

Sperm-mediated gene transfer
Marialuisa Lavitrano A B , Marco Busnelli A , Maria Grazia Cerrito A , Roberto Giovannoni A , Stefano Manzini A and Alessia Vargiolu AA Dipartimento di Scienze Chirurgiche e Terapia Intensiva, Università di Milano, Bicocca, 20052 Milano, Italy.
B Corresponding author. Email: marialuisa.lavitrano@unimib.it
Reproduction, Fertility and Development 18(2) 19-23 https://doi.org/10.1071/RD05124
Submitted: 21 September 2005 Accepted: 21 September 2005 Published: 14 December 2005
Abstract
Since 1989, a new method for the production of transgenic animals has been available, namely sperm-mediated gene transfer (SMGT), based on the intrinsic ability of sperm cells to bind and internalise exogenous DNA molecules and to transfer them into the oocyte at fertilisation. We first described the SMGT procedure in a small animal model, with high efficiency reported in the mouse. In addition, we successfully adapted and optimised the technique for use in large animals; it was, in fact, highly efficient in the generation of human decay accelerating factor transgenic pig lines, as well as multigene transgenic pigs in which three different reporter genes, namely enhanced green fluorescent protein, enhanced blue fluorescent protein and red fluorescent protein, were introduced. The major benefits of the SMGT technique were found to be its high efficiency, low cost and ease of use compared with other methods. Furthermore, SMGT does not require embryo handling or expensive equipment. Sperm-mediated gene transfer could also be used to generate multigene transgenic pigs that would be of benefit as large animal models for medical research, for agricultural and pharmaceutical applications and, in particular, for xenotransplantation, which requires extensive genetic manipulation of donor pigs to make them suitable for grafting to humans.
Extra keywords: sperm cells, transgenic animals.
Introduction
Transgenic technology is a potent biotechnological tool that allows the generation of genetically modified animals useful for biomedical, veterinary and agricultural applications.
Over the past 25 years, a variety of methods that allow the generation of transgenic animals have been developed (Wall 2001, 2002) and the first transgenic livestock were born 20 years ago (Hammer et al. 1985). Today, the most widely used methods for the production of transgenic farm animals are: (1) direct microinjection of foreign DNA into the pronuclei of fertilised eggs; (2) nuclear transfer using genetically modified embryonic or somatic donor cells; and (3) viral-based constructs as vectors for the introduction of exogenous DNA into embryos (Kues and Niemann 2004). These methods are afflicted by low efficiency, elevated management outlay and, last but not least, the manipulation of embryos at early stages of development. Moreover, it must also be noted that the use of retroviral vectors is affected by safety issues.
In 1989, we described a new method for the production of transgenic animals, sperm-mediated gene transfer (SMGT), which it is based on the intrinsic ability of sperm cells to bind and internalise exogenous DNA and to transfer it into the egg at fertilisation (Lavitrano et al. 1989, 1992, 1997b; Francolini et al. 1993; Zani et al. 1995), as illustrated in Fig. 1. The capacity of sperm cells to capture foreign DNA had been reported in the pioneer study of Brackett et al. (1971); nevertheless, this finding and its very important implications were ignored for approximately 20 years and rediscovered after the publication of our first paper.
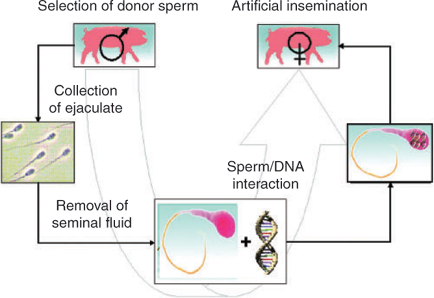
|
We first described the SMGT procedure in a small animal model, with high efficiency reported in the mouse (Lavitrano et al. 1989; Maione et al. 1998). Subsequently, we successfully adapted and optimised the technique for use in large animals; in fact, it was highly efficient for the generation of human decay accelerating factor (hDAF) transgenic pig lines (Lavitrano et al. 1997a, 2002, 2003), as well as multigene transgenic pigs, by simultaneously introducing three reporter genes, namely enhanced green fluorescent protein (EGFP), enhanced blue fluorescent protein (EBFP) and red fluorescent protein (DsRed2; Webster et al. 2005).
Several studies have proven the broad applicability of this technique to different animal species, from sea urchin to cattle (Lavitrano et al. 1989; Rottmann et al. 1991; Khoo et al. 1992; Schellander et al. 1995; Sperandio et al. 1996; Maione et al. 1998; Smith and Spadafora 2005). If we consider that SMGT can be applied to every animal species that undergoes sexual reproduction mediated by gametes, this strategy can be considered as potentially universal.
Molecular mechanism of sperm-mediated gene transfer
The beauty of SMGT is that the method uses the ‘natural’ vector of genetic material, namely the sperm cell, for transporting exogenous DNA.
The exogenous DNA molecules bind to the sperm cell’s head in the subacrosomal region and in the proximity of the equatorial area. Once bound to the cell membrane, DNA molecules are taken up (Francolini et al. 1993).
The process of DNA binding and internalisation is not a random event. The exogenous DNA interacts with DNA-binding proteins (DBPs) of 30–35 kDa, which are present on the sperm cell surface (Lavitrano et al. 1992; Zani et al. 1995) and we have demonstrated that major histocompatibility complex (MHC) class II and CD4 molecules also play a role in the process of sperm/DNA interaction (Lavitrano et al. 1997b). In fact, sperm cells from MHC class II-knockout mice have a reduced ability to bind DNA compared with sperm cells from wild-type animals (Mori et al. 1990; Wu et al. 1990). Conversely, spermatozoa from CD4-knockout mice are fully capable of binding exogenous DNA, yet lose the ability to internalise it. To further support the role of CD4 in DNA transfer, we have shown that nuclear internalisation of exogenous DNA is prevented in wild-type sperm cells pre-incubated with anti-CD4 monoclonal antibodies (mAbs). Thus, although not present in mature sperm cells, MHC class II expression appears to be required during spermatogenesis to produce sperm cells capable of taking up foreign DNA, whereas CD4 molecules present on sperm cells mediate the nuclear internalisation of sperm-bound DNA (Lavitrano et al. 1997b).
Nevertheless mature spermatozoa are naturally protected against the intrusion of foreign DNA molecules; in fact, we identified a factor (inhibitory factor 1, IF-1) present in the seminal fluid of mammals that blocks the binding of exogenous DNA to sperm cells. This factor also exerted a powerful inhibitory effect on DNA uptake in sperm cells of heterologous species. The DBPs appeared to be the specific target through which the inhibition is mediated because, in the presence of the inhibitory factor, the DBPs lose their ability to bind exogenous DNA (Zani et al. 1995). Thus, the interaction of exogenous DNA with sperm cells does not appear to be a casual event but, in contrast, relies on a molecular mechanism based on the cooperation of specific protein factors. In the absence of IF-1, DBPs are able to interact with DNA and the complex can translocate the DNA into the cell in a CD4-dependent manner (Lavitrano et al. 1997b). Thus, it is of utmost importance that seminal fluid is removed from sperm samples by extensive washing as soon as possible after ejaculation.
After DNA–sperm cell interaction and internalisation, the subsequent step involves integration of the exogenous DNA into the genome.
It has been suggested that various mechanisms may underlie DNA integration after the microinjection procedure (Brinster et al. 1985; Coffin 1990). DNA may be integrated when chromatin is freed from the nuclear context at fertilisation, at oocyte activation, at nucleus decondensation or even later at the formation of the pronuclei. Some groups have suggested that an integration event could result from the activation of an endogenous enzymatic machinery, such as during DNA repair (Spadafora 1998). Nonetheless, all data imply that this event arises after the penetration of the spermatozoon into the oocyte, but it is not known whether the DNA integration event happens before fertilisation. Interestingly, using the SMGT technique, it has been found that foreign DNA sequences are tightly bound to the sperm nuclear scaffold and that integration of the exogenous DNA occurs preferentially in the long interspersed nuclear element type-1 elements, repeated sequences interspersed into the genome. Moreover the presence of a topoisomerase II consensus sequence at one end of the integration site suggests a possible role for this enzyme in the integration process (Spadafora 1998).
Sequence analysis of randomly selected clones from a library of sperm genomic DNA incubated with pSV2CAT plasmid showed that foreign sequences were integrated in a unique site of the sperm genome (Magnano et al. 1998).
Technical aspects of the sperm-mediated gene transfer method
The initial experiments for the production of transgenic animals with SMGT were performed in the mouse using epididymal sperm cells. These experiments revealed two different problems: (1) a lack of reproducibility in some laboratories; and (2) rearranged banding patterns of the transgene (Wall 1999). Since then, our research group shifted its attention to the study of SMGT in the pig, which provides the advantage of using ejaculated instead of epididymal sperm. In the case of epididymal sperm, the donor animal can be used only once, whereas with ejaculated sperm the same animal can be used for its entire reproductive life. This is particularly important because we found that there are considerable differences in the capability of sperm from different animals to take up exogenous DNA (Lavitrano et al. 1996).
We have demonstrated that there are two important parameters that must be optimal for the SMGT technique to be effective: (1) quality of semen sample; and (2) DNA uptake, which is dependent primarily on viability and motility (Lavitrano et al. 2003). We found that the standard parameters that boar semen should possess in order to serve as a good vector for exogenous DNA are quality semen, based on standard parameters used in conventional animal breeding programmes (volume, concentration, presence of abnormal sperm, motility etc.), and the ability of the sperm cells to take up and internalise exogenous DNA.
Sperm quality is influenced by many factors, such as the season of the year (semen quality declines significantly during the hot season), collection frequency and age of the donor. Fertility results obtained at breeding should also be evaluated. In addition to the criteria used routinely by pig farmers to choose good boars for breeding, we paid particular attention to the acquisition of sperm with high progressive motility, which correlates with their fertilisation capacity. Motility should be at least 80% initially and not less that 65% after washing procedures. A critical parameter in sperm selection is also the ability of sperm cells to bind exogenous DNA and to internalise it into their nuclei. Interestingly, we found that DNA uptake correlates with semen quality, particularly in terms of high progressive motility.
We spent considerable effort determining the parameters that are important for the optimisation of DNA uptake by sperm. To optimise the protocol for generating transgenic animals, we established when, for how long and in what quantity DNA must be added to the sperm so that the exogenous DNA is being taken up by the majority of sperm cells and nuclei. In optimised experiments, exogenous DNA binds to approximately 90% of sperm cells and binding is followed by nuclear internalisation in 70% of these cells; DNA–sperm interaction is completed within 2–4 h (Lavitrano et al. 2003).
Deoxyribonucleic acid should be added within 30 min after washing and not later than 60 min; in fact, there is a window of opportunity that coincides with the early stage of capacitation (Lavitrano et al. 2003). It is important to optimise the amount of exogenous DNA per sperm cells in order to obtain the highest number of sperm cells containing DNA without overloading. Deoxyribonucleic acid-overloaded sperm could be damaged or disadvantaged in terms of fertilisation compared with normal spermatozoa and artificial insemination could amplify this disadvantage.
Interestingly, the uptake of the DNA, at least in mammals, does not generally interfere with physiological semen parameters, such as motility at time of collection and progressive motility. In some cases, treated sperm cells perform better than untreated cells (Chan 2000).
Production of transgenic pigs
Transgenic animals have, for the most part, been produced using the microinjection of exogenous DNA into the male pronuclei of zygotes (Wall 2001). This technique, although highly successful in mice, is not as efficient in farm animals, a fact that limits its general usefulness.
Given our interest in xenotransplantation and the possibility that a donor animal will need to express several transgenes, we used SMGT to produce pigs transgenic for hDAF, which has been shown to help overcome the first rejection barrier in pig-to-primate transplantation models. We have generated over 100 hDAF transgenic pigs to be used for xenotransplantation research and the efficiency of transgenesis obtained with SMGT was much greater than with any other method. We have reported that up to 80% of pigs had the transgene integrated into the genome. In eight experiments, 53 of 93 pigs generated were transgenic (57%; Lavitrano et al. 2002, 2003). This result contrasts with the reported efficiencies of 0.5–4% in pigs using microinjection (Niemann and Kues 2003); thus, the SGMT technique offers a 25-fold improvement. Most of the pigs carrying the hDAF gene transcribed it in a stable manner (64%) and the majority of pigs that transcribed the gene also expressed the protein (83%). The hDAF gene was transmitted to the progeny. Expression was stable and hDAF protein was found in caveolae, as described for normally expressed DAF proteins in humans. Moreover, the level of expression of hDAF protein in transgenic pigs was comparable with the level in human tissues. The expressed gene was functional, as assessed by in vitro experiments performed on peripheral blood mononuclear cells, because monocytes prepared from hDAF transgenic pigs were able to survive when challenged with fresh human serum (Lavitrano et al. 2002).
Multitransgenic pigs
A simple and efficient method for producing multitransgenic animals is required for medical and veterinary applications. Recently, nuclear transfer has been used to clone large animals and this could allow multiple genetic manipulations to be undertaken in vitro, before a single nuclear transfer, rather than having to go through complex and time-consuming breeding programmes (Niemann and Kues 2003). However, at present, the frequency of success in cloning large animals is very low and this technique is very expensive. We have used the SMGT method to produce, with high efficiency, multigene transgenic pigs using three genes coding for fluorescent proteins: EBFP, EGFP and DsRed2 (Webster et al. 2005). All three fluorescent proteins were expressed in 171 of 195 normally developed morula/blastocysts examined at Day 6 after insemination (88%). The genomic DNA of 18 piglets born from two litters was screened by polymerase chain reaction, showing that all piglets were transgenic with at least one gene, seven of 18 piglets were triple transgenic, seven of 18 were double transgenic and four of 18 were single transgenic. The RNA and protein expression was found in muscle, heart, liver, hair and peripheral blood mononuclear cells. These results indicate that SMGT is an effective method for introducing multiple genes into pigs, as shown by the simultaneous expression of the three fluorescent proteins.
Modifications of sperm-mediated gene transfer
Sperm-mediated gene transfer in vertebrates has undergone various developments over the past few years in different laboratories.
Numerous attempts have been made to improve the SMGT method, which have not always been worth the effort because of an increase in the procedural steps, equipment and/or personal skills required, as well as an increase in costs. One method that proved to be of interest in species for which there is a need for a more powerful technique is restriction enzyme-mediated integration (REMI). Sperm transfection is achieved with liposomes containing linearised plasmid molecules having cohesive ends and the restriction enzyme used for the linearisation. What drives the integration in the SMGT procedure is the ‘natural’ machinery already present in the cells’ nuclei, whereas in the REMI experiments it is an exogenous enzyme that mediates significant, but yet uninvestigated, genomic rearrangements (Shemesh et al. 2000).
Electroporation is a procedure that has been introduced into SMGT to ameliorate the number of DNA molecules taken up by the sperm cells (Rieth et al. 2000). Even though it seems not to affect the fertilisation capability of spermatozoa (Tsai et al. 1997), electroporation allows the sperm cells to take up more DNA molecules than unelectroporated spermatozoa, but does not yield a higher percentage of ‘transfected’ cells (Muller et al. 1992). Therefore, electroporation should be performed only in those species in which the setting up of an efficient protocol for DNA uptake is difficult and should be avoided when possible in order to diminish the handling stress to sperm cells. This could also be a drawback because DNA-overloaded sperm perform less efficiently compared with spermatozoa carrying an ‘optimum’ quantity of DNA (approximately 500 ng per 106 spermatozoa; Sciamanna et al. 2000).
The idea that forms the basis of the SMGT technique has been taken to extremes by producing transgenic sperm cells directly in seminiferous tubules using liposomes or electroporation. In this way, transgenic spermatozoa-producing animals have been generated, but the yield (7–13%) was not comparable with the classical ex vivo SMGT technique (5–60%; Kim et al. 1997). Very recently, the impact of electroporation treatment on the organism has also been assessed, showing that damage inflicted to spermatogenesis by electroporation can severely impair the production of sperm cells (Umemoto et al. 2005). Celebi et al. (2002) microinjected a circular plasmid, carrying the lacZ reporter gene mixed with non-commercial cationic lipids, into the seminiferous tubules of anaesthetised adult mice and showed that the transgene was transmitted to the offspring but remained episomal, because it was found in the tail of the young animals but not in that of adult animals. Therefore, the plasmid seemed to be lost during germ cell division.
Conclusions
Over the past decades, SMGT has drawn the attention of many researchers in the ongoing debate on animal transgenesis. In spite of its having become a well-accepted and established method, SMGT is still subject to development and new approaches are being developed to make the technique even more widespread and reliable. Given the wide availability of livestock semen, SMGT can be considered the method of choice for the production of genetically modified farm animals and represents the most powerful tool available today for medical research purposes. First, the overall frequency of transgenic offspring among species using SMGT is in the range of 5–60%, contrasting with efficiencies of 0.5–4% using microinjection (Niemann and Kues 2003). Second, with SMGT, DNA-treated sperm cells are used in artificial insemination, as normally practiced on the farm, whereas microinjection requires expensive equipment and has many steps (such as superovulation, collection and injection of zygotes), with the ensuing waste of egg donors and animal embryos. Sperm-mediated gene transfer overcomes several disadvantages of microinjection in the production of transgenic pigs. Furthermore, single transgenic expressor pig offspring obtained by microinjection cost approximately US$25 000 (Niemann and Kues 2003), whereas the cost using SMGT is less than US$1000 (Lavitrano et al. 2003). In conclusion, SMGT is an efficient and inexpensive method that will hopefully facilitate the implementation of strategies for securing the benefits that can be expected to arise from the introduction of transgenic livestock.
Acknowledgments
The authors’ work reported herein was funded by the Italian Ministry of Agriculture (DM 581/7240/96, DM 564/7240/97, DM 404/7240/99, DM 36/7303/02) and by the Italian Minister of Research and University (DD 21.09.99, n462 ric). The authors thank Elena Giovannoni for assistance and the care of animals.
Brackett, B. G. , Baranska, W. , Sawichi, W. , and Koprowski, H. (1971). Uptake of heterologous genome by mammalian spermatozoa and its transfer to ova through fertilization. Proc. Natl Acad. Sci. USA 68, 353–357.
Lavitrano, M. , Forni, M. , Varzi, V. , Pucci, L. , and Bacci, M. L. , et al. (1997a). Sperm-mediated gene transfer: production of pigs transgenic for a human regulator of complement activation. Transplant. Proc. 29, 3508–3509.
| Crossref | GoogleScholarGoogle Scholar | PubMed |

Lavitrano, M. , Maione, B. , Forte, E. , Francolini, M. , Sperandio, S. , Testi, R. , and Spadafora, C. (1997b). The interaction of sperm cells with exogenous DNA: a role of CD4 and major histocompatibility complex class II molecules. Exp. Cell Res. 233, 56–62.
| Crossref | GoogleScholarGoogle Scholar | PubMed |
Lavitrano, M. , Bacci, M. L. , Forni, M. , Lazzereschi, D. , and Di Stefano, C. , et al. (2002). Production by sperm-mediated gene transfer of human decay accelerating factor (hDAF) transgenic pigs for xenotransplantation. Proc. Natl Acad. Sci. USA 99, 14 230–14 235.
| Crossref | GoogleScholarGoogle Scholar |
Lavitrano, M. , Forni, M. , Bacci, M. L. , Di Stefano, C. , Varzi, V. , Wang, H. , and Seren, E. (2003). Sperm mediated gene transfer in pig: selection of donor boars and optimization of DNA uptake. Mol. Reprod. Dev. 64, 284–291.
| Crossref | GoogleScholarGoogle Scholar | PubMed |
Magnano, A. R. , Giordano, R. , Moscufo, N. , Baccetti, B. , and Spadafora, C. (1998). Sperm/DNA interaction: integration of foreign DNA sequences in the mouse sperm genome. J. Reprod. Immunol. 41, 187–196.
| Crossref | GoogleScholarGoogle Scholar | PubMed |
Maione, B. , Lavitrano, M. , Spadafora, C. , and Kiessling, A. A. (1998). Sperm-mediated gene transfer in mice. Mol. Reprod. Dev. 50, 406–409.
| Crossref | GoogleScholarGoogle Scholar | PubMed |
Mori, T. , Guo, M. W. , Mori, E. , Shindo, Y. , Mori, N. , Fukuda, I. , and Mori, T. (1990). Expression of class II major histocompatibility complex antigen on sperm and its role in fertilization. Am. J. Reprod. Immunol. 24, 9–10.
| PubMed |
Muller, F. , Ivics, Z. , Erdelyi, F. , Papp, T. , Varadi, L. , Horvath, L. , and Maclean, N. (1992). Introducing foreign genes into fish eggs with electroporated sperm as a carrier. Mol. Mar. Biol. Biotechnol. 1, 276–281.
| PubMed |
Niemann, H. , and Kues, W. A. (2003). Application of transgenesis in livestock for agriculture and biomedicine. Anim. Reprod. Sci. 79, 291–317.
| Crossref | GoogleScholarGoogle Scholar | PubMed |
Rieth, A. , Pothier, F. , and Sirard, M. A. (2000). Electroporation of bovine spermatozoa to carry DNA containing highly repetitive sequences into oocytes and detection of homologous recombination events. Mol. Reprod. Dev. 57, 338–345.
| Crossref | GoogleScholarGoogle Scholar | PubMed |
Rottmann, O. J. , Antes, R. , Hoefer, P. , and Maierhofer, G. (1991). Liposomes mediate gene transfer via spermatozoa into avian egg cells. J. Anim. Breed. Genet. 109, 64–70.
Schellander, K. , Peli, J. , Schmall, F. , and Brem, G. (1995). Artificial insemination in cattle with DNA-treated sperm. Anim. Biotech. 6, 41–50.
Sciamanna, I. , Piccoli, S. , Barberi, L. , Zaccagnini, G. , Magnano, A. R. , Giordano, R. , Campedelli, P. , Hodgson, C. , Lorenzini, R. , and Spadafora, C. (2000). DNA dose and sequence dependence in sperm-mediated gene transfer. Mol. Reprod. Dev. 56, 301–305.
| Crossref | GoogleScholarGoogle Scholar |
Shemesh, M. , Gurevich, M. , Harel-Markowitz, E. , Benvenisti, L. , Shore, L. S. , and Stram, Y. (2000). Gene integration into bovine sperm genome and its expression in transgenic offspring. Mol. Reprod. Dev. 56(Suppl. 2), 306–308.
| Crossref | GoogleScholarGoogle Scholar |
Smith, K. , and Spadafora, C. (2005). Sperm-mediated gene transfer: applications and implications. Bioessays 27, 551–562.
| Crossref | GoogleScholarGoogle Scholar | PubMed |
Spadafora, C. (1998). Sperm cells and foreign DNA: a controversial relation. BioEssays 20, 955–964.
| Crossref | GoogleScholarGoogle Scholar | PubMed |
Sperandio, S. , Lulli, V. , Bacci, M. L. , Forni, M. , Maione, B. , Spadafora, C. , and Lavitrano, M. (1996). Sperm mediated DNA transfer in bovine and swine species. Anim. Biotechnol. 7, 59–77.
Tsai, H. J. , Lai, C. H. , and Yang, H. S. (1997). Sperm as a carrier to introduce an exogenous DNA fragment into the oocyte of Japanese abalone (Haliotis divorsicolor suportexta). Transgenic Res. 6, 85–95.
| Crossref | GoogleScholarGoogle Scholar | PubMed |
Umemoto, Y. , Sasaki, S. , Kojima, Y. , Kubota, H. , Kaneko, T. , Hayashi, Y. , and Kohri, K. (2005). Gene transfer to mouse testes by electroporation and its influence on spermatogenesis. J. Androl. 26, 264–271.
| PubMed |
Wall, R. J. (1999). Sperm-mediated gene transfer: advances in sperm cell research and applications. Transgenic Res. 8, 313–315.
| Crossref | GoogleScholarGoogle Scholar | PubMed |
Wall, R. J. (2001). Pronuclear microinjection. Cloning Stem Cells 3, 209–220.
| Crossref | GoogleScholarGoogle Scholar | PubMed |
Wall, R. J. (2002). New gene transfer methods. Theriogenology 57, 189–201.
| Crossref | GoogleScholarGoogle Scholar | PubMed |
Webster, N. L. , Forni, M. , Bacci, M. L. , Giovannoni, R. , and Razzini, R. , et al. (2005). Multi-transgenic pigs expressing three fluorescent proteins produced with high efficiency by sperm mediated gene transfer. Mol. Reprod. Dev. 72, 68–76.
| Crossref | GoogleScholarGoogle Scholar | PubMed |
Wu, G. M. , Nose, K. , Mori, E. , and Mori, T. (1990). Binding of foreign DNA to mouse sperm mediated by its MHC class II structure. Am. J. Reprod. Immunol. 24, 120–126.
| PubMed |
Zani, M. , Lavitrano, M. , French, D. , Lulli, V. , Maione, B. , Sperandio, S. , and Spadafora, C. (1995). The mechanism of binding of exogenous DNA to sperm cells: factors controlling the DNA uptake. Exp. Cell Res. 217, 57–64.
| Crossref | GoogleScholarGoogle Scholar | PubMed |