

Analysis of DAZ gene expression in a partial AZFc deletion of the human Y chromosome
Byunghyuk Kim A , Wonkyung Lee A , Kunsoo Rhee A C , Soo Woong Kim B and Jae-Seung Paick B CA Department of Biological Sciences, Seoul National University, 599 Gwanak-ro, Gwanak-gu, Seoul 151-747, Korea.
B Department of Urology, College of Medicine, Seoul National University, 101 Daehak-ro, Jongno-gu, Seoul 110-799, Korea.
C Corresponding authors. Emails: rheek@snu.ac.kr; jspaick@snu.ac.kr
Reproduction, Fertility and Development 26(2) 307-315 https://doi.org/10.1071/RD12290
Submitted: 6 September 2012 Accepted: 10 January 2013 Published: 20 February 2013
Abstract
The azoospermia factor c (AZFc) region of the Y chromosome consists of repetitive amplicons and is therefore highly susceptible to structural rearrangements, such as deletions and duplications. The b2/b3 deletion is a partial AZFc deletion that is conventionally determined by the selective absence of sY1191 in sequence-tagged site polymerase chain reaction (PCR) and is generally believed to retain two of the four deleted in azoospermia (DAZ) genes on the Y chromosome. In the present study we determined the copy number and expression of DAZ genes in sY1191-negative individuals. Using a DAZ dosage PCR assay and Southern blot analysis we evaluated the expression of four DAZ genes in five of six sY1191-negative individuals. Furthermore, cloning and immunoblot analyses revealed that three or more DAZ genes are expressed in sY1191-negative testes with germ cells. The results indicate that the selective absence of sY1191 not only means b2/b3 deletion with two DAZ genes, but also includes another AZFc configuration with four DAZ genes. These results exemplify the prevalence of variations in the AZFc region of the human Y chromosome.
Additional keywords: male infertility, microdeletion, testis.
Introduction
The azoospermia factor c (AZFc) locus of the human Y chromosome contains massive sequence repeats called amplicons (Kuroda-Kawaguchi et al. 2001). The amplicons are organised in palindromic domains that harbour a cohort of genes for spermatogenesis (Kuroda-Kawaguchi et al. 2001). Because the ampliconic domains are highly susceptible to intra- and inter-chromosomal recombinations, microdeletions frequently occur in the AZFc region, often leading to spermatogenic failure in men (Kuroda-Kawaguchi et al. 2001; Skaletsky et al. 2003). For example, total deletion of AZFc results from recombination of the b2 and b4 domains, and spermatogenic failure is frequently observed in individuals with b2/b4 deletions (Reijo et al. 1995; Kuroda-Kawaguchi et al. 2001). The gr/gr deletion eliminates 1.6 Mb of sequence in the AZFc region and is considered to be a significant risk factor for impaired spermatogenesis (Repping et al. 2003). The b2/b3 deletion is another frequent deletion pattern in which an inversion and homologous recombination may occur sequentially within the AZFc locus (Fernandes et al. 2004; Repping et al. 2004). However, the effects of partial deletions on male fertility remain contentious. For example, some researchers have found an association between the b2/b3 deletion and infertility (Wu et al. 2007; Lu et al. 2009; Eloualid et al. 2012), whereas others have not (Repping et al. 2004; Hucklenbroich et al. 2005).
Sequence redundancy in AZFc makes it difficult to analyse the deletions with direct sequencing. Instead, unique sequence-tagged sites (STSs) at the AZFc region have been widely used to detect partial deletions within the region. For example, the gr/gr and b2/b3 deletions are determined in individuals with a selective absence of STS markers sY1291 and sY1191, respectively (Repping et al. 2003, 2004). The AZFc configuration resulting from these deletions has been further supported by the results of interphase fluorescent in situ hybridisation (FISH), Southern blot analyses and sequence family variant (SFV)-based assays (Fernandes et al. 2002, 2004; Repping et al. 2003, 2004; Lin et al. 2005, 2006). In this context, several association studies for the phenotypic consequences of partial AZFc deletions have relied largely on STS-PCR assays to detect such deletions (Hucklenbroich et al. 2005; Lynch et al. 2005; Ravel et al. 2006; Eloualid et al. 2012). However, this simple method may not provide information as to exactly which genes or gene copies are deleted in the chromosomes.
Deleted in azoospermia (DAZ) is a well known male infertility gene in the AZFc region of the Y chromosome (Saxena et al. 1996). DAZ and two autosomal homologues, namely deleted in azoospermia-like (DAZL) and bol, boule-like (Drosophila) (BOULE), are critical for germ cell development in humans and other animals, such as mice, Drosophila melanogaster and Caenorhabditis elegans (Reijo et al. 1995; Eberhart et al. 1996; Ruggiu et al. 1997; Karashima et al. 2000; VanGompel and Xu 2010). The human AZFc region includes four DAZ genes with over 99% sequence identity (Saxena et al. 1996, 2000) and it has been shown that all the DAZ genes are actively transcribed and translated in the human testis (Kim et al. 2009). The DAZ proteins contain one or more RNA recognition motifs (RRMs) and multiple DAZ repeats (Yen et al. 1997; Saxena et al. 2000). It is of note that the DAZ proteins are polymorphic, containing different numbers of the RRMs and DAZ repeats (Kim et al. 2009).
By screening AZFc deletions in a previous study (Kim et al. 2009), we found a sample with a selective absence of sY1191 on STS-PCR that was indicative of the b2/b3 deletion. Interestingly, immunoblot analysis with a testicular sample revealed three discrete DAZ bands of different sizes (Kim et al. 2009). This result was unexpected because the model of the b2/b3 deletion predicts that two copies of DAZ are eliminated, leaving only two DAZ genes on the Y chromosome (Fernandes et al. 2004; Repping et al. 2004). In the present study we analysed the copy number of DAZ genes in sY1191-negative individuals and examined DAZ expression in their testes at the transcriptional and translational levels. The results unexpectedly revealed that most of the sY1191-negative individuals examined have more than two DAZ genes.
Materials and methods
Human samples and deletion screening of the AZFc region
All the subjects in the present study were ethnic Korean men who were recruited from an infertility clinic. Semen analyses and physical and clinical examination were performed according to conventional procedures performed in the clinic (Kim et al. 1999). In all, there were 150 subjects in the present study, comprising 148 infertile men who were diagnosed with either azoospermia or obstructive azoospermia and two fertile men (#001 and #155). Human samples for bloods and testes were obtained from all 150 subjects after they had provided informed consent following study approval by the Institutional Review Board of Seoul National University Hospital. All samples were stored at –80°C until analysis.
Genomic DNA was isolated from the blood according to the protocol described by Sambrook et al. (1989). Briefly, 10 mL fresh blood was centrifuged at 1300g for 15 min at 4°C and the buffy coat was obtained. The buffy coat was treated with 100 μg mL−1 proteinase K for 3 h. An equal volume of phenol equilibrated with Tris-HCl (pH 8) was added to the samples and gently mixed. The aqueous phase was collected after centrifugation at 5000g for 15 min at room temperature and mixed with an equal volume of chloroform. The aqueous phase was ethanol precipitated after mixing with 0.4 volumes of 5 M ammonium acetate. The precipitates were suspended with 10 mM TE8 and the DNA concentration was determined using a spectrophotometer.
The screening of AZFc deletions was conducted by STS-PCR with PCR conditions available at GenBank (http://www.ncbi.nlm.nih.gov/genbank/; accessed 9 December 2010) under the following accession numbers: sY142 (G38345), sY254 (G38349), sY255 (G65827), sY1191 (G73809), sY1197 (G67168), sY1201 (G67170), sY1206 (G67171), sY1291 (G72340). All STS-PCR were performed separately for each STS.
For the validation of the absence of a PCR product for sY1191, three different annealing temperatures (50, 56 and 61°C) were tested in the PCR for sY1191. In addition, two different pairs of primers were used to confirm the sY1191 deletion: (1) sY1191–2 (490 bp; tcagacactattttggcaatttt (forward) and ggcagcagaatcgcttgtat (reverse)), which targets the sY1191 locus and includes the original sY1191 targeting sequence of 385 bp in size; and (2) u3 (193 bp; o1276: agtctgagtggctagtgcac; o1277: gaagcaaagttcagctgtgc), which is part of u3 and close to sY1191 (Machev et al. 2004). A pair of PCR primers for SRY was also used as a control (Simoni et al. 2004). The PCR were performed using the following conditions: initial heating at 95°C for 15 min followed by 30 cycles of 94°C for 30 s, 50–61°C for 30 s and 72°C for 30 s, with a final extension at 72°C for 4 min. The annealing temperatures were 50, 56 and 61°C for sY1191, 60°C for sY1191–2, 60°C for u3 and 55°C for SRY.
Reverse transcription–polymerase chain reaction
Testicular RNAs were extracted from human testicular samples using TRIzol (Invitrogen, Carlsbad, CA, USA) according to the manufacturer’s instructions. Total RNAs were reverse transcribed using random hexamers. The resultant cDNAs were amplified by PCR using the pairs of primers for the DAZ 3′-untranslated region (UTR), DAZL, GAPDH and the DAZ coding region. The pair of primers for the DAZ coding region was designed to amplify all four DAZ transcripts with one PCR reaction. The sequence information for the primers and the PCR conditions have been described in detail elsewhere (Kim et al. 2009).
Immunoblot analysis
Human testicular biopsy samples were homogenised in 1× sodium dodecyl sulfate (SDS) sample buffer (50 mM Tris-HCl, pH 6.8, 100 mM dithiothreitol, 2% SDS, 0.1% bromophenol blue, 10% glycerol). The SDS–polyacrylamide gel electrophoresis (PAGE) and immunoblotting assays were conducted as previously described (Kim et al. 2009) with some modifications. Briefly, protein extracts were resolved on 10% polyacrylamide gels and then transferred to nitrocellulose membranes. Membranes were blocked in 5% skim milk in TBST (20 mM Tris, 150 mM NaCl, 0.1% Tween 20) for 30 min, and then incubated overnight with anti-DAZ antibody (1 : 100 diluted in blocking solution; Kim et al. 2009) at 4°C. After three washes with TBST for 10 min each time, membranes were incubated with a peroxidase-conjugated anti-rabbit IgG (1 : 10 000; Calbiochem, La Jolla, CA, USA) for 1 h at room temperature. The membranes were then washed another three times with TBST for 10 min each time, and peroxidase activity was detected using enhanced chemiluminescence (ECL) reagent. As a loading control, membranes were reprobed with anti-β-tubulin antibody (1 : 1000; Sigma, St Louis, MO, USA).
Assays to determine DAZ dosage
In the present study, we used the DAZ-dosage PCR assay developed and described by Lin et al. (2006) using primer pairs PrDAZ109 and PrDAZ110 and some modifications in the composition of PCR reaction mix. Briefly, the 10 μL PCR reaction volume contained 100 ng genomic DNA, 4 pmol each primers, 0.25 mM each dNTPs, 1 μL of 10× PCR buffer and 0.5 units NovacleanTaq polymerase (Genenmed, Seoul, Korea). The PCR was performed using the following conditions: initial heating at 95°C for 5 min followed by 25 cycles of 95°C for 20 s, 49°C for 20 s and 72°C for 30 s, with a final extension at 72°C for 5 min. The PCR products were analysed on 2% agarose gels. Signal intensities were measured using ImageJ (National Institutes of Health (NIH), Besthesda, MA, USA).
Southern blot analysis
The probe for a dosage Southern blot was a mixture of two 1.4-kb PCR fragments containing the 3′ UTRs of DAZ and DAZL, and was prepared as described previously (Lin et al. 2006). The probe for Southern blot analysis of the RRM region of the DAZ genes was a 1.0-kb PstI fragment containing the 3′ end of intron 1 of the DAZ genomic clone 7A69G (Lin et al. 2005).
Briefly, 3 μg genomic DNA was digested with 50 units NsiI (restriction enzyme) for 6 h. The DNA fragments were separated on 0.5% agarose gels in 1× TAE buffer (40 mM Tris-acetate, 1 mM EDTA, pH 8.0). Using denaturation/transfer solution (0.4 M NaOH, 0.8 M NaCl), the DNA fragments in the gels were denatured for 30 min and the transferred to nylon membranes for 7 h by capillary action with frequent towel changes. Membranes were soaked in neutralisation solution (0.5 M Tris-HCl, pH 7.2, 1 M NaCl) for 15 min and then baked at 80°C for 2 h. The membranes were then incubated for 1 h at 65°C in prehybridisation buffer (5× saline-sodium citrate (SSC), 5× Denhardt’s reagent, 250 μg mL−1 salmon sperm DNA) and then hybridised with a radiolabelled probe overnight at 65°C in hybridisation buffer (5× SSC, 1× Denhardt’s reagent, 250 μg mL−1 salmon sperm DNA, 10% dextran sulfate, 0.1% SDS). The DNA probes were 32P labelled by random priming using the Prime-a-Gene labelling system (Promega, Madison, WI, USA) according to the manufacturer’s instructions. At the end of hybridisation, the membranes were washed four times at 65°C with washing buffer (50 mM Tris, 1 mM EDTA, 1× Denhardt’s reagent, 0.1% SDS, pH 8.0) over a 1 h period. After the final wash, the membranes were dried and exposed to a film at –80°C for 2 days. Signal intensities for a dosage Southern blot were measured using ImageJ (NIH).
Cloning and expression of DAZ transcripts
The PCR products amplified using primers for the DAZ coding region were used to clone DAZ transcripts from three sY1191-negative testicular cDNAs. After extraction with phenol–chloroform, the DAZ cDNAs were ligated into pGEM-T Easy Vector (Promega). The cloned DAZ transcripts were verified by sequence analyses. For mammalian expression of the DAZ cDNAs, the pGEM-T Easy Vector containing an entire protein coding region of DAZ was digested by EcoRI, and the resulting fragment was subcloned into pcDNA3.1(+) vector (Invitrogen). These constructs were subsequently transfected into 293T cells.
Cell culture and transfection
The culture of 293T cells and transient transfection using polyethylenimine have been described in detail elsewhere (Kim et al. 2009). Transfected cells were lysed and subjected to immunoblot analysis as described above.
Results
Screening for sY1191 deletion
To detect partial deletions in the AZFc locus, we performed genomic PCR analyses using specific STSs. We initially screened 150 subjects using three STSs (sY255, sY1191 and sY1291) and identified six individuals who lacked sY1191 only. We confirmed that the six sY1191-negative individuals had five additional STSs (sY142, sY254, sY1197, sY1201 and sY1206; Table 1). As controls, we included two individuals with an intact AZFc, two individuals with the sY1291-negative gr/gr deletions and two individuals with total AZFc deletions (Table 1; Kim et al. 2009).

|
The absence of a PCR product for sY1191 was further tested in order to validate that the result was due to genuine deletion rather than PCR failure. First, we examined the effect of lowered annealing temperatures in the PCR for sY1191, because non-optimal PCR conditions could result in false identification of deletion. Second, we used two different pairs of primers targeting the sY1191 locus (sY1191–2) and a sequence that is close to the sY1191 in the u3 region of AZFc (u3), because it is possible that STS-PCR failure may occur due to single nucleotide substitutions in primer sequences for STSs (Thornhill et al. 2002; Wu et al. 2011). In both tests, PCR fragments were amplified in the intact AZFc and gr/gr deletion samples, but not in the total AZFc deletion and six sY1191-negative samples (Fig. 1). These results strongly suggest that the sY1191-negative individuals have genuine deletions in the sY1191 region.
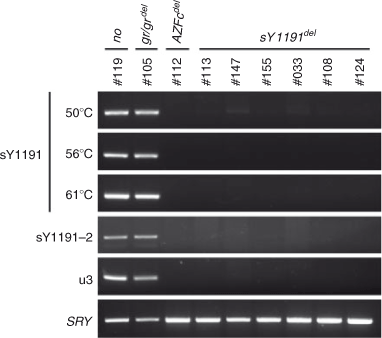
|
Expression of DAZ in sY1191-negative testes
To detect DAZ expression in AZFc-deleted testes, we performed reverse transcription–polymerase chain reaction (RT-PCR) analysis using a primer set targeting the 3′ UTR of all the DAZ cDNAs. A DAZ-specific band was detected in three of six sY1191-negative samples (Fig. 2a). In addition, the DAZ-specific band was detected in the intact AZFc and gr/gr deletion samples, but not in the total AZFc deletion sample (Fig. 2a). As a control, we determined the expression of DAZL, an autosomal homologue of DAZ with germ cell-specific expression (Yen et al. 1996). A DAZL-specific band was detected in all testicular samples except for the sY1191-negative samples that lacked the DAZ-specific band (Fig. 2a). The PCR fragment of GAPDH was detected in all samples examined (Fig. 2a). The RT-PCR results indicate that the sY1191-negative individuals without the DAZ- and DAZL-specific bands lack male germ cells in their testes (Fig. 2a). In support of this interpretation, the sY1191-negative individuals without the DAZ-specific bands were diagnosed with azoospermia (#033, #108 and #124), whereas those with the DAZ-specific bands were diagnosed with obstructive azoospermia (#113 and #147) or normal (#155).
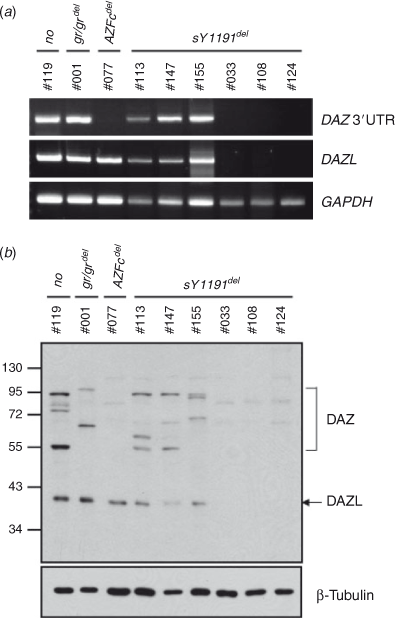
|
We then analysed DAZ protein expression in the sY1191-negative testes using a well-characterised DAZ antibody that detects both the DAZ and DAZL proteins in human testicular samples (Kim et al. 2009). As reported previously (Kim et al. 2009), four and two DAZ-specific bands were detected in the testicular samples with no AZFc deletion and gr/gr deletion, respectively (Fig. 2b). No DAZ-specific band was detected in the sample with a total AZFc deletion (Fig. 2b; Kim et al. 2009). Under our study conditions, three discrete DAZ bands were detected in the sY1191-negative samples that had germ cells (Fig. 2b). Consistent with the RT-PCR results, the DAZL-specific band was not detected in sY1191-negative samples without germ cells (Fig. 2b). Furthermore, the diverse DAZ-specific bands in the sY1191-negative samples confirmed the polymorphic nature of DAZ proteins (Fig. 2b; Kim et al. 2009). These results suggest that more than two DAZ genes are expressed in the sY1191-negative testes.
Determination of DAZ copy numbers in sY1191-negative individuals
We performed gene dosage assays to estimate the copy numbers of DAZ genes in individuals with a selective absence of sY1191. The autosomal DAZL gene was used as an internal dosage control for the determination of DAZ copy number. As comparative dosage controls, genomic samples with intact AZFc (#119; four DAZ genes), the gr/gr deletion (#105; two DAZ genes) and the AZFc deletion (#112; no DAZ gene) were used (Fig. 3). First, we determined the PCR band intensities of DAZ and DAZL using the six sY1191-negative subjects as well as the control samples, and compared their DAZ : DAZL signal ratios (Fig. 3a; Lin et al. 2006). Similarly, using dosage Southern blot, we determined DAZ : DAZL signal ratios in the same samples (Fig. 3b; Lin et al. 2006). Using both measures, the signal ratios of five sY1191-negative subjects (#113, #147, #155, #033 and #124) were similar to those of #119 with four DAZ genes; the exception was #108, which showed similar signal ratios as seen in #105 with two DAZ genes (Fig. 3). These results suggest that most of the sY1191-negative individuals examined have four DAZ genes.
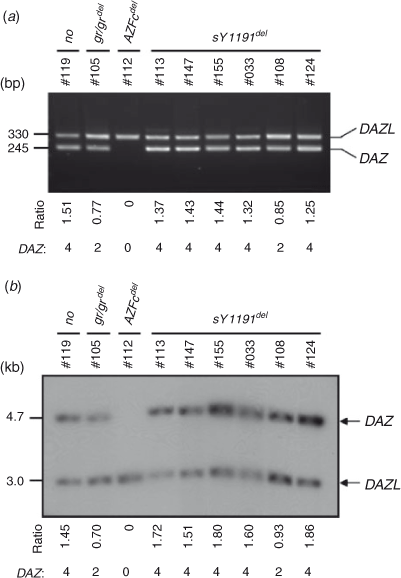
|
Identification of DAZ genes expressed in sY1191-negative testes
To gain an insight into the structure of DAZ genes in the sY1191-negative individuals, we undertook Southern blot analysis of the RRM region of the DAZ genes. The probe used was designed to detect NsiI fragments that are 31, 20 and 9 kb in size when the DAZ genes contain three, two and one RRM, respectively (Lin et al. 2005). Consistent with our previous report (Kim et al. 2009), the #119 sample had a DAZ gene with three RRMs (DAZ1), a DAZ gene with two RRMs (DAZ4) and two DAZ genes with a single RRM (DAZ2 and DAZ3), whereas the #105 sample had a DAZ gene with three RRMs (DAZ4) and a DAZ gene with a single RRM (DAZ3; Fig. 4). Hybridisation patterns of the six sY1191-negative subjects showed that three subjects (#113, #147 and #033) have at least three DAZ genes with three, two and one RRM each, whereas the others (#155, #108 and #124) have at least two DAZ genes with three and one RRM (Fig. 4). This suggests that the numbers and kinds of DAZ genes in sY1191-negative subjects are heterogeneous.
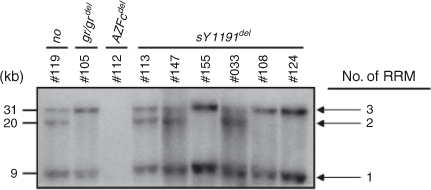
|
To identify which DAZ genes are expressed in the sY1191-negative individuals, we cloned the DAZ transcripts expressed in the three sY1191-negative testes with germ cells. We performed RT-PCR with primers targeting the full-length coding region of DAZ and isolated the multiple PCR fragments that ranged from 2 to 4 kb (data not shown). These PCR fragments were then cloned and sequenced. As reported previously (Kim et al. 2009), this method enables us to identify the original DAZ transcripts in subjects, but cannot avoid derivatives resulting from PCR artefacts due to the repetitive nature of DAZ genes. Therefore, we determined a cDNA clone as the real transcript from a specific DAZ gene rather than a PCR artefact as follows: (1) the origins of the DAZ cDNAs were sorted out, based on the sequence characteristics of the DAZ copies (Fig. 5; Saxena et al. 2000; Fernandes et al. 2002); and (2) the largest clone of a specific DAZ gene was counted as the original transcript and the smaller clones were considered artefacts with deletions during PCR amplification (Yen et al. 1997; Kim et al. 2009). We observed that all the DAZ cDNA clones originated from three or four different DAZ genes in each sY1191-negative testicular sample (Fig. 5). The DAZ cDNAs in the #113 and #147 samples originated from DAZ1, DAZ2, DAZ3 and DAZ4, whereas those in the #155 sample originated from DAZ1, DAZ2 and DAZ4 (Fig. 5). The #113 and #147 samples included the original DAZ transcripts with one, two and three RRMs, whereas the #155 sample included the DAZ transcripts with only one and three RRMs (Fig. 5). Of note, these results are consistent with the Southern blot analysis of the RRM region in Fig. 4.

|
We then identified the DAZ transcripts that were actually expressed in the testis. The DAZ cDNA clones were expressed ectopically in 293T cells, and their sizes were compared with the endogenous DAZ proteins of the same testicular samples. Among the DAZ cDNA clones isolated, we found that three of the clones matched the DAZ-specific bands in each testicular sample (Fig. 5). In the #113 and #147 testes, expression of DAZ1, DAZ2 and DAZ3 was detected; we could not detect endogenous DAZ4 proteins although DAZ4 transcripts were identified in both cases (Fig. 5a, b). Consistent with the cloning data, DAZ1, DAZ2 and DAZ4 were expressed in the #155 testis (Fig. 5c). Interestingly, the testicular DAZ protein always matched the largest clone of each of three DAZ genes (Fig. 5), suggesting that the largest clone is the genuine transcript of each DAZ gene. Together, these results indicate that at least three or more DAZ genes are expressed in the sY1191-negative testes.
Discussion
In the present study, we determined the DAZ copy number in Y chromosomes with selective deletion of the sY1191 STS marker. The gene dosage assays predicted that one subject (#108) has two DAZ genes, whereas the remaining five subjects (#113, #147, #155, #033 and #124) have four DAZ genes. Similar results have been reported in a study in the Taiwanese population (Lin et al. 2007). Using expression analysis, we further confirmed that more than two DAZ genes with polymorphic structures are indeed expressed in the three sY1191-negative testes. Together, these results suggest that the selective absence of sY1191 not only means b2/b3 deletion with two DAZ genes, but also includes another AZFc configuration with four DAZ genes.
The b2/b3 deletion model for selective absence of sY1191 has been supported by FISH analysis (Repping et al. 2004). The same report also suggested that subsequent duplication in the b2/b3 deletion would restore the copy number of DAZ genes to four while retaining sY1191 negativity (Repping et al. 2004). The results of RRM Southern blot analysis in the present study suggest that, of the five sY1191-negative subjects with four DAZ genes, #124 could carry such a b2/b3 deletion–duplication. However, our results do not support that the remaining four subjects with four DAZ genes (#113, #147, #155 and #033) underwent the b2/b3 deletion and subsequent duplication, because they have three or more different DAZ genes rather than two types of DAZ genes. Thus, it is possible that non-homology-based recombination may contribute to the AZFc partial deletions in our subjects (for a review, see Navarro-Costa et al. 2010). The most striking example may be the possible presence of the DAZ genes on the short arm of the Y chromosome (Premi et al. 2010). Alternatively, it is speculated that sister chromatid exchanges following the deletion–duplication enabled sY1191-negative subjects to have four different DAZ genes (Lin et al. 2005, 2007).
We determined the expression of DAZ proteins in the three sY1191-negative subjects with male germ cells and observed at least three different kinds of DAZ proteins. The cDNA cloning results revealed that the #113 and #147 subjects express four DAZ transcripts. However, we failed to detect DAZ4 protein, probably because the DAZ4 band may be too weak to be detected under our immunoblot conditions. In samples from #155, we detected three DAZ transcripts and the corresponding proteins. In any case, we observed the presence of more than two DAZ genes, which indicates that sY1191 negativity is not confined to the b2/b3 deletion with two DAZ genes.
Several association studies have relied on the selective absence of sY1191 to identify b2/b3 deletions in large populations (Hucklenbroich et al. 2005; Wu et al. 2007; Lu et al. 2009; Eloualid et al. 2012). However, our results suggest that the selective absence of sY1191 represents heterogeneous AZFc configurations with different gene copy numbers. In this regard, the uncertainty of associations between b2/b3 deletion and male infertility may be due, in part, to heterogeneous AZFc structures of the sY1191-negative subjects used in previous studies. Large-scale variations in the AZFc region are widespread across human Y chromosome lineages (Repping et al. 2006). Our results also show the prevalence of variations in the AZFc region with respect to DAZ genes. Therefore, it is necessary to characterise the detailed organisation of the AZFc region in sY1191-negative subjects by developing and identifying deletions with novel amplicon-specific sequence markers (Navarro-Costa et al. 2007).
Acknowledgements
The authors thank P. H. Yen (Institute of Biomedical Sciences, Academia Sinica, Taipei, Taiwan) for generously providing us with the probes for Southern blot analyses. This study was supported by grants from the BioImaging Research Center at Gwangju Institute of Science and Technology (GIST), the Basic Research Program (grant no. 2010-0022423 and 2012R1A2A201003512) and the Science Research Center Program (grant no. 2011-0006425) of the Ministry of Education, Science and Technology of Korea. B. Kim was supported by the second stage of the Brain Korea 21 Project in 2012.
References
Eberhart, C. G., Maines, J. Z., and Wasserman, S. A. (1996). Meiotic cell cycle requirement for a fly homologue of human Deleted in Azoospermia. Nature 381, 783–785.| Meiotic cell cycle requirement for a fly homologue of human Deleted in Azoospermia.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaK28XjvVKgu7c%3D&md5=0970ac7dad57310ebefd6772ff8b1a96CAS | 8657280PubMed |
Eloualid, A., Rhaissi, H., Reguig, A., Bounaceur, S., El Houate, B., Abidi, O., Charif, M., Louanjli, N., Chadli, E., Barakat, A., Bashamboo, A., McElreavey, K., and Rouba, H. (2012). Association of spermatogenic failure with the b2/b3 partial AZFc deletion. PLoS One 7, e34902.
| Association of spermatogenic failure with the b2/b3 partial AZFc deletion.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC38Xmt1ehtLg%3D&md5=9da994c15e0e9cc6b9653dbf2f4d9260CAS | 22514689PubMed |
Fernandes, S., Huellen, K., Goncalves, J., Dukal, H., Zeisler, J., Rajpert De Meyts, E., Skakkebaek, N. E., Habermann, B., Krause, W., Sousa, M., Barros, A., and Vogt, P. H. (2002). High frequency of DAZ1/DAZ2 gene deletions in patients with severe oligozoospermia. Mol. Hum. Reprod. 8, 286–298.
| High frequency of DAZ1/DAZ2 gene deletions in patients with severe oligozoospermia.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD38XislSmsLw%3D&md5=97e13e89a9765634a65082b8203c6b34CAS | 11870237PubMed |
Fernandes, S., Paracchini, S., Meyer, L. H., Floridia, G., Tyler-Smith, C., and Vogt, P. H. (2004). A large AZFc deletion removes DAZ3/DAZ4 and nearby genes from men in Y haplogroup N. Am. J. Hum. Genet. 74, 180–187.
| A large AZFc deletion removes DAZ3/DAZ4 and nearby genes from men in Y haplogroup N.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2cXlvVOqsg%3D%3D&md5=5f7af30a03c744d64ac7ef7e132cde13CAS | 14639527PubMed |
Hucklenbroich, K., Gromoll, J., Heinrich, M., Hohoff, C., Nieschlag, E., and Simoni, M. (2005). Partial deletions in the AZFc region of the Y chromosome occur in men with impaired as well as normal spermatogenesis. Hum. Reprod. 20, 191–197.
| Partial deletions in the AZFc region of the Y chromosome occur in men with impaired as well as normal spermatogenesis.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2MXisVShtg%3D%3D&md5=20fdc512b687a984a830f86f5ae49f3eCAS | 15498781PubMed |
Karashima, T., Sugimoto, A., and Yamamoto, M. (2000). Caenorhabditis elegans homologue of the human azoospermia factor DAZ is required for oogenesis but not for spermatogenesis. Development 127, 1069–1079.
| 1:CAS:528:DC%2BD3cXit1aqu7c%3D&md5=b200eae6771efae43a0ccd91a4997c37CAS | 10662646PubMed |
Kim, B., Lee, Y., Kim, Y., Lee, K. H., Chun, S., Rhee, K., Seo, J. T., Kim, S. W., and Paick, J. S. (2009). Polymorphic expression of DAZ proteins in the human testis. Hum. Reprod. 24, 1507–1515.
| Polymorphic expression of DAZ proteins in the human testis.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD1MXmtlKqur0%3D&md5=64a4f13969c6b05d8321506c3c9d5248CAS | 19223287PubMed |
Kim, S. W., Kim, K. D., and Paick, J. S. (1999). Microdeletions within the azoospermia factor subregions of the Y chromosome in patients with idiopathic azoospermia. Fertil. Steril. 72, 349–353.
| Microdeletions within the azoospermia factor subregions of the Y chromosome in patients with idiopathic azoospermia.Crossref | GoogleScholarGoogle Scholar | 1:STN:280:DyaK1Mzmslyltg%3D%3D&md5=142b9b1ba639590e400bcdfb78acc822CAS | 10439009PubMed |
Kuroda-Kawaguchi, T., Skaletsky, H., Brown, L. G., Minx, P. J., Cordum, H. S., Waterston, R. H., Wilson, R. K., Silber, S., Oates, R., Rozen, S., and Page, D. C. (2001). The AZFc region of the Y chromosome features massive palindromes and uniform recurrent deletions in infertile men. Nat. Genet. 29, 279–286.
| The AZFc region of the Y chromosome features massive palindromes and uniform recurrent deletions in infertile men.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD3MXotlWhsLY%3D&md5=f1e80b5f342d42df4eb0188c99dcd4b2CAS | 11687796PubMed |
Lin, Y. W., Thi, D. A., Kuo, P. L., Hsu, C. C., Huang, B. D., Yu, Y. H., Vogt, P. H., Krause, W., Ferlin, A., Foresta, C., Bienvenu, T., Schempp, W., and Yen, P. H. (2005). Polymorphisms associated with the DAZ genes on the human Y chromosome. Genomics 86, 431–438.
| Polymorphisms associated with the DAZ genes on the human Y chromosome.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2MXpsFyqsb0%3D&md5=c23cd4e4bd606129a29e86f9599f7948CAS | 16085382PubMed |
Lin, Y. W., Hsu, C. L., and Yen, P. H. (2006). A two-step protocol for the detection of rearrangements at the AZFc region on the human Y chromosome. Mol. Hum. Reprod. 12, 347–351.
| A two-step protocol for the detection of rearrangements at the AZFc region on the human Y chromosome.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD28XkvVSrsrc%3D&md5=09a7f67bed31c4fc38b3f5d7facfb193CAS | 16608905PubMed |
Lin, Y. W., Hsu, L. C., Kuo, P. L., Huang, W. J., Chiang, H. S., Yeh, S. D., Hsu, T. Y., Yu, Y. H., Hsiao, K. N., Cantor, R. M., and Yen, P. H. (2007). Partial duplication at AZFc on the Y chromosome is a risk factor for impaired spermatogenesis in Han Chinese in Taiwan. Hum. Mutat. 28, 486–494.
| Partial duplication at AZFc on the Y chromosome is a risk factor for impaired spermatogenesis in Han Chinese in Taiwan.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2sXlsVyqu74%3D&md5=5404f553bbb8a15bb158cad36dbb8bf1CAS | 17285591PubMed |
Lu, C., Zhang, J., Li, Y., Xia, Y., Zhang, F., Wu, B., Wu, W., Ji, G., Gu, A., Wang, S., Jin, L., and Wang, X. (2009). The b2/b3 subdeletion shows higher risk of spermatogenic failure and higher frequency of complete AZFc deletion than the gr/gr subdeletion in a Chinese population. Hum. Mol. Genet. 18, 1122–1130.
| The b2/b3 subdeletion shows higher risk of spermatogenic failure and higher frequency of complete AZFc deletion than the gr/gr subdeletion in a Chinese population.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD1MXisFSmtLs%3D&md5=5fffb9300a87c0875d314329a6e575e1CAS | 19088127PubMed |
Lynch, M., Cram, D. S., Reilly, A., O’Bryan, M. K., Baker, H. W., de Kretser, D. M., and McLachlan, R. I. (2005). The Y chromosome gr/gr subdeletion is associated with male infertility. Mol. Hum. Reprod. 11, 507–512.
| The Y chromosome gr/gr subdeletion is associated with male infertility.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2MXpsVaksb0%3D&md5=f8380984494e2f91e554d0a5e655978dCAS | 16123079PubMed |
Machev, N., Saut, N., Longepied, G., Terriou, P., Navarro, A., Levy, N., Guichaoua, M., Metzler-Guillemain, C., Collignon, P., Frances, A. M., Belougne, J., Clemente, E., Chiaroni, J., Chevillard, C., Durand, C., Ducourneau, A., Pech, N., McElreavey, K., Mattei, M. G., and Mitchell, M. J. (2004). Sequence family variant loss from the AZFc interval of the human Y chromosome, but not gene copy loss, is strongly associated with male infertility. J. Med. Genet. 41, 814–825.
| Sequence family variant loss from the AZFc interval of the human Y chromosome, but not gene copy loss, is strongly associated with male infertility.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2cXhtFSjur%2FK&md5=9e48631ad0adc99516b5955ed1ecbb7eCAS | 15520406PubMed |
Navarro-Costa, P., Pereira, L., Alves, C., Gusmão, L., Proença, C., Marques-Vidal, P., Rocha, T., Correia, S. C., Jorge, S., Neves, A., Soares, A. P., Nunes, J., Calhaz-Jorge, C., Amorim, A., Plancha, C. E., and Gonçalves, J. (2007). Characterizing partial AZFc deletions of the Y chromosome with amplicon-specific sequence markers. BMC Genomics 8, 342.
| Characterizing partial AZFc deletions of the Y chromosome with amplicon-specific sequence markers.Crossref | GoogleScholarGoogle Scholar | 17903263PubMed |
Navarro-Costa, P., Gonçalves, J., and Plancha, C. E. (2010). The AZFc region of the Y chromosome: at the crossroads between genetic diversity and male infertility. Hum. Reprod. Update 16, 525–542.
| The AZFc region of the Y chromosome: at the crossroads between genetic diversity and male infertility.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3cXhtVektr7J&md5=da9bd105f01d73c3ed08b741e8f6ccc1CAS | 20304777PubMed |
Premi, S., Srivastava, J., Epplen, J. T., and Ali, S. (2010). AZFc region of the Y chromosome shows singular structural organization. Chromosome Res. 18, 419–430.
| AZFc region of the Y chromosome shows singular structural organization.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3cXnsFGksbc%3D&md5=5d9b6e0c03c888bed28b350ea49b939aCAS | 20373140PubMed |
Ravel, C., Chantot-Bastaraud, S., El Houate, B., Mandelbaum, J., Siffroi, J. P., and McElreavey, K. (2006). GR/GR deletions within the azoospermia factor c region on the Y chromosome might not be associated with spermatogenic failure. Fertil. Steril. 85, 229–231.
| GR/GR deletions within the azoospermia factor c region on the Y chromosome might not be associated with spermatogenic failure.Crossref | GoogleScholarGoogle Scholar | 16412764PubMed |
Reijo, R., Lee, T. Y., Salo, P., Allagappan, R., Brown, L. G., Rosenberg, M., Rozen, S., Jaffe, T., Straus, D., Hovatta, O., de la Chapelle, A., Silber, S., and Page, D. C. (1995). Diverse spermatogenic defects in humans caused by Y chromosome deletions encompassing a novel RNA-binding protein gene. Nat. Genet. 10, 383–393.
| Diverse spermatogenic defects in humans caused by Y chromosome deletions encompassing a novel RNA-binding protein gene.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaK2MXnsVWlu7g%3D&md5=117c213735d9f70e2d8f765252c649c0CAS | 7670487PubMed |
Repping, S., Skaletsky, H., Brown, L., van Daalen, S. K., Korver, C. M., Pyntikova, T., Kuroda-Kawaguchi, T., de Vries, J. W., Oates, R. D., Silber, S., van der Veen, F., Page, D. C., and Rozen, S. (2003). Polymorphism for a 1.6-Mb deletion of the human Y chromosome persists through balance between recurrent mutation and haploid selection. Nat. Genet. 35, 247–251.
| Polymorphism for a 1.6-Mb deletion of the human Y chromosome persists through balance between recurrent mutation and haploid selection.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD3sXosFWrtrw%3D&md5=e64676038f75f821cf8e3f01e84d34ecCAS | 14528305PubMed |
Repping, S., van Daalen, S. K., Korver, C. M., Brown, L. G., Marszalek, J. D., Gianotten, J., Oates, R. D., Silber, S., van der Veen, F., Page, D. C., and Rozen, S. (2004). A family of human Y chromosomes has dispersed throughout northern Eurasia despite a 1.8-Mb deletion in the azoospermia factor c region. Genomics 83, 1046–1052.
| A family of human Y chromosomes has dispersed throughout northern Eurasia despite a 1.8-Mb deletion in the azoospermia factor c region.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2cXksFKisLc%3D&md5=df030e62f5125548b339d204544cb503CAS | 15177557PubMed |
Repping, S., van Daalen, S. K., Brown, L. G., Korver, C. M., Lange, J., Marszalek, J. D., Pyntikova, T., van der Veen, F., Skaletsky, H., Page, D. C., and Rozen, S. (2006). High mutation rates have driven extensive structural polymorphism among human Y chromosomes. Nat. Genet. 38, 463–467.
| High mutation rates have driven extensive structural polymorphism among human Y chromosomes.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD28XivFSht7s%3D&md5=aaf6c27fa64a01c256e0e326b27c4bf6CAS | 16501575PubMed |
Ruggiu, M., Speed, R., Taggart, M., McKay, S. J., Kilanowski, F., Saunders, P., Dorin, J., and Cooke, H. J. (1997). The mouse Dazla gene encodes a cytoplasmic protein essential for gametogenesis. Nature 389, 73–77.
| The mouse Dazla gene encodes a cytoplasmic protein essential for gametogenesis.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaK2sXlvVCksr8%3D&md5=7527e5adc50fe3322afd5580a140fe3aCAS | 9288969PubMed |
Sambrook, J., Fritsch, E. F., and Maniatis, T. (1989). ‘Molecular Cloning: A Laboratory Manual.’ (Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY.)
Saxena, R., Brown, L. G., Hawkins, T., Alagappan, R. K., Skaletsky, H., Reeve, M. P., Reijo, R., Rozen, S., Dinulos, M. B., Disteche, C. M., and Page, D. C. (1996). The DAZ gene cluster on the human Y chromosome arose from an autosomal gene that was transposed, repeatedly amplified and pruned. Nat. Genet. 14, 292–299.
| The DAZ gene cluster on the human Y chromosome arose from an autosomal gene that was transposed, repeatedly amplified and pruned.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaK28XntVClsbo%3D&md5=27e2f5ab61886e77dd1c9f81b19ec6e6CAS | 8896558PubMed |
Saxena, R., de Vries, J. W., Repping, S., Alagappan, R. K., Skaletsky, H., Brown, L. G., Ma, P., Chen, E., Hoovers, J. M., and Page, D. C. (2000). Four DAZ genes in two clusters found in the AZFc region of the human Y chromosome. Genomics 67, 256–267.
| Four DAZ genes in two clusters found in the AZFc region of the human Y chromosome.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD3cXlsVKlu78%3D&md5=fb9648eb752d348ff96438b6ee54b1d3CAS | 10936047PubMed |
Simoni, M., Bakker, E., and Krausz, C. (2004). EAA/EMQN best practice guidelines for molecular diagnosis of Y-chromosomal microdeletions. State of the art 2004. Int. J. Androl. 27, 240–249.
| EAA/EMQN best practice guidelines for molecular diagnosis of Y-chromosomal microdeletions. State of the art 2004.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2cXns1GmsLc%3D&md5=c52e87d6976c0b06b3bb53c0e5dfba5dCAS | 15271204PubMed |
Skaletsky, H., Kuroda-Kawaguchi, T., Minx, P. J., Cordum, H. S., Hillier, L., Brown, L. G., Repping, S., Pyntikova, T., Ali, J., Bieri, T., et al. (2003). The male-specific region of the human Y chromosome is a mosaic of discrete sequence classes. Nature 423, 825–837.
| The male-specific region of the human Y chromosome is a mosaic of discrete sequence classes.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD3sXks1Kmtbk%3D&md5=b9b577ec71a80ba1eaa240ab9bcd061eCAS | 12815422PubMed |
Thornhill, A. R., Guenther, A. J., Barbarotto, G. M., Session, D. R., Damario, M. A., Dumesic, D. A., and Snow, K. (2002). False-positive Y-microdeletion result for a fertile male caused by an alteration under a PCR primer. Int. J. Androl. 25, 352–357.
| False-positive Y-microdeletion result for a fertile male caused by an alteration under a PCR primer.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD38XptlSmt74%3D&md5=d7e7c1ed666fa35bb3b4022502c2d336CAS | 12406367PubMed |
VanGompel, M. J., and Xu, E. Y. (2010). A novel requirement in mammalian spermatid differentiation for the DAZ-family protein Boule. Hum. Mol. Genet. 19, 2360–2369.
| A novel requirement in mammalian spermatid differentiation for the DAZ-family protein Boule.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3cXms1Oks70%3D&md5=e2c3110cc8b77e2d370c835c7b72bb02CAS | 20335278PubMed |
Wu, B., Lu, N. X., Xia, Y. K., Gu, A. H., Lu, C. C., Wang, W., Song, L., Wang, S. L., Shen, H. B., and Wang, X. R. (2007). A frequent Y chromosome b2/b3 subdeletion shows strong association with male infertility in Han-Chinese population. Hum. Reprod. 22, 1107–1113.
| A frequent Y chromosome b2/b3 subdeletion shows strong association with male infertility in Han-Chinese population.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2sXlvVOis7w%3D&md5=a07c8bf9ab3b0099c9d1ed4c5a9dcf65CAS | 17204527PubMed |
Wu, Q., Chen, G. W., Yan, T. F., Wang, H., Liu, Y. L., Li, Z., Duan, S. W., Sun, F., Feng, Y., and Shi, H. J. (2011). Prevalent false positives of azoospermia factor a (AZFa) microdeletions caused by single-nucleotide polymorphism rs72609647 in the sY84 screening of male infertility. Asian J. Androl. 13, 877–880.
| Prevalent false positives of azoospermia factor a (AZFa) microdeletions caused by single-nucleotide polymorphism rs72609647 in the sY84 screening of male infertility.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3MXhsVarsbnJ&md5=01b653e8b5b0c89807f82f54f4eacb92CAS | 21765443PubMed |
Yen, P. H., Chai, N. N., and Salido, E. C. (1996). The human autosomal gene DAZLA: testis specificity and a candidate for male infertility. Hum. Mol. Genet. 5, 2013–2017.
| The human autosomal gene DAZLA: testis specificity and a candidate for male infertility.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaK28XnsFOhsrg%3D&md5=65882498a580f5a035d764148d19b6e4CAS | 8968756PubMed |
Yen, P. H., Chai, N. N., and Salido, E. C. (1997). The human DAZ genes, a putative male infertility factor on the Y chromosome, are highly polymorphic in the DAZ repeat regions. Mamm. Genome 8, 756–759.
| The human DAZ genes, a putative male infertility factor on the Y chromosome, are highly polymorphic in the DAZ repeat regions.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaK2sXmsleitbg%3D&md5=bcbb4889b59bcf5fb9f4cf2e5e1c7edfCAS | 9321470PubMed |