

Evolution of growth-promoting plant hormones
John J. Ross A and James B. Reid A BA School of Plant Science, University of Tasmania, Hobart, Tas. 7001, Australia.
B Corresponding author. Email: jim.reid@utas.edu.au
This paper is part of an ongoing series: ‘The Evolution of Plant Functions’.
Functional Plant Biology 37(9) 795-805 https://doi.org/10.1071/FP10063
Submitted: 22 March 2010 Accepted: 14 May 2010 Published: 24 August 2010
Abstract
The plant growth hormones auxin, gibberellins (GAs) and brassinosteroids (BRs) are major determinants of plant growth and development. Recently, key signalling components for these hormones have been identified in vascular plants and, at least for the GAs and BRs, biosynthetic pathways have been clarified. The genome sequencing of a range of species, including a few non-flowering plants, has allowed insight into the evolution of the hormone systems. It appears that the moss Physcomitrella patens can respond to auxin and contains key elements of the auxin signalling pathway, although there is some doubt as to whether it shows a fully developed rapid auxin response. On the other hand, P. patens does not show a GA response, even though it contains genes for components of GA signalling. The GA response system appears to be more advanced in the lycophyte Selaginella moellendorffii than in P. patens. Signalling systems for BRs probably arose after the evolutionary divergence of the mosses and vascular plants, although detailed information is limited. Certainly, the processes affected by the growth hormones (e.g. GAs) can differ in the different plant groups, and there is evidence that with the evolution of the angiosperms, the hormone systems have become more complex at the gene level. The intermediate nature of mosses in terms of overall hormone biology allows us to speculate about the possible relationship between the evolution of plant growth hormones and the evolution of terrestrial vascular plants in general.
Additional keywords: auxin, brassinosteroid, gibberellin, moss.
Introduction
Plant hormones play key roles in regulating many aspects of plant growth and development, including shoot elongation, plant architecture, fruit growth and seed development, all of which are crucial for food and biomass production. A key subset of the plant hormones comprises the growth-promoting compounds: auxins, gibberellins (GAs) and brassinosteroids (BRs). Through a combination of mutant analysis and molecular studies, we now possess a good understanding of the perception of each of these hormones and some of the key elements of their signal transduction pathways (Santner and Estelle 2009). We also understand GA and BR biosynthesis in several model species, such as Arabidopsis thaliana, tomato (Lycopersicon esculentum Mill.), pea (Pisum sativum L.) and rice (Orysa sativa L.) (Yokota 1997; Nomura and Bishop 2006; Yamaguchi 2008), and progress is being made in untangling auxin biosynthesis (Normanly 2010). While the presence of growth-promoting hormones across the many groups of plants has been documented extensively (e.g. GAs; MacMillan 2002), it is only in the last few years that sufficient genomes have been sequenced to allow insight into the possible evolutionary origins of the biosynthetic and signal transaction pathways.
An analysis of the evolutionary origins of plant hormone systems must determine the time and plant group in which these occurred. Inevitably, this will not be straightforward, as there are cases where these compounds exist in plants only as secondary metabolites or as pathogenic compounds. The role of a compound as a hormone probably developed by the recruitment of existing signalling components and may have occurred independently for the same group of compounds in different phylogenetic sequences. This may be the case for the steroids in plants and animals (see the discussion of BR below). Although recent work with model species is frequently used to generalise about the roles of plant hormones, there is also a growing body of evidence that there are major differences in hormone physiology across the families represented by the different model species. This occurs both for the biosynthetic pathways involved (e.g. auxin), the specificity of the receptor systems (e.g. BRs) and the developmental processes regulated (e.g. GAs). Whether these differing characteristics have evolved independently in the separate groups or have been lost in some groups has not been explored, but certainly requires detailed examination and will be addressed later. While the evolutionary origin of these three core hormone systems has received considerable attention and is reviewed below, there are still clear gaps in our knowledge, due to the lack of sequenced genomes, which will hopefully be filled as more genomes are sequenced in the near future.
Auxin
Occurrence, biosynthesis and biological activity
A wide range of species synthesise the main naturally-occurring auxin, indole-3-acetic acid (IAA; Cooke et al. 2002). These species represent all the major groups of land plants, including mosses, liverworts, lycophytes, ferns, gymnosperms and angiosperms. At least some bacterial species, including pathogens, also produce IAA, and there are also reports that certain fungi can synthesise this compound (Gay et al. 1994; Splivallo et al. 2009). However, evidence for IAA in microalgae, including Chlamydomonas spp., is not convincing at present (Dutcher et al. 1992). Consistent with that, Rensing et al. (2008) reported that genes thought to be related to auxin homeostasis were not found in the genomes of the aquatic unicellular green algae Ostreococcus tauri, Ostreococcus lucimarinus and Chlamydomonas reinhardtii. However, IAA has been identified in multicellular algae, including Fucus distichus and Ectocarpus siliculosus (Basu et al. 2002; Le Bail et al. 2010), and importantly, Nitella spp. (Sztein et al. 2000). Nitella is a member of the Charophyta, the algal group that is thought to have included the ancestors of land plants (Judd et al. 2002). It is possible that unicellular algae ancestral to all multicellular plants may have once had the capacity to synthesise auxin but have since lost that capacity. Alternatively, the ancestral unicellular algae lacked the ability to synthesise auxin but that capacity has since arisen independently in at least two different multicellular algal groups (and hence in land plants).
Lau et al. (2009) reviewed the evidence that auxin can have physiological effects on algal growth, as well as evidence to the contrary, and concluded by doubting the functional relevance of auxin in algae, suggesting instead that IAA may simply be a by-product of tryptophan. However, a recent paper on the brown alga E. siliculosus suggests a role for IAA in development, although the response pathway appears to be different to that in vascular plants (Le Bail et al. 2010).
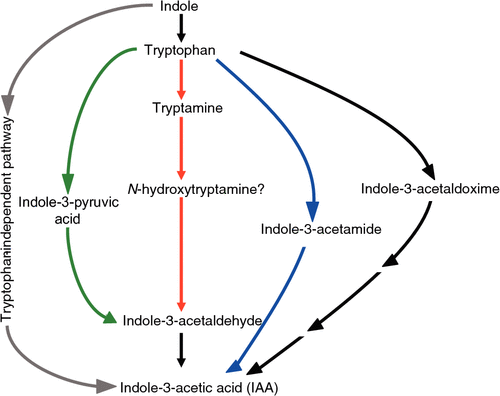
The actual presence or absence of IAA itself is not difficult to establish, since the compound is amenable to physicochemical analysis. However, our understanding of auxin synthesis pathways is still far from satisfactory. For angiosperms, five pathways have been proposed: one independent of and four dependent on the amino acid tryptophan (Fig. 1; Woodward and Bartel 2005; Sugawara et al. 2009). The main difficulty is determining which pathway or pathways predominate in a given species or plant part, or in a given environment. Another problem is that each pathway remains incompletely characterised at the biochemical and molecular levels.
|
In view of these gaps in our knowledge, it is difficult to discern evolutionary trends in auxin synthesis pathways. However, recent evidence indicates that two tryptophan-dependent pathways that operate in bacteria may also play significant roles in plants. One of these involves indole-3-pyruvic acid as the first product of tryptophan (Koga et al. 1991; Stepanova et al. 2008; Tao et al. 2008), and the other, indole-3-acetamide (Pollmann et al. 2009; Mano et al. 2010). Genes encoding enzymes for converting indole-3-acetamide to IAA in plants are phylogenetically related to those from bacteria (albeit distantly), indicating their ancient origin (Mano et al. 2010).
A third tryptophan-dependent pathway, via tryptamine, was brought to researchers’ attention in 2001 by the discovery of the YUC genes, which are thought to encode the enzyme for converting tryptamine to the next product in the pathway, N-hydroxytryptamine (Zhao et al. 2001). The YUC genes appear to be widespread in plants, including moss (Zhao 2010), although they appear not to have been identified in bacteria. Mutations in or overexpression of YUC genes result in auxin-related phenotypes, but there is some doubt about N-hydroxytryptamine as an intermediate (Quittenden et al. 2009) and about whether tryptamine is the in vivo substrate for the YUC proteins (Zhao 2010).
A fourth pathway, with indole-3-acetaldoxime as the first product of tryptophan, appears to be restricted to the Brassicaceae (Quittenden et al. 2009; Sugawara et al. 2009) and may therefore be of relatively recent origin. Indole-3-acetaldoxime is also a precursor of the defence compounds, indole glucosinolates (Grubb and Abel 2006).
Recently, Eklund et al. (2010) suggested that the tryptamine and indole-3-pyruvic acid pathways might operate in Physcomitrella patens, based on the presence of genes usually associated with these pathways (YUCCA genes for the tryptamine pathway and TAA1-like genes for the indole-3-pyruvic acid pathway).
Auxin signalling
Much of the recent literature on evolutionary aspects of auxin concerns the development of auxin signalling and, to a lesser extent, auxin transport, rather than auxin synthesis. This might, to some extent, reflect our better understanding of these aspects.
Genome sequencing in a range of species has permitted searches to be made for the auxin perception and signal transduction genes originally identified in angiosperms. Key proteins from those plants are the TIR1-AFB family, which function in auxin reception, and the Aux-IAA proteins, which, when auxin levels are low, interact with proteins known as auxin response factors (ARFs) to prevent these ARFs from regulating genes required for auxin responses (Santner et al. 2009). When auxin levels are high, the auxin–TIR1 complex destabilises Aux-IAA proteins, and the resulting low level of Aux-IAAs cannot prevent ARFs from regulating the auxin response genes.
The central issue is whether these auxin signalling components are also present in plants such as algae and mosses, and, if so, whether they interact in the way described above to rapidly regulate gene transcription. Lau et al. (2009) could find no complete Aux-IAA or ARF genes in the genomes of several algal species, including C. reinhardtii, even though Palenik et al. (2007) previously reported the presence of two Aux-IAA genes in that species.
Turning to the mosses, Cove et al. (e.g. Ashton et al. 1988) demonstrated that the caulenomata of putative auxin-deficient P. patens mutants could be rescued by applied auxin. This indicates that in the moss, auxin plays a role in development and that the components of auxin signalling operate, at least to some extent. Recently, Eklund et al. (2010) provided further confirmation of auxin action in P. patens when they showed that the A. thaliana SHI/STY gene family (positive regulators of auxin biosynthesis genes) also regulate auxin levels, and affect growth and development in the moss. These observations are consistent with the suggestion that land plants acquired functional or potentially functional components of auxin signalling before the divergence of the bryophyte groups from the lineage leading to vascular plants, but after the algae diverged from that lineage (Lau et al. 2009).
Pivotal for that suggestion has been the sequencing of the P. patens genome (Rensing et al. 2008). Previously, Cove et al. (2006) had commented that little was known about auxin-signalling pathways in moss. The sequencing indicated that auxin receptors, transporters and transcriptional regulators are encoded in the P. patens genome (Lau et al. 2008; Rensing et al. 2008). A total of 55 auxin-related genes were detected in P. patens by Rensing et al. (2008). Interestingly, however, there were fewer of these genes than in the flowering plants analysed: A. thaliana contains 174, Populus trichocarpa 230 and rice 175. (This represents a smaller proportion in the moss: e.g. 0.14% of the total genes compared with 0.65% in A. thaliana.)
There is some question, however, about whether auxin signalling components actually function in mosses to cause a ‘rapid transcriptional response to auxin’ (Paponov et al. 2009). According to Paponov et al. (2009), such a response, a feature of auxin signalling in angiosperms, does not occur in P. patens. Certainly, it appears at present that a rapid transcriptional response to auxin has not actually been demonstrated in the moss. While Paponov et al. (2009) acknowledge that Aux-IAAs in P. patens are degraded in an ‘auxin-dependent manner’, they suggest that this might not lead to rapid changes in gene expression. They note that Imaizumi et al. (2002) reported a relatively slow gene expression response to auxin in the moss; however, there appear to be no direct comparisons (in the one study) between P. patens and flowering plants in relation to the rate of transcriptional responses to auxin.
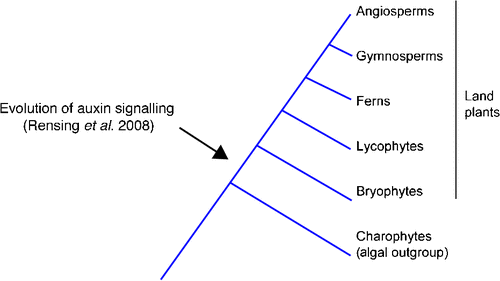
Paponov et al. (2009) suggest that a rapid transcriptional response to auxin evolved in vascular plants after their divergence from the last common ancestor shared with mosses (Fig. 2). In other words, they suggest that P. patens represents an intermediate stage between the green algae, which lack a functional auxin signalling pathway, and flowering plants, in which such a pathway is fully functional.
|
The intermediate nature of auxin signalling in P. patens might also be reflected in the nature and number of Aux-IAA proteins (Paponov et al. 2009). The moss Aux-IAAs all contain a particular motif in domain I, termed an LxLxPP motif, whereas in A. thaliana and other flowering plants, most Aux-IAAs contain an LxLxL motif that appears to be essential for strong transcriptional repression (Tiwari et al. 2004). Paponov et al. (2009) suggest that in flowering plants, the LxLxPP motif has been superseded by the LxLxL domain. Furthermore, P. patens has only three Aux-IAA genes, many less than the flowering plant species for which the genomes have been sequenced. For example, A. thaliana has 29; P. trichocarpa 35; rice 33.
Thus the Aux-IAA family has dramatically expanded and diversified during the evolution of flowering plants from the ancestors of land plants, and this is thought to be a major reason for the great diversity of responses to the relatively simple auxin molecule. Remington et al. (2004) suggest that some Aux-IAA genes predate the divergence of lineages leading to A. thaliana and rice 136 to 168 million years ago, while other duplication events appear to have occurred more recently, e.g. after the divergence of the Arabidopsis and Medicago lineages, around 96 to 113 million years ago. It appears also that many of the more recent duplication events in A. thaliana have been block duplications, although Remington et al. (2004) note that most of the early branching points in the Aux-IAA phylogeny involved tandem duplications.
Auxin transport
In angiosperms, auxin is transported basipetally in both the shoot and root via a dedicated polar transport system (Vieten et al. 2007). As for other aspects of auxin biology, a crucial question is: does polar auxin transport occur in the ‘lower’ plants, particularly in bryophytes? Previously, Cooke et al. (2002) found that polar IAA transport occurs in gametophytes of the liverwort Marchantia polymorpha and the moss Funaria hygrometrica, and suggested that polar auxin transport may have helped to regulate gametophyte development in the earliest land plants. However, Fujita et al. (2008) could not detect polar auxin transport in gametophores of P. patens, although, importantly, they did find that polar auxin transport might function in sporophyte development in that species.
Interestingly, Mravec et al. (2009) show that the ‘typical’ PIN protein in P. patens localises not to the plasma membrane but to the endoplasmic reticulum (ER). PIN5 from A. thaliana also localises to the ER, while other PINs from that species operate at the plasma membrane to orchestrate polar auxin transport. Mravec et al. (2009) suggested that an ER-based role for PIN proteins appeared very early in land plant evolution and might represent the ancestral function of these proteins. The ER-based PIN proteins are probably more concerned with intra-cellular than cell-to-cell transport, possibly explaining the difficulty of detecting polar auxin transport in the moss.
Gibberellins
Occurrence, biosynthesis and biological activity
GAs have been detected in a range of terrestrial vascular plants, including ferns and gymnosperms, and are also present in some fungi and bacteria (MacMillan 2002). Compared with auxin, our understanding of GA synthesis is markedly superior. We know the principal pathways involved, and most of the GA synthesis and deactivation genes have been cloned, at least in model species such as A. thaliana, rice and pea (Yamaguchi 2008). In angiosperms, the later stages of GA biosynthesis, and also the deactivation of bioactive GAs, are catalysed by members of the 2-oxoglutarate-dependent dioxygenase (2-ODD) group, and are known as GA 20-oxidases, GA 3-oxidases and GA 2-oxidases. Despite our extensive knowledge, however, there are very few analyses of the evolution of these genes, possibly reflecting the fact that the P. patens genome has not been sequenced for long. The three groups of GA-related 2-ODDs do form distinct groups in phylogenetic analyses, but it is interesting that the two synthesis groups (20-oxidases and 3-oxidases) do not cluster together in a sub-group distinct from the deactivation (2-oxidase) genes, regardless of whether the analysis is performed on genes from a large range of species (Sakakibara et al. 2008) or mainly on those from A. thaliana (Hedden and Phillips 2000). This may reflect the fact that the GA 2- and 3-oxidases oxidise neighbouring carbon atoms on the same ring of the GA molecule (the ‘A ring’), even though their actions have opposite effects on the content of bioactive GA.
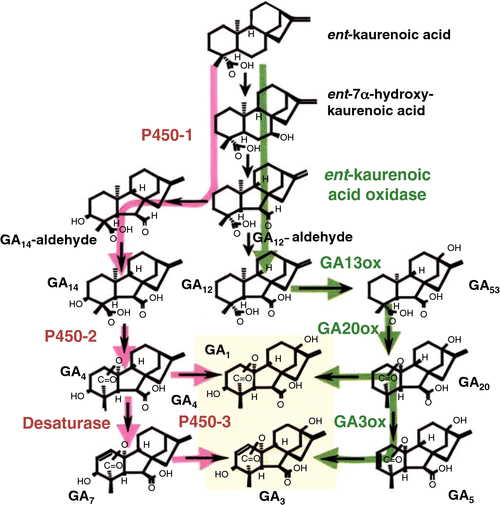
There appears to be little evidence from physicochemical studies that the algae produce GAs. Some fungal species, on the other hand, are copious producers of these hormones, and indeed this feature of certain pathogenic fungi led to the initial discovery of the GAs. However, consistent with the apparent gap in GA genes in algae, and the wide phylogenetic distance between fungi and land plants, it appears that the seed plants have not ‘inherited’ the same set of GA synthesis genes as the fungi (Bömke and Tudzynski 2009). This is because in fungi, the enzymes catalysing the later steps in GA synthesis are not 2-oxoglutarate-dependent dioxygenases but are cytochrome p450s instead (Bömke and Tudzynski 2009; Fig. 3). This is an excellent example of convergent evolution, because the phenotype in chemical terms is identical (GA3, for example, is present in seed plants as well as fungi) but has been arrived at by different evolutionary pathways.
|
A further difference concerns the biosynthesis of the early GA precursor ent-kaurene. In fungi (Hedden et al. 2002) and the moss P. patens (Hayashi et al. 2006), a single bifunctional enzyme catalyses the two steps involved in converting geranylgeranyl diphosphate to ent-kaurene (Davidson et al. 2006). In contrast, in the angiosperms, there is a separate monofunctional enzyme for each step (Davidson et al. 2006). Recently, it was shown that the gymnosperm white spruce (Picea glauca) employs two enzymes (Keeling et al. 2010), like angiosperms, and it appears that the two-enzyme system in plants evolved after the divergence of the mosses but before the divergence of the angiosperms from gymnosperms. Keeling et al. (2010) suggest that duplication and subfunctionalisation of the ancestral gene have resulted in the monofunctional enzymes found in seed plants.
The observation that in a phylogram of GA 2-ODD genes (Sakakibara et al. 2008), there are clustered representatives from P. patens in the 2-oxidase and 20-oxidase groups, suggests that these two groups originated and began to differentiate from each other before the divergence of the moss from the vascular lineage. In other words, the 2-ODD GA genes are very ancient. It should be noted, however, that there is no published evidence as yet that the P. patens ‘GA’ genes actually encode functional proteins. Two of these genes have been tested in this respect, with no functional activity detected (Hirano et al. 2007). Consistent with that, GAs appear not to have been detected in mosses, even though state-of-the-art physicochemical techniques have been used for P. patens gametophytes (Hirano et al. 2007). It has been suggested, however, that it is the spores or sporophyte – not the gametophyte – that might contain GAs in the moss (Anterola et al. 2009). A bioactive GA (GA4) has been detected in the lycophyte S. moellendorffii (Hirano et al. 2007) and several GAs have been found in ferns (e.g. Cibotium glaucum; MacMillan 2002).
It is interesting to review information on the functions of GAs and GA-related compounds in the various plant groups. Anterola (2008) develops the theme that as plants evolved, there were changes in the roles played by GAs, from promoters of spore germination in mosses, to antheridiogens in ferns and to growth promoters in angiosperms. Possibly, this should be amended to include GA-related compounds and GA precursors, since GAs themselves have not yet been identified in P. patens. The evidence for the involvement of GA-related compounds in the moss comes from the effects of AMO1618, which inhibits the formation of the early GA precursor ent-kaurene. AMO1618 inhibits spore germination in P. patens (Anterola et al. 2009). Ent-kaurene was able to substantially, but not completely, reverse this effect, while GA3 did not reverse the inhibition at all.
Another GA synthesis inhibitor, paclobutrazol, inhibited growth of the leafy P. patens gametophyte, an effect that again was not reversed by GA3. A third GA synthesis inhibitor, uniconazole, inhibited elongation in S. moellendorffii sporophytes and, yet again, a bioactive GA, GA4 could not reverse that inhibition.
One explanation for these observations is that a GA-like compound or compounds, derived from ent-kaurene, exhibits bioactivity in these species. This compound(s) is not ent-kaurene itself, because paclobutrazol inhibits growth but not ent-kaurene production. Neither does it appear to be a ‘normal’ bioactive GA, because GA3 could not restore growth or spore germination, and such GAs have not been identified in the moss. Possibly, the capacity of ent-kaurene to stimulate the germination of P. patens spores is attributable to a conversion by those spores of ent-kaurene to the bioactive compound. Further discussion on ent-kaurene-derived bioactive compounds was recently provided by Hayashi et al. (2010).
It is also relevant that the bifunctional ent-kaurene-producing enzyme in P. patens, when expressed in Escherichia coli (Anterola et al. 2009) or in a different insect cell-based system (Hayashi et al. 2006), produced ent-kaurene and ent-16α hydroxykaurene (called 16α hydroxykaurane in Anterola et al. 2009). It appears that P. patens produces large amounts of this latter compound, which has been implicated in plant defence (von Schwartzenberg et al. 2004).
In the ferns, there are also responses to GAs that are normally considered inactive in flowering plants. For these plants, unlike for mosses, there is no doubt that the GAs themselves are detectable (MacMillan 2002). In the ferns, the main role of the GA-related compounds is to promote the formation of antheridia at the expense of archegonia; the name given to such compounds is ‘antheridiogen’. In the ferns Lygodium circinnatum and Lygodium flexuosm, the main antheridiogen is GA73 methyl ester (Yamauchi et al. 1996). Other antheridiogens are GA9 methyl ester and 3-epi-GA63 (Yamauchi et al. 1995).
Thus, throughout evolutionary history, there may have been considerable change with regard to the roles played by GAs and related compounds, as well as in the actual compounds that activate the responses.
Regulation of GA levels by auxin and by GA signalling
In seed plants, it is well known that GA levels can be tightly regulated (Yamaguchi 2008), and we can speculate about the antiquity of the regulatory mechanisms involved. Two such regulatory factors are auxin and the ‘DELLA’ proteins, both of which upregulate GA synthesis and downregulate GA deactivation by controlling gene transcription. The effect of auxin is one of the clearer examples of plant hormone interactions. It appears to be ancient within the angiosperms, occurring in both monocots (Wolbang et al. 2004) and eudicots (Ross et al. 2000). One possible scenario is that the capacity of auxin to upregulate the GA genes arose only once, before the divergence of the three ODD groups; for the synthesis genes, this effect has persisted throughout subsequent evolution. After the divergence of the 2-oxidases, modification occurred such that at least some members of this group are now downregulated, not upregulated, by the level of auxin normally found in the plant. Interestingly, the putative ancestral condition (upregulation by normal auxin levels) can still be observed for some 2-oxidase genes in some circumstances (O’Neill and Ross 2002). A similar scenario might also apply in the case of the DELLA proteins, which mediate the capacity of bioactive GA to downregulate its synthesis and to upregulate its deactivation. The ancient nature of this phenomenon is indicated by evidence that it occurs in S. moellendorffii (Hirano et al. 2007).
GA signalling
Our understanding of GA signalling, as for GA synthesis, is well advanced, again enabling an examination of evolutionary trends. GAs operate by first interacting with a GA receptor, termed GID1 in rice, to form a complex that destabilises the growth-inhibitory DELLA proteins. This degradation occurs after the DELLAs are targeted to the 26S-proteasome (Ueguchi-Tanaka et al. 2007). Clearly, there are interesting parallels between GA and auxin signalling: GID1 is analogous to TIR1 and the DELLAs to Aux-IAA proteins, and protein destabilisation is a major feature of both systems (Santner et al. 2009). It has been claimed, somewhat controversially, that DELLA stability is affected also by hormones other than GA. Auxin is included in that category, but the evidence comes from a single report (Fu and Harberd 2003) that has yet to be confirmed. More recently, it has been suggested that usually, hormones other than GAs affect DELLA stability indirectly, by first affecting GA levels (Achard and Genschik 2009).
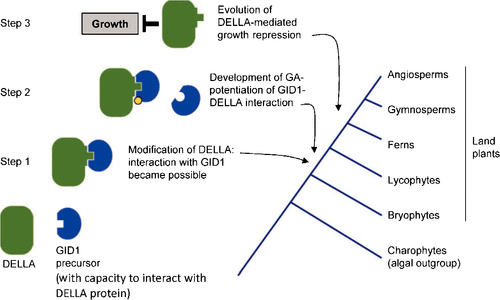
It appears that the lycophyte S. moellendorffii has a GID1-DELLA GA response system resembling that of seed plants (Hirano et al. 2007; Vandenbussche et al. 2007), whereas in the moss P. patens, such a system is not functional, even though genes for GID1-like proteins and for DELLAs are present. Yasumura et al. (2007) also concluded that there were significant developments in GA signalling between the divergences of the bryophytes and the lycophytes (Fig. 4). They found that the moss receptor and moss DELLA proteins did not interact with each other in a yeast two-hybrid assay, even when bioactive GA (GA3) was present. However, the corresponding proteins from the lycophyte Selaginella kraussiana did interact with each other, and this interaction was enhanced by GA3. Next, Yasumura et al. (2007) showed that the moss receptor interacted with the S. kraussiana DELLA (although this was not GA3-dependent), but the reciprocal interaction between lycophyte receptor and moss DELLA did not occur. They interpreted this observation as showing that the ancestral form of receptor, as present in the moss, was capable of interacting with DELLAs and that this capacity has subsequently persisted. The DELLAs, on the other hand, had to acquire the capacity to interact, and this occurred between the bryophyte and lycophyte divergences (Yasumura et al. 2007). Consistent with that, Hirano et al. (2007) also found that moss DELLAs did not interact with any receptors in the yeast two-hybrid assay.
|
However, Hirano et al. (2007) found that the same P. patens receptor as used by Yasumura et al. (2007) did not interact with any DELLA that they tested, including one from S. moellendorffi, although they did not test the S. kraussiana DELLA. Therefore, the interesting assertion by Yasumura et al. (2007) that the ancestral receptor possessed the capacity to interact with DELLAs does rest on only one interaction (with a DELLA from S. kraussiana). Furthermore, Hirano et al. (2007) reported that the rice DELLA protein SLR1 can interact with the lycophyte receptor but not with the moss receptor, suggesting that the moss receptor might not, after all, possess a ‘pre-existing’ capacity to interact with DELLAs.
Yasumura et al. (2007) suggest that the capacity of GA to stimulate the receptor–DELLA interaction also originated between the bryophyte and lycophyte divergences. Interestingly, the moss DELLAs, when transformed into certain A. thaliana genotypes, were able to inhibit growth but they did not do so in their native species; at least, not in the gametophyte. On the basis of this observation, Yasumura et al. concluded that the inhibitory function of the DELLAs evolved after the capacity to interact with the GA receptor. They contend that as far as the evolution of the DELLA growth-inhibiting capacity is concerned, changes in the responding system (involving the transcription of DELLA-regulated genes) have been more important than changes in the DELLA proteins themselves. Nevertheless, the DELLA proteins may also have undergone changes to enhance their growth-inhibiting capacity, as lycophyte DELLAs, but not those of moss, inhibited growth when overexpressed in rice (Hirano et al. 2007).
Interestingly, several GAs that are classified as inactive in flowering plants were highly effective at stimulating the receptor–DELLA interaction in S. moellendorffii (Hirano et al. 2007). Chief amongst these were GA37 and GA9, which were much more effective than GA1 or GA3, especially in the case of one of the two lycophyte receptors (SmGID1b). GA1 and GA3 meet the structural requirements for activity in flowering plants, in part because they have a hydroxyl group at the 3β position. GA4 (with a hydroxyl group at the 3β position but not at the 13 position) was more effective than GA1, in the S. moellendorffii system, leading to the suggestion that SmGID1b can discriminate between GAs based on the presence or absence of hydroxylation at the 13 position (Hirano et al. 2008). In contrast, however, in S. kraussiana, GA1, GA3 and GA4 were equally effective at stimulating a receptor–DELLA interaction (Yasumura et al. 2007).
Shimada et al. (2008), reporting on the crystal structure of GID1, noted that in the course of evolution from the more ancestral SmGID1-type receptors, replacement of certain key amino acid residues refined the receptor to give high affinity and specificity to bioactive seed plant GAs such as GA4 and GA1. The GID1-like receptors from the lycophyes and mosses appear to have evolved from proteins known as hormone-sensitive lipases, an ancient group present in both plants and animals. These enzymes have, during the course of evolution, lost their catalytic activity and instead have evolved a pocket in which the GA molecule fits. An amino acid ‘lid’, which holds the GA in place, has also developed, at least in angiosperms (Shimada et al. 2008).
Brassinosteroids
Occurrence, biosynthesis and biological activity
BRs have been indentified from ~60 plant species (Bajguz and Tretyn 2003). While the majority of the recent work has been performed in the model species A. thaliana, tomato, rice and pea, many of the early identifications were from cultured cells of Catharanthus roseus or tissues with high concentrations of the compounds (e.g. pollen) (Yokota 1997). Over 65 BRs have now been identified but elegant work using mutants late in the biosynthetic pathway suggests that only two C28 BRs are active, brassinolide and its precursor, castasterone (Nomura et al. 2005; Nomura and Bishop 2006). BRs have been identified from many angiosperm and gymnosperm families and also from a pteridophyte (Equisetum avense), a bryophyte (Marchantia polymorpha) and a chlorophyte (Hydrodictyou reticulatum) (Kim et al. 2002; Bajguz and Tretyn 2003). However, P450 genes with reasonable homologies to those involved with BR biosynthesis in angiosperms have not been found in S. moellendorffii or P. patens even though BRs were present (T. Yokota, pers. comm.).
The structure of BRs is derived from the 5-cholestone skeleton and BRs have strong similarities to the steroid hormones of animals. They consist of four rings and a variable-length side chain with a wide range of hydroxylation states (Yokota 1997; Bajguz and Tretyn 2003). The biosynthetic pathways for sterols from the cycloartenol precursor appear to be conserved across all land plants (Morikawa et al. 2009). The steps leading to the BRs have also been clarified recently with the predominant C28 pathway leading from 24-methylenecholesterol to castasterone, and then to brassinolide in certain species and tissues (Fujita et al. 2006; Nomura and Bishop 2006; Jager et al. 2007). While a complex matrix of steps is possible, recent work has suggested a likely dominant pathway. This has been possible, as the genes involved with this pathway have been identified in several model angiosperms, with cytochome P450 monooxygenases being responsible for the changing hydroxylation patterns. Indeed, C26 hydroxylation, one of the key deactivation processes for the BRs, may also result in the deactivation of insect steroidal hormones and the removal of cholesterol in mammals, showing strong evolutionary conservation in pathways that metabolise potentially dangerous sterols (Meaney 2005).
The isolation and characterisation of dwarf BR mutants was fundamental to the acceptance of BRs as growth-promoting hormones during the 1990s. Indeed, it was the complementation of the det2 mutant in A. thaliana by an animal 5α-reductase gene that really confirmed the hormonal status of BRs in plants (Li et al. 1997). Clear evidence was provided by the dark green dwarf phenotype of BR-deficient mutants in several angiosperm species (e.g. A. thaliana, pea, rice and tomato; Li et al. 1996; Bishop 2003; Nomura et al. 2004). However, while many processes have been linked with BR function (e.g. cell elongation, cell division, reproductive and vascular development, stress responses, senescence and etiolation), relatively few of these have been examined in sufficient depth to show a clear endogenous physiological role for BRs. This requires evidence that changes in endogenous BR levels, caused by environmental factors or autonomously during development, regulate the process in both a positive and negative fashion. Such evidence is only present for a few developmental processes including stem elongation (e.g. Nomura et al. 1997), xylem differentiation (Yamamoto et al. 2001) and fruit development (Symons et al. 2006), and then only in a select number of angiosperm species. Such physiological evidence is lacking from other plant groups.
In fact, information on BRs in plants other than the angiosperms is too patchy to indicate whether the biosynthetic pathways are identical to those in angiosperms but does suggest a similar range of compounds are present, especially in the gymnosperms (Fujioka 1999). In most cases, one or both of the known active molecules, castasterone or brassinolide, is (are) present. This at least provides the potential for BRs to be acting as a hormone in these systems, although they could simply be present as secondary metabolites. However, at present, we do not possess BR mutants in plant groups other than the angiosperms. There do not appear to be reports of BRs in fungi or bacteria, or in plant pathogens in general, which contrasts to what is known for auxin and the GAs.
BR signalling
The BR receptor was identified at a fairly early stage from A. thaliana via the isolation of the BR-insensitive mutant, bri1 (Clouse et al. 1996; Li and Chory 1997). Similar mutants have subsequently been identified in other species including pea (Nomura et al. 2003) and rice (Yamamuro et al. 2000; Bai et al. 2007), providing confirmation of a similar mode of perception in monocots and eudicots. Substantial progress on identifying elements of the transduction pathway has also been made using mutant analysis and molecular studies in A. thaliana, and it has emerged that the transduction pathway directly influences the expression of response genes (Vert and Chory 2006; Li and Jin 2007). The BRI1 receptor is a leucine-rich repeat receptor-like kinase that is located at the cell membrane, with the receptor domain on the external side of the membrane and with an intracellular kinase domain that is activated by BR binding. This domain is capable of transferring the external perception of the BR signal to the intracellular transduction pathway, which consists of several genetically-defined components, including a soluble glycogen synthase kinase 3-like kinase (BIN2), a phosphatase (BSU1) and two transcription factors (BZR1 and BZR2) (Bai et al. 2007; Li and Jin 2007). Bai et al. (2007) demonstrate that this transduction pathway from the cell surface to the regulation of transcription is also conserved between monocots and eudicots, although there has been some duplication of components in the different lineages. The BR receptor system is related to the transmembrane receptor kinase signal transduction pathways in animals (Torii 2004). There are similar receptor systems in bacteria and components may well have been recruited from elements of such systems. The BRI1 receptor in tomatoes also binds the proteinaceous hormone systemin, as well as BRs, but does not appear to be essential for systemin-induced responses (Scheer and Ryan 2002; Holton et al. 2008). Santner and Estelle (2009) conclude that signalling systems for BRs probably did not evolve until after the evolutionary split of mosses and vascular plants. Consistent with this view, Sasaki et al. (2007) did not show any relationship of plant receptor-like kinase genes in M. polymorpha to BR receptors. While several M. polymorpha gene sequences contained domains related to those in the BRI1 receptor, they did not contain the key extracellular domain involved with BR binding (S. E. Davidson, unpubl. data).
The system of perception and signal transduction for plant steroid hormones is often contrasted with that of animal systems, where an intracellular receptor–steroid complex is formed and directly regulates gene expression in the nucleus (Revelli et al. 1998). However, while some recent reports have also suggested that steroid reception may occur at the cell membrane in certain animal systems (Thomas et al. 2007), preliminary searches of animal genome databases do not reveal BRI1-like sequences. Further, receptor sequences similar to those in animals have not been found in plants (Clouse 2002). These observations imply that although similar compounds (steroids) are used by both plants and animals to regulate development, signalling systems for the compounds may have originated independently in the two life forms. This is possible, as it is hypothesised that recruitment of a control mechanism can occur backwards from the functional gene level.
Another difference between the plant and animal steroid receptors is that the main BR receptor gene in plants, BRI1, is expressed across all tissue types (Nomura et al. 2003), whereas in animals, the expression of the steroid receptors is tightly regulated in distinct tissue types (Williams 1997). However, BRI homologues in plants have gained functional specificity in the control of some developmental processes such as vascular differentiation (Caño-Delgado et al. 2004; Kim and Wang 2010). There is clearly an interesting area of research needed to clarify and confirm that the steroid hormone systems in plants and animals have evolved independently, even though some of the genes involved in BR biosynthesis in plants are similar to those in human systems (e.g. 5α reductase; Li et al. 1997).
There appears to be little interaction between bioactive GA and BR levels, in contrast to the effect of IAA on bioactive GA content (Jager et al. 2005). However, there have been numerous reports of interactions between IAA and BRs. Certainly, the expression of some genes is regulated by both hormones, although the number affected in this way is a small subset of the genes affected by the individual hormones (Goda et al. 2004). There is also evidence that bioactive BRs influence auxin transport (Li et al. 2005; Symons et al. 2008). A recent paper suggests that a member of the BR response pathway, BIN2 kinase, inactivates ARF2, leading to increased expression of auxin-induced genes (Vert et al. 2008). However, the purported synergistic enhancement of auxin-induced elongation by BR (Vert et al. 2008) is only weakly supported by the growth response data, raising questions about the overall conclusions of the study. Whatever the auxin–BR interactions, if any, prove to be, it is too early to speculate on their evolutionary origin until a clear understanding of the interactions is achieved in model angiosperms.
Conclusions and future directions
Our understanding of plant hormone evolution has improved rapidly over the last decade due to the identification of many of the hormone receptors and elements of the response pathways, and the availability of genome sequences for several model species. As next generation sequencing tools become widespread, we will see a plethora of genome sequences from all plant groups over the next few years that, when combined with detailed molecular data on the synthesis and response genes from angiosperm models, will allow a precise understanding of the evolution of all the plant hormone systems. It will be interesting to see how these systems have evolved to suit the different adaptive traits of the various life cycles that occur across the plant groups. Furthermore, it will be important to determine whether there are major differences between the gametophyte and sporophyte generations.
One intriguing question is whether the development of hormone systems was driven by adaptation to the terrestrial environment, was an essential precursor to terrestrial colonisation or was driven by the development of advanced multicellular growth. The recent identification of auxin and its proposed actions in a brown algal model, Ectocarpus siliculosus (Le Bail et al. 2010), may lend support to the latter hypothesis. However, this raises the question of how the systems were recruited in different phylogenetic groups (e.g. auxin in brown and green algae, and steroids in plants and animals). Once sufficient genomes are available, answers to these questions will be forthcoming. However, the lack of certain hormone-related genes in unicellular plants has already been suggested in some cases (e.g. for auxin synthesis genes; Rensing et al. 2008), probably arguing strongly for independent evolution of the systems in different multicellular groups.
What does appear to be clear is that many of the plant hormone systems have been exploited by phylogenetically distinct pathogens (e.g. auxin by Agrobacterium and certain insects, and GAs by Gibberella fujikuroi). While co-evolution of hosts and pathogens has been examined in detail for many processes, especially those related to resistance mechanisms, some hormone groups (e.g. BRs) still require examination. Given the central role that the plant growth hormones play in plant development, they are an ideal system for pathogen manipulation, since a plant is unlikely to be able to easily evolve resistance to major disruptions in these hormone systems. Furthermore, pathogens need to produce only small amounts of the hormone in question in order to dramatically affect resource allocation or structural development.
A further productive line of research will be to confirm the impression, drawn from the limited number of genomes sequenced outside the angiosperms, that, at least for auxin and the GAs, there has been considerable expansion of the number of genes involved with these hormone systems. Not only do the processes influenced by these groups of hormones appear to change, but the system has become genetically more complex and, possibly, the responses more rapid. Either the developmental processes themselves have been recruited and adapted to new roles, with the original hormone response system remaining in place, or the hormone response has been recruited by pre-existing developmental processes previously not involved with hormones. Future research should resolve this question.
Acknowledgements
We thank Drs Sandra Davidson and Gregory Jordan for critical comments on the manuscript, and the Australian Research Council for financial support.
Achard P, Genschik P
(2009) Releasing the brakes of plant growth: how GAs shutdown DELLA proteins. Journal of Experimental Botany 60, 1085–1092.
| Crossref | GoogleScholarGoogle Scholar | PubMed |

Anterola A,
Shanle E,
Mansouri K,
Schuette S, Karen R
(2009) Gibberellin precursor is involved in spore germination in the moss Physcomitrella patens. Planta 229, 1003–1007.
| Crossref | GoogleScholarGoogle Scholar | PubMed |
Bai M,
Zhang L,
Gampala SS,
Zhu S,
Song W,
Chong K, Wang Z
(2007) Functions of OsBZR1 and 14–3-3 proteins in brassinosteroid signaling in rice. Proceedings of the National Academy of Sciences of the United States of America 104, 13 839–13 844.
| Crossref | GoogleScholarGoogle Scholar | PubMed |
Bajguz A, Tretyn A
(2003) The chemical characteristics and distribution of brassinosteroids in plants. Phytochemistry 62, 1027–1046.
| Crossref | GoogleScholarGoogle Scholar | PubMed |
Basu S,
Sun H,
Brian L,
Quatrano RL, Muday GK
(2002) Early embryo development in Fucus distichus is auxin sensitive. Plant Physiology 130, 292–302.
| Crossref | GoogleScholarGoogle Scholar | PubMed |
Bishop GJ
(2003) Brassinosteroid mutants of crops. Journal of Plant Growth Regulation 22, 325–335.
| Crossref | GoogleScholarGoogle Scholar | PubMed |
Bömke C, Tudzynski B
(2009) Diversity, regulation, and evolution of the gibberellin biosynthetic pathway in fungi compared to plants and bacteria. Phytochemistry 70, 1876–1893.
| Crossref | GoogleScholarGoogle Scholar | PubMed |
Caño-Delgado A,
Yin Y,
Yu C,
Vafeados D,
Mora-García S,
Cheng JC,
Nam KH,
Li J, Chory J
(2004) BRL1 and BRL3 are novel brassinosteroid receptors that function in vascular differentiation in Arabidopsis. Development 131, 5341–5351.
| Crossref | GoogleScholarGoogle Scholar | PubMed |
Clouse SD
(2002) Brassinsoteroids: plant counterparts to animal steroid hormones? Vitamins and Hormones 65, 195–223.
| Crossref | GoogleScholarGoogle Scholar | PubMed |
Clouse SD,
Langford M, McMorris TC
(1996) A brassinosteroid-insensitive mutant in Arabidopsis thaliana exhibits multiple defects in growth and development. Plant Physiology 111, 671–678.
| Crossref | GoogleScholarGoogle Scholar | PubMed |
Cooke TJ,
Poli DB,
Sztein AE, Cohen JD
(2002) Evolutionary patterns in auxin action. Plant Molecular Biology 49, 319–338.
| Crossref | GoogleScholarGoogle Scholar | PubMed |
Cove D,
Bezanilla M,
Harries P, Quatrano R
(2006) Mosses as model systems for the study of metabolism and development. Annual Review of Plant Biology 57, 497–520.
| Crossref | GoogleScholarGoogle Scholar | PubMed |
Davidson SE,
Reid JB, Helliwell CA
(2006) Cytochromes P450 in gibberellin biosynthesis. Phytochemistry Reviews 5, 405–419.
| Crossref | GoogleScholarGoogle Scholar |
Dutcher SK,
Galloway RE,
Barclay WR, Poortinga G
(1992) Tryptophan analog resistance mutations in Chlamydomonas reinhardtii. Genetics 131, 593–607.
| PubMed |
Eklund DM,
Thelander M,
Landberg K,
Staldal V,
Nilsson A,
Johansson M,
Valsecchi I, Pederson ERA
(2010) Homologues of the Arabidopsis thaliana SHI/STY/LRP1 genes control auxin biosynthesis and affect growth and development in the moss Physcomitrella patens. Development 137, 1275–1284.
| Crossref | GoogleScholarGoogle Scholar | PubMed |
Fu X, Harberd NP
(2003) Auxin promotes Arabidopsis root growth by modulating gibberellin response. Nature 421, 740–743.
| Crossref | GoogleScholarGoogle Scholar | PubMed |
Fujita S,
Ohnishi T,
Watanabe B,
Yokota T,
Takatsuto S,
Fujioka S,
Yoshida S,
Sakata K, Mizutani M
(2006) Arabidopsis CYP90B1 catalyses the early C-22 hydroxylation of C27, C28 and C29 sterols. The Plant Journal 45, 765–774.
| Crossref | GoogleScholarGoogle Scholar | PubMed |
Fujita T,
Sakaguchi H,
Hiwatashi Y,
Wagstaff SJ,
Ito M,
Deguchi H,
Sato T, Hasebe M
(2008) Convergent evolution of shoots in land plants: lack of auxin polar transport in moss shoots. Evolution & Development 10, 176–186.
| Crossref |
PubMed |
Gay G,
Normand L,
Marmeisse R,
Sotta B, Debaud JC
(1994) Auxin overproducer mutants of Hebeloma cylindrosporum Romagnési have increased mycorrhizal activity. New Phytologist 128, 645–657.
| Crossref | GoogleScholarGoogle Scholar |
Goda H,
Sawa S,
Asami T,
Fujioka S,
Shimada Y, Yoshida S
(2004) Comprehensive comparison of auxin-regulated and brassinosteroid-regulated genes in Arabidopsis. Plant Physiology 134, 1555–1573.
| Crossref | GoogleScholarGoogle Scholar | PubMed |
Grubb CD, Abel S
(2006) Glucosinolate metabolism and its control. Trends in Plant Science 11, 89–100.
| Crossref | GoogleScholarGoogle Scholar | PubMed |
Hayashi K,
Kawaide H,
Notomi M,
Sakigi Y,
Matsuo A, Nozaki H
(2006) Identification and functional analysis of bifunctional ent-kaurene synthase from the moss Physcomitrella patens. FEBS Letters 580, 6175–6181.
| Crossref | GoogleScholarGoogle Scholar | PubMed |
Hayashi K,
Horie K,
Hiwatashi Y,
Kawaide H, Yamaguchi S , et al.
(2010) Endogenous diterpenes derived from ent-kaurene, a common gibberellin precursor, regulate protonema differentiation of the moss Physcomitrella patens. Plant Physiology 153, 1085–1097.
| Crossref | GoogleScholarGoogle Scholar | PubMed |
Hedden P, Phillips AL
(2000) Gibberellin metabolism: new insights revealed by the genes. Trends in Plant Science 5, 523–530.
| Crossref | GoogleScholarGoogle Scholar | PubMed |
Hedden P,
Phillips AL,
Rojas MC,
Carrera E, Tudzynski B
(2002) Gibberellin biosynthesis in plants and fungi: a case of convergent evolution? Journal of Plant Growth Regulation 20, 319–331.
| Crossref | GoogleScholarGoogle Scholar |
Hirano K,
Nakajima M,
Asano K,
Nishiyama T, Sakakibara H , et al.
(2007) The GID1-mediated gibberellin perception mechanism is conserved in the lycophyte Selaginella moellendorffii but not in the bryophyte Physcomitrella patens. The Plant Cell 19, 3058–3079.
| Crossref | GoogleScholarGoogle Scholar | PubMed |
Hirano K,
Ueguchi-Tanaka M, Matsuoka M
(2008) GID1-mediated gibberellin signaling in plants. Trends in Plant Science 13, 192–199.
| Crossref | GoogleScholarGoogle Scholar | PubMed |
Holton N,
Harrison K,
Yokota T, Bishop GJ
(2008) Tomato BRI1 and systemin wound signaling. Plant Signaling & Behavior 3, 54–55.
| PubMed |
Imaizumi T,
Kadota A,
Hasebe M, Wada M
(2002) Cryptochrome light signals control development to suppress auxin sensitivity in the moss Physcomitrella patens. The Plant Cell 14, 373–386.
| Crossref | GoogleScholarGoogle Scholar | PubMed |
Jager CE,
Symons GM,
Ross JJ,
Smith JJ, Reid JB
(2005) The brassinosteroid growth response in pea is not mediated by changes in gibberellin content. Planta 221, 141–148.
| Crossref | GoogleScholarGoogle Scholar | PubMed |
Jager CE,
Symons GM,
Nomura T,
Yamada Y,
Smith JJ,
Yamaguchi S,
Kamiya Y,
Weller JL,
Yokota T, Reid JB
(2007) Characterisation of two brassinosteroid C-6 oxidase genes in pea. Plant Physiology 143, 1894–1904.
| Crossref | GoogleScholarGoogle Scholar | PubMed |
Keeling CI,
Dullat HK,
Yuen M,
Ralph SG,
Jancsik S, Bohlmann J
(2010) Identification and functional characterization of monofunctional ent-copalyl diphosphate and ent-kaurene synthases in white spruce reveal different patterns for diterpene synthase evolution for primary and secondary metabolism in gymnosperms. Plant Physiology 152, 1197–1208.
| Crossref | GoogleScholarGoogle Scholar | PubMed |
Kim T-W, Wang Z-Y
(2010) Brassinosteroid signal transduction from receptor kinases to transcription factors. Annual Review of Plant Biology 61, 681–704.
| Crossref | GoogleScholarGoogle Scholar | PubMed |
Kim Y-S,
Sup YH,
Kim S-H, Kim S-K
(2002) Identification of a brassinosteroid, castasterone from Marchantia polymorpha. Bulletin of the Korean Chemical Society 23, 941–942.
| Crossref |
Koga J,
Adachi T, Hidaka H
(1991) IAA biosynthetic pathway from tryptophan via indole-3-pyruvic acid in Enterobacter cloacae. Agricultural and Biological Chemistry 55, 701–706.
Lau S,
Jürgens G, De Smet I
(2008) The evolving complexity of the auxin pathway. The Plant Cell 20, 1738–1746.
| Crossref | GoogleScholarGoogle Scholar | PubMed |
Lau S,
Shao N,
Bock R,
Jurgens G, De Smet I
(2009) Auxin signaling in algal lineages: fact or myth? Trends in Plant Science 14, 182–188.
| Crossref | GoogleScholarGoogle Scholar | PubMed |
Le Bail A,
Billoud B,
Kowalczyk N,
Kowalczyk M,
Gicquel M,
.
(2010) Auxin metabolism and function in the multicellular brown alga Ectocarpus siliculosus. Plant Physiology 153, 128–144.
| Crossref | GoogleScholarGoogle Scholar | PubMed |
Li J, Chory J
(1997) A putative leucine-rich repeat receptor kinase involved in brassinosteroid signal transduction. Cell 90, 929–938.
| Crossref | GoogleScholarGoogle Scholar | PubMed |
Li J, Jin H
(2007) Regulation of brassinosteroid signaling. Trends in Plant Science 12, 37–41.
| Crossref | GoogleScholarGoogle Scholar | PubMed |
Li J,
Nagpal P,
Vitart V,
McMorris TC, Chory J
(1996) A role for brassinosteroids in light-dependent development of Arabidopsis. Science 272, 398–401.
| Crossref | GoogleScholarGoogle Scholar | PubMed |
Li J,
Biswas MB,
Chao A,
Russell DW, Chory J
(1997) Conservation of function between mammalian and plant steroid 5α-reductases. Proceedings of the National Academy of Sciences of the United States of America 94, 3554–3559.
| Crossref | GoogleScholarGoogle Scholar | PubMed |
Li L,
Xu J,
Xu Z-H, Xue H-W
(2005) Brassinosteroids stimulate plant tropisms through modulation of polar auxin transport in Brassica and Arabidopsis. The Plant Cell 17, 2738–2753.
| Crossref | GoogleScholarGoogle Scholar | PubMed |
MacMillan J
(2002) Occurrence of gibberellins in vascular plants, fungi, and bacteria. Journal of Plant Growth Regulation 20, 387–442.
| Crossref | GoogleScholarGoogle Scholar |
Mano Y,
Nemoto K,
Suzuki M,
Seki H,
Fujii I, Muranaka T
(2010) The AMI1 gene family: indole-3-acetamide hydrolase functions in auxin biosynthesis in plants. Journal of Experimental Botany 61, 25–32.
| Crossref | GoogleScholarGoogle Scholar | PubMed |
Meaney S
(2005) Is C-26 hydroxylation an evolutionarily conserved steroid inactivation mechanism? The FASEB Journal 19, 1220–1224.
| Crossref | GoogleScholarGoogle Scholar | PubMed |
Morikawa T,
Saga H,
Hashizume H, Ohta D
(2009) CYP710A genes encoding sterol C22-desaturase in Physcomitrella patens as molecular evidence for the evolutionary conservation of a sterol biosynthetic pathway in plants. Planta 229, 1311–1322.
| Crossref | GoogleScholarGoogle Scholar | PubMed |
Mravec J,
Skupa P,
Bailly A,
Hoyerova K, Krecek P , et al.
(2009) Subcellular homeostasis of phytohormone auxin is mediated by the ER-localized PIN5 transporter. Nature 459, 1136–1140.
| Crossref | GoogleScholarGoogle Scholar | PubMed |
Nomura T, Bishop GJ
(2006) Cytochrome P450s in plant steroid hormone synthesis and metabolism. Phytochemistry Reviews 5, 421–432.
| Crossref | GoogleScholarGoogle Scholar |
Nomura T,
Nakayama M,
Reid JB,
Takeuchi Y, Yokota T
(1997) Blockage of brassinosteriod biosynthesis and sensitivity causes dwarfism in garden pea. Plant Physiology 113, 31–37.
| PubMed |
Nomura T,
Bishop GJ,
Kaneta T,
Reid JB,
Chory J, Yokota T
(2003) The LKA gene is a BRASSINOSTEROID INSENSITIVE 1 homolog of pea. The Plant Journal 36, 291–300.
| Crossref | GoogleScholarGoogle Scholar | PubMed |
Nomura T,
Jager CE,
Kitasaka Y,
Takeuchi K, Fukami M , et al.
(2004) Brassinosteroid deficiency due to truncated steroid 5α-reductase causes dwarfism in the lk mutant of pea. Plant Physiology 135, 2220–2229.
| Crossref | GoogleScholarGoogle Scholar | PubMed |
Nomura T,
Kushiro T,
Yokota T,
Kamiya Y,
Bishop GJ, Yamaguchi S
(2005) The last reaction producing brassinolide is catalyzed by cytochrome P-450s, CYP85A3 in tomato and CYP85A2 in Arabidopsis. The Journal of Biological Chemistry 280, 17 873–17 879.
| Crossref | GoogleScholarGoogle Scholar | PubMed |
Normanly J
(2010) Approaching cellular and molecular resolution of auxin biosynthesis and metabolism. Cold Spring Harbor Perspectives in Biology 2, a001594.
| Crossref | GoogleScholarGoogle Scholar | PubMed |
O’Neill DP, Ross JJ
(2002) Auxin regulation of the gibberellin pathway in pea. Plant Physiology 130, 1974–1982.
| Crossref | GoogleScholarGoogle Scholar | PubMed |
Palenik B,
Grimwood J,
Aerts A,
Rouze P, Salamov A , et al.
(2007) The tiny eukaryote Ostreococcus provides genomic insights into the paradox of plankton speciation. Plant Biology 104, 7705–7710.
Paponov IA,
Teale W,
Lang D,
Paponov M,
Reski R,
Rensing SA, Palme K
(2009) The evolution of nuclear auxin signaling. BMC Evolutionary Biology 9, 126.
| Crossref | GoogleScholarGoogle Scholar |
Pollmann S,
Duchting P, Weiler EW
(2009) Tryptophan-dependent indole-3-acetic acid biosynthesis by “IAA-synthase” proceeds via indole-acetamide. Phytochemistry 70, 523–531.
| Crossref | GoogleScholarGoogle Scholar | PubMed |
Quittenden LJ,
Davies NW,
Smith JA,
Molesworth PP,
Tivendale ND, Ross JJ
(2009) Auxin biosynthesis in pea: characterization of the tryptamine pathway. Plant Physiology 151, 1130–1138.
| Crossref | GoogleScholarGoogle Scholar | PubMed |
Remington DL,
Vision TJ,
Guilfoyle TJ, Reed JW
(2004) Contrasting modes of diversification in the Aux/IAA and ARF gene families. Plant Physiology 135, 1738–1752.
| Crossref | GoogleScholarGoogle Scholar | PubMed |
Rensing SA,
Lang D,
Zimmer AD,
Terry A, Salamov A , et al.
(2008) The Physcomitrella genome reveals evolutionary insights into the conquest of land by plants. Science 319, 64–69.
| Crossref | GoogleScholarGoogle Scholar | PubMed |
Revelli A,
Massobrio M, Tesarik J
(1998) Nongenomic actions of steroid hormones in reproductive tissues. Endocrine Reviews 19, 3–17.
| Crossref | GoogleScholarGoogle Scholar | PubMed |
Ross JJ,
O’Neill DP,
Smith JJ,
Kerckhoffs LHJ, Elliott RC
(2000) Evidence that auxin promotes gibberellin A1 biosynthesis in pea. The Plant Journal 21, 547–552.
| Crossref | GoogleScholarGoogle Scholar | PubMed |
Sakakibara K,
Nishiyama T,
Deguchi H, Hasebe M
(2008) Class 1 KNOX genes are not involved in shoot development in the moss Physcomitrella patens but do function in sporophyte development. Evolution & Development 10, 555–566.
| Crossref | GoogleScholarGoogle Scholar | PubMed |
Santner A, Estelle M
(2009) Recent advances and emerging trends in plant hormone signaling. Nature 459, 1071–1078.
| Crossref | GoogleScholarGoogle Scholar | PubMed |
Santner A,
Calderon-Villalobos LIA, Estelle M
(2009) Plant hormones are versatile chemical regulators of plant growth. Nature Chemical Biology 5, 301–307.
| Crossref | GoogleScholarGoogle Scholar | PubMed |
Sasaki G,
Katoh K,
Hirose N,
Suga H,
Kuma K,
Miyata T, Su Z-H
(2007) Multiple receptor-like kinase cDNAs from liverwort Marchantia polymorpha and two charophycean green algae, Closterium ehrenbergii and Nitella axillaris: extensive gene duplications and gene shufflings in the early evolution of streptophytes. Gene 401, 135–144.
| Crossref | GoogleScholarGoogle Scholar | PubMed |
Scheer JM, Ryan CA
(2002) The systemin receptor SR160 from Lycopersicon peruvianum is a member of the LRR receptor kinase family. Proceedings of the National Academy of Sciences of the United States of America 99, 9585–9590.
| Crossref | GoogleScholarGoogle Scholar | PubMed |
Shimada A,
Ueguchi-Tanaka M,
Nakatsu T,
Nakajima M,
Naoe Y,
Ohmiya H,
Kato H, Matsuoka M
(2008) Structural basis for gibberellin recognition by its receptor GID1. Nature 456, 520–523.
| Crossref | GoogleScholarGoogle Scholar | PubMed |
Splivallo R,
Fischer U,
Göbel C,
Feussner I, Karlovsky P
(2009) Truffles regulate plant root morphogenesis via the production of auxin and ethylene. Plant Physiology 150, 2018–2029.
| Crossref | GoogleScholarGoogle Scholar | PubMed |
Stepanova AN,
Robertson-Hoyt J,
Yun J,
Benavente LM,
Xie D-Y,
Dolezal K,
Schlereth A,
Jurgens G, Alonso JM
(2008) TAA1-mediated auxin biosynthesis is essential for hormone crosstalk and plant development. Cell 133, 177–191.
| Crossref | GoogleScholarGoogle Scholar | PubMed |
Sugawara S,
Hishiyama S,
Jikumaru Y,
Hanada A,
Nishimura T,
Koshiba T,
Zhao Y,
Kamiya Y, Kasahara H
(2009) Biochemical analyses of indole-3-acetaldoxime-dependent auxin biosynthesis Arabidopsis. Proceedings of the National Academy of Sciences of the United States of America 106, 5430–5435.
| Crossref | GoogleScholarGoogle Scholar | PubMed |
Symons GM,
Davies C,
Shavrukov Y,
Dry AB,
Reid JB, Thomas MR
(2006) Grapes on steroids. Brassinosteroids are involved in grape berry ripening. Plant Physiology 140, 150–158.
| Crossref | GoogleScholarGoogle Scholar | PubMed |
Symons GM,
Ross JJ,
Jager CE, Reid JB
(2008) Brassinosteroid transport. Journal of Experimental Botany 59, 17–24.
| Crossref | GoogleScholarGoogle Scholar | PubMed |
Sztein AE,
Cohen JD, Cooke TJ
(2000) Evolutionary patterns in the auxin metabolism of green plants. International Journal of Plant Sciences 161, 849–859.
| Crossref | GoogleScholarGoogle Scholar |
Tao Y,
Ferrer JL,
Ljung K,
Pojer F, Hong F , et al.
(2008) Rapid synthesis of auxin via a new tryptophan-dependent pathway is required for shade avoidance in plants. Cell 133, 164–176.
| Crossref | GoogleScholarGoogle Scholar | PubMed |
Thomas P,
Pang Y,
Dong J,
Groenen P,
Kelder J,
Vlieg J,
Zhu Y, Tubbs C
(2007) Steroid and G protein binding characteristics of the seatrout and human progestin membrane receptor α subtypes and their evolutionary origins. Endocrinology 148, 705–718.
| Crossref | GoogleScholarGoogle Scholar | PubMed |
Tiwari SB,
Hagen G, Guilfoyle TJ
(2004) Aux/IAA proteins contain a potent transcriptional repression domain. The Plant Cell 16, 533–543.
| Crossref | GoogleScholarGoogle Scholar | PubMed |
Torii KU
(2004) Leucine-rich repeat receptor kinases in plants: structure, function, and signal transduction pathways. International Review of Cytology 234, 1–46.
| Crossref | GoogleScholarGoogle Scholar | PubMed |
Ueguchi-Tanaka M,
Nakajima M,
Motoyuki A, Matsuoka M
(2007) Gibberellin receptor and its role in gibberellin signaling in plants. Annual Review of Plant Biology 58, 183–198.
| Crossref | GoogleScholarGoogle Scholar | PubMed |
Vandenbussche F,
Fierro AC,
Wiedemann G,
Reski R, Van Der Straeten D
(2007) Evolutionary conservation of plant gibberellin signaling pathway components. BMC Plant Biology 7, 65.
| Crossref | GoogleScholarGoogle Scholar | PubMed |
Vert G, Chory J
(2006) Downstream nuclear events in brassinosteroid signaling. Nature 441, 96–100.
| Crossref | GoogleScholarGoogle Scholar | PubMed |
Vert G,
Walche CL,
Chory J, Nemhauser JL
(2008) Integration of auxin and brassinosteroid pathways by auxin response factor 2. Proceedings of the National Academy of Sciences of the United States of America 105, 9829–9834.
| Crossref | GoogleScholarGoogle Scholar | PubMed |
Vieten A,
Sauer M,
Brewer PB, Friml J
(2007) Molecular and cellular aspects of auxin-transport-mediated development. Trends in Plant Science 12, 160–168.
| Crossref | GoogleScholarGoogle Scholar | PubMed |
von Schwartzenberg K,
Schultze W, Kassner H
(2004) The moss Physcomitrella patens releases a tetracyclic diterpene. Plant Cell Reports 22, 780–786.
| Crossref |
PubMed |
Williams GH
(1997) Steroid receptor specificity with reference to the estrogen receptor. Osteoporosis International 7(Suppl. 1), 35–39.
| Crossref | GoogleScholarGoogle Scholar |
Wolbang CM,
Chandler PM,
Smith JJ, Ross JJ
(2004) Auxin from the developing in fluorescence is required for the biosynthesis of active gibberellins in barley stems. Plant Physiology 134, 769–776.
| Crossref | GoogleScholarGoogle Scholar | PubMed |
Woodward AW, Bartel B
(2005) Auxin: regulation, action, and interaction. Annals of Botany 95, 707–735.
| Crossref | GoogleScholarGoogle Scholar | PubMed |
Yamaguchi S
(2008) Gibberellin metabolism and its regulation. Annual Review of Plant Biology 59, 225–251.
| Crossref | GoogleScholarGoogle Scholar | PubMed |
Yamamoto R,
Fujioka S,
Demura T,
Takatsuto S,
Yoshida S, Fukuda H
(2001) Brassinosteroid levels increase drastically prior to morphogenesis of tracheary elements. Plant Physiology 125, 556–563.
| Crossref | GoogleScholarGoogle Scholar | PubMed |
Yamamuro C,
Ihara Y,
Wu X,
Noguchi T,
Fujioka S,
Takatsuto S,
Ashikari M,
Kitano H, Matsuoka M
(2000) Loss of function of a rice brassinosteroid insensitive1 homolog prevents internode elongation and bending of the lamina joint. The Plant Cell 12, 1591–1606.
| Crossref |
PubMed |
Yamauchi T,
Oyama N,
Yamane H,
Murofushi N,
Schraudolf H,
Owen D, Mander LN
(1995) 3-epi-GA63, antheridiogen in Anemia phyllitidis. Phytochemistry 38, 1345–1348.
| Crossref | GoogleScholarGoogle Scholar |
Yamauchi T,
Oyama N,
Yamane H,
Murofushi N,
Schraudolf H,
Pour M,
Furber M, Mander LN
(1996) Identification of antheridiogens in Lygodium círcinnatum and Lygodium flexuosum. Plant Physiology 111, 741–745.
| PubMed |
Yasumura Y,
Crumpton-Taylor M,
Fuentes S, Harberd NP
(2007) Step-by-step acquisition of the gibberellin–DELLA growth-regulatory mechanism during land-plant evolution. Current Biology 17, 1225–1230.
| Crossref | GoogleScholarGoogle Scholar | PubMed |
Yokota T
(1997) The structure, biosynthesis and function of brassinosteroids. Trends in Plant Science 2, 137–143.
| Crossref | GoogleScholarGoogle Scholar |
Zhao Y
(2010) Auxin biosynthesis and its role in plant development. Annual Review of Plant Biology 61, 49–64.
| Crossref | GoogleScholarGoogle Scholar | PubMed |
Zhao Y,
Christensen SK,
Fankhauser C,
Cashman JR,
Cohen JD,
Weigel D, Chory J
(2001) A role for flavin monooxygenase-like enzymes in auxin biosynthesis. Science 291, 306–309.
| Crossref | GoogleScholarGoogle Scholar | PubMed |