

Effect of replacing a commercial pelleted calf meal with lucerne leaf-meal on performance of neonatal and transitional Holstein heifer calves
Joyce L. Marumo A C , Florence V. Nherera-Chokuda B , Jones W. Ng’ambi A and Mukengela C. Muya BA University of Limpopo, Private Bag X1106, Sovenga, 0727, South Africa.
B Agriculture Research Council, Private Bag X02 Irene, 0062, Pretoria, South Africa.
C Corresponding author. Email: joyceledilemangena0@gmail.com
Animal Production Science 58(5) 834-840 https://doi.org/10.1071/AN16529
Submitted: 2 August 2016 Accepted: 1 October 2016 Published: 28 November 2016
Journal compilation © CSIRO 2018 Open Access CC BY-NC-ND
Abstract
A study was conducted to (i) determine in vitro ruminal dry matter (DM) fermentation kinetics, effective rumen degradation of DM (EDDM), (ii) estimate energy fractions supply of lucerne leaf-meal (LLM) and composite diets of LLM with commercial pelleted calf meal and also (iii) determine the effects of substituting commercial pelleted calf meal with LLM on the intake, % ruminal nitrogen balance (RNB) and growth of the neonates (21–42 days old) and transition (43–56 days old) Holstein heifer calves. Forty-eight Holstein heifer calves were randomly assigned to three different dietary treatments in a complete randomised design of: (a) pelleted concentrate (PEL), (b) 65% pelleted concentrate: 35% LLM (P65 L35); (c) and 50% pelleted concentrate: 50% LLM (P50 L50). The study comprised of two experiments: neonatal (Experiment 1) and transition (Experiment 2) phases. Lucerne leaf-meal had gross energy of 16.2 MJ/kg and 25% crude protein DM. PEL diet was high in starch and bound protein compared with other diets. Inclusion of LLM in diets increased calcium levels but tended to decrease phosphorus levels. Calves were weaned at the age of 56 days. The feeds were incubated for 0, 4, 10, 18, 24 and 48 h using a DaisyII incubator. Rumen fluid was obtained from calves <50 days old. Large Ruminant Nutrition System was used to predict %RNB and energy density of the diets during neonatal and transition phases. Higher mean EDDM levels were found with LLM inclusions whereas fractions a, a + b and c did not vary. Neonates on diet C had higher (P < 0.05) daily DM and crude protein intakes, %RNB, total digestible nutrients, net energy at maintenance and net energy at gain during neonatal phase. Performance of calves was similar during the transition phase. LLM should be considered as a concentrate replacement in diets of neonates and calves.
Additional keywords: concentrate feed, performance, pre-weaning, ruminal nitrogen balance, transition.
Introduction
Protein nutrition is an essential component of neonatal dairy heifers. Commercial concentrates that are high in protein are often the main solid feed for dairy calves. Protein-rich forages are also potential protein sources for neonates. Castells et al. (2012, 2013), Jones and Heinrichs (2007) and Göncü et al. (2010) showed that forages improve rumen development in calves. Castells et al. (2012, 2013) reported increased rumen fill, pH, acetate production, and high population of Ruminococcus albus in calves supplemented with lucerne (Medicago sativa) hay. Lucerne is utilised widely in dairy cattle production and leaf-meals for monogastrics (Guenthner et al. 1973; Lindberg et al. 1995; Rechulicz et al. 2014). Lucerne leaf-meal (LLM) is also a herbal supplement for humans because the leaves are high in essential nutrients and antioxidants (Xie et al. 2008) and low in fibre. They are also high in methionine and lysine (Homolka et al. 2008). Lucerne hay is, however, low in essential nutrients such as phosphorus, non-fibre carbohydrates such as starch (Mason 1998) and has low digestibility compared with some grasses like tall fescue and red clover (Cherney et al. 2004; Homolka et al. 2012). Among all other leguminous forages lucerne is peculiar because of its high nutrients and antioxidant levels (Xie et al. 2008). However, information on LLM supplementation for optimal feed intake and digestion in pre- and post-weaned dairy calves is limited and inconclusive. Kuan et al. (1983) reported that high proportion of LLM in the diet resulted in a linear decrease in the digestibility of dry matter, crude protein and crude fat. However, Lindberg et al. (1995) did not report any negative effects on diet digestibility. The low fibre, high β-carotene and protein contents in LLM make it an attractive substitute forage for commercial calf feeding.
The aim of the study was, therefore, to assess nutrient intake, rumen degradability and growth of Holstein heifers during neonatal and weaning periods.
Materials and methods
Study location
The study was conducted at Irene, Animal Production Institute of the Agricultural Research Council of South Africa in 2014. Irene is located at longitude 28°13S, latitude 25°55E, altitude of 1524 m.
Forage processing and dietary nutrient composition
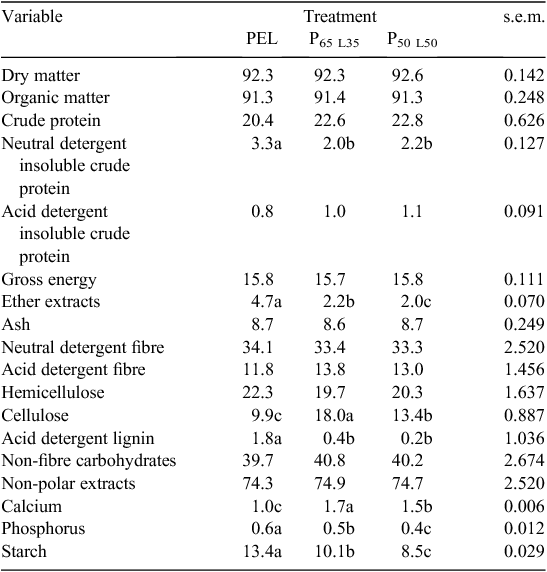
Six-week regrowth lucerne was hand-harvested at 5 cm above the ground and air-dried under shade to prevent nutrient loss (Smit 2014) over 7 days, with daily turning to prevent moulding (Mujumdar 1997). After drying, petioles were separated from stems and small twigs were crumbled manually to form LLM and stored in a dry area. Three diets were prepared using mixtures of LLM and a commercial pelleted calf meal concentrate (PEL). The diets were as follows: 100% PEL; 65% PEL with 35% LLM (P65 L35) and 50% PEL with 50% LLM (P50 L50). The nutrient contents of the dietary treatments are presented in Table 1. Lucerne leaf-meal had 95% DM, 90.5% organic matter, 25% crude protein, 0.6% neutral detergent insoluble crude protein, 1.3% acid detergent insoluble crude protein, 16.2 MJ/kg DM gross energy, 1.6% ether extract, 9.5% ash, 22.5% neutral detergent fibre, 7.7% acid detergent fibre, 14.0% hemicellulose, 14.8 cellulose, 0.8% acid detergent lignin, 40.8% non-structural fibre 1.4% calcium, 0.8% phosphorus and 0.2% starch. All the data were expressed on % DM.
|
In vitro degradation
Ground samples of 0.25 g of the three dietary treatments (PEL, P65 L35 or P50 L50) and LLM were weighed into ash-free and N-free nylon bags (Ankom F57, Ankom Technology Corp., Fairpost, NY, USA) and then heat sealed. In vitro micro mineral, macro mineral, reducing and buffer solutions based on ANKOM (Ankom Technology Corp.) procedures were prepared. Rumen fluid was collected from non-experimental dairy Holstein calves aged 50 days old using an oesophageal tube under mild vacuum suction (Tufarelli et al. 2010) into a pre-warmed CO2-filled thermos flask. Rumen fluid was collected before calves were fed.
In vitro fermentation was assessed at 0, 4, 10, 24 and 48 h using a Daisy II incubator (D200, Ankom Technology Corp.). The incubator maintained a constant temperature of 39°C. All samples were analysed in triplicate. Zero-hour bags were washed with water. At termination bags were washed under running water. Samples were placed into a 100°C oven for 48 h, weighed for DM and ashed to determine organic degradability values. The NDF and crude protein disappearance were estimated at 24 and 48 h because these are slowly degradable fractions.
Animals and data collection
Intake and growth study was divided into two experiments: neonatal phase (Experiment 1: involving 24 calves aged 21–41 days) and transition phase (Experiment 2: involving 24 calves aged 42–56 days).
Experiment 1: calves were randomly allocated to the three diets at 21 days old, fed 4 L of milk/day, and assessed for nutrient intake and growth until 42 days old. Experiment 2: 24 calves aged 42 days were randomly allocated to the three diets and milk supply reduced to 2 L/calf.day. Nutrient intake and growth were assessed until weaning at 56 days old. Calves in both experiments were individually housed in 2 × 5-m2 concrete-floored pens without bedding with ad libitum access to fresh water throughout the experimental periods. Milk and solid feed consumption were recorded daily. Bodyweight was recorded weekly before the morning feeding. Milk contained 3.8% fat, 3.5% protein and 4.8% lactose.
Simulation of LLM and diets
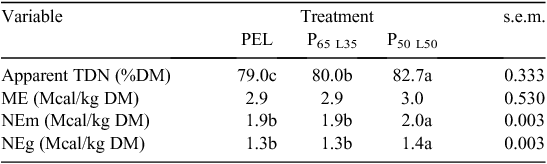
Large Ruminant Nutrition System (LRNS) model of the University of California-Davis (LRNS version 1.0.31, 2014) was used to predict energy nutrient density supply and balances of the three diets of LLM and commercial concentrates. The LRNS is based on the Cornell Net Carbohydrate and Protein System Version 5.0.40 (Fox et al. 2004). Level 1 solutions of the LRNS model were used to predict % ruminal nitrogen balance (RNB) (Table 2) and diet concentrations (Table 3). During the transition phase, diets were not predicted for energy density because the diets and chemical composition were similar to the diets used in neonatal phase and only quantity varied. The LRNS predictions were based on tropical climatic conditions of 22°C, 30% relative humidity and wind-speed of zero for calves housed in a 10-m2 cubicle with no exposure to storms and sunlight.

|
|
Chemical analyses
Dry matter of ingredients and diets was determined according to AOAC (2000) (Procedure 930.15). Ash contents and organic matter were determined according to AOAC (2000) (Procedure 942.05) at 550°C for 8 h. Ether extract was determined according to AOAC (2006) (Procedure 2003.05). Crude protein was determined using the Kjeldahl procedure (AOAC 2000) (Procedure 968.06). Gross energy values of the feed samples were determined by combustion in an adiabatic bomb calorimeter (PARR model 2081). Starch was determined by a modification of the method of Holm et al. (1986) as cited by Hall (2000). Calcium was determined according to Giron (1973) using a Perkin Elmer atomic spectrophotometer. Phosphorus was assayed according to AOAC (2000) (Procedure 965.17). Neutral detergent fibre and acid detergent fibre were determined according to Van Soest et al. (1991) and acid detergent lignin was determined according to Goering and Van Soest (1970). Acid detergent insoluble crude protein and neutral detergent insoluble crude protein (Licitra et al. 1996) were determined by measuring the crude protein content of the acid detergent fibre and neutral detergent fibre residue by Kjeldhal analysis and contents were expressed as a percentage of total N (Van Soest et al. 1991). All samples were analysed in triplicates.
Non-linear procedures (PROC NLIN) in SAS (2009) were used to estimate in vitro degradation kinetics in the rumen. Data were fitted into exponential model without lag time (Ørskov and McDonald 1979) to determine the rate constants and potential degradation according to the exponential model:
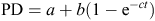
where PD is the potential degradability after time ‘t’, a is the soluble fraction (%; fraction washed out at t = 0; this value resulted from the incubation of 0 h bags, b is the insoluble but degradable fraction after time ‘t’, c is the rate of degradation of slowly degradable fraction b and t is the incubation time (h).
Effective degradability (EDDM; %) was calculated from the aforementioned parameters (a, b and c) assuming a fractional passage rate (kp) of 5%/h for the forage diets and 8%/h for the PEL:
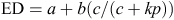
Statistical analyses
All the data on degradation, energy fractions, nutrient intake from milk and solid feed, and growth were subjected to analysis of variance using GLM procedures in Minitab Statistical Software, Version 17 (Minitab 2010) and covariance analysis was done using initial bodyweight of the calves as covariate. Treatment means were compared using a Fishers’ least significant difference and significant differences were declared at P < 0.05. Fitted non-linear regression degradation parameters and ED were analysed as completely randomised design with diet as a factor using analysis of variance procedures in SAS (2009).
Results and discussions
All the diets had similar crude protein contents (P > 0.05). However, diets containing LLM tended to have higher crude protein contents, which were not statistically significant at 5% level. The minimum crude protein required to support rumen bacterial growth for dairy calves is 18% (NRC 2001; Drackley et al. 2002). All the diets in the present study were above the minimum crude protein requirements. Bach (2014) noted that a starter feed above 20% crude protein positively influences growth of the calves after weaning. The neutral detergent fibre contents across all dietary treatments were higher than the recommended neutral detergent fibre content for dairy calf starter of 15–25% (Davis and Drackley 1998), depending on the digestibility of the feed. The LLM fibre in the present was highly degradable and, therefore, would not impact on nutrient and hence availability. The diets were also low in acid detergent lignin which, according to Goering and Van Soest (1970) and Buxton et al. (1996), would not negatively affect digestibility. There were no differences observed in acid detergent insoluble crude protein in both dietary treatments. Sniffen et al. (1992) reported that acid detergent fibre-bound N represents the portion of protein in a feedstuff that is unavailable for use by the animal because it is completely indigestible. Additionally, Van Soest (1994) stated that as the concentration of acid detergent fibre-bound N increases, total N digestibility decreases. This negative association has been observed in several studies (Weiss et al. 1986; Van Soest 1994). In addition, an increase in inclusion of LLM in the diets tended to decrease neutral detergent fibre and acid detergent lignin contents of the diets. Therefore, there would be more ruminal microbe attachment to the feed particles, as well as an increase in microbial growth and enzyme activity or intestinal activity (McAllister et al. 1994; McSweeney et al. 2001).
In vitro DM digestibility estimates increased with incubation time for all diets and no differences (P > 0.05) were observed during the 48 h of incubation (Table 4). The higher In vitro DM digestibility in the LLM substituted diets may probably be due to more cell contents in the diets, which are almost completely digestible and are not affected by lignin concentration in cell walls (Van Soest and Moore 1965). Greater organic matter degradation observed with LLM substituted diets at 24 h incubation may result from an increase in microbial protein synthesis (Verbic and Babnik 1997). This is based on the view that microbial protein synthesis is highly dependent on the availability of the rumen degradable organic matter. It has been reported that the rate of N and carbohydrates degradation, especially from the mixture of forage and concentrates, increases the efficiency of microbial protein synthesis due to an improved rumen environment (Karshi and Russell 2001). In vitro neutral detergent fibre and crude protein degradation values were higher in LLM substituted diets at 24 and 48 h than Diet PEL. LLM substituted diets had higher effective degradability values than Diet PEL. Concentrate feeding for calves has risk of acidosis (Nocek 1997). However, the results of the present study showed that LLM substituted diets were more digestible in the rumen compared with PEL. Thus, LLM may provide more digestible energy, but have higher risk of acidosis compared with PEL.

|
The predicted metabolisable energy densities of the diets during the neonatal phase were similar and within the range of 2.9–3.2 Mcal/kg DM as recommended by NRC (1989) for dairy calves (Table 3). These results are similar to those reported by Chester-Jones and Broadwater (2009) with a mean of 2.7 Mcal/kg DM and are within the recommendations by NRC (1989). Higher total digestible nutrients, net energy at maintenance and net energy at gain were observed with LLM substituted diets compared with diet PEL.
Average daily solid feed consumption during the neonatal phase was lower than the 680 g per calf recommended by NRC (2001) at ~41–42 days of age (Table 2). However, higher (P < 0.05) intakes of DM, crude protein, neutral detergent fibre and acid detergent fibre were observed in calves fed Diet P50 L50 during neonatal and transition phases and this may be ascribed to the differences in ruminal development and thus the calf’s metabolic and structural capacity to accommodate and digest solid feed (Khan et al. 2007). However, higher starch intake by calves on Diet PEL might be due to high starch content of the feed (13.4%) presented in Table 1. In addition, the higher average daily crude protein and neutral detergent fibre intakes during neonatal phase in calves fed Diet P50 L50 were the function of higher starter feed consumption. These findings are supported by higher average daily gain and better feed conversion ratio observed in calves on Diet P50 L50. However, Quigley et al. (2000) reported higher feed conversion ratio values of 3.10 in Holstein calves fed on a calf starter ration.
During the neonatal and transition phase, the DM and crude protein intakes were within the recommended range for the dairy calves as reported by Van Amburgh and Drackley (2005); however, a lower (P < 0.05) crude protein intake was observed with calves fed a 50% LLM inclusion during the transition phase. Total DM intake and feed conversion ratio for the solid feed during the transition phase did not differ across treatments (P > 0.05). It was reported that feeding a fibre source to young dairy calves was necessary because it improved rumen health (Thomas and Hinks 1982). This is also supported by recent literature that inclusion of forage in the diet of the calves during the pre-weaning period improved rumen health (Khan et al. 2011; Castells et al. 2012, 2013; Montoro et al. 2013). The higher (P < 0.05) average daily gain observed for LLM substituted diets than calf starter concentrate pellets (PEL) during the pre-weaning phase could be attributed to a better development of the rumen. This higher average daily gain observed during neonatal phase may be an indication that calves would reach puberty by 12 months and bred to calve by 22 months. Kertz et al. (1979) stated that high starter intake before weaning helps to ensure intake and sustain a desirable growth rate after weaning which improves the age at first calving. The results of the present study, indicate greater growth rate with LLM inclusion and they are higher than the target growth rate of 0.7 kg for Holstein for the first 12-week period recommended, as reported by (Dawson 2006).
Calves on LLM diets had higher (P < 0.05) DM intakes and average daily gain. The higher DM intake of lucerne results in higher protein intakes hence saving of protein concentrates (Kirilov 2001). The PEL diet had lower (P < 0.05) RNB values than LLM substituted diets. This is a serious hindrance for early growth as calves require more protein for muscle accretion and immune development. The slower growth for calves on this diet confirmed these observations. During the transition phase, a higher (P < 0.05) RNB was observed in calves on Diet P50 L50 and this indicates opportunities to improve the efficiency of N utilisation by rumen microbes. However, a lower value (P < 0.05) of RNB observed with Diet PEL indicates lower rumen fermentation, lower microbial yield and insufficient rumen-degradable N relative to carbohydrate supply (Maglione and Russell 1997). Ruminal N limitation can decrease microbial flow from the rumen (NRC 1985; Russell et al. 1992), depress fibre fermentation (Russell et al. 1992) and reduce DM intake (NRC 1987; Van Soest 1994).
Chester-Jones and Broadwater (2006) reported the highest average daily calf starter intakes of 0.09, 0.51, 1.12 and 2.18 kg for periods 1–14, 15–28, 29–42 and 43–56 days, respectively, for Holstein heifer calves. These results are higher than those of the present study at 21–42 and 43–56 days with intake values of 0.91–1.03 and 1.29–1.34 kg/calf.day, respectively. The higher the quality, the faster the rate of digestion and the higher the potential intake of the LLM.
Conclusions
The results of the present study indicate that nutrient composition, effective degradability of DM and energy density improved with higher LLM inclusion in the calf diet. This was confirmed by improved diet intake, % ruminal N balance and growth of Holstein heifer calves on LLM substituted diets. However, there is a need for further studies to investigate rumen microbial dynamics of a rapidly degradable LLM, production of volatile fatty acids and determine the effects of LLM on rumen papillae development and the outflows of microbial protein to the lower gut of neonates fed higher proportions of LLM.
Acknowledgements
We thank the Agricultural Research Council (ARC) Dairy Section staff members who were involved in the feeding trials and ARC-Professional Development Program for financial support towards this study.
References
Association of Analytical Chemists (AOAC) (2000) ‘Official methods of analysis.’ 17th edn. (Association of Official Analytical Chemists: Washington, DC)Association of Analytical Chemists (AOAC) (2006) ‘Official methods of analysis.’ 18th edn. (Association of Official Analytical Chemists: Washington, DC)
Bach A (2014) Effective forage and starter feeding strategies for pre-weaned calves. Advances in Dairy Technology 26, 153–163.
Buxton DR, Mertens DR, Fisher DS (1996) Forage quality and ruminant utilisation. In ‘Cool-season forage grasses’. (Eds LE Moser, DR Buxton, MD Casler) pp. 229–266. (ASA, CSSA, and SSSA: Madison, WI)
Castells L, Bach A, Araujo G, Montoro C, Terre M (2012) Effect of different forage sources on performance and feeding behaviour of Holstein calves. Journal of Dairy Science 95, 286–293.
| Effect of different forage sources on performance and feeding behaviour of Holstein calves.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3MXhs1OlsrjL&md5=84f8ed248fbb49255ea221c39fa1fefbCAS |
Castells L, Bach A, Arias A, Terre M (2013) Effects of forage provision to young calves on rumen fermentation and development of the gastrointestinal tract. Journal of Dairy Science 96, 5226–5236.
| Effects of forage provision to young calves on rumen fermentation and development of the gastrointestinal tract.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3sXot1KlsLc%3D&md5=d1065a473ba09b85a2b6bae9faa8ee7dCAS |
Cherney DJR, Cherney JH, Chase LE (2004) Lactation performance of Holstein cows fed fescue, orchard grass, or alfalfa silage. Journal of Dairy Science 87, 2268–2276.
| Lactation performance of Holstein cows fed fescue, orchard grass, or alfalfa silage.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2cXlvFKlu70%3D&md5=9617e567eb95597f10fe11efc9798356CAS |
Chester-Jones H, Broadwater N (2006) Research using standard diets for dairy calves result in very good performance to two months of age. Dairy Star Volume 8, No. 15, September 23, 2006, Sauk Center, MN and University of Minnesota Extension Dairy Connection (www.extension.umn.edu/dairy), University of Minnesota Extension Service, Saint Paul, MN.
Chester-Jones H, Broadwater N (2009) ‘Calf starters.’ (University of Minnesota Dairy Extension Service: Saint Paul, MN) pp. 1–40.
Davis CL, Drackley JK (1998) ‘The development, nutrition, and management of the young calf.’ (Iowa State University Press: Ames, IA)
Dawson LER (2006) Beef production from the dairy herd. Occasional Publication, Beef Open Day, Sustainable Beef Production, Charting the Way Forward, Oak Park, Carlow.
Drackley JK, Bartlett KS, Blome RM (2002) Protein content of milk replacers and calf starters for replacement calves. Available at http://www.livestocktrail.uiuc.edu/dairynet/paperDisplay.cfm?ContentID=339 [Verified 22 August 2014]
Fox DG, Tedeschi LO, Tylutki TP (2004) The Cornell Net Carbohydrate and Protein System model for evaluating herd nutrition and nutrient excretion. Animal Feed Science and Technology 112, 29–78.
| The Cornell Net Carbohydrate and Protein System model for evaluating herd nutrition and nutrient excretion.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2cXmvFymsQ%3D%3D&md5=146b04f26de9a677bb67d7a8c43e7af4CAS |
Giron HC (1973) Comparison between dry ashing and wet absorption analysis. Atomic Absorption Newsletter 12, 28–29.
Goering HK, Van Soest PJ (1970) ‘Forage fibre analyses (apparatus, reagents, procedures, and some applications).’ Agricultural Handbook 379. (ARS, USDA: Washington, DC)
Göncü S, Boğac M, Kiliç Ü, Görgülü M, Doran F (2010) Effects of feeding regime without roughage on performances and rumen development of calves during pre-weaning period. The Journal of Agricultural Science 16, 123–128.
Guenthner E, Carlson CW, Olson OE, Kohler GO, Livingstone AL (1973) Pigmentation of egg yolks by xanthophylls from corn, marigold, alfalfa and synthetic sources. Poultry Science 52, 1787–1798.
| Pigmentation of egg yolks by xanthophylls from corn, marigold, alfalfa and synthetic sources.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaE2cXlvVOnsQ%3D%3D&md5=d529a285a023f47f31ce5318c89393c2CAS |
Hall MB (2000) Neutral detergent soluble carbohydrates – nutritional relevance and analysis. Bulletin number 339, University of Florida., Gainesville.
Holm J, Bjorck I, Drews A, Asp NG (1986) A rapid method for the analysis of starch. Starch 38, 224–226.
| A rapid method for the analysis of starch.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaL28XltFekt7s%3D&md5=5da965b3f80fb09e38dd313ba51d0cebCAS |
Homolka P, Koukolova V, Němec Z, Mudřik Z, Hučko B, Sales J (2008) Amino acid contents and intestinal digestibility of lucerne in ruminants as influenced by growth stage. Czech Journal of Animal Science 53, 499–505.
Homolka P, Koukolova V, Podsedniček M, Hlavačkova A (2012) Nutritive value of red clover and lucerne forages for ruminants estimated by in vitro and in vivo digestibility methods. Czech Journal of Animal Science 57, 454–568.
Jones C, Heinrichs J (2007) Early weaning strategies. Available at http://www.extension.org/pages/Early_Weaning_Strategies [Verified 7 October 2015]
Karshi MA, Russell JB (2001) Effects of some dietary factors on ruminal microbial protein synthesis. Turkish Journal of Veterinary and Animal Sciences 25, 681–686.
Kertz AF, Prewitt LR, Everett JP (1979) An early weaning calf program: summarization and review. Journal of Dairy Science 62, 1835–1843.
| An early weaning calf program: summarization and review.Crossref | GoogleScholarGoogle Scholar |
Khan MA, Lee HJ, Lee WS, Kim HS, Kim SB, Ki KS, Park SJ, Ha JK, Choi YJ (2007) Starch source evaluation in calf starter: I. feed consumption, body weight gain, structural growth, and blood metabolites in Holstein Calves. Journal of Dairy Science 90, 5259–5268.
| Starch source evaluation in calf starter: I. feed consumption, body weight gain, structural growth, and blood metabolites in Holstein Calves.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2sXht1Kru73E&md5=54d9c5d2051e4c20ff474a15103178d8CAS |
Khan MA, Weary DM, Keyserlingk MAGV (2011) Hay intake improves performance and rumen development of calves fed higher quantities of milk. Journal of Dairy Science 94, 3547–3553.
| Hay intake improves performance and rumen development of calves fed higher quantities of milk.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3MXnvFahsLk%3D&md5=63a494240cb1c2fadadb6416e8600b76CAS |
Kirilov A (2001) Lucerne quality and possibilities for its estimation. In ‘Quality in lucerne and medics for animal production’. (Eds I Delgado, J Lloveras) pp. 231–234. (Options Méditerranéennes: Série A. Séminaires Méditerranéens: Zaragoza, Spain)
Kuan KK, Stanogias G, Dunkin AC (1983) The effect of proportion of cell-wall material from lucerne leaf meal on apparent digestibility, rate of passage and gut characteristics in pigs. Animal Production 36, 201–209.
Licitra G, Hernandez TM, Van Soest PJ (1996) Standardization of procedures for nitrogen fractionation of ruminant feeds. Animal Feed Science and Technology 57, 347–358.
| Standardization of procedures for nitrogen fractionation of ruminant feeds.Crossref | GoogleScholarGoogle Scholar |
Lindberg JE, Cortova Z, Thomke S (1995) The nutritive value of lucerne leaf meal for pigs based on digestibility and nitrogen utilisation. Journal of Animal Science 45, 245–251.
Maglione G, Russell JB (1997) The adverse effect of nitrogen limitation and excess-cellobiose on Fibrobacter succinogenes S85. Applied Microbiology and Biotechnology 48, 720–725.
| The adverse effect of nitrogen limitation and excess-cellobiose on Fibrobacter succinogenes S85.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaK1cXht1altb8%3D&md5=b4febc05fcc2871dd760ba08f4d2ec71CAS |
Mason S (1998) Alfalfa protein. Alberta Dairy Management 1A1, 1–2.
McAllister TA, Bae HD, Jines GA, Cheng KJ (1994) Microbial attachment and feed digestion in the rumen. Journal of Animal Science 72, 3004–3018.
McSweeney CS, Palmer B, McNeill DM, Krause DO (2001) Microbial interactions with tannins: rrnutritional consequences for ruminants. Animal Feed Science and Technology 91, 83–93.
| Microbial interactions with tannins: rrnutritional consequences for ruminants.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD3MXjslSms7Y%3D&md5=e0b2580344353d7e20a142cda4713e12CAS |
Minitab (2010) ‘Minitab statistical software version 17.’ (Minitab, Inc.: State College, PA). Available at www.minitab.com [Accessed 10 May 2014]
Montoro C, Miller-Cushon EK, De TJ, Vries TJ, Bach A (2013) Effect of physical form of forage on performance, feeding behavior, and digestibilities in Holstein calves. Journal of Dairy Science 96, 1117–1124.
| Effect of physical form of forage on performance, feeding behavior, and digestibilities in Holstein calves.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC38Xhsl2ju7bN&md5=08ac66659686b1d29c88e92b1f180bb4CAS |
Mujumdar AS (1997) Drying fundamentals. In ‘Industrial drying of foods’. (Ed. CGJ Baker) pp. 7–30. (Chapman and Hall: London)
National Research Council (NRC) (1985) ‘Ruminant nitrogen usage.’ (National Academy Press: Washington, DC)
National Research Council (NRC) (1987) ‘Predicting feed intake of food producing animals.’ (National Academy Press: Washington, DC)
National Research Council (NRC) (1989) ‘Nutrient requirements of dairy cattle.’ 6th revised edn. (National Academy Press: Washington, DC)
National Research Council (NRC) (2001) ‘Nutrient requirements of dairy cattle.’ 7th revised edn. (National Academy Press: Washington, DC)
Nocek JE (1997) Bovine acidosis: implications on Laminitis. Journal of Dairy Science 80, 1005–1028.
| Bovine acidosis: implications on Laminitis.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaK2sXjsVOkt74%3D&md5=1afbb0d47cd15d37cc7db3aa57e594a0CAS |
Ørskov ER, McDonald Y (1979) The estimation of protein degradability in the rumen from determining the digestibility of feeds in the rumen. Journal Agricultural Science 92, 499–503.
Quigley JD, Jaynes CA, Miller ML, Schanus E, Chester-Jones H, Marx GD, Allen DM (2000) Effects of hydrolyzed spray dried red blood cells in milk replacer on calf intake, body weight gain and efficiency. Journal of Dairy Science 83, 788–794.
| Effects of hydrolyzed spray dried red blood cells in milk replacer on calf intake, body weight gain and efficiency.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD3cXivFWlt7Y%3D&md5=cdd99c9ce45e3e1ef521478fbc8b4000CAS |
Rechulicz J, Katarzyna Ognik K, Grela ER (2014) The effect of adding protein-xanthophylls concentrate (PX) from lucerne (Medicago sativa) on growth parameters and redox profile in muscles of carp, Cyprinus carpio (L.). Turkish Journal of Fisheries and Aquatic Sciences 14, 697–703.
| The effect of adding protein-xanthophylls concentrate (PX) from lucerne (Medicago sativa) on growth parameters and redox profile in muscles of carp, Cyprinus carpio (L.).Crossref | GoogleScholarGoogle Scholar |
Russell JB, O’Connor JD, Fox DG, Van Soest PJ, Sniffen CJ (1992) A net carbohydrate and protein system for evaluating cattle diets: I. Ruminal fermentation. Journal of Animal Science 70, 3551–3561.
| A net carbohydrate and protein system for evaluating cattle diets: I. Ruminal fermentation.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaK3sXnvF2iug%3D%3D&md5=41ceaab8592091aad760c36ffe3883ecCAS |
SAS (Statistical Analysis System) (2009) ‘User’s guide: statistics, Version 9.2.’ (SAS Institute, Inc.: Cary, NC)
Smit CJ (2014) Effects of sweet potato forage meals on protein and energy supply, beta-carotene and blood glucose content of dairy cattle milk. Master Thesis, University of South Africa., Pretoria.
Sniffen CJ, O’Connor JD, Van Soest PJ, Fox DG, Russell JB (1992) A net carbohydrate and protein system for evaluating cattle diets: II carbohydrate and protein availability. Journal of Dairy Science 70, 3562–3577.
Thomas DB, Hinks CE (1982) The effect of changing the physical form of roughage on the performance of the early-weaned calf. Animal Production 35, 375–384.
| The effect of changing the physical form of roughage on the performance of the early-weaned calf.Crossref | GoogleScholarGoogle Scholar |
Tufarelli V, Cazzato E, Ficco A, Laudadio V (2010) Evaluation of chemical composition and in vitro digestibility of Appennine pasture plants using Yak rumen fluid or faecal extracts as inoculum source. Asian-Australasian Journal of Animal Sciences 23, 1587–1593.
| Evaluation of chemical composition and in vitro digestibility of Appennine pasture plants using Yak rumen fluid or faecal extracts as inoculum source.Crossref | GoogleScholarGoogle Scholar |
Van Amburgh ME, Drackley JK (2005) Current perspectives on the energy and protein requirements of the pre-weaned calf. In ‘Calf and heifer rearing: principles of rearing the modern dairy heifer from calf to calving’. (Ed. PC Garnsworthy) Chapter 5, pp. 67–82. (Nottingham University Press: UK)
Van Soest PJ (1994) ‘Nutritional ecology of the ruminant.’ 2nd edn. (Cornell University Press: Ithaca, NY)
Van Soest PJ, Moore LA (1965) New chemical methods for analysis of forage for the purpose of predicting nutritive value. In ‘Proceedings of the 9th International Grassland Congress’. pp. 783–789. (The Congress: Sao Paulo, Brazil)
Van Soest PJ, Roberston JB, Lewis BA (1991) Methods for dietary fibre, neutral detergent fibre and non-starch polysaccharides in relation to animal nutrition. Journal of Dairy Science 74, 3583–3597.
| Methods for dietary fibre, neutral detergent fibre and non-starch polysaccharides in relation to animal nutrition.Crossref | GoogleScholarGoogle Scholar | 1:STN:280:DyaK38%2FnvVCltA%3D%3D&md5=5ac74045d70ec4fdb1b477d24fb39c1dCAS |
Verbic J, Babnik D (1997) Evaluation of the protein supply in ruminants 3. The proposal of a system for Slovenia. Sodobno Kmetijstvo 30, 147–197.
Weiss WP, Conrad HR, Shockey WL (1986) Digestibility of nitrogen in heat damaged alfalfa. Journal of Dairy Science 69, 2658–2670.
| Digestibility of nitrogen in heat damaged alfalfa.Crossref | GoogleScholarGoogle Scholar | 1:STN:280:DyaL2s7htVSqug%3D%3D&md5=87459d93936aa77ba7e0f040217d7186CAS |
Xie Z, Huang J, Xu X, Jin Z (2008) Antioxidant activity of peptides isolated from alfalfa leaf protein hydrolysate. Food Chemistry 111, 370–376.
| Antioxidant activity of peptides isolated from alfalfa leaf protein hydrolysate.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD1cXnsFehtbg%3D&md5=4dc0527cc609e82deb726a45ff018cbbCAS |