of MBM limit the rate at which its mass can be converted to soluble hydrolysate. Nitrogen analysis of the soluble fraction compared with the nitrogen in the original MBM indicated that all of the protein was converted to a soluble form. The soluble MBM hydrolysates gave both a relatively high MW flocculant and relatively low MW coagulant. This is the first report of two different MW kaolin settling materials being prepared from a single starting material. In addition to its use as a flocculant and coagulant, the MBM-soluble hydrolysate may also be used for animal or human nutrition in some countries (Alvarez et al. 2012) and as a high quality media ingredient for industrial fermentation.
Acknowledgement
Technical assistance was provided by F. Fox.
References
Alvarez C, Rendueles M, Diaz M (2012) The yield of peptides and amino acids following acid hydrolysis of haemoglobin from porcine blood. Animal Production Science 52, 313–320.| The yield of peptides and amino acids following acid hydrolysis of haemoglobin from porcine blood.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC38XmvVKitrY%3D&md5=7a453b6ccfe69bb9520c64cbb204962dCAS |
Arvanitoyannis IS, Ladas D (2008) Meat waste treatment methods and potential uses. International Journal of Food Science & Technology 43, 543–559.
| Meat waste treatment methods and potential uses.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD1cXisFyktLg%3D&md5=a86e9066dbdef415f89002bf00f97b48CAS |
Bhaskar N, Modi VK, Govindaraju K, Radha C, Lalitha RG (2007) Utilization of meat industry by products: protein hydrolysate from sheep visceral mass. Bioresource Technology 98, 388–394.
| Utilization of meat industry by products: protein hydrolysate from sheep visceral mass.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD28XptlCqsL0%3D&md5=a409f5428890e023f08b978522583b29CAS | 16457999PubMed |
Bolto B, Gregory J (2007) Organic polyelectrolytes in water treatment. Water Research 41, 2301–2324.
| Organic polyelectrolytes in water treatment.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2sXkvFWnur4%3D&md5=a5f2fff8b7de3ca05325d3494d866c66CAS | 17462699PubMed |
Bratby J (2006) ‘Coagulation and flocculation in water and wastewater treatment.’ 2nd edn. (IWA Publishing: London)
Bruckner P, Prockop DJ (1981) Proteolytic enzymes as probes for the triple-helical conformation of procollagen. Analytical Biochemistry 110, 360–368.
| Proteolytic enzymes as probes for the triple-helical conformation of procollagen.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaL3MXhtlGntbo%3D&md5=b5ac4676c648a5c974731836f3b49380CAS | 7015914PubMed |
Church FC, Porter DH, Catignani GL, Swaisgood HE (1985) An o-phthalaldehyde spectrophotometric assay for proteinases. Analytical Biochemistry 146, 343–348.
| An o-phthalaldehyde spectrophotometric assay for proteinases.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaL2MXhs1altL0%3D&md5=97e55534e7c678f604cc912a299257b9CAS | 3896021PubMed |
Garcia RA, Rosentrater KA, Flores RA (2006) Characteristics of North American meat and bone meal relevant to the development of non-feed applications. Applied Engineering in Agriculture 22, 729–736.
Garcia RA, Pyle DJ, Piazza GJ, Wen Z (2011) Hydrolysis of animal protein meals for improved utility in non-feed applications. Applied Engineering in Agriculture 27, 269–275.
Gehrke CW, Wall LL, Absheer JS (1985) Sample preparation for chromatography of amino acids: acid hydrolysis of proteins. Journal – Association of Official Analytical Chemists 68, 811–821.
Guérard F, Dufossé L, De La Broise D, Binet A (2001) Enzymatic hydrolysis of proteins from yellowfin tuna (Thunnus albacores) wastes using Alcalase. Journal of Molecular Catalysis. B, Enzymatic 11, 1051–1059.
| Enzymatic hydrolysis of proteins from yellowfin tuna (Thunnus albacores) wastes using Alcalase.Crossref | GoogleScholarGoogle Scholar |
Hill RL, Schmidt WR (1962) The complete enzymatic hydrolysis of proteins. The Journal of Biological Chemistry 237, 389–396.
Jayathilakan K, Sudtana K, Radhakrishna K, Bawa AS (2012) Utilization of byproducts and waste materials from neat, poultry and fish processing industries: a review. Journal of Food Science and Technology 49, 278–293.
| Utilization of byproducts and waste materials from neat, poultry and fish processing industries: a review.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC38Xktlemu7w%3D&md5=0593a1f04f2e45cdd02d94af17e669a4CAS |
Lammens TM, Franssen MCR, Scott EL, Sanders JPM (2012) Availability of protein-derived amino acids as feedstock for the production of bio-based chemicals. Biomass and Bioenergy 44, 168–181.
| Availability of protein-derived amino acids as feedstock for the production of bio-based chemicals.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC38Xptl2mtrg%3D&md5=8789f24c1489f37d7e5bb895f5db9e68CAS |
Liang Y, Garcia RA, Piazza GJ, Wen Z (2011) Nonfeed application of rendered animal proteins for microbial production of eicosapentaenoic acid by the fungus Pythium irregular. Journal of Agricultural and Food Chemistry 59, 11 990–11 996.
| Nonfeed application of rendered animal proteins for microbial production of eicosapentaenoic acid by the fungus Pythium irregular.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3MXhtlaqsLjK&md5=cc8c17dcf6b540cdb2d86b6b401ac022CAS |
Liu Y, Li K (2007) Development and characterization of adhesives from soy protein for bonding wood. International Journal of Adhesion and Adhesives 27, 59–67.
| Development and characterization of adhesives from soy protein for bonding wood.Crossref | GoogleScholarGoogle Scholar |
Nielsen PM, Petersen D, Dambmann C (2001) Improved method for determining food protein degree of hydrolysis. Food and Chemical Toxicology 66, 642–646.
Park SK, Bae DH, Hettiarachchy NS (2000) Protein concentrate and adhesives from meat and bone meal. Journal of the American Oil Chemists’. Society 77, 1223–1227.
Piazza GJ, Garcia RA (2010a) Meat and bone meal extract and gelatin as renewable flocculants. Bioresource Technology 101, 781–787.
| Meat and bone meal extract and gelatin as renewable flocculants.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD1MXht1amu7vO&md5=549b7e0d2880aca9f22fdc980732ef37CAS | 19734043PubMed |
Piazza GJ, Garcia RA (2010b) Proteins and peptides as renewable flocculants. Bioresource Technology 101, 5759–5766.
| Proteins and peptides as renewable flocculants.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3cXlt1SisLc%3D&md5=95c19bfb4b33316fb92822ede88a672eCAS | 20236820PubMed |
Rayner CJ (1985) Protein hydrolysis of animal feeds for amino acid content. Journal of Agricultural and Food Chemistry 33, 722–725.
| Protein hydrolysis of animal feeds for amino acid content.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaL2MXktlKhtrg%3D&md5=99f7c20fe1c76d7dfeb5ccbc2a86a28fCAS |
Salehizadeh H, Shojaosadati SA (2001) Extracellular biopolymeric flocculants, recent trends and biotechnological importance. Biotechnology Advances 19, 371–385.
| Extracellular biopolymeric flocculants, recent trends and biotechnological importance.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD3MXms1OksL8%3D&md5=d91913c83124d3179700e862c21e356eCAS | 14538073PubMed |
Solaiman DKY, Garcia RA, Ashby RD, Piazza GJ, Steinbüchel A (2011) Rendered-protein hydrolysates for microbial synthesis of cyanophycin biopolymer. New Biotechnology 28, 552–558.
| Rendered-protein hydrolysates for microbial synthesis of cyanophycin biopolymer.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3MXhtFymt7fI&md5=6857db8745d7f46678414ee75015ee9fCAS |
Sriperm N, Pesti GM, Tillman PB (2011) Evaluation of the fixed nitrogen-to-protein (N : P) conversion factor (6.25) versus ingredient specific N : P conversion factors in feedstuffs. Journal of the Science of Food and Agriculture 91, 1182–1186.
| Evaluation of the fixed nitrogen-to-protein (N : P) conversion factor (6.25) versus ingredient specific N : P conversion factors in feedstuffs.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3MXkt12hu7c%3D&md5=d2011be3540f2938d1bc54640787b7f0CAS | 21305546PubMed |
Swain SN, Biswal SM, Nanda PK, Nayak PJ (2004) Biodegradable soy-based plastics: opportunities and challenges. Journal of Polymers and the Environment 12, 35–42.
| Biodegradable soy-based plastics: opportunities and challenges.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD3sXovV2ks7w%3D&md5=8b007e2fa8cff1e2ffbb592128ff1315CAS |
Thomas AW, Seymour-Jones FL (1923) The hydrolysis of collagen by trypsin. Journal of the American Chemical Society 45, 1515–1522.
| The hydrolysis of collagen by trypsin.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaB3sXhsFyksQ%3D%3D&md5=605589780e375ddfe0e8077d3660a50aCAS |
Toldrá F, Aristoy M-C, Mora L, Reig M (2012) Innovations in value-addition of edible meat by-products. Meat Science 92, 290–296.
| Innovations in value-addition of edible meat by-products.Crossref | GoogleScholarGoogle Scholar | 22560456PubMed |
Vasileva-Tonkova E, Nustorova M, Gushterova A (2007) New protein hydrolysates from collagen wastes used as peptone for bacterial growth. Current Microbiology 54, 54–57.
| New protein hydrolysates from collagen wastes used as peptone for bacterial growth.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD28XhtlalurvF&md5=92444adead09d11de27f72cbb0c9005bCAS | 17171464PubMed |
Webster JD, Ledward DA, Lawrie RA (1982) Protein hydrolysates from meat industry by-products. Meat Science 7, 147–157.
| Protein hydrolysates from meat industry by-products.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaL3sXjs1On&md5=96393819e147f31e11f9ecf0e1170c82CAS | 22055137PubMed |
Yin S-W, Tang C-H, Cao J-S, Hu E-K, Wen Q-B, Yang X-B (2008) Effects of limited enzymatic hydrolysis with trypsin on the functional properties of hemp (Cannabis sativa L.) protein isolate. Food Chemistry 106, 1004–1013.
| Effects of limited enzymatic hydrolysis with trypsin on the functional properties of hemp (Cannabis sativa L.) protein isolate.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2sXhtVerur7E&md5=464dfc03e952e79f0047cadc3e058347CAS |
Yuanlong C, Min C, Xiaoju C, Wenhua Z, Xuepin L, Bi S (2012) Enzymatic hydrolysis of skin shavings for preparation of collagen hydrolysates with specified molecular weight distribution. Journal of the Society of Leather Technologies and Chemists 96, 16–20.
Zhang G, Sun A, Li W, Liu T, Zhiguo S (2006) Mass spectrometric analysis of enzymatic digestion of denatured collagen for identification of collagen type. Journal of Chromatography. A 1114, 274–277.
| Mass spectrometric analysis of enzymatic digestion of denatured collagen for identification of collagen type.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD28XktVyis7Y%3D&md5=058fa52b9b4b76e9686646fd25def7abCAS | 16600269PubMed |
1 Mention of trade names or commercial products in this publication is solely for the purpose of providing specific information and does not imply recommendation or endorsement by the US Department of Agriculture (USDA). USDA is an equal opportunity provider and employer.