

Breeding lucerne for persistence
J. H. BoutonForage Improvement Division, The Samuel Roberts Noble Foundation, 2510 Sam Noble Parkway, Ardmore, OK 73401, USA. Email: jhbouton@noble.org
Crop and Pasture Science 63(2) 95-106 https://doi.org/10.1071/CP12009
Submitted: 9 January 2012 Accepted: 8 March 2012 Published: 17 April 2012
Journal Compilation © CSIRO Publishing 2012 Open Access CC BY-NC-ND
Abstract
Cultivated lucerne is the most widely grown forage legume in pastoral agriculture. Persistence is critical for most pastoral production systems and its definition includes concepts of productivity, but maintenance of adequate plant numbers is essential. There were three important eras in lucerne persistence breeding: species introduction leading to local varieties and land races (adaptation), development of multiple pest-resistant, autumn dormancy-specific cultivars, and introducing complex traits and the use of biotechnologies. Today’s persistent cultivar needs, at a minimum, adaptation, proper autumn dormancy, and targeted pest resistances. Adding complex, ‘persistence-limiting’ traits to these minimum base traits, such as tolerance to grazing, acid, aluminum-toxic soils, and drought, is successfully being achieved via traditional selection, but biotechnologies and inter-specific hybridisations are also being employed in some cases. The main issues around biotechnologies are public perception and regulatory issues which continue to hamper transgene deployment while genetic marker programs need to lower costs and concentrate on successful application. There is not one persistent ‘ideotype’ that will fill all situations, but specific ones need to be developed and targeted for geographies such as the subtropics. Finally, breeders need to understand what persistence traits lucerne producers are willing to pay a premium to obtain.
Additional keywords: acid soils, alfalfa, drought, grazing, Medicago, pest resistance.
Introduction
Cultivated lucerne (aka alfalfa), Medicago sativa L. ssp. sativa, is the most important and widely grown forage legume in the world with ~32 million hectares estimated to be cultivated mainly in the temperate regions of both the northern and southern hemispheres (Bouton 2001; Irwin et al. 2001). Cultivated lucerne is an autotetraploid with 2n = 4x = 32 (Stanford 1951). Most Medicago species originated in the Caucasus, north-western Iran, and north-eastern Turkey with M. sativa believed to be a complex of several perennial subspecies, both diploids and tetraploids, which possess the same karyotype (Quiros and Bauchan 1988). Of these subspecies, M. sativa ssp. sativa, M. sativa ssp. falcata, and M. sativa ssp. glutinosa are inter-fertile tetraploids while M. sativa ssp. coerulea is reported to be the diploid progenitor to cultivated lucerne. Diploids are also found among M. sativa ssp. falcata.
Lucerne produces more protein per hectare than grain or oilseed crops, making it highly desirable for hay production and pasture for livestock (Barnes et al. 1988). Some important specific characteristics of cultivated alfalfa, which enhance its position as one of the most widely used forage crops, are listed as follows (Bouton 2001): (i) high nutritional quality for livestock (protein, energy, vitamins, and minerals), (ii) high biomass production (the record yield for one hectare of lucerne is over 22 400 kg ha–1 of forage), (iii) ability to fix atmospheric nitrogen (N2), (iv) wide adaptation to various environments, (v) improvement of soil tilth when growing the crop (excellent basis for sustainable agricultural systems), (vi) utility as a model system for genetic studies of autotetraploid species, and (vii) its ease of use with the new biotechnologies.
Lucerne’s traditional role is as a hay, silage, pasture and dehydration crop. New uses include sprouts for salads, nutritional supplements for human diets, a bioenergy feedstock, a bioremediation system for removal of harmful nitrates, a source of pulp for paper manufacturing, and a ‘factory’ for production of industrial enzymes (Bouton 1996a).
In the past, lucerne was a key forage species in Australia because of its wide adaption range (Clements et al. 1984; Clements 1989; Gramshaw et al. 1989; Irwin et al. 2001). Due to its commercial value and high management requirements, coupled with the restriction of seed importation, lucerne also experienced a sustained breeding effort in Australia (Clements et al. 1984; Clements 1989; Irwin et al. 2001). Recently, when 91 perennial legumes and herbs representing 47 species in 21 genera were evaluated over 3 years for herbage production, persistence and the potential to reduce ground water recharge at sites in New South Wales, South Australia and Western Australia, lucerne was reported to be the best performing species across all sites (Li et al. 2008). Therefore, lucerne’s role as the best performing forage legume in Australian pastoral agriculture has been, and continues to be, high. This role as a major crop should continue to increase the need for breeding new cultivars for the diverse and challenging Australian environments (Venkatanagappa 2008).
Pastoral agriculture, and the forage crop improvement programs that underpin it, is unique in world agriculture. Final breeding products are generally consumed by livestock, so their economic value to man, although high, is mainly indirect (Bouton 2007). Direct value is accomplished when governed by per unit value of the harvested product (e.g. ‘cut-and-carry’ commercial hay sold into the open market or seed production also for sale directly into the market), or when planted for conservation purposes. The interesting aspect about lucerne is its dominant role across the entire range of pastoral agricultural production systems due to an ability to impart economic value to all of its direct and indirect uses.
Persistence in pastoral agriculture
Pastoral acreage is dominated worldwide by native range and low input, extensively managed systems, which are mainly polycultures and not monocultures as found for the major grain crops. Animal grazing imparts a unique form of biting and shredding-type defoliation, along with defecating, urinating, and treading effects on the forage crops and makes any pastoral environment challenging. In a general sense, developing new species, as well as cultivars within those species, requires an assessment of the effect of the plants on the animals, but important for this review, an assessment of the effect of the animal on the plant. Therefore, traits like dependability and persistence are critical for most pastoral systems, and even for intensively managed, high production, monoculture perennial systems, have their place as better persistence allows one to reduce initial establishment costs via proration over years.
In addition to basic genetics and breeding principles, forage improvement programs are also governed by range, forage, and pasture management principles in order to have desired impact. This too is a very unique feature of forage breeding found rarely in breeding the major grain crops. Diversity of species also makes pastoral agriculture unique. The American Seed Trade estimated in 2005 that seed of 5500 varieties representing 1500 grass and 4000 legume species were produced for sale annually in the USA under their ‘forage seeds’ category and over 300 varieties included in their ‘alfalfa’ estimates (Bouton 2007). Relative numbers can be extrapolated for Australasia. Therefore, based on world seed sales, one can definitely identify some major forage legume species such as lucerne and white clover.
Another unique aspect of pastoral agriculture is its complicated economic system; due mainly to the indirect value of the final saleable products. With few exceptions, forages are relegated to poorer soils where land prices are low. The tendency then is to undervalue their contribution to the final saleable product.
As a general rule, grasses are more persistent than legumes. Therefore, in pastoral agriculture, it is the grasses that supply the needed persistence and dependability to the overall system; whether at the farm level or at the individual pasture level. The role of the legume is to replace N fertiliser and supplement nutrition (protein and digestible energy) for the grass base. Finally, perennial species are used where risk can be minimised at the expense of animal performance, and annual systems used where short-term animal production and performance requirements are high and more risk can be accepted. Again, lucerne is unique in that it is a perennial that can be used for both of these production outcomes.
The Trilateral Workshop
An important publication on legume persistence is the Proceedings of the Trilateral Workshop held in Honolulu, Hawaii, on 18–22 July 1988. It was published as a book by the American Society of Agronomy Tri-Societies under the title ‘Persistence of Forage Legumes’ (Marten et al. 1989). This workshop is a good synopsis for where this general issue was in 1988 for the three countries involved, Australia, New Zealand, and USA, and especially for the main country at the centre of the Farrer Review, Australia.
Each individual definition or concept of persistence was a function of: (i) the country or even the region within the country, (ii) the animal system involved and their nutritional needs, (iii) economics, (iv) the prevailing production system, (v) ecological situation and pest problems, (vi) climate, (vii) edaphic situation, and (viii) availability of high nutritional feed supplements. Therefore, persistence seemed to be easily understood as a research objective, but was the most difficult of all pasture production concepts to reach a consensus.
Thus for the workshop participants, the main debate was whether to base persistence on the New Zealand view of stability and productivity in any given environment or the USA view of survival of plant material (Marten 1989). A good compromise was offered by the Australian R. J. Clements: ‘… persistence can include concepts of productivity, but the maintenance of adequate plant numbers is the essential criterion’.
As summarised by the workshop participants, there were 20 primary causes of poor persistence listed in non-prioritised order (Marten 1989). Therefore, these causes are many and span a range from mostly tangible ones to some intangible ones as far as extrapolation to concrete breeding objectives and specific persistence traits is concerned. Some tangible ones were poor seedling vigour or premature germination, lack of hard seed, inadequate seed production of self-reseeding legumes, poor competitive ability with associated grasses or invaders, poor shade tolerance, poor resistance to cumulative pest stress loads, poor drought tolerance, poor heat or cold tolerance, poor tolerance to anaerobic conditions, poor tolerance to frequent defoliation or to heavy grazing pressures, lack of rhizomes, stolons, or protected growing points, inadequate inoculation with rhizobia or low N2 fixation, allelopathy, lack of ‘farmer-resistant’ cultivars, inadequate root systems, and lack of overall ‘plasticity’ of current germplasm.
Lucerne cultivar development
Most modern lucerne cultivars are synthetics. The current approach is to identify the individual parental genotypes which contain the traits of interest using the various screening procedures. Most synthetics are broad-based usually containing more than 100 parents. This trend of using large numbers of parents began in the early 1970s and was based on the theoretical considerations of alfalfa’s autotetraploid genetics indicating inbreeding was reduced by large parental numbers via achievement of maximum heterozygosity for intra-allelic interactions (Busbice and Wilsie 1966; Hill 1987). A reduction of complementary gene interactions rather than intra-allelic interactions was later proposed to be responsible for inbreeding depression in alfalfa (Bingham et al. 1994).
There were three important eras in lucerne breeding in the USA (Bouton 2001). First, was the introduction of the species into a defined geography that led to local varieties and land races which in turn were used as base populations in initial cultivar development. Second, was the concentration on development of multiple pest-resistant and autumn dormancy-specific cultivars. Third, and recently, was the breeding for complex traits and the development and use of biotechnologies as breeding tools.
In Australia, similar eras were recorded, but described as (i) the time up until the mid 1970s that was predominated by introductions but especially by the proliferation and use of the cultivar Hunter River; (ii) the time since the 1970s dominated by development and use of multiple pest-resistant cultivars; and (iii) the recent time that is placing an emphasis on new breeding objectives, use of technologies, and introduction of new germplasm (Irwin et al. 2001).
Land races and local varieties
An important development during the past century for lucerne breeding programs was the use of unique introductions such as winter hardy spp. falactas in North America. Local varieties and land races served as base populations for cultivar improvement in all geographies (Bouton 2001; Irwin et al. 2001). The approach capitalised on natural selection for adaptation and demonstrates the important role adaptation plays in the success of lucerne cultivars. It was also responsible for overcoming the main persistence-limiting problem in the early days such as winter hardiness and cold survival. In Australia, it was likewise reported that most early cultivars were also developed by collecting, testing, and commercialising natural ecotypes again demonstrating the value of natural selection in developing adaptative, persistent cultivars (Clements 1989; Irwin et al. 2001). Therefore, in the Americas, cultivars like Vernal and CUF101 became widely used, while in Australia, it was Hunter River.
Multiple pest-resistant, dormancy-specific cultivars
Biotic factors such as diseases, insects, nematodes, weed pressure, and vertebrates are all known to negatively alter lucerne persistence (Beuselinck et al. 1994). Of these factors, disease and insect pests, and to a lesser extent nematodes, have been studied the most as seen from the last monograph dedicated solely to lucerne where whole chapters are devoted to their biology, control, and resistance breeding approaches (Hanson et al. 1988).
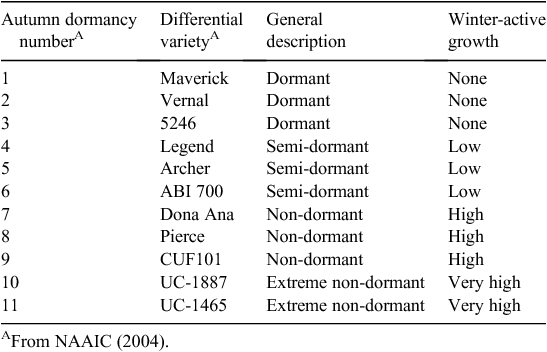
Since insect and disease pests are numerous in lucerne (Table 1), development during the late 1900s of modern cultivars with the proper autumn dormancy (Table 2), along with a broad genetic base for pest resistance was pursued in many countries within adapted germplasm pools (Bouton 2001). Of great value during this time, was the creation of the North American Alfalfa Improvement Conference (NAAIC; www.naaic.org, accessed 9 March 2012) and its role in the development and use by breeders worldwide of the standard tests developed for screening and selection of various diseases, insects, and nematode pests (NAAIC 2004). These standard tests, along with seed for their differentials (checks), are still widely used in current cultivar development programs.

|
|
Incorporation of multiple pest-resistant, autumn dormancy-specific traits definitely enhanced persistence and performance for specific regions as seen for lucerne performance trials that still compare new cultivars with older cultivars in the USA (www.naaic.org/Resources/yields.html, accessed 9 March 2012). These trials demonstrate both high yields with longer stand life for the newer cultivars due mainly to improving persistence via proper autumn dormancy and multiple pest resistance. For just one specific example, in the University of California 2010 trials (Putnam et al. 2010), recently released cultivars outyielded the Vernal and CUF101 checks by as much as 57% with an average increase of ~25% for most trials. Likewise, in eastern Australia, cultivars were successfully developed from Hunter River for resistance to aphids and anthracnose and phytopthora disease (Clements et al. 1984). These cultivars were later reported to outyield Hunter River by over 300% in disease-infested trials therefore leading to the conclusion that incorporation of multiple pest resistance has substantially increased the productivity and persistence of the crop in that region (Irwin et al. 2001).
Complex traits and the use of biotechnologies
Using the NAAIC ‘Standard Tests to Characterise Alfalfa Cultivars’ manual as a guide (NAAIC 2004), one sees screening tests for the following complex traits being added in recent times: forage production under salt stress in 1991, salt tolerance of germinating seedlings in 1991, winter survival in 1993 and revised in 2003, acid detergent fibre, neutral detergent fibre, and relative feed value in 1995, grazing tolerance in 1998, multifoliolate leaf expression in 1995, and standability or lodging resistance in 2006. Cultivars with these traits are now being sold into the seed market especially in the USA.
There are two general biotechnologies that show great promise for use in forage crop breeding programs; transgenics and genomics. However, from the author’s experience, everything with the new biotechnologies is more expensive when compared with the old ‘seed, feed, and fertiliser’ model of past forage breeding and agronomy programs; not just marginally expensive, but 2-fold more expensive. This high cost, especially the development and personnel costs, has been made somewhat bearable due to government investment, but for most private forage breeding companies, it is a non-starter from a financial standpoint. Costs are also responsible for the confinement, at least at this writing, of these technologies to a limited number of forage legume species that have the most profit potential with lucerne being among them (Bouton 2007).
The transgenic approach has already demonstrated success in introducing genes which have already made many important row crops resistant to insects, viruses, and herbicides. These successes also demonstrate the power of these transgenes to transform agricultural management. Although there were severe legal issues during the deregulation process, the recent commercialisation of herbicide resistance via the Roundup Ready gene is an example that transgenes can be commercially introduced into lucerne (Anonymous 2011). If weed pressure is a detriment to persistence during stand establishment or afterwards due to weed invasion, then this transgenic trait is an example that transgenics can be used to improve lucerne persistence. The deployment of this transgene also demonstrates that lucerne is now the major forage crop of choice for future transgenic traits.
The main uses of genomic technologies are assessment and characterisation of genetic diversity and increasing the speed and efficiency of the breeding process via marker-based selection (Irwin et al. 2001; Brummer et al. 2007). Development of molecular markers to select for simple traits appears easy, but selection for complex traits governed by quantitative trait loci (QTL), especially those surrounding persistence, has proven as difficult as they are important, and accurate phenotyping still remains the most difficult aspect (Bouton 2010). For example, QTL have been identified that explain up to 63% of the variation for anthracnose (Mackie et al. 2007), but only 8–17% of the variation (depending on the linkage group) for Stagnospora root and crown rot (Musial et al. 2007). Although these studies demonstrate the potential of using genotyping via molecular markers as a breeding tool for selection for resistance to these two pests, it also shows the need for more intensive mapping studies to actually identify all of the QTL needed to ensure genotyping will be better than conventional phenotyping.
Therefore, the main issues for the continued use of biotechnologies are that the most promising transgenic traits remain problematic due to regulatory issues; while current genetic marker programs have been slow in lowering development costs, but especially in their application for the complex persistence traits. However, for lucerne, the cost of producing biotechnology-based cultivars is economically attainable now, but will continue to be governed in the future by cost; especially by regulatory costs and public acceptance. For other legume species of lower economic value, it may be difficult to justify costs; hopefully, technologies developed for the major species will have some carry-over value for use in those species.
Breeding for persistence in lucerne
Reseeding, survival of the main plant, and possession of stolons or rhizomes that allow revegetation are the main morphological traits to ensure plant persistence (Beuselinck et al. 1994). Species like white clover with multiple strategies such as ability to both reseed and possession of a high number of stolons (Caradus and Williams 1989) have a persistence advantage especially compared with crown formers such as lucerne where survival of the established plant is critical (Leach and Clements 1984). Natural reseeding is also poor in lucerne, leaving stand persistence dependent on survival of a high number of the initial established plants. As discussed above in the ‘Multiple pest-resistant, dormancy-specific cultivar’ section, the most important development to increase plant and stand persistence in lucerne was made possible via selection and use of multiple pest-resistant cultivars. Therefore, to develop persistent lucerne cultivars, one must increase individual plant survival in the target environment by building on a good base of elite, multiple pest-resistant, autumn dormancy-specific germplasm and adding ‘persistence-limiting traits’ targeted for that specific environment. Three of the main persistence-limiting traits on a global basis are drought tolerance to mitigate problems projected due to climate change, grazing tolerance for use in intensively grazed pastures, and tolerance to acid, aluminum (Al)-toxic soils; the latter two being critical for the subtropics (Leach and Clements 1984). These three problems are now reviewed due to global importance, but also because of varying success in achieving tolerant cultivars. Approaches for these three traits also demonstrate a contrast in their breeding methods with traditional selection protocols being successfully used to develop grazing-tolerant cultivars, with protocols that moved from traditional to more biotechnology-based protocols being used to achieve important, but as yet preliminary success from a cultivar development perspective, for acid, Al-toxic soils tolerance and drought tolerance. Finally, the use of inter-specific hybridisation will also be considered.
Grazing tolerance
Lucerne as a grazing crop is limited due to rapid stand loss especially under intensive and long-term grazing (Leach and Clements 1984; Lodge 1991; Smith et al. 2000). Breeding objectives to enhance stolon or rhizome production and reseeding ability are a good initial strategy to enhance persistence (Beuselinck et al. 1994) and were the main reasons for the development of lucerne cultivars with the ‘creeping-rooted’ trait (Heinrichs 1963). However, grazing trials with these cultivars showed poor results for increased grazing persistence (Leach 1978; Gramshaw et al. 1982). The trait lacked expression in most environments and was also found to be associated with low herbage yield further reducing its use in breeding (Busbice and Hanson 1969). Although another Italian report showed similar poor persistence results for the creeping-rooted trait, ‘rhizomatous’ genotypes were felt to be more persistent in comparison (Piano et al. 1996). In follow-up research by the same group, it was concluded that it was the ‘prostrate’ types (with rhizomatous types found among these) that were the most grazing persistent types, and some ‘semi-erect’ types were reported to achieve a good balance between yield and grazing tolerance (Pecetti et al. 2008). Similar results reported prostrate types, especially those with wide crowns, demonstrated the best tolerance under rangeland conditions in Northern Great Plains region of the USA (Berdahl et al. 1989) and also under sheep grazing in Australia, but again, some tolerance could be found among upright, even winter-active, types in those studies (Humphries et al. 2001). Another similarity between these two studies was the role that ‘adaptation’ to the target environment played in achieving positive results.
Grazing tolerance, pest resistance and winter survival are all considered to be the major interacting factors affecting lucerne stand persistence under grazing (Lodge 1991; Smith et al. 2000). However, it was the association of grazing with diseases, especially during prolonged grazing in waterlogged or moisture-stressed conditions that was felt to be responsible for lucerne stand decline during grazing (Lodge 1991). Cultivars with high levels of disease resistance, however, are not necessarily grazing tolerant (Smith and Bouton 1993). Therefore, it was concluded that ‘grazing tolerance’ was a primary trait affecting stand persistence under grazing, and disease resistance was important, but secondary (Smith et al. 2000).
By merging the persistence definition from the Trilateral Workshop with the concept of individual plant survival, lucerne grazing tolerance is defined as ‘the maintenance of adequate plant numbers in the grazing system desired by the grazier to produce the desired productivity of animal gains or hay or silage yields for an economically sustainable timeframe’.
The development and use of Alfagraze (Bouton et al. 1991), and subsequent cultivars that followed using similar selection protocols (Moutray 2000), fit this definition and demonstrated that grazing tolerance could be achieved with good yield (‘dual purpose’), proper autumn dormancy, and pest resistance. This cultivar increased interest and use of lucerne for all grazing situations (Hoveland 1992; Henning 2000).
The procedure used to develop Alfagraze and other grazing-tolerant cultivars incorporated the selection of genotypes to be used as parents for new synthetics, or to composite the next population for further selection, after intensive grazing with continuous stocking by beef cattle (Fig. 1) (Smith et al. 2000). It created an anomaly because it imposed a situation different from current management standpoints of never recommending overgrazing lucerne for long durations with the common breeding approach for most pest resistances of exposing all plants equally to high pathogen inoculum levels and insect loads (the cattle can be viewed as big aphids or bacteria) in order to prevent ‘escapes’.

|
Based on the results with Alfagraze and other research documenting successful use across a range of germplasm from different dormancy groups (Smith and Bouton 1993; Moutray 2000), a standard test to screen lucerne cultivars for grazing tolerance has been included in the NAAIC ‘Standard Tests to Characterise Alfalfa Cultivars’ manual (Bouton and Smith 1998). Again, the test relies on overgrazing during screening and is not intended to be used as a grazing recommendation for producers (Fig. 1). Likewise, reselection and inter-mating of surviving plants after conducting the test with any base germplasm increased grazing tolerance in the selected populations as measured by procedures identical to the standard test (Smith and Bouton 1993; Humphries et al. 2001; Pecetti et al. 2008). The better survival of cultivars from this selection procedure under the three main management strategies, continuous stocking, rotational stocking, and hay cutting, also indicated its ability to increase general persistence via higher plant survival (Bouton and Gates 2003).
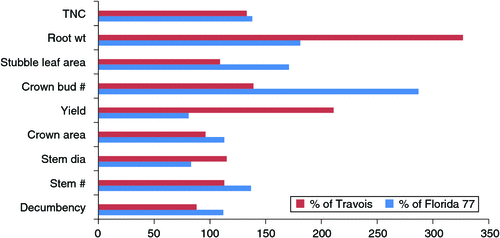
With Alfagraze, researchers also had a differential check to investigate individual morphological and physiological characteristics important for grazing tolerance. Research compared Alfagraze to other distinct, non-grazing-tolerant cultivars for these characteristics (Brummer and Bouton 1991, 1992). This research is summarised in Fig. 2 where Alfagraze’s response to frequent defoliation and/or intensive grazing for each characteristic is calculated as % of two distinct, non-tolerant cultivars; one an autumn dormancy 1 prostate type (Travois) and the other an autumn dormancy 8, narrow crowned upright type (Florida 77). Characteristics that showed substantial increase above or below 100% were Alfagraze’s larger roots that contained more total non-structural carbohydrates along with a much higher number of crown buds when compared with both checks. Decumbency score for Alfagraze showed it more prostrate than Florida 77, but a bit less prostrate than Travois, leading possibly, to resultant higher herbage yield compared with Travois and more like that of Florida 77. Alfagraze’s ability to maintain a higher stubble leaf than Florida 77, and one more like Travois, was another notable characteristic.
|
The indirect results of selection for grazing tolerance were increased resistance to Phoma crown rot, reduced resistance to blue alfalfa aphid, and little changes in other pest and disease resistance levels (Moutray 2000). Autumn dormancy may also be affected by selection for grazing tolerance because after two cycles of selection under grazing, selected populations became more autumn dormant (Bouton et al. 1998). Similarly, in Australia, persistent lucernes had long stems in the summer and short stems in the winter (Boschma and Williams 2008); stem growth patterns indicative of autumn-dormant plants.
Two recent trends with the grazing tolerance trait are also noteworthy. First, the trait was extrapolated to provide ‘traffic tolerance’ with better survival found after the effects of wheel traffic imposed by hay harvesting equipment (Lawton 2002); and second, the production and current commercialisation of two new Alfagraze-type cultivars, Alfagraze 300RR and Alfagraze 600RR, which contain the Roundup Ready trait with an expanded pest resistance package and in the 3 and 6 autumn dormancy categories, respectively (Bouton et al. 2006).
Acid, Al-toxic soils
Soil acidity is common to soils where rainfall is high enough to leach appreciable amounts of exchangeable bases from the soil surface layers with this leaching removing the buffering capacity of the soil, encouraging acidity by percolating organic acids into the soil profile, thereby causing a drop in pH, which leads ultimately to Al becoming soluble in the soil in the toxic Al3+ or Al(OH)2+ forms (Brady 1974). When soil pH is moved towards neutrality with liming, the toxicity of Al is suppressed by changing to less toxic forms such as Al(OH)0. Al can also complex with phosphates, sulfates, or organic matter components such as tartrates or citrates at any pH and be converted to non-toxic forms. The most common effect of Al on plant growth is the reduction of root elongation and proliferation (Fig. 3a), thereby leading to poor water and nutrient extraction.

|
Of the problems facing lucerne growers, acid, Al-toxic soils are the most widespread and limiting of all because lucerne is very sensitive to these conditions and acid, Al-toxic soils are found in large acreages on every continent (Devine et al. 1990; Bouton and Parrott 1997). Although liming and fertilisation are practiced to overcome these conditions, for many developing countries it is a prohibitive expense. Even where liming is practised, subsoils remain acidic; thus, the plant’s ability to penetrate the soil profile deeply and extract water and nutrients is reduced. In fact, subsoil liming was found to increase alfalfa yield by almost 50% over conventionally limed controls indicating that deeper rooting achieved through an ability to overcome subsoil toxicity has a significant effect on both stand life and productivity (Bouton et al. 1986; Sumner et al. 1986).
Plant breeding programs were pursued to develop alfalfa germplasms tolerant to these conditions. The screening and selection protocols employed a direct use of acid, Al-toxic soil and whole-plant expression was mainly based on herbage yield and/or root growth to identify tolerant genotypes to use as parents for synthetics (Devine et al. 1990; Dall’agnol et al. 1996); although later, screening in cell culture (Parrot and Bouton 1990; Bouton and Parrott 1997) and solution culture (Zhang et al. 2007) were also found to be useful as screening protocols.
Using soil screening procedures at the University of Georgia (USA), selection within cultivated alfalfa cultivars achieved some success in improving alfalfa’s tolerance to acid soils (Bouton and Sumner 1983; Bouton et al. 1986; Bouton and Radcliffe 1989; Hartel and Bouton 1989, 1991). The Georgia Aluminium Tolerant (GA-AT) germplasm developed during this research was the best result.
The GA-AT germplasm was found to possess better acid soil and Al tolerance than the most appropriate checks as determined by plant growth and N-fixing capacity when grown in both greenhouse (Hartel and Bouton 1989) and field soil conditions (Hartel and Bouton 1991) and to root deeper and extract more subsoil moisture during periods of limited rainfall (Bouton and Radcliffe 1989). This tolerance was also seen at the cellular level, as calli derived from several GA-AT genotypes showed less depression when grown in an Al-toxic medium than the check (Parrot and Bouton 1990). Finally, the USDA core collection plant introductions (PI) were tested against GA-AT and none demonstrated tolerance significantly (P < 0.05) better than GA-AT, indicating no source of high impact tolerance suitable for release as a commercial cultivar was currently available in the primary-cultivated, tetraploid alfalfa gene pool at that time (Bouton 1996b).
These results suggested that the trait is genetically complex. It also showed that use of tolerant Rhizobia is also critical adding another level of complexity when developing tolerant plant germplasm (Hartel and Bouton 1989, 1991). Recent work also found similar complexity, but the existence of significant general combining ability variances for root growth in solution culture suggested phenotypic recurrent mass selection to pyramid desirable Al-tolerant genes, focusing on parental lines and/or elite individual plants expressing long regrowth roots, should be feasible (Zhang et al. 2007). From the response of GA-AT, it is also appropriate to question why it was not commercialised as a tolerant cultivar. The answer lies in the fact that its annual dry matter yields achieved in acid soils was still only 20–30% of its yield compared with it being grown in the same soil conventionally limed (Hartel and Bouton 1991) For the farmer, surface liming to increase yield 5-fold is still a very economical undertaking. Therefore, because of genetic complexity, as well as no availability of tolerant cultivars from whole-plant selection, research efforts evolved into the use of biotechnologies (Bouton and Parrott 1997).
In the initial technology-based approach, one genotype was identified from a diploid spp. coerulea PI (PI464724 from Turkey), which was extremely Al tolerant. In several assays it grew as well in tissue culture with Al as in the absence of Al (Sledge et al. 2002). QTL mapping identified several genomic regions associated with Al tolerance in this diploid genotype based on callus assays (Sledge et al. 2002; Narasimhamoorthy et al. 2007). This genotype therefore possessed a unique source of Al tolerance genes, which had promise for being tagged and transferred via molecular markers into cultivated, tetraploid genotypes and finally becoming parental material to develop tolerant cultivars. However, to accomplish this, the genes needed initially to be transferred from the diploid to the tetraploid level. A diploid by tetraploid crossing method taking advantage of 2n eggs in the diploid parent (Bingham 1990) was used to accomplish this with several tetraploid hybrids produced. One of the tetraploid genotypes, Altet-4, was found to contain better whole-plant expression (Fig. 3b) as well as QTL alleles for Al tolerance (D.-M. Khu, R. Reyano, P. X. Zhao, J. H. Bouton, E. C. Brummer, M. J. Monteros, unpubl.). Initial mapping reported some of these QTL are detected in multiple environments, but others were environment-specific, suggesting that multiple mechanisms of Al tolerance exist (R. Reyano, D.-M. Khu, J. H. Bouton, M. J. Monteros, E. C. Brummer, unpubl.).
A second approach used transformation technology and was based the hypothesis that overexpression of organic acids in the roots may sequester and neutralise the toxic Al ions. Tobacco and papaya plants overexpressing a bacterial citrate synthase (CS) gene exhibited citrate overproduction and enhanced tolerance to Al (de la Fuente et al. 1997). Lucerne was similarly engineered by introducing the CS gene controlled by both a constitutive and a root-specific promoter (Barone et al. 2008). Several transgenic lucerne plants were produced, but when several assays were performed, including those examining Al exclusion, the only demonstrable enhanced Al tolerance was in the soil growth assays where a few plants demonstrated a moderate level of tolerance. In another study with lucerne, overexpression of malate dehydrogenase induced a significant increase of the concentration of malate and other organic acids in tissues of some transgenic lucerne lines, resulting again in a moderate level of Al tolerance (Tesfaye et al. 2001). This approach of overexpressing organic acids remains promising, but higher levels of Al tolerance will need to be achieved.
Drought tolerance
Drought tolerance is a trait that is always associated with persistence and is an inherent trait in lucerne, and one that has historically been associated with the crop (Barnes et al. 1988). This view was supported recently during the 2006–07 droughts in South Australia where established lucerne stands showed some stand decline, but were still able to maintain acceptable ground cover (Marshall et al. 2008). An even more dramatic response was seen after the record 2011 summer heat wave and corresponding drought in the southern Great Plains area of USA where lucerne paddocks showed complete autumn recovery while the adjacent tall fescue paddocks did not (Fig. 4).

|
Although lucerne is a drought-tolerant species, some of its acreage is produced under irrigated dryland conditions where reduced rainfall limits productivity. Therefore, either increasing lucerne’s ability to be more productive under limited rainfall or to demonstrate better water-use efficiency are being investigated. To this end, gene candidates for drought tolerance are now being expressed in lucerne. One of these is the WXP1 transgene responsible for production of cuticular wax and where ‘proof of concept’ experiments demonstrated that insertion of WXP1 increased lucerne’s ability to be productive, and even recover more quickly, after periods of limited water (Zhang et al. 2005).
In a related genomics project, research is underway to identify genetic mechanisms associated with drought tolerance within cultivated lucerne. Initial results indicated that molecular markers associated with yield under drought conditions were identified and they should be useful for selecting genotypes with an ability to yield better under water-limiting conditions (Han et al. 2008).
Using inter-specific hybrids
The three main driving forces in the evolution of higher plants are inter-specific hybridisation, mutation with Mendelian segregation, and polyploidy (Stebbins 1950). These same driving forces make up the basic approaches used by most plant breeders in the modern era with selection for natural mutations being the most used, but inter-specific hybridisation, especially for clonally propagated hybrids, also achieving some success in forage breeding (Bouton 2007).
Using related perennial Medicago species as novel sources of new and unique traits is therefore important to improve lucerne performance and persistence via inter-specific hybridisation with cultivated lucerne. Use of spp. coerulea genotype mentioned in the ‘Acid, Al-toxic soils’ section (see above) is a good example of this approach. However, an important example with commercial implications was the incorporation of glandular hairs from M. glandulosa and M. prostrata with the resulting stable populations demonstrating resistance to potato leaf hopper and improved nutritive quality (Sorensen et al. 1994; Elden and McCaslin 1997). There are now important cultivars in the commercial seed trade with a practical level of resistance to potato leafhopper (see http://m.hayandforage.com/news/farming_hopperfighting_alfalfas_job, accessed 9 March 2012).
Another recent example of the use of inter-specific hybridisation includes using M. sativa × M. arborea asymmetric hybrids and highly winter-active Omani landraces to produce new high yielding experimentals for potential use in the subtropics (Irwin et al. 2010).
Conclusions
Breeding goals to increase lucerne persistence are best defined when based on the ‘reference population of environments and the reference population of species and genotypes’ fundamental concept learned in basic plant breeding classes. For example, the greatest potential for lucerne expansion globally, and expansion in Australia particularly, is to increase its use in the subtropics. The crop has always been subject to ‘catastrophic losses’ in these geographic zones due to root and crown diseases, periodic high rainfall leading to saturated soils followed (ironically) by periodic drought, high summer temperatures, competition from volunteer aggressive grasses, and poor grazing management (Leach and Clements 1984). The subtropics and tropics also contain the highest concentration of acid, Al-toxic soils with over 1.6 billion ha estimated worldwide (Sanchez and Salinas 1981). So, for this example, the reference population of environments would be subtropical climates and soils in a specific geography while the reference population of genotypes could come from the different highly winter-active, multiple pest-resistant, grazing-tolerant, and acid-tolerant elite cultivars and breeding populations now available.
It is also apparent that many persistence-limiting traits are genetically complex requiring very specific screening and selection protocols, or even the use of the biotechnologies, to increase precision where normal genetic gains are incremental. However, biotechnologies should be considered only where its high development and regulatory costs can be borne.
Is there a persistent lucerne phenotype or ideotype?
The easy answer to this question is probably ‘no there is not, but different types predominate depending on the natural and managed environment’. For example, in intensively grazed paddocks, a plant that occupies space, and prevents other plants from occupying the same space, is at an advantage. It must occupy space and at the same time protect its top-growth (its photosynthetic apparatus) from herbivory. Therefore, it was not surprising that grazing tolerant plants that were prostrate or decumbent in their growth habit, had large roots and crowns, yet maintained both a high carbohydrate energy pool for growth, but at the same time a high leaf area in the low growing stems or crowns in order to replenish its energy pool, were found to be the predominate type (Fig. 2). Unfortunately, just due to the concept of harvest index, all these grazing persistence traits lead to low herbage dry matter yield. So, it is not surprising that the highest hay yields are found in well managed, less stressful conditions (good soils, with long growing seasons) with narrow crowned and winter-active plants that regrow quickly by rapidly using, and then replenishing, their energy pool.
In summary, breeding programs to increase lucerne persistence must be well defined with good base germplasm, long-term funding, and structured around a specific region’s animal production systems, climate, soils, and especially, accepted forage management practices. Finally, understanding what producers want and need, especially how much they are willing to pay for certain persistence traits will be crucial for a lucerne cultivar development program’s commercial success. This has particular bearing on the use of biotechnologies to achieve the desired phenotype because at this writing their development costs remain high and cost effective applications poor for most breeding efforts. If producer interest and willingness to pay are not assessed properly, failure in the marketplace is likely, and the wastage of a great deal of effort and resources could probably have been avoided by simply using a cheaper forage species more adapted to the stress.
References
Anonymous (2011) Roundup ready alfalfa. Available at: www.monsanto.com/newsviews/Pages/roundup-ready-alfalfa-supreme-court.aspx (accessed 3 January 2012).Barnes DK, Goplen BP, Baylor JE (1988) Highlights in the USA and Canada. In ‘Alfalfa and alfalfa improvement’. (Eds AA Hanson, DK Barnes, RR Hill, Jr.) pp. 1–24. (American Society of Agronomy: Madison, WI)
Barone P, Rosellini D, LaFayette P, Bouton J, Veronesi F, Parrot W (2008) Bacterial citrate synthase expression and soil aluminum tolerance in transgenic alfalfa. Plant Cell Reports 27, 893–901.
| Bacterial citrate synthase expression and soil aluminum tolerance in transgenic alfalfa.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD1cXksFygsrw%3D&md5=73dd83ca8655d65ec4baee903e53182eCAS |
Berdahl JD, Wilton AC, Frank AB (1989) Survival and agronomic performance of 25 alfalfa cultivars and strains interseeded into rangelands. Journal of Range Management 42, 312–316.
| Survival and agronomic performance of 25 alfalfa cultivars and strains interseeded into rangelands.Crossref | GoogleScholarGoogle Scholar |
Beuselinck PR, Bouton JH, Lamp WO, Matches AG, McCaslin MH, Nelson CJ, Rhodes LH, Sheaffer CC, Volenec JJ (1994) Improving legume persistence in forage crop systems. Journal of Production Agriculture 7, 311–322.
Bingham ET (1990) Backcrossing tetraploidy into diploid Medicago falcata L. using 2n eggs. Crop Science 30, 1353–1354.
| Backcrossing tetraploidy into diploid Medicago falcata L. using 2n eggs.Crossref | GoogleScholarGoogle Scholar |
Bingham ET, Groose RW, Woodfield DR, Kidwell KK (1994) Complementary gene interactions in alfalfa are greater in autotetraploids than diploids. Crop Science 34, 823–828.
| Complementary gene interactions in alfalfa are greater in autotetraploids than diploids.Crossref | GoogleScholarGoogle Scholar |
Boschma SP, Williams RW (2008) Using morphological traits to identify persistent lucernes for dryland agriculture in NSW, Australia. Australian Journal of Agricultural Research 59, 69–79.
| Using morphological traits to identify persistent lucernes for dryland agriculture in NSW, Australia.Crossref | GoogleScholarGoogle Scholar |
Bouton JH (1996a) New uses for alfalfa and other ‘old’ forage legumes. In ‘Progress in new crops’. (Ed. J Janick) pp. 251–259. (American Society of Horticultural Science Press: Alexandria, VA)
Bouton JH (1996b) Screening the alfalfa core collection for acid toil tolerance. Crop Science 36, 198–200.
| Screening the alfalfa core collection for acid toil tolerance.Crossref | GoogleScholarGoogle Scholar |
Bouton JH (2001) Alfalfa. In ‘Proceedings XIX International Grassland Congress’. (Eds JA Gomide, WRS Mattos, SC d’Silva) pp. 545–547. (FEALQ, Piracicaba, SP, Brazil: Sao Pedro, Sao Paulo, Brazil)
Bouton J (2007) The economic benefits of forage improvement in the United States. Euphytica 154, 263–270.
| The economic benefits of forage improvement in the United States.Crossref | GoogleScholarGoogle Scholar |
Bouton JH (2010) Future developments and uses. In ‘Handbook of plant breeding: Volume 5. Fodder crops and amenity grasses’. (Eds B Boller, UK Posselt, F Veronesi) pp. 201–209. (Springer: New York)
Bouton JH, Gates RN (2003) Grazing-tolerant alfalfa cultivars perform well under rotational stocking and hay management. Agronomy Journal 95, 1461–1464.
| Grazing-tolerant alfalfa cultivars perform well under rotational stocking and hay management.Crossref | GoogleScholarGoogle Scholar |
Bouton JH, Parrott WA (1997) ‘Salinity and aluminium stress.’. In ‘Biotechnology and the improvement of forage legumes’. (Eds BD McKersie, DCW Brown) pp. 203–226. (CAB International: Wallingford, UK)
Bouton JH, Radcliffe DE (1989) ‘Effects of acid soil selection on agronomically important traits in alfalfa.’ In ‘Proceedings XVI International Grassland Congress’. Nice, France, 4–11 October 1989. pp. 377–378.
Bouton JH, Smith SRJ (1998) Standard test to characterize alfalfa cultivar tolerance to intensive grazing with continuous stocking. Available at: www.naaic.org/stdtests/Grazing.html (accessed 29 December 2011).
Bouton JH, Sumner ME (1983) Alfalfa, Medicago sativa L., in highly weathered, acid soils. V. Field performance of alfalfa selected for acid tolerance. Plant and Soil 74, 431–436.
| Alfalfa, Medicago sativa L., in highly weathered, acid soils. V. Field performance of alfalfa selected for acid tolerance.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaL2cXhtVahsrg%3D&md5=2a4879cd7652295e5dc5a4f341f26636CAS |
Bouton JH, Sumner ME, Hammel JE, Shahandeh H (1986) Yield of an alfalfa germplasm selected for acid soil tolerance when grown in soil with modified subsoils. Crop Science 26, 334–336.
| Yield of an alfalfa germplasm selected for acid soil tolerance when grown in soil with modified subsoils.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaL28Xhs1Omtrw%3D&md5=0f85749a7d97c66aa47f94ad294d9acbCAS |
Bouton JH, Smith SR, Wood DT, Hoveland CS, Brummer EC (1991) Registration of ‘Alfagraze’ alfalfa. Crop Science 31, 479
| Registration of ‘Alfagraze’ alfalfa.Crossref | GoogleScholarGoogle Scholar |
Bouton JH, Gates RN, Utley PR (1998) Persistence and yield among nondormant alfalfas selected for grazing tolerance. Journal of Production Agriculture 11, 314–318.
Bouton J, McCaslin M, Fitzpatrick S, Temple S (2006) Roundup Ready Alfagraze. Available at: www.naaic.org/Meetings/National/2006meeting/procedings/Bouton(oral).PDF (accessed 29 December 2011).
Brady NC (1974) ‘The nature and property of soils.’ (Macmillan Publishing Co. Inc.: New York)
Brummer EC, Bouton JH (1991) Plant traits associated with grazing-tolerant alfalfa. Agronomy Journal 83, 996–1000.
| Plant traits associated with grazing-tolerant alfalfa.Crossref | GoogleScholarGoogle Scholar |
Brummer EC, Bouton JH (1992) Physiological traits associated with grazing-tolerant alfalfa. Agronomy Journal 84, 138–143.
| Physiological traits associated with grazing-tolerant alfalfa.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaK38XktVCnsLs%3D&md5=913f97cfc7486c54908d7ce09f6c8151CAS |
Brummer EC, Bouton JH, Sledge M (2007) Biotechnology and molecular approaches to forage improvement. In ‘Forages. Volume II – the science of grassland agriculture’. 6th edn. (Eds RF Barnes, CJ Nelson, KJ Moore, M Collins) pp. 439–451. (Blackwell Publishing: Ames, IA)
Busbice TH, Hanson AA (1969) Selection for improved creeping-rooted characteristics in alfalfa. Crop Science 9, 244–246.
| Selection for improved creeping-rooted characteristics in alfalfa.Crossref | GoogleScholarGoogle Scholar |
Busbice TH, Wilsie CP (1966) Inbreeding depression and heterosis in autotetraploids with application to Medicago sativa L. Euphytica 15, 52–67.
Caradus JR, Williams WM (1989) Breeding for legume persistence in New Zealand. In ‘Persistence of forage legumes. Proceedings of a Trilateral Workshop’. 18–22 July 1988, Honolulu, Hawaii. (Eds GC Marten, AG Matches, RF Barnes, RW Brougham, RJ Clements, GW Sheath) pp. 523–537. (ASA, CSSA, SSSA: Madison, WI)
Clements RJ (1989) Developing persistent pasture legume cultivars for Australia. In ‘Persistence of forage legumes. Proceedings of a Trilateral Workshop’. 18–22 July 1988, Honolulu, Hawaii. (Eds GC Marten, AG Matches, RF Barnes, RW Brougham, RJ Clements, GW Sheath) pp. 505–521. (ASA, CSSA, SSSA: Madison, WI)
Clements RJ, Turner JW, Irwin JAG, Langdon PW, Bray RA (1984) Breeding disease resistant aphid resistant lucerne Medicago-sativa for subtropical Queensland Australia. Australian Journal of Experimental Agriculture and Animal Husbandry 24, 178–188.
| Breeding disease resistant aphid resistant lucerne Medicago-sativa for subtropical Queensland Australia.Crossref | GoogleScholarGoogle Scholar |
Dall’agnol M, Bouton JH, Parrott WA (1996) Screening methods to develop alfalfa germplasms tolerant of acid, aluminum toxic soils. Crop Science 36, 64–70.
| Screening methods to develop alfalfa germplasms tolerant of acid, aluminum toxic soils.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaK28XotVyjsA%3D%3D&md5=0c6999a8c8c8f784210ffa98043dc5e7CAS |
de la Fuente JM, Ramirez-Rodriguez V, Cabrera-Ponce JL, Herrera-Estrella L (1997) Aluminum tolerance in transgenic plants by alteration of citrate synthesis. Science 276, 1566–1568.
| Aluminum tolerance in transgenic plants by alteration of citrate synthesis.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaK2sXjslGku7Y%3D&md5=ab5ab7b9c0c9a1789a36cef93a3b829bCAS |
Devine TE, Bouton JH, Mabrahtu T (1990) Legume genetics and breeding for stress tolerance and nutrient efficiency. In ‘Crops as enhancers of nutrient use’. (Eds VC Baligar, RR Duncan) pp. 211–252. (Academic Press: San Diego, CA)
Elden TC, McCaslin M (1997) Potato leafhopper (Homoptera: Cicadellidae) resistance in perennial glandular-haired alfalfa clones. Journal of Economic Entomology 90, 842–847.
Gramshaw D, Bishop HG, Ludke DH (1982) Performance of grazed creeping-rooted lucernes on two soils in central Queensland. Australian Journal of Experimental Agriculture and Animal Husbandry 22, 177–181.
| Performance of grazed creeping-rooted lucernes on two soils in central Queensland.Crossref | GoogleScholarGoogle Scholar |
Gramshaw D, Read JW, Collins WJ, Carter ED (1989) Sown pastures and legume persistence: an Australian overview. In ‘Persistence of forage legumes. Proceedings of a Trilateral Workshop’. Honolulu, Hawaii, 18–22 July 1988. (Eds GC Marten, AG Matches, RF Barnes, RW Brougham, RJ Clements, GW Sheath) pp. 1–22. (ASA, CSSA, SSSA: Madison, WI)
Han Y, Ray IM, Sledge MK, Bouton JH, Monteros MJ, Han YH (2008) Drought tolerance in tetraploid alfalfa. In ‘Multifunctional grasslands in a changing world, Volume II: XXI International Grassland Congress and VIII International Rangeland Congress’. Hohhot, China, 29 June–5 July 2008. p.419. (Guangdong People’s Publishing House: Guangzhou, China)
Hanson AA, Barnes DK, Hill RR (1988) ‘Alfalfa and alfalfa improvement.’ (American Society of Agronomy: Madison, WI)
Hartel PG, Bouton JH (1989) Rhizobium meliloti inoculation of alfalfa selected for tolerance to acid aluminum-rich soils. Plant and Soil 116, 283–285.
| Rhizobium meliloti inoculation of alfalfa selected for tolerance to acid aluminum-rich soils.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaL1MXlsFaluro%3D&md5=fbb850757e8bfd9507a2b9c9582e48f5CAS |
Hartel PG, Bouton JH (1991) Rhizobium meliloti inoculation of alfalfa selected for tolerance to acid, aluminum-rich soils. In ‘Plant–soil interactions at low pH’. (Eds RJ Wright, VC Baligar, RP Murrmann) pp. 597–601. (Kluwer Academic Publishers: Dordrecht, The Netherlands)
Heinrichs DH (1963) Creeping alfalfas. Advances in Agronomy 15, 317–337.
| Creeping alfalfas.Crossref | GoogleScholarGoogle Scholar |
Henning JC (2000) On-farm use of alfalfa for grazing. In ‘Proceedings/Reports of the American Forage and Grassland Council, 37th North American Alfalfa Improvement Conference’. Madison, Wisconsin, 16–19 July 2000. pp. 359–363.
Hill RR, Jr. (1987) Alfalfa. In ‘Principles of cultivar development, Vol. 2’. (Ed. WR Fehr) pp. 11–39. (Macmillan: New York)
Hoveland CS (1992) Grazing systems for humid regions. Journal of Production Agriculture 5, 23–27.
Humphries AW, Auricht GC, Kobelt ET (2001) Developing grazing tolerant lucerne. Available at: www.regional.org.au/au/asa/2001/6/a/humphries2.htm (accessed 2 January 2012).
Irwin JAG, Lloyd DL, Lowe KF (2001) Lucerne biology and genetic improvement: an analysis of past activities and future goals in Australia. Australian Journal of Agricultural Research 52, 699–712.
| Lucerne biology and genetic improvement: an analysis of past activities and future goals in Australia.Crossref | GoogleScholarGoogle Scholar |
Irwin JAG, Armour DJ, Pepper PM, Lowe KF (2010) Heterosis in lucerne testcrosses with Medicago arborea introgressions and Omani landraces and their performance in synthetics. Crop & Pasture Science 61, 450–463.
| Heterosis in lucerne testcrosses with Medicago arborea introgressions and Omani landraces and their performance in synthetics.Crossref | GoogleScholarGoogle Scholar |
Lawton K (2002) Traffic tolerance. Available at: http://farmindustrynews.com/traffic-tolerance (accessed 29 December 2011).
Leach GJ (1978) The ecology of lucerne pastures. In ‘Plant relations in pastures’. (Ed. JR Wilson) pp. 290–308. (CSIRO Publishing: Melbourne)
Leach GJ, Clements RJ (1984) Ecology and grazing management of alfalfa pastures in the subtropics. Advances in Agronomy 37, 127–154.
| Ecology and grazing management of alfalfa pastures in the subtropics.Crossref | GoogleScholarGoogle Scholar |
Li GD, Lodge GM, Moore GA, Craig AD, Dear BS, Boschma SP, Albertsen TO, Miller SM, Harden S, Hayes RC, Hughes SJ, Snowball R, Smith AB, Cullis BC (2008) Evaluation of perennial pasture legumes and herbs to identify species with high herbage production and persistence in mixed farming zones in southern Australia. Australian Journal of Experimental Agriculture 48, 449–466.
| Evaluation of perennial pasture legumes and herbs to identify species with high herbage production and persistence in mixed farming zones in southern Australia.Crossref | GoogleScholarGoogle Scholar |
Lodge GM (1991) Management practices and other factors contributing to the decline in persistence of grazed lucerne in temperate Australia: a review. Australian Journal of Experimental Agriculture 31, 713–724.
| Management practices and other factors contributing to the decline in persistence of grazed lucerne in temperate Australia: a review.Crossref | GoogleScholarGoogle Scholar |
Mackie JM, Musial JM, Armour DJ, Phan HTT, Ellwood SE, Aitken KS, Irwin JAG (2007) Identification of QTL for reaction to three races of Colletotrichum trifolii and further analysis of inheritance of resistance in autotetraploid lucerne. Theoretical and Applied Genetics 114, 1417–1426.
| Identification of QTL for reaction to three races of Colletotrichum trifolii and further analysis of inheritance of resistance in autotetraploid lucerne.Crossref | GoogleScholarGoogle Scholar | 1:STN:280:DC%2BD2s3nsVWisA%3D%3D&md5=d3a0ab9797d487c6d5645c0ff689af3fCAS |
Marshall EM, Humphries AW, Kobelt ET, Rowe TD, Auricht GC (2008) Drought response of lucerne in South East and Mallee environments in SA. Available at: www.sardi.sa.gov.au/__data/assets/pdf_file/0003/116553/Persistence_Drought_response_of_Lucerne.pdf (accessed 30 December 2011).
Marten GC (1989) Summary of the trilateral workshop on persistence of forage legumes. In ‘Persistence of forage legumes. Proceedings of a Trilateral Workshop’. 18–22 July 1988, Honolulu, Hawaii. (Eds GC Marten, AG Matches, RF Barnes, RW Brougham, RJ Clements, GW Sheath) pp. 569–572. (ASA, CSSA, SSSA: Madison, WI)
Marten GC, Matches AG, Barnes RF, Brougham RW, Clements RJ, Sheath GW (Eds) (1989) Persistence of forage legumes. In ‘Proceedings of a Trilateral Workshop’. Honolulu, Hawaii, 18–22 July 1988. (American Society of Agronomy: Madison, WI)
Moutray JB (2000) Future of alfalfa as a grazing crop: grazing tolerance. In ‘Proceedings/Reports of the American Forage and Grassland Council, 37th North American Alfalfa Improvement Conference’. Madison, Wisconsin, 16–19 July 2000. pp. 345–350.
Musial JM, Mackie JM, Armour DJ, Phan HTT, Ellwood SE, Aitken KS, Irwin JAG (2007) Identification of QTL for resistance and susceptibility to Stagonospora meliloti in autotetraploid lucerne. Theoretical and Applied Genetics 114, 1427–1435.
| Identification of QTL for resistance and susceptibility to Stagonospora meliloti in autotetraploid lucerne.Crossref | GoogleScholarGoogle Scholar | 1:STN:280:DC%2BD2s3nsVWisw%3D%3D&md5=9294d7505d7efe8e45ed5f84b2f94173CAS |
NAAIC (2004) ‘Standard tests to characterize alfalfa cultivars.’ 3rd edn. Available at: www.naaic.org/stdtests/index.html (accessed 29 December 2011).
Narasimhamoorthy B, Bouton JH, Olsen KM, Sledge MK (2007) Quantitative trait loci and candidate gene mapping of aluminum tolerance in diploid alfalfa. Theoretical and Applied Genetics 114, 901–913.
| Quantitative trait loci and candidate gene mapping of aluminum tolerance in diploid alfalfa.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2sXitVOltrs%3D&md5=93f3219a2d7a58d85d764ee15fe15575CAS |
Parrot WA, Bouton JH (1990) Aluminum tolerance in alfalfa as expressed in tissue culture. Crop Science 30, 387–389.
| Aluminum tolerance in alfalfa as expressed in tissue culture.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaK3cXksFGqtLs%3D&md5=e46e85926f683046800b2fea090bd848CAS |
Pecetti L, Romani M, De Rosa L, Piano E (2008) Selection of grazing-tolerant lucerne cultivars. Grass and Forage Science 63, 360–368.
| Selection of grazing-tolerant lucerne cultivars.Crossref | GoogleScholarGoogle Scholar |
Piano E, Valentini P, Pecetti L, Romani M (1996) Evaluation of a lucerne germplasm collection in relation to traits conferring grazing tolerance. Euphytica 89, 279–288.
| Evaluation of a lucerne germplasm collection in relation to traits conferring grazing tolerance.Crossref | GoogleScholarGoogle Scholar |
Putnam D, Giannini C, Maciel F, Orloff S, Kirby D, DeBen C, Pattigan D (2010) California alfalfa variety trial yield results including round-up ready varieties. Available at: http://alfalfa.ucdavis.edu/+producing/variety/apr/APR302-2010data.pdf (accessed 30 December 2011).
Quiros CF, Bauchan GR (1988) The genus Medicago and the origin of the Medicago sativa complex. In ‘Alfalfa and alfalfa improvement’. (Eds AA Hanson, DK Barnes, RR Hill, Jr.) pp. 93–124. (American Society of Agronomy: Madison, WI)
Sanchez PA, Salinas JG (1981) Low-input technology for managing oxisols and ultisols in tropical America. Advances in Agronomy 34, 279–406.
| Low-input technology for managing oxisols and ultisols in tropical America.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaL38XhtVaqt7s%3D&md5=d06f72108883c0834ecf7e5900eae254CAS |
Sledge MK, Bouton JH, Dall’Agnoll M, Parrott WA, Kochert G (2002) Identification and confirmation of aluminum tolerance QTL in diploid Medicago sativa subsp. coerulea. Crop Science 42, 1121–1128.
| Identification and confirmation of aluminum tolerance QTL in diploid Medicago sativa subsp. coerulea.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD38XmsVant70%3D&md5=5da7be09e3c6ad854228f66249e7392aCAS |
Smith SR, Bouton JH (1993) Selection within alfalfa cultivars for persistence under continuous stocking. Crop Science 33, 1321–1328.
| Selection within alfalfa cultivars for persistence under continuous stocking.Crossref | GoogleScholarGoogle Scholar |
Smith SR, Bouton JH, Singh A, McCaughey WP (2000) Development and evaluation of grazing-tolerant alfalfa cultivars: a review. Canadian Journal of Plant Science 80, 503–512.
| Development and evaluation of grazing-tolerant alfalfa cultivars: a review.Crossref | GoogleScholarGoogle Scholar |
Sorensen EL, Stuteville DL, Horber EK, Skinner DZ (1994) Registration of KS224 glandular-haired alfalfa germplasm with multiple pest resistance. Crop Science 34, 544
| Registration of KS224 glandular-haired alfalfa germplasm with multiple pest resistance.Crossref | GoogleScholarGoogle Scholar |
Stanford EH (1951) Tetrasomic inheritance in alfalfa. Agronomy Journal 43, 222–225.
| Tetrasomic inheritance in alfalfa.Crossref | GoogleScholarGoogle Scholar |
Stebbins GL (1950) ‘Variation and evolution in plants.’ (Columbia University Press: New York)
Sumner ME, Shahandeh H, Bouton JH, Hammel JE (1986) Amelioration of acid soil profile through deep liming and surface application of gypsum. Soil Science Society of America Journal 50, 1255–1258.
Tesfaye M, Temple SJ, Allan DL, Vance CP, Samac DA (2001) Overexpression of malate dehydrogenase in transgenic alfalfa enhances organic acid synthesis and confers tolerance to aluminum. Plant Physiology 127, 1836–1844.
| Overexpression of malate dehydrogenase in transgenic alfalfa enhances organic acid synthesis and confers tolerance to aluminum.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD38XjtVWksA%3D%3D&md5=cbab0b4b0f08178000e32d4010732621CAS |
Venkatanagappa S (2008) Breeding lucerne for the Australian environment through a Private Public Partnership. Available at: www.naaic.org/Meetings/National/2008meeting/proceedings/Venkatanagappa.pdf (accessed 29 December 2011).
Zhang J-Y, Broeckling CD, Blancaflor EB, Sledge MK, Sumner LW, Wang Z-Y (2005) Overexpression of WXP1, a putative Medicago truncatula AP2 domain-containing transcription factor gene, increases cuticular wax accumulation and enhances drought tolerance in transgenic alfalfa (Medicago sativa). The Plant Journal 42, 689–707.
| Overexpression of WXP1, a putative Medicago truncatula AP2 domain-containing transcription factor gene, increases cuticular wax accumulation and enhances drought tolerance in transgenic alfalfa (Medicago sativa).Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2MXlt1yns7w%3D&md5=09ebf4f880f4d3ad329173c29160848aCAS |
Zhang X, Humphries A, Auricht G (2007) Genetic variability and inheritance of aluminium tolerance as indicated by long root regrowth in lucerne (Medicago sativa L.). Euphytica 157, 177–184.
| Genetic variability and inheritance of aluminium tolerance as indicated by long root regrowth in lucerne (Medicago sativa L.).Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2sXpslaqu78%3D&md5=39cc446b4bdd26dc237941a8b31f985bCAS |