

Identifying crucial gaps in our knowledge of the life-history of avian influenza viruses – an Australian perspective
Marcel Klaassen A B C , Bethany J. Hoye B and David A. Roshier AA Centre for Integrative Ecology, School of Life and Environmental Sciences, Deakin University, Locked Bag 20000, Geelong, VIC 3220, Australia.
B Netherlands Institute of Ecology (NIOO-KNAW), Department of Animal Ecology, PO Box 50, 6700 AB Wageningen, the Netherlands.
C Corresponding author. Email: marcel.klaassen@deakin.edu.au
Emu 111(2) 103-112 https://doi.org/10.1071/MU10042
Submitted: 2 June 2010 Accepted: 22 November 2010 Published: 3 May 2011
Abstract
We review our current knowledge of the epidemiology and ecology of avian influenza viruses (AIVs) in Australia in relation to the ecology of their hosts. Understanding the transmission and maintenance of low-pathogenic avian influenza (LPAI) viruses deserves scientific scrutiny because some of these may evolve to a high-pathogenic AIV (HPAI) phenotype. That the HPAI H5N1 has not been detected in Australia is thought to be a result of the low level of migratory connectivity between Asia and Australia. Some AIV strains are endemic to Australia, with Australian birds acting as a reservoir for these viruses. However, given the phylogenetic relationships between Australian and Eurasian strains, both avian migrants and resident birds within the continent must play a role in the ecology and epidemiology of AIVs in Australia. The extent to which individual variation in susceptibility to infection, previous infections, and behavioural changes in response to infection determine AIV epidemiology is little understood. Prevalence of AIVs among Australian avifauna is apparently low but, given their specific ecology and Australian conditions, prevalence may be higher in little-researched species and under specific environmental conditions.
Additional keywords: ecology, epidemiology, host species, HPAI, LPAI, migration, wild birds.
Introduction
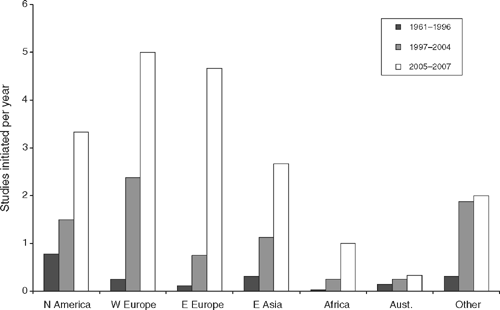
The presence of avian influenza viruses (AIVs; see Box 1 for background information) in wild populations of Australian birds has been known since the early 1970s (see Downie and Laver 1973). The first studies into the occurrence of avian influenza viruses found a wide variety of subtypes. However, prevalence appeared low in comparison to that of North America and Europe (Sims and Turner 2008). This low prevalence, in combination with relatively few outbreaks in poultry in comparison to South-East Asia, North America and Europe (Alexander 2007b), has resulted in AIV research in Australia being focussed on external sources of infection, notably from Asia (Tracey et al. 2004; East et al. 2008a, 2008b; McCallum et al. 2008; Sims and Turner 2008). Outbreaks of high-pathogenic AIV (HPAI) H5N1 in South-east Asia from 1996, and subsequent intercontinental spread of this strain (see http://www.who.int/csr/disease/avian_influenza/en/) has further served to reinforce the outward focus of Australian research. Although transport of poultry and other live birds and their products have been identified as the main cause of dispersal of AIVs, the role of wild migratory birds remains contentious (Van Borm et al. 2005; Kilpatrick et al. 2006; Feare 2007; Gauthier-Clerc et al. 2007). Several studies have reported temporal and spatial overlap between global H5N1 outbreaks and bird migration patterns (e.g. Gilbert et al. 2006; Si et al. 2009) and some species held under laboratory conditions have been seen to shed these viruses without overt clinical symptoms (Keawcharoen et al. 2008). Yet most isolations of HPAI viruses in wild birds are from dead or moribund individuals, with only three reported cases of HPAI H5N1 from apparently healthy birds (Feare 2010). In fact, the death of wild birds as a result of HPAI infection remains one of the hallmarks of local incursions of the disease for the poultry industry (Feare 2007; Gauthier-Clerc et al. 2007). In the wake of such speculation there has been a surge in surveillance of wild birds as an early warning system (Spackman 2009; Hoye et al. 2010b) and increased interest in the ecology of AIVs in wild birds globally over the last decade, although Australia remains underrepresented (Fig. 1).
| Box 1. A brief introduction to avian influenza viruses and immunology |
| Influenza viruses belong to a group of RNA viruses called the Orthomyxoviridae. There are three types of influenza viruses: A, B and C. Type B and C only infect humans, whereas influenza A viruses have been found to infect a wide range of host species, including humans, domestic livestock, other mammals, and a wide variety of domesticated and wild birds. Given that influenza A viruses have been found in over 100 species of wild birds across the globe, it is generally accepted that wild birds form the principal natural reservoir (hence ‘avian influenza viruses’), and that these viruses may, on occasion, be transmitted from this reservoir to other hosts such as domestic birds, livestock, marine mammals and humans, either directly or indirectly (Webster et al. 1992). |
| Avian influenza viruses (AIVs) are classified on the basis of the antigenic properties of two surface proteins: haemagglutinin (HA) and neuraminidase (NA). There are currently 16 HA (H1 to H16) and 9 NA (N1 to N9) known antigenic subtypes, and the majority of possible combinations of these have been detected in wild birds (Fouchier et al. 2005; Olsen et al. 2006). In addition, AIVs are further classified on the basis of their ability to cause disease in Chickens. Highly pathogenic avian influenza (HPAI) viruses are virulent viruses that induce mortality in up to 100% of infected Chickens (Alexander 2000). All other AIVs cause much milder disease, and are designated low-pathogenic avian influenza (LPAI) virus status. Only AIVs of subtypes H5 and H7 have been found to have the potential to become highly pathogenic and cause outbreaks of HPAI. Yet importantly, not all viruses of the H5 and H7 subtypes cause highly pathogenic disease. To date, HPAI isolates have been obtained primarily from domesticated ducks and poultry, with a few notable isolations from dead or moribund wild birds (Alexander 2007a). A virus’s pathogenicity may also show marked variation between host species. A virus that causes severe clinical signs and high mortality in Chickens may not induce the same effect in other bird species or mammals (Isoda et al. 2006) and the effects may even differ between closely related wild bird species (Keawcharoen et al. 2008). |
| Replication of LPAI viruses is thought to be restricted to the gastrointestinal and respiratory tracts, sampled by swabbing the cloaca or droppings and the oropharynx respectively (Brown and Stallknecht 2008). HPAI viruses, however, are able to replicate throughout the bird in clinical cases of the disease, causing systemic organ and tissue damage (Alexander 2000). Under laboratory conditions, birds infected with LPAI will shed virus for between 1 and 2 weeks (Homme and Easterday 1970; Kida et al. 1980), though the period of viral shedding appears to be significantly shorter when the individual has previously experienced an AIV (Kida et al. 1980; Fereidouni et al. 2009; Latorre-Margalef et al. 2009; Jourdain et al. 2010). The transmission of viruses between avian hosts is not well understood but there are various potential routes. A faecal – oral route is one of the most generally assumed routes, but infection via aerosols and perhaps even cloacal drinking may occur (Fouchier and Munster 2009). |
| Three surface proteins of influenza A viruses (HA, NA and matrix 2) induce a specific immune response in the host. The host may also produce antibodies to some internal proteins, especially nucleoprotein (NP) and matrix 1 proteins. Unlike the surface proteins, these internal proteins have a highly conserved genetic sequence, which allows the detection of antibodies from birds infected with any influenza A virus (Suarez and Schultz-Cherry 2000). Antibody titre has been seen to increase rapidly as a result of re-infections, and is thought to be responsible for the decreased shedding period seen in re-infected individuals (Kida et al. 1980; Fereidouni et al. 2009, 2010; Jourdain et al. 2010). However, there is very little knowledge on how long these antibodies persist, with current estimates somewhere between a few months to a year (Nishiura et al. 2009; Fereidouni et al. 2010; Hoye et al. 2010a). |
|
In light of these concerns, Sims (2006) drafted an agenda for future AIV research in Australia. Many of these recommendations do not directly relate to wild birds but those that do call for a focus on the ecology and epidemiology of AIVs within Australia and the region, as well as HPAI H5N1 in South-east Asia. Here we use Sims’ recommendations to structure our review of the current knowledge regarding life-history of AIVs in wild birds. Specifically:
-
Which AIVs are found in Australian wild birds?
-
What role does avian migration play in the long-distance dispersal of AIVs to Australia?
-
What effect might AIV infection have on transmission-relevant host behaviour?
-
How are AIVs transmitted among wild birds, and between wild birds and poultry?
Topics that were not addressed by Sims (2006) but that we feel should be included are:
-
What role do resident birds play in the ecology and epidemiology of AIVs in Australia?
-
What are the temporal and spatial dynamics of infection?
-
Does infection occur at random within a population or only in certain individuals, and if so, what effect does this have on transmission and maintenance?
There are notable differences in the biology and ecology of many Australian birds compared with northern hemisphere species, in which most research on AIVs has been conducted. As such, we pursue an Australian perspective where current knowledge and hypotheses regarding the epidemiology and ecology of AIVs in Australia is reviewed in light of the special characteristics of the Australian avian hosts and their environment.
Which AIVs are found in Australian wild birds?
Of all the LPAI subtypes identified in wild birds from other parts of the world, most have also been found in Australia (Downie and Laver 1973; Downie et al. 1977; Mackenzie et al. 1984, 1985; Nestorowicz et al. 1987; Peroulis and O’Riley 2004; Hurt et al. 2006; Haynes et al. 2009), including subtypes H5 and H7 (which have potential for HPAI phenotype; see Box 1). Studies on AIV gene segments isolated from wild birds in Australia indicate these viruses maintain phylogenetic relationships with those from Eurasia and, to a lesser extent, North America (Donis et al. 1989; Banks et al. 2000; Kishida et al. 2008). However, there is also evidence for the existence of distinct Australian sublineages of AIVs, suggesting that the maintenance host or hosts of at least some AIV types reside in Australia (Banks et al. 2000; Bulach et al. 2010). There is little information on the degree to which H5 subtypes found in Australia are related to the recent HPAI strains in Asia, or on the rate of exchange of viral material between the two continents. To resolve the rate of exchange of AIVs between Australia and other parts of the world, whole genome sequencing and subsequent phylogenetic analyses are required and are now being conducted (Bulach et al. 2010).
What role does avian migration play in the long-distance dispersal of AIVs to Australia?
Considerable focus has been placed on the role of avian migrants in the ecology and epidemiology of AIVs globally. Given the apparent similarity of viral diversity to the rest of Eurasia (see above) it may be prudent to assume avian movements and migration to Australia play a role in the ecology and epidemiology of AIVs on the continent.
Less clear is in which bird groups AIV of Eurasian origin could enter Australia and how it would subsequently spread within the Australian avifauna. Aquatic birds, such as waterfowl (Anseriformes) and shorebirds (Charadriiformes), are thought to be the natural reservoir for LPAI, primarily because AIVs are susceptible to desiccation but persist well in aquatic environments (Stallknecht et al. 1990; Ito et al. 1995). Australia sits at the southern end of the East Asian–Australasian Flyway but only 35 or so species of shorebird and seven species of landbird, from an avifauna numbering more than 700 species, are regular long-distance migrants between Eurasia and Australia (Palaearctic migrants) (Dingle 2004). Some of these migrants spend part of the year in areas of South-east Asia where HPAI H5N1 has become endemic (Tracey et al. 2004). Although the possibility remains that migratory shorebirds are a vector for AIVs to Australia, the evidence to date suggests the frequency of such ingress is very low (see Haynes et al. 2009).
Of the many Palaearctic anseriforms, only the Northern Shoveler (Anas clypeata), Northern Pintail (Anas acuta) and Garganey (Anas querquedula) occur but they are either vagrants or uncommon, irregular migrants to the Australo-Papuan region (Beehler et al. 1986; Marchant and Higgins 1990; Simpson and Day 2010). The vast floodplains and coastal swamps of southern New Guinea are a focal point for Palaearctic migrants that make it to the region (Bishop 2006), as well as large numbers of Australo-Papuan waterfowl (Halse et al. 1996), providing the opportunity for transmission between Palaearctic and Australo-Papuan species. In this context, common tropical species such as Wandering Whistling-Duck (Dendrocygna arcuata) and Magpie Goose (Anseranas semipalmata) may act as a bridge between hemispheres for emerging infectious diseases when they mix with temperate species for which the tropics are extralimital (see McCallum et al. 2008). However, there is no evidence yet to suggest that interspecific contact is a mode of transmission for dispersal of AIVs through the archipelagos to Australia’s north. Analyses of migration tracks of waterfowl, in combination with data from experimentally inoculated birds in laboratory conditions and the period for which they remain asymptomatic, suggests that intercontinental dispersal of HPAI H5N1 is dependent on successive infections of individual birds along the migration path (Gaidet et al. 2010). Moreover, it was estimated that the period for which an infected individual could disperse HPAI H5N1 more than 500 km was on average only 5–15 days per year. Although the potential to disperse HPAI H5N1 long distances remains if infection coincides with the migration period, more likely is the relay infection of LPAI viruses that circulate at high prevalence in migratory waterfowl (Gaidet et al. 2010). Thus, there is a critical lack of understanding of the interplay between the ecology of AIVs and the movement ecology of their avian hosts.
What role do resident birds play in the ecology and epidemiology of AIVs in Australia?
Initially, AIV research in Australia concentrated on endemic strains of AIVs (Sims and Turner 2008). With the advent of HPAI H5N1 outbreaks in Asia, the focus of research shifted primarily to monitoring H5 prevalence among wild birds, and refining our knowledge of the itineraries of birds migrating between Asia and Australia (Tracey et al. 2004; McCallum et al. 2008; Haynes et al. 2009). However, LPAI viruses of certain haemagglutinin (HA) subtypes may switch to an HPAI phenotype (Box 1) and within-continental strains may therefore also be a source of new HPAI strains, as illustrated by the 1976 outbreaks in poultry (Sims and Turner 2008). As such, understanding transmission and maintenance of AIVs among wild birds and between wild birds and poultry within Australia deserves scientific scrutiny (Box 2).
| Box 2. Australian drought cycles, waterfowl population dynamics, and HPAI outbreaks in poultry |
| Climatic conditions in Australia are characterised by periods of intense rainfall followed by periods of intense drought. During the wet, waterfowl numbers increase with many serologically naïve juveniles entering the population. During the subsequent period of drought, bird densities increase on the few remaining wetlands (Kingsford and Norman 2002; Chambers and Loyn 2006; Kingsford et al. 2010; Norman and Chambers 2010). We hypothesise that it is during this period of increasing densities of serologically naïve birds that AIV prevalence increases dramatically within wild populations and provide the source of infection in domestic poultry flocks. Australia has experienced five outbreaks of HPAI in poultry since 1976 all of which were caused by H7 viruses (Selleck et al. 2003; Hamilton et al. 2009), for which the ancestral reservoir is hypothesised to reside within Australia (Bulach et al. 2010). Following the introduction of LPAI viruses (H7 or H5) into poultry flocks via wild birds, conditions in poultry farms then provide ideal conditions for development of HPAI strains (Alexander 2007a). In at least two of the five outbreaks, direct contact with Australian wild waterfowl was implicated as the source of the outbreak (Selleck et al. 2003; Hamilton et al. 2009). Besides direct contact between waterfowl and poultry, intermediate avian hosts from a wide range of bird species that have high contact rates with both freshwater habitats and poultry may play a role in the transmission of these viruses between waterfowl and poultry (Olsen et al. 2006; Gronesova et al. 2008; Peterson et al. 2008; Hoye et al. 2010b). |
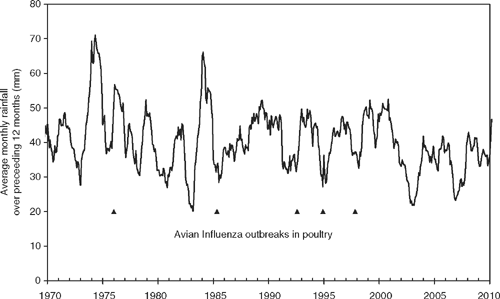
| A highly evocative correlation (Fig. 2) exists between outbreaks of HPAI in poultry in south-eastern Australia and rainfall (average monthly rainfall over the preceding 12 months) across the Murray–Darling Basin and strongly suggests outbreaks occur during periods of drought (when one expects waterfowl to concentrate on remaining wetlands) following a wet period (in which populations have grown and many birds may be immunologically naïve). |
|
Viral prevalence in Australian host species appears to be low in comparison to findings from related species in North America and Europe (Sims and Turner 2008). However, the available data for the Australian continent is spatially, temporally and phylogenetically patchy. Given this heterogeneity, and the modest number of samples taken within any location, time period or species (Haynes et al. 2009) it is extremely difficult to conclude that prevalence is in fact lower than that seen in Eurasia and North America. Further, the estimates of prevalence in North America and Europe may be upwardly biased owing to sampling being conducted at specific times and locations and in particular species and individuals in order to maximise the number of positive samples (Spackman 2009; Hoye et al. 2010b).
Sampling in Australia (and worldwide) has targeted Anseriformes and Charadriiformes (Senne 2003; Arzey 2004a, 2004b, 2005; Bunn 2004; Tracey et al. 2004; Turner 2004; East et al. 2008a, 2008b; McCallum et al. 2008; Sims and Turner 2008; Hamilton et al. 2009), despite LPAI viruses having been found to infect more than 100 host species, in 26 families, including many whose primary habitat is not aquatic (Olsen et al. 2006). Indeed, the question yet to be addressed is which species are reservoirs or temporary hosts for these viruses in Australia. Research into patterns of occurrence of antibodies to AIVs may provide a way to resolve this issue (see below).
Any species or avian group that have either high levels of viral prevalence, long infectious periods or behaviour that induces high inter specific and intraspecific rates of contact may be candidates for the maintenance of AIVs (Nishiura et al. 2009). Contact rates are highly dependent on density and, therefore, birds that regularly congregate in large numbers, such as many waterbirds, are of considerable interest in our efforts to understand the ecology of AIVs in birds. In Australia there are also species outside of the oft-tested Anseriformes and Charadriiformes orders that congregate in large numbers, including some parrots (e.g. Budgerigar (Melopsittacus undulatus) and Little Corella (Cacatua sanguinea)) and Australian Pelicans (Pelecanus conspicillatus) that congregate in the arid interior to breed following major rainfall events (Kingsford and Norman 2002; Reid 2009), as well as many pelagic seabirds that are colonial in behaviour and have also been found to be hosts of influenza viruses (Laver and Webster 1972; Downie and Laver 1973; Downie et al. 1977).
Many Australian species differ from related species at northern temperate latitudes in the nature and extent of their annual movements. Nonetheless, some species could be considered ecologically equivalent with respect to habitat requirements, feeding ecology, social behaviour and dispersal distances (e.g. waterfowl: Briggs 1992; Kingsford et al. 2010). Even if some species are not strictly LPAI reservoir hosts, they may be temporary hosts and may also transmit any introduced HPAI strains (Mackenzie et al. 1984). Thus the transmission and maintenance of AIVs within the Australian avian community, including the rate of contact (and viral exchange) between species deserves further study.
In light of the differences found among northern temperate duck species (Box 1), the susceptibility of a range of Australian species to HPAI H5N1 also requires investigation. It is of particular interest to ascertain whether some wild birds are resistant to the effects of the viruses and, if so, why, as recommended by Sims (2006). To date only one such study exists, in Emus (Dromaius novaehollandiae), which, unlike many waterfowl, showed no marked clinical signs upon infection with a HPAI (Heckert et al. 1999).
What are the temporal and spatial dynamics of infection?
An added complication to the assessment of AIV diversity, and the role of intercontinental migrants, nomads and residents in the dispersal and maintenance of these viruses, is seasonality of infection. Seasonal changes in the prevalence of infection are a common phenomenon among infectious diseases, both in humans and wildlife (Altizer et al. 2006). Several studies in northern temperate biomes have suggested that the frequency of AIV infections in wild birds may also exhibit seasonal fluctuations, with a yearly peak in late summer and early autumn, followed by low prevalence during the winter period (Hinshaw et al. 1980; Krauss et al. 2004; Munster et al. 2007; Wallensten et al. 2007).
Three mechanisms have been postulated to drive seasonal variation in infection as seen in North America and western Europe. First, an increased number of infected individuals may, in part, be a result of the pulsed entering of immunologically naïve young into the population at the end of summer (Hinshaw et al. 1985). Second, the intensity of population congregations of migratory birds often reaches its annual peak during migration or shortly after arrival on the wintering grounds, most notably in waterbirds. Finally, events within the annual cycle of birds may alter the susceptibility of individual hosts, such as changes in immune defences resulting from energetic trade-offs with moult, breeding or long-distance migration (Hasselquist 2007; Buehler et al. 2008; Martin II et al. 2008). Marked differences in viral prevalence have also been found between years (e.g. Krauss et al. 2004). These patterns are little understood, but a periodic variation in immunity has been implicated (Hinshaw et al. 1985).
However, such generalisations are based on a small number of species (migratory dabbling ducks, Anatinae), sampled predominantly in autumn at specific locations. The behaviour of many Australian species, including species already known to experience infection with AIV, is spatially and temporally more diverse than the seasonally driven migration patterns seen in other parts of the world (Kingsford and Norman 2002) where the majority of AIV research has been conducted. As noted by the Food and Agriculture Organisation (FAO 2007) ‘northern hemisphere stereotypes regarding migration in waterfowl and many other water bird species do not [necessarily] apply to southern hemisphere species. South African and Australian waterfowl tend to be nomadic, their movements dictated by available food supplies and rainfall, rather than truly migratory’. Yet, although the realised patterns of movement and migration are markedly different, the feeding and breeding biology of many Australian species are remarkably similar to that of their northern temperate counterparts (Briggs 1992). Some species are closely related to northern counterparts, such as the Pacific Black Duck (Anas superciliosa) and Northern Mallard (Anas platyrhynchos), and are likely to differ little in their susceptibility and transmission of AIVs. Current interpretation of infection dynamics in Australia is also hampered by a restricted spatial coverage of sampling owing to the large areas used by nomadic species that move across the whole continent in response to resource availability (Tracey 2010). There are also sampling problems with temporal coverage because movements of the most susceptible species are not as seasonally predictable as in other biomes. Further, in light of the somewhat sporadic (opportunistic) nature of most Australian surveillance to date, and because virus tends to be shed for only a matter of days, we cannot disregard the possibility that peak infection events have not been sampled.
Since sampling large numbers of wild birds is logistically challenging in a dynamic, event-driven system, as a first step, Sims (2006) called for longitudinal studies of viral prevalence in an Australian setting, where prevalence is systematically measured at regular intervals. This approach offers an opportunity to dissect seasonal and biological drivers of infection dynamics. It would ascertain whether infection follows seasonal cycles or behavioural cycles such as congregation and introduction of naïve individuals.
The specific environmental conditions in which birds reside may also play an important role in their exposure to infection and their ability to maintain infection within the population. For instance, open marine environments and the Arctic tundra have been hypothesised to be characterised by a particularly low density and diversity of pathogens (Piersma 1997; Mendes et al. 2005). Given that environmental transmission in aquatic environments is considered critical to the maintenance of infection within the wild bird population (Breban et al. 2009; Rohani et al. 2009), species inhabiting aquatic habitats have received most interest. In Australia, many such habitats are ephemeral (Roshier et al. 2001) and concentrations of waterbirds temporary (Kingsford et al. 2010). Drought–rainfall cycles in Australia are non-seasonal, running over several years and result in major fluctuations in aquatic bird population dynamics and densities beyond what we see in seasonal environments (Box 2).
Although there are several hypotheses as to why viral prevalence may vary over time and space (as above), firm data on temporal and spatial variation are currently lacking, notably outside North America and Europe. Therefore, in addition to establishing the spatial and temporal extent of variation in AIV prevalence in Australia, there is also the prospect that the contrasting environmental and ecological conditions in Australia could be a model system to help unravel the underlying mechanisms responsible for these patterns globally.
Does infection occur at random or only in certain individuals?
The risk of infection may not only vary spatially and temporally, but also between individuals. Contact rates and infection risks also depend on the susceptibility and the behaviour of individual birds, which may vary. It is well established that different personalities with concomitant behavioural characteristics exist in birds (Drent et al. 2003), which might have an effect on their sociality and contact rates. Condition and immune function may also vary across individuals (e.g. Ots et al. 1998; Forsman et al. 2008). In particular, prior exposure to an HA-homologous virus of LPAI phenotype has been shown to be sufficient to reduce clinical symptoms and viral shedding (Fereidouni et al. 2009). Even prior infection with an HA-heterologous virus may result in partially reduced symptoms and viral shedding (Fereidouni et al. 2009; Jourdain et al. 2010). Therefore, infection history may importantly determine an individual’s susceptibility to HPAI viruses. Whereas it is unlikely for migratory birds clinically affected by HPAI viruses to disperse HPAI over large distances (e.g. from Asia to Australia), the phenomenon of partial cross-protective immunity increases the potential for migrants to disperse HPAI over large distances. To date, most infection experiments have been conducted with seronegative birds; the studies by Fereidouni et al. (2009) and Jourdain et al. (2010) highlight the importance of including infection history in the design of experiments.
What effect might infection have on transmission-relevant behaviour by hosts?
Transmission-relevant behaviour may be different between infected and non-infected birds, either as a result of the infection or because underlying differences may make some individuals more susceptible to infection in the first place. Infection with HPAI viruses can induce severe clinical signs that may have a considerable effect on the rate of transmission of the infections. Also LPAI infections lacking clinical signs may nevertheless have an effect on the behaviour of free-living birds and, thus, transmission rate (van Gils et al. 2007; Latorre-Margalef et al. 2009). Further, the number of secondary infections caused by one infected individual (related to the amount of virus being shed and the duration of infection) may differ between individuals owing to variation in condition, immune function, infection history and age, although this has received little empirical attention (Sims 2006; Costa et al. 2010; see above). Most contemporary epidemiological models assume a single transmission coefficient and, as such, assume uniform contact rates, susceptibility and infectiousness across all individuals in a population, but it is increasingly realised that this may be a too coarse an approximation (McCallum et al. 2001; Beldomenico and Begon 2010). It is of considerable importance that more empirical data on individual variations in transmission become available to develop parameters for more sophisticated epidemiological models.
How are influenza viruses transmitted among wild birds, and between wild birds and poultry?
We are still in the very early stages of understanding the epidemiology of AIVs among wild birds (e.g. Nishiura et al. 2009) and currently rely on many assumptions regarding the mechanisms of transmission and the AIV-specific immunobiology of the species involved. Even basic knowledge, such as contact-rates between conspecifics and heterospecifics and the mechanisms of transmission (see Box 1) are lacking. How long previously infected birds remain immune from re-infection by homologous and heterologous AIV subtypes is likewise poorly known (Fereidouni et al. 2010).
Globally, few studies have examined the extent to which links exist between the circulation of LPAI viruses in wild bird and poultry. Even the temporal and spatial overlap of strains isolated from poultry and wild birds has received little attention (e.g. Halvorson et al. 1985; Hinshaw et al. 1986). Because of the lack of knowledge on exchange rates of viruses between wildlife and poultry, the fear of such exchange is great. Given the economic consequences and risks to public health associated with AIV outbreaks in poultry, the quest for an understanding of the environmental drivers (Box 2) for these outbreaks is ongoing. Currently, risk assessment relies on geographical information systems (GIS) based analyses, combining bird-banding and avian count-data with the distribution of poultry farms (Cumming et al. 2008; East et al. 2008a, 2008b). Although these analyses identify the potential for wildlife–poultry contacts, they do not take into account actual contact rates and pathways for infection. To our knowledge, no study has measured these, nor examined them in the context of the spatial and temporal dynamics of wild bird movements.
Challenges to the investigation of AIVs in Australian wild birds
Five years after Sims drafted his agenda for Australia’s influenza research (Sims 2006), and despite strong political impetus for understanding AIVs in Australian wild birds, we are still just scratching the surface. Many gaps remain in our understanding of AIV epidemiology globally and the gaps are notably wider in this part of the world. Since the outbreak of HPAI H5N1 in Asia, many birds have been screened in Australia but these have mainly served to confirm earlier findings implying waterfowl and shorebirds being the main carriers of AIVs and prevalence levels being low. Furthermore, the insight into the movements of some target species has notably increased with the development of satellite tracking technologies and advances in inferring migratory connectivity using molecular markers. But these movement studies mostly lack concomitant data on viral or antibody dynamics. In addition, many potential AIV host species may have been overlooked. There is an irrefutable need for directed ornithological and virological studies in light of specific scientific questions, and for interdisciplinary collaboration, so that advances in our understanding of avian movements feed into studies of host-range, contacts and infection dynamics and vice versa.
One of the greatest limitations to the study of LPAI viruses in wild birds is the short period during which an individual may be infected. Thus large numbers of birds must be sampled in order to find evidence of infection in most populations. Because there are often difficulties in obtaining such large quantities of birds one alternative is to use methods that expand the period of time in which infection can be detected, that is to investigate individual infection history, as indicated by antibodies to AIVs. Certain antibodies may be detected for several months after infection (see Box 1). Prevalence levels of antibodies are thus considerably higher than prevalence levels of virus and investigation of seroprevalence is potentially an effective means to screen for potential host species (Hoye et al. 2010b). Beyond the identification of host species, analysis of seroprevalence, combined with information on contemporary infection (sampling virus directly) can shed considerable light on the temporal dynamics of infection (Hoye et al. 2010a). Laboratory kits for this type of work have been developed for poultry and some species of waterfowl. Because not all species of bird may develop identical antibodies to AIVs, these kits need further laboratory and field validation to be useful for the study of a wider range of birds (Brown et al. 2010).
Given the low density of researchers, the remoteness of most locations where large numbers of birds reside, the erratic temporal dynamics in the movements of some target (waterbird) species and the short period of time during which infected birds shed virions, it is extremely difficult to obtain a holistic view of the composition of the viral community within Australia. Establishing a continent-wide early warning system for potentially hazardous AIV types within Australia also seems impossible. A more effective approach to further our understanding of AIV dynamics within Australia and potential threats to wildlife, poultry and humans alike, is highly focussed, hypothesis-driven research targeted at unravelling AIV host–pathogen and host–host interactions. For example, rather than sampling migrant and resident species everywhere they occur, one could sample before and after different groups come into contact with one another to determine sources and sinks of LPAI viruses (Langstaff et al. 2009). Preferably, future research should also be complementary to research done elsewhere, focussing on specific pathways and mechanisms for the maintenance of AIV infections and their relationships to specific environmental and ecological conditions prevailing in Australia.
Such studies are not only of immediate importance in understanding the ecology and epidemiology of AIVs and other zoonotic diseases for agricultural and human health. Progress in this field of research also contributes to building our fundamental understanding of wildlife disease ecology in general. Many pathogenic microorganisms are carried by birds that, by their nature, may disperse these pathogens over wide areas (Hubalek 2004). In order to evaluate the risks involved, and ultimately develop mitigation strategies, there is great need for enhanced knowledge on the interactive ecology of birds and bird-mediated pathogens. Resolving the many gaps in our knowledge identified above is also of great interest from a fundamental evolutionary ecological perspective since diseases are considered to play a key role in the evolution of life histories of organisms, potentially having great effects on their fitness.
Acknowledgements
We thank two anonymous reviewers for their helpful and stimulating comments on an earlier version of the manuscript. This study was supported through the Bird Health program within the International Polar Year by the Netherlands Organisation for Scientific Research (grant no. 851.40.073), Intramural Research Program of the National Institutes of Health (contract NIAIDNIH HHSN266200700010C), and the Australia Research Council (LP07775076).
References
Alexander, D. J. (2000). A review of avian influenza in different bird species. Veterinary Microbiology 74, 3–13.| A review of avian influenza in different bird species.Crossref | GoogleScholarGoogle Scholar | 1:STN:280:DC%2BD3c3mtF2isg%3D%3D&md5=3f463e0ba545c58ccb3a7b66a4ff1655CAS | 10799774PubMed |
Alexander, D. J. (2007a). An overview of the epidemiology of avian influenza. Vaccine 25, 5637–5644.
| An overview of the epidemiology of avian influenza.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2sXnslWns7w%3D&md5=d1734346c880910079b4336752a6b46cCAS | 17126960PubMed |
Alexander, D. J. (2007b). Summary of avian influenza activity in Europe, Asia, Africa, and Australasia, 2002–2006. Avian Diseases 51, 161–166.
| Summary of avian influenza activity in Europe, Asia, Africa, and Australasia, 2002–2006.Crossref | GoogleScholarGoogle Scholar | 17494548PubMed |
Altizer, S., Dobson, A., Hosseini, P., Hudson, P., Pascual, M., and Rohani, P. (2006). Seasonality and the dynamics of infectious diseases. Ecology Letters 9, 467–484.
| Seasonality and the dynamics of infectious diseases.Crossref | GoogleScholarGoogle Scholar | 16623732PubMed |
Arzey, G. (2004a). The role of wild aquatic birds in the epidemiology of avian influenza in Australia. Australian Veterinary Journal 82, 377–378.
| The role of wild aquatic birds in the epidemiology of avian influenza in Australia.Crossref | GoogleScholarGoogle Scholar | 1:STN:280:DC%2BD2czmsVSlug%3D%3D&md5=4312ccc84adcbcaa89d81264e2ee20a7CAS | 15267098PubMed |
Arzey, G. (2004b). The role of wild aquatic birds in the epidemiology of avian influenza in Australia – Reply. Australian Veterinary Journal 82, 644–645.
| The role of wild aquatic birds in the epidemiology of avian influenza in Australia – Reply.Crossref | GoogleScholarGoogle Scholar | 15887393PubMed |
Arzey, G. (2005). The role of wild waterfowl in the epidemiology of AI in Australia. Australian Veterinary Journal 83, 445.
| The role of wild waterfowl in the epidemiology of AI in Australia.Crossref | GoogleScholarGoogle Scholar | 1:STN:280:DC%2BD2MznslKksw%3D%3D&md5=7e64c7decad30036184e8a3ab11c2952CAS | 16035188PubMed |
Banks, J., Speidel, E. C., McCauley, J. W., and Alexander, D. J. (2000). Phylogenetic analysis of H7 haemagglutinin subtype influenza A viruses. Archives of Virology 145, 1047–1058.
| Phylogenetic analysis of H7 haemagglutinin subtype influenza A viruses.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD3cXksVeksLk%3D&md5=0558c120c3f8cccccc806710088c48aeCAS | 10881690PubMed |
Beehler, B., Pratt, T., and Zimmerman, D. (1986). ‘Birds of New Guinea.’ (Princeton University Press: Princeton, NJ.)
Beldomenico, P. M., and Begon, M. (2010). Disease spread, susceptibility and infection intensity: vicious circles? Trends in Ecology & Evolution 25, 21–27.
| Disease spread, susceptibility and infection intensity: vicious circles?Crossref | GoogleScholarGoogle Scholar |
Bishop, K.D. (2006). A review of the avifauna of the TransFly Eco-region: the status, distribution, habitats and conservation of the region’s birds. South Pacific Program of World Wildlife Fund, Report No. 9S0739.02.
Breban, R., Drake, J. M., Stallknecht, D. E., and Rohani, P. (2009). The role of environmental transmission in recurrent avian influenza epidemics. PLoS Computational Biology 5, e1000346.
| The role of environmental transmission in recurrent avian influenza epidemics.Crossref | GoogleScholarGoogle Scholar | 19360126PubMed |
Briggs, S. V. (1992). Movement patterns and breeding characteristics of arid zone ducks. Corella 16, 15–22.
Brown, J. D., and Stallknecht, D. E. (2008). Wild bird surveillance for the avian influenza virus. In ‘Methods in Molecular Biology. Vol. 436’. (Ed. E. Spackman.) pp. 85–97. (Humana Press: Totowa, NJ.)
Brown, J. D., Luttrell, M. P., Berghaus, R. D., Kistler, W., Keeler, S. P., Howey, A., Wilcox, B., Hall, J., Niles, L., Dey, A., Knutsen, G., Fritz, K., and Stallknecht, D. E. (2010). Prevalence of antibodies to type A influenza virus in wild avian species using two serologic assays. Journal of Wildlife Diseases 46, 896–911.
| 20688695PubMed |
Buehler, D. M., Piersma, T., Matson, K., and Tieleman, B. I. (2008). Seasonal redistribution of immune function in a migrant shorebird: annual-cycle effects override adjustments to thermal regime. American Naturalist 172, 783–796.
| Seasonal redistribution of immune function in a migrant shorebird: annual-cycle effects override adjustments to thermal regime.Crossref | GoogleScholarGoogle Scholar | 18999941PubMed |
Bulach, D., Halpin, R., Spiro, D., Pomeroy, L., Janies, D., and Boyle, D. B. (2010). Molecular analysis of H7 avian influenza viruses from Australia and New Zealand: genetic diversity and relationships from 1976 to 2007. Journal of Virology 84, 9957–9966.
| Molecular analysis of H7 avian influenza viruses from Australia and New Zealand: genetic diversity and relationships from 1976 to 2007.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3cXhtlarsrbJ&md5=dec5b4cad3d2e7538a65d37abd74397cCAS | 20668069PubMed |
Bunn, C. M. (2004). The role of wild aquatic birds in the epidemiology of avian influenza in Australia. Australian Veterinary Journal 82, 644.
| The role of wild aquatic birds in the epidemiology of avian influenza in Australia.Crossref | GoogleScholarGoogle Scholar | 1:STN:280:DC%2BD2M3kvVemsA%3D%3D&md5=062cce705bc2688a4f7474c8f2c62c44CAS | 15887393PubMed |
Chambers, L. E., and Loyn, R. H. (2006). The influence of climate variability on numbers of three waterbird species in Western Port, Victoria, 1973–2002. International Journal of Biometeorology 50, 292–304.
| The influence of climate variability on numbers of three waterbird species in Western Port, Victoria, 1973–2002.Crossref | GoogleScholarGoogle Scholar | 16435107PubMed |
Costa, T. P., Brown, J. D., Howerth, E. W., and Stallknecht, D. E. (2010). The effect of age on avian influenza viral shedding in Mallards (Anas platyrhynchos). Avian Diseases 54, 581–585.
| The effect of age on avian influenza viral shedding in Mallards (Anas platyrhynchos).Crossref | GoogleScholarGoogle Scholar | 1:STN:280:DC%2BC3czpsF2qug%3D%3D&md5=373145d2ab696c17698d2fadf88034d0CAS | 20521698PubMed |
Cumming, G. S., Hockey, P. A. R., Bruinzeel, L. W., and Du Plessis, M. A. (2008). Wild bird movements and avian influenza risk mapping in southern Africa. Ecology and Society 13, 26.
Dingle, H. (2004). The Australo-Papuan bird migration system: another consequence of Wallace’s Line. Emu 104, 95–108.
| The Australo-Papuan bird migration system: another consequence of Wallace’s Line.Crossref | GoogleScholarGoogle Scholar |
Donis, R. O., Bean, W. J., Kawaoka, Y., and Webster, R. G. (1989). Distinct lineages of influenza-virus H4 hemagglutinin genes in different regions of the world. Virology 169, 408–417.
| Distinct lineages of influenza-virus H4 hemagglutinin genes in different regions of the world.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaL1MXlt1OrsL0%3D&md5=751422c17a05b97eb7256fd2740cb1b5CAS | 2705304PubMed |
Downie, J. C., and Laver, W. G. (1973). Isolation of a type A influenza-virus from an Australian pelagic bird. Virology 51, 259–269.
| Isolation of a type A influenza-virus from an Australian pelagic bird.Crossref | GoogleScholarGoogle Scholar | 1:STN:280:DyaE3s7ksFWksg%3D%3D&md5=1f00d6452157995ba3224791fd5b3353CAS | 4632652PubMed |
Downie, J. C., Hinshaw, V., and Laver, W. G. (1977). Ecology of influenza-isolation of type-A influenza viruses from Australian pelagic birds. Australian Journal of Experimental Biology and Medical Science 55, 635–643.
| Ecology of influenza-isolation of type-A influenza viruses from Australian pelagic birds.Crossref | GoogleScholarGoogle Scholar | 1:STN:280:DyaE1c3htVagsw%3D%3D&md5=2340c3a27d9d7a43d20a9704789b4ad2CAS | 614832PubMed |
Drent, P. J., van Oers, K., and van Noordwijk, A. J. (2003). Realized heritability of personalities in the Great Tit (Parus major). Proceedings of the Royal Society of London. Series B. Biological Sciences 270, 45–51.
| Realized heritability of personalities in the Great Tit (Parus major).Crossref | GoogleScholarGoogle Scholar |
East, I. J., Hamilton, S., and Garner, G. (2008a). Identifying areas of Australia at risk of H5N1 avian influenza infection from exposure to migratory birds: a spatial analysis. Geospatial Health 2, 203–213.
| 18686269PubMed |
East, I. J., Hamilton, S. A., Sharp, L. A., and Garner, M. G. (2008b). Identifying areas of Australia at risk for H5N1 avian influenza infection from exposure to nomadic waterfowl moving throughout the Australo-Papuan region. Geospatial Health 3, 17–27.
| 19021105PubMed |
FAO (2007). ‘Wildbirds and Avian Influenza: An Introduction to Applied Field Research and Disease Sampling Techniques.’ (Food and Agriculture Organization: Rome.)
Feare, C. J. (2007). The spread of avian influenza. Ibis 149, 424–425.
| The spread of avian influenza.Crossref | GoogleScholarGoogle Scholar |
Feare, C. J. (2010). Role of wild birds in the spread of highly pathogenic avian influenza virus H5N1 and implications for global surveillance. Avian Diseases 54, 201–212.
| Role of wild birds in the spread of highly pathogenic avian influenza virus H5N1 and implications for global surveillance.Crossref | GoogleScholarGoogle Scholar | 20521633PubMed |
Fereidouni, S. R., Starick, E., Beer, M., Wilking, H., Kalthoff, D., Grund, C., Häuslaigner, R., Breithaupt, A., Lange, E., and Harder, T. C. (2009). Highly pathogenic avian influenza virus infection of Mallards with homo- and heterosubtypic immunity induced by low pathogenic avian influenza viruses. PLoS ONE 4, e6706.
| Highly pathogenic avian influenza virus infection of Mallards with homo- and heterosubtypic immunity induced by low pathogenic avian influenza viruses.Crossref | GoogleScholarGoogle Scholar | 19693268PubMed |
Fereidouni, S. R., Grund, C., Haeuslaigner, R., Lange, E., Wilking, H., Harder, T. C., Beer, M., and Starick, E. (2010). Dynamics of specific antibody responses induced in Mallards after infection by or immunization with low pathogenicity avian influenza viruses. Avian Diseases 54, 79–85.
| Dynamics of specific antibody responses induced in Mallards after infection by or immunization with low pathogenicity avian influenza viruses.Crossref | GoogleScholarGoogle Scholar | 20408403PubMed |
Forsman, A. M., Vogel, L. A., Sakaluk, S. K., Grindstaff, J. L., and Thompson, C. F. (2008). Immune-challenged House Wren broods differ in the relative strengths of their responses among different axes of the immune system. Journal of Evolutionary Biology 21, 873–878.
| Immune-challenged House Wren broods differ in the relative strengths of their responses among different axes of the immune system.Crossref | GoogleScholarGoogle Scholar | 1:STN:280:DC%2BD1c3kvVOhuw%3D%3D&md5=d444e8356f60239a98923998b7ea7a28CAS | 18298497PubMed |
Fouchier, R. A. M., and Munster, V. J. (2009). Epidemiology of low pathogenic avian influenza viruses in wild birds. Revue Scientifique Et Technique-Office International Des Epizooties 28, 49–58.
| 1:STN:280:DC%2BD1Mvps1Ckuw%3D%3D&md5=f0c2a398481497f9f5ae80f0a11679c5CAS |
Fouchier, R., Munster, V. J., Wallensten, A., Bestebroer, T. M., Herfst, S., Smith, D. L., Rimmelzwaan, G., Olsen, B., and Osterhaus, A. (2005). Characterisation of a novel influenza A virus hemagglutinin subtype (H16) obtained from Black-headed Gulls. Journal of Virology 79, 2814–2822.
| Characterisation of a novel influenza A virus hemagglutinin subtype (H16) obtained from Black-headed Gulls.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2MXitlaqurs%3D&md5=18bb265c01b48a07314b864505e4f778CAS | 15709000PubMed |
Gaidet, N., Cappelle, J., Takekawa, J. Y., Prosser, D. J., Iverson, S. A., Douglas, D. C., Perry, W. M., Mundkur, T., and Newman, S. H. (2010). Potential spread of highly pathogenic avian influenza H5N1 by wildfowl: dispersal ranges and rates determined from large-scale satellite telemetry. Journal of Applied Ecology 47, 1147–1157.
| Potential spread of highly pathogenic avian influenza H5N1 by wildfowl: dispersal ranges and rates determined from large-scale satellite telemetry.Crossref | GoogleScholarGoogle Scholar |
Gauthier-Clerc, M., Lebarbenchon, C., and Thomas, F. (2007). Recent expansion of highly pathogenic avian influenza H5N1: a critical review. Ibis 149, 202–214.
| Recent expansion of highly pathogenic avian influenza H5N1: a critical review.Crossref | GoogleScholarGoogle Scholar |
Gilbert, M., Xiao, X. M., Domenech, J., Lubroth, J., Martin, V., and Slingenbergh, J. (2006). Anatidae migration in the western Palearctic and spread of highly pathogenic avian influenza H5N1 virus. Emerging Infectious Diseases 12, 1650–1656.
| 17283613PubMed |
Gronesova, P., Kabat, P., Trnka, A., and Betakova, T. (2008). Using nested RT-PCR analyses to determine the prevalence of avian influenza viruses in passerines in western Slovakia, during summer 2007. Scandinavian Journal of Infectious Diseases 40, 954–957.
| Using nested RT-PCR analyses to determine the prevalence of avian influenza viruses in passerines in western Slovakia, during summer 2007.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD1cXhtl2qtL7M&md5=40a09523acced9e3b517efe9c5fd711eCAS | 18777249PubMed |
Halse, S. A., Pearson, G. B., Jaensch, R. P., Kulmoi, P., Gregory, P., Kay, W. R., and Storey, A. W. (1996). Waterbird surveys of the Middle Fly River floodplain, Papua New Guinea. Wildlife Research 23, 557–569.
| Waterbird surveys of the Middle Fly River floodplain, Papua New Guinea.Crossref | GoogleScholarGoogle Scholar |
Halvorson, D. A., Kelleher, C. J., and Senne, D. A. (1985). Epizootiology of avian influenza – effect of season on incidence in sentinel ducks and domestic Turkeys in Minnesota. Applied and Environmental Microbiology 49, 914–919.
| 1:STN:280:DyaL2M3isVWhsQ%3D%3D&md5=8c0e62e962934e4fc0e49bbb286c6e77CAS | 4004223PubMed |
Hamilton, S. A., East, I. J., Toribio, J. A., and Garner, M. G. (2009). Are the Australian poultry industries vulnerable to large outbreaks of highly pathogenic avian influenza? Australian Veterinary Journal 87, 165–174.
| Are the Australian poultry industries vulnerable to large outbreaks of highly pathogenic avian influenza?Crossref | GoogleScholarGoogle Scholar | 19382922PubMed |
Hasselquist, D. (2007). Comparative immunoecology in birds: hypotheses and tests. Journal of Ornithology 148, S571–S582.
| Comparative immunoecology in birds: hypotheses and tests.Crossref | GoogleScholarGoogle Scholar |
Haynes, L., Arzey, E., Bell, C., Buchanan, N., Burgess, G., Cronan, V., Dickason, C., Field, H., Gibbs, S., Hansbro, P. M., Hollingsworth, T., Hurt, A. C., Kirkland, P., McCracken, H., O’Connor, J., Tracey, J., Wallner, J., Warner, S., Woods, R., and Bunn, C. (2009). Australian surveillance for avian influenza viruses in wild birds between July 2005 and June 2007 Australian Veterinary Journal 87, 266–272.
| Australian surveillance for avian influenza viruses in wild birds between July 2005 and June 2007Crossref | GoogleScholarGoogle Scholar | 1:STN:280:DC%2BD1MvmtlCmtw%3D%3D&md5=bfbc4eb25486f44f5cf35311284e1c02CAS | 19573149PubMed |
Heckert, R. A., McIsacc, M., Chan, M., and Zhou, E.-M. (1999). Experimental infection of Emus (Dromaiius novaehollandiae) with avian influenza viruses of varying virulence: clinical signs, virus shedding and serology. Avian Pathology 28, 13–16.
| Experimental infection of Emus (Dromaiius novaehollandiae) with avian influenza viruses of varying virulence: clinical signs, virus shedding and serology.Crossref | GoogleScholarGoogle Scholar | 1:STN:280:DC%2BD2MvnslOmsw%3D%3D&md5=7faf85cc987015e9d59df4aff4f436d1CAS | 16147545PubMed |
Hinshaw, V. S., Webster, R. G., and Turner, B. (1980). The perpetuation of orthomyxoviruses and paramyxoviruses in Canadian waterfowl. Canadian Journal of Microbiology 26, 622–629.
| The perpetuation of orthomyxoviruses and paramyxoviruses in Canadian waterfowl.Crossref | GoogleScholarGoogle Scholar | 1:STN:280:DyaL3c3ktlOqsA%3D%3D&md5=38077d7a16aa2e87e0bb250660257c76CAS | 7397605PubMed |
Hinshaw, V. S., Wood, J. M., Webster, R. G., Deibel, R., and Turner, B. (1985). Circulation of influenza-viruses and paramyxoviruses in waterfowl originating from two different areas of North America. Bulletin of the World Health Organization 63, 711–719.
| 1:STN:280:DyaL287gs1GntA%3D%3D&md5=2b41dbedb2a19f85501aebaad49369c5CAS | 3878741PubMed |
Hinshaw, V. S., Nettles, V. F., Schorr, L. F., Wood, J. M., and Webster, R. G. (1986). Influenza-virus surveillance in waterfowl in Pennsylvania after the H5N2 avian outbreak. Avian Diseases 30, 207–212.
| Influenza-virus surveillance in waterfowl in Pennsylvania after the H5N2 avian outbreak.Crossref | GoogleScholarGoogle Scholar | 1:STN:280:DyaL283mvVyisQ%3D%3D&md5=90de5bc8a691dcb001171af883b2db84CAS | 3015104PubMed |
Homme, P. J., and Easterday, B. C. (1970). Avian influenza virus infections. 4. Response of pheasants, ducks, and geese to influenza A/Turkey/Wisconsin/1966 Virus. Avian Diseases 14, 285–290.
| Avian influenza virus infections. 4. Response of pheasants, ducks, and geese to influenza A/Turkey/Wisconsin/1966 Virus.Crossref | GoogleScholarGoogle Scholar | 1:STN:280:DyaE3c3jtVSlsw%3D%3D&md5=b1551b5b2b275dadf165b8a30bf3c5b3CAS | 5427235PubMed |
Hoye, B. J., Munster, V. J., Nishiura, H., Fouchier, R. A. M., Madsen, J., and Klaassen, M. (2010a). Reconstructing an annual cycle of interaction: natural infection and antibody dynamics to avian influenza along a migratory flyway. Oikos , .
| Reconstructing an annual cycle of interaction: natural infection and antibody dynamics to avian influenza along a migratory flyway.Crossref | GoogleScholarGoogle Scholar |
Hoye, B. J., Munster, V. J., Nishiura, H., Klaassen, M., and Fouchier, R. A. M. (2010b). Surveillance of wild birds for avian influenza virus. Emerging Infectious Diseases 16, 1827–1834.
| 21122209PubMed |
Hubalek, Z. (2004). An annotated checklist of pathogenic microorganisms associated with migratory birds. Journal of Wildlife Diseases 40, 639–659.
| 15650082PubMed |
Hurt, A. C., Hansbro, P. M., Selleck, P., Olsen, B., Minton, C., Hampson, A. W., and Barr, I. G. (2006). Isolation of avian influenza viruses from two different transhemispheric migratory shorebird species in Australia. Archives of Virology 151, 2301–2309.
| Isolation of avian influenza viruses from two different transhemispheric migratory shorebird species in Australia.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD28XhtFSnsLzK&md5=68f069234b47bed6ccf508b26c1ef55aCAS | 16732420PubMed |
Isoda, N., Sakoda, Y., Kishida, N., Bai, G. R., Matsuda, K., Umemura, T., and Kida, H. (2006). Pathogenicity of a highly pathogenic avian influenza virus, A/chicken/Yamaguchi/7/04 (H5N1) in different species of birds and mammals. Archives of Virology 151, 1267–1279.
| Pathogenicity of a highly pathogenic avian influenza virus, A/chicken/Yamaguchi/7/04 (H5N1) in different species of birds and mammals.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD28XmtFGqurc%3D&md5=7b32fd506522162648b600715ae42458CAS | 16502281PubMed |
Ito, T., Okazaki, K., Kawaoka, Y., Takada, A., Webster, R. G., and Kida, H. (1995). Perpetuation of influenza-A viruses in Alaskan waterfowl reservoirs. Archives of Virology 140, 1163–1172.
| Perpetuation of influenza-A viruses in Alaskan waterfowl reservoirs.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaK2MXns1WjtL0%3D&md5=8ce99a426228b3a888562397eb1afda6CAS | 7646350PubMed |
Jourdain, E., Gunnarsson, G., Wahlgren, J., Latorre-Margalef, N., Brojer, C., Sahlin, S., Svensson, L., Waldenstrom, J., Lundkvist, A., and Olsen, B. (2010). Influenza virus in a natural host, the Mallard: experimental infection data. PLoS ONE 5, e8935.
| Influenza virus in a natural host, the Mallard: experimental infection data.Crossref | GoogleScholarGoogle Scholar | 20126617PubMed |
Keawcharoen, J., van Riel, D., van Amerongen, G., Bestebroer, T., Beyer, W. E., van Lavieren, R., Osterhaus, A. D. M. E., Fouchier, R. A. M., and Kuiken, T. (2008). Wild ducks as long-distance vectors of highly pathogenic avian influenza virus (H5N1). Emerging Infectious Diseases 14, 600–607.
| Wild ducks as long-distance vectors of highly pathogenic avian influenza virus (H5N1).Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD1cXltFamtL4%3D&md5=5b8652804170a6b2456cf83c2e7947baCAS | 18394278PubMed |
Kida, H., Yanagawa, R., and Matsuoka, Y. (1980). Duck influenza lacking evidence of disease signs and immune-response. Infection and Immunity 30, 547–553.
| 1:STN:280:DyaL3M%2FmvVKgsg%3D%3D&md5=eaec10e88ff80e443092b3648dd1c3f0CAS | 7439994PubMed |
Kilpatrick, A. M., Chmura, A. A., Gibbons, D. W., Fleischer, R. C., Marra, P. P., and Daszak, P. (2006). Predicting the global spread of H5N1 avian influenza. Proceedings of the National Academy of Sciences of the United States of America 103, 19 368–19 373.
| Predicting the global spread of H5N1 avian influenza.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2sXhs1ylug%3D%3D&md5=81f2b1baf597e78fb5cf765066320f37CAS |
Kingsford, R. T., and Norman, F. I. (2002). Australian waterbirds – products of the continent’s ecology. Emu 102, 47–69.
| Australian waterbirds – products of the continent’s ecology.Crossref | GoogleScholarGoogle Scholar |
Kingsford, R. T., Roshier, D. A., and Porter, J. L. (2010). Australian waterbirds – time and space travellers in dynamic desert landscapes. Marine and Freshwater Research 61, 875–884.
| Australian waterbirds – time and space travellers in dynamic desert landscapes.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3cXhtVansL%2FI&md5=49f8afb7500c57938acb8ba6ce1fa7e3CAS |
Kishida, N., Sakoda, Y., Shiromoto, M., Bai, G. R., Isoda, N., Takada, A., Laver, G., and Kida, H. (2008). H2N5 influenza virus isolates from terns in Australia: genetic reassortants between those of the Eurasian and American lineages. Virus Genes 37, 16–21.
| H2N5 influenza virus isolates from terns in Australia: genetic reassortants between those of the Eurasian and American lineages.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD1cXntFKmurg%3D&md5=c16c2a88625bd625bb85e53e9fac4eabCAS | 18454312PubMed |
Krauss, S., Walker, D., Pryor, S. P., Niles, L., Li, C. H., Hinshaw, V. S., and Webster, R. G. (2004). Influenza A viruses of migrating wild aquatic birds in North America. Vector Borne and Zoonotic Diseases (Larchmont, N.Y.) 4, 177–189.
| Influenza A viruses of migrating wild aquatic birds in North America.Crossref | GoogleScholarGoogle Scholar | 15631061PubMed |
Langstaff, I. G., McKenzie, J. S., Stanislawek, W. L., Reed, C. E. M., Poland, R., and Cork, S. C. (2009). Surveillance for highly pathogenic avian influenza in migratory shorebirds at the terminus of the East Asian–Australasian Flyway. New Zealand Veterinary Journal 57, 160–165.
| Surveillance for highly pathogenic avian influenza in migratory shorebirds at the terminus of the East Asian–Australasian Flyway.Crossref | GoogleScholarGoogle Scholar | 1:STN:280:DC%2BD1MvhsVGjtw%3D%3D&md5=469f55d4db1d1293ce89d35bb8cd6e11CAS | 19521465PubMed |
Latorre-Margalef, N., Gunnarsson, G., Munster, V. J., Fouchier, R. A. M., Osterhaus, A., Elmberg, J., Olsen, B., Wallensten, A., Haemig, P. D., Fransson, T., Brudin, L., and Waldenstrom, J. (2009). Effects of influenza A virus infection on migrating mallard ducks. Proceedings of the Royal Society of London. Series B. Biological Sciences 276, 1029–1036.
| Effects of influenza A virus infection on migrating mallard ducks.Crossref | GoogleScholarGoogle Scholar |
Laver, W. G., and Webster, R. G. (1972). Antibodies to human influenza-virus neuraminidase (a/Asian/57 H2n2 Strain) in sera from Australian pelagic birds. Bulletin of the World Health Organization 47, 535–541.
| 1:STN:280:DyaE3s7osVWitg%3D%3D&md5=344b2efd20e3c97ded0cd477f9598d1fCAS | 4541008PubMed |
Mackenzie, J. S., Edwards, E. C., Holmes, R. M., and Hinshaw, V. S. (1984). Isolation of ortho- and paramoxyviruses from wild birds in Western Australia, and the characterisation of novel influenza A viruses. Australian Journal of Experimental Biology and Medical Science 62, 89–99.
| Isolation of ortho- and paramoxyviruses from wild birds in Western Australia, and the characterisation of novel influenza A viruses.Crossref | GoogleScholarGoogle Scholar | 6430260PubMed |
Mackenzie, J. S., Britten, D., Hinshaw, V., and Wood, J. (1985). Isolation of avian influenza and paramyxoviruses from wild birds in Western Australia. In ‘Veterinary Viral Diseases: Their Significance in South-east Asia and the Western Pacific’. (Ed. A. J. Delaa-Porta.) pp. 336–339. (Academic Press: Orlando, FL.)
Marchant, S., and Higgins, P. (Eds) (1990). ‘Handbook of Australian, New Zealand and Antarctic Birds. Vol. 1: Ratites to Ducks.’ (Oxford University Press: Melbourne.)
Martin, L. B., Weil, Z. M., and Nelson, R. J. (2008). Seasonal changes in vertebrate immune activity: mediation by physiological trade-offs. Philosophical Transactions of the Royal Society of London. Series B. Biological Sciences 363, 321–339.
| Seasonal changes in vertebrate immune activity: mediation by physiological trade-offs.Crossref | GoogleScholarGoogle Scholar |
McCallum, H., Barlow, N., and Hone, J. (2001). How should pathogen transmission be modelled? Trends in Ecology & Evolution 16, 295–300.
| How should pathogen transmission be modelled?Crossref | GoogleScholarGoogle Scholar |
McCallum, H. I., Roshier, D. A., Tracey, J. P., Joseph, L., and Heinsohn, R. (2008). Will Wallace’s Line save Australia from avian influenza? Ecology and Society 13, 41.
Mendes, L., Piersma, T., Lecoq, M., Spaans, B., and Ricklefs, R. E. (2005). Disease-limited distributions? Contrasts in the prevalence of avian malaria in shorebird species using marine and freshwater habitats. Oikos 109, 396–404.
| Disease-limited distributions? Contrasts in the prevalence of avian malaria in shorebird species using marine and freshwater habitats.Crossref | GoogleScholarGoogle Scholar |
Munster, V. J., Baas, C., Lexmond, P., Waldenström, J., Wallensten, A., Fransson, T., Rimmelzwaan, G. F., Beyer, W. E. P., Schutten, M., Olsen, B., Osterhaus, A. D. M. E., and Fouchier, R. A. M. (2007). Spatial, temporal, and species variation in prevalence of influenza A viruses in wild migratory birds. PLoS Pathogens 3, e61.
| Spatial, temporal, and species variation in prevalence of influenza A viruses in wild migratory birds.Crossref | GoogleScholarGoogle Scholar | 17500589PubMed |
Nestorowicz, A., Kawaoka, Y., Bean, W. J., and Webster, R. G. (1987). Molecular analysis of the hemagglutinin genes of Australian H7N7 influenza-viruses – role of passerine birds in maintenance or transmission. Virology 160, 411–418.
| Molecular analysis of the hemagglutinin genes of Australian H7N7 influenza-viruses – role of passerine birds in maintenance or transmission.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaL1cXmvFOktg%3D%3D&md5=b4f80b42259a088f56474335e82c6a2eCAS | 3660587PubMed |
Nishiura, H., Hoye, B., Klaassen, M., Bauer, S., and Heesterbeek, H. (2009). How to find natural reservoir hosts from endemic prevalence in a multi-host population: a study of influenza in waterfowl. Epidemics 1, 118–128.
| How to find natural reservoir hosts from endemic prevalence in a multi-host population: a study of influenza in waterfowl.Crossref | GoogleScholarGoogle Scholar | 21352759PubMed |
Norman, F. I., and Chambers, L. E. (2010). Counts of selected duck species at Corner Inlet, Victoria: changes in relation to local and distant meteorological variations. International Journal of Biometeorology 54, 269–282.
| Counts of selected duck species at Corner Inlet, Victoria: changes in relation to local and distant meteorological variations.Crossref | GoogleScholarGoogle Scholar | 19937455PubMed |
Olsen, B., Munster, V. J., Wallensten, A., Waldenstrom, J., Osterhaus, A., and Fouchier, R. (2006). Global patterns of influenza A virus in wild birds. Science 312, 384–388.
| Global patterns of influenza A virus in wild birds.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD28XjslSktbc%3D&md5=f2014d26adbf0a8bd95f424e66e72dd2CAS | 16627734PubMed |
Ots, I., Murumagi, A., and Horak, P. (1998). Haematological health state indices of reproducing Great Tits: methodology and sources of natural variation. Functional Ecology 12, 700–707.
| Haematological health state indices of reproducing Great Tits: methodology and sources of natural variation.Crossref | GoogleScholarGoogle Scholar |
Peroulis, I., and O’Riley, K. (2004). Detection of avian paramyxoviruses and influenza viruses amongst wild bird populations in Victoria. Australian Veterinary Journal 82, 79–82.
| Detection of avian paramyxoviruses and influenza viruses amongst wild bird populations in Victoria.Crossref | GoogleScholarGoogle Scholar | 1:STN:280:DC%2BD2c7pt12ktA%3D%3D&md5=eadb8bac3ce8ff241241951381c34f91CAS | 15088965PubMed |
Peterson, A. T., Bush, S. E., Spackman, E., Swayne, D. E., and Ip, H. S. (2008). Influenza A virus infections in land birds, People’s Republic of China. Emerging Infectious Diseases 14, 1644–1646.
| Influenza A virus infections in land birds, People’s Republic of China.Crossref | GoogleScholarGoogle Scholar | 18826836PubMed |
Piersma, T. (1997). Do global patterns of habitat use and migration strategies co-evolve with relative investments in immunocompetence due to spatial variation in parasite pressure? Oikos 80, 623–631.
| Do global patterns of habitat use and migration strategies co-evolve with relative investments in immunocompetence due to spatial variation in parasite pressure?Crossref | GoogleScholarGoogle Scholar |
Reid, J. (2009) Australian Pelican: flexible responses to uncertainty. In ‘Boom and Bust – Bird Stories for a Dry Country’. (Eds L. Robin, R. Heinsohn and L. Joseph.) pp. 95–120. (CSIRO Publishing: Melbourne.)
Rohani, P., Breban, R., Stallknecht, D. E., and Drake, J. M. (2009). Environmental transmission of low pathogenicity avian influenza viruses and its implications for pathogen invasion. Proceedings of the National Academy of Sciences of the United States of America 106, 10 365–10 369.
| Environmental transmission of low pathogenicity avian influenza viruses and its implications for pathogen invasion.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD1MXot1Gnsbc%3D&md5=12ca99a2b031554bbf86fd7a2d56771eCAS |
Roshier, D. A., Whetton, P. H., Allan, R. J., and Robertson, A. I. (2001). Distribution and persistence of temporary wetland habitats in arid Australia in relation to climate. Austral Ecology 26, 371–384.
| Distribution and persistence of temporary wetland habitats in arid Australia in relation to climate.Crossref | GoogleScholarGoogle Scholar |
Selleck, P. W., Arzey, G., Kirkland, P. D., Reece, R. L., Gould, A. R., Daniels, P. W., and Westbury, H. A. (2003). An outbreak of highly pathogenic avian influenza in Australia in 1997 caused by an H7N4 virus. Avian Diseases 47, 806–811.
| An outbreak of highly pathogenic avian influenza in Australia in 1997 caused by an H7N4 virus.Crossref | GoogleScholarGoogle Scholar | 1:STN:280:DC%2BD3svpslOnsw%3D%3D&md5=b80d693268da70096790f363aeb023f0CAS | 14575068PubMed |
Senne, D. A. (2003). Avian influenza in the Western Hemisphere including the Pacific Islands and Australia. Avian Diseases 47, 798–805.
| Avian influenza in the Western Hemisphere including the Pacific Islands and Australia.Crossref | GoogleScholarGoogle Scholar | 1:STN:280:DC%2BD3svpslOnsg%3D%3D&md5=a5f8c8bbb558ab2671b81743e5acb3d6CAS | 14575067PubMed |
Si, Y. L., Skidmore, A. K., Wang, T. J., de Boer, W. F., Debba, P., Toxopeus, A. G., Li, L., and Prins, H. H. T. (2009). Spatio-temporal dynamics of global H5N1 outbreaks match bird migration patterns. Geospatial Health 4, 65–78.
| 19908191PubMed |
Simpson, K., and Day, N. (2010). ‘Field Guide to the Birds of Australia.’ 8th edn. (Viking: Melbourne.)
Sims, L. D. (2006). A risk-based approach to avian influenza virus research priorities. (Australian Biosecurity Cooperative Research Centre: Manuda, QLD.) Available at http://www.abcrc.org.au/UPLOADS/PUBLICATIONS/PUBLICATION_250.pdf [Verified 5 April 2011].
Sims, L. D., and Turner, A. J. (2008). Avian influenza in Australia. In ‘Avian Influenza’. (Ed. D. E. Swayne.) pp. 239–250. (Blackwell: Ames, IA.)
Spackman, E. (2009). The ecology of avian influenza virus in wild birds: what does this mean for poultry? Poultry Science 88, 847–850.
| The ecology of avian influenza virus in wild birds: what does this mean for poultry?Crossref | GoogleScholarGoogle Scholar | 1:STN:280:DC%2BD1M7pvF2ktQ%3D%3D&md5=ad874c667ae96f0e753163989e6e4fe3CAS | 19276433PubMed |
Stallknecht, D. E., Kearney, M. T., Shane, S. M., and Zwank, P. J. (1990). Effects of pH, temperature, and salinity on persistence of avian influenza viruses in water. Avian Diseases 34, 412–418.
| Effects of pH, temperature, and salinity on persistence of avian influenza viruses in water.Crossref | GoogleScholarGoogle Scholar | 1:STN:280:DyaK3czhsV2juw%3D%3D&md5=e5dc1fe0fb488e01ebbb63250ed3c9d8CAS | 2142421PubMed |
Suarez, D. L., and Schultz-Cherry, S. (2000). Immunology of avian influenza virus: a review. Developmental and Comparative Immunology 24, 269–283.
| Immunology of avian influenza virus: a review.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD3cXisleisb8%3D&md5=99c694895119ca9598315f3f74e4b1bfCAS | 10717293PubMed |
Tracey, J. P. (2010). Risk-based surveillance of avian influenza in Australia’s wild birds. Wildlife Research 37, 134–144.
| Risk-based surveillance of avian influenza in Australia’s wild birds.Crossref | GoogleScholarGoogle Scholar |
Tracey, J. P., Woods, R., Roshier, D., West, P., and Saunders, G. R. (2004). The role of wild birds in the transmission of avian influenza for Australia: an ecological perspective. Emu 104, 109–124.
| The role of wild birds in the transmission of avian influenza for Australia: an ecological perspective.Crossref | GoogleScholarGoogle Scholar |
Turner, A. J. (2004). The role of wild aquatic birds in the epidemiology of avian influenza in Australia. Australian Veterinary Journal 82, 713.
| The role of wild aquatic birds in the epidemiology of avian influenza in Australia.Crossref | GoogleScholarGoogle Scholar | 1:STN:280:DC%2BD2Mzisleksg%3D%3D&md5=17e0356977dd0fd2295960892d3620f7CAS | 15977620PubMed |
Van Borm, S., Thomas, I., Hanquet, G., Lambrecht, N., Boschmans, M., Dupont, G., Decaestecker, M., Snacken, R., and van den Berg, T. (2005). Highly pathogenic H5N1 influenza virus in smuggled Thai eagles, Belgium. Emerging Infectious Diseases 11, 702–705.
| 15890123PubMed |
van Gils, J. A., Munster, V. J., Radersma, R., Liefhebber, D., Fouchier, R. A. M., and Klaassen, M. (2007). Hampered foraging and migratory performance in swans infected with low-pathogenic avain influenza A virus. PLoS ONE 2, e184.
| Hampered foraging and migratory performance in swans infected with low-pathogenic avain influenza A virus.Crossref | GoogleScholarGoogle Scholar | 17264886PubMed |
Wallensten, A., Munster, V. J., Latorre-Margalef, N., Brytting, M., Elmberg, J., Fouchier, R. A. M., Fransson, T., Haemig, P. D., Karlsson, M., Lundkvist, A., Osterhaus, A., Stervander, M., Waldenstrom, J., and Olsen, B. (2007). Surveillance of influenza A virus in migratory watefowl in northern Europe. Emerging Infectious Diseases 13, 404–411.
| Surveillance of influenza A virus in migratory watefowl in northern Europe.Crossref | GoogleScholarGoogle Scholar | 17552093PubMed |
Webster, R. G., Bean, W. J., Gorman, O. T., Chambers, T. M., and Kawaoka, Y. (1992). Evolution and ecology of influenza-A viruses. Microbiological Reviews 56, 152–179.
| 1:STN:280:DyaK383lt1OqtQ%3D%3D&md5=9a3750f4f5cf96b67fb5a8233ab16f2aCAS | 1579108PubMed |