

A comment on the quantitative significance of aerobic methane release by plants
Miko U. F. Kirschbaum A B H , Dan Bruhn A C , David M. Etheridge D , John R. Evans A B , Graham D. Farquhar A B , Roger M. Gifford A E , Keryn I. Paul F and Anthony J. Winters A GA Cooperative Research Centre for Greenhouse Accounting, GPO Box 475, Canberra, ACT 2601, Australia.
B Environmental Biology Group, Australian National University, GPO Box 475, Canberra, ACT 2601, Australia.
C Ecosystems Dynamics Group, Australian National University, GPO Box 475, Canberra, ACT 2601, Australia.
D CSIRO Marine and Atmospheric Research, PMB 1, Aspendale, Vic. 3195, Australia.
E CSIRO Plant Industry, GPO Box 1600, Canberra, ACT 2601, Australia.
F Ensis, PO Box E4008, Kingston, ACT 2604, Australia.
G School of Biological, Earth and Environmental Sciences, University of New South Wales, Sydney, NSW 2052, Australia.
H Corresponding author. Email: Miko@Kirschbaum.id.au
Functional Plant Biology 33(6) 521-530 https://doi.org/10.1071/FP06051
Submitted: 8 March 2006 Accepted: 28 March 2006 Published: 2 May 2006
Abstract
A recent study by Keppler et al. (2006; Nature 439, 187–191) demonstrated CH4 emission from living and dead plant tissues under aerobic conditions. This work included some calculations to extrapolate the findings from the laboratory to the global scale and led various commentators to question the value of planting trees as a greenhouse mitigation option. The experimental work of Keppler et al. (2006) appears to be largely sound, although some concerns remain about the quantification of emission rates. However, whilst accepting their basic findings, we are critical of the method used for extrapolating results to a global scale. Using the same basic information, we present alternative calculations to estimate global aerobic plant CH4 emissions as 10–60 Mt CH4 year–1. This estimate is much smaller than the 62–236 Mt CH4 year–1 reported in the original study and can be more readily reconciled within the uncertainties in the established sources and sinks in the global CH4 budget. We also assessed their findings in terms of their possible relevance for planting trees as a greenhouse mitigation option. We conclude that consideration of aerobic CH4 emissions from plants would reduce the benefit of planting trees by between 0 and 4.4%. Hence, any offset from CH4 emission is small in comparison to the significant benefit from carbon sequestration. However, much critical information is still lacking about aerobic CH4 emission from plants. For example, we do not yet know the underlying mechanism for aerobic CH4 emission, how CH4 emissions change with light, temperature and the physiological state of leaves, whether emissions change over time under constant conditions, whether they are related to photosynthesis and how they relate to the chemical composition of biomass. Therefore, the present calculations must be seen as a preliminary attempt to assess the global significance from a basis of limited information and are likely to be revised as further information becomes available.
Keywords: climate change, greenhouse gas, methane, plants, source.
Introduction
The atmospheric CH4 concentration has increased from a pre-industrial concentration of less than 700 to ~1750 ppb at present, with little change over the last five years [Ehhalt et al. 2001; see http://www.cmdl.noaa.gov/ccgg/iadv/ or http://gaw.kishou.go.jp/wdcgg.html (both verified 29 March 2006) for the most recent data]. It had been thought that all the major sources and sinks had been identified, although with significant uncertainties still remaining about the exact magnitude of each identified source and sink (Ehhalt et al. 2001; Wang et al. 2004; Frankenberg et al. 2005; do Carmo et al. 2006).
Keppler et al. (2006) recently reported that CH4 was emitted from dead and living plant material under aerobic conditions. This was a surprising finding as CH4 emission and uptake from various systems has been studied for a long time and it had always been thought that CH4 production occurs only under anaerobic conditions, such as in flooded soils, guts of ruminant animals, or during incomplete combustion in fires. Keppler et al. (2006) also attempted to use their data to provide an estimate of the possible global significance of aerobic CH4 emissions from plants. They derived a surprisingly large estimate of 62–236 Mt CH4 year–1, which suggested that the aerobic release by plants could constitute one quarter of total current global CH4 emissions.
Hence, the new findings caused a flood of media statements, partly spurred on by an opinion piece by Lowe (2006) that accompanied the original article in Nature. This opinion article implied that the new finding by Keppler et al. (2006) will require a major rethink of the global CH4 budget and a re-assessment of the value of planting trees as a greenhouse mitigation strategy.
The original study by Keppler et al. (2006) provided only scant data for extrapolation to the global scale. Nonetheless, such scaling up is required to evaluate whether the new findings might necessitate an immediate re-evaluation of current climate-change mitigation options, especially in relation to planting trees.
In this paper, we attempt to provide an estimate of the global significance of aerobic CH4 emission from plants because we believe that the approach used by Keppler et al. (2006) contained some methodological inconsistencies. In our alternative approach, we used two different methods for estimating global emissions. We also attempt to quantitatively assess the significance of aerobic plant CH4 emissions in modifying the value of planting trees for climate mitigation.
Methodology
Estimate of global methane emission
The Keppler approach
The only information currently available on aerobic CH4 emissions is that published by Keppler et al. (2006) in their paper and its accompanying information. They found that CH4 was emitted from dead and living tissue of several species, and that emission rates increased with increasing temperature or exposure to sunlight. However, we believe that their method for scaling to global emissions is inappropriate and dimensionally inconsistent. They multiplied their observed CH4 emission rates (in units of CH4 per unit of dry mass per unit of time), by estimates of net primary production (NPP in units of carbon per unit of area per unit of time) to estimate global CH4 emissions per year. As both parameters are rates, their product attains the units of CH4 per unit of time squared.
Keppler et al. (2006) used NPP as the basis of their calculations to provide an estimate of the growth of new tissue formed within a given year. This estimate of functionally active biomass, they believed, could be equated with the biomass in their measuring system. Implicit in this calculation method are the assumptions that all newly formed tissue contributes equally to total CH4 emissions and that emission rates are constant for a whole season and then cease.
However, this is likely to overestimate the amount of aerobic CH4 emission because (1) a large part of NPP is allocated to roots which do not emit CH4 at the same rate as light-exposed tissue, (2) woody material receives little light and is metabolically inert so that it is likely to also emit less CH4 than light-exposed and more metabolically-active tissue, and (3) a proportion of soft tissue, such as leaves, is shed in senescence or eaten by herbivores so that the duration of possible CH4 emission extends for much less than the full growing season.
We, therefore, used two alternative methods that are methodologically consistent for scaling to the global rate. The first assumes that leaves are responsible for most aerobic CH4 emissions, and we estimated global CH4 emission by multiplying the observed emission rates per unit dry mass by estimates of the standing biomass of leaves in different biomes. The second method assumes that the ratio of photosynthesis to aerobic CH4 emission is relatively constant and that global CH4 emissions can be calculated from estimates of photosynthesis.
Leaf-mass-based estimates
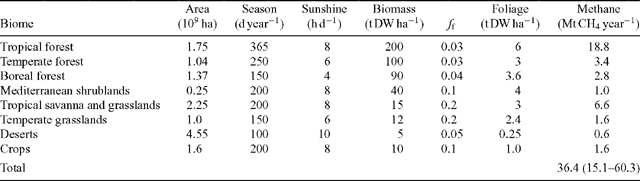
For each of eight major biomes, we estimated CH4 emissions, mb, as:
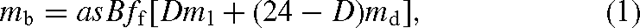
where a is the area of each biome (ha), B an estimate of biomass density in each biome (kg DW ha–1), ff the fraction of biomass contained in foliage, D effective daylight hours (h d–1), ml and md (kg CH4 kg–1 DW h–1) the respective CH4 emission rates in the light and dark as determined by Keppler et al. (2006) and s is season length (d year–1), an estimate of the time in each year over which conditions are conducive for metabolic activity when temperatures are moderate and water is available for plants. We used the same categorisation into different biomes as Keppler et al. (2006) and scaled up to a global estimate of CH4 emissions by simply summing the contributions from all biomes.
Photosynthesis-based estimates
The second method for estimating the global budget makes the assumption that the light-stimulated component of aerobic CH4 emission is linked to photosynthesis. Hence, this provides a methodologically consistent means of scaling up based on the rate of net primary production instead of leaf mass in different biomes. This provides a convenient means of integrating across day and season length with conditions more or less conducive to metabolic activity. Hence, we expressed the rates observed by Keppler et al. (2006) as a function of the assumed photosynthetic rates of the leaves in their measuring chambers. We calculated the ratio of photosynthesis to CH4 emission, r, in molar units as:
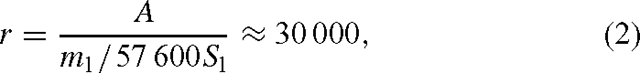
where A is photosynthetic rate, conservatively estimated as 10 µmol m–2 s–1 (e.g. Peterson et al. 1999), ml the average CH4 emission rate from attached leaves in the light, measured at 374 µg kg–1 DW h–1 by Keppler et al. (2006), Sl is specific leaf area, taken to be 20 m2 kg–1 as an average for a range of species (Vile et al. 2005) and 57 600 is a constant to convert from hours to seconds and mass-based to molar units for CH4 emission.
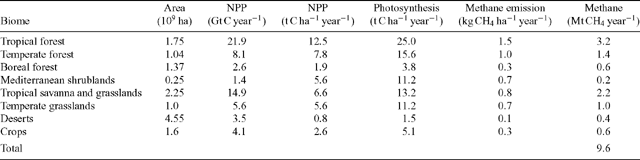
We then calculated CH4 emission for each biome, mb, as:
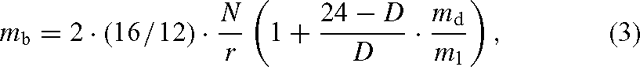
where N is NPP (kg C ha–1 year–1) of each biome. The constants 16 and 12 convert from mass-based carbon and CH4 units to molar units. The constant ‘2’ estimates annual photosynthesis from NPP. This implies that half of photosynthetically fixed carbon is lost again in autotrophic respiration (e.g. Gower et al. 1999). The term in the second set of brackets estimates the additional contribution from nighttime CH4 emission, with the symbols the same as in Eqn (1).
Estimating the relevance for tree plantings
For estimating the relevance of CH4 emissions for tree plantations, it is necessary to compare the difference in aerobic CH4 emission from alternative vegetation types with the benefit from carbon sequestration.
We used representative global average tree-growth rates to estimate the carbon sequestration potential of tree plantings. For pasture, we assumed the system to be in steady state, with no change in carbon stocks (e.g. Murty et al. 2002). Calculations were based on either an estimate of standing leaf mass in the two systems, or estimates of photosynthesis as the basis for estimating CH4 emissions. So, the difference in CH4 emissions between a tree plantation and a pasture, mΔ, was calculated as:
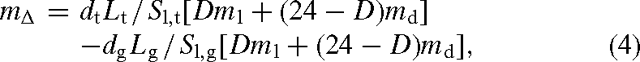
where dt and dg are the number of days in the growing season (d year–1) for trees and grass, respectively, Lt and Lg are leaf area indices (m2 m–2) for plantation trees (8.7) and grass (2.5; Asner et al. 2003), Sl,t and Sl, g specific leaf areas (m2 kg–1 DW) for trees (8.3) and grass (23; Vile et al. 2005), ml and md and D the same parameters as used above. The same daylength was used for pasture and plantations, but season length could be different owing to the longer persistence of tree foliage with access to deeper water supplies in seasonally dry environments. Details used in respective simulations are given in the Tables below.
The calculated differences in CH4 emissions could then be used to calculate a CH4 offset, mo, of any carbon sequestration benefit. This was calculated as:

where Ce is the carbon stored in a plantation in CO2 equivalents (CO2e), and w is the relative greenhouse warming potential of CH4 (23 for an assessment horizon of 100 years; Ramaswamy et al. 2001). The carbon storage benefit was calculated as:
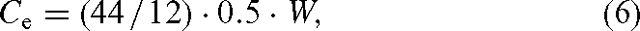
where W is the carbon storage benefit calculated as the biomass stored in wood, 0.5 calculates the fraction of woody dry mass that is carbon and 44 / 12 converts from carbon to CO2 to allow a dimensionally consistent comparison between carbon storage and CH4 emission effects. A generic carbon storage potential of 10 t DW ha–1 year–1 was assumed here (Schlamadinger et al. 2000). Adopting one global figure hides a great diversity across the globe, with boreal regions generally having lower growth rates, but growth rates in tropical regions being able to exceed that figure, especially on soils with good nutrition (Schlamadinger et al. 2000).
The effect of tree plantations was also assessed using photosynthesis as the means of calculating aerobic CH4 emission. Hence,
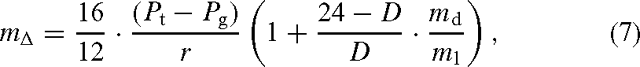
where r is the ratio of photosynthesis to CH4 emission as defined before, Pt and Pg are the photosynthetic rates (kg C ha–1 year–1) of trees and grass, respectively and the constants 16/12 convert between mass and molar units for carbon and methane. The term in the second bracket accounts for the additional nighttime CH4 emission as in Eqn (3). Photosynthesis of trees, Pt, was assumed to be related to carbon sequestration in wood by:

where W is the annual wood increment (kg DW ha–1 year–1), with the assumptions that one-third of NPP is stored in sequestered wood, that NPP is half of annual photosynthesis (e.g. Gower et al. 1999) and that half of dry mass is carbon. For grasslands, a range of assumptions was used as given in the Tables below. This was also combined with extremes in the assumption about the ratio of photosynthesis to CH4 emission as shown below.
Results
Global up-scaling
Our first estimate of global aerobic CH4 emissions used an approach similar to that of Keppler et al. (2006), except that we based our estimate on leaf mass in different biomes instead of NPP (Table 1).
|
With leaf mass as the basis for up-scaling, we derived an estimate of global CH4 emissions of 36 (range 15–60) Mt CH4 year–1, with about half of estimated global emissions attributed to tropical forests. This estimate is substantially lower than the 149 (range 62–236) Mt CH4 year–1 estimated by Keppler et al. (2006).
The second approach relates CH4 emissions to photosynthesis and uses independent estimates of photosynthesis as the basis for up-scaling. Using the same data for NPP as used by Keppler et al. (2006), and making a conservative assumption about photosynthetic rates in their plants resulted in a global emissions estimate of 9.6 Mt CH4 year–1 (Table 2), which was even lower than the leaf-mass-based estimate.
|
These two approaches are based on different assumptions, all of which are consistent with the currently available data. Both approaches result in estimates of global emissions that are much lower than that estimated by Keppler et al. (2006).
Tree plantings
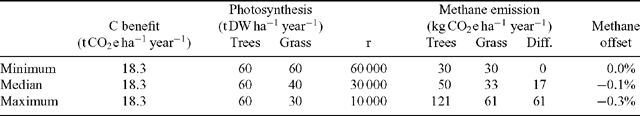
The extent by which CH4 emissions negate the benefit of carbon sequestration by tree plantings is shown in Table 3 with calculations based on leaf-mass estimates. For minimum, median and maximum calculations, the range of specific emission rates in the dark and light measured by Keppler et al. (2006) were used. In addition, different assumptions were used with respect to season length and sunshine hours. For the minimum calculation, the same short season length was assumed for trees and pasture. This might be applicable for a cool-temperate climate where cold winter temperatures equally inhibit grass and forest metabolic activity. For the maximum calculation, it was assumed that trees would be active all year, whereas grass would be limited by drought conditions to only half a year. A longer daylength was also assumed for these conditions.

|
These detailed calculations showed that consideration of CH4 emissions did little to negate the benefit from planting trees. Under our most likely assumptions, consideration of changes in CH4 emission reduced the carbon sequestration benefit by only 1.1%. The greatest reduction in sequestration benefit by CH4 emission was 4.4% when we employed the most extreme assumptions with respect to emission factors and season length.
Alternatively, the calculations were based on photosynthesis. We varied the assumed molar ratio of photosynthesis to CH4 emissions between 60 000 and 10 000, and the assumption about photosynthesis of grasslands relative to that of forest stands to cover a range of possibilities.
With these assumptions, the benefit of planting trees would be negated by between 0 and 0.3% through consideration of CH4 emissions (Table 4). Differences in methane emissions occur only when photosynthetic carbon gain from grasses is less than that from forests, which might be the case in seasonally dry environments where grass dies back at the onset of a dry season while trees might be able to continue to photosynthesise with access to water deeper in the profile. In systems without water limitations, photosynthetic rates of trees and grasses are likely to be similar, with consequently no expected differences in CH4 emissions.
|
Using this calculation method, we found the effect of including CH4 emissions to be negligible under all combinations of assumptions, with a 0.1% reduction as the most likely estimate, and an estimate of a 0.3% reduction only under the most extreme assumptions.
Discussion
We believe that the procedure used by Keppler et al. (2006) to calculate global aerobic plant CH4 emissions from their chamber measurements overestimates the likely magnitude of emissions. We, therefore, used two different methods with independent assumptions to scale up from their measurements to the globe.
Leaf-mass-based estimation
Keppler et al. (2006) presented rates of aerobic methane release per unit of plant dry mass, which in most cases was mainly leaf material. We therefore calculated global fluxes on the basis of estimates of leaf mass, which is dimensionally consistent.
This approach was based on estimated leaf mass in different biomes and resulted in an estimate of 36 Mt CH4 year–1. The NPP-based estimate of Keppler et al. (2006) greatly exceeds the leaf-mass-based estimate because the numeric value of NPP considerably exceeds the average amount of living foliage. For tropical forests, for example, we estimated an average leaf mass of 6 t DW ha–1 (Table 1), whereas NPP is estimated at 25 t DW ha–1 year–1 (equivalent to 12.5 t C ha–1 year–1; Table 2). Consequently, the leaf-mass-based estimate of CH4 emissions is only about one quarter of the NPP-based estimate.
For these calculations, we had to assume that leaves emit methane at a constant rate throughout their life. One could also estimate CH4 emissions on the basis of annually produced new leaf mass. This would be appropriate if the mechanism of aerobic CH4 emission were such that the source of carbon of the emitted CH4, such as available methoxyl side chains, is exhausted within one year of formation. CH4 emissions might cease even earlier, such as upon cessation of growth processes in newly formed leaves. As no information about the time course of CH4 emission is yet available, it was not possible to use this calculation method. CH4 might also be emitted from other light-exposed soft tissues, such as flowers, but the quantitative contribution of such tissues is likely to be small.
Our estimate is also based on assumptions of season length and effective daylength because methane emissions were stimulated by sunlight. We used the same season and daylength estimates as Keppler et al. (2006), which implicitly assumed that the CH4 release rates in the light have a high light threshold. Effects of temperature or other physiological variables had to be ignored because there is no information yet about their possible effect on emissions.
Photosynthesis-based estimation
Our second approach for scaling up is based on the ratio of photosynthesis to CH4 emissions. This approach assumes that CH4 emissions are in some way linked to the production of new carbohydrate, or directly involve the electron transport chain so that ratio of photosynthetic carbon gain to CH4 emission might be conserved across different physiological states and conditions.
If photosynthesis were directly involved in CH4 production it could be by providing reducing equivalents for methane production which would provide a mechanistic explanation for the light stimulation of emissions. Highest CH4 emission rates were also observed for the C4 plants maize and sorghum and the lowest rates for Norway spruce, a C3 plant, which would be in line with expected photosynthetic rates for these species.
This method thus requires no assumptions about season length as this is implicitly included in the estimates of photosynthesis for different biomes. An assumption about daylength was only required to estimate the (smaller) contribution from nighttime CH4 emissions. Estimates of daylength and NPP for different biomes, which we used to estimate photosynthesis, were taken from Keppler et al. (2006).
We used a conservative estimate of photosynthesis to calculate the ratio of photosynthesis to CH4 emission so that our estimate is more likely to overestimate CH4 emissions. Nonetheless, despite assumptions that were biased towards higher emissions, our derived estimate of global aerobic CH4 emissions was only 10 Mt CH4 year–1, and thus even lower than the leaf-mass-based estimate.
Methane oxidation in soils
Soils of most ecosystems also oxidise CH4. This is estimated to consume between 10–44 Mt CH4 year–1 globally (Ehhalt et al. 2001), with oxidation rates strongly dependent on soil moisture conditions, being highest at intermediate soil moisture (e.g. MacDonald et al. 1996; Price et al. 2004). When soils are too wet, soil micro-sites become anaerobic and CH4 is produced rather than oxidised.
Oxidation rates are generally in the range of 1–5 kg CH4 ha–1 year–1 (Smith et al. 2000; Mosier et al. 2004), with a reduction by about two-thirds when soils are cultivated (Smith et al. 2000). Oxidation rates can increase again under forests when soils remain uncultivated, but the recovery generally takes decades to centuries (Smith et al. 2000).
Hence, forests have the benefit, at least compared with cultivated soils, of encouraging CH4 oxidation in the soil by ≈1 kg CH4 ha–1 year–1 (Smith et al. 2000), corresponding to ≈23 kg CO2e ha–1 year–1 for a greenhouse warming potential of 23 for methane relative to CO2 based on an assessment horizon of 100 years (Ramaswamy et al. 2001). This approximately balances the estimated extra aerobic CH4 emission by trees relative to grass calculated by the photosynthesis-based method (Table 4).
The global methane budget
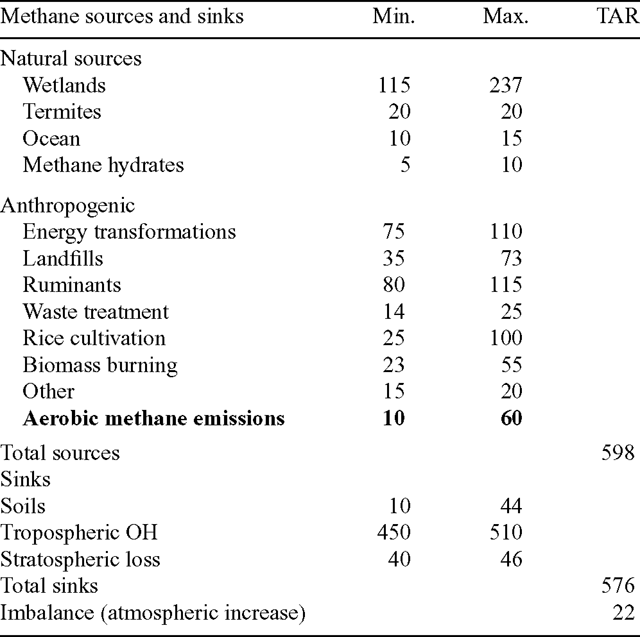
Our estimate for global aerobic CH4 emissions can be readily accommodated within the estimates of the established sources and sinks in the present global budget (Table 5). The uncertainty in emissions from wetlands alone has been calculated by different workers to be over 100 Mt CH4 year–1. An additional source term of up to 60 Mt CH4 year–1, as calculated here, would require no adjustment to any of the more established terms. By contrast, an additional flux of the order of 149 Mt CH4 year–1, as calculated by Keppler et al. (2006), could not be reconciled with the other terms in the budget without requiring substantial re-assessment of their magnitudes.
|
The possible magnitude of aerobic CH4 emissions from plants must also be consistent with the pre-industrial / agricultural CH4 budget when aerobic plant CH4 emissions were presumably no less than they are at present but emissions from burning fossil fuels, domestic livestock, flooded rice cultivation and landfills were much less than at present. Based on the global mean CH4 concentration of 695 ppb for the period 1000–1800 AD and estimates of its atmospheric lifetime of 8–11 years, the total of all CH4 sources must have been in the range of 200–250 Mt CH4 year–1 (Etheridge et al. 1998). Houweling et al. (2000) estimated the pre-industrial emissions from wetlands as 163 (uncertainty range 130–194) Mt CH4 year–1 and the sum of all other sources (termites, rice farming, ruminants, biomass burning, waste treatment, oceans, volcanoes) at 88.5 (53.5–143.5) Mt CH4 year–1, for a total of 251.5 (183.5–337.5) Mt CH4 year–1.
This suggests that an additional source of 10–60 Mt CH4 year–1, as calculated here, could be accommodated within the uncertainties of the pre-industrial / agricultural CH4 budget provided that the estimates of all other individual source terms and the atmospheric lifetime of CH4 were at the lower end of estimates. Aerobic CH4 emissions rates of 149 (62–236) Mt CH4 year–1, as estimated by Keppler et al. (2006), in contrast, would be too large to be reconciled with the current understanding of the pre-industrial / agricultural budget.
Frankenberg et al. (2005, 2006) recently reported space-based observations of the CH4 profile across the globe and a comparison with concentrations calculated from global sources and sinks. The comparison pointed to higher than expected concentrations over tropical forest regions, especially in South America. Aerobic CH4 emission from plants may resolve that discrepancy, and our calculations suggest that aerobic CH4 emissions are of the same magnitude as total CH4 emissions reported in a field study from Brazil by do Carmo et al. (2006). Other components of the global budget, such as emissions from biomass burning, termites, wetlands, and the CH4 production / oxidation balance in partly wet soils, as well as oxidation by OH in the atmosphere, however, are all sufficiently uncertain to further account for any remaining discrepancies.
The value of tree plantings
Our quantitative assessment of the value of plantation establishment as a greenhouse mitigation option is also only marginally affected by consideration of aerobic CH4 emissions. Our best estimate was that the carbon sequestration benefit might be negated by between 0.1 and 1.1% by such CH4 emissions. At the same time, long-established forests provide the additional benefit of encouraging greater CH4 oxidation in soils than in cultivated soils. Hence, there is no justification for questioning the value of planting trees on the grounds of aerobic CH4 emissions.
Carbon sequestration, the process of removing CO2 from the atmosphere and storing it in an increasing pool of woody biomass does, however, continue only over the growth phase of forests. After forests reach maturity there is no further carbon uptake whereas any difference in CH4 emission can be maintained indefinitely. Nonetheless, because the effect of CH4 emissions is of such minor importance, the value of tree plantings is overwhelmingly determined by considerations of its carbon balance.
The observations by Keppler et al. and their interpretation
The primary focus of this paper is to question the methods that Keppler et al. (2006) have used to scale up from their chamber observations to estimate global aerobic CH4 emissions from plants. Given that their experimental results form the basis of these global estimates, it is appropriate to scrutinise the experimental methods that underpin these estimates.
We accept that the experimental methods did indeed demonstrate that CH4 could be emitted under aerobic conditions. The most obvious alternative sources of methane could be either microbial or atmospheric. The authors were able to satisfactorily exclude anaerobic processes as the source of CH4 emissions by irradiating plant material yet still observing CH4 emissions. In principle, it is also possible that the observed CH4 emissions were the result of desorption of atmospheric CH4 following the flushing of chambers containing plant material with CH4-free air. This possibility was partly discounted by demonstrating that emitted CH4 had an isotopic signature similar to the esterified methoxyl groups (Keppler et al. 2004) of the C3 and C4 plants that were being studied.
However, that isotopic work does not fully allay our concerns about the reliability of measured methane emission rates because of the use of static chambers and CH4-free air. This approach inherently introduces some limitations into emission rate studies. The study used static chambers that were generally flushed with CH4-free air, and emission rates were calculated from the build-up of CH4. This inevitably necessitated plant exposure to a wide range of CH4 concentrations over the course of the observation period. Lowest CH4 concentrations observed were in the parts per billion range after flushing the chambers with CH4-free air, while the highest concentrations were attained at the end of the observation period. These levels were often in excess of normal ambient concentrations.
Over this range in concentrations, CH4 may have physically absorbed or desorbed from plant surfaces or the chamber materials, especially the plexiglass chambers used for experiments with intact plants, thus adding to or subtracting from plant-derived CH4 emissions. The change in CH4 concentration might also have stimulated or inhibited plant emissions. The importance and magnitude of these processes remains unknown until an underlying mechanism responsible for CH4 emissions can be demonstrated.
Concerns also relate to the draw-down in CO2 concentrations while CH4 was building up. Keppler et al. (2006) reported that CO2 concentrations did not fall below 250 ppm but did not report an upper limit of CO2 concentrations. If there is a metabolic link between photosynthetic and methane pathways, any changes in CO2 concentration could have affected CH4 emission rates.
A further concern relates to the light-exposure experiments with intact plants. The authors suggest that exposure to sunlight greatly increased CH4 emission rates. However, temperature was not tightly controlled in this set of experiments, and the possibility cannot be excluded that the apparent response to sunlight was, at least in part, a response to temperature, which had been shown to have a highly stimulatory effect on emissions.
If one accepts the basic veracity of the observations, the question turns to its possible interpretation. CH4 is a reduced compound, and some bacteria can generate energy from its oxidisation to CO2 and water. CH4 production under aerobic conditions, however, runs counter to the expected direction of chemical reactions in an oxidising atmosphere. The production of CH4 from purified pectin is particularly difficult to understand, given that it involves the reduction of pectin to the more highly reduced CH4. The continued increase in the rate of CH4 emission up to 70°C suggests that no enzymes are involved.
Rates of CH4 emission from both live and dead material, as well as from pectin, were also stimulated by light. The rates of release were, however, nearly 100 times greater from attached than detached or dried tissues. It may simply mean that rates occur faster in the more metabolically reactive conditions of a living cell or that two independent processes are involved. The slower process may be able to occur in dead tissues, but higher rates might require living tissues where the cell’s metabolism, especially in the presence of light, can facilitate the reduction of CH4. No specific mechanism has yet been suggested for the production of CH4 under aerobic conditions.
Future work
The calculations reported here are carried out under considerable uncertainty as much critical information that is required for up-scaling is not yet known. Additional research is needed to overcome this uncertainty. We consider the following as deserving priority.
Independent confirmation of the observations of aerobic methane release
The original study by Keppler et al. (2006) appears to have been carried out with care and considerable attention to possible problems and artefacts although only scant details of methods were given. Nonetheless, we urge caution in accepting its implications until there is independent confirmation of the findings. The emission rates also need to be quantified more reliably. Given this, any extrapolation of these findings to the global scale needs to be done with extra care and mindful of the possibility of artefacts.
Identification of the underlying mechanism
Elucidation of the underlying biochemical pathway for aerobic CH4 production is crucial for gaining a full understanding of its role and possible global significance. Other experiments suggested here will provide the observational base to allow identification of these underlying processes.
Is there a temporal component?
Once leaves are exposed to light, does the rate remain constant? If it does it would indicate that CH4 is released as a by-product of some on-going reaction. Alternatively, rates may decrease over time. If rates decrease slowly (in the order of days to weeks), it might indicate that CH4 is released slowly from a large pool. If rates change relatively quickly (over minutes to hours) after a change in conditions, then it lends support to the possibility that one might simply be observing an artefact of CH4 absorption / desorption.
Any possible change in emission rates over time is crucially important for up-scaling. If rates change over days to weeks, it would imply that annual rates cannot be estimated simply by multiplying short-term rates by the length of time over which plants are assumed to remain under the same condition.
From which plant tissues or compounds does CH4 originate?
Keppler et al. (2006) found that CH4 can be emitted from pure pectin, although at much smaller rates than from intact plants. It would be useful to test for any possible release from other compounds as well, including some with and some without methoxyl groups to help identify the chemical reactions that are involved in aerobic CH4 production.
At the whole-plant level, it is important to know whether CH4 is emitted from photosynthesising leaves, from growing tissues or from all metabolically active tissue. Do metabolically inert tissues, such as fully formed xylem, or tissue shielded from light, such as roots, also emit CH4? This again will be important for estimating annual emissions.
More species comparisons
The number of species tested is still very small. Only one tree species has been assayed using intact tissue and no species from tropical forests. While CH4 emission rates were measured from detached leaves of a greater number of species, these rates were nearly two orders of magnitude lower than those measured for intact plants. Hence, scaling to the globe is uncertain when a large part of the overall flux is likely to originate from species for which no measurements have yet been taken.
Temperature dependence of CH4 emission in the light
Keppler et al. (2006) presented the temperature response of CH4 emission in the dark, which showed a continued increase in rates up to 70°C, suggesting a non-enzymatic reaction mechanism. The temperature response of CH4 emission in the light needs to be measured to indicate whether plant metabolism is involved. If the light stimulation in intact leaves were to continue up to high temperatures, it would exclude an enzymatic process and indicate a non-enzymatic reaction as in non-living organic matter.
Irradiance dependence of CH4 emission
Keppler et al. (2006) have shown a dark–light modulation of emission rates. This response needs to be confirmed under more tightly controlled conditions. If it can be confirmed, it would be valuable to know more about the light-response curve. Do CH4 emissions increase linearly with light? Do they saturate? Is there some low-light threshold?
Action spectrum of CH4 emission
Is UV radiation involved in CH4 emission, or does the spectral dependence of CH4 emission mirror the action spectrum of photosynthesis? If it is driven by UV, then we might be dealing with a non-biological action, but if it mirrors the action spectrum of photosynthesis, it would indicate the involvement of photosynthetic electron transport.
It is noteworthy that the experimental work on dead plant materials was done in glass vials whereas the work on intact plants used plexiglass chambers (Keppler et al. 2006). As these chambers could possibly have different UV transmission properties, it raises the possibility that the apparent difference in CH4 emissions between dead and intact plants might have simply been due to the different amounts of UV the materials were subjected to. A carefully constructed action spectrum of CH4 emissions should show whether different amounts of UV could have played a role in these findings.
Do rates change with the physiological state of leaves?
Are rates higher or lower in water stressed leaves? How do rates change with plant nutrition? Are rates more constant if they are expressed as a function of photosynthetic rates across differences in nutrition or stress levels?
Conclusions
The work of Keppler et al. (2006) has identified a new source of CH4 emission from plants under aerobic conditions. If this can be confirmed by independent research, preferably by different methodologies, it will need to be incorporated into our understanding of CH4 budgets. Identifying the mechanism of aerobic methane formation would assist in scaling from the laboratory to the globe.
We reassessed here the global significance of aerobic CH4 emission and derived a much smaller estimate than that of Keppler et al. (2006). While the magnitude of the contribution from this additional source to the global CH4 budget is uncertain, our estimate can be readily accommodated within existing uncertainty in the established sources and sinks of both the present and pre-industrial / agricultural global CH4 budget.
For mitigating climate change, we also believe that the carbon sequestration benefit from planting trees far outweighs the warming associated with aerobic CH4 emissions by a factor of about 100. Statements in the media that have questioned the benefit of trees as a mitigation strategy are therefore unsubstantiated.
Acknowledgments
We thank Murray Badger, Trevor Booth, Ray Leuning, Michael Robinson, Stephen Roxburgh and Bruce Wright for useful discussions and specific comments and suggestions on this work.
Asner GP,
Scurlock JMO, Hicke JA
(2003) Global synthesis of leaf area index observations: implications for ecological and remote sensing studies. Global Ecology and Biogeography 12, 191–205.
| Crossref | GoogleScholarGoogle Scholar |

do Carmo JB,
Keller M,
Dias JD,
de Camargo PB, Crill P
(2006) A source of methane from upland forests in the Brazilian Amazon. Geophysical Research Letters 33, L04809.
| Crossref | GoogleScholarGoogle Scholar |
Etheridge DM,
Steele LP,
Francey RJ, Langenfelds RL
(1998) Atmospheric methane between 1000 A.D. and present: evidence of anthropogenic emissions and climate variability. Journal of Geophysical Research 103, 15979–15993.
| Crossref | GoogleScholarGoogle Scholar |
Foley JA
(1994) Net primary productivity in the terrestrial biosphere: the application of a global model. Journal of Geophysical Research 99, 20773–20783.
| Crossref | GoogleScholarGoogle Scholar |
Frankenberg C,
Meirink JF,
van Weele M,
Platt U, Wagner T
(2005) Assessing methane emissions from global space-borne observations. Science 308, 1010–1014.
| Crossref | GoogleScholarGoogle Scholar | PubMed |
Frankenberg C,
Meirink JF,
Bergamaschi P,
Goede APH,
Heimann M,
Körner S,
Platt U,
van Weele M, Wagner T
(2006) Satellite chartography of atmospheric methane from SCIAMACHY onboard ENVISAT: analysis of the years 2003 and 2004. Journal of Geophysical Research In press
,
Gower ST,
Kucharik CJ, Norman JM
(1999) Direct and indirect estimation of leaf area index, f
APAR and net primary production of terrestrial ecosystems. Remote Sensing of Environment 70, 29–51.
| Crossref | GoogleScholarGoogle Scholar |
House JI,
Prentice IC, Le Quéré C
(2002) Maximum impacts of future reforestation or deforestation on atmospheric CO2. Global Change Biology 8, 1047–1052.
| Crossref | GoogleScholarGoogle Scholar |
Houweling S,
Dentener F, Lelieveld J
(2000) Simulation of preindustrial methane to constrain the global source strength of natural wetlands. Journal of Geophysical Research 105, 17243–17255.
| Crossref | GoogleScholarGoogle Scholar |
Keppler F,
Kalin RM,
Harper DB,
McRoberst WC, Hamilton JTG
(2004) Dramatic 13C depletion in the plant methoxyl pool and its biogeochemical implications. Biogeosciences 1, 123–131.
Keppler F,
Hamilton JTG,
Braß M, Röckmann T
(2006) Methane emissions from terrestrial plants under aerobic conditions. Nature 439, 187–191.
| Crossref | GoogleScholarGoogle Scholar | PubMed |
Lowe DC
(2006) Global change: a green source of surprise. Nature 439, 148–149.
| Crossref | GoogleScholarGoogle Scholar | PubMed |
MacDonald JA,
Skiba U,
Sheppard LJ,
Hargreaves KJ,
Smith KA, Fowler D
(1996) Soil environmental variables affecting the flux of methane from a range of forest, moorland and agricultural soils. Biogeochemistry 34, 113–132.
| Crossref | GoogleScholarGoogle Scholar |
Mosier A,
Wassmann R,
Verchot L,
King J, Palm C
(2004) Methane and nitrogen oxide fluxes in tropical agricultural soils: sources, sinks and mechanisms. Environment, Development and Sustainability 6, 11–49.
| Crossref | GoogleScholarGoogle Scholar |
Murty D,
Kirschbaum MUF,
McMurtrie RE, McGilvray H
(2002) Does forest conversion to agricultural land change soil organic carbon and nitrogen? A review of the literature. Global Change Biology 8, 105–123.
| Crossref | GoogleScholarGoogle Scholar |
Peterson AG,
Ball JT,
Luo YQ,
Field CB, Reich PB , et al.
(1999) The photosynthesis-leaf nitrogen relationship at ambient and elevated atmospheric carbon dioxide: a meta-analysis. Global Change Biology 5, 331–346.
| Crossref | GoogleScholarGoogle Scholar |
Price SJ,
Kelliher FM,
Sherlock RR,
Tate KR, Condron LM
(2004) Environmental and chemical factors regulating methane oxidation in a New Zealand forest soil. Australian Journal of Soil Research 42, 767–776.
| Crossref | GoogleScholarGoogle Scholar |
Smith KA,
Dobbie KE,
Ball BC,
Bakken LR, Sitaula BK , et al.
(2000) Oxidation of atmospheric methane in Northern European soils, comparison with other ecosystems, and uncertainties in the global terrestrial sink. Global Change Biology 6, 791–803.
| Crossref | GoogleScholarGoogle Scholar |
Vile D,
Garnier E,
Shipley B,
Laurent G, Navas M-L , et al.
(2005) Specific leaf area and dry matter content estimate thickness in laminar leaves. Annals of Botany 96, 1129–1136.
| Crossref | GoogleScholarGoogle Scholar | PubMed |
Wang JS,
Logan JA,
McElroy MB,
Duncan BN,
Megretskaia IA, Yantosca RM
(2004) A 3-D model analysis of the slowdown and interannual variability in the methane growth rate from 1988 to 1997. Global Biogeochemical Cycles 18, GB3011.
| Crossref | GoogleScholarGoogle Scholar |