

Business partner or simple catch? The economic value of the sicklefin lemon shark in French Polynesia
E. Clua A D , N. Buray B , P. Legendre C , J. Mourier B and S. Planes BA Secretariat of the Pacific Community, BP D5, Noumea, New Caledonia.
B Centre de Recherches Insulaires et Observatoire de l’Environnement (CRIOBE – USR 3278 EPHE-CNRS), BP 1013, 98729 Moorea, French Polynesia.
C Département de Sciences biologiques, Université de Montréal, C.P. 6128, succ. Centre-ville, Montréal, Québec H3C 3J7, Canada.
D Corresponding author. Email: Ericc@spc.int
Marine and Freshwater Research 62(6) 764-770 https://doi.org/10.1071/MF10163
Submitted: 19 June 2010 Accepted: 15 December 2010 Published: 24 June 2011
Journal Compilation © CSIRO Publishing 2011 Open Access CC BY-NC-ND
Abstract
Most arguments invoked so far by the scientific community in favour of shark conservation rely on the ecological importance of sharks, and have little impact on management policies. During a 57-month study, we were able to individually recognise 39 sicklefin lemon sharks that support a shark-feeding ecotourism activity in Moorea Island, French Polynesia. We calculated the direct global revenue generated by the provisioning site, based on the expenses of local and international divers. The total yearly revenue was around USD5.4 million and the 13 sharks most often observed at the site had an average contribution each of around USD316 699. Any one of these sharks represents a potential contribution of USD2.64 million during its life span. We argue that publicising economic values per individual will be more effective than general declarations about their ecological importance for convincing policy makers and fishers that a live shark is more valuable than a dead shark for the local economy. Studies monitoring the potential negative ecological effects of long-term feeding of sharks should, however, be conducted to ensure these are also considered. Besides declarations about the non-consumptive direct-use value of sharks, as promoted by ecotourism, the calculation of their other economic values should also benefit shark conservation.
Additional keywords: conservation, ecotourism, Negaprion acutidens, shark-feeding, total economic value (TEV).
Introduction
There is a general consensus that the ongoing decimation of shark populations by commercial fishing is putting many species in danger of extinction (Baum et al. 2003; Clarke et al. 2006; Ferretti et al. 2008). The situation is alarming because of the critical role played by sharks as top predators in marine ecosystems and the potential consequences of their depletion through cascading effects (Stevens et al. 2000; Sergio et al. 2006; Heithaus et al. 2008; Baum and Worm 2009). As an example, the decline in abundance of 11 shark species along the east coast of North America has led to increased populations of cownose ray (Rhinoptera bonasus), which has significantly impacted the scallops on which the rays feed (Heithaus et al. 2008). In reef systems, the absence of apex predators has a significant impact on the total fish biomass and the structure of the trophic pyramid, altering the health of the ecosystems (Friedlander and De Martini 2002). In spite of this well-demonstrated role, and the arguments of the scientific community in favour of shark conservation, the status levels of shark populations around the world have not improved. At the moment, 17% of the shark and ray species are listed within the Critically Endangered, Endangered, and Vulnerable categories of the International Union for Conservation of Nature Red List (IUCN 2008), and a further 47% are listed as ‘data deficient’ (Polidaro et al. 2008).
The extractive activity of fishing relies on the market value for consumption, mainly motivated by the shark fin trade (Brierley 2007). Estimates of the world annual revenues from the fin trade (including the sale of shark carcasses and other by-products) range from USD400 million to USD550 million (Clarke et al. 2007). However, sharks have other attractive economic values, as shown through the development of ecotourism of the whale shark, Rhincodon typus, in the Republic of Seychelles (Rowat and Engelhardt 2007), and in Australia where the world’s first whale shark tourism industry was established in 1989. In 2006, tourist expenditures in the region of Ningaloo Reef (Western Australia) were USD758 per trip, and the total expenditure was USD5.9 million (Catlin et al. 2009). Recreational activities in South Africa, focussing on snorkelling in the Aliwal Shoal region (KwalaZulu Land) with tiger sharks, Galeocerdo cuvier, generated around USD1.62 million per year (Dicken and Hosking 2009). In the Republic of the Fiji Islands, Brunnschweiler (2009) showed that, in addition to the direct income of the two diving centres involved in the ecotourism activity, a shark feeding site was providing the eligible Fijian villages around Beqa Island (South Viti Levu) with around USD60 000 per year in compensation for not using the marine reserve where bull sharks, Carcharhinus leucas, were aggregated. However, in these studies, the economic valuation was conducted globally, with no specific references to the number of sharks, and no study has ever been done of the economic value of individual animals.
Marine mammal conservation is supported by the worldwide whale-watching industry; the economic value of that industry was estimated at USD160 million in the US in the late 1990s (Hoyt 2001). By attaching economic value to wildlife and educating tourists about the need for conservation, researchers can use wildlife tourism as leverage for the conservation of vulnerable and endangered species (Bookbinder et al. 1998; Higginbottom et al. 2003). At present, whale-watching takes place on all continents in more than 500 communities in over 70 countries, and targets over 600 regional populations of cetaceans (Hoyt 2007). Any information promoting a similar development for the benefit of shark conservation appears timely. In French Polynesia, shark fishing was officially banned in 2006 (Arrêté 396/CM, 28 April 2006), but poaching and attempts to cull sharks by local fishers are still seen (see Accessory Publication), and could jeopardise the ecotourism activity. This effect occurred in the Republic of Maldives where fishers, removing ∼20 grey reef sharks Carcharhinus amblyrhynchos with a market value of only about USD1000, caused diver operators to suspend their activity, resulting in a loss of USD500 000 per year for the local economy (Anderson and Waheed 1999). Given the audience of fishers and resource managers, this goal of convincing them about the higher indirect economic interest of maintaining sharks alive instead of fishing them for a direct income could be better met by providing information about individual sharks instead of a shark population in general.
In the present paper, we calculated the direct revenues to Moorea Island, French Polynesia, derived from tourist divers visiting a sicklefin lemon shark (Negaprion acutidens) feeding site. Based on a long-term study, where 39 lemon sharks involved in ecotourism activity were individually identified and followed through time, we were able to estimate the contributions of individual sharks to the overall income, and to assess their potential contribution throughout their life span.
Materials and methods
Study of the shark-feeding activity in Moorea Island
Shark feeding consists of using a small amount of natural bait to attract and aggregate animals at a given site where divers can observe them. This type of activity began in Moorea Island, in the Society Islands Archipelago, French Polynesia, in the late 1980s. Under public pressure regarding the risk of increasing aggressiveness towards humans, local authorities limited shark-feeding activities to two zones on the outer slope of the reef, and banned provisioning of sharks in the lagoon where most human activity is concentrated. The two zones where provisioning is allowed are located in the northern part of the island, close to several international seaside resorts. Our specific study area was at the diving spot called ‘Opunohu’ (17°30′S, 149°51′W), on the outer slope off Opunohu Bay, which was selected for its abundance of sicklefin lemon sharks, Negaprion acutidens. This tropical coastal species is very attractive in the context of ecotourism because its large size impresses tourists; N. acutidens can reach 3.5 m in total length (TL) (Compagno 1984). Although they usually display a shy and solitary behaviour, lemon sharks can aggregate in significant numbers through feeding (up to 15 animals in French Polynesia) (see Accessory Publication). Five different diving centres bring tourists daily to the Opunohu site, but only three feed the sharks. For almost five years (57 months), from January 1st, 2005 to September 31st, 2009, we monitored the feeding sessions implemented by the TopDive Center in the presence of tourist divers, at depths of 20–25 m; each session lasted 60–100 min. Sessions consisted of hiding a small cage containing tuna heads and flesh on the substrate to lure and keep the sharks around to ensure close observation. The food was released at the end of the dive. Data on presence/absence of individual sharks were recorded for each dive using photo-identification, the size and gender, and natural marks on shark bodies such as scars, notches, missing tissues or coloured spots (Buray et al. 2009).
Calculation of the direct non-consumptive use value of sharks
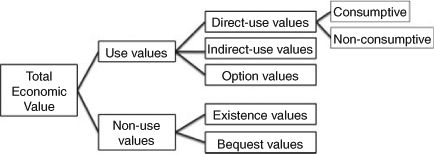
After the seminal study of Costanza et al. (1997) valuing the world natural capital and ecosystem services, the concept of Total Economic Value (TEV) was developed, in particular through the Millenium Ecosystem Assessment (MEA 2005). Following this methodology, the economic value of an entire ecosystem or a single species can be divided between use and non-use values. The former can be split into non-consumptive and consumptive direct use values, indirect use values, and option values. The non-use values are shared between existence and bequest values (Fig. 1). Although fisheries focus on the consumptive direct use value of sharks, the development of ecotourism activities involving sharks fits into the direct non-consumptive use values. For assessing this value, we collected data on the total expenditures that can be directly linked to the purchase of a dive that allows visitors to watch lemon sharks in their natural environment. Divers were divided in two groups composed of local divers and international divers. For the local divers group, expenditures are limited to the diving fees. For the international divers group, expenditures cover the average cost for a diving session (including two dives), accommodation for one night before and one night after diving, four meals and, if they came to Moorea with the intent to see sharks, a return flight ticket between Tahiti Island (where international flights land) and Moorea Island. Based on this approach, we obtained a total expenditure, which we combined with lemon shark information in three ways: (1) we first calculated an average value per shark by dividing this amount by the number of sharks that contributed to the attractiveness of the Opunohu site; (2) we then did a more accurate calculation by dispatching this same amount per individual shark, based on the visitation of the Opunohu site by individual sharks; and (3) finally, based on the sicklefin lemon shark life history traits, we calculated the average economic value of a shark for its life span, with a yearly discount rate of 8% as is usually done in the South Pacific for adjusting a future value to a present one, by taking off a certain percentage for each given year of the concerned period (Johnston and Pickering 2003).
|
Assessment of the diving activity in Moorea Island and the attractiveness of sharks for divers
French Polynesia is a well-known destination for tourism, with a strong reputation in the specific market of ‘honeymooners’; in 2009, 160 447 tourists visited this French territory, which has a total resident population of 259 600 people (IEOM 2010). To obtain an accurate number of dives per year at the Opunohu site, where the lemon sharks are aggregated through provisioning, we contacted the five diving centres that take divers to this location. We requested figures about the total number of dives at the Opunohu site between 2005 and 2008 (four years), including information about the origin of divers (either local or international). We then calculated the average number of dives as the mean for the four years. The importance of the lemon sharks in the divers’ decision to purchase a dive from one of the five diving centres was assessed through a survey conducted by sending emails to over 300 divers who had had shark experiences with the TopDive Center asking ‘did the potential presence of sharks during the dive play a positive role in: (i) your choice for coming purposely to dive in Moorea?; (ii) purchasing a dive at the Opunohu site while you already were in Moorea (for another reason than to specifically watch sharks)?; and (iii) the quality of your dive experience?’ The divers were also asked to mention their country of residence. If they were living in Moorea, they were asked to answer only the last two questions.
Results
Direct income generated by lemon sharks
The five diving centres bringing divers to the Opunohu site for the four years totalled 61 050 person-dives in this location, for an average of 15 262 divers per year (s.d. = 3158). A total of 151 persons answered the email survey, comprising 26% of local divers and 74% of international divers. Amongst the international divers, 27% of them had came purposely to Moorea for shark watching and 68% of the remaining ones did not originally come to Moorea for sharks but made the decision to dive with sharks during their stay; 100% of the local divers declared that sharks had significantly contributed to their decision to purchase a dive. Based on these proportions, we estimated that a total of 3968 local and 3049 international divers came to Moorea each year with the specific purpose to see sharks, and an additional 5606 international divers made the decision to go and dive with sharks while they were staying in Moorea.
A diving session comprises two dives, and costs USD150 for an international diver and USD125 for a local diver. The average cost for a night in Moorea is around USD186 (including breakfast); an average meal (either lunch or dinner) costs USD26 and the two-way ticket from Tahiti to Moorea costs USD62. The cost of transportation on the island (taxis, car rental) is not included in our calculations. The average expenditure for an international diver who purposely came for shark watching is around USD688; it drops down to USD626 if the choice of diving for shark watching was made in Moorea (as we do not include the cost of airfare). If we multiply these figures with the number of international and local divers mentioned above, we obtain a total annual generated income around USD248 015 for local divers and USD5 156 349 for international divers, respectively. The total amount is USD5 404 365.
Individual and life-span economic value of sharks
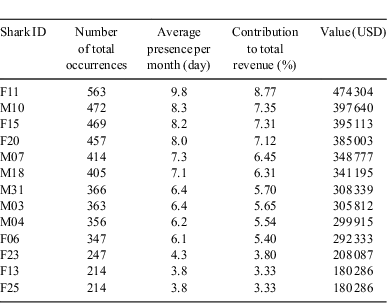
We studied 1013 diving sessions that allowed 6415 observations of 39 individual lemon sharks. The contribution of each shark to the total revenue is estimated by the relative importance of its participation to the feedings. It varied from 8.77% (563 sightings) of the total occurrences for the most regular animal, down to 0.02% for a female shark that showed up only once (Fig. 2). The 13 most commonly occurring animals, which represent 33% of the population, accounted for 73% of the occurrences and were considered to be resident sharks. Based on the total revenue of USD5.404 million, their decreasing contributions, depending on their sightings, ranged from USD474 304 to USD180 286 (Table 1). These 13 sharks at the site each had an average contribution of USD316 699.

|
|
In the absence of accurate data on the life history of the sicklefin lemon shark, we used the life history traits of the Atlantic lemon shark, Negaprion brevirostris. A growth curve gives an asymptotic maximum age of 27 years for this species, with sexual maturity reached at about 11 years of age (Brown and Gruber 1988), corresponding to the average size of 2.4-m TL. Given that the youngest sharks observed at the feeding site were around 2.1-m TL (corresponding to an age of ∼7 years), we hypothesise that a shark could remain associated with to a feeding site for 20 years. Based on the average contribution of a shark in the present study of USD138 573 (equal to the mean yearly financial contribution of the 39 sharks), a lemon shark would represent a direct non-consumptive use value for its entire life of USD2.64 million in the present context of Moorea Island.
Discussion
Individual and life-span economic values
Diving with sharks is estimated to have contributed millions of dollars to local and regional economies for decades. The annual revenue per site or country ranges from USD2.2–7.4 million, depending on year and region (Table 2). Our findings of USD5.4 million for Moorea, a single island of French Polynesia, are consistent with these figures. However, in the previous studies reported in Table 2, the economic valuation was globally conducted, with no specific reference to the number of sharks. No study had ever been done of the economic value of individual sharks. Identification of individual sharks allowed us to estimate their individual contributions to the total revenue. A parallel study on the effects of shark feeding on the behaviour of these sicklefin lemon sharks allowed us to demonstrate a significantly increasing residency pattern, in particular for the 13 most commonly sighted sharks on which the diving tourism relies (Clua et al. 2010). In addition to assessing a revenue total for Moorea Island, we were able to show that a single resident lemon shark can generate up to USD474 000 per year in revenue and is worth up to USD2.64 million for the local economy during its entire life. We believe that this type of information is critical, and more efficient than total figures, in the process of advocating shark conservation, in particular for local and coastal fishers. The objective is to convince them and other resource users that a shark alive is more valuable than a dead shark for the local economy, which can obtain continuous direct and indirect income from live sharks. This process will never be fully successful until some compensatory mechanism is established that allows all stakeholders, in particular fishers, to benefit indirectly from ecotourism revenues (Wilson 2003; Dobson 2006).

|
There are two other islands with similar international reputations for shark diving tourism in French Polynesia: Bora-Bora and Rangiroa Islands. Sharks and their potential for attracting tourists appear to be an important asset to be preserved, mainly in the context of a declining tourism industry, such as in French Polynesia, which faced a drop of 29% in activity between 2006 and 2009 (IEOM 2010). The shark tourism industry in the Polynesian islands and elsewhere should benefit from our findings.
Monitoring the potential negative effects of feeding sharks
The calculation of the economic value of sharks will help demonstrate that non-consumptive wildlife tourism better assists the conservation of vulnerable and endangered species, by attaching economic value to wildlife, which speaks to decision makers, and by educating tourists about the need for conservation (Halpenny 2003; Higginbottom et al. 2003; Topelko and Dearden 2005). However, care must be taken to anticipate and alleviate possible negative effects of shark diving ecotourism on shark behaviour, habitat and ecology (Orams 2002; Walker 2002). Operators who specialise in wildlife-based tourism tend to be characterised by small and medium-sized enterprises that are often locked into dependence relationships with external creditors. Therefore, issues of profitability will often take precedence over issues of environmental protection (Williams and Montanari 1999; Issacs 2000). In the best cases, the balancing of anthropocentric and ecocentric concerns in the management of shark-based tourism seems to often result in compromises that do not provide full support to conservation (Dobson 2006). Given the lack of knowledge on these issues, which has been one of the major pitfalls in the assessment of marine ecotourism (Garrod and Wilson 2004), studies about the effects of tourism activity upon the health and well-being of the target species must be developed, to ensure that the entertainment of tourists will not prevail on the well-being and reproduction of wild animals (Newsome and Rodger 2008). Our preliminary results from Moorea Island in 2005 show a risk of behavioural shift for the lemon sharks (Clua et al. 2010). In spite of some movements of males and females, probably related to mating, the general trend is that residency significantly increased during the study, showing that inbreeding is a potential risk as a result of the reduction of shark mobility, in particular for males. A resulting lack of genetic variability would decrease the resilience of the lemon shark population, and presents a strong case for rejecting any proposal or compromise on the part of fishers to keep alive resident sharks while harvesting the other individuals. Intra and inter-specific aggressiveness was also witnessed during the study, leading to a risk of potentially severe bites on humans, which led us to suggest a yearly closure period during the mating period in French Polynesia to decrease shark behavioural modifications owing to long-term uninterrupted shark-feeding activities (Clua et al. 2010).
Assessment of other values that comprise the Total Economic Value of sharks
In a world dominated by the concept of Homo oeconomicus (Smith 1986), it appears critical to transfer the issue of shark conservation from the ecological domain to that of the economy. In this respect, our efforts should go further than the simple calculation of the direct non-consumptive use of sharks (Fig. 1) mainly promoted by ecotourism. In that perspective, the direct consumptive use value, mainly based on fisheries, does not have to be demonstrated (see Clarke et al. 2007). Given its consumptive nature, this value is not supporting shark conservation, except for the suggestion that fisheries should be made sustainable to ensure long-term profits.
The indirect-use value (Fig. 1) mainly relies on the added value obtained by maintaining sharks in marine ecosystems. An ecosystem with apex predators leads to higher productivity and biomass (Friedlander and DeMartini 2002), and this should benefit finfish fisheries. Through underwater visual censuses conducted on sites with a gradient of densities of top predators, Sandin et al. (2010) showed that fish species targeted by sharks (and potentially by fishermen) tended to allocate more energy to reproduction than to somatic storage; this phenomenon led to a gain in biomass based on more individuals, in spite of their smaller size compared with those not subjected to shark predation. This gain of biomass, which has not yet been assessed in economic terms, could be of indirect benefit to fishers. As a complementary approach, some information is available about the cascading effects following the depletion of sharks, including the economic consequences of the process. Following the collapse of scallop stocks, which were over-consumed by cownose rays after their predators, large sharks, had disappeared from the coast of North Carolina, Myers et al. (2007) described the destruction of a century-old small-scale traditional fishery that depended on the scallops. The indirect economic consequences of either keeping sharks or making them disappear from marine ecosystems both suggest that their conservation is more economically favourable, and these consequences should be more thoroughly addressed.
In the same spirit, it would be interesting to assess the option values of sharks (Fig. 1), based for example on the potential profit linked to the discovery or commercial use of medically active substances present in their skeleton or organs. For instance, squalene is an antioxidant decreasing the probability of cancer. Its preventive effect on tumours has been clinically proven (Kelly 1999), in particular for the colon (Rao et al. 1998). Extracts from shark cartilage, already marketed under the brand name NeovastatND, inhibit tumour progression by limiting angiogenesis (Sheu et al. 1998). Shark immune systems are much more sophisticated than was previously thought and radically different from that of other vertebrates (Streltsov et al. 2004). Sharks seem to retain a non-specific immune response throughout their lives, allowing them to detoxify many potentially harmful compounds quickly without needing prior exposure. Criscitiello et al. (2010) just obtained the first cellular identification of T-Cell Receptor-expressing lymphocytes in a cartilaginous fish, the nurse shark Ginglymostoma cirratum. These studies may produce critical improvements of the mediated human immune response to viruses such as HIV. Based on these examples in the domain of health, it seems obvious that the option values of sharks should also be considered from the perspective of avoiding their extinction.
The non-use value is also far from negligible for sharks, which have a privileged status in several cultures, especially in the Pacific. The bequest value refers to the importance that current generations place on ensuring the presence of sharks to future generations. In the Melanesian culture, the shark often has the status of a totem animal which hosts the soul of ancestors; people never cull it as it holds the destiny of the tribe (Riesenfeld 1950). The existence value relies on the willingness of people to maintain sharks as part of the shared natural heritage, without any intention of using them now or in the future. A study conducted in Canada assessed the willingness by four groups of the population to pay to ensure the conservation of endangered aquatic animals, including the right whales Eubalaena spp., the leatherback turtle Dermochelys coriacea and the porbeagle shark Lamna nasus. The study showed that the youngest segment of the population, despite ranking it at the end of the list, was ready to spend about USD15 per person per year to save this shark (Rudd 2007).
Any attempt to calculate non-use values seems legitimate, and not considering these values in tandem with use values underestimates the real value of marine wildlife (Whitehead 1993). However, there is controversy about the reliability of the contingent valuation method (Cameron and Carson 1989), which appears to be the most widely used approach for assessing non-use values, as applied for example by Rudd (2007). Limitations include the subjectiveness of human perceptions involved in the process, which prevents one from being able to accurately replicate a study (Boardman et al. 2001; Bateman et al. 2002). In this context, we suggest a focus on the calculation of use values, which are less subject to criticism and are powerful enough to support objective arguments in favour of shark and other wildlife conservation.
Acknowledgements
This study benefited from the financial support of the Direction à l’Environnement (DIREN) of French Polynesia and the scientific support of the Coordination Unit of the Coral Reef Initiatives for the Pacific (CRISP Program) based in Noumea, New Caledonia. We thank the private diving company TopDive Center (now TopDive-Bathys) in Moorea for logistic support and René Galzin, Centre de Recherche Insulaire et Observatoire de l’Environnement (CRIOBE), for scientific support. We also thank the Guest Editor and the anonymous referees for very helpful comments, which improved the quality of this paper.
References
Anderson, R. C., and Ahmed, H. (1993). The shark fisheries in the Maldives. Ministry of Fisheries and Agriculture, Male, and FAO, Rome.Anderson, R. C., and Waheed, A. (1999). Management of shark fisheries in the Maldives. In ‘Case Studies of the Management of Elasmobranch Fisheries’. (Ed. R. Shotton.) FAO Fisheries Technical Paper 378/2. (FAO: Rome.)
Anonymous (1998). White Shark Exploitation in South Africa. Newsletter of the IUCN Shark Specialist Group, N°12. IUCN Shark Specialist Group, Berkshire, UK.
Bateman, I., Carson, R. T., Day, B., Hanemann, M., Hanley, N., et al. (2002). ‘Economic Valuation with Stated Preference Techniques: A Manual.’ (Edward Elgar: Cheltenham, UK.)
Baum, J. K., and Worm, B. (2009). Cascading top-down effects of changing oceanic predator abundances. Journal of Animal Ecology 78, 699–714.
| Cascading top-down effects of changing oceanic predator abundances.Crossref | GoogleScholarGoogle Scholar | 19298616PubMed |
Baum, J. K., Meyer, R. A., Kehler, D. G., Worm, B., Harley, S. J., et al. (2003). Collapse and conservation of shark populations in the Northwest Atlantic. Science 299, 389–392.
| Collapse and conservation of shark populations in the Northwest Atlantic.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD3sXjsF2kuw%3D%3D&md5=924bb8fc3c277e1376697b925d80c33aCAS | 12532016PubMed |
Boardman, A. E., Greenberg, D. H., Vining, A. R., and Weimer, D. L. (2001). ‘Cost-Benefit Analysis.’ 2nd edn. (Prentice Hall: Upper Saddle River, NJ.)
Bookbinder, M. P., Dinerstein, E., Rijal, A., Cauley, H., and Rajouria, A. (1998). Ecotourism’s support of biodiversity conservation. Conservation Biology 12, 1399–1404.
| Ecotourism’s support of biodiversity conservation.Crossref | GoogleScholarGoogle Scholar |
Brierley, A. S. (2007). Fisheries ecology: hunger for shark fin soup drives clam chowder off the menu. Current Biology 17, R555–R557.
| Fisheries ecology: hunger for shark fin soup drives clam chowder off the menu.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2sXnslWisrs%3D&md5=c52dbfe2979bd63b30537fc1ea1f088bCAS | 17637357PubMed |
Brown, C. A., and Gruber, S. H. (1988). Age assessment of the lemon shark, Negaprion brevirostris, using tetracycline validated vertebral centra. Copeia 1988, 747–753.
| Age assessment of the lemon shark, Negaprion brevirostris, using tetracycline validated vertebral centra.Crossref | GoogleScholarGoogle Scholar |
Brunnschweiler, J. M. (2009). The Shark Reef Marine Reserve: a marine tourism project in Fiji involving local communities. Journal of Sustainable Tourism 18, 1–14.
Buray, N., Mourier, J., Planes, S., and Clua, E. (2009). Underwater photo-identification of sicklefin lemon sharks, Negaprion acutidens, at Moorea (French Polynesia). Cybium 33, 21–27.
Cameron, M. C., and Carson, R. T. (1989). ‘Using Surveys to Value Public Goods: The Contingent Valuation Method.’ (Resources for the Future: Washington, DC.)
Catlin, J., Jones, T., Norman, B., and Wood, D. (2009). Consolidation in a wildlife tourism industry: the changing impact of whale shark tourist expenditure in the Ningaloo coast region. International Journal of Tourism Research 12, 134–148.
Clarke, S. C., McAllister, M. K., Milner-Gulland, E. J., Kirkwood, G. P., Michielsens, C. G. J., et al. (2006). Global estimates of shark catches using trade records from commercial markets. Ecology Letters 9, 1115–1126.
| Global estimates of shark catches using trade records from commercial markets.Crossref | GoogleScholarGoogle Scholar | 16972875PubMed |
Clarke, S. C., Milner-Gulland, E. J., and Bjørndal, T. (2007). Perspective: social, economic and regulatory drivers of the shark fin trade. Marine Resource Economics 22, 305–327.
Clua, E., Buray, N., Legendre, P., Mourier, J., and Planes, S. (2010). Behavioural response of sicklefin lemon sharks Negaprion acutidens to underwater feeding for ecotourism purposes. Marine Ecology Progress Series 414, 257–266.
| Behavioural response of sicklefin lemon sharks Negaprion acutidens to underwater feeding for ecotourism purposes.Crossref | GoogleScholarGoogle Scholar |
Compagno, L. J. V. (1984). FAO Species Catalogue. Sharks of the World. An Annotated and Illustrated Catalogue of Shark Species Known to Date: Part 2. Carcharhiniformes FAO Fisheries Synopsis 125, Vol. 4, Part 2, 251–655.
Costanza, R., d’Arge, R., de Groot, R., Farber, S., Grasso, M., et al. (1997). The value of the world’s ecosystem services and natural capital. Nature 387, 253–260.
| The value of the world’s ecosystem services and natural capital.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaK2sXjtlShtbs%3D&md5=d5cb7d6a9fdb002637f22093622cf50eCAS |
Criscitiello, M. F., Ohta, Y., Saltis, M., McKinney, E. C., and Flajnik, M. F. (2010). Evolutionarily conserved TCR binding sites, identification of T-Cells in primary lymphoid tissues, and surprising trans-rearrangements in nurse shark. Journal of Immunology 184, 6950–6960.
| Evolutionarily conserved TCR binding sites, identification of T-Cells in primary lymphoid tissues, and surprising trans-rearrangements in nurse shark.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3cXmvFejsL8%3D&md5=0c2796e6a2e7060fc27d9592935f0cbaCAS |
Davis, D., Banks, S., Birtles, A., Valentine, P., and Cuthill, M. (1997). Whale sharks in Ningaloo Marine Park: managing tourism in an Australian marine protected area. Tourism Management 18, 259–271.
| Whale sharks in Ningaloo Marine Park: managing tourism in an Australian marine protected area.Crossref | GoogleScholarGoogle Scholar |
Dicken, M. L., and Hosking, S. G. (2009). Socio-economic aspects of the tiger shark diving industry within the Aliwal Shoal Marine Protected Area, South Africa. African Journal of Marine Science 31, 227–232.
| Socio-economic aspects of the tiger shark diving industry within the Aliwal Shoal Marine Protected Area, South Africa.Crossref | GoogleScholarGoogle Scholar |
Dobson, J. (2006). Sharks, wildlife tourism, and state regulation. Tourism in Marine Environments 3, 15–23.
| Sharks, wildlife tourism, and state regulation.Crossref | GoogleScholarGoogle Scholar |
Ferretti, F., Ransom, A. M., Serena, F., and Lotze, H. K. (2008). Loss of large predatory sharks from the Mediterranean Sea. Conservation Biology 22, 952–964.
| Loss of large predatory sharks from the Mediterranean Sea.Crossref | GoogleScholarGoogle Scholar | 18544092PubMed |
Friedlander, A. M., and DeMartini, E. E. (2002). Contrasts on density, size, and biomass of reef fishes between the northwestern and the main Hawaiian islands: the effects of fishing down apex predators. Marine Ecology Progress Series 230, 253–264.
| Contrasts on density, size, and biomass of reef fishes between the northwestern and the main Hawaiian islands: the effects of fishing down apex predators.Crossref | GoogleScholarGoogle Scholar |
Garrod, B., and Wilson, J. C. (2004). Nature on the edge? Marine ecotourism in peripheral coastal areas. Journal of Ecotourism 12, 95–120.
Halpenny, E. A. (2003). NGOs as conservation agents: Achieving conservation through marine ecotourism. In ‘Marine Ecotourism: Issues and Experience’. (Eds B. Garrod and J. C. Wilson.) pp. 107–121. (Channel View Publications: Clevedon.)
Heithaus, M. R., Frid, A., Wirsing, A. J., and Worm, B. (2008). Predicting ecological consequences of marine top predator declines. Trends in Ecology & Evolution 23, 202–210.
| Predicting ecological consequences of marine top predator declines.Crossref | GoogleScholarGoogle Scholar | 18308421PubMed |
Higginbottom, K., Tribe, A., and Booth, R. (2003). Contributions of non-consumptive wildlife tourism to conservation. In ‘Nature-based Tourism, Environment and Land Management’. (Eds R. Buckley, C. Pickering and D. B.Weaver.) pp. 181–196. (CABI Publishing: London.)
Hoyt, E. (2001). ‘Whale Watching: Worldwide Tourism Numbers, Expenditures, and Expanding Socioeconomic Benefits’, pp. i–vi; 1–158. (International Fund for Animal Welfare: Yarmouth Port, MA.)
Hoyt, E. (2007). ‘A Blueprint for Dolphin and Whale Watching Development.’ (Humane Society International: Washington, DC.)
IEOM (Institut d’Emission d’Outre-Mer) (2010). La Polynésie française en 2009. (Eds Institut d’Emission d’Outre-Mer: Papeete, Polynésie Française.)
Issacs, J. C. (2000). The limited potential for ecotourism to contribute towards wildlife conservation. Wildlife Society Bulletin 28, 61–69.
IUCN (2008). 2008 IUCN Red List of Threatened Species. Available at www.iucnredlist.org [Verified 14 March 2011].
Johnston, B., and Pickering, T. (2003). The economics of aquaculture in comparison with other rural development opportunities in Pacific island countries. Marine studies technical report 2003/07. 24 pp. (University of the South Pacific, Suva.)
Kelly, G. S. (1999). Squalene and its potential clinical uses. Alternative Medicine Review 4, 29–36.
| 1:STN:280:DyaK1M7ktlekug%3D%3D&md5=5233dff5d44bc14f454ff36d6cda3becCAS | 9988781PubMed |
Martin, R. A., and Hakeem, A. A. A. (2006). Development of a sustainable shark diving ecotourism industry in the Maldives: Challenges and Opportunities. Maldives Marine Research Bulletin No 8.
MEA (Millennium Ecosystem Assessment) (2005). ‘Ecosystems and Human Well-being: Biodiversity Synthesis.’ (World Resources Institute, Island Press: Washington, DC.)
Myers, R. A., Baum, J. K., Shepherd, T. D., Powers, S. P., and Peterson, C. H. (2007). Cascading effects of the loss of apex predatory sharks from a coastal ocean. Science 315, 1846–1850.
| Cascading effects of the loss of apex predatory sharks from a coastal ocean.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2sXjsFSitrw%3D&md5=cb95aeccbb3f469fcd3ed09861c4e3b9CAS | 17395829PubMed |
Newsome, D., and Rodger, K. (2008). To feed or not to feed: a contentious issue in wildlife tourism. In ‘Too Close For Comfort: Contentious Issues in Human–Wildlife Encounters’. (Eds D. Lunney, A. Munn and W. Meikle.) pp. 255–270. (Royal Zoological Society of New South Wales: Mosman.)
Orams, M. (2002). Feeding wildlife as a tourist attraction. Tourism Management 23, 281–293.
| Feeding wildlife as a tourist attraction.Crossref | GoogleScholarGoogle Scholar |
Polidaro, B. A., Livingstone, S. R., Carpenter, K. E., Hutchinson, B., Mast, R. B., et al. (2008). Status of the World’s Marine Species. Available at: http://iucn.org/about/work/programmes/species/red_list/review/ [Verified 14 March 2011].
Rao, C. V., Newmark, H. L., and Reddy, B. S. (1998). Chemopreventive effect of squalene on colon cancer. Carcinogenesis 19, 287–290.
| Chemopreventive effect of squalene on colon cancer.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaK1cXhs1ykurc%3D&md5=d26902a02423c58677dfc9ddc180b7d1CAS | 9498278PubMed |
Riesenfeld, A. (1950). ‘The Megalithic Culture of Melanesia.’ (E. J. Brill: Leiden.)
Rowat, D., and Engelhardt, U. (2007). Seychelles: a case study of community involvement in the development of whale shark ecotourism and its socio–economic impact. Fisheries Research 84, 109–113.
| Seychelles: a case study of community involvement in the development of whale shark ecotourism and its socio–economic impact.Crossref | GoogleScholarGoogle Scholar |
Rudd, M. A. (2007). Public preferences and willingness to pay for aquatic species at risk conservation programs in Canada. SWGC Environmental Valuation and Policy Laboratory. EVPL Working Paper 07–001. (Memorial University of Newfoundland, Canada.)
Sandin, S. A., Walsh, S. M., and Jackson, J. B. C. (2010). Prey release, trophic cascades, and phase shifts in tropical nearshore marine ecosystems. In ‘Trophic Cascades: Predators, Prey, and the Changing Dynamics of Nature’. (Eds J. Terborgh and J. A. Estes.) pp. 71–90. (Island Press: Washington, DC.)
Sergio, F., Newton, I., Marchesi, L., and Pedrini, P. (2006). Ecological justified charisma: preservation of top predators delivers biodiversity conservation. Journal of Applied Ecology 43, 1049–1055.
| Ecological justified charisma: preservation of top predators delivers biodiversity conservation.Crossref | GoogleScholarGoogle Scholar |
Sheu, J. R., Fu, C. C., Tsai, M. L., and Chung, W. J. (1998). Effect of U-995, a potent shark cartilage-derived angiogenesis inhibitor, on anti-angiogenesis and anti-tumor activities. Anticancer Research 18, 4435–4441.
| 1:CAS:528:DyaK1MXmvVahug%3D%3D&md5=932b217bedb9b908d382fadf4c494f0fCAS | 9891506PubMed |
Smith, A. (1986). ‘The Wealth of Nations.’ Books I–III. (Penguin Classics: New York.)
Stevens, J., Bonfil, R., Dulvy, N., and Walker, P. (2000). The effects of fishing on sharks, rays, and chimaeras (chondrichthyans), and the implications for marine ecosystems. ICES Journal of Marine Science 57, 476–494.
| The effects of fishing on sharks, rays, and chimaeras (chondrichthyans), and the implications for marine ecosystems.Crossref | GoogleScholarGoogle Scholar |
Streltsov, V. A., Varghese, J. N., Carmichael, J. A., Irving, R. A., Hudson, P. J., et al. (2004). Structural evidence for evolution of shark Ig new antigen receptor variable domain antibodies from a cell-surface receptor. Proceedings of the National Academy of Science of the United States of America 101, 12 444–12 449.
| Structural evidence for evolution of shark Ig new antigen receptor variable domain antibodies from a cell-surface receptor.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2cXnsVens7w%3D&md5=265183c503cf95c7008643a0cd45bfbeCAS |
Topelko, K. N., and Dearden, P. (2005). The shark watching industry and its potential contribution to shark conservation. Journal of Ecotourism 4, 108–128.
| The shark watching industry and its potential contribution to shark conservation.Crossref | GoogleScholarGoogle Scholar |
Walker, T. I. (2002). Review of fisheries and processes impacting shark populations of the world. In ‘Elasmobranch biodiversity, conservation and management: proceedings of the International Seminar and Workshop, Sabah, Malaysia 1997’, pp. 220–229. (Eds S. L. Fowler, T. M. Reed and F. A. Dipper.) Occasional Paper of the IUCN Species Survival Commission 25, IUCN/SSC Shark Specialist Group, IUCN, Gland, Switzerland.
Whitehead, J. C. (1993). Total economic values for coastal and marine wildlife: specification, validity, and valuation issues. Marine Resource Economics 8, 119–132.
Williams, A. M., and Montanari, A. (1999). Sustainability and self-regulation: critical perspectives. Tourism Geographies 1, 26–40.
| Sustainability and self-regulation: critical perspectives.Crossref | GoogleScholarGoogle Scholar |
Wilson, J. (2003). Planning policy issues for marine ecotourism. In ‘Marine Ecotourism: Issues and Experiences’. (Eds B. Garrod and J. Wilson.) pp. 48–65. (Channel View Publications: Clevedon.)