

Climate change and its implications for Australia’s freshwater fish
John R. Morrongiello A B M N , Stephen J. Beatty C , James C. Bennett D E , David A. Crook B F , David N. E. N. Ikedife D , Mark J. Kennard G H , Adam Kerezsy H I , Mark Lintermans J , Dale G. McNeil B K , Bradley J. Pusey G H and Thomas Rayner LA School of Biological Sciences, Monash University, Clayton, Vic. 3800, Australia.
B eWater Cooperative Research Centre, Canberra, ACT 2601, Australia.
C Centre for Fish, Fisheries and Aquatic Ecosystem Research, School of Biological Sciences, Murdoch University, Murdoch, WA 6150, Australia.
D Entura-Hydro Tasmania, Hobart, Tas. 7001, Australia.
E Antarctic Climate and Ecosystems Cooperative Research Centre, University of Tasmania, Sandy Bay, Tas. 7005, Australia.
F Department of Sustainability and Environment, Arthur Rylah Institute for Environmental Research, Heidelberg, Vic. 3084, Australia.
G Tropical Rivers and Coastal Knowledge Commonwealth Environmental Research Facility, Griffith University, Nathan, Qld 4111, Australia.
H Australian Rivers Institute, Griffith University, Brisbane, Nathan, Qld 4111, Australia.
I Bush Heritage Australia, Melbourne, Vic. 3000, Australia.
J Institute for Applied Ecology, University of Canberra, Canberra, ACT 2601, Australia.
K South Australian Research and Development Institute, Aquatic Sciences, Henley Beach, SA 5022, Australia.
L Australian Wetlands and Rivers Institute, the University of New South Wales, Sydney, NSW 2052, Australia.
M Present address: CSIRO Marine and Atmospheric Research, GPO Box 1538, Hobart, Tas. 7001, Australia.
N Corresponding author. Email: john.morrongiello@csiro.au
Marine and Freshwater Research 62(9) 1082-1098 https://doi.org/10.1071/MF10308
Submitted: 7 December 2010 Accepted: 8 May 2011 Published: 21 September 2011
Journal Compilation © CSIRO Publishing 2011 Open Access CC BY-NC-ND
Abstract
Freshwater environments and their fishes are particularly vulnerable to climate change because the persistence and quality of aquatic habitat depend heavily on climatic and hydrologic regimes. In Australia, projections indicate that the rate and magnitude of climate change will vary across the continent. We review the likely effects of these changes on Australian freshwater fishes across geographic regions encompassing a diversity of habitats and climatic variability. Commonalities in the predicted implications of climate change on fish included habitat loss and fragmentation, surpassing of physiological tolerances and spread of alien species. Existing anthropogenic stressors in more developed regions are likely to compound these impacts because of the already reduced resilience of fish assemblages. Many Australian freshwater fish species are adapted to variable or unpredictable flow conditions and, in some cases, this evolutionary history may confer resistance or resilience to the impacts of climate change. However, the rate and magnitude of projected change will outpace the adaptive capacities of many species. Climate change therefore seriously threatens the persistence of many of Australia’s freshwater fish species, especially of those with limited ranges or specific habitat requirements, or of those that are already occurring close to physiological tolerance limits. Human responses to climate change should be proactive and focus on maintaining population resilience through the protection of habitat, mitigation of current anthropogenic stressors, adequate planning and provisioning of environmental flows and the consideration of more interventionist options such as managed translocations.
Additional keywords: arid zone, drought, environmental flows, freshwater fish, life history, local adaptation, refugia, threatened species.
Introduction
Climate change is a major threat to global biodiversity and ecosystem functioning (Thomas et al. 2004; IPCC 2007) and its effects are already evident across a range of environments and biota (Parmesan and Yohe 2003). The rate and severity of climate changes have, and will, vary across the globe, with particular regions, ecosystems and taxa being differentially affected depending on their susceptibility and level of exposure. Freshwater environments, and the organisms that inhabit them, are particularly vulnerable because they are isolated and fragmented within a terrestrial landscape (Fausch et al. 2002). Furthermore, surface water, which determines the quality and availability of aquatic habitat, depends heavily on rainfall and temperature regimes that will be drastically affected by climate change (Carpenter et al. 1992; Hobday and Lough 2011). Relatively few studies have explored the implications of climate change for freshwater biota, with the majority of these focussing on northern hemisphere or high-latitude freshwaters (e.g. Xenopoulos and Lodge 2006; Buisson et al. 2008; Graham and Harrod 2009; Heino et al. 2009; McCullough et al. 2009; but see Carpenter et al. 1992, for global perspectives, and Chessman 2009 and Kingsford et al. 2011, for Australian perspectives). Projected global warming that surpasses or optimises thermal tolerances and requirements is often emphasised as a major driver of assemblage turnover, range shifts and range expansions in these systems because surface water is seldom limiting (but see Xenopoulos and Lodge 2006, for hydrological example). Much less, however, is known of how climate change will affect freshwaters in arid and semiarid regions where surface water is already scarce and likely to become more so (Bates et al. 2008).
Australia encompasses a diversity of climates and geography. Its aquatic biota inhabits a broad range of freshwater environments, spanning the naturally variable and unpredictable hydrology of arid and semiarid regions that cover much of the continent, to the highly seasonal yet predictable flows of tropical regions and the more stable and consistent baseflow regimes of some temperate and subalpine regions (Puckridge et al. 1998; Kennard et al. 2010). Australia’s freshwater fish species (~206 native species Allen et al. 2002), consequently, exhibit a diverse array of reproductive, morphological and physiological adaptations that facilitate persistence in particular environmental circumstances (e.g. Humphries et al. 1999; Pusey et al. 2004; Crook et al. 2010a). However, the extent to which current populations and assemblages will persist into the future remains unclear. Never before have freshwater fish faced such a magnitude and rate of climate change, coupled with the added pressure of human disturbance (reviewed in Dudgeon et al. 2006). It is clear that the projected reductions in surface-water availability resulting from climate change, despite high uncertainty associated with greenhouse gas-scenario selection and model downscaling (CSIRO and Bureau of Meteorology 2007; Hobday and Lough 2011), pose a significant threat to the viability of freshwater fish populations in many regions of Australia.
At present, information regarding the nature and severity of this threat is fragmented and largely incidental or anecdotal. The aim of the present review is to synthesise this existing information and begin predicting the likely impacts of climate change on Australia’s freshwater fish species (Fig. 1). We conduct regional assessments to explore differences and commonalities in potential fish responses to climate change across the continent, and discuss these in the context of existing anthropogenic stressors. We also consider the potential for species to adapt to projected conditions and discuss management options to some of the issues raised. The review will provide a contextual background to facilitate further research through the identification of key knowledge gaps and will assist managers in forming proactive and effective responses to the threats posed by climate change.

|
Regional comparisons
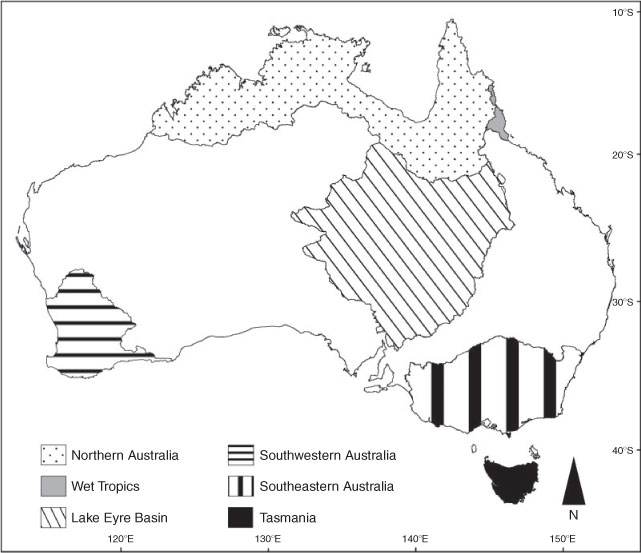
Across Australia, significant climatic changes have been observed over the past 50 years (CSIRO and Bureau of Meteorology 2007). Average annual air temperatures have risen by 0.9°C, which is faster than the global average (Lough et al. 2011), and rainfall patterns have changed such that some regions are experiencing significant deficits and others increased variability. Droughts are also becoming more severe and less precipitation is falling as snow. Although there are some general patterns associated with climate change, the impacts on hydrology and thus aquatic fauna have not been, and will not be, uniform across the continent (Fig. 2; CSIRO and Bureau of Meteorology 2007; Hobday and Lough 2011; Lough et al. 2011). In the following sections, we explore the potential impacts of climate change on freshwater fish across six representative geographic regions of Australia (Table 1, Fig. 3) that loosely correspond to major drainage divisions, hydrological regimes (Haines et al. 1988; Kennard et al. 2010) and biogeographical provinces of freshwater fish (Unmack 2001). The implications of climate change in the Murray–Darling Basin are considered in more detail by others (Aldous et al. 2011; Kingsford et al. 2011; Pittock and Finlayson 2011; Balcombe et al. 2011; Pratchett et al. 2011).

|

|
|
Northern Australia
The freshwaters of northern Australia are diverse, ranging from the complex floodplain-river and billabong systems in the Gulf of Carpentaria, to deeply incised bedrock-controlled rivers of the Kimberley region and geographically isolated streams on upland plateaus. Flow regimes are summer-dominated and vary according to the extent of seasonality, predictability and degree of flow permanence (Kennard et al. 2010). Northern Australia, as defined here, constitutes only 17% of the continental area, yet contains ~60% of Australia’s freshwater-fish biodiversity (Unmack 2001; Pusey et al. 2004). Although most of the floodplain ecosystems are in good ecological condition and reflect the integrity of the surrounding savanna and limited extent of past water-resource development (Woinarski et al. 2007; Pusey and Kennard 2009), the fish fauna of the region is increasingly threatened by hydrological alteration and a range of diffuse threats.
The precise nature of changes in northern Australia’s rainfall and runoff under various climate scenarios has been notoriously difficult to quantify with high certainty (Cresswell et al. 2009). In general, however, the projected trends of slightly reduced discharge, a minor increase in the number of days of zero flow, and increased rates of evapotranspiration (Table 1) suggest that the current trend for increasing intermittency in inland river reaches will intensify. More intense cyclones and individual rainfall events are likely to steepen the flood hydrograph and increase recharge rates of shallow groundwater aquifers (Cresswell et al. 2009). The impacts of these minor hydrological changes on the freshwater fish species of the region are difficult to predict, except that any reduction in dry-season flows may potentially exacerbate droughts, reduce the availability of flowing habitats (e.g. riffle habitats used by members of the Terapontidae (grunters) as nursery areas), decrease the extent to which migratory species may move within individual river systems (Chan et al. 2011), and decrease the number of refugial habitats if spell lengths exceed refugia permanence. Further, increased rainfall intensity, unaccompanied by an overall substantial increase in total rainfall, may mean that flood events are shorter in duration. This may result in intermittent tributary streams that flow during floods being less likely to provide suitable spawning habitat for a sufficient time to allow hatching, development and migration back to permanent water for species such as plotosid catfishes.
Northern Australia’s coastal freshwater floodplains are extensive (~30% of the region’s area) and occur close to the current sea level (D. Ward, pers. comm.); thus, they are vulnerable to sea-level rise. For example, the extensive coastal wetlands of Kakadu are only 0.2–1.2 m above the mean high water level (Eliot et al. 1999), which is within the bounds of projected sea-level rises of ~0.3 m by 2030 (Table 1). Likewise, increased intensity of cyclones and associated storm surges further increases the likelihood of saltwater intrusion into coastal freshwater wetlands, with the attendant risk of floristic change (e.g. loss of Melaleuca forests and upstream migration of mangroves) and geomorphological transition from freshwater wetlands to saline mudflats (Eliot et al. 1999). The transition of wetlands to saline mudflats would occur rapidly (Woodroffe 1995) and locally extirpate wetland-dependent species such as pennyfish (Denariusa bandata) and blue-eyes (Pseudomugil tennellus and P. gertrudae) and potentially reduce population sizes of many other species, including the diadromous barramundi (Lates calcarifer) for which coastal floodplain wetlands are an important nursery habitat. A shift from a wetland to saline mudflat habitat would have widespread effects on primary production, foodwebs, fish-assemblage structure and overall diversity.
Mean annual air temperatures in northern Australia already range from 30°C to 33°C, and maximum instantaneous water temperatures frequently exceed this range (e.g. >37°C), particularly during the dry season (Pusey et al. 2004; Hamilton 2010). Projected warming in northern Australia, coupled with increased intermittency in rivers that regularly cease to flow (Cresswell et al. 2009; Kennard et al. 2010), may result in refugial water temperatures exceeding the physiological tolerances, already near their lethal limit, of many species. Although altered thermal regimes might cause a contraction in the distributions of some species in northern Australia, others may experience geographic range shifts and expansions when dispersal is not restricted by barriers. For example, the broad-scale distribution of spangled perch (Leiopotherapon unicolor) was suggested to be limited by the 4°C winter isotherm (Llewellyn 1973), whereas more recent research has extended the distribution further south (Schiller et al. 1997). Whether this is due to a shift southward of the limiting isotherm, or extension beyond this thermal limit, is unknown. However, range shifts in many other species restricted to this region may be constrained, given the presence of arid zones to the immediate south and the presence of the Great Dividing Range in the east.
Importantly, the generally low levels of human development, near-natural condition of the catchments and flow regimes coupled with the few artificial barriers to fish movement and dispersal mean rivers of northern Australia provide a circumstance where the native fauna is less constrained by human factors in its potential capacity to adjust to system-wide changes as a result of global climate change.
Wet Tropics
Australia’s Wet Tropics region is confined to a small and isolated coastal strip in the continent’s north-east (Fig. 3) characterised by forested mountain ranges in the upper reaches and cleared alluvial floodplains in the lowlands (Russell et al. 1996). The current climate is highly seasonal, with low interannual variability (Table 1; Kennard et al. 2010). During the monsoonal summer months (December–March), air temperature, rainfall and floodplain inundation are at their peaks, whereas stable baseflows extend through the dry season (April–November), maintained by rainfall-recharged groundwater and cloud capture in upland rainforests (McJannet et al. 2007). The Wet Tropics region hosts a highly diverse and endemic freshwater fish fauna (~107 species, ~50% of Australia’s freshwater fish biodiversity), including representatives from 37 families (Pusey et al. 2008). These families can be divided into the following two broad groups: those distinguished by low within-family generic and specific diversity (i.e. one species per family, e.g. Scorpaenidae (bullrout)), and those characterised by higher specific or generic diversity and containing endemics (e.g. Melanotaeniidae (rainbowfishes)). Most of the fish biodiversity is found on the narrow coastal plain, in short, lowland main-channel sections of rivers, owing to natural and anthropogenic instream barriers that limit upstream movement (Pusey and Kennard 1996). However, upland streams are important reservoirs of endemic species because of their prolonged isolation and persistence as refugia through past glacial maxima (Schneider and Moritz 1999).
Climate-change modelling suggests that warming and rainfall changes will be less pronounced on the north-eastern coast than elsewhere in Australia (CSIRO and Bureau of Meteorology 2007). However, the effects of increased flow seasonality and duration of dry periods (Table 1) is likely to result in greater upstream tidal penetration, longer water-residence times and increased water temperatures (Rayner et al. 2008). Increased water extraction for human use also has the potential to exacerbate these effects, with implications for water quality, habitat availability, riparian vegetation structure, instream productivity and ultimately fish assemblages (Pusey et al. 2008; Rayner et al. 2009). For example, climate-induced alterations to the structure of riparian vegetation communities could affect the availability and consumption of riparian fruits by khaki grunter (Hephaestus tulliensis) (Rayner et al. 2009). Increases in water-residence time, instream temperature and autochthonous production could advantage alien species such as black mangrove cichlid (Tilapia mariae) that consume filamentous algae – a niche that appears under-utilised by native species (Rayner et al. 2009). Increases in the intensity of cyclones and rainfall events will result in a concurrent change in flood-disturbance regimes (Eliot et al. 1999; Williams et al. 2003), and disrupt the seasonal cycle of fish-assemblage structure and function (T. Rayner, unpubl. data).
Cyclonic storm surges will exacerbate the effects of saltwater intrusion associated with sea-level rise (CSIRO and Bureau of Meteorology 2007). Projected sea-level rises are likely to have analogous impacts on the habitat and fishes of the Wet Tropics to those associated with past sea-level rises (e.g. during the Holocene) where between 50 and 400 km of main channel habitat in each catchment was lost (Chivas et al. 2001; Yokoyama et al. 2001). For example, the lesser salmon catfish (Arius graeffei) is not found in the Wet Tropics, whereas it is present in all surrounding systems, where long, low-gradient main channels were maintained during past climate-change events (Pusey et al. 2004). In contrast, the presence of high mountains in the region is likely to have buffered fishes from past climate-change events by maintaining flows through cloud capture, especially in small streams. This hypothesis is consistent with the work of Tedesco et al. (2005) who found the highest fish species richness in Central and South American and West African drainages that were connected to rainforest refugia during the last glacial maximum. Increases in the basal elevation of orographic cloud formation of 100 m per degree of warming are predicted, with major implications for flow volumes in upland tropical streams (Still et al. 1999). Fish species that are restricted to shallow riffle habitats (e.g. Allen’s cling-goby (Stiphodon alleni), scaleless goby (Schismatogobius sp.) and Glossogobius bellendenensis; Pusey et al. 2004) are particularly vulnerable to extended periods of low rainfall, because their habitats are likely to experience more frequent and extreme drying. Never before have Wet Tropics fish faced the challenge of rapid climate change with the added pressure of human disturbance.
Lake Eyre Basin
Riverine systems in the arid and semiarid interior of Australia are characterised by unpredictable and highly variable hydrology (Puckridge et al. 1998). Consequently, rivers persist as a chain of isolated waterholes for most of the time. In the major catchments, such as the Diamantina/Warburton, Georgina and Cooper, flow and flood events generally occur in summer and occasionally fill large terminal lakes, such as Lake Eyre. The 23 native and 2 alien fish species present in central Australian rivers often have life-history characteristics adapted for these variable habitats (Pusey et al. 2004; Balcombe et al. 2006; Kerezsy 2010), with the majority of species reproducing throughout the year in permanent waterholes (Balcombe and Arthington 2009). Many fish species from central Australian waterways are also capable of migrating long distances (at least 300 km in the case of L. unicolor) in ephemeral desert systems when flow and connection pathways are re-instated (Kerezsy 2010). Additionally, some species such as desert goby (Chlamydogobius eremius) and Lake Eyre hardyhead (Craterocephalus eyresii) possess specific traits, such as tolerance to hypersaline and hypoxic conditions and the ability to take up oxygen from the air, that enable them to persist in arid environments (Thompson and Withers 2002; McNeil and Schmarr 2009).
Given that arid-zone fish must persist in isolated waterholes for the majority of the time, any prolonged drying of central Australia that exceeds waterhole permanence thresholds would reduce ranges for species in systems where there is already little surface water (Silcock 2009). In extreme cases, such as the Neales catchment in South Australia where only a single permanent waterhole exists (McNeil and Schmarr 2009), prolonged drying could result in catchment-wide extinction of all species. Although most arid-zone fish species have a wide distribution, some are range-limited, and these species would obviously be most at risk if catchments experienced increased drying. Examples include the endemic Cooper Creek catfish (Neosiluroides cooperensis) and arid-zone populations of more widespread species such as Australian smelt (Retropinna semoni), carp gudgeon (Hypseleotris spp.), barred grunter (Amniataba percoides) and golden goby (Glossogobius aureus). It is important, however, to acknowledge the role of natural environmental variability with, for example, an ephemeral catchment such as the Mulligan containing no water in a dry year and at least seven species of fish following a flood (Kerezsy 2010). Consequently, determining the precise impact of climate change (as opposed to the overall impact of on-going climatic variability) on these arid riverine systems is likely to be difficult.
In contrast, the responses of isolated fish populations in spring complexes to a reduction in the availability of artesian water, associated with less rainfall and groundwater recharge (Fairfax et al. 2007), are more predictable. These include range reductions and possible extinction of endangered and endemic species in springs such as Dalhousie in South Australia and Edgbaston and Elizabeth Springs in Queensland. Undoubtedly, the species at most risk from spring drying include the red-finned blue eye (Scaturiginichthys vermeilipinnis), which is currently present in only four springs at Edgbaston in western Queensland (Fairfax et al. 2007), the Dalhousie endemics (Neosilurus sp., Cratocephalus dalhousiensis, Chlamydogobius sp.) and the Flinders Ranges gudgeon (Mogurnda clivicola), which is restricted to two springs in the Northern Flinders Ranges (McNeil et al. 2010). These fish species, however, currently face more immediate threats such as alien species (Kerezsy 2009), agricultural development and disease (McNeil and Schmarr 2009). Additionally, water-resource development in the Lake Eyre Basin is likely to increase under drying climatic conditions, with the possible exploitation of artesian spring resources posing a catastrophic threat to the persistence of these groundwater-dependent species.
South-western Australia
The Mediterranean climate of south-western Australia is characterised by highly seasonal rainfall (Nicholls et al. 1997), resulting in seasonal stream flow and significant periods of natural intermittency. As a consequence, the aquatic fauna has evolved under a regime of seasonal inundation of wetlands and annual periods of no-flow in rivers (Bunn et al. 1986). However, since 1975, a climatic phase shift has seen significant reductions in rainfall and stream flows (Table 1) (CSIRO 2009a). Furthermore, 80–90% of native vegetation has been cleared for agriculture (Halse et al. 2003), resulting in >70% of the Australian continent’s secondary salinised areas. Now only ~44% of flow in the largest 30 rivers is fresh (Mayer et al. 2005). These current stressors have already caused reductions in the range of aquatic fauna from upstream areas of catchments (e.g. Morgan et al. 1998; Halse et al. 2003; Davies 2010) and the greatest diversity of freshwater fishes is now found in less-degraded, forested systems (Morgan et al. 1998).
Although this region hosts a relatively low-diversity native fish assemblage (10 native, 10 alien species), 80% of these natives are endemic (Morgan et al. 1998). Many species have highly restricted distributions; e.g. salamanderfish (Lepidogalaxias salamandroides), black-stripe minnow (Galaxiella nigrostriata) and mud minnow (G. munda) are found almost exclusively in seasonal peat-wetland habitats of the far south-west (Pusey and Edward 1990; Morgan et al. 2000). Endemic freshwater fish species of the region breed during high flow periods in winter and spring (Allen et al. 2002) and this is the period projected to continue to experience substantial declines in rainfall and discharge (CSIRO 2009a). Many species also move into seasonally inundated tributary or wetland habitats to spawn (e.g. Pen and Potter 1991; Beatty et al. 2009). Durational suitability of these habitats has already declined since 1975 and will continue to decline in the face of an additional 25% reduction in annual flow (CSIRO 2009a). The projected additional decreases in rainfall, stream flow and groundwater recharge (CSIRO 2009a, 2009b), and increases in extreme weather events associated with climate change (Table 1), coupled with increased human exploitation of water resources (CSIRO 2009a, 2009b) will only exacerbate current impacts on an already stressed system.
Substantial reductions in groundwater levels will have major implications for habitat availability. For example, groundwater-derived baseflow in the Blackwood River is important in maintaining access to feeding areas (riffles) for freshwater cobbler (Tandanus bostocki), with a reduction of just 8% in baseflow discharge predicted to prevent access to riffle zones (Beatty et al. 2010b). Further reductions in rainfall will potentially prevent access to these habitats because baseflow magnitude depends on the previous year’s rainfall (Golder and Associates 2008). Likewise, projected reductions in groundwater discharge may result in the loss of tributaries as freshwater refugia during baseflow periods (Beatty et al. 2009). Further rainfall and groundwater reductions in the northern region of south-western Australia will threaten the habitats of outlying populations of both Galaxiella species (300 km north of common range Morgan et al. 1998; Beatty et al. 2010a) which are already under pressure from recent flow reductions (Durrant 2009). Reductions in rainfall, delayed onset of the wet season and lowered groundwater levels are likely to seriously threaten the two aestivating species, namely L. salamandroides and G. nigrostriata (Pusey and Edward 1990). Both species are very small, with limited energetic capacity to persist for many months in the dormant condition; prolongation of this state increases mortality and has serious consequences for subsequent reproductive viability after the onset of winter rains (Pusey 1990).
Salinisation of lotic systems has already altered fish assemblages, causing upstream colonisation by estuarine species and downstream contraction of more sensitive species into remaining fresher habitats (Beatty et al. 2009; Beatty et al. 2010a). Along with maintaining habitat connectivity, fresh groundwater discharge buffers salinities in the main channel of the Blackwood River (Beatty et al. 2010b), resulting in several fishes utilising only the main channel during baseflow periods when most freshwater tributaries cease to flow (Beatty et al. 2009). Although the relationship between future rainfall reductions and trends in salinisation of rivers throughout this region is largely unknown (Mayer et al. 2005), further reductions in fresh groundwater discharge have the potential to result in the loss of main channel habitat for freshwater fish in many south-western rivers. Some species, such as western minnow (Galaxias occidentalis), may be resilient to additional hydrological changes compared with other fish species that have undergone drastic range declines or have highly specialised habitat requirements. This fish has an acute salinity tolerance of 14 g L–1 (Beatty et al. 2011) and continues to occupy large areas of inland salinised catchments (Morgan et al. 1998; Beatty et al. 2011).
Projected increases in the number of extreme-temperature days (i.e. >40°C, Suppiah et al. 2007) will increase the risk of the thermal tolerances of fish being exceeded in the small refuge-pool habitats typical throughout the south-west (Davies 2010), which could result in southerly contractions of the region’s fish fauna (Booth et al. 2011). Temperature is also a critical factor in the reproduction and growth of many freshwater fish species (e.g. Pen and Potter 1991) and changes in prevailing average temperature regimes may lead to a decoupling of the thermal and hydrological conditions essential for reproductive success. Reductions in discharge and elevated temperatures are likely to favour the continued establishment and spread of alien species in south-western Australia, such as mosquitofish (Gambusia holbrooki) and goldfish (Carassius auratus). In particular, G. holbrooki is predicted to increase in prevalence because of its life-history characteristics and high thermal and salinity tolerances (Pyke 2005), with its likely increased dominance of remaining refugia further imperilling the persistence of native species into the future.
South-eastern Australia
South-eastern Australia has a diversity of freshwaters, ranging from coastal rivers to alpine streams (Table 1, Fig. 3) and spans several biogeographical provinces (Unmack 2001). The seasonally variable climate experiences precipitation peaks in winter and spring, and is frequently punctuated by drought (Murphy and Timbal 2008), with many lowland streams experiencing cease-to-flow conditions for significant periods in most years (Kennard et al. 2010). Large areas of this region are heavily developed for agriculture and forestry, or urbanised such that much of the available water is exploited for human consumption. Despite this extensive human disturbance, south-eastern Australia continues to host a relatively high number of native fish species (~54 species, Allen et al. 2002; Lintermans 2007). Climate change is predicted to severely affect the freshwaters of this region, with significant declines in rainfall and increases in temperature resulting in less runoff, increased evapotranspiration and decreased groundwater recharge, as well as increased frequency and severity of drought, bushfire and individual rainfall events (Table 1). These additional climatic impacts will add to, or compound, the existing anthropogenic stressors that have already degraded the region’s freshwaters.
South-eastern Australia has recently experienced one of the most severe and prolonged droughts on record (Murphy and Timbal 2008). During this ‘millennium drought’, many small and headwater streams ceased flowing or completely dried out, resulting in the mortality of local fish assemblages and loss of species at the regional level (Bond and Lake 2005; Morrongiello et al. 2006). Species particularly affected were river blackfish (Gadopsis marmoratus), mountain galaxias (Galaxias olidus), southern pygmy perch (Nannoperca australis) and the cool-water alien salmonids Salmo trutta and Oncorhynchus mykiss. These drought conditions reflect projections of longer dry spells and reduced runoff (CSIRO and Bureau of Meteorology 2007) that will intensify cease-to-flow events and potentially lower ground-water levels, thus rendering isolated pool habitats more vulnerable to complete desiccation via evaporation (Lake 2003). Increased drying may also expose acid sulfate soils (Kingsford et al. 2011), that when rewetted can seriously affect water quality. The occurrence of locally catastrophic fish kills may increase in the future because flow pulses following extended dry spells have commonly resulted in ‘blackwater’ events that cause mass mortality of fish because of low dissolved oxygen (Howitt et al. 2007) and toxic polyphenols (McMaster and Bond 2008). Reductions in river discharge will increase ephemerality and lead to greater fragmentation and isolation of habitat, potentially limiting genetic exchange among local populations. Although this can result in high levels of genetic structuring within ephemeral streams (e.g. N. australis Cook et al. 2007), any larger-scale drying may increase the incidence of genetic bottlenecks and the loss of locally adapted genotypes from whole regions.
Fish in many regulated lowland systems of south-eastern Australia probably face fewer acute risks of a drier climate than those in ephemeral systems, although they are still likely to be negatively affected by reduced runoff and increased human water demands. These fish assemblages are already in a generally degraded state and composed of resistant or resilient species (Humphries and Lake 2000). Further reductions in discharge during spring are likely to result in a loss of flow peaks that act as spawning cues for several lowland river fish species (Humphries et al. 1999). Reduced flood frequency and magnitude could have a negative impact on riparian trees such as river red gum (Eucalyptus camaldulensis) (Horner et al. 2009) and impede the transport of terrestrial organic carbon into the river channel, which helps support the food chain on which fish rely (Robertson et al. 1999).
In coastal river systems, most fish species are diadromous and spawn from autumn to winter, coinciding with peaks in river discharge. Recent studies, for example, have found that tupong (Pseudaphritis urvillii) (Crook et al. 2010b) and Australian grayling (Prototroctes maraena) (Koster and Dawson 2009) undertake downstream spawning migrations in response to within-channel flow peaks. O’Connor and Mahoney (2004) also showed that P. maraena will reabsorb their eggs and fail to spawn if appropriate flows do not occur during the autumn–winter period. Reductions in the frequency of flow peaks in autumn–winter that trigger spawning behaviour, therefore, present a potential threat to reproduction and recruitment of diadromous species in coastal river systems. Reduced flows are also associated with increased periods of estuary mouth closure and species that exhibit obligate diadromy will not be able to complete their life cycles if they are unable to access the sea at the appropriate time of year (Gillanders et al. 2011).
Climatic warming may compound stressors associated with stream drying through raised water temperatures and associated reduced dissolved oxygen, resulting in the physiological tolerances of at least some species being surpassed (McNeil and Closs 2007; McMaster and Bond 2008). Successful spawning and recruitment under such conditions is highly unlikely, even if a pool ultimately retains enough water to permit survival of the resident fish. For example, the maximum sustained tolerable temperatures for adult G. marmoratus, N. australis and R. semoni are within the range 27–29°C, whereas gamete viability and larval survival of N. australis and R. semoni are very low at these levels (Harasymiw 1983). Projected increases in fire frequency and severity in upland areas could also have catastrophic impacts on fishes, because of acute thermal stress, water quality degradation, habitat loss and sedimentation (Lyon and O’Connor 2008). Species such as barred galaxias (G. fuscus), two-spined blackfish (G. bispinosus) and Macquarie perch (Macquaria australasica) are particularly vulnerable to local extinction because of their already fragmented ranges (Lintermans 2007), whereas aliens such as salmonids will experience range contractions as a result of widespread warming (Bond et al. 2011).
A warmer climate may also facilitate increases in the range or abundance of some species in south-eastern Australia normally associated with warmer or more northern freshwaters (Bond et al. 2011; Booth et al. 2011). For example, the southerly range of golden perch (M. ambigua) is partly temperature-limited. Morrongiello et al. (2011) found that during the recent drought, the annual growth of M. ambigua in south-eastern Australian lakes was negatively correlated with declining water levels; however, this effect was offset by increased growth during warmer years. Despite climatic models projecting significant declines in future water availability, fish growth may increase because of a disproportionate lengthening of the growing season, and thus make these higher latitudes more favourable habitat. Such a pattern is supported by modelling of M. ambigua’s distribution under future climate-change scenarios (Bond et al. 2011). The potential for such range shifts may, however, be retarded by in-stream barriers or geographic features (Booth et al. 2011; Morrongiello et al. 2011).
Tasmania
Tasmania’s unique freshwater environment is characterised by an extensive network of lentic water bodies, particularly in the state’s Central Plateau region (Table 1; Hardie et al. 2006), resulting in a highly endemic fish fauna (Allen et al. 2002). This region’s 10 endemic galaxiids naturally have restricted distributions; however, the additional impacts of predation by alien fish species and anthropogenic catchment and waterway manipulations for hydro-electricity generation (Hardie et al. 2006; Stuart-Smith et al. 2007), irrigation and recreation have resulted in distributions being further constrained. Limited life-history and physiological-tolerance information is available for most of the galaxiid endemics; however, they are thought to be adapted to predictable and relatively benign conditions, and thus to be sensitive to environmental change.
In Tasmania, climate change is projected to result in more seasonally and spatially variable rainfall, with concordant shifts in local hydrology (Fig. 2). In the upper Derwent and Lake St Clair regions, catchment runoff is projected to significantly decline, particularly during summer (Bennett et al. 2010). In the Lake Pedder region, rainfall is projected to increase over winter and decline over summer, with no significant change to mean annual levels (Grose et al. 2010). Swamp galaxias (Galaxias parvus, Lake Pedder) and Clarence galaxias (G. johnstonii, Upper Derwent) are endemic to these regions of Tasmania’s south-west and rely on marshes and headwater rivulets as important refuges from predation and competition from brown trout (S. trutta) and the more widespread climbing galaxias (G. brevipinnis) (Crook and Sanger 1998a; Threatened Species Section 2006). Reduced summer runoff is likely to diminish habitat availability and connectivity, which may lead to further population declines for both species, as evidenced by recent surveys that indicate negative drought impacts on several G. johnstonii populations (Inland Fisheries Service, unpubl. data). Further, although projected increases in winter runoff may facilitate population connectivity, it may also heighten the risk of refuge invasion by S. trutta and G. brevipinnis because of increased stream flow and barrier inundation (Crook and Sanger 1998a).
Freshwaters on Tasmania’s Central Plateau will experience the most significant declines in runoff, with average projected decreases of between 15% and 35% by 2100 (Bennett et al. 2010). Inflow changes for three representative lakes in the Central Plateau region (Great Lake and Lakes Crescent and Sorell) are included in Fig. 2. This area supports six endemic fish species, all likely to be affected by climate change, in addition to significant existing stressors. Declining lake levels have the potential to reduce refuge and spawning habitat for paragalaxiids (Paragalaxias julianus, P. dissimilis, P. eleotroides and P. mesotes) and saddled galaxias (G. tanycephalus), in turn leading to increased predation, increased risk of recruitment failure and a subsequent reduction in their abundance. Inflow decreases are also likely to influence power generation from these lakes, resulting in increased abstraction and altered water level-management regimes, as well as causing acute and chronic water-quality deterioration within shallow lakes and lagoons.
Spawning of golden galaxias (G. auratus) in Lakes Crescent and Sorell has been linked to rising water levels in late autumn to winter, which are needed to inundate the critical spawning habitat of cobbled littoral areas (Hardie et al. 2007). Spawning takes place in water temperatures ranging from 1.4°C to 9.7°C, with spawning peaks recorded in winter when temperatures were below 5°C (Hardie et al. 2007). Decreased winter inflows, coupled with projected warming of up to 2.5°C, may reduce spawning success by limiting habitat access and altering thermal cues.
One endemic galaxiid that may not be negatively affected by climate change is Swan galaxias (G. fontanus), which inhabits a restricted number of headwater streams around Tasmania’s eastern highlands and midlands region (Crook and Sanger 1998b). This species appears to be remarkably tolerant of high temperatures, low oxygen and high chemical concentrations associated with low flows in residual stream-pool habitats (Threatened Species Section 2006); however, a significant number of G. fontanus monitoring sites were dry in 2008–2009 (Inland Fisheries Service, unpubl. data). Nonetheless, although climate-change projections indicate that mean annual temperatures will increase in the region by several degrees, modelling (Fig. 2) indicates that inflows are projected to increase in the region and this is likely to decrease the threat of habitat loss resulting from headwater drying. In general, climate change is likely to increase the risk of population decline for many of Tasmania’s endemic galaxiids proportional to projected hydrological change. These risks, however, need to be put into perspective with existing threatening processes (Hardie et al. 2006), particularly alien species, irrigation development and hydro-electricity generation.
Adaptive potential among and within species
The evolution of Australia’s fish species in a variable and unpredictable environment has led, in part, to a depauperate fauna (Allen et al. 2002). Yet this has also meant that many species have adaptations that enable survival, exploitation and even dependence on these unique conditions. Equally important, however, is the acknowledgement that other fish species have very localised ranges and narrow tolerances because of isolation and habitat or physiological specificity (Unmack 2001). Population and species persistence under climate change depends on the interplay between ecological and evolutionary processes (Fig. 1; Kinnison and Hairston 2007); the study of variation in life-history traits provides a valuable evolutionary perspective to current and historical ecological processes and facilitates the prediction of species’ responses to future climatic scenarios.
Functional traits, such as life-history strategies, have been used to characterise groups of species and predict their response and vulnerability to environmental change (Winemiller 2005). For example, Humphries et al. (1999) used reproductive and larval developmental traits to categorise the responses of Murray–Darling species to flow parameters, whereas Crook et al. (2010a) explored the sensitivity of south-eastern Australian fish species to the impacts of drought by characterising resistance and resilience traits. The approach of this latter study is potentially useful for exploring the impacts of climate change, as increasingly drought-like conditions have been projected for much of inland Australia (CSIRO and Bureau of Meteorology 2007). Among-species trait-based approaches have also been used for comparisons among regions such as along hydrologic gradients in southern USA and eastern Australia (Olden and Kennard 2010).
For species to persist in the face of climate change, they require traits that allow them to resist its impacts (1) in situ (e.g. thermal tolerances, plastic or adapted life-history characteristics), (2) by moving to more favourable locations (e.g. individual dispersal potential, scope for range shifts) or (3) by rapidly evolving mechanisms that confer resistance or resilience (Parmesan 2006). Australia’s variable environment means that many fish species are exposed to different conditions across their range, such that preferential phenotypes in one population may be unfavourable in others. In the absence of constraints, divergent natural selection can result in the adaptation of populations, through specialist genotypes, to their local conditions (reviewed by Kawecki and Ebert 2004). Conversely, temporal environmental heterogeneity or significant gene flow may result in set trait values not being beneficial in unpredictable conditions, or adaptations diluted by immigrants. These conditions favour the evolution of adaptive phenotypic plasticity where genotypes produce different phenotypes of relatively high fitness under all environments, or dominant generalist genotypes with moderate fitness in all environments.
The exploration of within-species trait and genetic variation has only recently begun in Australian freshwater fish species and initial results indicate that significant variation in both is the norm (e.g. N. australis reproductive investment, colouration, genetics: Llewellyn 1974; Humphries 1995; Cook et al. 2007; Morrongiello et al. 2010; M. ambigua reproductive behaviour, genetics: Pusey et al. 2004; Balcombe et al. 2006; King et al. 2009; Faulks et al. 2010). The degree to which observed trait variability is related to local adaptation or phenotypic plasticity remains largely unknown; however, it does raise the possibility that some populations have an inbuilt adaptive potential to persist under changing environmental conditions. Nonetheless, the rate of climate change and the existence of other current stressors such as habitat fragmentation may render any trait or genetic variability ineffective in saving species from range reductions or extinction.
Management considerations
Future climate impacts on freshwater fish biodiversity and the required adaptation responses will depend not only on the rate and magnitude of climate change, but on the ability to reduce system vulnerability and increase system resilience by treating existing impacts and conserving freshwater habitat (Fig. 1).
Alien species
The introduction and spread of alien species has significantly impacted on freshwater environments worldwide (Dudgeon et al. 2006). These impacts now need to be considered in concert with climate change. Alterations to the thermal and hydrological regime of freshwaters will significantly interact with natural and anthropogenic factors, the ability of alien species to become established, their interactions with native fish species and ecosystem functioning, as well as the control strategies required to ameliorate impacts (Rahel and Olden 2008). Alien species already make up a significant proportion of Australia’s freshwater fish diversity (~206 natives: Allen et al. 2002; ~43 aliens: Koehn and MacKenzie 2004) and trends indicate that the proportion of aliens is likely to increase (Koehn and MacKenzie 2004; Lintermans 2004). Similarly, at least 53 native species are known to have been translocated, mostly outside their natural range (Lintermans 2004).
The changing impacts of alien species were identified as a major issue in the regional summaries above, suggesting that climate change will alter or intensify the nature of interactions between native and alien species, and increase the prevalence of novel or homogenised fish assemblages (Hobbs et al. 2006; Olden et al. 2008), which will raise further management implications. Some aliens will benefit from warmer temperatures, habitat degradation and increased modification of natural flow regimes (Bunn and Arthington 2002; Koehn 2004; Kennard et al. 2005), whereas others will be detrimentally affected by increases in flow ephemerality, disturbance and the surpassing of physiological tolerances (Closs and Lake 1996; Lyon and O’Connor 2008; Costelloe et al. 2010). Identifying the factors that give alien species competitive advantages or disadvantages over native species are key requirements for managing the threat of alien fish species under climate change.
The role of refugia
Refugia are integral properties of the abiotic environment that confer resistance or resilience on biota to disturbance and will play a vital role in facilitating the persistence of the species into the future. By definition, refuge habitats are a relative concept and depend on the species present, their adaptations and the spatial and temporal scale of disturbance (reviewed in Magoulick and Kobza 2003). Intertwined with the natural persistence of refugia into the future are anthropogenic impacts that increase climatic pressures on refuge biota (e.g. water allocation away from refuge habitats, catchment modification and habitat fragmentation). A significant increase in the awareness of refugia and their integration into legislative and management-policy frameworks is required to reduce impacts of future climate change (D. G. McNeil, S. K. C. Gehrig, G. Peters, J. Marshall, K. Cheshire, J. Lobegeiger, S. Balcombe, N. Bond, P. Reich and T. Barlow, unpubl. data). Likewise, more work is needed to fully understand the spatial and temporal extent over which current refugia are effective, the importance of connectivity among refugia, and their likely persistence and form under climate-change scenarios.
Learning from experience
The recent drought in south-eastern Australia has served as a ‘dry run’ for a drier climatic future, with the fate of many native fish populations dependent on emergency interventions by management agencies. Recently, in Victoria, environmental flows have been released to sustain populations of Murray hardyhead (Craterocephalus fluviatilis) and individuals from several G. fuscus populations were maintained in captivity to facilitate restocking after their catchments were badly burnt by bushfire. Although reactive rather than proactive, these measures are likely to have played a vital role in preventing the extinction of both species. In general, however, the allocation of water to protect threatened refuge populations has been difficult to obtain, particularly with increasing human demands. A further problem is that the majority of threatened species and critical refuge habitats exist within unregulated waterways and off-channel habitats, where water delivery may be impossible or unfeasible. Emergency responses such as fish rescues and captive-breeding programs have been widely undertaken in response to the recent drought (see the South Australian Drought Action Plan); however, they do not provide a long-term option for maintaining fish populations under a drying climate future. Management interventions around key fish habitats need long-term and proactive vision, with the aim of ameliorating the impacts of water abstraction and land-use practices. It is imperative that contingencies developed under the recent drought are not forgotten during intervening wet periods and that a focus is maintained on planning for the protection of native fish species under future drought and drying climatic regimes.
Preserving genetic diversity
Understanding the importance of genetic variability in determining how species will respond to climate change is critical to developing strategies to conserve the diversity of fish species in Australian freshwaters (sensu Adkison 1995). If trait variation is locally adaptive or dispersal potential low, then a range of populations would need to be protected to ensure that underlying genetic diversity is conserved (Adkison 1995; Hughes 2007). Conversely, if trait variation is plastic, then conservation management should focus on preserving a variety of habitats to ensure that a range of life histories continues to be expressed in wild populations (Beechie et al. 2006). Both scenarios bestow on species a capacity to adapt to climate change.
Managed translocation
Many freshwater fish species are particularly vulnerable to the impacts of climate change because they cannot resist in situ, lack adaptive potential or are prevented from moving to more favourable environments by natural and anthropogenic barriers. The future conservation of many of these species may therefore rely on proactive managed translocation of individuals to areas, either within or outside natural ranges, where conditions are more favourable (Richardson et al. 2009). Although such interventions have obvious appeal, there are ecological and management implications that need to be considered (Olden et al. 2011). For example, translocated species (e.g. translocated mouth almighty (Glossamia aprion) is suspected of causing the local extinction of Lake Eacham rainbowfish (Melanotaenia eachamensis) in Lake Eacham; Barlow et al. 1987) or genotypes within species (e.g. freshwater shrimp (Paratya australiensis); Hughes et al. 2003) may have a detrimental impact on local species or endemic genotypes. Conversely, translocated populations of the threatened M. australasica and Pedder galaxias (G. pedderensis) have played a vital role in preventing these species going extinct, although the impacts of these translocations on native species in receiving areas is unclear (Hamr 1995; Lintermans 2006). Olden et al. (2011) documented a series of recommendations that should be considered before any planned translocation occurs. These include assessing the trade-offs between the probability of extinction in the donor region versus the probability of causing declines of native species or loss of ecosystem function in the recipient region (e.g. by life-history and ecological traits), the preferential translocation of species within their natural range (dependent on genetic assessments), an examination of the trade-offs between actively facilitating a species’ range expansion through barrier removal and habitat restoration and the probability of range expansion by non-natives, and strategically targeting receiving waters to convey the best long-term resistance to climate change on translocated species.
We emphasise, however, that interventionist measures such as managed translocation, although a legitimate option, should be considered as a last resort in dealing with climate-change impacts. Instead, it is better if the resilience of a system or assemblage is maintained through the mitigation of current and well known stressors such as water abstraction, habitat fragmentation and loss, water-quality degradation and the spread of alien species, coupled with the conservation of representative aquatic habitats and fish populations (Kingsford 2011; Kingsford et al. 2011; Pittock and Finlayson 2011; Pratchett et al. 2011).
Conclusion
Climate change will have differential implications for Australia’s freshwater fish, in part dependent on current levels of anthropogenic stress. We predict that a drier, warmer and more variable climate will have a negative effect on many native fish species. Projected hydrological changes will be a particularly important driver because these will affect habitat quality, connectivity and persistence as well as disrupt key life-history processes. Our conclusion differs somewhat from those of other studies worldwide (e.g. Buisson et al. 2008; Graham and Harrod 2009; Heino et al. 2009; McCullough et al. 2009) that have strongly implicated warming as being the major impact on freshwater fish assemblages through the surpassing or optimisation of thermal tolerances. This key difference likely reflects the primary focus of many studies on northern hemisphere or high-latitude freshwater environments where water availability is currently not, and is unlikely to be, limiting. Further research into the implications of climate change on arid and semiarid freshwater systems worldwide is needed because these environments will continue to experience significant water deficits (Bates et al. 2008) that will threaten the persistence of fish assemblages into the future.
It is important to recognise that Australia’s fish fauna has often (but not always) evolved in a variable and unpredictable environment and this legacy may confer some degree of resistance or resilience to climate-change effects. The danger is that any such adaptive potential has already been eroded by current anthropogenic stress or will be swamped by the rate of climatic change. We encourage further research into the lethal (e.g. physiological tolerances) and sublethal (e.g. reproductive requirements) responses to environmental conditions by species and a more thorough examination of feedback loops involving climate drivers and anthropogenic pressures. This additional knowledge will facilitate more accurate predictions of climate-change impacts on fish and also pragmatic solutions to managers. However, it is vital that our present understanding of environment–biota interactions is not forgotten but incorporated into current actions via a proactive and adaptive approach to managing the effects of climate change.
Acknowledgements
We thank the Australian Society for Fish Biology for convening this special issue and John Koehn, Andrew Boulton, Nick Bond and Stephen Balcombe for helpful discussions on the scope and content of this manuscript. Andrew Boulton, John Koehn, Alistair Hobday, Helen Morrongiello and anonymous reviewers provided valuable comments that greatly improved the manuscript and Tom Daniel provided graphical assistance. We are grateful to our respective institutions and funding bodies for providing financial support during the writing of this review.
References
Adkison, M. D. (1995). Population differentiation in Pacific salmon: local adaptation, genetic drift, or the environment? Canadian Journal of Fisheries and Aquatic Sciences 52, 2762–2777.| Population differentiation in Pacific salmon: local adaptation, genetic drift, or the environment?Crossref | GoogleScholarGoogle Scholar |
Aldous, A., Fitzsimons, J., Richter, B., and Bach, L. (2011). Droughts, floods and freshwater ecosystems: evaluating climate change impacts and developing adaptation strategies. Marine and Freshwater Research 62, 223–231.
| Droughts, floods and freshwater ecosystems: evaluating climate change impacts and developing adaptation strategies.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3MXjsVKktrw%3D&md5=6a1ffe0cf925eda9340ec29be4eee8f8CAS |
Allen, G. R., Midgley, S. H., and Allen, M. (2002). ‘Field Guide to the Freshwater Fishes of Australia.’ (Western Australian Museum: Perth.)
Balcombe, S. R., Sheldon, F., Capon, S., Bond, N., Hadwen, W., Marsh, N., and Bernays, S. (2011). Climate change threats to native fish in degraded rivers and floodplains of the Murray–Darling Basin, Australia. Marine and Freshwater Research 62, 1099–1114.
| Climate change threats to native fish in degraded rivers and floodplains of the Murray–Darling Basin, Australia.Crossref | GoogleScholarGoogle Scholar |
Balcombe, S. R., and Arthington, A. H. (2009). Temporal changes in fish abundance in response to hydrological variability in a dryland floodplain river. Marine and Freshwater Research 60, 146–159.
| Temporal changes in fish abundance in response to hydrological variability in a dryland floodplain river.Crossref | GoogleScholarGoogle Scholar |
Balcombe, S. R., Arthington, A. A., Foster, N. D., Thoms, M. C., Wilson, G. G., and Bunn, S. E. (2006). Fish assemblages of an Australian dryland river: abundance, assemblage structure and recruitment patterns in the Warrego River, Murray–Darling Basin. Marine and Freshwater Research 57, 619–633.
| Fish assemblages of an Australian dryland river: abundance, assemblage structure and recruitment patterns in the Warrego River, Murray–Darling Basin.Crossref | GoogleScholarGoogle Scholar |
Barlow, C. G., Hogan, A. E., and Rodgers, L. J. (1987). Implication of translocated fishes in the apparent extinction in the wild of the Lake Eacham rainbowfish, Melanotaenia eachamensis. Australian Journal of Marine and Freshwater Research 38, 897–902.
| Implication of translocated fishes in the apparent extinction in the wild of the Lake Eacham rainbowfish, Melanotaenia eachamensis.Crossref | GoogleScholarGoogle Scholar |
Bates, B. C., Kundzewicz, Z. W., Wu, S., and Palutikof, J. P. (Eds) (2008). ‘Climate Change and Water. Technical Paper of the Intergovernmental Panel on Climate Change.’ (IPCC Secretariat: Geneva, The Switzerland.)
Beatty, S. J., McAleer, F. J., and Morgan, D. L. (2009). Migration patterns of fishes of the Blackwood River and relationships to groundwater intrusion. Centre for Fish & Fisheries Research, Murdoch University. Report to Department of Water, Government of Western Australia, Perth. Available at http://www.cffr.murdoch.edu.au/reports/FishMigrationsBlackwoodRiver_Nov%202009.pdf [accessed 6 February 2011].
Beatty, S. J., Morgan, D. L., Klunzinger, M., and Lymbery, A. J. (2010a). Aquatic macrofauna of Ellen Brook and the Brockman River: fresh water refuges in a salinised catchment. Report to the Ellen–Brockman Integrated Catchment Group. Centre for Fish & Fisheries Research, Murdoch University, Perth. Available at http://www.cffr.murdoch.edu.au/reports/EllenBrockmanFinalReportCFFR.pdf [accessed 7 February 2011].
Beatty, S. J., Morgan, D. L., McAleer, F. J., and Ramsay, A. (2010b). Groundwater contribution to baseflow maintains habitat connectivity for Tandanus bostocki (Teleosteii: Plotosidae) in a south-western Australian river. Ecology Freshwater Fish 19, 595–608.
| Groundwater contribution to baseflow maintains habitat connectivity for Tandanus bostocki (Teleosteii: Plotosidae) in a south-western Australian river.Crossref | GoogleScholarGoogle Scholar |
Beatty, S. J., Morgan, D. L., Rashnavadi, M., and Lymbery, A. J. (2011). Salinity tolerances of endemic freshwater fishes of south-western Australia: implications for conservation in a biodiversity hotspot. Marine and Freshwater Research 62, 91–100.
| Salinity tolerances of endemic freshwater fishes of south-western Australia: implications for conservation in a biodiversity hotspot.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3MXmtFGlsg%3D%3D&md5=f4f42b0fba0323a1e365f58a787e6776CAS |
Beechie, T., Buhle, E., Ruckelshaus, M., Fullerton, A., and Holsinger, L. (2006). Hydrologic regime and the conservation of salmon life history diversity. Biological Conservation 130, 560–572.
| Hydrologic regime and the conservation of salmon life history diversity.Crossref | GoogleScholarGoogle Scholar |
Bennett, J. C., Ling, F. L. N., Graham, B., Grose, M. R., Corney, S. P., White, C. J., Holz, G. K., Post, D. A., Gaynor, S. M., and Bindoff, N. L. (2010). Climate futures for Tasmania: water and catchments technical report. Antarctic Climate and Ecosystems Cooperative Research Centre, Hobart, Tasmania. Available at http://www.climatechange.tas.gov.au/government_action/climate_futures [accessed 23 March 2011].
Bond, N. R., and Lake, P. S. (2005). Ecological restoration and large-scale ecological disturbance: the effects of drought on the response by fish to a habitat restoration experiment. Restoration Ecology 13, 39–48.
| Ecological restoration and large-scale ecological disturbance: the effects of drought on the response by fish to a habitat restoration experiment.Crossref | GoogleScholarGoogle Scholar |
Bond, N., Thomson, J., Reich, P., and Stein, J. L. (2011). Using species distribution models to infer potential climate change-induced range shifts of freshwater fish in south-eastern Australia. Marine and Freshwater Research 62, 1043–1061.
| Using species distribution models to infer potential climate change-induced range shifts of freshwater fish in south-eastern Australia.Crossref | GoogleScholarGoogle Scholar |
Booth, D. J., Bond, N., and Macreadie, P. (2011). Detecting range shifts among Australian fishes in response to climate change. Marine and Freshwater Research 62, 1027–1042.
| Detecting range shifts among Australian fishes in response to climate change.Crossref | GoogleScholarGoogle Scholar |
Buisson, L., Thuiller, W., Lek, S., Lim, P., and Grenouillet, G. (2008). Climate change hastens the turnover of stream fish assemblages. Global Change Biology 14, 2232–2248.
| Climate change hastens the turnover of stream fish assemblages.Crossref | GoogleScholarGoogle Scholar |
Bunn, S. E., and Arthington, A. H. (2002). Basic principles and ecological consequences of altered flow regimes for aquatic biodiversity. Environmental Management 30, 492–507.
| Basic principles and ecological consequences of altered flow regimes for aquatic biodiversity.Crossref | GoogleScholarGoogle Scholar |
Bunn, S. E., Edward, D. H., and Loneragan, N. R. (1986). Spatial and temporal variation in the macroinvertebrate fauna of streams of the northern jarrah forest, Western Australia: community structure. Freshwater Biology 16, 67–91.
| Spatial and temporal variation in the macroinvertebrate fauna of streams of the northern jarrah forest, Western Australia: community structure.Crossref | GoogleScholarGoogle Scholar |
Carpenter, S. R., Fisher, S. G., Grimm, N. B., and Kitchel, J. F. (1992). Global change and freshwater ecosystems. Annual Review of Ecology and Systematics 23, 119–139.
| Global change and freshwater ecosystems.Crossref | GoogleScholarGoogle Scholar |
Chan T., Hart B., Kennard M., Pusey B., Shenton W., Douglas M. M., Valentine E., Patel S. (2011). Bayesian network models for environmental flow decision making in the Daly River, Northern Territory, Australia. River Research and Applications, in press. https://doi.org/10.1002/RRA.1456
Chessman, B. C. (2009). Climatic changes and 13-year trends in stream macroinvertebrate assemblages in New South Wales, Australia. Global Change Biology 15, 2791–2802.
| Climatic changes and 13-year trends in stream macroinvertebrate assemblages in New South Wales, Australia.Crossref | GoogleScholarGoogle Scholar |
Chivas, A. R., Garcia, A., van der Kaars, S., Couapel, M. J. J., Holt, S., Reeves, J. M., Wheeler, D. J., Switzer, A. D., Murray-Wallace, C. V., and Banerjee, D. (2001). Sea-level and environmental changes since the last interglacial in the Gulf of Carpentaria, Australia: an overview. Quaternary International 83–85, 19–46.
| Sea-level and environmental changes since the last interglacial in the Gulf of Carpentaria, Australia: an overview.Crossref | GoogleScholarGoogle Scholar |
Closs, G. P., and Lake, P. S. (1996). Drought, differential mortality and the coexistence of a native and an introduced fish species in a south east Australian intermittent stream. Environmental Biology of Fishes 47, 17–26.
| Drought, differential mortality and the coexistence of a native and an introduced fish species in a south east Australian intermittent stream.Crossref | GoogleScholarGoogle Scholar |
Cook, B. D., Bunn, S. E., and Hughes, J. M. (2007). Molecular genetic and stable isotope signatures reveal complementary patterns of population connectivity in the regionally vulnerable southern pygmy perch (Nannoperca australis). Biological Conservation 138, 60–72.
| Molecular genetic and stable isotope signatures reveal complementary patterns of population connectivity in the regionally vulnerable southern pygmy perch (Nannoperca australis).Crossref | GoogleScholarGoogle Scholar |
Costelloe, J. F., Reid, J. R. W., Pritchard, J. C., Puckridge, J. T., Bailey, V. E., and Hudson, P. J. (2010). Are alien fish disadvantaged by extremely variable flow regimes in arid-zone rivers? Marine and Freshwater Research 61, 857–863.
| Are alien fish disadvantaged by extremely variable flow regimes in arid-zone rivers?Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3cXhtVansL%2FJ&md5=bcf5cc3a4e9dcce11e63aad041f62c93CAS |
Cresswell, I., Petheram, C., Harrington, I., Buettikofer, H., Hodgen, M., (2009). Water resources of northern Australia. In ‘Northern Australia Land and Water Science Review. Final Report to the Northern Australia Land and Water Taskforce’. (Ed. P. Stone.) pp. 1–40. (Department of Infrastructure, Transport, Regional Development and Local Government: Canberra.) Available at http://www.nalwt.gov.au/files/Chapter_01-Water_Resources_in_northern_Australia_Final.pdf [accessed 20 September 2010].
Crook, D. A., and Sanger, A. C. (1998a). Threatened fishes of the world. Galaxias johnstonii Scott, 1936 (Galaxiidae). Environmental Biology of Fishes 53, 154.
Crook, D. A., and Sanger, A. C. (1998b). Threatened fishes of the world. Galaxias fontanus Fulton, 1978 (Galaxiidae). Environmental Biology of Fishes 53, 32.
Crook, D. A., Reich, P., Bond, N. R., McMaster, D., Koehn, J. D., and Lake, P. S. (2010a). Using biological information to support proactive strategies for managing freshwater fish during drought. Marine and Freshwater Research 61, 379–387.
| Using biological information to support proactive strategies for managing freshwater fish during drought.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3cXjvFSjs7g%3D&md5=80d269acaf21f989f18f33c88f071d64CAS |
Crook, D. A., Koster, W. M., Macdonald, J. I., Nicol, S. J., Belcher, C. A., Dawson, D. R., O’Mahony, D. J., Lovett, D., Walker, A., and Bannam, L. (2010b). Catadromous migrations by female tupong (Pseudaphritis urvillii) in coastal streams in Victoria, Australia. Marine and Freshwater Research 61, 474–483.
| Catadromous migrations by female tupong (Pseudaphritis urvillii) in coastal streams in Victoria, Australia.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3cXlt1Sjs7Y%3D&md5=b92114ba83e029acfe78d9bc12458fa9CAS |
CSIRO (2008). Water availability in the Murray–Darling Basin. A report to the Australian Government from the CSIRO Murray–Darling Basin Sustainable Yields Project. CSIRO, Australia. Available at http://www.csiro.au/files/files/po0n.pdf [accessed 23 March 2011].
CSIRO (2009a). Surface water yields in south-west Western Australia. A report to the Australian Government from the CSIRO South-West Western Australia Sustainable Yields Project. CSIRO Water for a Healthy Country Flagship, Australia. Available at http://www.clw.csiro.au/publications/waterforahealthycountry/swsy/pdf/SWSY-Main-Report-SurfaceWater.pdf [accessed 23 March 2011].
CSIRO (2009b). Groundwater yields in south-west Western Australia. A report to the Australian Government from the CSIRO South-West Western Australia Sustainable Yields Project. CSIRO Water for a Healthy Country Flagship, Australia. Available at http://www.clw.csiro.au/publications/waterforahealthycountry/swsy/pdf/SWSY-Main-Report-Groundwater.pdf [accessed 23 March 2011].
CSIRO and Bureau of Meteorology (2007). Climate change in Australia: technical report. CSIRO, Melbourne. Available at http://climatechangeinaustralia.com.au/technical_report.php [accessed 23 March 2011].
Davies, P. M. (2010). Climate change implications for river restoration in global biodiversity hotspots. Restoration Ecology 18, 261–268.
| Climate change implications for river restoration in global biodiversity hotspots.Crossref | GoogleScholarGoogle Scholar |
Dudgeon, D., Arthington, A. H., Gessner, M. O., Kawabata, Z. I., Knowler, D. J., Lévêque, C., Naiman, R. J., Prieur-Richard, A.-H., Soto, D., Stiassny, M. L. J., and Sullivan, C. A. (2006). Freshwater biodiversity: importance, threats, status and conservation challenges. Biological Reviews of the Cambridge Philosophical Society 81, 163–182.
| Freshwater biodiversity: importance, threats, status and conservation challenges.Crossref | GoogleScholarGoogle Scholar |
Durrant, J. (2009). Streamflow trends in south-western Western Australia. Surface water hydrology series – report no. HY32. Department of Water, Government of Western Australia. Available at http://www.water.wa.gov.au/PublicationStore/first/87846.pdf [accessed 23 March 2011].
Eliot, I., Finlayson, C. M., and Waterman, P. (1999). Predicted climate change, sea-level rise and wetland management in the Australian wet–dry tropics. Wetlands Ecology and Management 7, 63–81.
| Predicted climate change, sea-level rise and wetland management in the Australian wet–dry tropics.Crossref | GoogleScholarGoogle Scholar |
Fairfax, R., Fensham, R., Wager, R., Brooks, S., Webb, A., and Unmack, P. (2007). Recovery of the red-finned blue-eye: an endangered fish from springs of the Great Artesian Basin. Wildlife Research 34, 156–166.
| Recovery of the red-finned blue-eye: an endangered fish from springs of the Great Artesian Basin.Crossref | GoogleScholarGoogle Scholar |
Faulks, L. K., Gilligan, D. M., and Beheregaray, L. B. (2010). Clarifying an ambiguous evolutionary history: range-wide phylogeography of an Australian freshwater fish, the golden perch (Macquaria ambigua). Journal of Biogeography 37, 1329–1340.
| Clarifying an ambiguous evolutionary history: range-wide phylogeography of an Australian freshwater fish, the golden perch (Macquaria ambigua).Crossref | GoogleScholarGoogle Scholar |
Fausch, K. D., Torgersen, C. E., Baxter, C. V., and Li, H. W. (2002). Landscapes to riverscapes: bridging the gap between research and conservation of stream fishes. Bioscience 52, 483–498.
| Landscapes to riverscapes: bridging the gap between research and conservation of stream fishes.Crossref | GoogleScholarGoogle Scholar |
Gillanders, B. M., Elsdon, T. S., Halliday, I. A., Jenkins, G. P., Robins, J. B., and Valesini, F. J. (2011). Potential effects of climate change on Australian estuaries and fish-utilising estuaries: a review. Marine and Freshwater Research 62, 1115–1131.
| Potential effects of climate change on Australian estuaries and fish-utilising estuaries: a review.Crossref | GoogleScholarGoogle Scholar |
Golder and Associates (2008). Analysis of the lower Blackwood River and tributaries flow, groundwater and climate relationships and recommendation of management triggers. Report no. 077641626001 R Rev1. Report to the Department of Water, Government of Western Australia, Perth. Available at http://www.water.wa.gov.au/PublicationStore/first/97074.pdf [accessed 23 March 2011].
Graham, C. T., and Harrod, C. (2009). Implications of climate change for the fishes of the British Isles. Journal of Fish Biology 74, 1143–1205.
| Implications of climate change for the fishes of the British Isles.Crossref | GoogleScholarGoogle Scholar | 1:STN:280:DC%2BC3cjnt1ygug%3D%3D&md5=c45aed507470998e7e370afdb13d5565CAS |
Grose, M. R., Barnes-Keoghan, I., Corney, S. P., White, C. J., Holz, G. K., Bennett, J. B., Gaynor, S. M., and Bindoff, N. L. (2010). Climate futures for Tasmania: general climate impacts. Antarctic Climate and Ecosystems Cooperative Research Centre, Hobart. Available at http://www.climatechange.tas.gov.au/government_action/climate_futures [accessed 23 March 2011].
Haines, A. T., Finlayson, B. L., and McMahon, T. A. (1988). A global classification of river regimes. Applied Geography 8, 255–272.
| A global classification of river regimes.Crossref | GoogleScholarGoogle Scholar |
Halse, S. A., Ruprecht, J. K., and Pinder, A. M. (2003). Salinisation and prospects for biodiversity in rivers and wetlands of southwest Western Australia. Australian Journal of Botany 51, 673–688.
| Salinisation and prospects for biodiversity in rivers and wetlands of southwest Western Australia.Crossref | GoogleScholarGoogle Scholar |
Hamilton, S. K. (2010). Biogeochemical implications of climate change for tropical rivers and floodplains. Hydrobiologia 657, 19–35.
| Biogeochemical implications of climate change for tropical rivers and floodplains.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3cXht1WmtbrL&md5=329e376f867bf3722f564ca7133e14c9CAS |
Hamr, P. (1995). Threatened fishes of the world: Galaxias peaderensis Frankenberg, 1968 (Galaxiidae). Environmental Biology of Fishes 43, 406.
Harasymiw, B. J. (1983). ‘LaTrobe Valley Water Resources Biological Studies. Vol. IV: Effects of Temperature on Life Stages of LaTrobe River Fish Species.’ (State Electricity Commission of Victoria: Melbourne.)
Hardie, S. A., Jackson, J. E., Barmuta, L. A., and White, R. W. G. (2006). Status of galaxiid fishes in Tamania, Australia: conservation listings, threats and management issues. Aquatic Conservation: Marine and Freshwater Ecosystems 16, 235–250.
| Status of galaxiid fishes in Tamania, Australia: conservation listings, threats and management issues.Crossref | GoogleScholarGoogle Scholar |
Hardie, S. A., White, R. W. G., and Barmuta, L. A. (2007). Reproductive biology of the threatened golden galaxias Galaxias auratus Johnston and the influence of lake hydrology. Journal of Fish Biology 71, 1820–1840.
| Reproductive biology of the threatened golden galaxias Galaxias auratus Johnston and the influence of lake hydrology.Crossref | GoogleScholarGoogle Scholar |
Heino, J., Virkkala, R., and Toivonen, H. (2009). Climate change and freshwater biodiversity: detected patterns, future trends and adaptations in northern regions. Biological Reviews of the Cambridge Philosophical Society 84, 39–54.
| Climate change and freshwater biodiversity: detected patterns, future trends and adaptations in northern regions.Crossref | GoogleScholarGoogle Scholar |
Hobbs, R. J., Arico, S., Aronson, J., Baron, J. S., Bridgewater, P., Cramer, V. A., Epstein, P. R., Ewel, J. J., Klink, C. A., Lugo, A. E., Norton, D., Ojima, D., Richardson, D. M., Sanderson, E. W., Valladares, F., Vila, M., Zamora, R., and Zobel, M. (2006). Novel ecosystems: theoretical and management aspects of the new ecological world order. Global Ecology and Biogeography 15, 1–7.
| Novel ecosystems: theoretical and management aspects of the new ecological world order.Crossref | GoogleScholarGoogle Scholar |
Hobday, A. J., and Lough, J. M. (2011). Projected climate change in Australian marine and freshwater environments. Marine and Freshwater Research 62, 1000–1014.
| Projected climate change in Australian marine and freshwater environments.Crossref | GoogleScholarGoogle Scholar |
Horner, G. J., Baker, P. J., Mac Nally, R., Cunningham, S. C., Thomson, J. R., and Hamilton, F. (2009). Mortality of developing floodplain forests subjected to a drying climate and water extraction. Global Change Biology 15, 2176–2186.
| Mortality of developing floodplain forests subjected to a drying climate and water extraction.Crossref | GoogleScholarGoogle Scholar |
Howitt, J. A., Baldwin, D. S., Rees, G. N., and Williams, J. L. (2007). Modelling blackwater: predicting water quality during flooding of lowland river forests. Ecological Modelling 203, 229–242.
| Modelling blackwater: predicting water quality during flooding of lowland river forests.Crossref | GoogleScholarGoogle Scholar |
Hughes, J. M. (2007). Constraints on recovery: using molecular methods to study connectivity of aquatic biota in rivers and streams. Freshwater Biology 52, 616–631.
| Constraints on recovery: using molecular methods to study connectivity of aquatic biota in rivers and streams.Crossref | GoogleScholarGoogle Scholar |
Hughes, J., Goudkamp, K., Hurwood, D., Hancock, M., and Bunn, S. (2003). Translocation causes extinction of a local population of the freshwater shrimp Paratya australiensis. Conservation Biology 17, 1007–1012.
| Translocation causes extinction of a local population of the freshwater shrimp Paratya australiensis.Crossref | GoogleScholarGoogle Scholar |
Humphries, P. (1995). Life history, food and habitat of southern pygmy perch, Nannoperca australis, in the Macquarie River, Tasmania. Marine and Freshwater Research 46, 1159–1169.
| Life history, food and habitat of southern pygmy perch, Nannoperca australis, in the Macquarie River, Tasmania.Crossref | GoogleScholarGoogle Scholar |
Humphries, P., and Lake, P. S. (2000). Fish larvae and the management of regulated rivers. Regulated Rivers: Research and Management 16, 421–432.
| Fish larvae and the management of regulated rivers.Crossref | GoogleScholarGoogle Scholar |
Humphries, P., King, A. J., and Koehn, J. D. (1999). Fish, flows and flood plains: links between freshwater fishes and their environment in the Murray–Darling River system, Australia. Environmental Biology of Fishes 56, 129–151.
| Fish, flows and flood plains: links between freshwater fishes and their environment in the Murray–Darling River system, Australia.Crossref | GoogleScholarGoogle Scholar |
IOCI (2002). ‘Climate Variability and Change in South Western Australia.’ Indian Ocean Climate Initiative. Available at http://www.ioci.org.au/pdf/IOCI_TechnicalReport02.pdf [accessed 23 March 2011].
IPCC (2007). ‘Climate Change 2007: The Physical Science Basis.’ Contribution of Working Group I to the fourth assessment report of the Intergovernmental Panel on Climate Change. (Cambridge University Press: Cambridge, UK.)
Jones, R. N., and Durack, P. J. (2005). Estimating the impacts of climate change on Victoria’s runoff using a hydrological sensitivity model. CSIRO Marine and Atmospheric Research, Melbourne. Available at http://www.dpi.vic.gov.au/CA256F310024B628/0/1161F8AC6816BBD5CA25738400825BD1/$File/Climate+change+and+runoff.pdf [accessed 23 March 2011].
Kawecki, T. J., and Ebert, D. (2004). Conceptual issues in local adaptation. Ecology Letters 7, 1225–1241.
| Conceptual issues in local adaptation.Crossref | GoogleScholarGoogle Scholar |
Kennard, M. J., Arthington, A. H., Pusey, B. J., and Harch, B. D. (2005). Are alien fish a reliable indicator of river health? Freshwater Biology 50, 174–193.
| Are alien fish a reliable indicator of river health?Crossref | GoogleScholarGoogle Scholar |
Kennard, M. J., Pusey, B. J., Olden, J. D., MacKay, S. J., Stein, J. L., and Marsh, N. (2010). Classification of natural flow regimes in Australia to support environmental flow management. Freshwater Biology 55, 171–193.
| Classification of natural flow regimes in Australia to support environmental flow management.Crossref | GoogleScholarGoogle Scholar |
Kerezsy, A. (2009). ‘Gambusia Control in Spring Wetlands.’ (South Australian Arid Lands Natural Resources Management Board: Adelaide.)
Kerezsy, A. (2010). The distribution, recruitment and movement of fish in far western Queensland. Ph.D. Thesis, Griffith University, Brisbane.
King, A. J., Tonkin, Z., and Mahoney, J. (2009). Environmental flows enhance native fish spawning and recruitment in the Murray River, Australia. River Research and Applications 25, 1205–1218.
| Environmental flows enhance native fish spawning and recruitment in the Murray River, Australia.Crossref | GoogleScholarGoogle Scholar |
Kingsford, R. T. (2011). Conservation management of rivers and wetlands under climate change – a synthesis. Marine and Freshwater Research 62, 217–222.
| Conservation management of rivers and wetlands under climate change – a synthesis.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3MXjsVKksb0%3D&md5=f827938094a01b252709717279509058CAS |
Kingsford, R. T., Walker, K. F., Lester, R. E., Young, W. J., Fairweather, P. G., Sammut, J., and Geddes, C. (2011). A Ramsar wetland in crisis – the Coorong, Lower Lakes and Murray Mouth, Australia. Marine and Freshwater Research 62, 255–265.
| A Ramsar wetland in crisis – the Coorong, Lower Lakes and Murray Mouth, Australia.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3MXjsVKksbg%3D&md5=50a046216c7e92528af202f64520a64dCAS |
Kinnison, M. T., and Hairston, N. G. (2007). Eco-evolutionary conservation biology: contemporary evolution and the dynamics of persistence. Functional Ecology 21, 444–454.
| Eco-evolutionary conservation biology: contemporary evolution and the dynamics of persistence.Crossref | GoogleScholarGoogle Scholar |
Koehn, J. D. (2004). Carp (Cyprinus carpio) as a powerful invader in Australian waterways. Freshwater Biology 49, 882–894.
| Carp (Cyprinus carpio) as a powerful invader in Australian waterways.Crossref | GoogleScholarGoogle Scholar |
Koehn, J. D., and MacKenzie, R. F. (2004). Priority management actions for alien freshwater fish species in Australia. New Zealand Journal of Marine and Freshwater Research 38, 457–472.
| Priority management actions for alien freshwater fish species in Australia.Crossref | GoogleScholarGoogle Scholar |
Koster, W., and Dawson, D. (2009). Investigation of the movements of Australian grayling in the Bunyip River using acoustic telemetry. Unpublished report for Melbourne Water. Arthur Rylah Institute for Environmental Research Department of Sustainability and Environment, Melbourne.
Lake, P. S. (2003). Ecological effects of perturbation by drought in flowing waters. Freshwater Biology 48, 1161–1172.
| Ecological effects of perturbation by drought in flowing waters.Crossref | GoogleScholarGoogle Scholar |
Lintermans, M. (2004). Human-assisted dispersal of alien freshwater fish in Australia. New Zealand Journal of Marine and Freshwater Research 38, 481–501.
| Human-assisted dispersal of alien freshwater fish in Australia.Crossref | GoogleScholarGoogle Scholar |
Lintermans, M. (2006). The re-establishment of the endangered Macquarie perch, Macquaria australasica, in the Queanbeyan River, New South Wales, with an examination of dietary overlap with alien trout. CRC Freshwater Ecology Technical Report. Available at http://freshwater.canberra.edu.au/publications.nsf/f8748e6acfab1b7fca256f1e001536e1/9ffa733471131b0aca25725f00244a04?OpenDocument [accessed 23 March 2011].
Lintermans, M. (2007). ‘Fishes of the Murray–Darling Basin: An Introductory Guide.’ (Murray–Darling Basin Commission: Canberra.)
Llewellyn, L. C. (1973). Spawning development and temperature tolerance of the spangeld perch Madigania unicolor (Günther), from inland waters in Australia. Australian Journal of Marine and Freshwater Research 24, 73–94.
| Spawning development and temperature tolerance of the spangeld perch Madigania unicolor (Günther), from inland waters in Australia.Crossref | GoogleScholarGoogle Scholar |
Llewellyn, L. C. (1974). Spawning, development and distribution of the southern pigmy perch Nannoperca australis australis Günther from inland waters in eastern Australia. Australian Journal of Marine and Freshwater Research 25, 121–149.
| Spawning, development and distribution of the southern pigmy perch Nannoperca australis australis Günther from inland waters in eastern Australia.Crossref | GoogleScholarGoogle Scholar |
Lough, J. M., Hobday, A. J., and Jones, D. (2011). Observed climate change in Australian marine and freshwater environments. Marine and Freshwater Research 62, 984–999.
| Observed climate change in Australian marine and freshwater environments.Crossref | GoogleScholarGoogle Scholar |
Lyon, J. P., and O’Connor, J. P. (2008). Smoke on the water: can riverine fish populations recover following a catastrophic fire-related sediment slug? Austral Ecology 33, 794–806.
| Smoke on the water: can riverine fish populations recover following a catastrophic fire-related sediment slug?Crossref | GoogleScholarGoogle Scholar |
Magoulick, D. D., and Kobza, R. M. (2003). The role of refugia for fishes during drought: a review and synthesis. Freshwater Biology 48, 1186–1198.
| The role of refugia for fishes during drought: a review and synthesis.Crossref | GoogleScholarGoogle Scholar |
Mayer, X., Ruprecht, J., and Bari, M. (2005). Stream salinity status and trends in south-west Western Australia. Department of Environment. Available at http://www.water.wa.gov.au/PublicationStore/first/47530.pdf [accessed 23 March 2011].
McCullough, D. A., Bartholow, J. M., Jager, H. I., Beschta, R. L., Cheslak, E. F., Deas, M. L., Ebersole, J. L., Foott, J. S., Johnson, S. L., Marine, K. R., Mesa, M. G., Petersen, J. H., Souchon, Y., Tiffan, K. F., and Wurtsbaugh, W. A. (2009). Research in thermal biology: burning questions for coldwater stream fishes. Reviews in Fisheries Science 17, 90–115.
| Research in thermal biology: burning questions for coldwater stream fishes.Crossref | GoogleScholarGoogle Scholar |
McJannet, D., Wallace, J., and Reddell, P. (2007). Precipitation interception in Australian tropical rainforests: II. Altitudinal gradients of cloud interception, stemflow, throughfall and interception. Hydrological Processes 21, 1703–1718.
| Precipitation interception in Australian tropical rainforests: II. Altitudinal gradients of cloud interception, stemflow, throughfall and interception.Crossref | GoogleScholarGoogle Scholar |
McJannet, D. L., Wallace, J. W., Henderson, A., and McMahon, J. (2009). High and low flow regime changes at environmental assets across northern Australia under future climate and development scenarios. A report to the Australian Government from the CSIRO Northern Australian Sustainable Yields Project. CSIRO Water for a Healthy Country National Research Flagship, Division of Land and Water, Canberra. Available at http://www.clw.csiro.au/publications/waterforahealthycountry/nasy/documents/TechReports/NASY-flow-regime-changes-environmental.pdf [accessed 23 March 2011].
McMaster, D., and Bond, N. R. (2008). A field and experimental study on the tolerances of fish to Eucalyptus camaldulensis leachate and low dissolved oxygen concentrations. Marine and Freshwater Research 59, 177–185.
| A field and experimental study on the tolerances of fish to Eucalyptus camaldulensis leachate and low dissolved oxygen concentrations.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD1cXisVWitLo%3D&md5=56ae7ba9f86384ee8d9ff45067820052CAS |
McNeil, D. G., and Closs, G. P. (2007). Behavioural responses of a south-east Australian floodplain fish community to gradual hypoxia. Freshwater Biology 52, 412–420.
| Behavioural responses of a south-east Australian floodplain fish community to gradual hypoxia.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2sXjvFSit74%3D&md5=64100ed757c7d955309495275251de68CAS |
McNeil, D. G., and Schmarr, D. W. (2009). Recovery of Lake Eyre Basin fishes following drought: 2008/09 fish survey report. South Australian Research and Development Institute (Aquatic Sciences), Adelaide. SARDI Publication No. F2009/000407-1. SARDI Research Report Series No. 411. Available at http://saalnrm.sa.gov.au/Portals/8/pdf/Reports/rep_lebfish_sardi_march09.pdf [accessed 23 March 2011].
McNeil, D. G., Schmarr, D. W., and White, M. (2010). Assessment of native vegetation and endemic fish (Mogurnda clivicola) at springs in the Northern Flinders Ranges. SARDI Publication Series. South Australian Research and Development Institute, Aquatic Sciences, Adelaide.
Morgan, D. L., Gill, H. S., and Potter, I. C. (1998). Distribution, identification and biology of freshwater fishes in south-western Australia. Records of the Western Australian Museum Supp No. 56, 1–97.
Morgan, D. L., Gill, H. S., and Potter, I. C. (2000). Age composition, growth and reproductive biology of the salamanderfish Lepidogalaxias salamandroides: a re-examination. Environmental Biology of Fishes 57, 191–204.
| Age composition, growth and reproductive biology of the salamanderfish Lepidogalaxias salamandroides: a re-examination.Crossref | GoogleScholarGoogle Scholar |
Morrongiello, J., Elith, J., and Crook, D. (2006). Impacts of drought on fish in Victorian rivers and streams. Arthur Rylah Institute for Environmental Research, Melbourne.
Morrongiello, J. R., Bond, N. R., Crook, D. A., and Wong, B. B. M. (2010). Nuptial coloration varies with ambient light environment in a freshwater fish. Journal of Evolutionary Biology 23, 2718–2725.
| Nuptial coloration varies with ambient light environment in a freshwater fish.Crossref | GoogleScholarGoogle Scholar | 1:STN:280:DC%2BC3cbmvVWkuw%3D%3D&md5=9118803e681c7bc1cfdfbeb1faaecf03CAS |
Morrongiello, J. R., Crook, D. A., King, A. J., Ramsey, D. S. L., and Brown, P. (2011). Impacts of drought and predicted effects of climate change on fish growth in temperate Australian lakes. Global Change Biology 17, 745–755.
| Impacts of drought and predicted effects of climate change on fish growth in temperate Australian lakes.Crossref | GoogleScholarGoogle Scholar |
Murphy, B. F., and Timbal, B. (2008). A review of recent climate variability and climate change in southeastern Australia. International Journal of Climatology 28, 859–879.
| A review of recent climate variability and climate change in southeastern Australia.Crossref | GoogleScholarGoogle Scholar |
Nicholls, N. (2005). Climate variability, climate change and the Australian snow season. Australian Meteorological Magazine 54, 177–185.
Nicholls, N., Drosdowsky, W., and Lavery, B. (1997). Australian rainfall variability and change. Weather 52, 66–72.
O’Connor, J. P., and Mahoney, J. C. (2004). Observations of ovarian involution in the Australian grayling (Prototroctes maraena). Ecology Freshwater Fish 13, 70–73.
| Observations of ovarian involution in the Australian grayling (Prototroctes maraena).Crossref | GoogleScholarGoogle Scholar |
Olden, J. D., and Kennard, M. J. (2010). Intercontinental comparision of fish life-history strategies along a gradient of hydrologic variability. In ‘Community Ecology of Stream Fishes: Concepts, Approaches, and Techniques’. (Eds K. B. Gido and D. A. Jackson.) pp. 83–107. (American Fisheries Society: Bethesda, MA.)
Olden, J. D., Kennard, M. J., and Pusey, B. J. (2008). Species invasions and the changing biogeography of Australian freshwater fishes. Global Ecology and Biogeography 17, 25–37.
Olden, J. D., Kennard, M. J., Lawler, J. J., and Poff, N. L. (2011). Challenges and opportunities in implementing managed relocation for conservation of freshwater species. Conservation Biology 25, 40–47.
| Challenges and opportunities in implementing managed relocation for conservation of freshwater species.Crossref | GoogleScholarGoogle Scholar |
Parmesan, C. (2006). Ecological and evolutionary responses to recent climate change. Annual Review of Ecology Evolution and Systematics 37, 637–669.
| Ecological and evolutionary responses to recent climate change.Crossref | GoogleScholarGoogle Scholar |
Parmesan, C., and Yohe, G. (2003). A globally coherent fingerprint of climate change impacts across natural systems. Nature 421, 37–42.
| A globally coherent fingerprint of climate change impacts across natural systems.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD3sXoslM%3D&md5=881f3bd5bf4de5383d00e613bb3bd47fCAS |
Pen, L. J., and Potter, I. C. (1991). The biology of the western pygmy perch, Edelia vittata, and comparisons with two other teleost species endemic to south-western Australia. Environmental Biology of Fishes 31, 365–380.
| The biology of the western pygmy perch, Edelia vittata, and comparisons with two other teleost species endemic to south-western Australia.Crossref | GoogleScholarGoogle Scholar |
Pittock, J., and Finlayson, C. M. (2011). Australia’s Murray–Darling Basin: freshwater ecosystem conservation options in an era of climate change. Marine and Freshwater Research 62, 232–243.
| Australia’s Murray–Darling Basin: freshwater ecosystem conservation options in an era of climate change.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3MXjsVKksbo%3D&md5=c87b710ca2ef52d8bbf6ae5aad474251CAS |
Pratchett, M. S., Bay, L. K., Gehrke, P. C., Koehn, J. D., Osborne, K., Pressey, R. L., Sweatman, H. P. A., and Wachenfeld, D. (2011). Contribution of climate change to degradation and loss of critical fish habitats in Australian marine and freshwater environments. Marine and Freshwater Research 62, 1062–1081.
| Contribution of climate change to degradation and loss of critical fish habitats in Australian marine and freshwater environments.Crossref | GoogleScholarGoogle Scholar |
Puckridge, J. T., Sheldon, F., Walker, K. F., and Boulton, A. J. (1998). Flow variability and the ecology of large rivers. Marine and Freshwater Research 49, 55–72.
| Flow variability and the ecology of large rivers.Crossref | GoogleScholarGoogle Scholar |
Pusey, B. J. (1990). Seasonality, aestivation and the life history of the salamanderfish Lepidogalaxias salamandroides Mees (Pisces: Lepidogalaxiidae). Environmental Biology of Fishes 29, 15–26.
| Seasonality, aestivation and the life history of the salamanderfish Lepidogalaxias salamandroides Mees (Pisces: Lepidogalaxiidae).Crossref | GoogleScholarGoogle Scholar |
Pusey, B. J., and Edward, D. H. D. (1990). Structure of fish assemblages in waters of the southern acid peat flats, south-western Australia. Australian Journal of Marine and Freshwater Research 41, 721–734.
| Structure of fish assemblages in waters of the southern acid peat flats, south-western Australia.Crossref | GoogleScholarGoogle Scholar |
Pusey, B. J., and Kennard, M. J. (1996). Species richness and geographical variation in assemblage structure of the freshwater fish fauna of the Wet Tropics region of northern Queensland. Marine and Freshwater Research 47, 563–573.
| Species richness and geographical variation in assemblage structure of the freshwater fish fauna of the Wet Tropics region of northern Queensland.Crossref | GoogleScholarGoogle Scholar |
Pusey, B. J., and Kennard, M. J. (2009). Aquatic ecosystems of northern Australia. In ‘Northern Australia Land and Water Science Review. Final Report to the Northern Australia Land and Water Taskforce’. (Ed. P. Stone) pp. 1–73. (Department of Infrastructure, Transport, Regional Development and Local Government: Canberra). Available at http://www.nalwt.gov.au/files/Chapter_03-Aquatic_ecosystems_in_northern_Australia.pdf [accessed 20 September 2010].
Pusey, B. J., Kennard, M. J., and Arthington, A. H. (2004). ‘Freshwater Fishes of Northeastern Australia.’ (CSIRO Publishing: Melbourne.)
Pusey, B. J., Kennard, M. J., and Arthington, A. H. (2008). Origins and maintenance of freshwater fish biodiversity in the Wet Tropics region. In ‘Living in a Dynamic Tropical Forest Landscape’. (Eds N. Stork and S. Turton.) pp. 150–161. (Blackwell Publishing Ltd: Oxford, UK.)
Pyke, G. H. (2005). A review of the biology of Gambusia affinis and G. holbrooki. Reviews in Fish Biology and Fisheries 15, 339–365.
| A review of the biology of Gambusia affinis and G. holbrooki.Crossref | GoogleScholarGoogle Scholar |
Rahel, F. J., and Olden, J. D. (2008). Assessing the effects of climate change on aquatic invasive species. Conservation Biology 22, 521–533.
| Assessing the effects of climate change on aquatic invasive species.Crossref | GoogleScholarGoogle Scholar |
Rayner, T. S., Pusey, B. J., and Pearson, R. G. (2008). Seasonal flooding, instream habitat structure and fish assemblages in the Mulgrave River, north-east Queensland: towards a new conceptual framework for understanding fish-habitat dynamics in small tropical rivers. Marine and Freshwater Research 59, 97–116.
| Seasonal flooding, instream habitat structure and fish assemblages in the Mulgrave River, north-east Queensland: towards a new conceptual framework for understanding fish-habitat dynamics in small tropical rivers.Crossref | GoogleScholarGoogle Scholar |
Rayner, T. S., Pusey, B. J., and Pearson, R. G. (2009). Spatio-temporal dynamics of fish feeding in the lower Mulgrave River, north-eastern Queensland: the influence of seasonal flooding, instream productivity and invertebrate abundance. Marine and Freshwater Research 60, 97–111.
| Spatio-temporal dynamics of fish feeding in the lower Mulgrave River, north-eastern Queensland: the influence of seasonal flooding, instream productivity and invertebrate abundance.Crossref | GoogleScholarGoogle Scholar |
Richardson, D. M., Hellmann, J. J., McLachlan, J. S., Sax, D. F., Schwartz, M. W., Gonzalez, P., Brennan, E. J., Camacho, A., Root, T. L., Sala, O. E., Schneider, S. H., Ashe, D. M., Clark, J. R., Early, R., Etterson, J. R., Fielder, E. D., Gill, J. L., Minteer, B. A., Polasky, S., Safford, H. D., Thompson, A. R., and Vellend, M. (2009). Multidimensional evaluation of managed relocation. Proceedings of the National Academy of Sciences, USA 106, 9721–9724.
| Multidimensional evaluation of managed relocation.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD1MXotFGjsr8%3D&md5=f43b29491a927df51a27ef2656996925CAS |
Robertson, A. I., Bunn, S. E., Boon, P. I., and Walker, K. F. (1999). Sources, sinks and transformations of organic carbon in Australian floodplain rivers. Marine and Freshwater Research 50, 813–829.
| Sources, sinks and transformations of organic carbon in Australian floodplain rivers.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD3cXks1ymug%3D%3D&md5=b63884b07587f5fea3581c9b9fccdc0cCAS |
Russell, D. J., Hales, P. W., and Helmke, S. A. (1996). Stream habitat and fish resources in the Russell and Mulgrave Rivers Catchment. Queensland Department of Primary Industries, Northern Fisheries Centre, Brisbane.
Schiller, C. B., Bruce, A. M. G., and Gehrke, P. C. (1997). Distribution and abundance of native fish in New South Wales rivers. In ‘Fish and Rivers in Stress’. (Eds P. C. Gehrke and J. H. Harris.) pp. 71–103. (CRC for Freshwater Ecology and New South Wales Fisheries: Sydney.)
Schneider, C., and Moritz, C. (1999). Rainforest refugia and evolution in Australia’s Wet Tropics. Proceedings of the Royal Society of London. Series B. Biological Sciences 266, 191–196.
| Rainforest refugia and evolution in Australia’s Wet Tropics.Crossref | GoogleScholarGoogle Scholar |
Silcock, J. (2009). Identification of permanent refuge waterbodies in the Cooper Creek and Georgina–Diamantina River catchments for Queensland and South Australia. South Australian Arid Lands Natural Resources Management Board, Port Augusta. Available at http://www.saalnrm.sa.gov.au/Portals/8/pdf/Reports/rep_June09_IDpermanentwaterbodies.pdf [accessed 23 March 2011].
Still, C. J., Foster, P. N., and Schneider, S. H. (1999). Simulating the effects of climate change on tropical montane cloud forests. Nature 398, 608–610.
| Simulating the effects of climate change on tropical montane cloud forests.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaK1MXislykurg%3D&md5=c09335da9d7fa6994a7ed79de58b9179CAS |
Stuart-Smith, R. D., Stuart-Smith, J. F., White, R. W. G., and Barmuta, L. A. (2007). The impact of an introduced predator on a threatened galaxiid fish is reduced by the availability of complex habitats. Freshwater Biology 52, 1555–1563.
Suppiah, R., Hennessy, K. J., Whetton, P. H., McInnes, K., Macadam, I., Bathols, J., Ricketts, J., and Page, C. M. (2007). Australian climate change projections derived from simulations performed for the IPCC 4th assessment report. Australian Meteorological Magazine 131, 131–152.
Tedesco, P. A., Oberdorff, T., Lasso, C. A., Zapata, M., and Hugueny, B. (2005). Evidence of history in explaining diversity patterns in tropical fish. Journal of Biogeography 32, 1899–1907.
| Evidence of history in explaining diversity patterns in tropical fish.Crossref | GoogleScholarGoogle Scholar |
Thomas, C. D., Cameron, A., Green, R. E., Bakkenes, M., Beaumont, L. J., Collingham, Y. C., Erasmus, B. F. N., de Siqueira, M. F., Grainger, A., Hannah, L., Hughes, L., Huntley, B., van Jaarsveld, A. S., Midgley, G. F., Miles, L., Ortega-Huerta, M. A., Peterson, A. T., Phillips, O. L., and Williams, S. E. (2004). Extinction risk from climate change. Nature 427, 145–148.
| Extinction risk from climate change.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2cXhtFOgtQ%3D%3D&md5=ddd9396c5d742894b328ef14d3b5fd6aCAS |
Thompson, G. G., and Withers, P. C. (2002). Aerial and aquatic respiration of the Australian desert goby, Chlamydogobius eremius. Comparative Biochemistry and Physiology – Part A. Molecular and Integrated Physiology 131, 871–879.
| Aerial and aquatic respiration of the Australian desert goby, Chlamydogobius eremius.Crossref | GoogleScholarGoogle Scholar |
Threatened Species Section (2006). Recovery plan: Tasmanian Galaxiidae 2006–2010. Department of Primary Industries, Water, Hobart. Available at http://www.environment.gov.au/biodiversity/threatened/publications/pubs/tasmanian-galaxiidae.pdf [accessed 23 March 2011].
Unmack, P. J. (2001). Biogeography of Australian freshwater fishes. Journal of Biogeography 28, 1053–1089.
| Biogeography of Australian freshwater fishes.Crossref | GoogleScholarGoogle Scholar |
Williams, S. E., Bolitho, E. E., and Fox, S. (2003). Climate change in Australian tropical rainforests: an impending environmental catastrophe. Proceedings of the Royal Society B-Biological Sciences , 1887–1892.
Winemiller, K. O. (2005). Life history strategies, population regulation, and implications for fisheries management. Canadian Journal of Fisheries and Aquatic Sciences 62, 872–885.
| Life history strategies, population regulation, and implications for fisheries management.Crossref | GoogleScholarGoogle Scholar |
Woinarski, J., Mackey, B., Nix, H., and Traill, B. (2007). ‘The Nature of Northern Australia. Natural Values, Ecological Processes and Future Prospects.’ (ANUE Press: Canberra.)
Woodroffe, C. D. (1995). Response of tide-dominated mangrove shorelines in northern Australia to anticipated sea-level rise. Earth Surface Processes and Landforms 20, 65–85.
| Response of tide-dominated mangrove shorelines in northern Australia to anticipated sea-level rise.Crossref | GoogleScholarGoogle Scholar |
Xenopoulos, M. A., and Lodge, D. M. (2006). Going with the flow: using species–discharge relationships to forecast losses in fish diversity. Ecology 87, 1907–1914.
| Going with the flow: using species–discharge relationships to forecast losses in fish diversity.Crossref | GoogleScholarGoogle Scholar |
Yokoyama, Y., Purcell, A., Lambeck, K., and Johnston, P. (2001). Shore-line reconstruction around Australia during the last glacial maximum and late glacial stage. Quaternary International 83–85, 9–18.
| Shore-line reconstruction around Australia during the last glacial maximum and late glacial stage.Crossref | GoogleScholarGoogle Scholar |