

Low-phosphorus conditions affect the nitrogen nutrition and associated carbon costs of two legume tree species from a Mediterranean-type ecosystem
Anathi Magadlela A , Aleysia Kleinert A , Léanne L. Dreyer A and Alex J. Valentine A BA Botany and Zoology Department, University of Stellenbosch, Private Bag X1, Matieland 7602, South Africa.
B Corresponding author. Email: alexvalentine@mac.com
Australian Journal of Botany 62(1) 1-9 https://doi.org/10.1071/BT13264
Submitted: 29 October 2013 Accepted: 11 February 2014 Published: 14 April 2014
Abstract
The role of phosphorus nutrition in two-legume tree species from the Mediterranean-type ecosystem of the Cape Floristic Region (CFR) in South Africa was investigated. There is very little information about the functional adaptations of nitrogen (N) and phosphorus (P) nutrition in these legume trees growing in nutrient-poor soils. Nodulated Virgilia divaricata and V. oroboides tree saplings were grown in sterilised sand and supplied with Long Ashton nutrient solution, which was modified to contain either sufficient-phosphate (500 µM) or low-phosphate (5 µM) nutrient solution for 90 days. During low-P conditions, the growth of V. divaricata was not affected, whereas V. oroboides showed a decrease in growth. The decrease in V. oroboides under low-P conditions was related to the lower P uptake, which resulted in an alteration in belowground biomass allocation, which consequently affected on the N nutrition and carbon (C) cost of growth. In this regard, V. oroboides plants allocated less biomass to roots and nodules, as a proportion of whole plant growth. The impact of this was a decline in N nutrition, growth respiration and photosynthetic costs in V. oroboides. In contrast, V. divaricata maintained its P concentrations, photosynthetic costs and increased its nodule allocation under low-P conditions, to the benefit of N nutrition. The two CFR tree legumes appear to have different adaptations to low-P conditions, which may influence their N and P acquisition in their naturally low-P environment.
Additional keywords: acidic soils, biological nitrogen, fynbos, nutrient deficiencies fixation, Virgilia.
Introduction
The Mediterranean-type ecosystem of the Cape Floristic Region (CFR) in South Africa mainly grows on sandstone-derived soils (Goldblatt and Manning 2000), which are typically very acidic and nutrient poor (Kruger et al. 1983). The soils of this region bear a resemblance to the soils of the Western Australian heathlands rather than to those of other Mediterranean-climate regions (Groves 1983; Mitchell et al. 1984. Virgilia is one of tree legume genera that are endemic to these CFR (fynbos) acidic soils. Mediterranean acidic soils usually have different concentrations of elements, coupled with related nutrient deficiencies, associated with nitrogen (N) and phosphorus (P) deficiencies (Bordeleau and Prevost 1994; von Uexkull and Mutert 1998; Grigg et al. 2008). Soil acidity is a significant problem facing legume and agricultural production in many areas of the world, including southern Africa (Graham 1992; Bordeleau and Prevost 1994; Marschner 1995; Correa and Barneix 1997). Most legume plants require neutral to slightly acidic soils for growth, but experience problems with nodulation if the pH drops to a very acidic state (Lie 1981; Munns 1986; Graham 1992; Bordeleau and Prevost 1994; Marschner 1995; Correa and Barneix 1997). Both soil acidity and nutritional disorder adversely affect the survival, growth and N fixation of microorganisms because they affect legume–rhizobia symbiosis (Lie 1981; Munns 1986; Graham 1992). P remains mostly unavailable for plant uptake, specifically on acid-weathered soils in both tropical and subtropical regions (Bieleski 1973; Schachtman et al. 1998; von Uexkull and Mutert 1998; Vance 2001).
Phosphate is quite abundant in many soils, but because it forms insoluble complexes with cations (calcium and iron) and is bound to organic compounds by microbial action, it is often unavailable for plant uptake (Jungk et al. 1993; Richardson 1994; Hinsinger 2001; Dakora and Phillips 2002; Vance 2001). Low-P soils will limit legume growth to a greater extent than do low-N soils, because during symbiosis with rhizobia, legumes can utilise both atmospheric N and soil N acquired through rhizobia in their nodules (Mortimer et al. 2008). It has been reported that host legume nodules require comparatively high amounts of P and energy; thus, P deficiency can impair both nodulation and symbiotic N fixation, affecting photosynthesis, respiration, growth, organic-acid supply and production of the host (Drevon and Hartwig 1997; Vadez et al. 1997; Almeida et al. 2000; Olivera et al. 2004; Lynch and Ho 2005; Harrison et al. 2009). Despite these complications, legume species endemic to fynbos preferentially grow specifically in such acidic soils. They must, therefore, have evolved adaptations to function optimally under these limiting P conditions (Vance et al. 2003). Some strategies are aimed at conserving the use of P, whereas others are directed toward enhanced acquisition and uptake of P (Lajtha and Harrison 1995; Raghothama 1999; Horst et al. 2001; Vance 2001). Adaptations that conserve the use of P involve a decrease in growth rate, increased growth per unit of P uptake, remobilisation of internal inorganic P (Pi), modification in carbon (C) metabolism that bypass P-requiring steps and alternative respiratory pathways (Schachtman et al. 1998; Plaxton and Carswell 1999; Raghothama 1999; Uhde-Stone et al. 2003a, 2003b). In legumes, adaptations leading to enhanced P acquisition entail the expression of genes that result in the production of cluster roots. Cluster roots increase the root surface area. This enhances nodule efficiency for P utilisation (Le Roux et al. 2009), root exudation of organic acids and acid phosphatase, as well as the induction of numerous transporters (Gilbert et al. 1999; Gilroy and Jones 2000; Lynch and Brown 2001; Neumann and Martinoia 2002; Lamont 2003; Uhde-Stone et al. 2003a; Vance et al. 2003). The exudation of organic acids such as malate and citrate stimulated by P stress has mostly been reported in non-mycorrhizal species such as lupin (Dinkelaker et al. 1995; Jones 1998; Hinsinger 2001; Ryan et al. 2001; Le Roux et al. 2008).
The high sensitivity of legume plants, and indeed the N2-fixation process to environmental conditions such as acidic soils associated with P deficiency, may result in higher C costs (Mengel 1994). This concurs with Le Roux et al. (2009), who showed that lupin nodules under P stress acted as stronger C sinks. Nodules are known to have a strong sink capacity for P assimilation during P starvation (Hogh-Jensen et al. 2002). The enhanced nodule cost for P utilisation is considered to be an essential coping strategy during P stress (Le Roux et al. 2009). The C sink was found to be more pronounced in plants during double symbiosis under low-P conditions (Mortimer et al. 2008). This was shown by a greater growth respiration of low-P plants than high-P plants (Mortimer et al. 2008). The sink effect was also evidenced by the higher photosynthetic rates of host plant (Mortimer et al. 2008). In the case of P stress, the most direct currency is P itself or growth parameters related to P accumulation (Koide et al. 2000).
Virgilia is a small tree genus that includes two species (V. divaricata (Adamson) and V. oroboides (P.J.Bergius) Salter) and two subspecies. It is confined to the south-western and southern coastal regions of the CFR (Greinwald et al. 1989). Studies have been conducted on growth and adaptations of legume species native to Mediterranean-type fynbos ecosystems that occur on naturally acidic soils (Muofhe and Dakora 1999; Spriggs and Dakora 2008; Power et al. 2010; Kanu and Dakora 2012). However, information on the physiology of N and P uptake, efficiency and utilisation in legume trees in fynbos soils is largely unknown.
The mechanisms of legume adaptations to acquire N from both the atmosphere and soil in a low-P environment will assist in the understanding of the distribution of these trees in their native environment of the CFR. The aim of the present study was, therefore, to investigate how low P nutrition affects the N acquisition pathways and their associated costs in two indigenous fynbos legumes, V. divaricata and V. oroboides.
Materials and methods
Plant material and growth conditions
Seeds of both V. oroboides and V. divaricata were obtained from Kirstenbosch Botanical Gardens, Cape Town, South Africa, and scarified using an acid-scarification method that entails soaking the seeds in sulfuric acid (H2SO4) for 30 min and then rinsing them 10 times in distilled water. Hereafter, seeds were treated overnight with smoke water, also obtained from Kirstenbosch. The seeds were germinated in natural fynbos soil (natural inoculation), obtained from Stellenbosch Mountain, Stellenbosch, South Africa. Plants were grown under glasshouse conditions at the University of Stellenbosch, South Africa, and they were watered daily. The range of midday irradiances was between 600 and 800 µmol m–2 s–1 and the average night and day temperature ranges were 15−25°C. After 40 days, the plants were transferred to clean sand and initially watered with distilled water for a week to acclimatise. Hereafter, seedlings were supplied with low N (500 µM), quarter-strength Long Ashton nutrient solution modified to sufficient P (500 µM) and low P (5 µM) (pH 5.8) once a week and watered with distilled water in between nutrient-solution supply. The experiment was split between the two species, so that each species was subjected to low- and sufficient-P treatments. The combination of species and P concentrations resulted in four treatments, with eight replicates each.
Harvesting and nutrient analysis
Seedlings were harvested at 90 days after transplanting into the sand culture. On harvesting, the plants were separated into nodules, roots, stems and leaves. The harvested plant material was placed in a drying oven at 40°C for 3 days, and the dry weights (DWs) of plant parts were recorded. The dried material was milled with a ball mill. The milled samples were analysed for their respective C, N and P concentrations by a commercial laboratory, using inductively coupled plasma–mass spectrometry (ICP-MS) and a LECO-nitrogen analyser with suitable standards (BemLab, De Beers Road, Somerset West, South Africa).
Gas-exchange measurements
The photosynthetic response to varying levels of light was determined to measure maximum photosynthetic rate (Pmax) on a light-response curve. Measurements were performed on the youngest fully expanded leaves (5 replicates in each treatment per species), using an open gas-exchange system Li-6400 (LI-COR, IRGA, Lincoln, NE, USA). Measurements were taken from 0900 hours to 1600 hours. A full light-response curve took 60–90 min to complete. The leaves were enclosed in a leaf chamber and at a leaf temperature of 24°C, the light was varied from 0, 50, 100, 150, 200, 250, 350, 500, 650, 800, 900, 1000, 1500 to 2000 µmol m–2 s–1.
Carbon and nutrition cost calculations
-
Construction costs, CW (mmol C g–1 DW), were calculated according to the methods of Mortimer et al. (2005), modified from the equation used by Peng et al. (1993), as follows:
where CW is the construction cost of the tissue (mmol C g–1 DW), C is the carbon concentration (mmol C g–1), k is the reduction state of the N substrate (k = –3 for NH3) and N is the organic-N content of the tissue (g DW–1) (Williams et al. 1987). The constant (1/0.89) represents the fraction of the construction costs that provides reductant that is not incorporated into the biomass (Williams et al. 1987; Peng et al. 1993) and (6000/180) converts units of g glucose per DW to mmol C per g DW.
-
Specific N-absorption rate (SNAR; mg N g–1 root DW day–1) is the calculation of the net N-absorption rate per unit root DW (Nielsen et al. 2001), as follows:
where M is the N content per plant, t is the time and R is the root DW.
-
Specific N-utilisation rate (SNUR; g DW mg–1 N day–1) is a measure of the DW gained for the N taken up by the plant (Nielsen et al. 2001), as follows:
where M is the N content and W is the plant DW.
-
Growth respiration Rg (t) (µmol CO2 day–1) is the daily growth respiration of the plant (Peng et al. 1993), as follows:
where Ct (µmol CO2 day–1) is the C required for daily construction of new tissue. Ct was calculated by multiplying the root growth rate (g DW day–1) by tissue construction cost (Cw). ΔWc (µmol C day–1) is the change in root C content and was calculated by multiplying the root C content and root growth rate.
-
Belowground allocation represents the fraction of new biomass partitioned into new roots and nodules over the given growth period. This was calculated according to Bazzaz (1997), as follows:
where RGR is the relative growth rate (mg g–1 day–1) and δ is the fraction of new biomass gained during the growth period. Br/Bt is the root weight ratio, based on total plant biomass (Bt) and root biomass (Br).
Calculations of percentage N derived from the atmosphere (%NDFA)
The δ15N analyses were carried out at the Archeometry Department, University of Cape Town, South Africa. The isotopic ratio of δ15N was calculated as δ = 1000‰ (Rsample/Rstandard), where R is the molar ratio of the heavier to the lighter isotope of the samples and standards are as described by Farquhar et al. (1989). Between 2.100 and 2.200 mg of each milled sample were weighed into 8 mm × 5-mm tin capsules (Elemental Micro-analysis, Devon, UK) on a Sartorius microbalance (Goettingen, Germany). The samples were then combusted in a Fisons NA 1500 (Series 2) CHN analyser (Fisons Instruments SpA, Milan, Italy). The δ15N values for the N gas released were determined on a Finnigan Matt 252 mass spectrometer (Finnigan MAT GmbH, Bremen, Germany), which was connected to a CHN analyser by a Finnigan MAT Conflo control unit. Three standards were used to correct the samples for machine drift, namely, two in-house standards (Merck Gel and Nasturtium) and the IAEA (International Atomic Energy Agency) standard (NH4)2SO4.
%NDFA was calculated according to Shearer and Kohl (1986), as follows:

where the reference plant was non-nodulated V. divaricata, planted 2 weeks later than the experimental plants and grown under the same glasshouse conditions using 500 μM N in a Long Ashton nutrient solution (25% strength). The B value is the δ15N natural abundance of the N derived exclusively from biological N-fixation of nodulated V. divaricata. The seeds were germinated in the natural inoculum and, thereafter, the seedlings were grown with N-free 25% strength Long Ashton nutrient solution in sterile-sand culture. The B value of V. divaricata was determined as –2.58.
Statistical analysis
The effects of the factors and their interactions were tested with an ANOVA (Kaleidagraph, Synergy Software, Reading, PA, USA). Where the ANOVA revealed significant differences among treatments, the means (6–8) were separated using post hoc Tukey’s l.s.d. (SuperAnova for Macintosh, Abacus Concepts, Berkeley, CA, USA) (P ≤ 0.05).
Results
Biomass
Under normal P conditions (HP), V. oroboides is evidently better adapted than is V. divaricata, because it had greater total biomass for the duration of the experiment (Table 1). The greater biomass resulted from the shoot DW, whereas root and nodule DWs were consistently lower for the duration of the experiment. There were no significant differences between the leaf areas (Table 1) of the two species. Under low-P conditions (LP), V. divaricata maintained a constant plant biomass, whereas V. oroboides showed a 38% decline in plant DW (Table 1). This decline resulted from a 70% decline in nodule DW, and a 63% decline in shoot DW, respectively. Low-P conditions may have inhibited the growth of nodules, resulting in a decline in biological N fixation. Furthermore, V. oroboides also showed an increase in its root : shoot ratio, whereas V. divaricata remained unchanged (Table 1).

|
P and N Nutrition
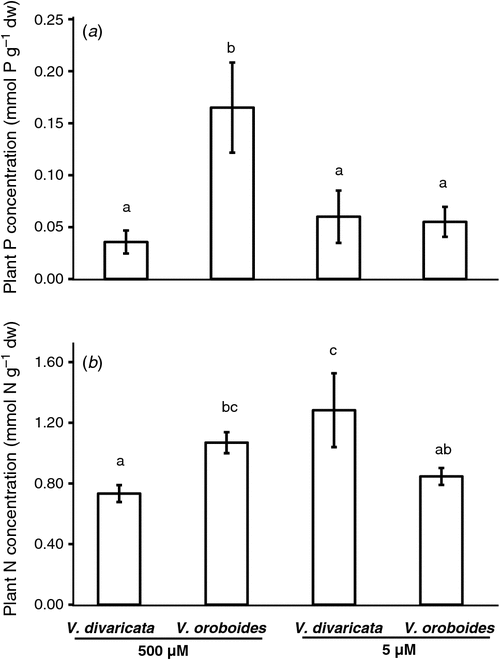
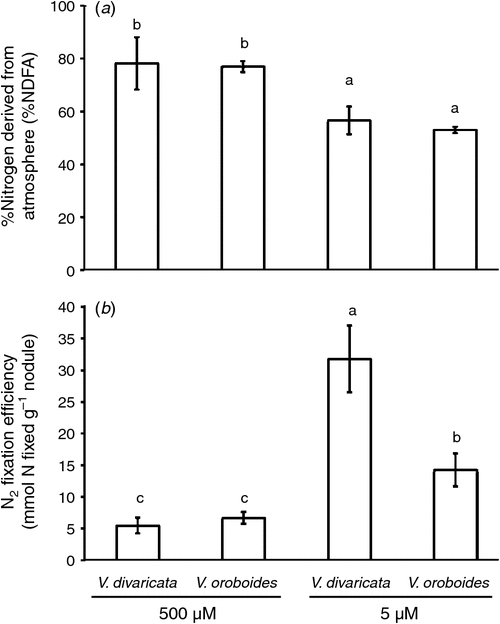
The P concentration (Fig. 1a) of plants exposed to adequate P concentrations followed a similar pattern as did the plant biomass. V. oroboides accumulated more P than did V. divaricata. The increased nodule DW of both Virgilia species and the sufficient plant P concentration of V. oroboides appear to favour biological N fixation (BNF), as shown by the increase in %NDFA (Fig. 2a). There was a significant difference in N uptake between the species during adequate P conditions. V. oroboides showed a greater SNAR than did V. divaricata (Table 1). This was also reflected by the higher plant N concentration (Fig. 2b). Specific N-utilisation rate was lower in V. oroboides than in V. divaricata under adequate P conditions (Table 1), suggesting that V. divaricata is more efficient at utilising N resources.
|
|
Phosphorus-stressed V. oroboides plants showed a decrease in P concentration, following a similar pattern of development of nodules, whereas V. divaricata maintained a constant P concentration. The decrease in P concentration of V. oroboides concurred with the decrease in %NDFA (Fig. 2a), which is an indicative decrease in BNF. Although V. divaricata maintained its P concentration during P stress, there was a decrease in %NDFA, whereas there was an increase in N concentration. This suggests that V. divaricata may be dependent on other N sources, e.g. soil N. In addition, V. oroboides had a lower specific N uptake (SNAR) under P stress than under adequate P conditions, but showed greater efficiency in N utilisation (SNUR) (Table 1). Although plants under P stress had a decreased BNF, they were more efficient in utilising atmospheric N per nodule, as indicated by the BNF-derived N per nodule (Fig. 2b).
Photosynthetic rate
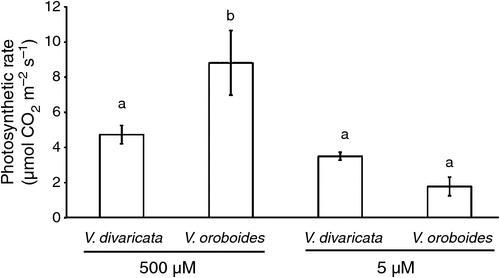
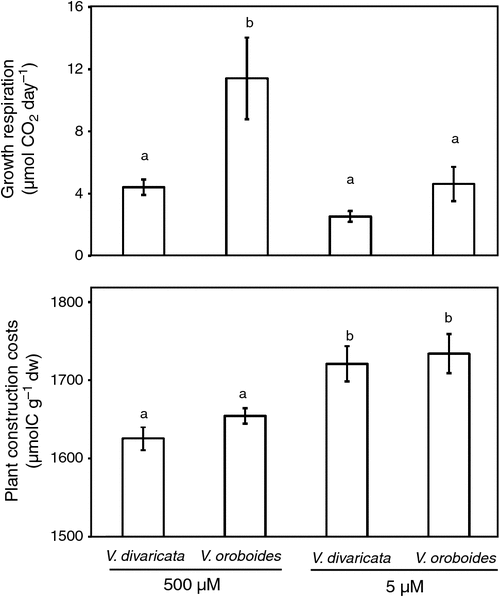
There was a significant difference in photosynthetic rate between the two species under sufficient P conditions. V. oroboides showed a higher photosynthesis rate (Fig. 3) to maintain its increased growth. The higher photosynthetic rate of V. oroboides was complemented by an increase in plant-growth respiration (Fig. 4a).
|
|
During P stress, V. oroboides had a decreased photosynthetic rate, shown by the significant difference between the treatments, whereas V. divaricata maintained its photosynthetic rate.
Respiratory carbon costs
Both species showed greater construction costs (Fig. 4b) during P stress. There was no significant difference in growth respiration (Fig. 4a) in V. divaricata during the two treatments. This may be because V. divaricata was able to maintain its DW and P nutrition during the experiment. In contrast, V. oroboides decreased growth respiration by 60% under P stress. During P stress, both plant species showed both decreased Rubisco and electron-transport activities (Fig. 4b, c) during photosynthesis.
Belowground allocation
During sufficient P supply, V. oroboides showed an increase in plant relative growth rate relative to V. divaricata (Fig. 5a). Under P stress, however, V. oroboides had a lower relative growth rate, whereas V. divaricata maintained its relative growth rate. Furthermore, V. oroboides increased its allocation to new root tissue during sufficient P supply, whereas there was no significant difference in both treatments in V. divaricata (Fig. 5b). In contrast, V. divaricata allocated more to new nodule-tissue growth during limited P supply than did V. oroboides (Fig. 5c).
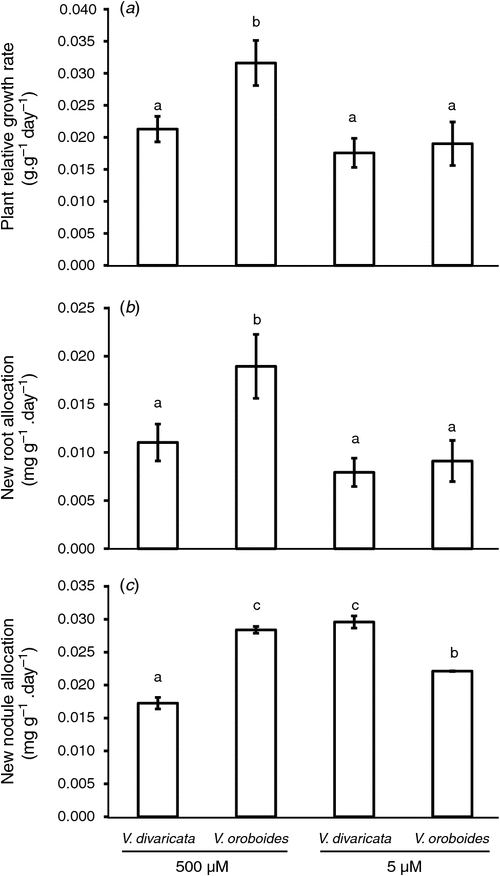
|
Discussion
The two tree legumes appear to have different adaptations to P starvation, in that during low-P supply, V. divaricata maintained its biomass relative to V. oroboides, by altering its biomass allocation to the belowground nutrient-acquiring structures.
The limited P supply reduced plant growth and may have limited the N demand and, consequently, BNF of the two legumes. This is consistent with results from previous studies where limited P nutrition reduced nodule growth, metabolism and N2 fixation and, subsequently, also plant growth (Sa and Israel 1991; Al Niemi et al. 1998; Almeida et al. 2000; Olivera et al. 2004; Hernández et al. 2007; Le Roux et al. 2008; Le Roux et al. 2009; Mortimer et al. 2009). The difference between these two indigenous legumes is that V. oroboides accumulated more P under P-sufficient conditions, which promoted nodular growth, BNF and SNAR. Consequently, it is likely that this improved BNF-derived N nutrition supported the increased biomass accumulation of V. oroboides, because it has been demonstrated in soybean that symbiotic N2 fixation requires more P for optimal functioning than for plant growth (Israel 1987). However, during P deficiency, V. divaricata appears to be physiologically better adapted for maintaining biomass and macro-element nutrition than is V. oroboides.
During P deficiency, the 48% decline in V. oroboides plant DW, coupled with a 64% decline in plant P concentration, concurs with results of previous work on P concentration in plants grown under P-deficient conditions (Hernández et al. (2007). Furthermore, the increase in the root : shoot ratio of V. oroboides during P deficiency represents a typical P-stress response of plants under P limitation (Rychter and Mikulska 1990; Juszczuk and Rychter 2002). This reduced growth was associated with a decline in the nodule DW, which concurs with previous findings (Gordon et al. 1990; Almeida et al. 2000; Tang et al. 2001; Høgh-Jensen et al. 2002). The decrease in biomass of V. oroboides may have reduced the reliance on BNF as a source of N for growth, as supported by findings of previous work (Sa and Israel 1991; Gordon et al. 1997; Almeida et al. 2000) where limiting P supply reduced the growth and nodule biomass, and caused a reduction in N demand and N2 fixation in the host plant. The higher N accumulation in V. divaricata under low-P conditions, in spite of similar reliance on BNF as for V. oroboides, suggested that V. divaricata may employ additional strategies for N acquisition under these conditions. One mechanism is the high specific N-absorption rate of the V. divaricata root system, which reflects the contribution of both roots and nodules. Another mechanism is the improved efficiency of BNF under low-P conditions in V. divaricata. In this regard, V. divaricata increased its allocation of resources to new nodule-tissue growth under P starvation, and also showed greater efficiency than did V. oroboides in acquiring atmospheric N2. It is known that enhanced nodule efficiency for nutrient utilisation is considered a pivotal coping strategy during P deficiency (Vadez et al. 1997; Raghothama 1999; Høgh-Jensen et al. 2002; Vance et al. 2003; Lynch and Ho 2005; Mortimer et al. 2008, 2009; Le Roux et al. 2009). However, enhanced nodule function may impose a cost on host reserves.
The increase in tissue-construction costs in both species under low P concurs with the findings of previous studies, which have shown an increase in C budget during low-P conditions (Lynch and Beebe 1995; Nielsen et al. 1998; Mortimer et al. 2008). This indicates that the belowground structures required more C per gram of tissue produced, although it is unclear what the precise structural nature of this additional C is (Mortimer et al. 2008). This increased construction did not appear to incur a large sink demand on photosynthetic products and instead it seems evident that growth respiration, representing the costs associated with the new tissue growth, had a larger sink effect. In V. oroboides, the higher photosynthetic rates at sufficient P were associated with the enhanced growth-respiration rates of the belowground nutrient-acquiring structures. The combined sink effect of the host plant roots and nodules can incur a drain of the host C reserves so as to maintain N and P nutrition. This may increase the photosynthetic rate of the host plant (Ainsworth et al. 2004; Kaschuk et al. 2009, 2010a, 2010b). However, under low-P conditions, the decline in photosynthesis in V. oroboides concurs with the lower investment in belowground growth, as evidenced by the decreased allocation and growth respiration of the root and nodule nutrient-acquiring structures. In contrast to this, the unchanged photosynthetic and growth-respiration rates of V. divaricata at both sufficient- and low-P treatments indicated an adaptation to low-P environments. In this regard, the increase in nodule allocation appears to have been achieved at the expense of root allocation. This trade-off has clearly benefited the efficiency of N nutrition of V. divaricata during low P supply.
Although the current study showed that legume performance in nutrient-poor soils may be dependent on different allocations of biomass to nutrient-acquiring structures such as root and nodules, other biological and biochemical factors may be equally important. In this regard, it is very likely that biological factors, such as mycorrhiza formation (Maseko and Dakora 2013), along with biochemical factors, such as the secretion of organic acids (Dakora and Phillips 2002) and acid phosphatases (Maseko and Dakora 2013), may also play important roles in nutrient poor soils.
Conclusions
During P starvation, V. divaricata saplings were better adapted than those of V. oroboides, by the alteration in biomass allocation to nodules and their improved efficiency of N acquisition and utilisation. The results of the present study suggested that V. divaricata is a more resilient tree, and would do well both in habitats with high- or low-P availability. This is interesting because V. oroboides trees are more specifically associated with P-poor CFR soils than is V. divaricata, the latter species often growing in richer P-nutrient soils. The implication of these findings is that the V. divaricata tree species may have a competitive advantage, if it should invade the more P-poor habitat of V. oroboides.
Acknowledgements
This work was funded by the DST/NRF–Center of Excellence for Tree Health and Biotechnology, based at the University of Pretoria. We acknowledge the Department of Botany and Zoology at the University of Stellenbosch for their research facilities.
References
Ainsworth EA, Rogers A, Nelson R, Long SP (2004) Testing the ‘source–sink’ hypothesis of down-regulation of hypothesis in elevated [CO2] in the field with single gene substitutions in Glycine max. Agricultural and Forest Meteorology 122, 85–94.| Testing the ‘source–sink’ hypothesis of down-regulation of hypothesis in elevated [CO2] in the field with single gene substitutions in Glycine max.Crossref | GoogleScholarGoogle Scholar |
Al-Niemi TS, Kahn ML, McDermott TR (1998) Phosphorus uptake by bean nodules. Plant and Soil 198, 71–78.
| Phosphorus uptake by bean nodules.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaK1cXhvFKmu7w%3D&md5=1cc871909d700caed192e593412dbb66CAS |
Almeida JPF, Hartwig UA, Frehner M, Nösberger J, Lüscher A (2000) Evidence that P deficiency induced N feedback regulation of symbiotic N2 fixation in white clover (Trifolium repens L.). Journal of Experimental Botany 51, 1289–1297.
| Evidence that P deficiency induced N feedback regulation of symbiotic N2 fixation in white clover (Trifolium repens L.).Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD3cXlsVyjtbs%3D&md5=f45d28c8aa18527143c9f7b7cd66bb9eCAS |
Bazzaz FA (1997) Allocation of resources in plants: state of science and critical questions. In ‘Plant resource allocation’. (Eds FA Bazzaz, J Grace) pp. 1–37. (Academic Press: San Diego, CA)
Bieleski RL (1973) Phosphate pools, phosphate transport, and phosphate availability. Annual Review of Plant Physiology 24, 225–252.
| Phosphate pools, phosphate transport, and phosphate availability.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaE3sXltFeitbY%3D&md5=19704fd5a74dea689619444df4652925CAS |
Bordeleau LM, Prevost D (1994) Nodulation and nitrogen fixation in extreme environments. Plant and Soil 161, 115–125.
| Nodulation and nitrogen fixation in extreme environments.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaK2cXlt12rt7Y%3D&md5=cb29945e294860beeee12c325d43281eCAS |
Correa OS, Barneix AJ (1997) Cellular mechanisms of pH tolerance in Rhizobium loti. World Journal of Microbiology and Biotechnology 13, 153–157.
| Cellular mechanisms of pH tolerance in Rhizobium loti.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaK2sXjtVGgtbs%3D&md5=ff236f28d202261a067e5d3bccad045cCAS |
Dakora FD, Phillips DA (2002) Root exudates as mediators of mineral acquisition in low-nutrient environments. Plant and Soil 245, 35–47.
| Root exudates as mediators of mineral acquisition in low-nutrient environments.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD38XnvVCit70%3D&md5=4ef7d1deaec3cd2fab8d11e8a08ce0b9CAS |
Dinkelaker B, Hengeler C, Marschner H (1995) Distribution and function of proteoid roots and other root clusters. Botanica Acta 108, 183–200.
| Distribution and function of proteoid roots and other root clusters.Crossref | GoogleScholarGoogle Scholar |
Drevon JJ, Hartwig UA (1997) Phosphorus deficiency increases the argon-induced decline of nodule nitrogenase activity in soybean and alfalfa. Planta 201, 463–469.
| Phosphorus deficiency increases the argon-induced decline of nodule nitrogenase activity in soybean and alfalfa.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaK2sXjtVehsro%3D&md5=f5988251fc8c21da4366d0f7475d4a86CAS |
Farquhar GD, Ehleringer JR, Hubick KT (1989) Carbon isotope discrimination and photosynthesis. Annual Review of Plant Physiology 40, 503–537.
| Carbon isotope discrimination and photosynthesis.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaL1MXktlKmu70%3D&md5=136c49a5bf26dfb3f52abb3d4ecc03e0CAS |
Gilbert GA, Knight JD, Vance CP, Allan DL (1999) Acid phosphatase activity in phosphorus-deficient white lupin roots. Plant, Cell and Environment 22, 801–810.
| Acid phosphatase activity in phosphorus-deficient white lupin roots.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaK1MXlsFGnsr8%3D&md5=05169b42c4e9ed233d861dd451292412CAS |
Gilroy S, Jones DL (2000) Through form to function: root hair development and nutrition uptake. Trends in Plant Science 5, 56–60.
| Through form to function: root hair development and nutrition uptake.Crossref | GoogleScholarGoogle Scholar | 1:STN:280:DC%2BD3M7jt12nsw%3D%3D&md5=5479f5308097a3c6166f4c9d3fe6f41aCAS | 10664614PubMed |
Goldblatt P, Manning J (2000) ‘Cape plants: a conspectus of the Cape flora of South Africa. Strelitzia, vol. 9.’ (National Botanical Institute: Pretoria, South Africa.)
Gordon AJ, Kessler W, Minchin FR (1990) Defoliation-induced stress in nodules of white clover. I. Changes in physiological parameters and protein synthesis. Journal of Experimental Botany 41, 1245–1253.
| Defoliation-induced stress in nodules of white clover. I. Changes in physiological parameters and protein synthesis.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaK3MXpvVGmsw%3D%3D&md5=37d37321fc35762f6fd4aac906dbd360CAS |
Gordon AJ, Kessler W, Minchin FR, Skot L, James CL (1997) Stress-induced decline in soybean N2 fixation is related to nodule sucrose synthase activity. Plant Physiology 114, 937–946.
Graham PH (1992) Stress tolerance in Rhizobium and Bradyrhizobium and nodulation under adverse soil conditions. Canadian Journal of Microbiology 38, 475–484.
| Stress tolerance in Rhizobium and Bradyrhizobium and nodulation under adverse soil conditions.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaK38Xlt1Ort7Y%3D&md5=a61de08eb1e49af81ac9bc5f42e15916CAS |
Greinwald R, Veen G, Van Wyk BE, Witte L, Czygan FC (1989) Distribution and taxonomic significance of major alkaloids in the genus Virgilia. Biochemical Systematics and Ecology 17, 231–238.
| Distribution and taxonomic significance of major alkaloids in the genus Virgilia.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaK3cXlt1WksA%3D%3D&md5=5c732f51570296b6c63a7fe2d56a8924CAS |
Grigg AM, Veneklass EJ, Lambers H (2008) Water relations and mineral nutrition of closely related woody plant species on desert dunes and interdunes. Australian Journal of Botany 56, 27–43.
| Water relations and mineral nutrition of closely related woody plant species on desert dunes and interdunes.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD1cXhsFymtLw%3D&md5=2c0a8b9e4974872b0b6faf050ab8bfc6CAS |
Groves RH (1983) ‘Nutrient cycling in Australian heath and South African fynbos.’ (Springer-Verlag: Berlin)
Harrison MT, Edwards EJ, Farquhar GD, Nicotra AB, Evans JR (2009) Nitrogen in cell walls of sclerophyllous leaves accounts for little of the variation in photosynthetic nitrogen-use-efficiency. Plant, Cell and Environment 32, 259–270.
| Nitrogen in cell walls of sclerophyllous leaves accounts for little of the variation in photosynthetic nitrogen-use-efficiency.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD1MXjsVKlsLw%3D&md5=83ccd3fda0b34fb22a37551d1a3cbaa9CAS | 19054350PubMed |
Hernández G, Ramírez M, Valdés-López O, Tesfaye M, Graham MA, Czechowski T, Schlereth A, Wandrey M, Erban A, Chueng F, Wu HC, Lara M, Town CD, Kopka J, Udvardi MK, Vance CP (2007) Phosphorus stress in common bean: root transcript and metabolic response. Plant Physiology 144, 752–767.
| Phosphorus stress in common bean: root transcript and metabolic response.Crossref | GoogleScholarGoogle Scholar | 17449651PubMed |
Hinsinger P (2001) Bioavailability of soil inorganic P in the rhizosphere as affected by root-induced chemical changes: a review. Plant and Soil 237, 173–195.
| Bioavailability of soil inorganic P in the rhizosphere as affected by root-induced chemical changes: a review.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD38XovVWlsQ%3D%3D&md5=9004d339d2a399d376e3f81cad08732cCAS |
Høgh-Jensen H, Schjoererring JK, Soussana JF (2002) The influence of phosphorus deficiency on growth and nitrogen fixation of white clover plants. Annals of Botany 90, 745–753.
| The influence of phosphorus deficiency on growth and nitrogen fixation of white clover plants.Crossref | GoogleScholarGoogle Scholar | 12451030PubMed |
Horst WJ, Kamh M, Jibrin JM, Chude VA (2001) Agronomic measures for increasing P availability to crops. Plant and Soil 237, 211–223.
| Agronomic measures for increasing P availability to crops.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD38XovVWltw%3D%3D&md5=c964d613c4589cc8e84e9f4d9cb6e700CAS |
Israel DW (1987) Investigation of the role of phosphorus in symbiotic dinitrogen. Plant Physiology 84, 835–840.
| Investigation of the role of phosphorus in symbiotic dinitrogen.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaL2sXltFakt7g%3D&md5=ac4f33538158ebe18fe135afe9973454CAS | 16665531PubMed |
Jones DL (1998) Organic acids in the rhizosphere: a critical review. Plant and Soil 205, 25–44.
| Organic acids in the rhizosphere: a critical review.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaK1MXhtlGjs78%3D&md5=2ba08e296cb8f7509773bf165f731cdeCAS |
Jungk A, Seeling B, Gerke J (1993) Mobilisation of different phosphate fractions in the rhizosphere. Plant and Soil 156, 91–94.
| Mobilisation of different phosphate fractions in the rhizosphere.Crossref | GoogleScholarGoogle Scholar |
Juszczuk IM, Rychter AM (2002) Pyruvate accumulation during phosphate deficiency stress of bean roots. Plant Physiology and Biochemistry 40, 783–788.
| Pyruvate accumulation during phosphate deficiency stress of bean roots.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD38Xnt1Ggtb4%3D&md5=0128286e6328fa925ce229aefe5b71a3CAS |
Kanu SA, Dakora FD (2012) Symbiotic nitrogen contribution and biodiversity of root-nodule bacteria nodulating Psoralea species in the Cape fynbos, South Africa. Soil Biology and Biochemistry 54, 68–76.
| Symbiotic nitrogen contribution and biodiversity of root-nodule bacteria nodulating Psoralea species in the Cape fynbos, South Africa.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC38XhtVSlsb7P&md5=599dee9ba7bdd7e6c2bd4a066d6a81f5CAS |
Kaschuk G, Kuyper TW, Leffelaar PA, Hungria M, Giller KE (2009) Are the rates of photosynthesis stimulated by the carbon sink strength of rhizobial and arbuscular mycorrhizal symbioses? Soil Biology and Biochemistry 41, 1233–1244.
| Are the rates of photosynthesis stimulated by the carbon sink strength of rhizobial and arbuscular mycorrhizal symbioses?Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD1MXmtVGktbo%3D&md5=3b8d661e5233b5c22f04d81431fa6305CAS |
Kaschuk G, Hungria M, Leffelaar PA, Giller KE, Kuyper TW (2010a) Differences in photosynthetic behaviour and leaf senescence of soybean (Glycine max [L.] Merrill) dependent on N2 fixation or nitrate supply. Plant Biology 12, 60–69.
| Differences in photosynthetic behaviour and leaf senescence of soybean (Glycine max [L.] Merrill) dependent on N2 fixation or nitrate supply.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3cXlslWqtrY%3D&md5=67e6f845c71c3e22cb1bc62060aebae3CAS | 20653888PubMed |
Kaschuk G, Leffelaar PA, Giller KE, Alberton O, Hungria M, Kuyper TW (2010b) Responses of legumes to rhizobia and arbuscular mycorrhizal fungi: a meta-analysis of potential photosynthate limitation of symbioses. Soil Biology and Biochemistry 42, 125–127.
| Responses of legumes to rhizobia and arbuscular mycorrhizal fungi: a meta-analysis of potential photosynthate limitation of symbioses.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD1MXhsVGrtLfE&md5=6c36cf4f2f9f90cb97b531426e2f3295CAS |
Koide RT, Goff MD, Dickie IA (2000) Components of growth efficiencies on mycorrhizal and non-mycorrhizal plants. New Phytologist 148, 163–168.
| Components of growth efficiencies on mycorrhizal and non-mycorrhizal plants.Crossref | GoogleScholarGoogle Scholar |
Kruger FJ, Mitchell DT, Jarvis JUM (1983) ‘Mediterranean-type ecosystems. The role of nutrients.’ (Springer-Verlag: Berlin)
Lajtha K, Harrison AF (1995) Strategies of phosphorus acquisition and conservation by plants species and communities. In ‘Phosphorus in the global environment’. (Ed. H Tissen) pp. 140–147. (John Wiley Sons: Chichester, UK)
Lamont BB (2003) Structure, ecology and physiology of root clusters: a review. Plant and Soil 248, 1–9.
| Structure, ecology and physiology of root clusters: a review.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD3sXhtFCqs7s%3D&md5=324b1c82a25b8a9ff21e780abd7022b7CAS |
Le Roux MR, Kahn S, Valentine AJ (2008) Organic acid accumulation inhibits N2 fixation in P stressed lupin nodules. New Phytologist 177, 956–964.
| Organic acid accumulation inhibits N2 fixation in P stressed lupin nodules.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD1cXktVSnsLk%3D&md5=070d3ad629e0ce4bb1aeefbd265e3efbCAS | 18069956PubMed |
Le Roux MR, Khan S, Valentine AJ (2009) Nitrogen and carbon costs of soybean and lupin root systems during phosphate starvation. Symbiosis 48, 102–109.
| Nitrogen and carbon costs of soybean and lupin root systems during phosphate starvation.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD1MXpsFenurY%3D&md5=36ec231aff485e82037767996aed37c2CAS |
Lie TA (1981) Environmental physiology of legume-Rhizobium symbiosis. In ‘Nitrogen fixation vol. 1: ecology’. (Ed. WJ Broughton) pp. 104–134. (Clarendon Press: Oxford, UK)
Lynch JP, Beebe SE (1995) Adaptations of bean (Phaseolus vulgaris L.) to low phosphorus availability. HortScience 30, 1165–1171.
Lynch JP, Brown KM (2001) Topsoil foraging-an architectural adaptations of plants to low phosphorus. Plant and Soil 237, 225–237.
| Topsoil foraging-an architectural adaptations of plants to low phosphorus.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD38XovVWltA%3D%3D&md5=9e634f2da28b841af93504bde22d38e5CAS |
Lynch JP, Ho MD (2005) Rhizoeconomics: carbon costs of phosphorus acquisition. Plant and Soil 269, 45–56.
| Rhizoeconomics: carbon costs of phosphorus acquisition.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2MXks1Oisrw%3D&md5=6c040ce5e9b44d5abec556913341f132CAS |
Marschner H (1995) ‘Mineral nutrition in plants.’ 2nd edn. (Academic: San Diego, CA)
Maseko ST, Dakora FP (2013) Plant Enzymes, root exudates, cluster roots and mycorrhizal symbiosis are the driver of P nutrition in native legumes growing in P deficient soil of the Cape Fynbos in South Africa. Journal of Agricultural Science and Technology 3, 331–340.
Mengel K (1994) Symbiotic dinitrogen fixation-its dependence on plant nutrition and its ecophysiological impact. Zeitschrift für Pflanzenernährung und Bodenkunde 157, 233–241.
Mitchell DT, Brown G, Jongens-Roberts SM (1984) Variation and forms of phosphorus in the sandy soils coastal fynbos, southern-western Cape. Journal of Ecology 72, 575–584.
| Variation and forms of phosphorus in the sandy soils coastal fynbos, southern-western Cape.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaL2cXls12rsrg%3D&md5=d520d7019f84b31ae7d26bffc8a24bcbCAS |
Mortimer PE, Archer E, Valentine AJ (2005) Mycorrhizal C costs and nutritional benefits in developing grapevines. Mycorrhiza 15, 159–165.
| Mycorrhizal C costs and nutritional benefits in developing grapevines.Crossref | GoogleScholarGoogle Scholar | 1:STN:280:DC%2BD2M3ksFWisw%3D%3D&md5=bc78e864e931d0c584b5365088782ba6CAS | 15883853PubMed |
Mortimer PE, Perez-Fernandez MA, Valentine AJ (2008) The role of arbuscular mycorrhizal colonisation in the carbon and nutrient economy of the tripartite symbiosis with nodulated Phaseolus vulgaris. Soil Biology and Biochemistry 40, 1019–1027.
| The role of arbuscular mycorrhizal colonisation in the carbon and nutrient economy of the tripartite symbiosis with nodulated Phaseolus vulgaris.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD1cXksVKru7c%3D&md5=34752c61c7b554ccf3ddb9269fc5f6f1CAS |
Mortimer PE, Perez-Fernandez MA, Valentine AJ (2009) Arbuscular mycorrhizae affects the N and C economy of nodulated Phaseolus vulgaris (L.) during NH4 + nutrition. Soil Biology and Biochemistry 41, 2115–2121.
| Arbuscular mycorrhizae affects the N and C economy of nodulated Phaseolus vulgaris (L.) during NH4 + nutrition.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD1MXhtFKnsrjF&md5=ecc6771783a6a288575898ca50353fc1CAS |
Munns DN (1986) Acid soil tolerance in legume and rhizobia. Advances in Plant Nutrition 2, 63–91.
Muofhe ML, Dakora FD (1999) Nitrogen nutrition in nodulated field plants of the shrub tea legume Aspalathus linearis assessing using 15N natural abundance. Plant and Soil 209, 181–186.
| Nitrogen nutrition in nodulated field plants of the shrub tea legume Aspalathus linearis assessing using 15N natural abundance.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaK1MXlt1emsrw%3D&md5=5ab695e397a06c4f6d71076d2a12050aCAS |
Neumann G, Martinoia E (2002) Cluster root- an underground adaptation for survival in extreme environments. Trends in Plant Science 7, 162–167.
| Cluster root- an underground adaptation for survival in extreme environments.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD38XjtVKnsbY%3D&md5=649c1b3cbf74458c659b6069af9a054bCAS | 11950612PubMed |
Nielsen KL, Bouma TJ, Lynch J, Eissenstat DM (1998) Effects of phosphorus availability and vesicular-arbuscular mycorrhizas on the carbon budget of common bean (Phaseolus vulgaris). New Phytologist 139, 647–656.
| Effects of phosphorus availability and vesicular-arbuscular mycorrhizas on the carbon budget of common bean (Phaseolus vulgaris).Crossref | GoogleScholarGoogle Scholar |
Nielsen KL, Amram E, Lynch JP (2001) The effect of phosphorus availability on the carbon economy of contrasting common bean (Phaseolus vulgaris L.) genotypes. Journal of Experimental Botany 52, 329–339.
| The effect of phosphorus availability on the carbon economy of contrasting common bean (Phaseolus vulgaris L.) genotypes.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD3MXjtVaju7w%3D&md5=ed2b7781fdd89fc3707f6464a5145f73CAS | 11283178PubMed |
Olivera M, Tejera N, Iribarne C, Ocana A, Lluch C (2004) Growth, nitrogen fixation and ammonium assimilation in common bean (Phaseolus vulgaris): effect of phosphorus. Physiologia Plantarum 121, 498–505.
| Growth, nitrogen fixation and ammonium assimilation in common bean (Phaseolus vulgaris): effect of phosphorus.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2cXlvFOiur8%3D&md5=1197313fe8a2f0f3f51cac250f2a1a59CAS |
Peng S, Eissenstat DM, Graham JH, Williams K, Hodge NC (1993) Growth depression in mycorrhizal citrus at high-phosphorus supply. Plant Physiology 101, 1063–1070.
Plaxton WC, Carswell MC (1999) Metabolic aspects of the phosphate starvation response in plants. In ‘Plant responses to environmental stress: from phytohormones to genome reorganisation’. (Ed. HR Lerner) pp. 350–372. (Marcel-Decker: New York)
Power SC, Cramer MD, Verboom GA, Chimphango SBM (2010) Does phosphate acquisition constraint legume persistence in the fynbos of the Cape Floristic Region? Plant and Soil 334, 33–46.
| Does phosphate acquisition constraint legume persistence in the fynbos of the Cape Floristic Region?Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3cXhtVaqur7F&md5=da909e8069c6f07d08ea30251e65f242CAS |
Raghothama KG (1999) Phosphate acquisition. Annual Review of Plant Physiology and Plant Molecular Biology 50, 665–693.
| Phosphate acquisition.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaK1MXkt1yktrs%3D&md5=d7c5df202ff4d3000a97d47a650e0d30CAS | 15012223PubMed |
Richardson AE (1994) Soil microorganisms and phosphorus availability. In ‘Soil biota: management in sustainable farming systems’. (Eds CE Pankhurst, BD Doube, VVSR Gupta, PR Grace) pp. 50–60. (CSIRO: Melbourne)
Ryan PR, Delhaise E, Jones DL (2001) Function and mechanism of organic anion exudation from roots. Annual Review of Plant Physiology and Plant Molecular Biology 52, 527–560.
| Function and mechanism of organic anion exudation from roots.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD3MXkslWgsbg%3D&md5=91e69caed65b0c103adcb7cc7e9acaaeCAS | 11337408PubMed |
Rychter AM, Mikulska M (1990) The relationship between phosphate status and cyanide-resistant respiration in bean roots. Physiologia Plantarum 79, 663–667.
| The relationship between phosphate status and cyanide-resistant respiration in bean roots.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaK3cXlslWjsro%3D&md5=426f9f2f3a34510286ae4cec9914464eCAS | 21087276PubMed |
Sa TM, Israel DW (1991) Energy status and function of phosphorus deficient soybeans nodules. Plant Physiology 97, 928–935.
| Energy status and function of phosphorus deficient soybeans nodules.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaK38XktVCjtA%3D%3D&md5=f82904af1edb3c00ef5edad0fa7fc116CAS | 16668533PubMed |
Schachtman DP, Reid RJ, Ayling SM (1998) Phosphorus uptake by plants: from soil to cell. Plant Physiology 116, 447–453.
| Phosphorus uptake by plants: from soil to cell.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaK1cXht1ajtbc%3D&md5=e9ced15bcd4b16b542a93014493e6f87CAS | 9490752PubMed |
Shearer G, Kohl DH (1986) N2-fixation in field settings: estimations based on natural 15N abundance. Australian Journal of Plant Physiology 13, 699–756.
Spriggs AC, Dakora FD (2008) Field assessment of symbiotic N2 fixation in wild and cultivated Cyclopia species in the South African fynbos by 15N natural abundance. Tree Physiology 29, 239–247.
| Field assessment of symbiotic N2 fixation in wild and cultivated Cyclopia species in the South African fynbos by 15N natural abundance.Crossref | GoogleScholarGoogle Scholar | 19203949PubMed |
Tang C, Hensinger P, Drevon J-J, Jaillard B (2001) Phosphorus deficiency impairs early nodule functioning and enhances proton release in roots of Medicago truncatula L. Annals of Botany 88, 131–138.
| Phosphorus deficiency impairs early nodule functioning and enhances proton release in roots of Medicago truncatula L.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD3MXkvVOltL4%3D&md5=afdf358e6bce256868489a53409a0b9aCAS |
Uhde-Stone C, Gilbert G, Johnson JMF, Litjens R, Zinn KE, Temple SJ, Vance CP, Allan DL (2003a) Acclimation of white lupin to phosphorus deficiency involves enhanced expression of genes related to organic acid metabolism. Plant and Soil 248, 99–116.
| Acclimation of white lupin to phosphorus deficiency involves enhanced expression of genes related to organic acid metabolism.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD3sXhtFCru7g%3D&md5=68daab1da024739c1c0f02b42a97de12CAS |
Uhde-Stone C, Zinn KE, Ramerez-Yanez M, Li A, Vance CP, Allan DL (2003b) Nylon filters arrays reveal differential gene expression in proteoid roots of white lupin in response to P deficiency. Plant Physiology 131, 1064–1079.
| Nylon filters arrays reveal differential gene expression in proteoid roots of white lupin in response to P deficiency.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD3sXisFemu74%3D&md5=6bff6b6bf8560e0975d341af0ce2f9cfCAS | 12644659PubMed |
Vadez V, Beck DP, Lasso JH, Drevon JJ (1997) Utilisation of acetylene reduction assay to screen for tolerance of symbiotic N2 fixation to limitation P nutrition in common bean. Physiologia Plantarum 99, 227–232.
| Utilisation of acetylene reduction assay to screen for tolerance of symbiotic N2 fixation to limitation P nutrition in common bean.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaK2sXjtFWjsLc%3D&md5=b101ea721581b8ee421316e48d7753d4CAS |
Vance CP (2001) Symbiotic nitrogen fixation and phosphorus acquisition. Plant nutrition in a world of declining renewable resources. Plant Physiology 127, 390–397.
| Symbiotic nitrogen fixation and phosphorus acquisition. Plant nutrition in a world of declining renewable resources.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD3MXnslGltbw%3D&md5=e0b55789efe26ce5ac76ae505c253c89CAS | 11598215PubMed |
Vance CP, Uhde-Stone C, Allan DL (2003) Phosphorus acquisition and use: critical adaptations by plants for securing a non-renewable resource. New Phytologist 157, 423–447.
| Phosphorus acquisition and use: critical adaptations by plants for securing a non-renewable resource.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD3sXisF2gu70%3D&md5=6475fd5c3e1e1eea82a67eb0e2194ff5CAS |
Von Uexkull HR, Mutert E (1998) Global extent, development and economic impact of acid soils. In ‘Plant–soil interaction at low pH: principles and management’. (Eds RA Date, NJ Grundon NJ, GE Payment, ME Probert) pp. 5–9. (Kluwer Academic Publisher: Dordrecth, The Netherlands)
Williams K, Percival F, Merino J, Mooney HA (1987) Estimation of tissue construction cost from heat of combustion and organic nitrogen content. Plant, Cell and Environment 10, 725–734.