

Going nowhere fast: a review of seed dispersal in eucalypts
Trevor H. BoothCSIRO Land and Water, GPO Box 1600, Canberra, ACT 2601, Australia. Email: Trevor.Booth@csiro.au
Australian Journal of Botany 65(5) 401-410 https://doi.org/10.1071/BT17019
Submitted: 25 January 2017 Accepted: 6 July 2017 Published: 8 August 2017
Abstract
Eucalypt species have several features that make them particularly well suited for climate change studies. A key assumption is that they have very limited powers of dispersal. If this is correct, it means that climate change analyses to the end of this century can concentrate mainly on assessing whether or not eucalypt species are likely to be able to survive at their existing sites. A recent major climate change study of more than 600 eucalypt species for the period 2014–2085 has used 5 km as a usual dispersal limit for the period to 2085, with the possibility of rare long-distance events. The review presented here considers how far natural stands of eucalypt species are likely to be able to migrate in the period to 2085. It is the first review to consider eucalypt seed dispersal as its major focus. It draws on evidence from millions of years ago to the present, and from eucalypt stands in Australia and around the world. Although rare long-distance events cannot be entirely ruled out, it is concluded that the great bulk of the evidence available indicates that the most likely potential dispersal rate is equivalent to about 1–2 m per year, i.e. ~70–140 m in the period to 2085. Over decades, this is likely to occur as a series of stepwise events, associated with disturbances such as bushfires. However, limitations such as inadequate remnant eucalypt stands and extensive agricultural developments may reduce actual migration rates below even this modest potential.
Additional keywords: Angophora, climate change, Corymbia, Eucalyptus, species distribution model.
Introduction
Eucalypt species (including a total of ~800 species from the genera Eucalyptus, Corymbia and Angophora) have been identified as being particularly useful for climate change impact studies (Booth et al. 2015). Hundreds of eucalypt species have been evaluated at trial sites in numerous countries around the world, using unimproved seed collected from natural stands (see, for example, Poynton 1979; Jacobs 1981). So, unlike most plant and animal groups, much is known about the ability of eucalypt species to tolerate climatic conditions different from those of their natural distributions. Another key characteristic that makes eucalypts particularly useful for climate change studies is the assumption that they have very limited dispersal powers. If it can be assumed that over decades eucalypts disperse at rates equivalent to about 1–2 m per year, then climate change studies to the end of this century can concentrate on analysing locations within existing distributions. This greatly simplifies climate change studies, because complicating factors, such as dispersal distances and land use patterns, as well as edaphic suitability and competitive establishment at new sites, can be ignored. If their dispersal distances are very limited, the key question for eucalypt species for the next 70 years or so becomes: ‘can they survive where they are presently located?’.
A recent major climate change study of eucalypts (González-Orozco et al. 2016) showed dramatic shifts in the ‘the predicted species distribution of the suitable climate space’ for 657 eucalypt species in the period from 2014 to 2085. This included many that involve changes of several hundred kilometres, with the greatest being ~1400 km for E. semota C.J.Macpherson & Grayling. However, González-Orozco et al. (2016) stated in their methods section that ‘most eucalypt species would not be expected to disperse beyond a single grid cell [5 × 5 km] during the time period in this study without a rare long-distance dispersal event’.
The purpose of the present review is to examine information about the seed dispersal capabilities of eucalypts, with a view to indicating how far most eucalypt stands are likely to disperse in the period to 2085. The difference between limited dispersal of ~100 m and dispersal up to several kilometres is important. Climatic and edaphic conditions are likely to be similar over very short distances, but the chances of experiencing different environmental conditions increases with greater distances.
Pollen movement (and hybridisation with other species) can contribute to migration of eucalypts. Dispersal studies suggest that eucalypt pollen regularly travels distances of over 1 km, although most is distributed within 200 m and seed crops are dominated by genetic material from nearby trees (Byrne et al. 2008; Broadhurst 2013). Pollen may well play a vital role in eucalypt gene flow over long periods (Potts and Wiltshire 1997), whereas seed dispersal is the critical means for movement of intact (i.e. non-hybridised) species over substantial distances in the next few decades. So, the focus here is on seed dispersal, particularly the distances likely to be achieved in the remainder of this century. Related aspects of the reproductive biology of eucalypts, such as flowering, pollination and seed maturation, are not considered here, but have been reviewed by House (1997). Factors affecting the establishment of eucalypt seedlings from seed have been previously reviewed by Stoneman (1994), so are only briefly outlined here. For any eucalypt species, seed dispersal is the critical issue for short-term migration. If this is very limited, its natural abilities to colonise new locations will be greatly limited.
Relevant references for the present review were located first in the Web of Science™ and Google Scholar™ by searches using ‘eucalypt’ and ‘seed dispersal’ as key terms. However, many of these papers related to the dispersal of seed of non-eucalypt species in environments where eucalypts are also present, so 118 additional references were examined by searching papers that cited a key early eucalypt seed dispersal paper (Cremer 1977). The review is structured in the following way: basic mechanisms of seed dispersal and seedling establishment, alternative dispersal methods, dispersal over very long periods, dispersal within Australia and dispersal from plantings outside Australia. A final section before the discussion considers likely problems of establishment for the dispersal of eucalypt species under climate change in Australia.
Basic mechanisms of seed dispersal and seedling establishment
Eucalypt species use the following four regenerative strategies: obligate seeding, lignotuber sprouting, stem sprouting and combination sprouting (Nicolle 2006). However, their broad-scale dispersal depends on seed production. The basic strategy of eucalypt seed dispersal is to produce very large numbers of seeds, which are simply released and fall to the ground (see Vittoz and Engler 2007, for a review of different seed dispersal strategies). Some fast-growing eucalypts may produce seeds in as little as 3 years, but slow-growing species such as E. regnans F.Muell. may take 15–20 years (Gill 1981). Eucalypt seeds are relatively light in relation to most tree species (see, for example, Greene and Johnson 1994). Most range in weight from 0.16 g per 1000 seeds for E. deglupta Blume to 4.95 g per 1000 seeds for E. globulus Labill. (see the Royal Botanic Gardens Seed Information Database, data.kew.org/sid/, verified 14 July 2017). Variations in published weights for seeds of the same species are often due to whether chaff is included, and also whether seeds are air-dried or not. Eucalypt seeds lack any aerodynamic features, such as wings, to assist their spread. So, their dispersal is largely influenced by factors such as tree height, canopy width, seed weight and wind strength.
Early studies of seed dispersal in eucalypts showed that the bulk of the seeds fall within a distance roughly equal to the height of the tree (Grose 1957; Cunningham 1960; Cremer 1966). Cremer (1977) refined this simple assessment, by calculating dispersal distances in relation to wind velocity, height of release and terminal velocity. For example, from a height of 40 m with a 10 km h−1 (2.78 m s–1) wind, the lightest seed tested (E. deglupta) was expected to disperse 52.9 m, whereas the heaviest tested (E. globulus) was expected to disperse 20.1 m.
The size of the seed crop varies in cycles between 2 and 4 years for many eucalypt species (Harwood 1990). Seed collection for research purposes usually involves sampling of mature dominant trees. This is performed by removing selected branches and placing them over plastic sheets. The heat of the sun causes the seed carrying capsules to open and seeds can be collected. Heavy-seeding species, such as E. camaldulensis Dehnh. and E. globulus, may yield 5–10 kg of seed per tree in good seeding years (Harwood 1990). Under natural conditions, seed release in eucalypt forests and woodlands ranges from ~1 to 2.3 million seeds ha–1 (Loneragan 1979) to ~49 million seeds ha–1 (Burrows and Burrows 1992). There is generally a low level of seed release throughout the year, which is slower under wet conditions, but increases during periods of drought or following fires (Cunningham 1960).
Seed release following bushfires is important, because fires tend to clear away potentially competing vegetation. The role of seeds and seedling establishment in relation to fires has been considered by Gill (1997) as part of a review of eucalypts and fires. In eucalypt woodlands or forests, bushfires can involve ground fire (e.g. underground fire in peat), surface fire or crown fire. In surface fires, the tree canopy may be scorched, but does not burn. It is crown fires that are most likely to kill existing trees, providing opportunities for new individuals to establish, although some existing trees may resprout depending on the severity of the fire (Burrows 2013; Denham et al. 2016). Judd (1993) assessed seed survival of E. regnans under experimental conditions and Mercer et al. (1994) developed a time-dependent model of seed survival under fire conditions. Judd (1993) reported that film evidence showed that canopies can burn for as little as 10–30 s, and Ashton (1986) demonstrated that capsules of E. obliqua L’Hér. provide adequate protection to ensure that some seeds survive a crown fire.
Ants often eat large proportions of fallen eucalypt seeds (Yates et al. 1995). For example, Wellington and Noble (1985) estimated a period of only 5 days before half the fallen seeds of E. incrassata Labill. were eaten in long-unburnt semiarid stands in south-eastern Australia. Following bushfires, numbers of seed eating ants have been shown to increase in an E. obliqua L’Hér.–Casuariana pusilla Macklin woodland in south-eastern Australia (Andersen 1988). In an E. delegatensis R.T.Baker forest, O’Dowd and Gill (1984) showed that although ant numbers increase following fire, the numbers per available seeds declined as a result of the massive release of seeds. In the absence of fire, Andersen (1987) showed that ants limit seedling recruitment in an E. baxteri (Benth.) Maiden & Blakely ex J.M.Black–C. pusilla woodland in south-eastern Australia; however, predator satiation following massive seed release after fire produced high numbers of seedlings. Similarly, mass release of eucalypt seeds after fires also appeared to help satiate ants, and improve the chances of successful establishment at two old-field recolonisation sites in Western Australia (Harris and Standish 2008).
Stoneman (1994) and Bell and Williams (1997) reviewed the ecology and physiology of establishment of native eucalypt forests and woodlands from seed. Factors affecting germination and emergence, mortality and survival, and growth were considered. In summary, abiotic factors including water, temperature and, in some cases, light, were found to be major factors influencing germination and emergence (Boland et al. 1980; Gibson and Bachelard 1989; Battaglia 1993). Biotic factors included seed harvesting, which has already been mentioned. In addition, soil pathogens may cause mortality (Mwanza and Kellas 1987), and allelopathic effects may restrict emergence levels, but its importance depends on the species, the amount and types of litter, soil properties and climate (May and Ash 1990). Direct effects included seedbed conditions and overstorey density. Emergence and survival of eucalypt seed is usually better on burnt than unburnt, and disturbed rather than undisturbed, seedbeds (Burrows et al. 1990). In experiments, eucalypt seed has performed best when planted at depths between ~0.5 and 5 cm (Free 1951; Cremer 1965). Not surprisingly, considering the small size of the eucalypt seed, mortality is often greater where competing vegetation is already present, although partial shelter in commercial forests can moderate climatic conditions and improve establishment (Keenan 1986). Despite these limitations, eucalypt seeds are so numerous that some survive to replace existing stands, even though eucalypt seed loses viability usually within a year (Grose 1960).
Research since 1997 has refined knowledge of the processes affecting seed germination and seedling survival. For example, López et al. (2000) showed that the larger seeds of E. globulus germinated better than smaller seeds across a range of temperatures. Close and Wilson (2002) showed that large E. regnans seeds germinated at a faster rate than small seeds, and chilling increased germination rates for high-altitude provenances. Schütz et al. (2002) showed that large-seeded species from the south-western Australia are able to penetrate the soil profile faster and deeper. A particularly interesting observation was in relation to seedling survival under environmental change. Eucalyptus pauciflora Sieber ex Spreng seedlings grown in the field under elevated CO2 conditions (2× ambient) had 10 times more necrotic leaf area after an autumn freezing event than those grown under ambient conditions (Barker et al. 2005).
Alternative dispersal methods
Whereas the seeds of most eucalypt species fall within a distance approximately equivalent to the height of the tree, there are a few exceptions. If a species is found in riverine locations, the seeds may float downstream, and some may be delivered onto areas suitable for germination and establishment. For example, the spread of E. camaldulensis as an invasive weed along perennial, seasonal and intermittent watercourses in South Africa has often been reported (see, for example, Rejmánek and Richardson 2011). However, genomic research on 990 trees from 97 E. camaldulensis populations across the natural distribution in Australia has shown that genetic similarity was greatest between the nearest populations, regardless of whether or not they occurred on the same river system (Butcher et al. 2009).
A small number of bird species, including Calyptorhynchus baudinii Carnaby (long-billed black cockatoo), Calyptorhynchus magnificus Latham (red-tailed black cockatoo) and Purpreicephalus spurius Kuhl (red-capped parrot), derive a significant part of their diet from the unusually large seeds (~0.10 g each) of Corymbia calophylla (Lindl.) K.D.Hill & L.A.S.Johnson (House 1997). Calyptorhynchus banksia naso Gould (forest red-tailed black cockatoo), similarly, eat seeds from C. calophylla and, to a lesser extent, from E. patens Benth. and E. staeri (Maiden) Kessell & C.A.Gardner (Cooper et al. 2003). It is assumed that uneaten seeds are sometimes dispersed; however, no published data are available for how often this occurs or how far seeds are transported.
Australian stingless bees (Tetragonula carbonaria Smith) are attracted to and collect resin from the fruits of Corymbia torelliana (F.Muell.) K.D.Hill & L.A.S.Johnson (Leonhardt et al. 2014). In doing so, they pick up seeds, which may be removed by grooming on return to the nest. This can result in dispersal distances of up to 300 m (Wallace and Trueman 1995).
Seed dispersal by ants is not usually significant for eucalypts. Ants either eat eucalypt seeds in situ (Wellington 1989) or take them short distances, generally less than a few metres, back to their nests (Ashton 1979; Harris and Standish 2008). Many species of harvesting ants maintain seed stores below 15 cm of the soil surface (Andersen 1982). However, the seeds of most eucalypt species are so small that they cannot usually successfully germinate and establish if buried below ~5 cm (Cremer 1965).
Outlying populations
Advances in genetics and genecology are providing insights into variations within and among populations of eucalypts (Potts and Wiltshire 1997). If long-distance eucalypt-seed dispersal were taking place frequently, it might be expected that outlying stands would be common and that these stands would be genetically similar to parts at least of current main distributions. For example, a stand of E. globulus subsp. bicostata (Maiden, Blakely & Simmonds) J.B.Kirkp. that exists at Mount Bryan (South Australia) is located more than 600 km from the nearest other population of this subspecies. Vaillancourt et al. (2001) used random amplified polymorphic DNA (RAPD) molecular-marker analysis to assess the population, which appears to originate from a very old individual (possibly as old as 4000 years) that predates European settlement. Whether this stand is a result of natural long-distance seed dispersal, seed dispersal by the Aboriginal population or is a survivor of a much broader previous distribution of the subspecies is not clear. However, Vaillancourt et al. (2001) concluded that the simplest explanation was that the stand was once connected to the main Victorian populations and became isolated as conditions in the region became more arid.
Genetic differentiation has also been studied for 13 stands of Eucalyptus diversicolor F.Muell., which is found in a limited high-rainfall area in the extreme south-west of Western Australia (Coates and Sokolowski 1989). Two southern coastal outliers (William Bay and Many Peaks) showed insignificant genetic divergence from the main distribution, suggesting that separation may be recent. In contrast, outlying populations to the north-east and west showed greater differences from the main forest. This suggests that they have persisted as survivors of a more extensive distribution as conditions became drier from ~5000 years before present.
Other examples of isolated disjunct populations are found in the south-west of Western Australia. For instance, Byrne and Hopper (2008) used restriction fragment-length polymorphism (RFLP) markers to examine material from 11 locations of E. caesia subsp. caesia Brooker & Hopper and three locations of E. caesia subsp. magna Brooker & Hopper. The outlying populations showed high divergence, suggesting isolation over long periods. The outliers appear to represent surviving islands rather than being the result of recent dispersal.
One of the largest disjunctions for eucalypt species is shown by Eucalyptus perriniana F.Muell. ex Rodway. The main populations are located in south-eastern mainland Australia, whereas three small isolated populations are more than 500 km away in southern Tasmania. Genetic analyses showed that the Tasmanian populations were genetically distinct from the mainland populations, suggesting that they are surviving remnants rather than the result of recent long-distance dispersal (Rathbone et al. 2007).
Dispersal over very long periods
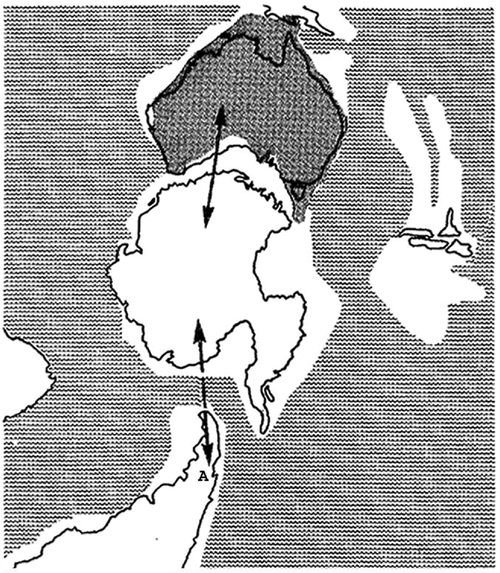
The current natural distribution of almost all eucalypt species is limited to the Australian continent, with a small number of species found to the north of Australia in Papua New Guinea, Indonesia and the Philippines (Boland et al. 1984). Intriguingly, the oldest Eucalyptus macrofossils have been found in Patagonia in Argentina, where the genus is no longer native (Gandolfo et al. 2011). These fossils have been dated to the early Eocene, ~51.9 million years ago. The earliest fossil eucalypt-type pollen from Myrtaceidites tenuis, which is considered to be a precursor of eucalypts, has been found not only in Australia, but also in eastern Antarctica (Macphail and Thornhill 2016). The point of origin of eucalypts is uncertain (Hill et al. 2016), but their dispersal to or from South America must have taken place when what has become Australia was still linked to South America by Antarctica (see Fig. 1). So, it is clear that eucalypts can disperse over vast distances given vast periods of time. But even the slowest rate of dispersal considered here (about 1 m per year), would be adequate given tens of millions of years.
|
There is a growing number of phylogeographic studies, which consider the historical and current distributions of eucalypt species in the light of genetics, and provide some insights into eucalypt dispersal over very long periods. In the Pleistocene epoch (from 2.588 million years ago to 11.7 thousand years ago), southern Australia experienced climatic oscillations with cyclic expansion and contraction of cool–dry (glacial) and warm–wet (interglacial) conditions (Markgraf et al. 1995). In contrast to much of Europe and North America, glaciation in southern Australia was not extensive, but was limited to the Central Plateau of Tasmania (Colhoun et al. 1996) and small parts of the Snowy Mountains in mainland Australia (Barrows et al. 2001).
Byrne (2008) reviewed phylogeographic studies during Pleistocene climatic cycles in southern Australia. In contrast to the major refugia found in the northern hemisphere, there appear to have been many localised refugia, with eucalypt species showing resilience rather than large-scale migration. For example, a phylogeographical study of E. loxophleba Benth. confirmed the importance of cyclic climatic expansions and contractions for eucalypts in the south-west of Western Australia (Byrne and Hines 2004). More recently, Nevill et al. (2010) used genetic analysis to show the long-term persistence of E. regnans in multiple refugia under harsh environmental conditions, which was followed by recolonisation of wider areas when more favourable conditions returned.
A study of E. globulus phylogeography in Tasmania and south-eastern mainland Australia showed genetic patterns corresponding to hypothesised glacial refugia (Freeman et al. 2001), which are consistent with limited seed based dispersal following deglaciation. Other studies in Tasmania have shown expansion of eucalypt species into previously glaciated inland plateau regions from eastern and south-eastern coastal refugia (McKinnon et al. 2001, 2004).
Seed dispersal from eucalypt sites in Australia
Dispersal distances of eucalypt seeds over periods of several decades are not surprisingly much more modest than is prehistoric spread over very long periods. For example, in 1932, E. cladocalyx F.Muell., a tree with some commercial uses but a limited natural distribution in South Australia, was introduced into King’s Park, Perth, Western Australia, as part of a series of war-memorial avenues. It became invasive and spread into the native bushland of the park, which is dominated by E. gomphocephala DC. and E. marginata Donn ex Sm. Some 70 years later, Ruthrof et al. (2003) reported that it had spread 70 m from the original planting sites. Seedling establishment was affected by fire events, with high numbers of seedlings being found in burnt areas and low numbers in unburnt areas, and with E. cladocalyx showing quicker canopy recovery following fires than locally native species.
A novel approach has made use of the many kilometres of cleared buffer strips around commercial plantations to examine the dispersal of E. globulus. This is the major hardwood plantation species used in Australia, with over 500 000 ha of plantations (ABARES 2015). Although comprising four major subspecies (recognised by some taxonomists as separate species), the commercial plantations are based on E. globulus subsp. globulus, which is native to Tasmania, as well as having very limited occurrences in southern coastal Victoria. Larcombe et al. (2013) inspected 290 km of plantation boundaries both within and outside the species’ natural distribution. They recorded nearly 5000 wildlings (i.e. plants growing outside plantation boundaries), with 98% occurring within 10 m of the plantation edge.
Evidence of seed dispersal from eucalypt stands outside Australia
Eucalypts have been very widely tested and extensively planted around the world for commercial-forestry purposes. One of the great advantages of overseas dispersal studies is that movements of eucalypts from planted locations into natural vegetation can be distinguished very easily. Plantations are generally located in areas where commercially satisfactory growth can be achieved, so climatic and edaphic conditions just outside plantation edges do not usually limit survival of wildlings. An important part of assessments outside Australia is evaluating the risks that eucalypt species may pose as potential weeds. These studies also provide some evidence relevant to factors affecting their natural spread.
Features influencing invasive characteristics of Australian tree species have been reviewed by Booth (2014). Even though many eucalypt species have been introduced around the world, there are very few species that are major problems as weeds. Where they are a problem, such as E. globulus in California, it is often because they have been widely planted in the past, so dispersal, although limited, now occurs from many locations (Wolf and DiTomaso 2016). Rejmánek and Richardson (2011) suggested that eucalypts are only weakly invasive because of (1) their relatively limited seed dispersal, (2) their high seedling mortality and (3) lack of compatible ectomycorrhizal fungi. The following recent studies, not included in the Booth (2014) review, have also emphasised the importance of the feeble powers of dispersal of eucalypts and their high seedling mortality.
For example, Fernandes et al. (2016) assessed natural regeneration of E. globulus from plantations into natural areas in Portugal. They found that 92% of seedlings were recorded less than 15 m from the plantation edge, and the presence of native forest or grassland was very resistant to eucalypt seedling establishment. Águas et al. (2014) studied the natural establishment of E. globulus in burnt stands in Portugal and also found that the existence of ‘close seed sources’ was important. Similarly, Calviño-Cancela and Rubido-Bará (2013) examined seed dispersal, seedling recruitment and survival in areas around plantations of E. globulus in Spain. Most seed fell within the first 5 m from plantation edges (84.3%) and 98.6% within the first 15 m. Both emergence (3.3% and 0.9%) and survival up to 6 months (17.5% and 3.7%) were greater in nearby shrublands than in native forest.
Lorentz and Minogue (2015) assessed the potential invasiveness of E. amplifolia Naudin, E. camaldulensis and E. grandis W.Hill in Florida. They found that no seedlings established outside two plantation stands, so they conducted seed addition studies to assess emergence rates and found the probability of emergence of added seed for different treatment combinations to be very low (P = 0.0 to 0.0032). They concluded that invasiveness risk was low, and a buffer strip of native grasses would be more effective than the bare-soil buffer strips usually used around plantations.
A few studies have examined the spread of eucalypt stands over several decades, particularly those of E. globulus in California. For example, the expansion of 16 mostly E. globulus plantations on Angel Island in San Francisco Bay were examined as part of a plan for their removal (California State Parks System 1988). The sketch map shown in Fig. 2a (McBride et al. 1988) covers a period of 73 years from 1915 to 1988, which is a length of time similar to those being considered in many current climate change analyses. The figure shows how wildlings spread from the original plantation, with the greatest extension being ~98 m in an easterly direction. This corresponds to the prevailing winds in the San Francisco Bay area, which are predominantly from the west or west–north-west (see Fig. 2b). Eucalypt expansion was also favoured by moist swales, and on road cuts where competition from annual grasses had been eliminated (J. McBride, pers. comm.). The total area shown in Fig. 2a increased by 3.7 times, from 0.24 ha of the original plantation area in 1915 to 0.89 ha in 1988. The report mapped and tabulated areas of expansion for all the 16 stands, with the mean increase in the total area in 1988 being 3.6 times that of the original plantation area. Greatest dispersal distances for individual stands, estimated from the report’s maps for this review, ranged from zero for roadside plantings (Site 14) to 133 m (Site 9). Similarly, Fork et al. (2015) analysed the spread of six eucalypt stands in the Elkholm Slough watershed (California). They found an average increase of 271% over 70 years (1931–2001), which was almost identical to that reported for the stands at Angel Island (i.e. 270%) over 73 years.

|
Problems of migratory eucalypt establishment in Australia
Although the present review has concentrated mainly on seed dispersal, it is appropriate to at least briefly consider the challenges of seedling establishment, because this is vital if individual eucalypt species are to respond to climate change by moving to new environments. The focus of interest here is on how difficult it is for eucalypts to migrate across landscapes, but much of the evidence comes from regeneration at or near existing sites.
For example, ~47 million hectares of the once extensive temperate eucalypt woodlands of southern Australia have been widely cleared for agriculture, with as little as 3% of the original areas of some types remaining in some areas (Department of the Environment and Water Resources 2007). The same problems that face restoration efforts limit the prospects of natural migration under climate change. These include clearing for agriculture, dryland salinity, excessive fertilisation, grazing, reduced fire regimes and exotic weeds (see review by Yates and Hobbs 1997).
Eucalypt woodlands have often been so effectively cleared that the remnants in many areas are inadequate to allow recolonisation of abandoned farmland. For instance, Standish et al. (2007) examined three farmland areas in the wheatbelt of Western Australia that had been abandoned for as much as 45 years. They found that two old fields were dominated by non-native annual grasses. They found declining levels of native seeds in the soil seedbank at 0 m, 50 m and 100 m into the old fields. They attributed the failure of eucalypt species to reclaim these areas to lack of seed availability and recruitment failure. Only in one old-field situation were eucalypts showing some signs of successfully re-establishing. Their even height suggested that one recruitment event, perhaps a bushfire in 1963, had facilitated their establishment.
Eucalypt regeneration has also been surveyed at 519 sites in grassy dry forests and grassy woodlands of central Victoria (Dorrough and Moxham 2005) and was observed at 27% of sites. Factors that reduced regeneration included intensive past land use (cultivation), regular livestock grazing and high cover of exotic annual vegetation (i.e. grasses, graminoids and forbs). A long history of agriculture limited future recruitment, even when grazing was removed. Increasing distance to remnant trees was also a factor, presumably being related to inadequate seed dispersal.
While regeneration is difficult even at sites where remnant trees are present, similar factors are likely to limit the abilities of eucalypt species to track rapidly changing climates across landscapes. Agricultural and urban developments, even major roadways, present significant dispersal barriers. Prior to European settlement, spread of eucalypts across landscapes was easier, particularly given long periods of time, occurrence of natural fires and the use of fire by the Aboriginal population (Yates and Hobbs 1997). Bushfires, mainly set off by lightning strikes, are still experienced regularly (about 1 year in 4 in any area) in the semiarid tropical savannas of Western Australia and the Northern Territory (Williams et al. 2009), so a high proportion of the land area burns every year.
However, the mean height of eucalypts in the wet–dry climate of the north is generally much less than in the milder conditions of high-rainfall regions of southern Australia (Boland et al. 1984). Because tree height plays a major role in determining dispersal distances, migration rates under the most favourable conditions may not be dramatically different. Hypothetically, in level topography, a representative eucalypt stand in northern Australia might take 5 years to grow to a height of 5 m and then disperse seeds over 5 m, whereas a stand in southern Australia might take 20 years to grow to 20 m dispersing seeds over 20 m. Over 100 years, the distance of each step is different, but the total migration distance would be similar. In reality, of course, the occurrence of such step movements are likely to be highly variable.
Whereas Australia is relatively flat in comparison to other continents, topography provides both opportunities and challenges for eucalypt dispersal in hilly or mountainous terrain. Austin and van Niel (2011) examined the importance of topography in determining the distribution of E. fastigata H.Deane & Maiden and discussed its shifting to different slopes and/or aspects as a response to climate change. They suggested that ‘species may need only to disperse across a valley to reach a suitable habitat, rather than to migrate hundreds of kilometres in latitude or hundreds of metres up an elevational gradient.’, p. 17. The possibility of species retreating to higher, cooler locations is a response often mentioned in climate change studies, but Green and Venn (2012) found that seedling establishment of E. pauciflora was rare uphill of existing sites, because of the limitations of seed dispersal. Simple geometry means that eucalypt seed dispersal distances are likely to be less on uphill slopes than on level ground.
Changing environmental conditions are likely to impose stresses on existing tree populations and contribute to dieback events (Ross and Brack 2015), which will reduce seed production. Even if they do not cause seedling or tree deaths, changing climatic conditions may interfere with reproductive processes. For example, E. nitens (H.Deane & Maiden) Maiden plantations grow successfully in South Africa under conditions that are warmer and drier than those within their natural distribution, but they do not flower and, hence, do not produce seeds (Booth et al. 2015). Similar problems have been observed with the majority of the more than 900 eucalypt species and subspecies being grown outside their natural distributions at the Currency Creek Arboretum in South Australia (Hay 2016).
Discussion
Studies described here have confirmed the general conclusions from pioneering eucalypt-dispersal studies of more than 50 years ago (Grose 1957; Cunningham 1960; Cremer 1966), which showed that eucalypts have very limited seed dispersal capabilities. It would be desirable to have more long-term studies of eucalypt dispersal, particularly from Australia. Nevertheless, the longer-term studies (see, for example, the studies from California and the Ruthrof et al. (2003) study from Australia) have suggested that eucalypt seed dispersal rates over periods of ~70 years are equivalent to about 1–2 m per year. Movements tend to take place in a stepwise fashion, taking advantage of disturbance events such as bushfires. Occasional long-distance dispersal events cannot be entirely ruled out, but the evidence presented here does not suggest that they are common events. Actual dispersal rates under climate change may well be below even the modest potential of 1–2 m per year, because obstacles such as inadequate remnants and extensive agricultural areas provide major impediments to natural migration.
There is no chance of natural migration of eucalypts by seed dispersal fully keeping pace with many of the distances indicated by even the moderate climate change scenario used by González-Orozco et al. (2016). Their fig. 1 was a reasonable way of summarising the large amount of data shown in their 657 supplementary maps. However, it would have been desirable to indicate in the caption that such great movements are not expected. If eucalypts are routinely capable of migrating only across landscapes in the order of ~71–142 m in 71 years, then no species should be capable of occupying any substantial part of the ‘2085 only’ areas shown in the 25 × 25 km resolution supplementary maps of González-Orozco et al. (2016).
Eucalypt pollen dispersal was mentioned in the introduction, but the possibility of long-distance migration over many kilometres by means of reticulate evolution should be mentioned. This involves pollen spread, hybridisation with related species, backcrossing and re-assembly, leaving evidence in the signatures of the chromosomal DNA. Examples include evolution of eastern grey box eucalypt species (Eucalyptus supraspecies Moluccanae) in the Murray–Darling Basin (Flores-Rentería et al. 2017), and E. globulus with related Symphyomyrtus species in south-eastern Tasmania (McKinnon et al. 2004). The latter study referred to movements over very long periods from glacial refugia, so it appears unlikely that such evolutionary processes can have any significant effect over periods as brief as that from 2014 to 2085.
Any tree species that experiences climatic conditions outside those of its current natural distribution as a result of climate change can respond in one of the following three ways: adapt (tolerate), move or die (Aitken et al. 2008). As mentioned in the introduction, there is a wealth of information available from commercial trials about how eucalypt species and provenances respond to climatic conditions different from those found within their natural distributions. Evolutionary change can be considered to be a special case of adaptation. However, as many eucalypt species can live for hundreds of years (see table 1.1 in Williams and Brooker 1997), the likelihood of any evolutionary adaptation to climate change in most stands is not great over the period to 2085. The ability of eucalypts to move by dispersal to new locations has been shown here to be very effective over very long periods, but very limited over periods of a few decades. Regional and local refugia have been of critical importance in allowing eucalypt species to recolonise regions when conditions become more favourable. As suggested by Austin and van Niel (2011), refugia at the landscape scale are likely to be important for eucalypt species to withstand the climate changes expected to occur this century. Finer-scale species distribution modelling tools are required that take into account, not only broad-scale climatic conditions, but also the effects of factors such as slope, position on slope and aspect on microsite conditions.
Trials outside Australia have demonstrated that many eucalypt species have climatic adaptability beyond that indicated by conventional analyses of their natural distributions (Booth et al. 2015). The analyses of González-Orozco et al. (2016) endeavoured to assess some of this adaptability, and Prober et al. (2016) reviewed recent studies of the climatic adaptability of eucalypts. However, more research is needed to determine the degree to which eucalypt species can rely on this adaptability as well as the existence of suitable refugia to cope with climate change, and the extent to which they will have to depend on their limited dispersal capabilities.
Conflicts of interest
The author declares no conflicts of interest.
Acknowledgements
Thanks go to Carlos González-Orozco and his colleagues for writing the paper that stimulated the preparation of this review. I am grateful to John Gerlach and Doug Johnson (California State Parks) for assistance with locating a copy of the 1988 Focussed environmental study (Angel Island) report. Thanks also go to Joe McBride (University of California, Berkeley) for his comments on that study. Ray Arrit (Iowa Environmental Mesonet, Iowa State University) assisted with obtaining approval to use the windrose figure. I am grateful to Suzanne Prober, Mike Austin and Libby Pinkard, as well as the anonymous referees, for their comments on earlier versions of this paper.
References
ABARES (2015) ‘Australia’s forests at a glance 2015.’ (Australian Bureau of Agricultural and Resource Economics and Sciences: Canberra)Águas A, Ferreira A, Maia P, Fernandes PM, Roxo L, Keizer J, Silva JS, Rego FC, Moreira F (2014) Natural establishment of Eucalyptus globulus Labill. in burnt stands in Portugal. Forest Ecology and Management 323, 47–56.
| Natural establishment of Eucalyptus globulus Labill. in burnt stands in Portugal.Crossref | GoogleScholarGoogle Scholar |
Aitken SN, Yeaman S, Holliday JA, Wang T, Curtis-McLane S (2008) Adaptation, migration or extirpation: climate change outcomes for tree populations. Evolutionary Applications 1, 95–111.
| Adaptation, migration or extirpation: climate change outcomes for tree populations.Crossref | GoogleScholarGoogle Scholar |
Andersen AN (1982) Seed removal by ants in the mallee of northwestern Victoria. In ‘Ant–plant interactions in Australia.’ (Ed. RC Buckley) pp. 31–44. (Dr W. Junk: The Hague)
Andersen AN (1987) Effects of seed predation by ants on seedling densities at a woodland site in SE Australia. Oikos 48, 171–174.
| Effects of seed predation by ants on seedling densities at a woodland site in SE Australia.Crossref | GoogleScholarGoogle Scholar |
Andersen AN (1988) Immediate and longer-term effects of fire on seed predation by ants in sclerophyllous vegetation in south-eastern Australia. Australian Journal of Ecology 13, 285–293.
| Immediate and longer-term effects of fire on seed predation by ants in sclerophyllous vegetation in south-eastern Australia.Crossref | GoogleScholarGoogle Scholar |
Ashton DH (1979) Seed harvesting by ants in forests of Eucalyptus regnans F.Muell. in central Victoria. Australian Journal of Ecology 4, 265–277.
| Seed harvesting by ants in forests of Eucalyptus regnans F.Muell. in central Victoria.Crossref | GoogleScholarGoogle Scholar |
Ashton DH (1986) Viability of seeds of Eucalyptus obliqua and Leptospermum juniperinum from capsules subjected to a crown fire. Australian Forestry 49, 28–35.
| Viability of seeds of Eucalyptus obliqua and Leptospermum juniperinum from capsules subjected to a crown fire.Crossref | GoogleScholarGoogle Scholar |
Austin MP, van Niel KP (2011) Impact of landscape predictors on climate change modelling of species distributions: a case study with Eucalyptus fastigata in southern New South Wales, Australia. Journal of Biogeography 38, 9–19.
| Impact of landscape predictors on climate change modelling of species distributions: a case study with Eucalyptus fastigata in southern New South Wales, Australia.Crossref | GoogleScholarGoogle Scholar |
Barker DH, Loveys BR, Egerton JJG, Gorton G, Williams WE, Ball MC (2005) CO2 enrichment predisposes foliage of a eucalypt to freezing injury and reduces spring growth. Plant, Cell & Environment 28, 1506–1515.
| CO2 enrichment predisposes foliage of a eucalypt to freezing injury and reduces spring growth.Crossref | GoogleScholarGoogle Scholar |
Barrows T, Stone J, Fifield L, Cresswell R (2001) Late Pleistocene glaciation of the Kosciuszko Massif, Snowy Mountains, Australia. Quaternary Research 55, 179–189.
| Late Pleistocene glaciation of the Kosciuszko Massif, Snowy Mountains, Australia.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD3MXhvV2iu7k%3D&md5=5a237734856e19d1d941650ce19019beCAS |
Battaglia M (1993) Seed germination physiology of Eucalyptus delegatensis R.T.Baker in Tasmania. Australian Journal of Botany 41, 119–136.
| Seed germination physiology of Eucalyptus delegatensis R.T.Baker in Tasmania.Crossref | GoogleScholarGoogle Scholar |
Bell DT, Williams JE (1997) Eucalypt ecophysiology. In ‘Eucalypt ecology’. (Eds JE Williams, JCZ Woinarski) pp. 168–196. (Cambridge University Press: Cambridge, UK)
Boland DJ, Brooker MIH, Turnbull JW (1980) ‘Eucalyptus seed.’ (CSIRO: Melbourne)
Boland DJ, Brooker MIH, Chippendale GM, Hall N, Hyland BPM, Johnston RD, Kleinig D, Turner JD (1984) ‘Forest Trees of Australia.’ (Nelson & CSIRO: Melbourne)
Booth TH (2014) Modern tree colonisers from Australia into the rest of the world. In ‘Invasion biology and ecological theory’. (Eds HHT Prins, IJ Gordon) pp. 304–323. (Cambridge University Press: Cambridge, UK)
Booth TH, Broadhurst LM, Pinkard E, Prober SM, Dillon SK, Bush D, Pinyopusarerk K, Doran JC, Ivkovich M, Young AG (2015) Native forests and climate change: lessons from eucalypts. Forest Ecology and Management 347, 18–29.
| Native forests and climate change: lessons from eucalypts.Crossref | GoogleScholarGoogle Scholar |
Broadhurst LM (2013) A genetic analysis of scattered yellow box (Eucalyptus melliodora A.Cunn. ex Schauer, Myrtaceae) and their restored cohorts. Biological Conservation 161, 48–57.
| A genetic analysis of scattered yellow box (Eucalyptus melliodora A.Cunn. ex Schauer, Myrtaceae) and their restored cohorts.Crossref | GoogleScholarGoogle Scholar |
Burrows GE (2013) Buds, bushfires and resprouting in eucalypts. Australian Journal of Botany 61, 331–349.
| Buds, bushfires and resprouting in eucalypts.Crossref | GoogleScholarGoogle Scholar |
Burrows DM, Burrows WH (1992) Seed production and litter fall in some eucalypt communities in central Queensland. Australian Journal of Botany 40, 389–403.
| Seed production and litter fall in some eucalypt communities in central Queensland.Crossref | GoogleScholarGoogle Scholar |
Burrows N, Gardiner G, Ward B, Robinson A (1990) Regeneration of Eucalyptus wandoo following fire. Australian Forestry 53, 248–258.
| Regeneration of Eucalyptus wandoo following fire.Crossref | GoogleScholarGoogle Scholar |
Butcher PA, McDonald MW, Bell JC (2009) Congruence between environmental parameters, morphology and genetic structure in Australia’s most widely distributed eucalypt, Eucalyptus camaldulensis. Tree Genetics & Genomes 5, 189–210.
| Congruence between environmental parameters, morphology and genetic structure in Australia’s most widely distributed eucalypt, Eucalyptus camaldulensis.Crossref | GoogleScholarGoogle Scholar |
Byrne M (2008) Evidence for multiple refugia at different time scales during Pleistocene climatic oscillations in southern Australia inferred from phylogeography. Quaternary Science Reviews 27, 2576–2585.
| Evidence for multiple refugia at different time scales during Pleistocene climatic oscillations in southern Australia inferred from phylogeography.Crossref | GoogleScholarGoogle Scholar |
Byrne M, Hines B (2004) Phylogeographical analysis of cpDNA variation in Eucalyptus loxophleba (Myrtaceae). Australian Journal of Botany 52, 459–470.
| Phylogeographical analysis of cpDNA variation in Eucalyptus loxophleba (Myrtaceae).Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2cXmsVChtLk%3D&md5=565817a718fa17b79ff9b798f35c93ddCAS |
Byrne M, Hopper SD (2008) Granite outcrops as ancient islands in old landscapes: evidence from the phylogeography and population genetics of Eucalyptus caesia (Myrtaceae) in Western Australia. Biological Journal of the Linnean Society. Linnean Society of London 93, 177–188.
| Granite outcrops as ancient islands in old landscapes: evidence from the phylogeography and population genetics of Eucalyptus caesia (Myrtaceae) in Western Australia.Crossref | GoogleScholarGoogle Scholar |
Byrne M, Elliott CP, Yates CJ, Coates DJ (2008) Maintenance of high pollen dispersal in Eucalyptus wandoo, a dominant tree of the fragmented agricultural region in Western Australia. Conservation Genetics 9, 97–105.
| Maintenance of high pollen dispersal in Eucalyptus wandoo, a dominant tree of the fragmented agricultural region in Western Australia.Crossref | GoogleScholarGoogle Scholar |
California State Parks System (1988) ‘Focussed environmental study: restoration of Angel Island natural areas affected by Eucalyptus.’ (California State Parks System: Sacramento, CA)
Calviño-Cancela M, Rubido-Bará M (2013) Invasive potential of Eucalyptus globulus: seed dispersal, seedling recruitment and survival in surrounding plantations. Forest Ecology and Management 305, 129–137.
| Invasive potential of Eucalyptus globulus: seed dispersal, seedling recruitment and survival in surrounding plantations.Crossref | GoogleScholarGoogle Scholar |
Close DC, Wilson SJ (2002) Provenance effects on pre-germination treatments for Eucalyptus regnans and E. delegatensis seed. Forest Ecology and Management 170, 299–305.
| Provenance effects on pre-germination treatments for Eucalyptus regnans and E. delegatensis seed.Crossref | GoogleScholarGoogle Scholar |
Coates DJ, Sokolowski RE (1989) Geographic patterns of genetic diversity in karri (Eucalyptus diversicolor F.Muell.). Australian Journal of Botany 37, 145–156.
| Geographic patterns of genetic diversity in karri (Eucalyptus diversicolor F.Muell.).Crossref | GoogleScholarGoogle Scholar |
Colhoun EA, Hannan D, Kiernan K (1996) Late Wisconsin glaciation of Tasmania. Papers and Proceedings of the Royal Society of Tasmania 130, 33–45.
Cooper CE, Withers PC, Mawson PR, Johnstone R, Kirby T, Prince J, Bradshaw SD, Robertson H (2003) Characteristics of marri (Corymbia calophylla) fruits in relation to the foraging behaviour of the forest red-tailed black cockatoo (Calyptorrynchus banksia naso). Journal of the Royal Society of Western Australia 86, 139–142.
Cremer KW (1965) Emergence of Eucalyptus regnans seedlings from buried seed. Australian Forestry 29, 119–124.
| Emergence of Eucalyptus regnans seedlings from buried seed.Crossref | GoogleScholarGoogle Scholar |
Cremer KW (1966) Dissemination of seed from Eucalyptus regnans. Australian Forestry 30, 33–37.
| Dissemination of seed from Eucalyptus regnans.Crossref | GoogleScholarGoogle Scholar |
Cremer KW (1977) Distance of seed dispersal in eucalypts estimated from seed weights. Australian Forest Research 7, 225–228.
Cunningham TM (1960) The natural regeneration of Eucalyptus regnans. School of Forestry Bulletin No. 1. (Melbourne University: Melbourne)
Denham AJ, Vincent BE, Clarke PJ, Auld TD (2016) Responses of tree species to a severe fire indicate major structural change to Eucalyptus–Callitris forests. Plant Ecology 217, 617–629.
| Responses of tree species to a severe fire indicate major structural change to Eucalyptus–Callitris forests.Crossref | GoogleScholarGoogle Scholar |
Department of the Environment and Water Resources (2007) ‘Australia’s native vegetation: a summary of Australia’s major vegetation groups.’ (Australian Government: Canberra)
Dorrough J, Moxham C (2005) Eucalypt establishment in agricultural landscapes and implications for landscape-scale restoration. Biological Conservation 123, 55–66.
| Eucalypt establishment in agricultural landscapes and implications for landscape-scale restoration.Crossref | GoogleScholarGoogle Scholar |
Fernandes P, Antunes C, Pinho P, Máguas C, Correia O (2016) Natural regeneration of Pinus pinaster and Eucalyptus globulus from plantation into adjacent natural habitats. Forest Ecology and Management 378, 91–102.
| Natural regeneration of Pinus pinaster and Eucalyptus globulus from plantation into adjacent natural habitats.Crossref | GoogleScholarGoogle Scholar |
Flores-Rentería L, Rymer PD, Riegler M (2017) Unpacking boxes: integration of molecular, morphological and ecological approaches reveals extensive patterns of reticulate evolution in box eucalypts. Molecular Phylogenetics and Evolution 108, 70–87.
| Unpacking boxes: integration of molecular, morphological and ecological approaches reveals extensive patterns of reticulate evolution in box eucalypts.Crossref | GoogleScholarGoogle Scholar |
Fork S, Woolfolk A, Akhavan A, van Dyke E, Murphy S, Candiloro B, Newberry T, Schreibman S, Salisbury J, Wasson K (2015) Biodiversity effects and rates of spread of non-native eucalypt woodlands in central California. Ecological Applications 25, 2306–2319.
| Biodiversity effects and rates of spread of non-native eucalypt woodlands in central California.Crossref | GoogleScholarGoogle Scholar |
Free RA (1951) Germination and emergence of eucalypt seed when sown at depth. Australian Forestry 15, 115–120.
| Germination and emergence of eucalypt seed when sown at depth.Crossref | GoogleScholarGoogle Scholar |
Freeman JS, Jackson HD, Steane DA, McKinnon GE, Dutkowski GW, Potts BM, Vaillancourt RE (2001) Chloroplast DNA phylogeography of Eucalyptus globulus. Australian Journal of Botany 49, 585–596.
| Chloroplast DNA phylogeography of Eucalyptus globulus.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD3MXptVemu7w%3D&md5=bb3a39a5d0a0144b47d8173e36118eb7CAS |
Gandolfo MA, Hermsen EJ, Zamaloa MC, Nixon KC, González CC, Wilf P, Cúneo NR (2011) Oldest known Eucalyptus macrofossils are from South America. PLoS One 6, e21084
| Oldest known Eucalyptus macrofossils are from South America.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3MXot1yqtLg%3D&md5=0582449abd1c2819855ec99ccea36e9eCAS |
Gibson A, Bachelard EP (1989) Variations in seed and seedling responses to water stress in three provenances of Eucalyptus camaldulensis Dehnh. Annals of Forest Science 46, 388s–392s.
| Variations in seed and seedling responses to water stress in three provenances of Eucalyptus camaldulensis Dehnh.Crossref | GoogleScholarGoogle Scholar |
Gill AM (1981) Adaptive responses of Australian vascular plant species to fire. In ‘Fire and the Australian biota.’ (Eds AM Gill, RH Groves, R Noble) pp. 243–272. (Australian Academy of Science: Canberra)
Gill AM (1997) Eucalypts and fire: interdependent or independent? In ‘Eucalypt ecology’. (Eds JE Williams, JCZ Woinarski) pp. 151–1167. (Cambridge University Press: Cambridge, UK)
González-Orozco C, Pollock LJ, Thornhill AH, Mischler BD, Knerr N, Laffan SW, Miller JT, Roscuer DF, Faith DP, Nipperess DA, Kujala H, Linke S, Butt N, Kűlheim C, Crisp MD, Gruber B (2016) Phylogenetic approaches reveal biodiversity threats under climate change. Nature Climate Change 6, 1110–1114.
| Phylogenetic approaches reveal biodiversity threats under climate change.Crossref | GoogleScholarGoogle Scholar |
Greene DF, Johnson EA (1994) Estimating the mean annual seed production of trees. Ecology 75, 642–647.
| Estimating the mean annual seed production of trees.Crossref | GoogleScholarGoogle Scholar |
Green K, Venn S (2012) Tree-limit ribbons in the Snowy Mountains, Australia: characterisation and recent seedling establishment. Arctic and Alpine Research 44, 180–187.
| Tree-limit ribbons in the Snowy Mountains, Australia: characterisation and recent seedling establishment.Crossref | GoogleScholarGoogle Scholar |
Grose RJ (1957) A study of some factors associated with the natural regeneration of Eucalyptus delegatensis. Forestry Commission Victoria Bulletin No. 4 (Forestry Commission Victoria: Melbourne)
Grose RJ (1960) Effective seed supply for the natural regeneration of Eucalyptus delegatensis R.T.Baker syn. Eucalyptus gigantean Hook.f. Journal of the Australian Pulp and Paper Industry Association 13, 131–147.
Harris RJ, Standish RJ (2008) Ant dispersal and predation affects the availability of seeds for old-field recolonization in Western Australia. Journal of the Royal Society of Western Australia 91, 301–311.
Harwood C (1990) Technical aspects of seed collection, storage and handling. In ‘Sowing the seeds: direct seeding and natural regeneration conference’. (Ed. Anon.) pp. 20–23. (Greening Australia: Canberra)
Hay A (2016) The forest at the edge of time. In ‘The best Australian science writing 2016’. (Ed. J. Chandler) pp. 52–73. (University of New South Wales Press: Sydney)
Hill RS, Beer YK, Hill KE, Maciunas E, Tarran MA, Wainman CC (2016) Evolution of the eucalypts – an interpretation of the macrofossil record. Australian Journal of Botany 64, 600–608.
| Evolution of the eucalypts – an interpretation of the macrofossil record.Crossref | GoogleScholarGoogle Scholar |
House SM (1997) Reproductive biology of eucalypts. In ‘Eucalypt ecology’. (Eds JE Williams, JCZ Woinarski) pp. 30–55. (Cambridge University Press: Cambridge, UK)
Jacobs MR (1981) ‘Eucalypts for planting.’ (FAO: Rome)
Judd TS (1993) Seed survival in small myrtaceous capsules subjected to experimental heating. Oecologia 93, 576–581.
| Seed survival in small myrtaceous capsules subjected to experimental heating.Crossref | GoogleScholarGoogle Scholar | 1:STN:280:DC%2BC1czot1egtQ%3D%3D&md5=1e261d16cf2721b24741da8fcdb7cc59CAS |
Keenan RJ (1986) Review of the shelterwood system and its potential for application in Tasmanian eucalypt forests. Australian Forestry 49, 226–235.
| Review of the shelterwood system and its potential for application in Tasmanian eucalypt forests.Crossref | GoogleScholarGoogle Scholar |
Larcombe MJ, Silva JS, Vaillancourt RE, Potts BM (2013) Assessing the invasive potential of Eucalyptus globulus in Australia: quantification of wildling establishment from plantations. Biological Invasions 15, 2763–2781.
| Assessing the invasive potential of Eucalyptus globulus in Australia: quantification of wildling establishment from plantations.Crossref | GoogleScholarGoogle Scholar |
Leonhardt SD, Baumann AM, Wallace HM, Brooks P, Schmitt T (2014) The chemistry of an unusual seed dispersal mutualism: bees use a complex set of olfactory cues to find their partner. Animal Behaviour 98, 41–51.
| The chemistry of an unusual seed dispersal mutualism: bees use a complex set of olfactory cues to find their partner.Crossref | GoogleScholarGoogle Scholar |
Loneragan OW (1979) ‘Karri (Eucalyptus diversicolor F.Muell) phenological studies in relation to reforestation.’ Bulletin no. 90. (Forests Department of Western Australia: Perth)
López M, Humara JM, Casares A, Majada J (2000) The effect of temperature and water stress on laboratory germination of Eucalyptus globulus Labill. seeds of different sizes. Annals of Forest Science 57, 245–250.
| The effect of temperature and water stress on laboratory germination of Eucalyptus globulus Labill. seeds of different sizes.Crossref | GoogleScholarGoogle Scholar |
Lorentz KA, Minogue PJ (2015) Potential invasiveness of Eucalyptus species in Florida. Invasive Plant Science and Management 8, 90–97.
| Potential invasiveness of Eucalyptus species in Florida.Crossref | GoogleScholarGoogle Scholar |
Macphail M, Thornhill AH (2016) How old are the eucalypts? A review of the microfossil and phylogenetic evidence. Australian Journal of Botany
| How old are the eucalypts? A review of the microfossil and phylogenetic evidence.Crossref | GoogleScholarGoogle Scholar |
Markgraf V, McGlone M, Hope G (1995) Neogene paleoclimatic change in southern temperate ecosystems – a southern perspective. Trends in Ecology & Evolution 10, 143–147.
| Neogene paleoclimatic change in southern temperate ecosystems – a southern perspective.Crossref | GoogleScholarGoogle Scholar | 1:STN:280:DC%2BC3M7itFajug%3D%3D&md5=a867688f927b90f99a954908e2c3f73aCAS |
May FE, Ash JE (1990) An assessment of the allelopathic potential of Eucalyptus. Australian Journal of Botany 38, 245–254.
| An assessment of the allelopathic potential of Eucalyptus.Crossref | GoogleScholarGoogle Scholar |
McBride JR, Sugihara N, Amme D (1988) The effects of Eucalyptus establishment on native plant communities In ‘Focused environmental study: restoration of Angel Island natural areas affected by Eucalyptus’. (Ed. D Boyd) pp. 43–85. (California Department of Parks and Recreation: Sacramento, CA)
McKinnon GE, Vaillancourt RE, Jackson HD, Potts BM (2001) Chloroplast sharing in Tasmanian eucalypts. Evolution 55, 703–711.
| Chloroplast sharing in Tasmanian eucalypts.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD3MXktleqtLs%3D&md5=902cd44d367872fdc57422a30e9a27e4CAS |
McKinnon GE, Jordan GJ, Vaillancourt RE, Steane DA, Potts BM (2004) Glacial refugia and reticulate evolution: the case for Tasmanian eucalypts. Proceedings of the Transactions of the Royal Society London B 359, 275–284.
| Glacial refugia and reticulate evolution: the case for Tasmanian eucalypts.Crossref | GoogleScholarGoogle Scholar |
Mercer GN, Gill AM, Weber RO (1994) A time-dependent model of fire impact on seed survival in woody fruits. Australian Journal of Botany 42, 71–81.
| A time-dependent model of fire impact on seed survival in woody fruits.Crossref | GoogleScholarGoogle Scholar |
Mwanza EJM, Kellas JD (1987) Identification of the fungi associated with damping-off in the regeneration of Eucalyptus obliqua and E. radiata in a central Victorian forest. European Journal of Forest Pathology 17, 237–245.
| Identification of the fungi associated with damping-off in the regeneration of Eucalyptus obliqua and E. radiata in a central Victorian forest.Crossref | GoogleScholarGoogle Scholar |
Nevill PG, Bossinger G, Ades PK (2010) Phylogeography of the world’s tallest angiosperm, Eucalyptus regnans: evidence for multiple isolated Quaternary refugia. Journal of Biogeography 37, 179–192.
| Phylogeography of the world’s tallest angiosperm, Eucalyptus regnans: evidence for multiple isolated Quaternary refugia.Crossref | GoogleScholarGoogle Scholar |
Nicolle D (2006) A classification and census of regenerative strategies in the eucalypts (Angophora, Corymbia and Eucalyptus – Myrtaceae), with special reference to the obligate seeders. Australian Journal of Botany 54, 391–407.
| A classification and census of regenerative strategies in the eucalypts (Angophora, Corymbia and Eucalyptus – Myrtaceae), with special reference to the obligate seeders.Crossref | GoogleScholarGoogle Scholar |
O’Dowd DJ, Gill AM (1984) Predator satiation and site alteration following fire: mass repdoduction of alpine ash (Eucalyptus delegatensis) in southeastern Australia. Ecology 65, 1052–1066.
| Predator satiation and site alteration following fire: mass repdoduction of alpine ash (Eucalyptus delegatensis) in southeastern Australia.Crossref | GoogleScholarGoogle Scholar |
Potts BM, Wiltshire RJE (1997) Eucalypt genetics and genecology. In ‘Eucalypt ecology’. (Eds JE Williams, JCZ Woinarski) pp. 56–91. (Cambridge University Press: Cambridge, UK)
Poynton RJ (1979) ‘Tree planting in southern Africa. Vol. 2. The eucalypts.’ (Department of Forestry, Republic of South Africa: Pretoria, South Africa)
Prober SM, Potts BM, Bailey T, Byrne M, Dillon S, Harrison PA, Hoffman AA, Jordan R, McLean EH, Steane DA, Stock WD, Vaillancourt RE (2016) Climate adaptation and ecological restoration in eucalypts. The Royal Society of Victoria 128, 40–53.
| Climate adaptation and ecological restoration in eucalypts.Crossref | GoogleScholarGoogle Scholar |
Rathbone DA, McKinnon GE, Potts BM, Steane DA, Vaillancourt RE (2007) Microsatellite and cpDNA variation in island and mainland populations of a regionally rare eucalypt, Eucalyptus perriniana (Myrtaceae). Australian Journal of Botany 55, 513–520.
| Microsatellite and cpDNA variation in island and mainland populations of a regionally rare eucalypt, Eucalyptus perriniana (Myrtaceae).Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2sXpt1SltbY%3D&md5=b5c755718cd63bcbc2833da92a747cfaCAS |
Rejmánek M, Richardson DM (2011) Eucalypts. In ‘Encyclopedia of invasive plants’. (Eds D Simberloff, M Rejmánek) pp. 203–209. (University of California Press: Berkeley, CA)
Ross C, Brack C (2015) Eucalyptus viminalis dieback in the Monaro region, NSW. Australian Forestry 78, 243–253.
| Eucalyptus viminalis dieback in the Monaro region, NSW.Crossref | GoogleScholarGoogle Scholar |
Ruthrof KX, Loneragan WA, Yates CJ (2003) Comparative population dynamics of Eucalyptus cladocalyx in its native habitat and as an invasive species in an urban bushland in south-western Australia. Diversity & Distributions 9, 469–483.
| Comparative population dynamics of Eucalyptus cladocalyx in its native habitat and as an invasive species in an urban bushland in south-western Australia.Crossref | GoogleScholarGoogle Scholar |
Schütz W, Milberg P, Lamont BB (2002) Germination requirements and seedling responses to water availability and soil type in four eucalypt species. Acta Oecologica 23, 23–30.
| Germination requirements and seedling responses to water availability and soil type in four eucalypt species.Crossref | GoogleScholarGoogle Scholar |
Standish RJ, Cramer VA, Wild SL, Hobbs RJ (2007) Seed dispersal and recruitment limitation are barriers to native colonization of old-fields in western Australia. Journal of Applied Ecology 44, 435–445.
| Seed dispersal and recruitment limitation are barriers to native colonization of old-fields in western Australia.Crossref | GoogleScholarGoogle Scholar |
Stoneman GL (1994) Ecology and physiology of establishment of eucalypt seedlings from seed: a review. Australian Forestry 57, 11–29.
| Ecology and physiology of establishment of eucalypt seedlings from seed: a review.Crossref | GoogleScholarGoogle Scholar |
Vaillancourt RE, Boomsma DB, Nicolle D (2001) A disjunct population of Eucalyptus globulus subsp. bicostata from South Australia. Transactions of the Royal Society of South Australia 126, 65–68.
Vittoz P, Engler R (2007) Seed dispersal distances: a typology based on dispersal modes and plant traits. Botanica Helvetica 117, 109–124.
| Seed dispersal distances: a typology based on dispersal modes and plant traits.Crossref | GoogleScholarGoogle Scholar |
Wallace HM, Trueman SJ (1995) Dispersal of Eucalyptus torelliana seeds by the resin-collecting sting-less bee, Trigona carbonaria. Oecologia 104, 12–16.
| Dispersal of Eucalyptus torelliana seeds by the resin-collecting sting-less bee, Trigona carbonaria.Crossref | GoogleScholarGoogle Scholar | 1:STN:280:DC%2BC1cznsFCitA%3D%3D&md5=0e8cac358efb65c2c55776ae0214db5fCAS |
Wellington AB (1989) Seedling regeneration and the population dynamics of eucalypts. In ‘Mediterranean landscapes in Australia: mallee ecosystems and their management’. (Eds JC Noble, RA Bradstock) pp. 155–167. (CSIRO: Canberra)
Wellington AB, Noble IR (1985) Seed dynamics and factors affecting recruitment of the mallee Eucalyptus incrassata in semi-arid, south-eastern Australia. Journal of Ecology 73, 657–666.
| Seed dynamics and factors affecting recruitment of the mallee Eucalyptus incrassata in semi-arid, south-eastern Australia.Crossref | GoogleScholarGoogle Scholar |
Williams JE, Brooker MIH (1997) Eucalypts: an introduction. In ‘Eucalypt ecology’. (Eds JE Williams, JCZ Woinarski) pp. 1–15. (Cambridge University Press: Cambridge, UK)
Williams RJ, Bradstock RA, Cary GJ, Enright NJ, Gill AM, Liedloff AC, Lucas C, Whelan RJ, Andersen AN, Bowman DJMS, Clarke PJ, Cook GD, Hennessy KJ, York A (2009) Interactions between climate change, fire regimes and biodiversity in Australia: a preliminary assessment. Report to the Department of Climate Change and Department of Environment, Water, Heritage and the Arts, Canberra.
Wolf KM, DiTomaso JM (2016) Management of blue gum eucalyptus in California requires region-specific consideration. California Agriculture 70, 39–47.
| Management of blue gum eucalyptus in California requires region-specific consideration.Crossref | GoogleScholarGoogle Scholar |
Yates CJ, Hobbs RJ (1997) Temperate eucalypt woodlands: a review of their status, processes threatening their persistence and techniques for restoration. Australian Journal of Botany 45, 949–973.
| Temperate eucalypt woodlands: a review of their status, processes threatening their persistence and techniques for restoration.Crossref | GoogleScholarGoogle Scholar |
Yates CJ, Taplin RH, Hobbs RJ, Bell RW (1995) Factors limiting the recruitment of Eucalyptus salmonophloia in remnant woodlands II. Post dispersal seed predation and soil seed reserves. Australian Journal of Botany 43, 145–155.
| Factors limiting the recruitment of Eucalyptus salmonophloia in remnant woodlands II. Post dispersal seed predation and soil seed reserves.Crossref | GoogleScholarGoogle Scholar |