

Tropical forest responses to increasing atmospheric CO2: current knowledge and opportunities for future research
Lucas A. Cernusak A H , Klaus Winter B , James W. Dalling C , Joseph A. M. Holtum B D , Carlos Jaramillo B , Christian Körner E , Andrew D. B. Leakey C , Richard J. Norby F , Benjamin Poulter G , Benjamin L. Turner B and S. Joseph Wright BA School of Marine and Tropical Biology, James Cook University, Cairns, Qld 4878, Australia.
B Smithsonian Tropical Research Institute, PO Box 0843-03092, Balboa, Ancon, Republic of Panama.
C Department of Plant Biology, University of Illinois, Urbana-Champaign, Urbana, IL 61801, USA.
D School of Marine and Tropical Biology, James Cook University, Townsville, Qld 4811, Australia.
E Institute of Botany, University of Basel, Basel, CH-4056, Switzerland.
F Environmental Sciences Division and Climate Change Science Institute, Oak Ridge National Laboratory, Oak Ridge, TN 37831, USA.
G Laboratoire des Sciences du Climat et de l’Environnement, Gif sur Yvette French Centre National de la Recherche Scientifique, the Atomic Energy Commission and the University of Versailles Saint-Quentin, 91191, France.
H Corresponding author. Email: lcernusak@gmail.com
Functional Plant Biology 40(6) 531-551 https://doi.org/10.1071/FP12309
Submitted: 20 October 2012 Accepted: 21 March 2013 Published: 16 May 2013
Journal Compilation © CSIRO Publishing 2013 Open Access CC BY-NC-ND
Abstract
Elevated atmospheric CO2 concentrations (ca) will undoubtedly affect the metabolism of tropical forests worldwide; however, critical aspects of how tropical forests will respond remain largely unknown. Here, we review the current state of knowledge about physiological and ecological responses, with the aim of providing a framework that can help to guide future experimental research. Modelling studies have indicated that elevated ca can potentially stimulate photosynthesis more in the tropics than at higher latitudes, because suppression of photorespiration by elevated ca increases with temperature. However, canopy leaves in tropical forests could also potentially reach a high temperature threshold under elevated ca that will moderate the rise in photosynthesis. Belowground responses, including fine root production, nutrient foraging and soil organic matter processing, will be especially important to the integrated ecosystem response to elevated ca. Water use efficiency will increase as ca rises, potentially impacting upon soil moisture status and nutrient availability. Recruitment may be differentially altered for some functional groups, potentially decreasing ecosystem carbon storage. Whole-forest CO2 enrichment experiments are urgently needed to test predictions of tropical forest functioning under elevated ca. Smaller scale experiments in the understorey and in gaps would also be informative, and could provide stepping stones towards stand-scale manipulations.
Additional keywords: carbon storage, CO2 enrichment, liana, phosphorus, succession, water use efficiency.
Introduction
The rise in atmospheric CO2 concentration (ca) caused by human industrialisation is unprecedented, rapid and ubiquitous. Like all vegetation on earth, tropical forests existed under a ca less than 300 parts per million (ppm) for at least 800 000 years before the start of the 20th century (Lüthi et al. 2008). The ca rose from 300 ppm early in the twentieth century to 392 ppm in 2011, and projections for intermediate emissions scenarios suggest it could exceed 800 ppm by the year 2100 (Intergovernmental Panel on Climate Change 2011). Because CO2 is the primary substrate for photosynthesis, this dramatic increase in ca will undoubtedly affect the metabolism of tropical forests worldwide. The qualitative and quantitative expression of such effects, however, is largely unknown and represents a major source of uncertainty that limits our capacity to understand tropical ecosystem processes, assess their vulnerabilities to climate change and improve their representation in Earth system models.
Tropical forests play a significant role in the global carbon cycle. They contain about half the carbon stored in plant biomass in the terrestrial biosphere and account for about one-third of global terrestrial productivity (Field et al. 1998; Malhi and Grace 2000; Roy et al. 2001; Beer et al. 2010; Pan et al. 2011; Saatchi et al. 2011). In general, responses to elevated ca have been studied far less in tropical forests than in temperate forests (Hogan et al. 1991; Stork et al. 2007; Körner 2009; Luo et al. 2011; Leakey et al. 2012). This gap in research effort may partly reflect the challenges associated with studying tropical forest communities, given their large stature and biological complexity, and logistical challenges associated with conducting research in tropical environments. Free air CO2 enrichment (FACE) experiments conducted in temperate forests indicated that enriching ca to 550 ppm caused a 23% increase in net primary productivity (NPP) compared with that observed at ambient ca of ~380 ppm (Norby et al. 2005), with one experiment showing a subsequent decrease to 9% NPP stimulation caused by limited nitrogen availability (Norby et al. 2010). No FACE experiment has been conducted in a tropical forest for comparison so far.
Tropical ecosystems differ from temperate ecosystems in important climatic, edaphic, floristic and ecological attributes, and these are likely to influence how they respond to rising ca. High tropical temperatures increase the potential for stimulation of net photosynthesis (A) by elevated ca through suppression of photorespiration compared with predictions for cooler temperate and boreal ecosystems (Farquhar et al. 1980; Long 1991). On the other hand, it has been argued that tropical forest plants may be near a high temperature threshold, beyond which A could decline (Doughty and Goulden 2008; Doughty 2011). It is not known how the negative effects of high temperature on A will interact with the positive effects of high ca. In addition, it is possible that biomass production and the competitive ability of tropical canopy trees are not carbon-limited at the current ca, such that increasing A may have little influence on overall growth performance (Körner 2003, Körner 2009).
FACE experiments have indicated that nitrogen availability plays an important role in constraining productivity responses to elevated ca in temperate forests (Norby and Zak 2011). However, nitrogen availability is high in many tropical forests (Hedin et al. 2009; Brookshire et al. 2012). Phosphorus or some other rock-derived nutrient, rather than nitrogen, could present the primary nutritional constraint on growth responses to elevated ca in tropical ecosystems (Quesada et al. 2010; Vitousek et al. 2010).
Shifts in floristic composition due to elevated ca exposure may also be more important in tropical ecosystems. For example, woody climbing plants (lianas) and potentially N2-fixing legumes are far more abundant in tropical forests than in temperate forests. Functional type-specific responses in these groups could have significant consequences for tropical forest structure and function under elevated ca, as could an increasing abundance of light-wooded and relatively short-lived pioneer species.
We explore these issues in more detail below, and highlight the challenges and opportunities associated with tackling them experimentally. Our goal is to provide a framework that can be used to guide future experimental research aimed at understanding how woody plants in tropical forests will respond to rising ca. Conjecture and speculation are necessary ingredients in this endeavour, due to the relatively small amount of research that has thus far been conducted into tropical forests’ responses to elevated ca.
Effects of elevated ca on leaf gas exchange
In this section, we discuss the effects of elevated ca on leaf-level A, typically expressed as CO2 uptake per unit leaf area per unit of time. For a fixed leaf area index, an increase in A will cause a proportional increase in gross primary productivity (GPP). GPP describes the rate of photosynthetic carbon uptake from the atmosphere by a plant canopy, typically expressed per unit of ground area per unit of time.
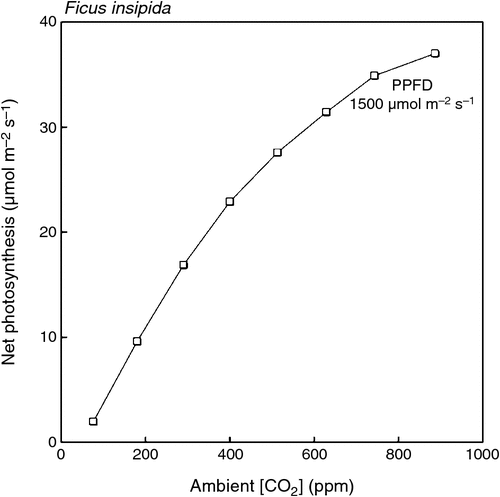
Elevated ca generally causes A to increase in plants that use the C3 photosynthetic pathway (Lloyd and Farquhar 1996; Drake et al. 1997). The overwhelming majority of tropical woody plants use this photosynthetic pathway, with notable exceptions in the genera Euphorbia (Pearcy and Troughton 1975) and Clusia (Holtum et al. 2004). An example of the short-term response of A to ca is shown in Fig. 1 for a seedling of a C3 tropical pioneer tree, Ficus insipida Willd. (K. Winter, unpublished). The measurements were made under near optimal conditions for photosynthesis in a tropical C3 plant. Fig. 1 clearly shows the potential for significant increases in A in response to rising ca.
|
In principle, the A of tropical woody plants has greater potential to respond positively to elevated ca than that of plants at higher latitudes, as a result of the higher leaf temperatures associated with tropical climates. Rubisco is the primary carboxylating enzyme in C3 plants. Competition between CO2 and O2 at the active sites of Rubisco causes the enzyme to catalyse the fixation of both of these substrates. Fixation of O2 by Rubisco leads to the release of CO2 from mitochondria through the process of photorespiration. As leaf temperature increases, the specificity of Rubisco for fixing CO2 instead of O2 decreases and the solubility of CO2 relative to O2 also decreases. Therefore, photorespiration increases as a proportion of gross photosynthesis with increasing leaf temperature (Farquhar et al. 1980; Long 1991). On the other hand, photorespiration can be suppressed by increasing the concentration of CO2 relative to that of O2 around the active sites of Rubisco. The upshot is that there is greater opportunity to increase net CO2 uptake by suppressing photorespiration at higher temperatures. Further predictions resulting from consideration of Rubisco kinetics that are relevant to tropical forest responses to elevated ca are: (1) the temperature optimum for A should increase with increasing ca; (2) the proportional increase in the maximum quantum yield of CO2 uptake caused by elevated ca should increase with increasing leaf temperature; and (3) the proportional decrease in the light compensation point caused by elevated ca should be larger at higher than at lower leaf temperatures (Farquhar et al. 1980; Long 1991).
These predictions of the interaction between elevated ca and temperature at the leaf level are also apparent when A is scaled up to the canopy (Long 1991). As a result, ecosystem models generally predict a larger proportional stimulation of NPP in warm tropical climates, compared with cooler, higher latitudes. For example, a global dynamic vegetation model employing a modified version of the Farquhar et al. photosynthesis model (Farquhar et al. 1980; Collatz et al. 1991) predicted a 35% increase in NPP for tropical forests at ca of 550 ppm relative to that at ca of 370 ppm, whereas the predicted increase for temperate forests was 26%. This geographic difference in the simulated proportional stimulation of NPP was largely caused by predicted differences in photorespiration, assuming sufficient nutrient and water availability to support increased NPP (Hickler et al. 2008).
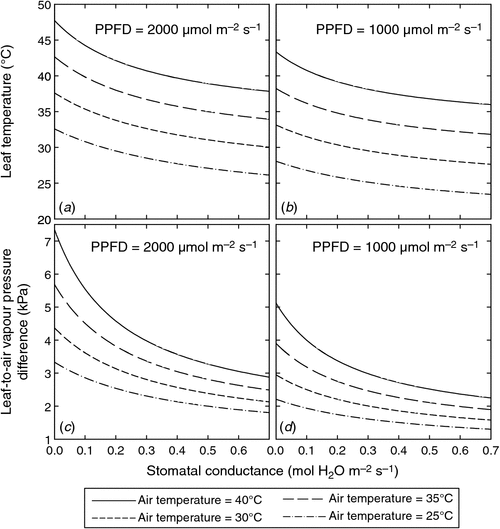
On the other hand, the proportional increase in A in response to ca may be damped if environmental conditions are not otherwise favourable for photosynthetic gas exchange. One reason this might occur is if the stomata close to slow the rate of water loss from leaves. Stomatal conductance (gs) typically decreases in response to increasing leaf-to-air vapour pressure difference (VPD), and A typically shows a linear or curvilinear relationship with gs (Wong et al. 1979; Cernusak and Marshall 2001; Cernusak et al. 2011a; Medlyn et al. 2011). Thus, a reduction in gs caused by increased VPD will also cause a reduction in A, with the proportional reduction in A likely to be somewhat less than that in gs (Farquhar and Sharkey 1982). The VPD can vary as a function of the vapour pressure of the air surrounding the leaf or as a function of the vapour pressure inside the leaf. Because the vapour pressure inside leaves is assumed to be at or near saturation, it is effectively controlled by leaf temperature. Thus, for a fixed vapour pressure outside the leaf, the VPD increases exponentially as leaf temperature increases, and gs and A are likely to decrease accordingly (Sage and Kubien 2007; Lloyd and Farquhar 2008).
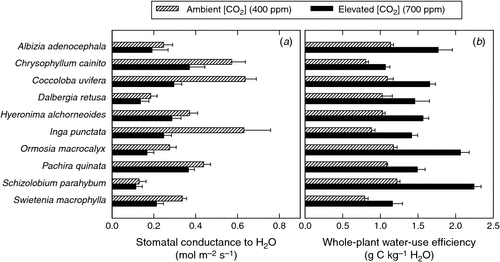
Leaf temperatures in tropical forests are expected to increase with increasing ca as a result of increasing air temperature and the increasing elevation of leaf temperature above air temperature. Air temperature is expected to increase due to the radiative effects of CO2 and other greenhouse gases in the atmosphere, and due to decreased evapotranspiration (Sellers et al. 1996). Over the last century, the global average surface temperature increased by 0.74°C, accompanying an increase in ca from 280 ppm to 380 ppm between 1750 and 2005 (Solomon et al. 2007). In tropical forest regions, the rate of surface warming since the mid-1970s has averaged 0.26°C per decade (Malhi and Wright 2004). At the leaf level, gs typically declines in response to elevated ca (Wullschleger et al. 2002; Ainsworth and Rogers 2007), although not always (Körner and Würth 1996; Keel et al. 2007). Declining gs in response to elevated ca has been observed in seedlings and saplings of tropical tree species (Berryman et al. 1994; Goodfellow et al. 1997; Cernusak et al. 2011b). An example of the response of gs to growth at elevated compared with ambient ca is shown in Fig. 2a for the seedlings of 10 tropical tree species. Here, it can be clearly seen that gs generally decreased in response to elevated ca but the response was variable among species. Lower gs results in a lower transpiration rate (E), causing an increase in leaf temperature (Fig. 3a, b), with associated increases in VPD (Fig. 3c, d).
|
|
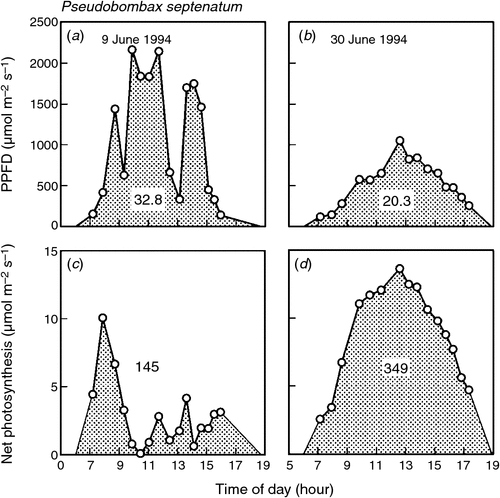
Many tropical forest trees display a pronounced midday depression in both gs and A on sunny days (Roy and Salager 1992; Koch et al. 1994; Zotz et al. 1995; Ishida et al. 1999; Kosugi et al. 2009). This has also been detected at the canopy scale with eddy covariance measurements (Goulden et al. 2004; Doughty and Goulden 2008; Kosugi et al. 2008). It coincides with the high VPD associated with increasing leaf temperatures under high irradiance (Fig. 4). The midday depression in gs and A occurs independently of soil moisture status (Kosugi et al. 2009). For example, the data shown in Fig. 4 were recorded during the rainy season in Panama, when soil moisture was high. Thus, as ca continues to rise over the coming century, higher VPD caused by higher leaf temperatures could cause midday depressions of gs and A to occur more frequently and for longer periods during the day. This would dampen the positive response of A to rising ca, although an overall increase in A is still predicted (Lloyd and Farquhar 2008).
|
Nonstomatal effects can also limit A at high leaf temperatures. The light-saturated potential electron transport rate appears to have a temperature optimum of ~40°C in tropical tree leaves (Lloyd et al. 1995; Mercado et al. 2006). Thus, A would be expected to decrease for a given chloroplastic CO2 concentration at leaf temperatures higher than 40°C. Exposure to leaf temperatures higher than 45°C can cause denaturation of Rubisco and other photosynthetic enzymes (Berry and Björkman 1980). Necrosis and tissue death typically occur after exposure to leaf temperatures between 50 and 53°C (Krause et al. 2010). Sun-exposed outer canopy leaves in a tropical forest reached temperatures between 46°C and 48°C, suggesting they may be operating near their limit of heat tolerance under current conditions (Krause et al. 2010).
An in situ warming experiment recently demonstrated that A in tropical tree and liana leaves was significantly reduced by a warming of 2–3°C over a 13-week period, with the leaf temperatures of warmed leaves reaching 45°C (Doughty 2011). This is within the range of leaf warming that could be expected to occur in response to elevated ca in the next few decades. The reduction in A was attributed mainly to nonstomatal limitations. It has been suggested that elevated ca could mitigate the adverse effects of elevated temperatures on the photosynthetic performance of tropical tree leaves (Hogan et al. 1991) through the suppression of photorespiration and the associated alleviation of photoinhibition under high irradiance (Kriedemann et al. 1976; Rasineni et al. 2011). Furthermore, the gs of tropical tree seedlings was observed to decline less in response to increasing leaf temperature for seedlings grown under elevated ca compared with those grown under ambient ca (Berryman et al. 1994). Thus, elevated ca has the potential to alleviate both the stomatal and nonstomatal limitations on A associated with high leaf temperatures. Future experiments that expose tropical tree leaves to both warming and elevated ca will be critical for testing this hypothesis further.
Growth under elevated ca has been shown to cause acclimation of photosynthetic capacity in a range of species, generally explained by reduced Rubisco activity, which is often correlated with reduced leaf nitrogen concentrations (Drake et al. 1997). In general, this type of acclimation allows plants to optimise overall performance by balancing sink and source activity. For example, when Rubisco expression is reduced, nitrogen can be reallocated away from the photosynthetic apparatus to structures such as fine roots that allow increased nutrient foraging (Moore et al. 1999; Long et al. 2004). In FACE experiments with temperate forest trees, the maximum Rubisco carboxylation velocity decreased by ~7% in response to growth at ca elevated to 200 ppm above ambient CO2 concentration (Ainsworth and Long 2005). With this lowered photosynthetic capacity, light-saturated photosynthesis was still stimulated by 47% in the elevated ca treatments. So far, no comparable data have been collected in tropical forest trees.
The above considerations suggest that both stomatal and nonstomatal limitations on A will increase with increasing leaf temperatures in tropical canopies, and there is further potential for acclimation or downregulation of photosynthetic capacity to balance source and sink activity. The nature and extent of the interaction between these limitations and the positive effects of elevated ca on A and GPP in tropical forests will have far-reaching consequences for tropical forest function under elevated ca.
Effects of elevated ca on growth
NPP is the net amount of carbon fixed into organic matter in a given time after accounting for autotrophic respiration (Eqn 1):

where Ra is autotrophic (plant) respiration. NPP can be calculated as the change in plant mass over time plus nonrespiratory carbon losses (i.e. tissue turnover, reproduction, herbivory, exudation of organic compounds from roots, biogenic volatile emissions, etc.), such that NPP = dM/dt + L, where M is the mass of carbon in a plant or a community of plants, t is time, dM/dt is change in mass over change in time, and L is nonrespiratory carbon losses (Lloyd and Farquhar 1996). Thus, an increase in NPP may or may not lead to an increase in ecosystem carbon storage in plant biomass (denoted by M), depending on how NPP is partitioned between dM/dt and L.
From the few datasets so far published, tropical forests appear to function with a relatively low carbon use efficiency (Chambers and Silver 2004; Malhi 2012), defined as the ratio of NPP to GPP. This indicates proportionally high Ra and the potential for a shift in Ra to significantly affect NPP (Metcalfe et al. 2010). To date, little is known about whether or how Ra will be affected by elevated ca in tropical forests. Increasing temperature or increasing supply of respiratory substrates associated with increasing ca could be expected to cause an increase in maintenance respiration (Leakey et al. 2009; Clark et al. 2010). On the other hand, responses to temperature may be tempered by acclimation, such that maintenance respiration rates are affected little by a gradual shift in temperature regime (Atkin et al. 2005). Nitrogen allocation to tropical tree leaves was observed to decrease under elevated ca (Berryman et al. 1993; Winter et al. 2000; Cernusak et al. 2011b), and this could cause a decrease in leaf maintenance respiration at a given temperature (Ryan 1995; Gonzàlez-Meler et al. 2009). Leaf dark respiration comprises more than a third of total Ra in tropical forests (Chambers and Silver 2004; Cavaleri et al. 2008). Not surprisingly, the way Ra is treated in ecosystem models has a large impact on predictions of the tropical forest NPP response to rising ca, especially with regard to the temperature dependence of Ra (Galbraith et al. 2010).
Shifting allocation patterns could affect how the change in biomass over time, dM/dt, responds to elevated ca in tropical forests. In artificial tropical miniecosystems, it was observed that A nearly doubled under elevated ca. However, the extra photosynthate produced in the elevated ca treatment was mostly allocated to increased fine root production and root exudation, thereby increasing L, rather than to aboveground biomass and coarse roots, which would have accelerated the increase in M (Körner and Arnone 1992). Similar responses were also observed in some temperate ecosystems (Körner et al. 2005; Norby et al. 2010), as originally foreshadowed by Strain and Bazzaz (1983). Across a broad range of tropical forest plots, a tradeoff was observed between NPP allocation to fine root production versus allocation to wood production, with allocation to the canopy remaining relatively invariant (Malhi et al. 2011). This suggests that increased allocation to fine root production under elevated ca could cause L to increase at the expense of increased allocation to the production of long-lived woody tissues, which would cause M to increase.
Much of the control over dM/dt in plants has traditionally been attributed to changes in A. This has recently been termed a ‘carbon-centric’ perspective (Sala et al. 2012). Alternatively, it has been suggested that demand for photosynthate at the sites of new tissue synthesis could exert greater control over dM/dt than does A (Körner 2003). This would require that the carbohydrates produced by A in excess of their consumption by anabolic processes must be lost from the plant through Ra or L. Under such a scenario, it is also likely that some fraction of the excess photosynthate would accumulate as nonstructural carbohydrates (NSC). Thus, analysis of the NSC concentrations in trees could provide a sensor that indicates carbon shortage or surplus for fuelling anabolic metabolism (Körner 2003). A steady and very high NSC concentration would indicate that the current level of A provides either a fully adequate supply or an oversupply of reduced carbon compounds to the plant.
In a seasonal forest in Panama, NSC concentrations were observed to be steady through the year or to increase during the dry season (Newell et al. 2002; Würth et al. 2005). This was interpreted to suggest that dM/dt was not limited by A (Körner 2003; Würth et al. 2005). Increasing ca around the canopy leaves at the same site led to increases in A and in NSC concentrations (Würth et al. 1998b; Lovelock et al. 1999), but did not affect the growth rates of branches. This further reinforced the idea that growth will not respond to increased A in these trees, although increased growth in the season following exposure to elevated ca could not be ruled out (Lovelock et al. 1999). This interpretation was not supported by the results of another experiment at the same site, in which supplemental lighting was provided to canopy trees under cloudy skies. In that case, growth increased, which was driven by increases in A (Graham et al. 2003). A definitive test of the hypothesis will require longer-term experiments on woody species growing under elevated ca in tropical forests.
It was recently suggested that NSC may play an important physiological role in maintaining the integrity of the vascular system of large woody plants (Sala et al. 2012). It was also suggested that increased allocation to the NSC pool may be a general response to stress, indicating more severe carbon limitation to growth, rather than vice versa (Wiley and Helliker 2012). Further, high soluble sugar concentrations in the leaves may improve their ability to photosynthesise at high temperatures (Hüve et al. 2006). If these assertions are correct, there may not be a direct negative relationship between the size of the NSC pool and the ability of tropical trees and lianas to increase growth in response to elevated ca. On the other hand, if NSC concentrations are generally high, and this reflects an oversupply of carbon for growth such that growth will not respond to a stimulation of A (Körner and Arnone 1992; Bader et al. 2010), this should be incorporated into coupled climate–carbon models, which treat carbon reserve pools very simply, if at all.
Growth at low nutrient availability consistently reduces the percentage growth response to elevated ca (McMurtrie et al. 2008). This pattern was demonstrated in experiments with seedlings of many tropical tree species (Oberbauer et al. 1985; Reekie and Bazzaz 1989; Ziska et al. 1991; Lovelock et al. 1998; Winter et al. 2000; Winter et al. 2001a, 2001b; Cernusak et al. 2011b; de Oliveira et al. 2012), although some exceptions also occurred (Körner and Arnone 1992; Arnone and Körner 1995; Carswell et al. 2000). Overall, it is likely that nutrient availability will play a major role in determining the productivity responses of woody tropical forest plants to rising ca.
Nitrogen appears to be relatively abundant in tropical forests, as indicated by high rates of nitrogen loss (Hedin et al. 2009; Brookshire et al. 2012) and increasing rates of long-term atmospheric nitrogen deposition (Chen et al. 2010; Hietz et al. 2011). However, vast areas of tropical forests occur on old, stable landscapes (e.g. large parts of South America, Africa, South-East Asia and Australia). In these regions, severely phosphorus-impoverished soils, a result of prolonged weathering in moist climates (Lambers et al. 2008), present a major constraint on plant growth (Vitousek et al. 2010). It was recently demonstrated that total soil phosphorus status was the measure of soil fertility that best predicted variation in productivity across a wide range of Amazonian forest plots (Quesada et al. 2012). In addition to low phosphorus availability in tropical soils, an apparent link between transpiration and phosphorus acquisition may further diminish the concentrations of phosphorus in plant tissues as ca rises (Cernusak et al. 2011c). On the other hand, it has been argued that phosphorus availability and uptake could be maintained under rising ca by the strong buffer power of soils for inorganic phosphorus, and by increased carbon allocation to mycorrhizal fungi and other specialised mechanisms for phosphorus acquisition (Lovelock et al. 1996; Lloyd et al. 2001; Lloyd and Farquhar 2008; Turner 2008). Organic phosphorus is abundant in tropical forest soils (Johnson et al. 2003; Vincent et al. 2010; Turner and Engelbrecht 2011) and may further support any increased growth of tropical forest trees under elevated ca.
Although phosphorus is likely to be the nutrient that constrains productivity in tropical forests in general, it is also worth pointing out that considerable heterogeneity exists across the tropical biome such that other nutrients can also be limiting. For example, nitrogen appeared to limit productivity in tropical forests on very young and very old soils (Fyllas et al. 2009; Mercado et al. 2011), in recently established secondary forests (Davidson et al. 2007) and in tropical montane forests (Tanner et al. 1998). Recent research has shown that the capacity for nitrate assimilation in the shoots of C3 plants decreases under elevated ca as a result of decreased photorespiration (Rachmilevitch et al. 2004; Bloom et al. 2012). If this pattern extends to woody tropical forest plants, the limitations imposed on tropical forest productivity by nitrogen availability could increase. Other nutrients may also play important roles in regulating productivity in tropical forests, for example, calcium, molybdenum and potassium (Vitousek 1984; Barron et al. 2009; Wright et al. 2011), and their availability could also constrain productivity responses to elevated ca.
It is clear that there are many unresolved issues associated with predicting whether and to what extent tropical forest NPP will be stimulated by future increases in ca. Tropical field experiments are urgently needed to address these issues (Leakey et al. 2012).
Effects of elevated ca on resistance to drought
Both seasonal and interannual droughts significantly impact upon the productivity and species composition of tropical forests (Condit et al. 1995; Engelbrecht et al. 2007; Nepstad et al. 2007; Brando et al. 2008; Phillips et al. 2009; da Costa et al. 2010). Elevated ca could make woody tropical forest plants more able to withstand drought in two ways. First, elevated ca could increase water use efficiency (WUE), thereby allowing a greater amount of photosynthesis for a given amount of water transpired to the atmosphere (Eamus 1991; Winter et al. 2001a; Battipaglia et al. 2013). This could lead to a reduction in soil water depletion due to reduced canopy-scale transpiration, which could sustain transpiration, and therefore photosynthesis, for a longer time between rain events (Morgan et al. 2004; Keel et al. 2007; Leuzinger and Körner 2007; Holtum and Winter 2010; Leuzinger and Körner 2010; Macinnis-Ng et al. 2011). Such water savings could also facilitate microbial activity and nutrient provision, and enable turgor pressure in meristem tissues to remain above the critical threshold required for cell expansion (Boyer 1968; Eamus et al. 1995), thereby facilitating growth. Second, elevated ca could increase the NSC pool, which, in turn, could be used to sustain plant metabolism for longer periods following stomatal closure and cessation of A in response to drought, according to the carbon-centric perspective (Sala et al. 2012).
Because A tends to increase under elevated ca for a given gs, leaf-level WUE typically increases. Leaf-level WUE can be defined as the ratio of photosynthesis to transpiration, A/E. A/E can be defined as the ratio of the diffusion gradient for CO2 to that for water vapour between the external air and the intercellular air spaces in the leaf (Farquhar and Richards 1984):

where ci is the intercellular CO2 concentration. The factor 1.6 in the denominator is the ratio of the diffusivity of water vapour to that of CO2 in the stomatal pores. Eqn (2) shows that if ci/ca remains constant, A/E will increase proportionally with increasing ca, so long as VPD is also constant. If VPD also increases as ca increases, as suggested above, this will damp the response of A/E to increasing ca (Barton et al. 2012). Variation in ci/ca in response to elevated ca can be assessed instantaneously by measuring changes in the CO2 and water vapour concentrations of air passing over a leaf (von Caemmerer and Farquhar 1981). In addition, time-integrated assessments of ci : ca can be obtained by measuring carbon-isotope discrimination (Δ13C) in plant biomass (Farquhar et al. 1982). Although Eqn (2) applies at the leaf level, the increase in WUE under elevated ca is also manifested at the whole-plant level, as shown in Fig. 2b for seedlings of 10 tropical tree species.
Measurement of Δ13C provides an opportunity to examine the historical responses of ci/ca to increasing ca in tropical trees since preindustrial times. This can be accomplished by analysing Δ13C in tree rings or in leaf dry matter preserved in herbaria. The few studies conducted so far on tropical trees show that ci/ca tended to remain constant as ca increased from preindustrial to present concentrations (Hietz et al. 2005; Nock et al. 2011; Bonal et al. 2011; Loader et al. 2011), or that ci/ca decreased in a tropical dry forest tree species (Brienen et al. 2011). Both trends (constant ci/ca and decreasing ci/ca), would indicate large increases in A/E as ca increased from 280 ppm to 380 ppm if VPD also remained constant. However, historical changes in VPD are more difficult to determine. As noted above, decreasing gs in response to increasing ca should lead to an increase in leaf temperature associated with a decrease in evaporative cooling of the leaf by transpiration (Fig. 3). This could lead to an increase in VPD, which would then dampen the increase in A/E caused by increasing ca. Overall, it seems likely that A/E has increased over the past century and will continue to increase as ca increases. This suggestion is consistent with the response of whole-plant WUE to elevated ca, as shown in Fig. 2b, because the whole-plant response incorporates the increase in VPD associated with lower gs.
It is critical to determine how stomatal responses to ca and VPD are likely to influence evapotranspiration, cloud formation and precipitation patterns in tropical regions as ca rises. At the continental scale, an increase in WUE in tropical forests could have important implications for the hydrological cycle, including increased runoff (Gedney et al. 2006). If WUE increases more than NPP, excess water is likely to enter riverine systems. This could accelerate weathering processes and the export of sediments and associated nutrients to the ocean. Conversely, a decrease in the amount of water returned to the atmosphere by transpiration could cause a decrease in cloud formation and precipitation (Betts et al. 2004). Models provide an opportunity to investigate this complex web of feedback over decadal to centennial time scales (Luo et al. 2011). Assessment of historical changes in ci/ca through analyses of Δ13C and experimental investigations of the responses of gs and ci/ca to elevated ca provide a means to parameterise or constrain such models (Buckley 2008; de Boer et al. 2011; Prentice et al. 2011).
Growth under water deficit generally causes NSC concentrations to increase, and this may be because cell expansion is more sensitive to water stress than is A (Hsiao 1973; Chaves et al. 2003; Muller et al. 2011). This, combined with the considerations described above, led Körner (2009) to predict that for tropical trees, ‘CO2 would have few if any effects under periodic drought, given the tendency for growth to be controlled by carbon sinks when water is in short supply.’ This prediction is based on the idea that the NSC pool represents a passive overflow or repository for carbon supply. However, it has been shown experimentally that growth can respond to elevated ca under water deficit in tropical tree seedlings (Cernusak et al. 2011b). Seedlings of two tropical tree species were grown at ambient and elevated ca, and at a volumetric soil water content of 0.27 or 0.08 m3 m–3. The low water supply was sufficient to reduce gs to less than half that observed at high water supply. The percentage increase in plant biomass caused by growth at elevated compared with ambient ca was larger on average for the plants grown under water deficit than for the well-watered plants. This agrees with the results obtained for temperate woody plant seedlings and saplings, which also showed positive growth responses to elevated ca under water deficit (Tolley and Strain 1984; Arp et al. 1998; Centritto et al. 1999), although not in every case (Guehl et al. 1994; Duursma et al. 2011).
The size of the NSC pool in plants generally increases in response to elevated ca (Drake et al. 1997; Ainsworth and Long 2005). There has recently been debate about whether drought-induced tree mortality in the absence of biotic agents results from carbon starvation, hydraulic failure, impaired carbon translocation or a combination of these processes (McDowell et al. 2008; Sala et al. 2010; Anderegg et al. 2012). To the extent that drought-induced mortality can be delayed by having a larger reserve of NSC, elevated ca should allow tropical woody plants to survive for longer periods under drought. On the other hand, this may not be the case in tropical trees with sunlit canopies if NSC storage is already high under present-day ca (Newell et al. 2002; Körner 2003; Würth et al. 2005). Thus it may be the heavily shaded individuals in the understorey that benefit most from higher NSC concentrations as ca rises (Würth et al. 1998a; Lloyd and Farquhar 2008).
The interaction between potentially increasing drought frequency and intensity in tropical forests and potentially increasing ability to withstand drought under elevated ca will play a critical role in defining the overall response of carbon cycling in these ecosystems as ca rises. Increased WUE was one of the most consistently observed responses to elevated ca in potted tropical tree seedlings (Ziska et al. 1991; Eamus et al. 1993; Winter et al. 2001a; Holtum and Winter 2010; Cernusak et al. 2011b). Experiments are now required to build upon these initial results, involving woody tropical forest plants growing in their native soil environments and exposed to either naturally occurring or experimentally imposed droughts in combination with elevated ca.
Effects of elevated ca on species composition
Much uncertainty surrounding the functioning of tropical forests under elevated ca derives from potential shifts in forest composition. Although it is currently unknown how changes in species regeneration success under elevated ca will alter the future carbon cycling of tropical forests, the potential effects may be large. For example, spatial variation in species composition within the single forest type of the 50-ha plot on Barro Colorado Island in Panama was associated with variation in standing dry biomass ranging from 180 Mg ha–1 to 440 Mg ha–1 (Chave et al. 2003). In a further analysis of compositional effects on ecosystem function, Bunker et al. (2005) showed that a range of potential extinction scenarios influencing tree species with different functional traits could result in declines in carbon storage of up to 70% on Barro Colorado Island. Clearly, the importance of ca effects on species regeneration success should be recognised and investigated as a critical driver of future carbon cycling.
The largest impact on forest composition and associated carbon storage will arise if forest disturbance regimes are altered. In mature tropical forests, typical adult tree mortality rates are 1–2% of trees dying per year (Lewis et al. 2004), with the resulting disturbance to the forest canopy ranging from single treefall gaps to large canopy openings. Tropical forest plant species lie along a continuum from extremely shade-tolerant to absolutely light-demanding (Wright et al. 2005). The most shade-tolerant recruit and survive everywhere. The most light-demanding species will only recruit in large forest openings. Whereas small forest disturbances often result in the replacement of canopy trees by slow-growing shade-tolerant juveniles characterised by high wood density and large adult stature, larger disturbances resulting from multiple tree falls favour the initial recruitment of fast-growing, light-demanding pioneer species, generally characterised by a low wood density and a short lifespan (Swaine and Whitmore 1988).
Elevated ca may influence forest disturbance regimes in three ways. First, elevated ca may cause forest turnover rates to increase if there is an increase in competition caused by higher resource availability (i.e. CO2) (Lewis et al. 2009a; Bugmann and Bigler 2011). Second, increases in surface temperature are expected to result in stronger convectional storms, such as those that propagate across the Amazon basin (Nelson et al. 1994; Garstang et al. 1998; Knutson et al. 2010). Even though these storms are rare events (Gloor et al. 2009; Lloyd et al. 2009), they can produce canopy blow-downs extending over hundreds to thousands of hectares, resulting in large patches of early successional vegetation (Negron-Juarez et al. 2010). Third, more severe or frequent climate anomalies (Timmermann et al. 1999; Neelin et al. 2006) can result in biome-wide increases in adult tree mortality. These were observed in the Amazon basin following severe drought in 1997 (Williamson et al. 2000) and in 2005 (Phillips et al. 2009), and are inferred to have also occurred in 2010 (Lewis et al. 2011). Depending on the spatial distribution of tree mortality within the stand, these events have the potential to promote the widespread recruitment of pioneer species.
Elevated ca may also promote the regeneration of pioneer species in the absence of changes in forest disturbance by influencing the competitive balance between early and late successional species in gaps. For small-seeded pioneers, a critical filter to recruitment success is survival through the early establishment phase (Dalling and Hubbell 2002). Germinating seeds and emerging seedlings are particularly susceptible to drought-induced mortality during short dry spells (Engelbrecht et al. 2006; Daws et al. 2008), and small seedlings can be smothered by falling litter (Dalling and Hubbell 2002). Elevated ca may promote seedling establishment if it accelerates seedling growth and effectively shortens this vulnerable establishment period or ameliorates the effects of short-term drought by increasing WUE.
Seedlings establishing in newly formed gaps also face competition from pre-existing recruits of shade-tolerant species (advance regeneration) and potentially from adult trees that surround the gap and contribute to the lateral in-filling of the canopy. If seedling growth rates are not constrained by nutrient availability, the higher maximal assimilation rates of pioneers relative to shade-tolerant species should translate to a greater growth stimulation and competitive advantage (Oberbauer et al. 1985). This may be amplified by greater stimulation of photosynthesis during sunflecks, which dominate understorey light environments (Leakey et al. 2002). Conversely, elevated ca may intensify competition between pioneer recruits and the advance regeneration. Elevated ca has been shown to significantly enhance the growth of shade-tolerant seedlings under very low light conditions (Würth et al. 1998a) and may be expected to also enhance seedling survival. Growth enhancements are also likely to be strong for shade-tolerant liana species that await gap formation to recruit to the canopy (Körner 2009).
Lianas appear to be increasing in tropical forests over recent decades (Phillips et al. 2002; Wright et al. 2004; Schnitzer and Bongers 2011). This change in the functional composition of tropical forests may be the result of rising ca (Lewis et al. 2009a; Schnitzer and Bongers 2011) or other processes such as shifting dynamics of seed dispersal caused by hunting that favours predominantly wind-dispersed lianas over predominantly animal-dispersed trees (Wright et al. 2007). Both liana and tree seedlings have been shown to profit from elevated CO2 when grown in deep shade (Körner 2009). Beneath the canopy, lianas are likely to exhibit better light foraging per unit of carbon gained than tree saplings, due to their flexible growth strategy. This provides a hypothesised mechanism by which lianas could benefit more from elevated ca than trees (Körner 2009). Should lianas become more vigorous due to a functional type-specific benefit from elevated ca, this would have far-reaching consequences for carbon storage (Phillips et al. 2002). Liana infestations increase tree mortality and suppress tree growth, whereas lianas themselves allocate relatively little biomass to wood.
Woody legumes are both abundant and diverse in tropical forests, especially in the Neotropics and Africa (Gentry 1988; Losos and Leigh 2004; ter Steege et al. 2006). Some of these leguminous tree and liana species have the ability to form bacterial nodules on their roots that can fix atmospheric N2 (de Souza Moreira et al. 1992; Sprent 2009). Such species may be able to respond more strongly to elevated ca than nonfixing species, especially in nitrogen-poor soils (Thomas et al. 1991; Tissue et al. 1997; Cernusak et al. 2011b). An ability to acquire nitrogen from the atmosphere may also provide an advantage for phosphorus acquisition by promoting production of nitrogen-rich phosphatase enzymes in roots (Houlton et al. 2008). Such enzymes can be bound to root surfaces or released into the rhizosphere to hydrolyse organically-bound phosphorus, making it available for plant uptake (Richardson et al. 2005; Turner 2008). On the other hand, a pan-tropical increase in nitrogen deposition (Chen et al. 2010; Hietz et al. 2011) might limit the relative advantage of legumes over plant species that are incapable of N2 fixation. In an in situ experiment in a tropical forest understorey, a nodulated legume, Tachigali versicolour Standl. & L.O.Williams, had a growth response to elevated ca similar to that of nonlegumes (Würth et al. 1998a). However, demand for nitrogen is low in plants in deep shade and the benefit of N2 fixation may therefore be greatest in a gap environment (McHargue 1999; Barron et al. 2011). Future experiments on the responses of nodulated legumes to elevated ca should consider interactions with irradiance.
Increased carbon allocation to reproduction was observed for temperate trees exposed to FACE (Ladeau and Clark 2006; Way et al. 2010). Increased reproductive effort under elevated ca in tropical forests could have important consequences for long-term population dynamics. In addition, increased carbon allocation to short-lived tissues, such as flowers and fruits, would not lead to the same increase in carbon storage as increased allocation to wood. A significant increase in flower production has been observed over 18 years in a tropical forest in Panama (Wright and Calderon 2006). If this pattern is related to increasing ca, this may portend further increases in carbon allocation to reproduction as ca continues to rise.
The Late Paleocene–Eocene Thermal Maximum (PETM) was a global warming event that occurred ~56 million years ago, in which the global mean temperature rapidly increased by ~5°C in ~10 000 years (Zachos et al. 2003). This global warming event was associated with a large injection of greenhouse gases into the atmosphere and a rise in ca to ~1000 ppm. The PETM may provide a historic analogue for anthropogenic climate change, although the latter is occurring at a much faster rate. Pollen assemblages in three stratigraphic sections in eastern Colombia and western Venezuela demonstrated an increase in the diversity of tropical woody plants in response to the PETM (Jaramillo et al. 2010). Importantly, the PETM was associated with an intensification of the hydrological cycle, which probably resulted in either increased precipitation in the tropics or at least no increase in aridity (Jaramillo et al. 2010; Clementz and Sewall 2011). It is not known to what extent similar conditions will prevail as anthropogenic climate change unfolds. Nevertheless, the diversification of tropical woody plants in response to the PETM provides an extremely valuable insight into potential responses of tropical forests to elevated ca and climate change.
Currently, our ability to predict changes in the species composition of tropical forests under elevated ca is poor for at least four reasons: (1) temperate ecosystems have been the focus of most of research effort to date; (2) elevated ca field studies have focussed on even-aged stands where gap dynamics and gap regeneration were absent; (3) many studies of seedlings have featured plants that were isolated from ecological interactions or grown with a disturbed soil–microbe–plant complex; and (4) the traits that drive variation in response to elevated ca are not well understood in general, particularly in tropical species (Leakey et al. 2012; Leakey and Lau 2012).
Effects of elevated ca on ecosystem carbon storage
Forests have been a significant carbon sink in recent decades, both globally and in the tropics (Lewis et al. 2009b; Pan et al. 2011). Although it is likely that rising ca has played at least some role in driving the increase in tropical forest biomass (Lewis et al. 2009a), several other mechanisms may also have contributed. These possibilities include secondary succession on abandoned agricultural lands, recovery from other anthropogenic uses, including timber and fire wood extraction, and increased deposition of limiting nutrients resulting from anthropogenic activities.
Of the carbon stored in tropical forests, more than half is in live biomass (Pan et al. 2011). Repeated censuses of forest inventory plots over time have indicated that live biomass is increasing in some old-growth tropical forests. The average annualised change in aboveground biomass (AGB) in 79 plots in African tropical forests was about +0.6 Mg C ha–1 per year during the period 1968–2007 (Lewis et al. 2009b). Similarly, the average AGB change across 59 Amazonian plots was about +0.6 Mg C ha–1 per year from the 1980s until the early 2000s (Baker et al. 2004). The global terrestrial carbon sink of ~2 Pg C per year for the 1990s (Solomon et al. 2007) implies an increase in ecosystem carbon storage of ~0.2 Mg C ha–1 per year, if it were spread evenly over the global vegetated land surface. Körner (2009) suggested one could optimistically allow for three times more missing carbon to be located in tropical forests than in extratropical regions. Thus, for a first approximation, the estimated increase in AGB of 0.6 Mg C ha–1 per year is seemingly consistent with the carbon balance of the Earth system.
About one-third of carbon stored in tropical forests is in soil organic matter (Pan et al. 2011). The mean residence time for soil organic matter is similar to that for live biomass in tropical forests, on the order of 10–15 years (Malhi et al. 1999). This differs markedly from high latitude forests, where the mean residence time for soil organic matter can be 10 times longer than that for live biomass. Little is known about how elevated ca will affect soil carbon storage in tropical forests as ca rises. Increased NPP under elevated ca could accelerate the decomposition of soil organic matter through a priming effect, in which increased litter fall provides the carbon that fuels the microbial decomposition of soil organic matter (Sayer et al. 2011). However, like other responses of tropical forests to elevated CO2, soil priming effects appear to be regulated by nutrient availability (Nottingham et al. 2013). In general, small changes in soil carbon are difficult to detect, and even the longest manipulative experiments may not provide sufficient time for directional changes to be detected.
FACE experiments conducted with temperate forest trees provide an important source of information to draw upon when considering the likely responses of ecosystem carbon storage in tropical forests in response to rising ca. In experiments conducted in young forest stands, there was either a sustained increase in AGB through the full course of the experiment (McCarthy et al. 2010); or there was a transitory increase in AGB, with the increase restricted to the first few years of the experiment, and an increased allocation to fine root production and increased carbon in the soil (Norby et al. 2010). In a 100-year-old deciduous forest 35 m tall, there was no increase in basal area increment during 8 years of FACE (Körner et al. 2005). These experiments demonstrate no consistent evidence for sustained increases in temperate ecosystem carbon storage under elevated ca (Norby and Zak 2011).
Increased NPP caused by elevated ca could potentially be associated with less carbon storage in tropical forests if species composition shifts in favour of lianas and shorter-lived, faster growing tree species with lower wood density (Phillips et al. 2002; Körner 2004; Laurance et al. 2004; Körner 2009). The current distribution of AGB and NPP across Amazonia indicates a negative relationship between the two, such that more productive forests tend be of a lower biomass (Malhi et al. 2006; Saatchi et al. 2011). Higher mortality rates and lower wood density both contribute to the lower AGB in the Amazonian sites with higher NPP (Malhi et al. 2006). Liana infestations increase tree mortality (Putz 1984; Ingwell et al. 2010), especially of late successional tree species (Schnitzer and Bongers 2011), which have high wood density compared with early successional tree species (Wright et al. 2010). Thus, if liana abundance continues to increase and tropical forests continue to become more dynamic under elevated ca, they may store less carbon as a result. Interestingly, Amazonian forest plots appear to be increasing in dynamism while increasing in AGB (Baker et al. 2004; Lewis et al. 2004; Phillips et al. 2004). Continued monitoring is required to determine whether this is a transient response that will eventually give way to declining AGB following a shift towards a more gap-dominated structure (Malhi 2012).
The future of tropical forest carbon storage under climate change remains a large source of uncertainty in global climate simulations. For the Amazon Basin, uncertainty in future carbon storage results both from variability in climate projections (Salazar et al. 2007; Poulter et al. 2010b) and from uncertainty associated with direct effects of elevated ca on the productivity and WUE of tropical forests (Lapola et al. 2009; Rammig et al. 2010). In simulations with the direct effects of increasing ca on plant physiological processes turned off, both rising temperature and precipitation reduction caused declines in Amazonian forest biomass. However, with direct effects of elevated ca on plant physiology turned on, rising ca mitigated much of the climate-driven decline in forest biomass (Galbraith et al. 2010). These modelling studies highlight the critical role that experimental research can play in reducing the uncertainty associated with the direct effects of elevated ca on the physiology of woody tropical forest plants.
Challenges and opportunities for tropical CO2 enrichment experiments
FACE studies were helpful for extending observations in temperate forests to the forest stand scale, but compromises in experimental design were necessary because the systems were expensive to construct and operate (Norby and Zak 2011). For example, although there is much interest in understanding the carbon cycling responses of intact mature forests, most of the FACE experiments were conducted in young monoculture plantations. An example is shown in Fig. 5a, b. The one exception, in which mature, temperate forest trees were exposed to elevated ca (Körner et al. 2005), necessarily required a different compromise: a focus on individual trees rather than the whole ecosystem (Fig. 5c, d). Tropical forests contain large trees and are species-rich. In studies of forest dynamics, representative plots consequently tend to be 1 ha or larger to adequately capture the abundance of co-occurring species. Given the greater stature, diversity and complexity of tropical forests, as well as many infrastructure constraints (e.g. roads, power, CO2 supply), a FACE experiment will be even more challenging to implement in a tropical forest. This may require the development of new approaches, different from those deployed in previous experiments in temperate forests. Despite these challenges, investigating stand-level responses to elevated ca in tropical forests must remain a long-term goal, as this is the scale relevant to predicting global climate change feedback over the coming century.

|
Field experiments to explore how elevated ca will affect establishment of early and late successional tree species could provide a tractable alternative in the near term, because compositional shifts in tropical forest canopies are likely to be driven by changes in seedling establishment success. Because FACE systems require wind to disperse CO2 and because wind velocities are often low in forest gaps, CO2 enrichment systems with forced ventilation, such as simple open-top chambers, would be a suitable tool for the study of gap dynamics, particularly in small treefall gaps. Any such experiment should pay careful attention to soil conditions in order to ensure undisturbed, intact plant–soil interactions. We note that the air within 1–2 m of the forest floor in tropical forests tends to be naturally enriched in CO2 compared with air above the canopy. However, during the day, when photosynthesis is possible, this natural CO2 enrichment rarely exceeds 50 ppm (Lloyd et al. 1996; Buchmann et al. 1997; Würth et al. 1998a; Holtum and Winter 2001).
Liana responses to elevated ca in the understorey could be successfully studied using open-top chambers or similar systems for CO2 augmentation. This type of experiment would further lend itself to differential tests across other plant functional types, for example, comparisons of N2-fixing tree and liana species with nonfixing species. Multifactor experiments under elevated ca could also be implemented in the understorey, including variable intensity of drought, irradiance or both. Interactions with irradiance may be particularly important, because elevated ca can lower the light compensation point in deep shade, with potentially large effects on plant carbon balance under extreme photon shortage.
Tree and liana branches in tropical forest canopies can be exposed to elevated ca with low-cost installations (Körner and Würth 1996; Würth et al. 1998b; Lovelock et al. 1999) and accessed for physiological measurements using an existing network of tropical forest canopy cranes (Basset et al. 2003). In addition, individual leaves or branches can be warmed in situ in order to examine responses to both elevated ca and elevated temperature. Liana responses to elevated ca along their growth trajectories from the forest floor to the canopy could be successfully studied in this way (Zotz et al. 2006). Experiments exposing only a part of the plant to elevated ca should focus on short-term responses, as interactions at the whole-plant scale will not be present in such experiments. In addition, the degree to which branches behave autonomously in their carbon relations with other parts of the plant could influence physiological responses, and this should be taken into account in experimental design and interpretation of the results (Sprugel et al. 1991; Lovelock et al. 1999).
The study of forest segments containing tall trees should be feasible in closed systems, a possibility that has been trialled but not extensively exploited in replicated experiments (Osmond et al. 2004). Examples of modern, large, naturally-lit enclosures that would accommodate tall forest trees include highly transparent ethylene tetrafluoroethylene covered domes (Fig. 5e, f) such as those in the Eden Project (Eden Project 2013). Closed systems would require temperature control, but use relatively little CO2 for augmentation compared with FACE systems. Moreover, plants can be maintained at above or below current ambient CO2 conditions. Temperature can also be manipulated. Unlike open CO2 enrichment systems in which CO2 concentrations exhibit pronounced short-term fluctuations around the target concentration (Holtum and Winter 2003; Bunce 2012), relatively stable CO2 concentrations can be maintained in enclosed systems. Although enclosed systems have disadvantages in terms of artificial rain and require sustained air mixing, this technology opens exciting possibilities for experimental ecosystem science, including the measurement of ecosystem gas exchange (Osmond et al. 2004), either by placing premanufactured enclosures over existing tropical vegetation or by establishing tree stands in purpose-built enclosures.
Belowground responses, including root deployment and function, nutrient turnover and soil organic matter processing, are especially important to the integrated ecosystem response to elevated CO2. Belowground processes are difficult to measure and are still poorly quantified. For example, because of the past focus on temperate forests, in which nitrogen is believed to be the major limiting nutrient, there has been little research on the effects of elevated ca on phosphorus cycling, and phosphorus cycling is poorly represented in models. Experimental approaches and site selection must carefully consider the importance of the belowground environment by avoiding artefacts associated with soil disturbance and include the capability for sufficient belowground measurements.
Many of the critical questions about the role of tropical forests in global carbon cycling are inherently long-term questions (e.g. 50–100 years). Furthermore, no experiment or small set of experiments can ever represent the full diversity of the tropical biome. Models that are well informed by experimental observations offer an opportunity to extrapolate through space and time. Hence an important strategy for designing experiments that will provide the most useful and needed data and the greatest understanding of processes is to engage a modelling perspective from the start.
Opportunities for reducing uncertainty in models
The current generation of ecosystem models demonstrates the potential for elevated ca to mitigate much of the climate-driven loss of tropical forest biomass that might otherwise occur by the year 2100 (Lapola et al. 2009; Galbraith et al. 2010; Poulter et al. 2010a; Rammig et al. 2010). However, model predictions are currently based on very limited information and omit what are likely to be critical modifying processes. Uncertainties in the representation of elevated ca effects on tropical forest vegetation include the potential for elevated ca to relieve the limitations on canopy photosynthesis caused by high-temperature stress, nutrient limitations on NPP responses to elevated ca, the effects of elevated ca on drought induced tree mortality and the effects of elevated ca on species composition. The results of existing ecosystem models represent testable hypotheses that can guide experimental design, and understanding the critical points of uncertainty in the models with regard to representation of elevated ca responses can help to identify the highest priority research needs.
Progress can be achieved in the representation of leaf and canopy responses to high temperature by comparing modelled response curves with those observed in leaf cuvettes and by eddy covariance (Lloyd et al. 1995; Doughty and Goulden 2008; Verbeeck et al. 2011). For example, observations in a tropical forest in Amazonia indicated that a 3°C rise in bulk air temperature above 28.5°C, caused by an increase in irradiance, resulted in a 40% reduction in whole-canopy gross gas exchange (Doughty and Goulden 2008). This was caused by increases in leaf temperatures of 5–8°C in the sunlit fraction of the canopy, and subsequent declines in gs and A in sunlit leaves. Fig. 6 shows an example of modelled response curves for canopy photosynthesis for two dynamic global vegetation models, Lund-Potsdam-Jena (LPJ) and Organizing Carbon and Hydrology in Dynamic Ecosystems (ORCHIDEE), which differ in their treatment of light diffusion through a canopy, gs, parameterisation of the Farquhar et al. (1980) photosynthesis model and soil–water balance (Sitch et al. 2003; Krinner et al. 2005). Although temperature and radiation responses differ between the two models, neither appears capable of capturing the pattern observed in the Amazonian forest, namely an almost linear decline in canopy photosynthesis between bulk air temperatures of 28.5 and 31.5°C. In order to extend the representation of temperature responses to elevated ca conditions, validation data from experimental manipulation experiments are essential.
|
Recent progress has been made towards including the effects of phosphorus availability in ecosystem models (Lloyd et al. 2001; Wang et al. 2007; Mercado et al. 2011; Goll et al. 2012). However, the representation of phosphorus dynamics in models is also limited by uncertainty about the extent to which tropical woody plants can access organic phosphorus, and the mobility of organic phosphorus compounds in tropical forest soils (Turner 2008; Cernusak et al. 2011c; Turner and Engelbrecht 2011). An improved understanding of phosphorus cycling in tropical forests is needed. Furthermore, the effects of phosphorus availability on the responsiveness of tropical vegetation to elevated ca have not been tested experimentally. Such experiments should be given a high priority.
Predicting drought-induced vegetation mortality remains a significant challenge (Nepstad et al. 2007; McDowell et al. 2011). It was recently shown that hydraulic impairment was a better predictor of drought-induced mortality than was the size of the NSC pool in a temperate deciduous tree species (Anderegg et al. 2012). Carbon starvation thus did not appear to be a useful predictor. However, if it is shown that the NSC pool plays a significant role in embolism repair (Sala et al. 2012), this could have implications for recovery from drought under elevated ca, because NSC concentrations are likely to increase. Further experimentation is required to develop a mechanistic model of mortality mechanisms and plant age structure that can account for the effects of elevated ca.
At the community scale, incorporating predictions regarding the effects of elevated ca on compositional change into ecosystem models represents a further challenge. If experimental manipulations of ca in tropical forests are limited to targeting juvenile stages, then models will need to upscale observed shifts in the recruitment success of plant functional types to the dynamics of entire forest assemblages. The development of models, such as the Ecosystem Demography model (Moorcroft et al. 2001; Medvigy et al. 2009; Fisher et al. 2010), which link mechanistic representations of ecophysiology and biogeochemistry to the size-structured competition and succession found in forest gap models will provide a potential tool to achieve this integration. In addition, a new approach to representing functional diversity in dynamic global vegetation models was recently developed, based on generating plant traits from distributions of growth strategies rather than from fixed traits (Pavlick et al. 2012). This approach could prove useful for simulating tropical forests, allowing for a more flexible representation of their structure and function through space and time. Model validation under ambient ca using existing long-term tropical forest dynamics data from large plots (Condit 1995) will be essential.
Conclusions
Model simulations suggest that tropical forest NPP will respond more strongly to elevated ca than that of temperate and boreal forests. This hypothesis could have significant implications for the global carbon cycle and for climate change predictions. It is sufficiently compelling to justify large-scale investment in experimental testing. Ultimately, we would like to know whether and to what extent elevated ca-induced increases in NPP in tropical forests will result in increased carbon storage and negative feedback to ca. Addressing this question will require a combination of experimental and model-based approaches. Additional supporting hypotheses that should also have a high priority for experimental research are the following: (1) elevated ca will increase the high temperature tolerance of photosynthesis in tropical tree leaves; (2) phosphorus availability will limit tropical forest NPP responses to elevated ca; (3) elevated ca will increase the drought resistance of tropical forests; and (4) elevated ca-induced changes in species composition will cause directional changes in carbon storage in tropical forests.
Carefully considered experiments that need not necessarily take place at the stand scale could make important contributions towards testing these latter hypotheses. Understorey experiments that quantify the potential for elevated ca to alter the regeneration success of species representing important functional groups, and account for interactions with soil nutrient and water status, would be achievable in the near term and cost-effective. Open-top chambers and branch bags installed in the canopy to elevate ca, in combination with in situ warming, could provide a useful method for answering questions about canopy leaf physiology in relation to temperature and drought. These experiments should not be viewed as replacements for stand-level experiments, but rather as tractable first steps. Stand-level CO2 enrichment experiments will provide invaluable results, and should be vigorously pursued. The critical role of tropical forests in the terrestrial carbon cycle and the paucity of experimental data so far available together should make tropical forest CO2 enrichment experiments a very high priority for global climate change research.
Acknowledgements
This review resulted from a symposium held at the Smithsonian Tropical Research Institute on 31 March and 1 April 2011. Funding for the symposium was provided by the Smithsonian Tropical Research Institute. LAC was supported by a Future Fellowship from the Australian Research Council (FT100100329).
References
Ainsworth EA, Long SP (2005) What have we learned from 15 years of free-air CO2 enrichment (FACE)? A meta-analytic review of the responses of photosynthesis, canopy properties and plant production to rising CO2. New Phytologist 165, 351–372.| What have we learned from 15 years of free-air CO2 enrichment (FACE)? A meta-analytic review of the responses of photosynthesis, canopy properties and plant production to rising CO2.Crossref | GoogleScholarGoogle Scholar | 15720649PubMed |
Ainsworth EA, Rogers A (2007) The response of photosynthesis and stomatal conductance to rising [CO2]: mechanisms and environmental interactions. Plant, Cell & Environment 30, 258–270.
| The response of photosynthesis and stomatal conductance to rising [CO2]: mechanisms and environmental interactions.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2sXjtlemu78%3D&md5=e7e789bd6ed2f1abf505f3139bacfcfdCAS |
Anderegg WRL, Berry JA, Smith DD, Sperry JS, Anderegg LDL, Field CB (2012) The roles of hydraulic and carbon stress in a widespread climate-induced forest die-off. Proceedings of the National Academy of Sciences of the United States of America 109, 233–237.
| The roles of hydraulic and carbon stress in a widespread climate-induced forest die-off.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC38XhsVehtrw%3D&md5=aefa22c82d5a186f8f5cf399b6c236d4CAS |
Arnone JA, Körner C (1995) Soil and biomass carbon pools in model communities of tropical plants under elevated CO2. Oecologia 104, 61–71.
| Soil and biomass carbon pools in model communities of tropical plants under elevated CO2.Crossref | GoogleScholarGoogle Scholar |
Arp WJ, Van Mierlo JEM, Berendse F, Snijders W (1998) Interactions between elevated CO2 concentration, nitrogen and water: effects on growth and water use of six perennial plant species. Plant, Cell & Environment 21, 1–11.
| Interactions between elevated CO2 concentration, nitrogen and water: effects on growth and water use of six perennial plant species.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaK1cXitlCgsLw%3D&md5=effdf774d8553c5bf39a63dc70d5d874CAS |
Atkin OK, Bruhn D, Hurry VM, Tjoelker MG (2005) The hot and the cold: unravelling the variable response of plant respiration to temperature. Functional Plant Biology 32, 87–105.
| The hot and the cold: unravelling the variable response of plant respiration to temperature.Crossref | GoogleScholarGoogle Scholar |
Bader MKF, Siegwolf R, Körner C (2010) Sustained enhancement of photosynthesis in mature deciduous forest trees after 8 years of free air CO2 enrichment. Planta 232, 1115–1125.
| Sustained enhancement of photosynthesis in mature deciduous forest trees after 8 years of free air CO2 enrichment.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3cXhtFKmur%2FP&md5=3f538196f9ef72829b67b608f9acc5c9CAS |
Baker TR, Phillips OL, Malhi Y, Almeida S, Arroyo L, Di Fiore A, Erwin T, Higuchi N, Killeen TJ, Laurance SG Lewis SL, Monteagudo A, Neill DA, Núñez Vargas P, Pitman NCA, Silva JNM, Vásquez Martinez R (2004) Increasing biomass in Amazonian forest plots. Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences 359, 353–365.
| Increasing biomass in Amazonian forest plots.Crossref | GoogleScholarGoogle Scholar | 15212090PubMed |
Barron AR, Würzburger N, Bellenger JP, Wright SJ, Kraepiel AML, Hedin LO (2009) Molybdenum limitation of asymbiotic nitrogen fixation in tropical forest soils. Nature Geoscience 2, 42–45.
| Molybdenum limitation of asymbiotic nitrogen fixation in tropical forest soils.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD1cXhsFartrnJ&md5=469f7c0ba568b4e52ac9ac4f9b9d812dCAS |
Barron AR, Purves DW, Hedin LO (2011) Facultative nitrogen fixation by canopy legumes in a lowland tropical forest. Oecologia 165, 511–520.
| Facultative nitrogen fixation by canopy legumes in a lowland tropical forest.Crossref | GoogleScholarGoogle Scholar | 21110206PubMed |
Barton CVM, Duursma RA, Medlyn BE, Ellsworth DS, Eamus D, Tissue DT, Adams MA, Conroy J, Crous KY, Liberloo M, Löw M, Linder S, McMurtrie RE (2012) Effects of elevated atmospheric CO2 on instantaneous transpiration efficiency at leaf and canopy scales in Eucalyptus saligna. Global Change Biology 18, 585–595.
| Effects of elevated atmospheric CO2 on instantaneous transpiration efficiency at leaf and canopy scales in Eucalyptus saligna.Crossref | GoogleScholarGoogle Scholar |
Basset Y, Horlyck V, Wright SJ (Eds) (2003) ‘Studying forest canopies from above: the international canopy crane network.’ (Smithsonian Tropical Research Institute, Panama and the United Nations Environmental Programme: Balboa)
Battipaglia G, Saurer M, Cherubini P, Calfapietra C, McCarthy HR, Norby RJ, Cotrufo MF (2013) Elevated CO2 increases tree-level intrinsic water use efficiency: insights from carbon and oxygen isotope analyses in tree rings across three forest FACE sites. New Phytologist 197, 544–554.
| Elevated CO2 increases tree-level intrinsic water use efficiency: insights from carbon and oxygen isotope analyses in tree rings across three forest FACE sites.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC38XhvVynsr3K&md5=43874dc99149b49d8df5e5bf905ff627CAS | 23215904PubMed |
Beer C, Reichstein M, Tomelleri E, Ciais P, Jung M, Carvalhais N, Rödenbeck C, Arain MA, Baldocchi D, Bonan GB, Bondeau A, Cescatti A, Lasslop G, Lindroth A, Lomas M, Luyssaert S, Margolis H, Oleson KW, Roupsard O, Veenendaal E, Viovy N, Williams C, Woodward FI, Papale D (2010) Terrestrial gross carbon dioxide uptake: global distribution and covariation with climate. Science 329, 834–838.
| Terrestrial gross carbon dioxide uptake: global distribution and covariation with climate.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3cXpvV2iu7k%3D&md5=29599db30c8bad24021f28b581515f1fCAS | 20603496PubMed |
Berry J, Björkman O (1980) Photosynthetic response and adaptation to temperature in higher plants. Annual Review of Plant Physiology and Plant Molecular Biology 31, 491–543.
| Photosynthetic response and adaptation to temperature in higher plants.Crossref | GoogleScholarGoogle Scholar |
Berryman CA, Eamus D, Duff GA (1993) The influence of CO2 enrichment on growth, nutrient content and biomass allocation of Maranthes corymbosa. Australian Journal of Botany 41, 195–209.
| The influence of CO2 enrichment on growth, nutrient content and biomass allocation of Maranthes corymbosa.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaK3sXlslOktbk%3D&md5=c7fdd07f7fd79570ebaf42b432651ac6CAS |
Berryman CA, Eamus D, Duff GA (1994) Stomatal responses to a range of variables in two tropical tree species grown with CO2 enrichment. Journal of Experimental Botany 45, 539–546.
| Stomatal responses to a range of variables in two tropical tree species grown with CO2 enrichment.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaK2cXlslGns7o%3D&md5=5c9afb5a886b9bc701e71ee570935f72CAS |
Betts RA, Cox PM, Collins M, Harris PP, Huntingford C, Jones CD (2004) The role of ecosystem–atmosphere interactions in simulated Amazonian precipitation decrease and forest dieback under global climate warming. Theoretical and Applied Climatology 78, 157–175.
| The role of ecosystem–atmosphere interactions in simulated Amazonian precipitation decrease and forest dieback under global climate warming.Crossref | GoogleScholarGoogle Scholar |
Bloom AJ, Rubio-Asensio JS, Randall L, Rachmilevitch S, Cousins AB, Carlisle EA (2012) CO2 enrichment inhibits shoot nitrate assimilation in C3 but not C4 plants and slows growth under nitrate in C3 plants. Ecology 93, 355–367.
| CO2 enrichment inhibits shoot nitrate assimilation in C3 but not C4 plants and slows growth under nitrate in C3 plants.Crossref | GoogleScholarGoogle Scholar | 22624317PubMed |
Bonal D, Ponton S, Le Thiec D, Richard B, Ningre N, Hérault B, Ogée J, Gonzalez S, Pignal M, Sabatier D, Guehl J-M (2011) Leaf functional response to increasing atmospheric CO2 concentrations over the last century in two northern Amazonian tree species: an historical δ13C and δ18O approach using herbarium samples. Plant, Cell & Environment 34, 1332–1344.
| Leaf functional response to increasing atmospheric CO2 concentrations over the last century in two northern Amazonian tree species: an historical δ13C and δ18O approach using herbarium samples.Crossref | GoogleScholarGoogle Scholar |
Boyer JS (1968) Relationship of water potential to growth of leaves. Plant Physiology 43, 1056–1062.
| Relationship of water potential to growth of leaves.Crossref | GoogleScholarGoogle Scholar | 1:STN:280:DC%2BC3cngvFSltw%3D%3D&md5=6bface797829489e24506e0a12846251CAS | 16656882PubMed |
Brando PM, Nepstad DC, Davidson EA, Trumbore SE, Ray D, Camargo P (2008) Drought effects on litterfall, wood production and belowground carbon cycling in an Amazon forest: results of a throughfall reduction experiment. Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences 363, 1839–1848.
| Drought effects on litterfall, wood production and belowground carbon cycling in an Amazon forest: results of a throughfall reduction experiment.Crossref | GoogleScholarGoogle Scholar | 18267902PubMed |
Brienen RJW, Wanek W, Hietz P (2011) Stable carbon isotopes in tree rings indicate improved water use efficiency and drought responses of a tropical dry forest tree species. Trees 25, 103–113.
| Stable carbon isotopes in tree rings indicate improved water use efficiency and drought responses of a tropical dry forest tree species.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3MXmsVWrtA%3D%3D&md5=cd2478cf8c5a7317077013521f9cafebCAS |
Brookshire ENJ, Gerber S, Menge DNL, Hedin LO (2012) Large losses of inorganic nitrogen from tropical rainforests suggest a lack of nitrogen limitation. Ecology Letters 15, 9–16.
| Large losses of inorganic nitrogen from tropical rainforests suggest a lack of nitrogen limitation.Crossref | GoogleScholarGoogle Scholar | 1:STN:280:DC%2BC38%2FjslGrsw%3D%3D&md5=2c3db3d14b6e773cb23799d701094e49CAS |
Buchmann N, Guehl JM, Barigah TS, Ehleringer JR (1997) Interseasonal comparison of CO2 concentrations, isotopic composition, and carbon dynamics in an Amazonian rainforest (French Guiana). Oecologia 110, 120–131.
| Interseasonal comparison of CO2 concentrations, isotopic composition, and carbon dynamics in an Amazonian rainforest (French Guiana).Crossref | GoogleScholarGoogle Scholar |
Buckley TN (2008) The role of stomatal acclimation in modelling tree adaptation to high CO2. Journal of Experimental Botany 59, 1951–1961.
| The role of stomatal acclimation in modelling tree adaptation to high CO2.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD1cXmtleltLk%3D&md5=0e58d52cd78d97a5c9eda63332624a41CAS | 18000018PubMed |
Bugmann H, Bigler C (2011) Will the CO2 fertilization effect in forests be offset by reduced tree longevity? Oecologia 165, 533–544.
| Will the CO2 fertilization effect in forests be offset by reduced tree longevity?Crossref | GoogleScholarGoogle Scholar | 21104278PubMed |
Bunce JA (2012) Responses of cotton and wheat photosynthesis and growth to cyclic variation in carbon dioxide concentration. Photosynthetica 50, 395–400.
| Responses of cotton and wheat photosynthesis and growth to cyclic variation in carbon dioxide concentration.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC38XhtFKis7vO&md5=4be8cd776ee6ab2a0fa6a2310cd2a369CAS |
Bunker DE, deClerck F, Bradford JC, Colwell RK, Perfecto I, Phillips OL, Sankaran M, Naeem S (2005) Species loss and aboveground carbon storage in a tropical forest. Science 310, 1029–1031.
| Species loss and aboveground carbon storage in a tropical forest.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2MXhtF2is7nL&md5=b8141b46f683deb60830a8c29aa8628dCAS | 16239439PubMed |
Campbell GS, Norman JM (1998) ‘An introduction to environmental biophysics.’ (Springer-Verlag: New York)
Carswell FE, Grace J, Lucas ME, Jarvis PG (2000) Interaction of nutrient limitation and elevated CO2 concentration on carbon assimilation of a tropical tree seedling (Cedrela odorata). Tree Physiology 20, 977–986.
| Interaction of nutrient limitation and elevated CO2 concentration on carbon assimilation of a tropical tree seedling (Cedrela odorata).Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD3cXlvVantL8%3D&md5=ee200fb0aad7d09e2b5d795230436d66CAS | 11303573PubMed |
Cavaleri MA, Oberbauer SF, Ryan MG (2008) Foliar and ecosystem respiration in an old-growth tropical rain forest. Plant, Cell & Environment 31, 473–483.
| Foliar and ecosystem respiration in an old-growth tropical rain forest.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD1cXlt1CksrY%3D&md5=3d962ddd4afd301a2a5bab969f15db48CAS |
Centritto M, Lee HSJ, Jarvis PG (1999) Interactive effects of elevated CO2 and drought on cherry (Prunus avium) seedlings I. Growth, whole-plant water use efficiency and water loss. New Phytologist 141, 129–140.
| Interactive effects of elevated CO2 and drought on cherry (Prunus avium) seedlings I. Growth, whole-plant water use efficiency and water loss.Crossref | GoogleScholarGoogle Scholar |
Cernusak LA, Marshall JD (2001) Responses of foliar δ13C, gas exchange, and leaf morphology to reduced hydraulic conductivity in Pinus monticola branches. Tree Physiology 21, 1215–1222.
| Responses of foliar δ13C, gas exchange, and leaf morphology to reduced hydraulic conductivity in Pinus monticola branches.Crossref | GoogleScholarGoogle Scholar | 1:STN:280:DC%2BD3MrlsVaqtg%3D%3D&md5=02ba376dbb91e5f5f96a34a9cb3b0cebCAS | 11600343PubMed |
Cernusak LA, Hutley LB, Beringer J, Holtum JAM, Turner BL (2011a) Photosynthetic physiology of eucalypts along a sub-continental rainfall gradient in northern Australia. Agricultural and Forest Meteorology 151, 1462–1470.
| Photosynthetic physiology of eucalypts along a sub-continental rainfall gradient in northern Australia.Crossref | GoogleScholarGoogle Scholar |
Cernusak LA, Winter K, Martinez C, Correa E, Aranda J, Garcia M, Jaramillo C, Turner BL (2011b) Responses of legume versus nonlegume tropical tree seedlings to elevated CO2 concentration. Plant Physiology 157, 372–385.
| Responses of legume versus nonlegume tropical tree seedlings to elevated CO2 concentration.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3MXht1Sit7zF&md5=fddaae83ba34e2d51d974bfa698f83c6CAS | 21788363PubMed |
Cernusak LA, Winter K, Turner BL (2011c) Transpiration modulates phosphorus acquisition in tropical tree seedlings. Tree Physiology 31, 878–885.
| Transpiration modulates phosphorus acquisition in tropical tree seedlings.Crossref | GoogleScholarGoogle Scholar | 21856654PubMed |
Chambers JQ, Silver WL (2004) Some aspects of ecophysiological and biogeochemical responses of tropical forests to atmospheric change. Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences 359, 463–476.
| Some aspects of ecophysiological and biogeochemical responses of tropical forests to atmospheric change.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2cXls1Sjs7w%3D&md5=2033bc1c8856ea1df648c3c95493231dCAS | 15212096PubMed |
Chave J, Condit R, Lao S, Caspersen JP, Foster RB, Hubbell SP (2003) Spatial and temporal variation of biomass in a tropical forest: results from a large census plot in Panama. Journal of Ecology 91, 240–252.
| Spatial and temporal variation of biomass in a tropical forest: results from a large census plot in Panama.Crossref | GoogleScholarGoogle Scholar |
Chaves MM, Maroco JP, Pereira JS (2003) Understanding plant responses to drought – from genes to the whole plant. Functional Plant Biology 30, 239–264.
| Understanding plant responses to drought – from genes to the whole plant.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD3sXjtVKlt7o%3D&md5=17b4b78fac11852745aa8deb5537afc3CAS |
Chen Y, Randerson JT, van der Werf GR, Morton DC, Mu MQ, Kasibhatla PS (2010) Nitrogen deposition in tropical forests from savanna and deforestation fires. Global Change Biology 16, 2024–2038.
| Nitrogen deposition in tropical forests from savanna and deforestation fires.Crossref | GoogleScholarGoogle Scholar |
Clark DB, Clark DA, Oberbauer SF (2010) Annual wood production in a tropical rain forest in NE Costa Rica linked to climatic variation but not to increasing CO2. Global Change Biology 16, 747–759.
| Annual wood production in a tropical rain forest in NE Costa Rica linked to climatic variation but not to increasing CO2.Crossref | GoogleScholarGoogle Scholar |
Clementz MT, Sewall JO (2011) Latitudinal gradients in greenhouse seawater δ18O: evidence from Eocene sirenian tooth enamel. Science 332, 455–458.
| Latitudinal gradients in greenhouse seawater δ18O: evidence from Eocene sirenian tooth enamel.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3MXkvValsr8%3D&md5=5fc4919d27e2d36803157b20384c47f0CAS | 21512030PubMed |
Collatz GJ, Ball JT, Grivet C, Berry JA (1991) Physiological and environmental regulation of stomatal conductance, photosynthesis and transpiration: a model that includes a laminar boundary layer. Agricultural and Forest Meteorology 54, 107–136.
| Physiological and environmental regulation of stomatal conductance, photosynthesis and transpiration: a model that includes a laminar boundary layer.Crossref | GoogleScholarGoogle Scholar |
Condit R (1995) Research in large, long-term tropical forest plots. Trends in Ecology & Evolution 10, 18–22.
| Research in large, long-term tropical forest plots.Crossref | GoogleScholarGoogle Scholar | 1:STN:280:DC%2BC3M7itFWltw%3D%3D&md5=b2d258a6c3244cb70f1923dea6260709CAS |
Condit R, Hubbell SP, Foster RB (1995) Mortality rates of 205 neotropical tree and shrub species and the impact of a severe drought. Ecological Monographs 65, 419–439.
| Mortality rates of 205 neotropical tree and shrub species and the impact of a severe drought.Crossref | GoogleScholarGoogle Scholar |
da Costa ACL, Galbraith D, Almeida S, Portela BTT, da Costa M, de Athaydes Silva Junior J, Braga AP, de Gonçalves PHL, de Oliveira AAR, Fisher R, Phillips OL, Metcalfe DB, Levy P, Meir P (2010) Effect of 7 yr of experimental drought on vegetation dynamics and biomass storage of an eastern Amazonian rainforest. New Phytologist 187, 579–591.
| Effect of 7 yr of experimental drought on vegetation dynamics and biomass storage of an eastern Amazonian rainforest.Crossref | GoogleScholarGoogle Scholar |
Dalling JW, Hubbell SP (2002) Seed size, growth rate and gap microsite conditions as determinants of recruitment success for pioneer species. Journal of Ecology 90, 557–568.
| Seed size, growth rate and gap microsite conditions as determinants of recruitment success for pioneer species.Crossref | GoogleScholarGoogle Scholar |
Davidson EA, de Carvalho CJR, Figueira AM, Ishida FY, Ometto JPHB, Nardoto GB, Sabá RT, Hayashi SN, Leal EC, Vieira ICG, Martinelli LA (2007) Recuperation of nitrogen cycling in Amazonian forests following agricultural abandonment. Nature 447, 995–998.
| Recuperation of nitrogen cycling in Amazonian forests following agricultural abandonment.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2sXms12jsr0%3D&md5=640420f51b41c6db682a714b1d3ebf75CAS | 17581583PubMed |
Daws MI, Crabtree LM, Dalling JW, Mullins CE, Burslem D (2008) Germination responses to water potential in neotropical pioneers suggest large-seeded species take more risks. Annals of Botany 102, 945–951.
| Germination responses to water potential in neotropical pioneers suggest large-seeded species take more risks.Crossref | GoogleScholarGoogle Scholar | 18840874PubMed |
de Boer HJ, Lammertsma EI, Wagner-Cremer F, Dilcher DL, Wassen MJ, Dekker SC (2011) Climate forcing due to optimization of maximal leaf conductance in subtropical vegetation under rising CO2. Proceedings of the National Academy of Sciences of the United States of America 108, 4041–4046.
| Climate forcing due to optimization of maximal leaf conductance in subtropical vegetation under rising CO2.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3MXjs1WqtLs%3D&md5=b4196dd39c6f27a401a7e92ff19c41f5CAS | 21330553PubMed |
de Oliveira EAD, Approbato AU, Legracie JR, Martinez CA (2012) Soil-nutrient availability modifies the response of young pioneer and late successional trees to elevated carbon dioxide in a Brazilian tropical environment. Environmental and Experimental Botany 77, 53–62.
| Soil-nutrient availability modifies the response of young pioneer and late successional trees to elevated carbon dioxide in a Brazilian tropical environment.Crossref | GoogleScholarGoogle Scholar |
de Souza Moreira FM, da Silva MF, de Faria SM (1992) Occurence of nodulation in legume species in the Amazon region of Brazil. New Phytologist 121, 563–570.
| Occurence of nodulation in legume species in the Amazon region of Brazil.Crossref | GoogleScholarGoogle Scholar |
Doughty CE (2011) An in situ leaf and branch warming experiment in the Amazon. Biotropica 43, 658–665.
| An in situ leaf and branch warming experiment in the Amazon.Crossref | GoogleScholarGoogle Scholar |
Doughty CE, Goulden ML (2008) Are tropical forests near a high temperature threshold? Journal of Geophysical Research 113, –G00B07.
| Are tropical forests near a high temperature threshold?Crossref | GoogleScholarGoogle Scholar |
Drake BG, Gonzàlez-Meler MA, Long SP (1997) More efficient plants: a consequence of rising atmospheric CO2? Annual Review of Plant Physiology and Plant Molecular Biology 48, 609–639.
| More efficient plants: a consequence of rising atmospheric CO2?Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaK2sXjs1eltbY%3D&md5=050ad0f7b4f2a39c914080ee055d75b9CAS | 15012276PubMed |
Duursma RA, Barton CVM, Eamus D, Medlyn BE, Ellsworth DS, Forster MA, Tissue DT, Linder S, McMurtrie RE (2011) Rooting depth explains CO2 × drought interaction in Eucalyptus saligna. Tree Physiology 31, 922–931.
| Rooting depth explains CO2 × drought interaction in Eucalyptus saligna.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3MXhsVCls7vL&md5=d159626ab68d83a69f937814fdffd25cCAS | 21571724PubMed |
Eamus D (1991) The interaction of rising CO2 and temperatures with water use efficiency. Plant, Cell & Environment 14, 843–852.
| The interaction of rising CO2 and temperatures with water use efficiency.Crossref | GoogleScholarGoogle Scholar |
Eamus D, Berryman CA, Duff GA (1993) Assimilation, stomatal conductance, specific leaf area and chlorophyll responses to elevated CO2 of Maranthes corymbosa, a tropical monsoon rain forest species. Australian Journal of Plant Physiology 20, 741–755.
| Assimilation, stomatal conductance, specific leaf area and chlorophyll responses to elevated CO2 of Maranthes corymbosa, a tropical monsoon rain forest species.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaK2cXitFart7o%3D&md5=47d51dd860f009f3bb4726fb20c486d2CAS |
Eamus D, Berryman CA, Duff GA (1995) The impact of CO2 enrichment on water relations in Maranthes corymbosa and Eucalyptus tetrodonta. Australian Journal of Botany 43, 273–282.
| The impact of CO2 enrichment on water relations in Maranthes corymbosa and Eucalyptus tetrodonta.Crossref | GoogleScholarGoogle Scholar |
Eden Project 2013 Eden Project home page. ( Eden Project: Bodelva UK ) Available online at: www.edenproject.com [Verified 8 April 2013]
Engelbrecht BMJ, Dalling JW, Pearson TRH, Wolf RL, Galvez DA, Koehler T, Tyree MT, Kursar TA (2006) Short dry spells in the wet season increase mortality of tropical pioneer seedlings. Oecologia 148, 258–269.
| Short dry spells in the wet season increase mortality of tropical pioneer seedlings.Crossref | GoogleScholarGoogle Scholar |
Engelbrecht BMJ, Comita LS, Condit R, Kursar TA, Tyree MT, Turner BL, Hubbell SP (2007) Drought sensitivity shapes species distribution patterns in tropical forests. Nature 447, 80–82.
| Drought sensitivity shapes species distribution patterns in tropical forests.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2sXkvVaju78%3D&md5=7c33f1b1c305c6667bf214ce5d360980CAS |
Farquhar GD, Richards RA (1984) Isotopic composition of plant carbon correlates with water-use efficiency in wheat genotypes. Australian Journal of Plant Physiology 11, 539–552.
| Isotopic composition of plant carbon correlates with water-use efficiency in wheat genotypes.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaL2MXhtFSju7w%3D&md5=6d68b1a5561d71b7f0f480fb6c702091CAS |
Farquhar GD, Sharkey TD (1982) Stomatal conductance and photosynthesis. Annual Review of Plant Physiology 33, 317–345.
| Stomatal conductance and photosynthesis.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaL38XktlKjs7o%3D&md5=829b005fabe66430031b82818f4c3429CAS |
Farquhar GD, von Caemmerer S, Berry JA (1980) A biochemical model of photosynthetic CO2 assimilation in leaves of C3 species. Planta 149, 78–90.
| A biochemical model of photosynthetic CO2 assimilation in leaves of C3 species.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaL3cXksVWrt7w%3D&md5=b0417117f3598df013ab17a3da1f67a8CAS |
Farquhar GD, O’Leary MH, Berry JA (1982) On the relationship between carbon isotope discrimination and the intercellular carbon dioxide concentration in leaves. Australian Journal of Plant Physiology 9, 121–137.
| On the relationship between carbon isotope discrimination and the intercellular carbon dioxide concentration in leaves.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaL38XhsF2ms70%3D&md5=b75c628f7553ab4d91e590fd2bab057aCAS |
Field CB, Behrenfeld MJ, Randerson JT, Falkowski P (1998) Primary production of the biosphere: integrating terrestrial and oceanic components. Science 281, 237–240.
| Primary production of the biosphere: integrating terrestrial and oceanic components.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaK1cXksFKitb0%3D&md5=0fd98279ba9ea8116c38792ff44e6a97CAS | 9657713PubMed |
Fisher R, McDowell N, Purves D, Moorcroft P, Sitch S, Cox P, Huntingford C, Meir P, Woodward FI (2010) Assessing uncertainties in a second-generation dynamic vegetation model caused by ecological scale limitations. New Phytologist 187, 666–681.
| Assessing uncertainties in a second-generation dynamic vegetation model caused by ecological scale limitations.Crossref | GoogleScholarGoogle Scholar | 20618912PubMed |
Fyllas NM, Patino S, Baker TR, Nardoto GB, Martinelli LA, Quesada CA, Paiva R, Schwarz M, Horna V, Mercado LM (2009) Basin-wide variations in foliar properties of Amazonian forest: phylogeny, soils and climate. Biogeosciences 6, 2677–2708.
| Basin-wide variations in foliar properties of Amazonian forest: phylogeny, soils and climate.Crossref | GoogleScholarGoogle Scholar |
Galbraith D, Levy PE, Sitch S, Huntingford C, Cox P, Williams M, Meir P (2010) Multiple mechanisms of Amazonian forest biomass losses in three dynamic global vegetation models under climate change. New Phytologist 187, 647–665.
| Multiple mechanisms of Amazonian forest biomass losses in three dynamic global vegetation models under climate change.Crossref | GoogleScholarGoogle Scholar | 20659253PubMed |
Garstang M, White S, Shugart HH, Halverson J (1998) Convective cloud downdrafts as the cause of large blowdowns in the Amazon rainforest. Meteorology and Atmospheric Physics 67, 199–212.
| Convective cloud downdrafts as the cause of large blowdowns in the Amazon rainforest.Crossref | GoogleScholarGoogle Scholar |
Gedney N, Cox PM, Betts RA, Boucher O, Huntingford C, Stott PA (2006) Detection of a direct carbon dioxide effect in continental river runoff records. Nature 439, 835–838.
| Detection of a direct carbon dioxide effect in continental river runoff records.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD28XhsVSku7o%3D&md5=6f5f41d85eeadeca2dbf44b54ef5d16aCAS | 16482155PubMed |
Gentry AH (1988) Changes in plant community diversity and floristic composition on environmental and geographical gradients. Annals of the Missouri Botanical Garden 75, 1–34.
| Changes in plant community diversity and floristic composition on environmental and geographical gradients.Crossref | GoogleScholarGoogle Scholar |
Gloor M, Phillips OL, Lloyd JJ, Lewis SL, Malhi Y, Baker TR, López-Gonzalez G, Peacock J, Almeida S, Alves de Oliveira AC, Alvarez E, Amaral I, Arroyo L, Aymard G, Banki O, Blanc L, Bonal D, Brando P, Chao KJ, Chave J, Dávila N, Erwin T, Silva J, Di Fiore A, Feldpausch TR, Freitas A, Herrera R, Higuchi N, Honorio E, Jiménez E, Killeen T, Laurance W, Mendoza C, Monteagudo A, Andrade A, Neill D, Nepstad D, Núñez Vargas P, Peñuela MC, Peña Cruz A, Prieto A, Pitman N, Quesada C, Salomão R, Silveira M, Schwarz M, Stropp J, Ramírez F, Ramírez H, Rudas A, Ter Steege H, Silva N, Torres A, Terborgh J, Vasquéz R, Van Der Heijden G (2009) Does the disturbance hypothesis explain the biomass increase in basin-wide Amazon forest plot data? Global Change Biology 15, 2418–2430.
| Does the disturbance hypothesis explain the biomass increase in basin-wide Amazon forest plot data?Crossref | GoogleScholarGoogle Scholar |
Goll DS, Brovkin V, Parida BR, Reick CH, Kattge J, Reich PB, van Bodegom PM, Niinemets U (2012) Nutrient limitation reduces land carbon uptake in simulations with a model of combined carbon, nitrogen and phosphorus cycling. Biogeosciences Discussions 9, 3173–3232.
| Nutrient limitation reduces land carbon uptake in simulations with a model of combined carbon, nitrogen and phosphorus cycling.Crossref | GoogleScholarGoogle Scholar |
Gonzàlez-Meler MA, Blanc-Betes E, Flower CE, Ward JK, Gomez-Casanovas N (2009) Plastic and adaptive responses of plant respiration to changes in atmospheric CO2 concentration. Physiologia Plantarum 137, 473–484.
| Plastic and adaptive responses of plant respiration to changes in atmospheric CO2 concentration.Crossref | GoogleScholarGoogle Scholar | 19671094PubMed |
Goodfellow J, Eamus D, Duff G (1997) Diurnal and seasonal changes in the impact of CO2 enrichment on assimilation, stomatal conductance and growth in a long-term study of Mangifera indica in the wet–dry tropics of Australia. Tree Physiology 17, 291–299.
| Diurnal and seasonal changes in the impact of CO2 enrichment on assimilation, stomatal conductance and growth in a long-term study of Mangifera indica in the wet–dry tropics of Australia.Crossref | GoogleScholarGoogle Scholar | 14759852PubMed |
Goulden ML, Miller SD, da Rocha HR, Menton MC, de Freitas HC, Figueira AMES, de Sousa CAD (2004) Diel and seasonal patterns of tropical forest CO2 exchange. Ecological Applications 14, 42–54.
| Diel and seasonal patterns of tropical forest CO2 exchange.Crossref | GoogleScholarGoogle Scholar |
Graham EA, Mulkey SS, Kitajima K, Phillips NG, Wright SJ (2003) Cloud cover limits net CO2 uptake and growth of a rainforest tree during tropical rainy seasons. Proceedings of the National Academy of Sciences of the United States of America 100, 572–576.
| Cloud cover limits net CO2 uptake and growth of a rainforest tree during tropical rainy seasons.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD3sXnvVKnug%3D%3D&md5=acc493a4400b2714942f4c1e94a364e5CAS | 12518044PubMed |
Guehl JM, Picon C, Aussenac G, Gross P (1994) Interactive effects of elevated CO2 and soil drought on growth and transpiration efficiency and its determinants in two European forest tree species. Tree Physiology 14, 707–724.
| Interactive effects of elevated CO2 and soil drought on growth and transpiration efficiency and its determinants in two European forest tree species.Crossref | GoogleScholarGoogle Scholar | 14967642PubMed |
Hedin LO, Brookshire ENJ, Menge DNL, Barron AR (2009) The nitrogen paradox in tropical forest ecosystems. Annual Review of Ecology Evolution and Systematics 40, 613–635.
| The nitrogen paradox in tropical forest ecosystems.Crossref | GoogleScholarGoogle Scholar |
Hickler T, Smith B, Prentice IC, Mjofors K, Miller P, Arneth A, Sykes MT (2008) CO2 fertilization in temperate FACE experiments not representative of boreal and tropical forests. Global Change Biology 14, 1531–1542.
| CO2 fertilization in temperate FACE experiments not representative of boreal and tropical forests.Crossref | GoogleScholarGoogle Scholar |
Hietz P, Wanek W, Dunisch O (2005) Long-term trends in cellulose δ13C and water-use efficiency of tropical Cedrela and Swietenia from Brazil. Tree Physiology 25, 745–752.
| Long-term trends in cellulose δ13C and water-use efficiency of tropical Cedrela and Swietenia from Brazil.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2MXlvVSgsLw%3D&md5=873752973a25a2c8422455fe0c7630f0CAS | 15805094PubMed |
Hietz P, Turner BL, Wanek W, Richter A, Nock CA, Wright SJ (2011) Long-term change in the nitrogen cycle of tropical forests. Science 334, 664–666.
| Long-term change in the nitrogen cycle of tropical forests.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3MXhtlyqu7jI&md5=f423260e7698d2c15f33a7e4d25c50faCAS | 22053047PubMed |
Hogan KP, Smith AP, Ziska LH (1991) Potential effects of elevated CO2 and changes in temperature on tropical plants. Plant, Cell & Environment 14, 763–778.
| Potential effects of elevated CO2 and changes in temperature on tropical plants.Crossref | GoogleScholarGoogle Scholar |
Holtum JAM, Winter K (2001) Are plants growing close to the floors of tropical forests exposed to markedly elevated concentrations of carbon dioxide? Australian Journal of Botany 49, 629–636.
| Are plants growing close to the floors of tropical forests exposed to markedly elevated concentrations of carbon dioxide?Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD3MXptVemu7g%3D&md5=99ead393ad6778ebb6fbfefab6db1187CAS |
Holtum JAM, Winter K (2003) Photosynthetic CO2 uptake in seedlings of two tropical tree species exposed to oscillating elevated concentrations of CO2. Planta 218, 152–158.
| Photosynthetic CO2 uptake in seedlings of two tropical tree species exposed to oscillating elevated concentrations of CO2.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD3sXovV2ltLg%3D&md5=97e41a95f0f7da07dfbe321acc17e603CAS |
Holtum JAM, Winter K (2010) Elevated [CO2] and forest vegetation: more a water issue than a carbon issue? Functional Plant Biology 37, 694–702.
| Elevated [CO2] and forest vegetation: more a water issue than a carbon issue?Crossref | GoogleScholarGoogle Scholar |
Holtum JAM, Aranda J, Virgo A, Gehrig HH, Winter K (2004) δ13C values and crassulacean acid metabolism in Clusia species from Panama. Trees 18, 658–668.
| δ13C values and crassulacean acid metabolism in Clusia species from Panama.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2cXovVKqsbY%3D&md5=1df634009ad188b60c23a0227ee9c9f9CAS |
Houlton BZ, Wang YP, Vitousek PM, Field CB (2008) A unifying framework for dinitrogen fixation in the terrestrial biosphere. Nature 454, 327–330.
| A unifying framework for dinitrogen fixation in the terrestrial biosphere.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD1cXosFCqs7w%3D&md5=e7b5140e3f12e571ecf7f3965bf6cb1aCAS | 18563086PubMed |
Hsiao TC (1973) Plant responses to water stress. Annual Review of Plant Physiology and Plant Molecular Biology 24, 519–570.
| Plant responses to water stress.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaE3sXlt1emurY%3D&md5=a5e5aaee17ed731cbfb99fe1c300e030CAS |
Hüve K, Bichele I, Tobias M, Niinemets U (2006) Heat sensitivity of photosynthetic electron transport varies during the day due to changes in sugars and osmotic potential. Plant, Cell & Environment 29, 212–228.
| Heat sensitivity of photosynthetic electron transport varies during the day due to changes in sugars and osmotic potential.Crossref | GoogleScholarGoogle Scholar |
Ingwell LL, Wright SJ, Becklund KK, Hubbell SP, Schnitzer SA (2010) The impact of lianas on 10 years of tree growth and mortality on Barro Colorado Island, Panama. Journal of Ecology 98, 879–887.
| The impact of lianas on 10 years of tree growth and mortality on Barro Colorado Island, Panama.Crossref | GoogleScholarGoogle Scholar |
Intergovernmental Panel on Climate Change (IPCC) (2011) Carbon dioxide: projected emissions and concentrations. (IPCC: Geneva) Available at: http://www.ipcc-data.org/ddc_co2.html [Verified 5 April 2013]
Ishida A, Toma T, Marjenah (1999) Limitation of leaf carbon gain by stomatal and photochemical processes in the top canopy of Macaranga conifera, a tropical pioneer tree. Tree Physiology 19, 467–473.
| Limitation of leaf carbon gain by stomatal and photochemical processes in the top canopy of Macaranga conifera, a tropical pioneer tree.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaK1MXjvVeitbg%3D&md5=67d6f9c893934838b5bcacdb33f5c21aCAS | 12651553PubMed |
Jaramillo C, Ochoa D, Contreras L, Pagani M, Carvajal-Ortiz H, Pratt LM, Krishnan S, Cardona A, Romero M, Quiroz L, Rodriguez G, Rueda MJ, de la Parra F, Morón S, Green W, Bayona G, Montes C, Quintero O, Ramirez R, Mora G, Schouten S, Bermudez H, Navarrete R, Parra F, Alvarán M, Osorno J, Crowley JL, Valencia V, Vervoort J (2010) Effects of rapid global warming at the Paleocene–Eocene boundary on Neotropical vegetation. Science 330, 957–961.
| Effects of rapid global warming at the Paleocene–Eocene boundary on Neotropical vegetation.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3cXhtl2isbvE&md5=5b5382f3a4fe30cd623bb50e4e9fc817CAS | 21071667PubMed |
Johnson AH, Frizano J, Vann DR (2003) Biogeochemical implications of labile phosphorus in forest soils determined by the Hedley fractionation procedure. Oecologia 135, 487–499.
Keel SG, Pepin S, Leuzinger S, Körner C (2007) Stomatal conductance in mature deciduous forest trees exposed to elevated CO2. Trees – Structure and Function 21, 151–159.
| Stomatal conductance in mature deciduous forest trees exposed to elevated CO2.Crossref | GoogleScholarGoogle Scholar |
Knutson T, McBride J, Chan J, Emanuel K, Holland G, Landsea C, Held I, Kossin JP, Srivastava AK, Sugi M (2010) Tropical cyclones and climate change. Nature Geoscience 3, 157–163.
| Tropical cyclones and climate change.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3cXisVahur4%3D&md5=dc9cf008ec1b2f5d0dd48465db8d9d2eCAS |
Koch GW, Amthor JS, Goulden ML (1994) Diurnal patterns of leaf photosynthesis, conductance and water potential at the top of a lowland rain forest canopy in Cameroon: measurements from the Radeau des Cimes. Tree Physiology 14, 347–360.
| Diurnal patterns of leaf photosynthesis, conductance and water potential at the top of a lowland rain forest canopy in Cameroon: measurements from the Radeau des Cimes.Crossref | GoogleScholarGoogle Scholar | 14967691PubMed |
Körner C (2003) Carbon limitation in trees. Journal of Ecology 91, 4–17.
| Carbon limitation in trees.Crossref | GoogleScholarGoogle Scholar |
Körner C (2004) Through enhanced tree dynamics carbon dioxide enrichment may cause tropical forests to lose carbon. Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences 359, 493–498.
| Through enhanced tree dynamics carbon dioxide enrichment may cause tropical forests to lose carbon.Crossref | GoogleScholarGoogle Scholar | 15212098PubMed |
Körner C (2009) Responses of humid tropical trees to rising CO2. Annual Review of Ecology Evolution and Systematics 40, 61–79.
| Responses of humid tropical trees to rising CO2.Crossref | GoogleScholarGoogle Scholar |
Körner C, Arnone JA (1992) Responses to elevated carbon dioxide in artificial tropical ecosystems. Science 257, 1672–1675.
| Responses to elevated carbon dioxide in artificial tropical ecosystems.Crossref | GoogleScholarGoogle Scholar | 17841166PubMed |
Körner C, Würth M (1996) A simple method for testing leaf responses of tall tropical forest trees to elevated CO2. Oecologia 107, 421–425.
| A simple method for testing leaf responses of tall tropical forest trees to elevated CO2.Crossref | GoogleScholarGoogle Scholar |
Körner C, Asshoff R, Bignucolo O, Hättenschwiler S, Keel SG, Pelaez-Riedl S, Pepin S, Siegwolf RTW, Zotz G (2005) Carbon flux and growth in mature deciduous forest trees exposed to elevated CO2. Science 309, 1360–1362.
| Carbon flux and growth in mature deciduous forest trees exposed to elevated CO2.Crossref | GoogleScholarGoogle Scholar | 16123297PubMed |
Kosugi Y, Takanashi S, Ohkubo S, Matsuo N, Tani M, Mitani T, Tsutsumi D, Nik AR (2008) CO2 exchange of a tropical rainforest at Pasoh in peninsular Malaysia. Agricultural and Forest Meteorology 148, 439–452.
| CO2 exchange of a tropical rainforest at Pasoh in peninsular Malaysia.Crossref | GoogleScholarGoogle Scholar |
Kosugi Y, Takanashi S, Matsuo N, Nik AR (2009) Midday depression of leaf CO2 exchange within the crown of Dipterocarpus sublamellatus in a lowland dipterocarp forest in peninsular Malaysia. Tree Physiology 29, 505–515.
| Midday depression of leaf CO2 exchange within the crown of Dipterocarpus sublamellatus in a lowland dipterocarp forest in peninsular Malaysia.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD1MXkslKnsLc%3D&md5=7150a5c1d7c2a14198579d8d2163fc47CAS | 19203974PubMed |
Krause GH, Winter K, Krause B, Jahns P, Garcia M, Aranda J, Virgo A (2010) High-temperature tolerance of a tropical tree, Ficus insipida: methodological reassessment and climate change considerations. Functional Plant Biology 37, 890–900.
| High-temperature tolerance of a tropical tree, Ficus insipida: methodological reassessment and climate change considerations.Crossref | GoogleScholarGoogle Scholar |
Kriedemann PE, Sward RJ, Downton WJS (1976) Vine response to carbon dioxide enrichment during heat therapy. Australian Journal of Plant Physiology 3, 605–618.
| Vine response to carbon dioxide enrichment during heat therapy.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaE28XlslCntb0%3D&md5=002be31e6e3b872e5cc76f28da09987bCAS |
Krinner G, Viovy N, de Noblet-Ducoudre N, Ogee J, Polcher J, Friedlingstein P, Ciais P, Sitch S, Prentice IC (2005) A dynamic global vegetation model for studies of the coupled atmosphere-biosphere system. Global Biogeochemical Cycles 19, GB1015
| A dynamic global vegetation model for studies of the coupled atmosphere-biosphere system.Crossref | GoogleScholarGoogle Scholar |
Ladeau SL, Clark JS (2006) Elevated CO2 and tree fecundity: the role of tree size, interannual variability, and population heterogeneity. Global Change Biology 12, 822–833.
| Elevated CO2 and tree fecundity: the role of tree size, interannual variability, and population heterogeneity.Crossref | GoogleScholarGoogle Scholar |
Lambers H, Raven JA, Shaver GR, Smith SE (2008) Plant nutrient-acquisition strategies change with soil age. Trends in Ecology & Evolution 23, 95–103.
| Plant nutrient-acquisition strategies change with soil age.Crossref | GoogleScholarGoogle Scholar |
Lapola DM, Oyama MD, Nobre CA (2009) Exploring the range of climate biome projections for tropical South America: the role of CO2 fertilization and seasonality. Global Biogeochemical Cycles 23, 1–16.
| Exploring the range of climate biome projections for tropical South America: the role of CO2 fertilization and seasonality.Crossref | GoogleScholarGoogle Scholar |
Laurance WF, Oliveira AA, Laurance SG, Condit R, Nascimento HEM, Sanchez-Thorin AC, Lovejoy TE, Andrade A, D’Angelo S, Ribeiro JE, Dick CW (2004) Pervasive alteration of tree communities in undisturbed Amazonian forests. Nature 428, 171–175.
| Pervasive alteration of tree communities in undisturbed Amazonian forests.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2cXhvFCls7s%3D&md5=c70889a78937653a71910a66c8054387CAS | 15014498PubMed |
Leakey ADB, Lau JA (2012) Evolutionary context for understanding and manipulating plant responses to past, present and future atmospheric [CO2]. Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences 367, 613–629.
| Evolutionary context for understanding and manipulating plant responses to past, present and future atmospheric [CO2].Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC38XltV2rur4%3D&md5=c1172bbc67a5164f8e75aa6ddf73590bCAS |
Leakey ADB, Press MC, Scholes JD, Watling JR (2002) Relative enhancement of photosynthesis and growth at elevated CO2 is greater under sunflecks than uniform irradiance in a tropical rain forest tree seedling. Plant, Cell & Environment 25, 1701–1714.
| Relative enhancement of photosynthesis and growth at elevated CO2 is greater under sunflecks than uniform irradiance in a tropical rain forest tree seedling.Crossref | GoogleScholarGoogle Scholar |
Leakey ADB, Xu F, Gillespie KM, McGrath JM, Ainsworth EA, Ort DR (2009) Genomic basis for stimulated respiration by plants growing under elevated carbon dioxide. Proceedings of the National Academy of Sciences of the United States of America 106, 3597–3602.
| Genomic basis for stimulated respiration by plants growing under elevated carbon dioxide.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD1MXivFKjtLY%3D&md5=87245c8a750770a9670602c9a5364b94CAS |
Leakey ADB, Bishop KA, Ainsworth EA (2012) A multi-biome gap in understanding of crop and ecosystem responses to elevated CO2. Current Opinion in Plant Biology 15, 228–236.
| A multi-biome gap in understanding of crop and ecosystem responses to elevated CO2.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC38Xnsl2rsro%3D&md5=790ad381ed5817b5a7d6f0679f92c957CAS |
Leuzinger S, Körner C (2007) Water savings in mature deciduous forest trees under elevated CO2. Global Change Biology 13, 2498–2508.
| Water savings in mature deciduous forest trees under elevated CO2.Crossref | GoogleScholarGoogle Scholar |
Leuzinger S, Körner C (2010) Rainfall distribution is the main driver of runoff under future CO2 concentration in a temperate deciduous forest. Global Change Biology 16, 246–254.
| Rainfall distribution is the main driver of runoff under future CO2 concentration in a temperate deciduous forest.Crossref | GoogleScholarGoogle Scholar |
Lewis SL, Phillips OL, Baker TR, Lloyd J, Malhi Y, Almeida S, Higuchi N, Laurance WF, Neill DA, Silva JNM, Terborgh J, Torres Lezama A, Vásquez Martinez R, Brown S, Chave J, Kuebler C, Núñez Vargas P, Vinceti B (2004) Concerted changes in tropical forest structure and dynamics: evidence from 50 South American long-term plots. Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences 359, 421–436.
| Concerted changes in tropical forest structure and dynamics: evidence from 50 South American long-term plots.Crossref | GoogleScholarGoogle Scholar | 1:STN:280:DC%2BD2czhtVSqtw%3D%3D&md5=d1e692a2681b576c03250c92512b567cCAS | 15212094PubMed |
Lewis SL, Lloyd J, Sitch S, Mitchard ETA, Laurance WF (2009a) Changing ecology of tropical forests: evidence and drivers. Annual Review of Ecology Evolution and Systematics 40, 529–549.
| Changing ecology of tropical forests: evidence and drivers.Crossref | GoogleScholarGoogle Scholar |
Lewis SL, Lopez-Gonzalez G, Sonké B, Affum-Baffoe K, Baker TR, Ojo LO, Phillips OL, Reitsma JM, White L, Comiskey JA, Djuikouo MN, Ewango CEN, Feldpausch TE, Hamilton AC, Gloor M, Hart T, Hladik A, Lloyd J, Lovett JC, Makana J-R, Malhi Y, Mbago FM, Ndangalasi HJ, Peacock J, Peh KS-H, Sheil D, Sunderland T, Swaine MD, Taplin J, Taylor D, Thomas SC, Votere R, Wöll H (2009b) Increasing carbon storage in intact African tropical forests. Nature 457, 1003–1006.
| Increasing carbon storage in intact African tropical forests.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD1MXitFKktrs%3D&md5=2d4a0ded0e011d2bce3f9f74fa53981bCAS | 19225523PubMed |
Lewis S, Brando P, Phillips O, van der Heijden G, Nepstad D (2011) The 2010 Amazon drought. Science 331, 554
| The 2010 Amazon drought.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3MXhtlWisLs%3D&md5=6371b4f3d4b99680d2e09038783dbd45CAS | 21292971PubMed |
Lloyd J, Farquhar GD (1996) The CO2 dependence of photosynthesis, plant growth responses to elevated atmospheric CO2 concentrations and their interaction with soil nutrient status. I. General principles and forest ecosystems. Functional Ecology 10, 4–32.
| The CO2 dependence of photosynthesis, plant growth responses to elevated atmospheric CO2 concentrations and their interaction with soil nutrient status. I. General principles and forest ecosystems.Crossref | GoogleScholarGoogle Scholar |
Lloyd J, Farquhar GD (2008) Effects of rising temperatures and [CO2] on the physiology of tropical forest trees. Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences 363, 1811–1817.
| Effects of rising temperatures and [CO2] on the physiology of tropical forest trees.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD1cXmsFGqt7w%3D&md5=8d9c0669037a9a362c57e30c2dba16dbCAS | 18267901PubMed |
Lloyd J, Grace J, Miranda AC, Meir P, Wong SC, Miranda BS, Wright IR, Gash JHC, McIntyre J (1995) A simple calibrated model of Amazon rainforest productivity based on leaf biochemical properties. Plant, Cell & Environment 18, 1129–1145.
| A simple calibrated model of Amazon rainforest productivity based on leaf biochemical properties.Crossref | GoogleScholarGoogle Scholar |
Lloyd J, Kruijt B, Hollinger DY, Grace J, Francey RJ, Wong SC, Kelliher FM, Miranda AC, Farquhar GD, Gash JHC, Vygodskaya NN, Wright IR, Miranda HS, Schulze ED (1996) Vegetation effects on the isotopic composition of atmospheric CO2 at local and regional scales: theoretical aspects and a comparison between rain forest in Amazonia and a boreal forest in Siberia. Australian Journal of Plant Physiology 23, 371–399.
| Vegetation effects on the isotopic composition of atmospheric CO2 at local and regional scales: theoretical aspects and a comparison between rain forest in Amazonia and a boreal forest in Siberia.Crossref | GoogleScholarGoogle Scholar |
Lloyd J, Bird MI, Veenendaal EM, Kruijt B (2001) Should phosphorus availability be constraining moist tropical forest responses to increasing CO2 concentrations? In ‘Global biogeochemical cycles in the climate system’. (Eds E-D Schulze, M Heimann, S Harrison, E Holland, J Lloyd, IC Prentice, D Schimel) pp. 95–114. (Academic Press: San Diego)
Lloyd J, Gloor EU, Lewis SL (2009) Are the dynamics of tropical forests dominated by large and rare disturbance events? Ecology Letters 12, E19–E21.
| Are the dynamics of tropical forests dominated by large and rare disturbance events?Crossref | GoogleScholarGoogle Scholar | 19930036PubMed |
Loader NJ, Walsh RPD, Robertson I, Bidin K, Ong RC, Reynolds G, McCarroll D, Gagen M, Young GHF (2011) Recent trends in the intrinsic water-use efficiency of ringless rainforest trees in Borneo. Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences 366, 3330–3339.
| Recent trends in the intrinsic water-use efficiency of ringless rainforest trees in Borneo.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3MXhs1KksbvN&md5=4af711844f72a8aa3f560834201f4de9CAS | 22006972PubMed |
Long SP (1991) Modification of the response of photosynthetic productivity to rising temperature by atmospheric CO2 concentrations – has its importance been underestimated. Plant, Cell & Environment 14, 729–739.
| Modification of the response of photosynthetic productivity to rising temperature by atmospheric CO2 concentrations – has its importance been underestimated.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaK38XhsVyns7s%3D&md5=aef20ae7edcbb9febebd7cd28e15e186CAS |
Long SP, Ainsworth EA, Rogers A, Ort DR (2004) Rising atmospheric carbon dioxide: plants face the future. Annual Review of Plant Biology 55, 591–628.
| Rising atmospheric carbon dioxide: plants face the future.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2cXlvFeisb8%3D&md5=90372ae3a3ad173142e8eb82fdb1595aCAS | 15377233PubMed |
Losos E, Leigh E (Eds) (2004) ‘Tropical forest diversity and dynamism: findings from a large-scale plot network.’ (University of Chicago Press: Chicago)
Lovelock CE, Kyllo D, Winter K (1996) Growth responses to vesicular-arbuscular mycorrhizae and elevated CO2 in seedlings of a tropical tree, Beilschmiedia pendula. Functional Ecology 10, 662–667.
| Growth responses to vesicular-arbuscular mycorrhizae and elevated CO2 in seedlings of a tropical tree, Beilschmiedia pendula.Crossref | GoogleScholarGoogle Scholar |
Lovelock CE, Winter K, Mersits R, Popp M (1998) Responses of communities of tropical tree species to elevated CO2 in a forest clearing. Oecologia 116, 207–218.
| Responses of communities of tropical tree species to elevated CO2 in a forest clearing.Crossref | GoogleScholarGoogle Scholar |
Lovelock CE, Virgo A, Popp M, Winter K (1999) Effects of elevated CO2 concentrations on photosynthesis, growth and reproduction of branches of the tropical canopy tree species, Luehea seemannii Tr. & Planch. Plant, Cell & Environment 22, 49–59.
| Effects of elevated CO2 concentrations on photosynthesis, growth and reproduction of branches of the tropical canopy tree species, Luehea seemannii Tr. & Planch.Crossref | GoogleScholarGoogle Scholar |
Luo YQ, Melillo J, Niu S, Beier C, Clark JS, Classen AT, Davidson E, Dukes JS, Evans RD, Field CB, Czimczik CI, Keller M, Kimball BA, Norby RJ, Pelini SL, Pendall E, Rastetter E, Six J, Smith M, Tjoelker MG, Torn MS (2011) Coordinated approaches to quantify long-term ecosystem dynamics in response to global change. Global Change Biology 17, 843–854.
| Coordinated approaches to quantify long-term ecosystem dynamics in response to global change.Crossref | GoogleScholarGoogle Scholar |
Lüthi D, Le Floch M, Bereiter B, Blunier T, Barnola J-M, Siegenthaler U, Raynaud D, Jouzel J, Fischer H, Kawamura K, Stocker TF (2008) High-resolution carbon dioxide concentration record 650,000–800,000 years before present. Nature 453, 379–382.
| High-resolution carbon dioxide concentration record 650,000–800,000 years before present.Crossref | GoogleScholarGoogle Scholar | 18480821PubMed |
Macinnis-Ng C, Zeppel M, Williams M, Eamus D (2011) Applying a SPA model to examine the impact of climate change on GPP of open woodlands and the potential for woody thickening. Ecohydrology 4, 379–393.
| Applying a SPA model to examine the impact of climate change on GPP of open woodlands and the potential for woody thickening.Crossref | GoogleScholarGoogle Scholar |
Malhi Y (2012) The productivity, metabolism and carbon cycle of tropical forest vegetation. Journal of Ecology 100, 65–75.
| The productivity, metabolism and carbon cycle of tropical forest vegetation.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC38Xit1ylurw%3D&md5=b60829e6187e1affe3d378f5f06b53ceCAS |
Malhi Y, Grace J (2000) Tropical forests and atmospheric carbon dioxide. Trends in Ecology & Evolution 15, 332–337.
| Tropical forests and atmospheric carbon dioxide.Crossref | GoogleScholarGoogle Scholar |
Malhi Y, Wright J (2004) Spatial patterns and recent trends in the climate of tropical forest regions. Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences 359, 311–329.
| Spatial patterns and recent trends in the climate of tropical forest regions.Crossref | GoogleScholarGoogle Scholar | 15212087PubMed |
Malhi Y, Baldocchi DD, Jarvis PG (1999) The carbon balance of tropical, temperate and boreal forests. Plant, Cell & Environment 22, 715–740.
| The carbon balance of tropical, temperate and boreal forests.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaK1MXksVartb8%3D&md5=4e27aaa56602a40c060701c1188ac1e3CAS |
Malhi Y, Wood D, Baker TR, Wright J, Phillips OL, Cochrane T, Meir P, Chave J, Almeida S, Arroyo L, Higuchi N, Killeen TJ, Laurance SG, Laurance WF, Lewis SL, Monteagudo A, Neill DA, Núñez Vargas P, Pitman NCA, Quesada CA, Salomão R, Silva JNM, Torres Lezama A, Terborgh J, Vásquez Martínez R, Vinceti B (2006) The regional variation of aboveground live biomass in old-growth Amazonian forests. Global Change Biology 12, 1107–1138.
| The regional variation of aboveground live biomass in old-growth Amazonian forests.Crossref | GoogleScholarGoogle Scholar |
Malhi Y, Doughty C, Galbraith D (2011) The allocation of ecosystem net primary productivity in tropical forests. Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences 366, 3225–3245.
| The allocation of ecosystem net primary productivity in tropical forests.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3MXhs1KksbvM&md5=8950600f46310cd4332a3bb4353ab546CAS | 22006964PubMed |
McCarthy HR, Oren R, Johnsen KH, Gallet-Budynek A, Pritchard SG, Cook CW, LaDeau SL, Jackson RB, Finzi AC (2010) Re-assessment of plant carbon dynamics at the Duke free-air CO2 enrichment site: interactions of atmospheric CO2 with nitrogen and water availability over stand development. New Phytologist 185, 514–528.
| Re-assessment of plant carbon dynamics at the Duke free-air CO2 enrichment site: interactions of atmospheric CO2 with nitrogen and water availability over stand development.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3cXhsVyrsLs%3D&md5=10d18073722c513f03b4e2f987005d87CAS | 19895671PubMed |
McDowell N, Pockman WT, Allen CD, Breshears DD, Cobb N, Kolb T, Plaut J, Sperry J, West A, Williams DG, Yepez EA (2008) Mechanisms of plant survival and mortality during drought: why do some plants survive while others succumb to drought? New Phytologist 178, 719–739.
| Mechanisms of plant survival and mortality during drought: why do some plants survive while others succumb to drought?Crossref | GoogleScholarGoogle Scholar | 18422905PubMed |
McDowell NG, Beerling DJ, Breshears DD, Fisher RA, Raffa KF, Stitt M (2011) The interdependence of mechanisms underlying climate-driven vegetation mortality. Trends in Ecology & Evolution 26, 523–532.
| The interdependence of mechanisms underlying climate-driven vegetation mortality.Crossref | GoogleScholarGoogle Scholar |
McHargue LA (1999) Factors affecting the nodulation and growth of tropical woody legume seedlings. PhD Thesis thesis, Florida International University, Miami, Florida.
McMurtrie RE, Norby RJ, Medlyn BE, Dewar RC, Pepper DA, Reich PB, Barton CVM (2008) Why is plant-growth response to elevated CO2 amplified when water is limiting, but reduced when nitrogen is limiting? A growth-optimisation hypothesis. Functional Plant Biology 35, 521–534.
| Why is plant-growth response to elevated CO2 amplified when water is limiting, but reduced when nitrogen is limiting? A growth-optimisation hypothesis.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD1cXpt1emtbY%3D&md5=299584b228de10c25b533b8e5c128527CAS |
Medlyn BE, Duursma RA, Eamus D, Ellsworth DS, Prentice IC, Barton CVM, Crous KY, de Angelis P, Freeman M, Wingate L (2011) Reconciling the optimal and empirical approaches to modelling stomatal conductance. Global Change Biology 17, 2134–2144.
| Reconciling the optimal and empirical approaches to modelling stomatal conductance.Crossref | GoogleScholarGoogle Scholar |
Medvigy D, Wofsy SC, Munger JW, Hollinger DY, Moorcroft PR (2009) Mechanistic scaling of ecosystem function and dynamics in space and time: ecosystem demography model version 2. Journal of Geophysical Research-Biogeosciences 114,
| Mechanistic scaling of ecosystem function and dynamics in space and time: ecosystem demography model version 2.Crossref | GoogleScholarGoogle Scholar |
Mercado L, Lloyd J, Carswell F, Malhi Y, Meir P, Nobre AD (2006) Modelling Amazonian forest eddy covariance data: a comparison of big leaf versus sun/shade models for the C-14 tower at Manaus I. Canopy photosynthesis. Acta Amazonica 36, 69–82.
| Modelling Amazonian forest eddy covariance data: a comparison of big leaf versus sun/shade models for the C-14 tower at Manaus I. Canopy photosynthesis.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD28XlsF2jtL8%3D&md5=4ffca3e9569d258e6dc70ab5bfe46886CAS |
Mercado LM, Patiño S, Domingues TF, Fyllas NM, Weedon GP, Sitch S, Quesada CA, Phillips OL, Aragão LEOC, Malhi Y, Dolman AJ, Restrepo-Coupe N, Saleska SR, Baker TR, Almeida S, Higuchi N, Lloyd J (2011) Variations in Amazon forest productivity correlated with foliar nutrients and modelled rates of photosynthetic carbon supply. Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences 366, 3316–3329.
| Variations in Amazon forest productivity correlated with foliar nutrients and modelled rates of photosynthetic carbon supply.Crossref | GoogleScholarGoogle Scholar | 22006971PubMed |
Metcalfe DB, Meir P, Aragão LEOC, Lobo-do-Vale R, Galbraith D, Fisher RA, Chaves MM, Maroco JP, da Costa ACL, de Almeida SS, Braga AP, Gonçalves PHL, de Athaydes J, da Costa M, Portela TTB, de Oliveira AAR, Malhi Y, Williams M (2010) Shifts in plant respiration and carbon use efficiency at a large-scale drought experiment in the eastern Amazon. New Phytologist 187, 608–621.
| Shifts in plant respiration and carbon use efficiency at a large-scale drought experiment in the eastern Amazon.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3cXhtFSmu7jO&md5=71a735abd1741520a7d65cfdf4fb931eCAS | 20553394PubMed |
Moorcroft PR, Hurtt GC, Pacala SW (2001) A method for scaling vegetation dynamics: the ecosystem demography model (ED). Ecological Monographs 71, 557–586.
| A method for scaling vegetation dynamics: the ecosystem demography model (ED).Crossref | GoogleScholarGoogle Scholar |
Moore BD, Cheng SH, Sims D, Seemann JR (1999) The biochemical and molecular basis for photosynthetic acclimation to elevated atmospheric CO2. Plant, Cell & Environment 22, 567–582.
| The biochemical and molecular basis for photosynthetic acclimation to elevated atmospheric CO2.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaK1MXksVartL0%3D&md5=e2bb5ed85001378589dea2d74ab2e033CAS |
Morgan JA, Pataki DE, Körner C, Clark H, Del Grosso SJ, Grünzweig JM, Knapp AK, Mosier AR, Newton PCD, Niklaus PA Nippert JB, Nowak RS, Parton WJ, Polley HW, Shaw MR (2004) Water relations in grassland and desert ecosystems exposed to elevated atmospheric CO2. Oecologia 140, 11–25.
| Water relations in grassland and desert ecosystems exposed to elevated atmospheric CO2.Crossref | GoogleScholarGoogle Scholar | 1:STN:280:DC%2BD2c3oslKksQ%3D%3D&md5=c9d27de38a6982156e281ab7fb7f0441CAS | 15156395PubMed |
Muller B, Pantin F, Genard M, Turc O, Freixes S, Piques M, Gibon Y (2011) Water deficits uncouple growth from photosynthesis, increase C content, and modify the relationships between C and growth in sink organs. Journal of Experimental Botany 62, 1715–1729.
| Water deficits uncouple growth from photosynthesis, increase C content, and modify the relationships between C and growth in sink organs.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3MXjsFyjtbc%3D&md5=fe5e08ba3fb9c3e8d83c6908268f2b2fCAS | 21239376PubMed |
Neelin J, Munnich M, Su H, Meyerson J, Holloway C (2006) Tropical drying trends in global warming models and observations. Proceedings of the National Academy of Sciences of the United States of America 103, 6110–6115.
| Tropical drying trends in global warming models and observations.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD28XktFamsr8%3D&md5=28f8474a6e6c7b8e6a6c65c6ad2abb7dCAS | 16606851PubMed |
Negron-Juarez RI, Chambers JQ, Guimaraes G, Zeng H, Raupp CFM, Marra DM, Ribeiro GHPM, Saatchi SS, Nelson BW, Higuchi N (2010) Widespread Amazon forest tree mortality from a single cross-basin squall line event. Geophysical Research Letters 37, L16701
| Widespread Amazon forest tree mortality from a single cross-basin squall line event.Crossref | GoogleScholarGoogle Scholar |
Nelson BW, Kapos V, Adams JB, Oliveira WJ, Braun OPG, Doamaral IL (1994) Forest disturbance by large blowdowns in the Brazilian Amazon. Ecology 75, 853–858.
| Forest disturbance by large blowdowns in the Brazilian Amazon.Crossref | GoogleScholarGoogle Scholar |
Nepstad DC, Tohver IM, Ray D, Moutinho P, Cardinot G (2007) Mortality of large trees and lianas following experimental drought in an Amazon forest. Ecology 88, 2259–2269.
| Mortality of large trees and lianas following experimental drought in an Amazon forest.Crossref | GoogleScholarGoogle Scholar | 17918404PubMed |
Newell EA, Mulkey SS, Wright SJ (2002) Seasonal patterns of carbohydrate storage in four tropical tree species. Oecologia 131, 333–342.
| Seasonal patterns of carbohydrate storage in four tropical tree species.Crossref | GoogleScholarGoogle Scholar |
Nock CA, Baker PJ, Wanek W, Leis A, Grabner M, Bunyavejchewin S, Hietz P (2011) Long-term increases in intrinsic water-use efficiency do not lead to increased stem growth in a tropical monsoon forest in western Thailand. Global Change Biology 17, 1049–1063.
| Long-term increases in intrinsic water-use efficiency do not lead to increased stem growth in a tropical monsoon forest in western Thailand.Crossref | GoogleScholarGoogle Scholar |
Norby RJ, Zak DR (2011) Ecological lessons from free-air CO2 enrichment (FACE) experiments. Annual Review of Ecology Evolution and Systematics 42, 181–203.
| Ecological lessons from free-air CO2 enrichment (FACE) experiments.Crossref | GoogleScholarGoogle Scholar |
Norby RJ, DeLucia EH, Gielen B, Calfapietra C, Giardina CP, King JS, Ledford J, McCarthy HR, Moore DJP, Ceulemans R, De Angelis P, Finzi AC, Karnosky DF, Kubiske ME, Lukac M, Pregitzer KS, Scarascia-Mugnozza GE, Schlesinger WH, Oren R (2005) Forest response to elevated CO2 is conserved across a broad range of productivity. Proceedings of the National Academy of Sciences of the United States of America 102, 18052–18056.
| Forest response to elevated CO2 is conserved across a broad range of productivity.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2MXhtlersr3N&md5=97507f7ae6945f8f4c4c3eaf5a9765d6CAS | 16330779PubMed |
Norby RJ, Warren JM, Iversen CM, Medlyn BE, McMurtrie RE (2010) CO2 enhancement of forest productivity constrained by limited nitrogen availability. Proceedings of the National Academy of Sciences of the United States of America 107, 19 368–19 373.
| CO2 enhancement of forest productivity constrained by limited nitrogen availability.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3cXhsVGlsrfP&md5=6b48ea79355f6b04a17d2bc2642b00c5CAS |
Nottingham AT, Turner BL, Chamberlain PM, Stott AW, Tanner EVJ (2013) Priming and microbial nutrient limitation in lowland tropical forest soils of contrasting fertility. Biogeochemistry 111, 219–237.
| Priming and microbial nutrient limitation in lowland tropical forest soils of contrasting fertility.Crossref | GoogleScholarGoogle Scholar |
Oberbauer SF, Strain BR, Fetcher N (1985) Effect of CO2-enrichment on seedling physiology and growth of two tropical tree species. Physiologia Plantarum 65, 352–356.
| Effect of CO2-enrichment on seedling physiology and growth of two tropical tree species.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaL28XosFGksQ%3D%3D&md5=aa76171f0817b88cb6c97e4fc2e0a423CAS |
Osmond CB, Ananyev G, Berry J, Langdon C, Kolber Z, Lin G, Monson R, Nichol C, Rascher U, Schurr U, Smith S, Yakir D (2004) Changing the way we think about global change research: scaling up in experimental ecosystem science. Global Change Biology 10, 393–407.
| Changing the way we think about global change research: scaling up in experimental ecosystem science.Crossref | GoogleScholarGoogle Scholar |
Pan YD, Birdsey RA, Fang J, Houghton R, Kauppi PE, Kurz WA, Phillips OL, Shvidenko A, Lewis SL, Canadell JG, Ciais P, Jackson RB, Pacala SW, McGuire AD, Piao S, Rautiainen A, Stitch S, Hayes D (2011) A large and persistent carbon sink in the world’s forests. Science 333, 988–993.
| A large and persistent carbon sink in the world’s forests.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3MXhtVWrtr%2FE&md5=029f5be6f15c1249af26e500034d2975CAS |
Pavlick R, Drewry DT, Bohn K, Reu B, Kleidon A (2012) The Jena diversity-dynamic global vegetation model (JeDi-DGVM): a diverse approach to representing terrestrial biogeography and biogeochemistry based on plant functional trade-offs. Biogeosciences Discussions 9, 4627–4726.
| The Jena diversity-dynamic global vegetation model (JeDi-DGVM): a diverse approach to representing terrestrial biogeography and biogeochemistry based on plant functional trade-offs.Crossref | GoogleScholarGoogle Scholar |
Pearcy RW, Troughton J (1975) C4 photosynthesis in tree form Euphorbia species from Hawaiian rainforest sites. Plant Physiology 55, 1054–1056.
| C4 photosynthesis in tree form Euphorbia species from Hawaiian rainforest sites.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaE2MXksVSgtL8%3D&md5=f2891fd1991e31264788039fc13fab6aCAS | 16659208PubMed |
Phillips OL, Martinez RV, Arroyo L, Baker TR, Killeen T, Lewis SL, Malhi Y, Mendoza AM, Neill D, Núãez Vargas P, Alexiades M, Cerón C, Di Fiore A, Erwin T, Jardim A, Palacios W, Saldias M, Vinceti B (2002) Increasing dominance of large lianas in Amazonian forests. Nature 418, 770–774.
| Increasing dominance of large lianas in Amazonian forests.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD38XmtV2gu7g%3D&md5=c10126a8c0717d47276353ba7eee73ccCAS | 12181565PubMed |
Phillips OL, Baker TR, Arroyo L, Higuchi N, Killeen TJ, Laurance WF, Lewis SL, Lloyd J, Malhi Y, Monteagudo A, Neill DA, Núñez Vargas P, Silva JNM, Terborgh J, Vásquez Martínez R, Alexiades M, Almeida S, Brown S, Chave J, Comiskey JA, Czimczik CI, Di Fiore A, Erwin T, Kuebler C, Laurance SG, Nascimento HEM, Olivier J, Palacios W, Patiño S, Pitman NCA, Quesada CA, Saldias M, Torres Lezama A, Vinceti B (2004) Pattern and process in Amazon tree turnover, 1976–2001. Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences 359, 381–407.
| Pattern and process in Amazon tree turnover, 1976–2001.Crossref | GoogleScholarGoogle Scholar | 1:STN:280:DC%2BD2czhtVSqsQ%3D%3D&md5=e731c05c81eeda6d66d420ef6bb80929CAS | 15212092PubMed |
Phillips OL, Aragao L, Lewis SL, Fisher JB, Lloyd J, López-González G, Malhi Y, Monteagudo A, Peacock J, Quesada CA, van der Heijden G, Almeida S, Amaral I, Arroyo L, Aymard G, Baker TR, Bánki O, Blanc L, Bonal D, Brando P, Chave J, Alvea de Oliveira AC, Dávila Cardozo N, Czimczik CI, Feldpausch TR, Freitas MA, Gloor E, Higuchi N, Jiménez E, Lloyd G, Meir P, Mendoza C, Morel A, Neill DA, Nepstad D, Patiño S, Peñuela MC, Preito A, Ramírez F, Schwarz M, Silva J, Silveira M, Sota Thomas A, ter Steege H, Stropp J, Vasquéz R, Zelazowski P, Alvarez Dávila E, Andelman S, Andrade A, Chao K-J, Erwin T, Di Fiore A, Honorio Coronado E, Keeling H, Killeen TJ, Laurance WF, Peña Cruz A, Pitman NCA, Núñez Vargas P, Ramírez-Angulo H, Rudas A, Salomão R, Silva N, Terborgh J, Torres-Lezama A (2009) Drought sensitivity of the Amazon rainforest. Science 323, 1344–1347.
| Drought sensitivity of the Amazon rainforest.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD1MXisFemt7Y%3D&md5=559b3511080f93aff6f6798fa747a8e9CAS | 19265020PubMed |
Poulter B, Aragao L, Heinke J, Gumpenberger M, Heyder U, Rammig A, Thonicke K, Cramer W (2010a) Net biome production of the Amazon Basin in the 21st century. Global Change Biology 16, 2062–2075.
| Net biome production of the Amazon Basin in the 21st century.Crossref | GoogleScholarGoogle Scholar |
Poulter B, Hattermann F, Hawkins E, Zaehle S, Sitch S, Coupe NR, Heyder U, Cramer W (2010b) Robust dynamics of Amazon dieback to climate change with perturbed ecosystem model parameters. Global Change Biology 16, 2476–2495.
Prentice IC, Harrison SP, Bartlein PJ (2011) Global vegetation and terrestrial carbon cycle changes after the last ice age. New Phytologist 189, 988–998.
| Global vegetation and terrestrial carbon cycle changes after the last ice age.Crossref | GoogleScholarGoogle Scholar | 1:STN:280:DC%2BC3M7msVCgsQ%3D%3D&md5=634b4167b0a88e851d7d5b63da8583dcCAS | 21288244PubMed |
Putz FE (1984) The natural history of lianas on Barro Colorado Island, Panama. Ecology 65, 1713–1724.
| The natural history of lianas on Barro Colorado Island, Panama.Crossref | GoogleScholarGoogle Scholar |
Quesada C, Lloyd J, Schwarz M, Patiño S, Baker TR, Czimczik C, Fyllas NM, Martinelli L, Nardoto GB, Schmerler J, Santos AJB, Hodnett MG, Herrera R, Luizão FJ, Arneth A, Lloyd G, Dezzeo N, Hilke I, Kuhlmann I, Raessler M, Brand WA, Geilmann H, Moraes Filho JO, Carvalho FP, Araujo Filho RN, Chaves JE, Cruz OF, Pimentel TP, Paiva R (2010) Variations in chemical and physical properties of Amazon forest soils in relation to their genesis. Biogeosciences 7, 1515–1541.
| Variations in chemical and physical properties of Amazon forest soils in relation to their genesis.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3cXhtV2rtLrL&md5=b63efefe377e344fd577136d9ae8bac8CAS |
Quesada CA, Phillips OL, Schwarz M, Czimczik CI, Baker TR, Patiño S, Fyllas NM, Hodnett MG, Herrera R, Almeida S, Alvarez Dávila E, Arneth A, Arroyo L, Chao KJ, Dezzeo N, Erwin T, di Fiore A, Higuchi N, Honorio Coronado E, Jiménez EM, Killeen T, Torres-Lezama A, Lloyd G, López-Gonzáles G, Luizão FJ, Malhi Y, Monteagudo A, Neill DA, Núñez Vargas P, Paiva R, Peacock J, Peñuela MC, Peña Cruz A, Pitman N, Priante Filho N, Prieto A, Ramírez H, Rudas A, Salomão R, Santos AJB, Schmerler J, Silva N, Silveira M, Vásquez R, Vieira I, Terborgh J, Lloyd J (2012) Basin-wide variations in Amazon forest structure and function are mediated by both soils and climate. Biogeosciences 9, 2203–2246.
| Basin-wide variations in Amazon forest structure and function are mediated by both soils and climate.Crossref | GoogleScholarGoogle Scholar |
Rachmilevitch S, Cousins AB, Bloom AJ (2004) Nitrate assimilation in plant shoots depends on photorespiration. Proceedings of the National Academy of Sciences of the United States of America 101, 11506–11510.
| Nitrate assimilation in plant shoots depends on photorespiration.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2cXmvVKgu7s%3D&md5=8b10e0e6b695ac9eba58afa085e78f10CAS | 15272076PubMed |
Rammig A, Jupp T, Thonicke K, Tietjen B, Heinke J, Ostberg S, Lucht W, Cramer W, Cox P (2010) Estimating the risk of Amazonian forest dieback. New Phytologist 187, 694–706.
| Estimating the risk of Amazonian forest dieback.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3cXhtFSmu7jP&md5=329656e4c1801643ccc5ad84b9b64e9eCAS | 20553387PubMed |
Rasineni GK, Guha A, Reddy AR (2011) Elevated atmospheric CO2 mitigated photoinhibition in a tropical tree species, Gmelina arborea. Journal of Photochemistry and Photobiology. B, Biology 103, 159–165.
| Elevated atmospheric CO2 mitigated photoinhibition in a tropical tree species, Gmelina arborea.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3MXkvVGqt7s%3D&md5=07dc215451d8238bf26ebf95dd8e2219CAS | 21441036PubMed |
Reekie EG, Bazzaz FA (1989) Competition and patterns of resource use among seedlings of five tropical trees grown at ambient and elevated CO2. Oecologia 79, 212–222.
| Competition and patterns of resource use among seedlings of five tropical trees grown at ambient and elevated CO2.Crossref | GoogleScholarGoogle Scholar |
Richardson AE, George TS, Hens M, Simpson RJ (2005) Utilization of soil organic phosphorus by higher plants. In ‘Organic phosphorus in the environment.’. (Eds BL Turner, E Frossard, DS Baldwin) pp. 165–184. (CABI Publishing: Wallingford)
Roy J, Salager J-L (1992) Midday depression of net CO2 exchange of leaves of an emergent rain forest tree in French Guiana. Journal of Tropical Ecology 8, 499–504.
| Midday depression of net CO2 exchange of leaves of an emergent rain forest tree in French Guiana.Crossref | GoogleScholarGoogle Scholar |
Roy J, Saugier B, Mooney HA (2001) ‘Terrestrial global productivity.’ (Academic Press: San Diego)
Ryan MG (1995) Foliar maintenance respiration of sub-alpine and boreal trees and shrubs in relation to nitrogen content. Plant, Cell & Environment 18, 765–772.
| Foliar maintenance respiration of sub-alpine and boreal trees and shrubs in relation to nitrogen content.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaK2MXnsFKmurg%3D&md5=1ea573f2b71713600c7ca5973dcd2254CAS |
Saatchi SS, Harris NL, Brown S, Lefsky M, Mitchard ETA, Salas W, Zutta BR, Buermann W, Lewis SL, Hagen S, Petrova S, White L, Silman M, Morel A (2011) Benchmark map of forest carbon stocks in tropical regions across three continents. Proceedings of the National Academy of Sciences of the United States of America 108, 9899–9904.
| Benchmark map of forest carbon stocks in tropical regions across three continents.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3MXnvVymtbY%3D&md5=a46b78928f1c8354be72adb264ba2bbaCAS | 21628575PubMed |
Sage RF, Kubien DS (2007) The temperature response of C3 and C4 photosynthesis. Plant, Cell & Environment 30, 1086–1106.
| The temperature response of C3 and C4 photosynthesis.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2sXhtVeiurrP&md5=e94f29c8de8a7fc666f92a7e0859b4d0CAS |
Sala A, Piper F, Hoch G (2010) Physiological mechanisms of drought-induced tree mortality are far from being resolved. New Phytologist 186, 274–281.
| Physiological mechanisms of drought-induced tree mortality are far from being resolved.Crossref | GoogleScholarGoogle Scholar | 20409184PubMed |
Sala A, Woodruff DR, Meinzer FC (2012) Carbon dynamics in trees: feast or famine? Tree Physiology 32, 764–775.
| Carbon dynamics in trees: feast or famine?Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC38XhtFClu7fI&md5=5ebaea8041f9aff827d7003cab877dacCAS | 22302370PubMed |
Salazar LF, Nobre CA, Oyama MD (2007) Climate change consequences on the biome distribution in tropical South America. Geophysical Research Letters 34,
| Climate change consequences on the biome distribution in tropical South America.Crossref | GoogleScholarGoogle Scholar |
Sayer EJ, Heard MS, Grant HK, Marthews TR, Tanner EVJ (2011) Soil carbon release enhanced by increased tropical forest litterfall. Nature Climate Change 1, 304–307.
| Soil carbon release enhanced by increased tropical forest litterfall.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3MXhtVyltb3L&md5=6cec210b6c05f4b1474fe7afec8c8ffcCAS |
Schnitzer SA, Bongers F (2011) Increasing liana abundance and biomass in tropical forests: emerging patterns and putative mechanisms. Ecology Letters 14, 397–406.
| Increasing liana abundance and biomass in tropical forests: emerging patterns and putative mechanisms.Crossref | GoogleScholarGoogle Scholar | 21314879PubMed |
Sellers PJ, Bounoua L, Collatz GJ, Randall DA, Dazlich DA, Los SO, Berry JA, Fung I, Tucker CJ, Field CB, Jensen TG (1996) Comparison of radiative and physiological effects of doubled atmospheric CO2 on climate. Science 271, 1402–1406.
| Comparison of radiative and physiological effects of doubled atmospheric CO2 on climate.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaK28XhsFegtb8%3D&md5=6d752a4800b9f68855810b9a3aa06d51CAS |
Sitch S, Smith B, Prentice IC, Arneth A, Bondeau A, Cramer W, Kaplan JO, Levis S, Lucht W, Sykes MT, Thonicke K, Venevsky S (2003) Evaluation of ecosystem dynamics, plant geography and terrestrial carbon cycling in the LPJ dynamic global vegetation model. Global Change Biology 9, 161–185.
| Evaluation of ecosystem dynamics, plant geography and terrestrial carbon cycling in the LPJ dynamic global vegetation model.Crossref | GoogleScholarGoogle Scholar |
Solomon S, Qin D, Manning M, Chen Z, Marquis M, Averyt KB, Tignor M, Miller HL (Eds) (2007) ‘Climate change 2007: the physical science basis. Contribution of Working Group I to the Fourth Assessment Report of the Intergovermental Panel on Climate Change.’ (Cambridge University Press: Cambridge, UK)
Sprent JI (2009) ‘Legume nodulation: a global perspective.’ (Wiley-Blackwell: Oxford, UK)
Sprugel DG, Hinckley TM, Schaap W (1991) The theory and practice of branch autonomy. Annual Review of Ecology and Systematics 22, 309–334.
| The theory and practice of branch autonomy.Crossref | GoogleScholarGoogle Scholar |
Stork NE, Balston J, Farquhar GD, Franks PJ, Holtum JAM, Liddell MJ (2007) Tropical rainforest canopies and climate change. Austral Ecology 32, 105–112.
| Tropical rainforest canopies and climate change.Crossref | GoogleScholarGoogle Scholar |
Strain BR, Bazzaz FA (1983) Terrestrial plant communities. In ‘CO2 and plants’. (Ed. ER Lemon) pp. 177–122. (Westview: Boulder, CO)
Swaine MD, Whitmore TC (1988) On the definition of ecological species groups in tropical rain forests. Vegetatio 75, 81–86.
| On the definition of ecological species groups in tropical rain forests.Crossref | GoogleScholarGoogle Scholar |
Tanner EVJ, Vitousek PM, Cuevas E (1998) Experimental investigation of nutrient limitation of forest growth on wet tropical mountains. Ecology 79, 10–22.
| Experimental investigation of nutrient limitation of forest growth on wet tropical mountains.Crossref | GoogleScholarGoogle Scholar |
ter Steege H, Pitman NCA, Phillips OL, Chave J, Sabatier D, Duque A, Molino J-F, Prévost M-F, Spichiger R, Castellanos H, von Hildebrand P, Vásquez R (2006) Continental-scale patterns of canopy tree composition and function across Amazonia. Nature 443, 444–447.
| Continental-scale patterns of canopy tree composition and function across Amazonia.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD28XhtVSns7fL&md5=1de3862aace756a5f912de6ce5458791CAS | 17006512PubMed |
Thomas RB, Richter DD, Ye H, Heine PR, Strain BR (1991) Nitrogen dynamics and growth of seedlings of an N-fixing tree (Gliricidia sepium (Jacq.) Walp.) exposed to elevated atmospheric carbon dioxide. Oecologia 88, 415–421.
| Nitrogen dynamics and growth of seedlings of an N-fixing tree (Gliricidia sepium (Jacq.) Walp.) exposed to elevated atmospheric carbon dioxide.Crossref | GoogleScholarGoogle Scholar |
Timmermann A, Oberhuber J, Bacher A, Esch M, Latif M, Roeckner E (1999) Increased El Niño frequency in a climate model forced by future greenhouse warming. Nature 398, 694–697.
| Increased El Niño frequency in a climate model forced by future greenhouse warming.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaK1MXivVegtrc%3D&md5=86102392717cc4be9f37e3bf68283c3bCAS |
Tissue DT, Megonigal JP, Thomas RB (1997) Nitrogenase activity and N2 fixation are stimulated by elevated CO2 in a tropical N2-fixing tree. Oecologia 109, 28–33.
| Nitrogenase activity and N2 fixation are stimulated by elevated CO2 in a tropical N2-fixing tree.Crossref | GoogleScholarGoogle Scholar |
Tolley LC, Strain BR (1984) Effects of CO2 enrichment and water sress on growth of Liquidamber styraciflua and Pinus taeda seedlings. Canadian Journal of Botany 62, 2135–2139.
| Effects of CO2 enrichment and water sress on growth of Liquidamber styraciflua and Pinus taeda seedlings.Crossref | GoogleScholarGoogle Scholar |
Turner BL (2008) Resource partitioning for soil phosphorus: a hypothesis. Journal of Ecology 96, 698–702.
| Resource partitioning for soil phosphorus: a hypothesis.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD1cXptFygs70%3D&md5=d2d3f6f01993fc7c7c05f72ea671a8e8CAS |
Turner BL, Engelbrecht BMJ (2011) Soil organic phosphorus in tropical rainforests. Biogeochemistry 103, 297–315.
| Soil organic phosphorus in tropical rainforests.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3MXisVKiu70%3D&md5=9fc2a1857420a40bf2fc1fdbba426554CAS |
Verbeeck H, Peylin P, Bacour C, Bonal D, Steppe K, Ciais P (2011) Seasonal patterns of CO2 fluxes in Amazon forests: fusion of eddy covariance data and the ORCHIDEE model. Journal of Geophysical Research 116,
| Seasonal patterns of CO2 fluxes in Amazon forests: fusion of eddy covariance data and the ORCHIDEE model.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC38XnsVSku7s%3D&md5=f1525c3855bacd66cd52ae59fc8ac44cCAS |
Vincent AG, Turner BL, Tanner EVJ (2010) Soil organic phosphorus dynamics following perturbation of litter cycling in a tropical moist forest. European Journal of Soil Science 61, 48–57.
| Soil organic phosphorus dynamics following perturbation of litter cycling in a tropical moist forest.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3cXhvFajs70%3D&md5=5953d2b6fa58202ecf725c39cbb7c303CAS |
Vitousek PM (1984) Litterfall, nutrient cycling, and nutrient limitation in tropical forests. Ecology 65, 285–298.
| Litterfall, nutrient cycling, and nutrient limitation in tropical forests.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaL2cXhtlOqu7c%3D&md5=1407402cb91e53881f418d955d49fcaeCAS |
Vitousek PM, Porder S, Houlton BZ, Chadwick OA (2010) Terrestrial phosphorus limitation: mechanisms, implications, and nitrogen-phosphorus interactions. Ecological Applications 20, 5–15.
| Terrestrial phosphorus limitation: mechanisms, implications, and nitrogen-phosphorus interactions.Crossref | GoogleScholarGoogle Scholar | 20349827PubMed |
von Caemmerer S, Farquhar GD (1981) Some relationships between the biochemistry of photosynthesis and the gas exchange of leaves. Planta 153, 376–387.
| Some relationships between the biochemistry of photosynthesis and the gas exchange of leaves.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaL38XjtFyjug%3D%3D&md5=3387f565dfaf023128aa42635b34a338CAS |
Wang YP, Houlton BZ, Field CB (2007) A model of biogeochemical cycles of carbon, nitrogen, and phosphorus including symbiotic nitrogen fixation and phosphatase production. Global Biogeochemical Cycles 21, GB1018
| A model of biogeochemical cycles of carbon, nitrogen, and phosphorus including symbiotic nitrogen fixation and phosphatase production.Crossref | GoogleScholarGoogle Scholar |
Way DA, Ladeau SL, McCarthy HR, Clark JS, Oren R, Finzi AC, Jackson RB (2010) Greater seed production in elevated CO2 is not accompanied by reduced seed quality in Pinus taeda L. Global Change Biology 16, 1046–1056.
| Greater seed production in elevated CO2 is not accompanied by reduced seed quality in Pinus taeda L.Crossref | GoogleScholarGoogle Scholar |
Wiley E, Helliker B (2012) A re-evaluation of carbon storage in trees lends greater support for carbon limitation to growth. New Phytologist 195, 285–289.
| A re-evaluation of carbon storage in trees lends greater support for carbon limitation to growth.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC38XosFSntb4%3D&md5=1a28156c382e86df4fe3abdfeb91f041CAS | 22568553PubMed |
Williamson G, Laurance W, Oliveira A, Delamonica P, Gascon C, Lovejoy T, Pohl L (2000) Amazonian tree mortality during 1997 El Niño drought. Conservation Biology 14, 1538–1542.
| Amazonian tree mortality during 1997 El Niño drought.Crossref | GoogleScholarGoogle Scholar |
Winter K, Garcia M, Lovelock CE, Gottsberger R, Popp M (2000) Responses of model communities of two tropical tree species to elevated atmospheric CO2: growth on unfertilized soil. Flora 195, 289–302.
Winter K, Aranda J, Garcia M, Virgo A, Paton SR (2001a) Effect of elevated CO2 and soil fertilization on whole-plant growth and water use in seedlings of a tropical pioneer tree, Ficus insipida Willd. Flora 196, 458–464.
Winter K, Garcia M, Gottsberger R, Popp M (2001b) Marked growth response of communities of two tropical tree species to elevated CO2 when soil nutrient limitation is removed. Flora 196, 47–58.
Wong S-C, Cowan IR, Farquhar GD (1979) Stomatal conductance correlates with photosynthetic capacity. Nature 282, 424–426.
| Stomatal conductance correlates with photosynthetic capacity.Crossref | GoogleScholarGoogle Scholar |
Wright SJ, Calderon O (2006) Seasonal, El Niño and longer term changes in flower and seed production in a moist tropical forest. Ecology Letters 9, 35–44.
Wright SJ, Calderon O, Hernandez A, Paton S (2004) Are lianas increasing in importance in tropical forests? A 17-year record from Panama. Ecology 85, 484–489.
| Are lianas increasing in importance in tropical forests? A 17-year record from Panama.Crossref | GoogleScholarGoogle Scholar |
Wright SJ, Muller-Landau HC, Calderon O, Hernandez A (2005) Annual and spatial variation in seedfall and seedling recruitment in a neotropical forest. Ecology 86, 848–860.
| Annual and spatial variation in seedfall and seedling recruitment in a neotropical forest.Crossref | GoogleScholarGoogle Scholar |
Wright SJ, Hernandez A, Condit R (2007) The bushmeat harvest alters seedling banks by favoring lianas, large seeds, and seeds dispersed by bats, birds, and wind. Biotropica 39, 363–371.
| The bushmeat harvest alters seedling banks by favoring lianas, large seeds, and seeds dispersed by bats, birds, and wind.Crossref | GoogleScholarGoogle Scholar |
Wright SJ, Kitajima K, Kraft NJB, Reich PB, Wright IJ, Bunker DE, Condit R, Dalling JW, Davies SJ, Díaz S, et al (2010) Functional traits and the growth-mortality trade-off in tropical trees. Ecology 91, 3664–3674.
| Functional traits and the growth-mortality trade-off in tropical trees.Crossref | GoogleScholarGoogle Scholar | 21302837PubMed |
Wright SJ, Yavitt JB, Wurzburger N, Turner BL, Tanner EVJ, Sayer EJ, Santiago LS, Kaspari M, Hedin LO, Harms KE, Garcia MN, Corre MD (2011) Potassium, phosphorus, or nitrogen limit root allocation, tree growth, or litter production in a lowland tropical forest. Ecology 92, 1616–1625.
| Potassium, phosphorus, or nitrogen limit root allocation, tree growth, or litter production in a lowland tropical forest.Crossref | GoogleScholarGoogle Scholar | 21905428PubMed |
Wullschleger SD, Tschaplinski TJ, Norby RJ (2002) Plant water relations at elevated CO2 – implications for water-limited environments. Plant, Cell & Environment 25, 319–331.
| Plant water relations at elevated CO2 – implications for water-limited environments.Crossref | GoogleScholarGoogle Scholar |
Würth MKR, Winter K, Körner C (1998a) In situ responses to elevated CO2 in tropical forest understorey plants. Functional Ecology 12, 886–895.
| In situ responses to elevated CO2 in tropical forest understorey plants.Crossref | GoogleScholarGoogle Scholar |
Würth MKR, Winter K, Körner C (1998b) Leaf carbohydrate responses to CO2 enrichment at the top of a tropical forest. Oecologia 116, 18–25.
Würth MKR, Pelaez-Riedl S, Wright SJ, Körner C (2005) Non-structural carbohydrate pools in a tropical forest. Oecologia 143, 11–24.
| Non-structural carbohydrate pools in a tropical forest.Crossref | GoogleScholarGoogle Scholar |
Zachos JC, Wara MW, Bohaty S, Delaney ML, Petrizzo MR, Brill A, Bralower TJ, Premoli-Silva I (2003) A transient rise in tropical sea surface temperature during the Paleocene–Eocene thermal maximum. Science 302, 1551–1554.
| A transient rise in tropical sea surface temperature during the Paleocene–Eocene thermal maximum.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD3sXpt1SmsL8%3D&md5=a3d86d365423730e0f952eda38a65727CAS | 14576441PubMed |
Ziska LH, Hogan KP, Smith AP, Drake BG (1991) Growth and photosynthetic response of 9 tropical species with long-term exposure to elevated carbon dioxide. Oecologia 86, 383–389.
| Growth and photosynthetic response of 9 tropical species with long-term exposure to elevated carbon dioxide.Crossref | GoogleScholarGoogle Scholar |
Zotz G, Harris G, Königer M, Winter K (1995) High rates of photosynthesis in the tropical pioneer tree, Ficus insipida Willd. Flora 190, 265–272.
Zotz G, Cueni N, Körner C (2006) In situ growth stimulation of a temperate zone liana (Hedera helix) in elevated CO2. Functional Ecology 20, 763–769.
| In situ growth stimulation of a temperate zone liana (Hedera helix) in elevated CO2.Crossref | GoogleScholarGoogle Scholar |