

Nitrification rates and associated nitrous oxide emissions from agricultural soils – a synopsis
Ryan FarquharsonCSIRO Agriculture, PMB2 Glen Osmond, SA 5064, Australia. Email: ryan.farquharson@csiro.au
Soil Research 54(5) 469-480 https://doi.org/10.1071/SR15304
Submitted: 23 October 2015 Accepted: 15 February 2016 Published: 27 June 2016
Journal Compilation © CSIRO Publishing 2016 Open Access CC BY-NC-ND
Abstract
Laboratory incubations were performed to estimate nitrification rates and the associated nitrous oxide (N2O) emissions under aerobic conditions on a range of soils from National Agricultural Nitrous Oxide Research Program field sites. Significant site-to-site variability in nitrification rates and associated N2O emissions was observed under standardised conditions, indicating the need for site-specific model parameterisation. Generally, nitrification rates and N2O emissions increased with higher water content, ammonium concentration and temperature, although there were exceptions. It is recommended that site-specific model parameterisation be informed by such data. Importantly, the ratio of N2O emitted to net nitrified N under aerobic conditions was small (<0.2% for the majority of measurements) but did vary from 0.03% to 1%. Some models now include variation in the proportion of nitrified N emitted as N2O as a function of water content; however, strong support for this was not found across all of our experiments, and the results demonstrate a potential role of pH and ammonium availability. Further research into fluctuating oxygen availability and the coupling of biotic and abiotic processes will be required to progress the process understanding of N2O emissions from nitrification.
Additional keywords: agriculture, ammonia oxidation, modelling, nitrogen.
Introduction
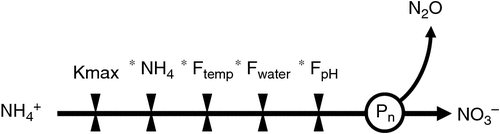
Nitrous oxide (N2O) emissions from agricultural systems are thought to result principally from the microbial-mediated processes of nitrification (the oxidation of ammonium to nitrate) and denitrification (the reduction of nitrate to N2O and dinitrogen) in soil, although other processes and combinations thereof exist (Butterbach-Bahl et al. 2013). Most agro-ecological models (e.g. DayCent, APSIM, DNDC, WNMM, NOE) explicitly deal with each of these processes with varying degrees of process understanding and empiricism (Li et al. 1992; Parton et al. 1998; Hénault et al. 2005; Li et al. 2007; Thorburn et al. 2010). By necessity, these processes are often simplistically modelled using potential process rates that then are modified by drivers such as substrate availability, water content, pH and temperature (Fig. 1) following the ‘hole-in-the-pipe’ schema of Firestone and Davidson (1989). Although this approach may be suitable for a particular site in a given year, such models may not be portable because the true mechanisms, which can vary over space and time, have not been captured.
|
Nitrification in agricultural systems is thought to be mostly carried out by autotrophic bacteria (Jia and Conrad 2009), although some autotrophic archaea and heterotrophs can also nitrify. The mechanism of N2O production from nitrification has not been systematically determined (Khalil et al. 2004; Shaw et al. 2006). The oxidative decomposition of hydroxylamine (Bremner et al. 1980; Hooper and Terry 1979) and nitrifier denitrification (Poth and Focht 1985; Zhu et al. 2013a) (i.e. denitrification by ammonia oxidising bacteria, as proposed by Baggs (2011)) have been implicated, with a key role for nitrite suggested (Mørkved et al. 2007). The roles of abiotic factors such as metal ion species, including iron III and manganese II, in the oxidation of hydroxylamine (Hooper and Terry 1979; Bremner et al. 1980; Zhu et al. 2013b; Heil et al. 2015), chemical or thermal decomposition of hydroxylamine, and processes such as nitrifier denitrification under conditions of oxygen stress have not been widely acknowledged or included in models. Consequently, most models adopt a grey-box approach to predicting N2O from nitrification, allocating a set proportion of nitrified N to N2O emission (Pn in Fig. 1). Some exceptions to the use of fixed proportions exist. For example, the empirically based NOE model (Bessou et al. 2010) now incorporates a function based on water content as a rudimentary proxy for the influence of oxygen availability on the proportion of nitrified N emitted as N2O (Pn), as observed by Khalil et al. (2004), and the ecosys model explicitly represents the role of nitrite in the generation of N2O (Grant 1995). However, there remains a paucity of data to support these approaches across a range of soils.
Few studies have explicitly reported nitrification rates and the associated N2O emissions. Of the reports we could find in the literature (Table 1), the varying methodologies used, range of experimental systems involved and dearth of data preclude a meta-analysis to determine which factors influence the proportion or ratio of N2O produced to nitrified N across a range of soils. Studies have used pure and mixed cultures of various nitrifiers, soil slurries, loose soil incubations, repacked cores, intact cores and in situ techniques. Measurements have relied on the use of inhibitors or strictly aerobic conditions to prevent denitrification, and isotopic labels, but rarely has it been clearly stated whether the calculations were based on net or gross rates, which may account for some of the variance in observed product ratios. Of the factors examined, a negative relationship between oxygen availability and Pn was the most common observation, with the most decisive work being done by using cultures (Goreau et al. 1980; Lipschultz et al. 1981; Hynes and Knowles 1984; Kester et al. 1997) with varying oxygen contents, although cell density was also important (Remde and Conrad 1990; Frame and Casciotti 2010). In addition, oxygen availability has been shown to be important in soil, either directly (Goodroad and Keeney 1984; Khalil et al. 2004; Mørkved et al. 2006; Zhu et al. 2013a) or via water content (Maag and Vinther 1996; Garrido et al. 2002; Ambus 2005; Mathieu et al. 2006; Chen et al. 2010), although some studies have found conflicting responses (Bateman and Baggs 2005). Interestingly, Hynes and Knowles (1978) found that the addition of sterile soil to a culture increased N2O emission 10-fold and broadened the range of oxygen concentrations over which high N2O emissions occurred. There were conflicting responses of Pn to temperature (Goodroad and Keeney 1984; Maag and Vinther 1996; Ingwersen et al. 1999). Martikainen (1985) found an exponential increase in Pn as pH declined from 4.7 to 4.1 in some acidic forest soils, and Mørkved et al. (2007) reported N2O emissions two orders of magnitude higher in soils of pH <5, possibly due to chemodenitrification. However, other studies were less conclusive about the influence of pH (Goodroad and Keeney 1984; Mørkved et al. 2006). Jiang and Bakken (1999) noted that Pn increased when conditions were unfavourable to nitrification, i.e. because of acid and low ammonium availability. The source of nitrogen also seems to be important, with urine and urea giving higher Pn values than ammonium (Carter 2007; Zhu et al. 2013a). Recent work has again highlighted other abiotic factors such as metal species (Zhu et al. 2013b; Heil et al. 2015) as being important determinants of N2O emissions from nitrification. In addition, the role of ammonia sorption in moderating the inhibitory effect of ammonia on nitrite oxidation and hence N2O from nitrification was recently identified (Venterea et al. 2015).

|
It has been suggested that, in many Australian environments, nitrification may be a significant contributor to N2O emissions because much of our broadacre agriculture occurs in a semi-arid climate and soil fertility is relatively low (e.g. Barton et al. 2008, 2010; 2011). However, few studies attribute N2O emissions to the actual processes that produce them, notwithstanding the diversity in soils, climate and management practices. Without this knowledge, estimates of the contribution of nitrification v. denitrification to total N2O fluxes can vary widely, and the ability to develop robust mitigation strategies remains compromised.
This study used laboratory-based incubations to estimate nitrification rates and associated N2O emissions under standard conditions in a range of soils from field sites within the National Agricultural Nitrous Oxide Research Program (see papers from this special journal issue for details). The objectives of the study were to provide evidence for site-specific parameterisation of models and to test model assumptions. In addition, incubation conditions were modified to explore whether any factors result in changes to the proportion of nitrified N emitted as N2O, which for many models has been a fixed value with limited experimental evidence. The goal remains to derive new algorithms where sufficient information is available so that models can be modified to account for the known variability in potential nitrification rates and the proportion of nitrified N emitted as N2O. Such models will be crucial for developing mitigation strategies and performing life-cycle analyses and are likely to have a role in improving inventories of N2O emissions for the agricultural sector.
Materials and methods
A simple incubation technique was used to allow an approximation of potential nitrification rates and the associated N2O emissions in standardised laboratory incubations, adopting similar methods to Garrido et al. (2002) and Freney et al. (1979), who conducted extensive method testing. The method relies on maintaining aerobic conditions in which case denitrification does not occur. Assuming that no nitrate is immobilised and abiotic production of N2O is non-existent, the increase in nitrate is used to calculate the nitrification rate, and all N2O produced can be attributed to nitrification. Although net nitrification rates were measured, we anticipate that the net nitrification rates approximate the gross rates under the conditions of the experiments. To test whether the assumption of no denitrification holds, acetylene at 10 kPa was added to a subset of samples representative of each treatment within each experiment to inhibit both nitrification and the reduction of N2O to N2 by denitrifiers (Davidson et al. 1986; Klemedtsson et al. 1988). This test allowed the detection of denitrification for each experimental treatment to give some confidence in the validity of the results. A pre-incubation period allowed the soil biota to resuscitate after soil sampling, transport and preparation, as well as avoiding flushes of activity on wetting up.
Soils and incubation
Soil sampling
Soils were sampled from each of the sites in Table 2. Soil samples from all sites were collected at 0–10 cm depth, and an additional sample at the Camden site was collected at 0–2 cm depth. Samples were air-dried at 40°C in a draught oven (except for Lucaston where field-moist samples were immediately transported on ice to preserve the temperature sensitivity of the microbial community) before transportation to the laboratory.

|
Soil preparation
Air-dried soil was sieved to ≤5 mm (the Hamilton soil was also sieved to ≤2 mm) to exclude large chunks of root material and gravel. The gravimetric water content of the sieved soil at 100 cm of water suction or 10 kPa (denoted θ10kPa) was measured by using suction plates (Table 2).
Experimental units were prepared by weighing 7 g (oven-dry equivalent) of soil into 50-mL centrifuge tubes and then tapping the tube gently on a workbench to aid settling of the soil and give an even surface. The experiments are therefore best described as loose soil incubations, although some settling to a natural bulk density occurred, but was not quantified.
Pre-incubation
Soils were pre-incubated to allow microbial activity to recover after sampling and processing, and to avoid flushes of activity on wetting up. Soils were pre-incubated by wetting up to a defined water content with Milli-Q water, which allowed the addition of 250 μL substrate at the commencement of the incubation proper (see below). Moisture and air exchange were maintained for 2–3 weeks at 22°C in a constant-temperature room. In some experiments, additional treatments involving the pre-incubation phase (e.g. no pre-incubation to examine initial flushes of activity and thermal acclimation) were included.
Incubation proper
At the start of the incubation proper, water contents were adjusted and substrate added, giving the final water content as defined in Table 3 as a percentage of θ10kPa and 60% θ10kPa for the Kingaroy and Lucaston experiments. For most experiments, ammonium sulfate at 100 μg N g–1 dry soil was added, but ammonium concentrations were altered in some experiments as per Table 3. All solutions were added by pipetting evenly onto the soil surface. Tubes were immediately capped with Suba-Seals (Sigma-Aldrich, St Louis, MO, USA) then equilibrated to atmospheric pressure by using an open-ended needle. Soils were incubated for 2–4 days. Various treatments were imposed at time zero, usually in a balanced factorial design with four replicates.

|
The consumption of oxygen was quantified in preliminary experiments using CO2 measurements and found to be well below the oxygen available in the headspace, although it is acknowledged that localised anaerobiosis in microsites is possible. Hence, acetylene was used in a subset of samples to detect denitrification. Within each replicate an additional sample of each treatment was injected with 10 kPa of acetylene (10% of an atmosphere), with the accumulation of N2O being indicative of denitrification, because nitrous oxide reductase activity was blocked (Yoshinari and Knowles 1976) simultaneously with ammonia oxidase (Hynes and Knowles 1978; Bremner and Blackmer 1979; Walter et al. 1979).
Time zero samples were sampled immediately as described below. Remaining samples were incubated at 22°C in a constant temperature room, except for the Lucaston experiment in which water baths were used to modify the temperatures (see Table 3).
Incubation sampling
Prior to gas sampling, the soil atmosphere was mixed with the headspace by vigorously bashing each tube on the bench. Tubes were sampled by taking gas samples in excess of 20 mL through the Suba-Seal using a Hamilton 50-mL gas-tight syringe (Hamilton Company, Reno, NV, USA) equipped with a valve and side-port taper needles. The use of an oversized syringe and negative pressure also assisted with mixing the soil and headspace atmospheres. Gas (20 mL at atmospheric pressure) was then injected into evacuated glass 12-mL Exetainers (Labco, Lampeter, UK). Extracts of the soil (1 : 5 KCl) were performed by adding 35 mL of 2 M KCl directly to the tubes immediately after gas sampling and shaking for 2 h. The tubes were centrifuged and the supernatant was filtered before freezing until further analysis.
Analyses
The gas samples were analysed for N2O, CO2 and CH4 on an Agilent 7890A gas chromatograph (Agilent Technologies, Santa Clara, CA, USA) fitted with an electron capture detector (ECD), thermal conductivity detector and flame ionisation detector and equipped with HayeSep packed columns (HayeSeparations Inc., Bandera, TX, USA) using helium carrier gas with 10% CH4 in argon as a makeup gas for the ECD. KCl extracts were analysed for NH4-N and NOx-N by flow injection colourimetry on a Lachat QuikChem 8500 series 2 (Lachat, Loveland, CO, USA). Nitrate was reduced to nitrite by Cd reduction, and the resultant nitrite was reacted with N-1-naphthylethylenediamine dihydrochloride with sulphanilamide; ammonium was determined after reaction with sodium salicylate and dichloro-isocyanurate as per Rayment and Lyons (2011). A subset of samples was analysed for nitrite alone, but levels were negligible so all NOx-N was assumed to be NO3-N.
Calculations
The method used assumes that no denitrification occurred and a check was performed using acetylene to block both nitrification and the reduction of N2O to N2 by denitrifiers. No increase in N2O concentration over time in tubes with acetylene for a particular treatment provided confidence that no denitrification occurred within that treatment.
Where no denitrification occurred, the increase in nitrate from time zero approximates the nitrification rate because the nitrate was not consumed by denitrification, and NO loss and NO2– accumulation (tested in preliminary experiments) were considered minimal. Immobilisation of nitrate was also considered minimal given the pre-incubation period and this was supported by measurements of mineral N at each stage of the experiment. The increase in headspace N2O from time zero was considered to be the N2O produced via nitrification, because no denitrification occurred and N2O production from other sources was assumed to be non-existent.
Both the nitrate accumulation (nitrification) and associated N2O emissions are presented as a mass of nitrogen transformed g–1 soil day–1.
In addition to N2O in the soil atmosphere and headspace, dissolved N2O was calculated by assuming equilibration with the headspace and using the Henry’s law constant, the concentration of N2O in the headspace and the temperature of the sample (Hudson 2004). Thus, the total amounts of N2O produced (soil atmosphere and headspace plus dissolved N2O) are presented.
Finally, the N2O from nitrification was normalised by dividing the N2O production rate by the apparent nitrification rate. Most publications express N2O as a proportion of nitrified N; however, this may be misleading in situations where only net nitrification rates were measured. This metric should be reserved for experiments where the gross nitrification rate was measured. Regardless, in our work, there should be only a minor variance between the ratio and the proportion because the production of N2O from nitrification was generally three orders of magnitude lower than the nitrification rate, and gross nitrification should be reasonably close to the net nitrification given the conditions of the experiment.
Statistical analyses
All statistical analysis was done using Genstat 16 (VSN International, Hemel Hempstead, UK). Each experiment was analysed separately by using a balanced design and a 1-, 2- or 3-factor analysis of variance, depending on the number of treatments. Data were checked for homogeneity of variance and normality. In some experiments, the treatments were non-orthogonal, in which case subsets of treatments were tested separately. In some experiments, obvious outliers were identified and with additional evidence were omitted from the analysis. In these cases, an unbalanced analysis of variance was performed.
Results
Potential nitrification rates ranged from <1 to 33 μg N g–1 soil day–1, with the majority of cases being <6 μg N g–1 soil day–1. The potential nitrification rate varied by site and by treatments such as soil depth, water content, temperature and ammonium concentration.
The N2O emissions associated with nitrification ranged from <1 to 50 ηg N g–1 soil day–1, although the upper values were subject to large within-replicate variability. Generally, when potential nitrification rates were high, so too were the associated N2O emissions, although there were some discrepancies resulting in variation in the ratio of N2O produced to the nitrification rate.
The ratio of N2O to nitrification rate ranged from 0.03% to 1%, although the high values were associated with greater variability within replicates and could be considered an overestimate. There was significant site-to-site variation in the ratio (see Table 3), and factors such as water content, ammonium availability, pH and temperature also affected it, but often in interactions with other treatments and sometimes inconsistently across experiments. Relationships between the nitrification rate and N2O grouped by site can be seen in Fig. 2.

|
Water content
In general, the nitrification rates increased with water content, as did the associated N2O emissions. In some cases (Tamworth, Kingsthorpe), the nitrification-derived N2O increased more than the nitrification rate itself with increasing water content, so the ratio of the N2O produced to the nitrification rate (i.e. Pn) was greater at higher water contents. However, the increase in Pn with water content was not widespread as demonstrated by the Camden, Griffith lysimeters soils, Terang and Noorat experiments.
Ammonium availability
In the Tamworth experiment, increasing ammonium availability up to 50 μg N g–1 soil did result in higher nitrification rates for soil at 60% θ10kPa but not at 40% θ10kPa. The N2O associated with nitrification and the ratio of N2O produced to the nitrification rate appeared to increase with N addition, although there was significant variability in these data. There was also an increase in N2O from nitrification with increasing NH4 addition in the Kingsthorpe soil at 50% θ10kPa.
The addition of NH4-N at 100 μg N g–1 soil had minimal effects on nitrification rates compared with no additional ammonium in the Kingaroy experiment; however, higher N2O emissions and proportions of nitrified N emitted as N2O were evident with N addition, particularly on the soil with a legume history.
Soil depth
Potential nitrification rates were higher in the surface 0–2 cm than the 0–10 cm soil from the Camden site. Similarly, the rate of N2O production associated with nitrification was higher in the surface soil. Accordingly, there was little variance in the proportion of nitrified N emitted as N2O by depth.
Aggregate size
In the Hamilton experiment, the soil was sieved to ≤5 mm as in other experiments, and to ≤2 mm in an attempt to reduce variability thought to be associated with aggregates and particulate organic matter that was observed in a preliminary experiment. In this experiment, the sieve size had little effect on nitrification rates and associated N2O emissions.
pH
The pH of the Wagga Wagga soil was manipulated by the addition of calcium hydroxide, raising the pH from 5.5 to 7.5 during the 3-week pre-incubation period. The nitrification rate was only marginally higher in the higher pH soil, whereas the N2O from nitrification was significantly higher in the higher pH soil at 80% θ10kPa but not at 60% θ10kPa. The proportion of nitrified N emitted as N2O was on average higher in the higher pH soil (0.056% v. 0.035%), but these values were still much lower than in other soils.
History
In the Kingaroy experiment, soils with a legume or grass history were compared. Nitrification rates were higher in the soil with a legume history and this soil responded to added N compared with the soil with a grass history, which did not respond to added N.
Temperature
The Lucaston experiment demonstrated a peak in the nitrification rate at 35°C and very low nitrification rates at 45°C. By contrast, the associated N2O emissions were relatively consistent with the highest emission coming from the 45°C treatment. Accordingly, we calculated that 1% of the nitrified N was emitted as N2O at 45°C but acknowledge the variability in that treatment. The possibility of thermal decomposition of hydroxylamine to N2O at these high temperatures was noted.
Texture
The three soils of different texture (sand, loam and clay loam) from the Griffith lysimeters were compared within the same experiment, but water contents were set as proportions of water content at 10 kPa of suction to provide some semblance of attainable water contents in the field rather than on a volumetric or gravimetric basis. At similar proportions of θ10kPa, the loam soil had the highest nitrification rates, which increased with water content, but also the lowest proportion of nitrified N emitted as N2O. The high nitrification rates in the loam were consistent with lysimeter studies where this soil had higher mineralisation rates and accumulated more nitrate (Jamali et al. 2016).
Discussion
The contribution of nitrification to N2O emissions under aerobic conditions is likely to be small, with ratios of N2O emitted to nitrate accumulation generally peaking at 0.5% for some sites, but being <0.1% for most sites and conditions. Evidently, using a fixed value for the proportion of nitrified N emitted as N2O in models is not correct, although for some soils (e.g. Camden) the value calculated was reasonably consistent across treatments. The values we obtained for the ratio of N2O emitted to nitrate accumulated are more constrained in range than published proportions, which range from 0.01% to >20% (Table 1). It should be noted, though, that the values obtained in the present work are associated with aerobic conditions, and it is probable that the proportion of nitrified N emitted as N2O will increase with fluctuating aerobic and anaerobic conditions. In particular, Yu et al. (2010), using pure cultures, showed that N2O production from nitrifiers occurs on the transition from anaerobic to aerobic conditions and is related to the accumulation of ammonium during the anaerobic phase.
The nitrification rates estimated here are intended to set an upper limit on the rate of conversion of ammonium to nitrate for each soil at a standard temperature of 22°C. Although it is not possible to extrapolate directly from laboratory-based assays to rates in the field, such data can be used to modify potential nitrification rates in models in a relative sense and provide an evidence-based means to do so when consistent methodology is used. The site-to-site variation in potential nitrification rates demonstrates the need to adjust the models for each site, but to do so without evidence risks developing models that provide reasonable simulations for the wrong reasons and can limit the portability of such models through both space and time. Simple incubation methods such as used in this work provide independent evidence for site-specific calibration instead of relying purely on mathematical or non-objective means of calibrating models, and have been employed in some modelling strategies such as NOE (Hénault et al. 2005), where empirical evidence is used to parameterise the model.
Overall, between-site variation in potential nitrification rates and associated N2O emissions was greater than within-site variation, demonstrating the need to characterise and parameterise each site individually. Further analysis is required to determine which properties govern the site-to-site variation; however, from the results a few candidates emerge.
The Camden soil had higher potential nitrification rates across a range of water contents than other soils, indicative of rapid nitrogen cycling, which, if uncoupled from nitrogen uptake by plants or microorganisms (e.g. from land-use change), might result in high fluxes of N2O. This scenario can be seen from the field-scale measurements at the Hamilton site where cropping has been introduced into productive pasture (Belyaeva et al. 2016).
With the exception of Noorat, the cropping sites appear to have higher proportions of nitrified N emitted as N2O than the dairy pasture sites. Until a firm mechanism for N2O production via nitrification is identified, it is very difficult to determine why. Possible explanations may be related to soil type and longer term management such as soil disturbance, nitrogen addition or efficiency of plant uptake. The Kingaroy site in particular had a high proportion of nitrified N emitted as N2O. In this soil, the nitrification rate was not very sensitive to the addition of ammonium, with added ammonium at 0 and 100 μg N g–1 soil having similar but low rates of nitrification. However, the N2O from nitrification was sensitive to the addition of ammonium, especially in the soil from the legume site. We hypothesise that the presence of legumes altered the soil microbial community, resulting in the observed differences. The Tamworth site was notable for the relatively high N2O emissions associated with nitrification that was responsive to the addition of ammonium. The consistent presence of denitrification in the 80% θ10kPa treatment makes the Tamworth soil suitable for further studies, including examining water relations, oxygen availability and thresholds for the presence of denitrification.
Nitrification rates increased with increasing water content, even at water contents generally thought to inhibit aerobic processes (e.g. the 90% θ10kPa treatment in the Hanwood loam). Indeed, water content is widely used as a proxy for the aerobicity of soil even though water can contain significant amounts of dissolved oxygen, and despite decoupling of the actual drivers and water content having been demonstrated previously (e.g. Hall et al. 2013). Water also affects the diffusion of solutes, not just gases, so the capture of these multiple effects is challenging.
Water content was not as strong a predictor of the proportion of nitrified N emitted as N2O as anticipated. For example, in the Camden soil, the nitrification rate increased with water content, as did the N2O from nitrification. Hence, the proportion of nitrified N emitted as N2O remained constant with water content at that site. However, in other experiments, the proportion of nitrified N emitted as N2O was greater at higher water contents, which is consistent with other reports (Table 1).
In some soils, a clear threshold of water content was evident for denitrification being present. These soils were from Hamilton, Tamworth, Terang and Noorat, each at 80% θ10kPa. This study was not aimed at identifying thresholds for denitrification, although it would seem that acetylene at 10 kPa in laboratory incubations is useful for that purpose. Indeed, comparing the response of processes to water content across different soils is difficult because different metrics can be used, and water content is usually a proxy for several factors that influence process rates. With respect to N2O emissions and soil water metrics, some progress has been made since Farquharson and Baldock (2008) highlighted this issue, with metrics such as volumetric water content and relative diffusivity providing improvements in explaining N2O emissions (Castellano et al. 2010; van der Weerden et al. 2012; Balaine et al. 2013; Klefoth et al. 2015; Jamali et al. 2016). Ultimately, the modelling of oxygen demand and transport (Cook et al. 2013) is likely to provide the best prospects for improving simulations of N2O dynamics.
Apart from response functions based on water content and temperature, most models do not account for several abiotic interactions. At a basic level, some models invoke, for example, a physical protection of organic matter as a function of clay content, but to our knowledge, no model explicitly accounts for thermal decomposition of hydroxylamine or the catalysis of reactions by metal species, although DNDC purportedly tracks oxidation–reduction reactions over a range of potentials. The Lucaston experiment provided some evidence for a role of abiotic processes in producing N2O. In work yet to be published (T. Lai, M. Denton, R. Farquharson, unpubl. data), we have consistently observed a decline in nitrification rates above a certain temperature (usually ~35°C), whereas N2O emissions continue or decline to a lesser extent. Further investigation is required to unravel responses at high temperatures, considering that surface-soil temperatures >35°C are possible for much of the continent. Indeed, many models are built on data from temperate systems, resulting in poor model functions at the higher temperatures that can occur in Australian soils. There is also debate around thermal acclimation and adaptation, which is particularly pertinent when attempting to predict the influence of climate change on biogeochemical cycling.
Bulk soil pH has long been used to adjust potential nitrification rates in models, based on the observation that autotrophic nitrification rates tend to be lower on acidic soils. Indeed, nitrification and N2O emissions were low from the Wagga Wagga soil with a pHCaCl of 5.8; however, adjusting the pH to 7.5 over 3 weeks did not increase nitrification rates significantly. Like many soil properties, including redox potential, there are significant challenges in studying the influence of pH on microbial processes. Microsite pH where processes are active can be quite different from bulk soil pH and is in fact modified by the very processes we are interested in (Strong et al. 1997). In addition, issues of adaptation and buffering need to be considered over varying lengths of time (Bramley and White 1989). We did observe some differences with higher N2O emissions on higher pH soil from Wagga Wagga, but we also observed large changes in ammonia availability in tubes with acetylene, indicating significant loss or immobilisation of ammonium-N in the acidic soil and significant gain (most likely due to mineralisation of organic N) in the higher pH soil. This can have indirect effects on nitrification (specifically nitrite oxidation) and deserves further attention (Venterea et al. 2015).
Several model assumptions were tested during the course of this work and the outcomes are summarised below:
-
The proportion of nitrified N emitted as N2O (Pn in Fig. 1) is not constant as assumed in many models.
-
The proportion of nitrified N emitted as N2O can vary as a function of water content (as now captured in some models, e.g. NOE; Bessou et al. 2010) but not always.
-
Potential nitrification rates (Kmax in Fig. 1) do vary across sites. Potential nitrification rates should be parameterised using independent evidence because the cyclical nature of nitrogen dynamics makes it extremely difficult to resolve such parameters mathematically.
-
Potential nitrification rates do vary with depth, with the surface soil being more active than deeper soil.
-
Nitrification rates and N2O emissions can increase with ammonium addition, but can also be unresponsive in some soils. Use of ammonium concentration as a multiplicative factor in models (e.g. Fig. 1) may need to be reconsidered, especially where saturation kinetics are not adopted. The sorption of ammonium related to cation exchange capacity (Venterea et al. 2015), pH changes, mineralisation–immobilisation dynamics and other limiting factors may need to be taken into account.
-
The pH functions for nitrification rates in models are likely being misused given microsite issues, the existence of empirically derived potential nitrification rates, which would account for the differences, and the potential for soil microbial communities and functions to adapt.
-
Water functions are not universal. Water functions are problematic because they attempt to account for multiple processes or mechanisms simultaneously (Hall et al. 2013). As a solution, explicit modelling of oxygen transport and consumption (Cook et al. 2013), for example, could be implemented in models.
-
Microsites are prevalent in soils, with hotspots and hot moments potentially contributing significantly to the overall flux of N2O from soil (Parkin 1987; Nielsen and Revsbech 1998; Groffman et al. 2009). Modelling soils in homogenous layers, although common, can be problematic. Hotspots are not a simple function of a single driver, so capturing the interactions and recognising heterogeneity in soil is important (Korsaeth et al. 2001). Averaging is the antithesis of capturing heterogeneity and may result in erroneous predictions, especially in non-linear systems with interacting factors, as is common for biogeochemical cycling.
-
Many models have not been properly tested for responses at higher temperatures. We observed a thermal optimum for nitrification at ~35°C in one temperate soil, but we also note the possibility that thermal acclimation or adaptation can occur (Avrahami and Bohannan 2009; Gödde and Conrad 1999).
-
The proportion of nitrified N emitted as N2O appears to uncouple at high soil temperatures, which may be due to abiotic processes such as the decomposition of hydroxylamine to N2O.
-
Abiotic processes and some abiotic factors may be important and are not yet captured in most models. For example, the decomposition of hydroxylamine may be more rapid at high soil temperatures, which could explain the increase in Pn in the Lucaston experiment. As well, the role of metal ion species in the chemical oxidation of hydroxylamine (Heil et al. 2015) and the role of cation exchange capacity on ammonium sorption and nitrite oxidation (Venterea et al. 2015) require further investigation.
-
Land-use history might be important in governing nitrogen cycling and, in addition to the above factors, is probably linked to changes in soil biota in response to more active nitrogen cycling, and longer term legacy effects related to organic matter dynamics.
In some of the experiments, considerable heterogeneity was observed. Experimental studies aim to reduce heterogeneity; however, when it comes to N2O emissions, the heterogeneity itself might be an important determinant of process rates and hence emissions. The juxtaposition of nitrification and denitrification sites in soil is one example where heterogeneity itself is important. Yet, models and experimental methods either do not explicitly recognise or attempt to minimise heterogeneity, when instead it needs to be accounted for at a range of scales (Turner et al. 2008).
To explore further the production of N2O from nitrification when conditions simultaneously suit denitrification, alternative methods, including the use of stable isotopes, are required. We also suggest that some measurement of redox potential (e.g. Vorenhout et al. 2004) could be informative about which processes are active within a sample (e.g. Hernandez-Ramirez et al. 2009). Although microsite issues would remain (as indeed they do for measurements of pH), routine measurements of redox potential could provide valuable insights into which processes are responsible for the observations made (Husson 2013), as well as inform management practices to minimise emissions of N2O and methane (Yu and Patrick 2003). In the medium term, it is important that standardised techniques are used and additional data collected to enable relationships to be elucidated across a range of soils by using appropriate statistical techniques.
Although observations in field experiments are useful, the resulting information can only be used to inform basic empirical models because the multiple processes and drivers, underlain by soil heterogeneity, are still poorly understood and rarely quantified. Only by understanding such complexity can truly mechanistic models be built. These very models will be crucial to developing robust and tailored mitigation strategies.
Acknowledgements
This project was funded by The Australian Government Department of Agriculture and Water Resources, Filling the Research Gap program, with contributions from the Commonwealth Scientific and Industrial Research Organisation and oversight of the Grains Research and Development Corporation.
Technical assistance for soil processing and laboratory incubations was provided by Thomas Carter and Alana Massalsky. Damian Mowat performed water retention measurements. John Gouzos performed mineral nitrogen analyses. Gas measurements were made at the Central Analytical Facility operated by the Institute for Future Environments (QUT).
Warwick Dougherty, Kulan Thurton and Deidre Harvey provided extensive assistance with the Camden experiment. Ivan Kennedy and Rosalind Deaker from the University of Sydney and Gupta Vadakattu provided acetylene. Helen Suter, Deli Chen, Clemens Scheer and Louise Barton provided useful feedback on methods. Helen Suter also helped coordinate soil sampling and provided site information.
Staff and lead scientists from each of the field sites organised and undertook soil sampling, air-drying and transfer of soil to the Waite laboratory. Particular thanks to Graeme Ward, Kevin Kelly, Wendy Quayle, Hiz Jamali, Rob Harris, Mike Bell, Graeme Schwenke, Guangdi Li, Weijin Wang, Steven Reeves, Lawrence Di Bella, Melissa Royle, Nigel Swarts and their colleagues.
Chris Smith and Henrike Mielenz provided excellent advice on the manuscript and the input of anonymous reviewers further improved the manuscript. The ongoing support and advice of Jeff Baldock, Phil Price, Zaman Nuruzzaman, Peter Grace and the NANORP team are gratefully acknowledged.
References
Ambus P (2005) Relationship between gross nitrogen cycling and nitrous oxide emission in grass–clover pasture. Nutrient Cycling in Agroecosystems 72, 189–199.| Relationship between gross nitrogen cycling and nitrous oxide emission in grass–clover pasture.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2MXhtVKitrfF&md5=9d7b9bee293e692030e13062407864f1CAS |
Avrahami S, Bohannan BJM (2009) N2O emission rates in a California meadow soil are influenced by fertilizer level, soil moisture and the community structure of ammonia-oxidizing bacteria. Global Change Biology 15, 643–655.
| N2O emission rates in a California meadow soil are influenced by fertilizer level, soil moisture and the community structure of ammonia-oxidizing bacteria.Crossref | GoogleScholarGoogle Scholar |
Baggs E (2011) Soil microbial sources of nitrous oxide: recent advances in knowledge, emerging challenges and future direction. Current Opinion in Environmental Sustainability 3, 321–327.
| Soil microbial sources of nitrous oxide: recent advances in knowledge, emerging challenges and future direction.Crossref | GoogleScholarGoogle Scholar |
Balaine N, Clough TJ, Beare MH, Thomas SM, Meenken ED, Ross JG (2013) Changes in relative gas diffusivity explain soil nitrous oxide flux dynamics. Soil Science Society of America Journal 77, 1496–1505.
| Changes in relative gas diffusivity explain soil nitrous oxide flux dynamics.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3sXhsF2lurnM&md5=643a6315d899650a21bb3cb44c494a5cCAS |
Barton L, Kiese R, Gatter D, Butterbach-Bahl K, Buck R, Hinz C, Murphy DV (2008) Nitrous oxide emissions from a cropped soil in a semi-arid climate. Global Change Biology 14, 177–192.
Barton L, Murphy DV, Kiese R, Butterbach-Bahl K (2010) Soil nitrous oxide and methane fluxes are low from a bioenergy crop (canola) grown in a semi-arid climate. Global Change Biology. Bioenergy 2, 1–15.
| Soil nitrous oxide and methane fluxes are low from a bioenergy crop (canola) grown in a semi-arid climate.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3cXms1antb0%3D&md5=d1c7d24d4264220dbb973917781d5336CAS |
Barton L, Butterbach-Bahl K, Kiese R, Murphy DV (2011) Nitrous oxide fluxes from a grain–legume crop (narrow-leafed lupin) grown in a semiarid climate. Global Change Biology 17, 1153–1166.
| Nitrous oxide fluxes from a grain–legume crop (narrow-leafed lupin) grown in a semiarid climate.Crossref | GoogleScholarGoogle Scholar |
Bateman EJ, Baggs EM (2005) Contributions of nitrification and denitrification to N2O emissions from soils at different water-filled pore space. Biology and Fertility of Soils 41, 379–388.
| Contributions of nitrification and denitrification to N2O emissions from soils at different water-filled pore space.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2MXns1Wrs7c%3D&md5=24e8bc55257f9edb302edab19e3f4c83CAS |
Belyaeva ON, Officer SJ, Armstrong RD, Harris RH, Wallace A, Partington DL, Fogarty K, Phelan AJ (2016) Use of the agricultural practice of pasture termination in reducing soil N2O emissions in high-rainfall cropping systems of south-eastern Australia. Soil Research 54, 585–597.
| Use of the agricultural practice of pasture termination in reducing soil N2O emissions in high-rainfall cropping systems of south-eastern Australia.Crossref | GoogleScholarGoogle Scholar |
Bessou C, Mary B, Léonard J, Roussel M, Gréhan E, Gabrielle B (2010) Modelling soil compaction impacts on nitrous oxide emissions in arable fields. European Journal of Soil Science 61, 348–363.
| Modelling soil compaction impacts on nitrous oxide emissions in arable fields.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3cXot1artLg%3D&md5=cc15296f9a02f82b243e0ec9f28f771dCAS |
Bramley RGV, White RE (1989) The effect of pH, liming, moisture and temperature on the activity of nitrifiers in a soil under pasture. Australian Journal of Soil Research 27, 711–724.
| The effect of pH, liming, moisture and temperature on the activity of nitrifiers in a soil under pasture.Crossref | GoogleScholarGoogle Scholar |
Bremner JM, Blackmer AM (1979) Effects of acetylene and soil water content on emission of nitrous oxide from soils. Nature 280, 380–381.
| Effects of acetylene and soil water content on emission of nitrous oxide from soils.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaL3cXksFSntw%3D%3D&md5=ed2e3b6f61deb316e19f1508e76a13daCAS |
Bremner JM, Blackmer AM, Waring SA (1980) Formation of nitrous oxide and dinitrogen by chemical decomposition of hydroxylamine in soils. Soil Biology & Biochemistry 12, 263–269.
| Formation of nitrous oxide and dinitrogen by chemical decomposition of hydroxylamine in soils.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaL3cXltFaqsrg%3D&md5=742060bd0f6515be845634bc5c9cab26CAS |
Butterbach-Bahl K, Baggs EM, Dannenmann M, Kiese R, Zechmeister-Boltenstern S (2013) Nitrous oxide emissions from soils: how well do we understand the processes and their controls? Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences 368, 20130122
| Nitrous oxide emissions from soils: how well do we understand the processes and their controls?Crossref | GoogleScholarGoogle Scholar | 23713120PubMed |
Carter MS (2007) Contribution of nitrification and denitrification to N2O emissions from urine patches. Soil Biology & Biochemistry 39, 2091–2102.
| Contribution of nitrification and denitrification to N2O emissions from urine patches.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2sXls1ajs7s%3D&md5=34ef8f8e8bcb1d58b4775bdf616ca3a0CAS |
Castellano MJ, Schmidt JP, Kaye JP, Walker C, Graham CB, Lin H, Dell CJ (2010) Hydrological and biogeochemical controls on the timing and magnitude of nitrous oxide flux across an agricultural landscape. Global Change Biology 16, 2711–2720.
| Hydrological and biogeochemical controls on the timing and magnitude of nitrous oxide flux across an agricultural landscape.Crossref | GoogleScholarGoogle Scholar |
Chen D, Suter HC, Islam A, Edis R (2010) Influence of nitrification inhibitors on nitrification and nitrous oxide (N2O) emission from a clay loam soil fertilized with urea. Soil Biology & Biochemistry 42, 660–664.
| Influence of nitrification inhibitors on nitrification and nitrous oxide (N2O) emission from a clay loam soil fertilized with urea.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3cXis1Sgtrw%3D&md5=cb0fbf1d12401ca061462b81b933ca1fCAS |
Cheng W, Tsuruta H, Chen G, Yagi K (2004) N2O and NO production in various Chinese agricultural soils by nitrification. Soil Biology & Biochemistry 36, 953–963.
| N2O and NO production in various Chinese agricultural soils by nitrification.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2cXktVemsbY%3D&md5=cb88cb4e5f419a9569c5be75fc943e42CAS |
Cook FJ, Knight JH, Kelliher FM (2013) Modelling oxygen transport in soil with plant root and microbial oxygen consumption: depth of oxygen penetration. Soil Research 51, 539–553.
| Modelling oxygen transport in soil with plant root and microbial oxygen consumption: depth of oxygen penetration.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3sXhslyqtLbM&md5=b67a1bb89b74a12afc1c184548b7b789CAS |
Davidson EA, Swank WT, Perry TO (1986) Distinguishing between nitrification and denitrification as sources of gaseous nitrogen production in soil. Applied and Environmental Microbiology 52, 1280–1286.
Farquharson R, Baldock J (2008) Concepts in modelling N2O emissions from land use. Plant and Soil 309, 147–167.
| Concepts in modelling N2O emissions from land use.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD1cXosVyhs7w%3D&md5=b5d35e5e7b068d830cdd8d26705d8cc2CAS |
Firestone MK, Davidson EA (1989) Microbiological basis of NO and N2O production and consumption in soil. In ‘Exchange of trace gases between terrestrial ecosystems and the atmosphere’. (Eds MO Andreae, DS Schimel) pp. 7–21. (John Wiley & Sons Ltd: Chichester, New York)
Frame CH, Casciotti KL (2010) Biogeochemical controls and isotopic signatures of nitrous oxide production by a marine ammonia-oxidizing bacterium. Biogeosciences 7, 2695–2709.
| Biogeochemical controls and isotopic signatures of nitrous oxide production by a marine ammonia-oxidizing bacterium.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3cXhsFCns7vP&md5=5707cb2db4ad92f25881910d7c8be1e3CAS |
Freney JR, Denmead OT, Simpson JR (1979) Nitrous oxide emission from soils at low moisture contents. Soil Biology & Biochemistry 11, 167–173.
| Nitrous oxide emission from soils at low moisture contents.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaE1MXktVCns7g%3D&md5=5f69bf81335a48a7a6d3893f7b8baafeCAS |
Galbally IE, Meyer MCP, Wang YP, Smith CJ, Weeks IA (2010) Nitrous oxide emissions from a legume pasture and the influences of liming and urine addition. Agriculture, Ecosystems & Environment 136, 262–272.
| Nitrous oxide emissions from a legume pasture and the influences of liming and urine addition.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3cXisFentbk%3D&md5=70afceda0fbcf84514ae46810f7cc755CAS |
Garrido F, Hénault C, Gaillard H, Pérez S, Germon JC (2002) N2O and NO emissions by agricultural soils with low hydraulic potentials. Soil Biology & Biochemistry 34, 559–575.
| N2O and NO emissions by agricultural soils with low hydraulic potentials.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD38XivVahsrY%3D&md5=90fdfcd3eb5be0af1315066127659da8CAS |
Gödde M, Conrad R (1999) Immediate and adaptational temperature effects on nitric oxide production and nitrous oxide release from nitrification and denitrification in two soils. Biology and Fertility of Soils 30, 33–40.
| Immediate and adaptational temperature effects on nitric oxide production and nitrous oxide release from nitrification and denitrification in two soils.Crossref | GoogleScholarGoogle Scholar |
Goodroad LL, Keeney DR (1984) Nitrous oxide production in aerobic soils under varying pH, temperature and water content. Soil Biology & Biochemistry 16, 39–43.
| Nitrous oxide production in aerobic soils under varying pH, temperature and water content.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaL2cXksFeqs7s%3D&md5=ee160bd226cad2bbf91e2b637c7242ffCAS |
Goreau TJ, Kaplan WA, Wofsy SC, McElroy MB, Valois FW, Watson SW (1980) Production of NO2 – and N2O by nitrifying bacteria at reduced concentrations of oxygen. Applied and Environmental Microbiology 40, 526–532.
Grant RF (1995) Mathematical modelling of nitrous oxide evolution during nitrification. Soil Biology & Biochemistry 27, 1117–1125.
| Mathematical modelling of nitrous oxide evolution during nitrification.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaK2MXntlGqsL0%3D&md5=a26507f036e6cb5a2089f916309b1fd1CAS |
Groffman PM, Butterbach-Bahl K, Fulweiler RW, Gold AJ, Morse JL, Stander EK, Tague C, Tonitto C, Vidon P (2009) Challenges to incorporating spatially and temporally explicit phenomena (hotspots and hot moments) in denitrification models. Biogeochemistry 93, 49–77.
| Challenges to incorporating spatially and temporally explicit phenomena (hotspots and hot moments) in denitrification models.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD1MXivVSjs70%3D&md5=000f8bb77d84d8ed2dda6396f122a9b5CAS |
Hall S, McDowell W, Silver W (2013) When wet gets wetter: decoupling of moisture, redox biogeochemistry, and greenhouse gas fluxes in a humid tropical forest soil. Ecosystems 16, 576–589.
| When wet gets wetter: decoupling of moisture, redox biogeochemistry, and greenhouse gas fluxes in a humid tropical forest soil.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3sXnslemu7k%3D&md5=77edb54c0cf48f8dfc85c8ca5aa02c83CAS |
Heil J, Liu SR, Vereecken H, Brüggemann N (2015) Abiotic nitrous oxide production from hydroxylamine in soils and their dependence on soil properties. Soil Biology & Biochemistry 84, 107–115.
| Abiotic nitrous oxide production from hydroxylamine in soils and their dependence on soil properties.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC2MXjslynsLw%3D&md5=d3a2b3849e51c209d9113aef5da0fa23CAS |
Hénault C, Bizouarn F, Laville P, Gabrielle B, Nicoullaud B, Germon JC, Cellier P (2005) Predicting in situ soil N2O emission using NOE algorithm and soil database. Global Change Biology 11, 115–127.
| Predicting in situ soil N2O emission using NOE algorithm and soil database.Crossref | GoogleScholarGoogle Scholar |
Hernandez-Ramirez G, Brouder SM, Smith DR, Van Scoyoc GE, Michalski G (2009) Nitrous oxide production in an Eastern corn belt soil: sources and redox range. Soil Science Society of America Journal 73, 1182–1191.
| Nitrous oxide production in an Eastern corn belt soil: sources and redox range.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD1MXos1Ggs7g%3D&md5=8a37e5299232352a18d3897231bbd861CAS |
Hooper AB, Terry KR (1979) Hydroxylamine oxidoreductase of Nitrosomonas production of nitric-oxide from hydroxylamine. Biochimica et Biophysica Acta 571, 12–20.
| Hydroxylamine oxidoreductase of Nitrosomonas production of nitric-oxide from hydroxylamine.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaL3cXjs1Kkuw%3D%3D&md5=d7a776719cf72016d6aa36fe3b213b75CAS | 497235PubMed |
Hudson F (2004) ‘Sample preparation and calculation for dissolved gas analysis in water samples using a GC headspace equilibration technique.’ (US Environmental Protection Agency: Washington, DC)
Husson O (2013) Redox potential (Eh) and pH as drivers of soil/plant/microorganism systems: a transdisciplinary overview pointing to integrative opportunities for agronomy. Plant and Soil 362, 389–417.
| Redox potential (Eh) and pH as drivers of soil/plant/microorganism systems: a transdisciplinary overview pointing to integrative opportunities for agronomy.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC38XhvV2ks7bM&md5=ae9dc09091dd9ca22ba08e691e78734dCAS |
Hynes RK, Knowles R (1978) Inhibition by acetylene of ammonia oxidation in Nitrosomonas europaea. FEMS Microbiology Letters 4, 319–321.
| Inhibition by acetylene of ammonia oxidation in Nitrosomonas europaea.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaE1MXhtFGkurs%3D&md5=8702f6d553d6007ff7f72205b6d47cfbCAS |
Hynes RK, Knowles R (1984) Production of nitrous oxide by Nitrosomonas europaea: effects of acetylene, pH, and oxygen. Canadian Journal of Microbiology 30, 1397–1404.
| Production of nitrous oxide by Nitrosomonas europaea: effects of acetylene, pH, and oxygen.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaL2cXmt1Kmu7w%3D&md5=c2f59d6be0f8b532e15109f65840775aCAS |
Ingwersen J, Butterbach-Bahl K, Gasche R, Richter O, Papen H (1999) Barometric process separation: new method for quantifying nitrification, denitrification, and nitrous oxide sources in soils. Soil Science Society of America Journal 63, 117–128.
| Barometric process separation: new method for quantifying nitrification, denitrification, and nitrous oxide sources in soils.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaK1MXitFGgsbg%3D&md5=c779dbd466bb3f95a9f903d1f3391896CAS |
Jamali H, Quayle W, Scheer C, Rowlings D, Baldock J (2016) Effect of soil texture and wheat plants on N2O fluxes: A lysimeter study. Agricultural and Forest Meteorology 223, 17–29.
| Effect of soil texture and wheat plants on N2O fluxes: A lysimeter study.Crossref | GoogleScholarGoogle Scholar |
Jia Z, Conrad R (2009) Bacteria rather than Archaea dominate microbial ammonia oxidation in an agricultural soil. Environmental Microbiology 11, 1658–1671.
| Bacteria rather than Archaea dominate microbial ammonia oxidation in an agricultural soil.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD1MXptlyhurs%3D&md5=ae7cac4b4a2c87fb94d48a6a5491ebf4CAS | 19236445PubMed |
Jiang Q, Bakken LR (1999) Nitrous oxide production and methane oxidation by different ammonia-oxidizing bacteria. Applied and Environmental Microbiology 65, 2679–2684.
Jørgensen KS, Jensen HB, Sørensen J (1984) Nitrous oxide production from nitrification and denitrification in marine sediment at low oxygen concentrations. Canadian Journal of Microbiology 30, 1073–1078.
| Nitrous oxide production from nitrification and denitrification in marine sediment at low oxygen concentrations.Crossref | GoogleScholarGoogle Scholar |
Kester RA, De Boer W, Laanbroek HJ (1997) Production of NO and N2O by pure cultures of nitrifying and denitrifying bacteria during changes in aeration. Applied and Environmental Microbiology 63, 3872–3877.
Khalil K, Mary B, Renault P (2004) Nitrous oxide production by nitrification and denitrification in soil aggregates as affected by O2 concentration. Soil Biology & Biochemistry 36, 687–699.
| Nitrous oxide production by nitrification and denitrification in soil aggregates as affected by O2 concentration.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2cXisFOntrc%3D&md5=59ed8388503d18a5a1e48c5e017d362dCAS |
Klefoth RR, Clough TJ, Oenema O, Van Groenigen JW (2015) Soil bulk density and moisture content influence relative gas diffusivity and the reduction of nitrogen-15 nitrous oxide. Vadose Zone Journal 13,
Klemedtsson L, Svensson BH, Rosswall T (1988) A method of selective inhibition to distinguish between nitrification and denitrification as sources of nitrous oxide in soil. Biology and Fertility of Soils 6, 112–119.
Korsaeth A, Molstad L, Bakken LR (2001) Modelling the competition for nitrogen between plants and microflora as a function of soil heterogeneity. Soil Biology & Biochemistry 33, 215–226.
| Modelling the competition for nitrogen between plants and microflora as a function of soil heterogeneity.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD3MXht1Kmtr0%3D&md5=e52a2058dc0808ab03061e719f9fca8aCAS |
Li C, Frolking S, Frolking TA (1992) A model of nitrous oxide evolution from soil driven by rainfall events: 1. Model structure and sensitivity. Journal of Geophysical Research 97, 9759–9776.
| A model of nitrous oxide evolution from soil driven by rainfall events: 1. Model structure and sensitivity.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaK3sXkt1GktQ%3D%3D&md5=1980a7d8e0775da0d200c553730af6d9CAS |
Li Y, White R, Chen D, Zhang J, Li B, Zhang Y, Huang Y, Edis R (2007) A spatially referenced water and nitrogen management model (WNMM) for (irrigated) intensive cropping systems in the North China Plain. Ecological Modelling 203, 395–423.
| A spatially referenced water and nitrogen management model (WNMM) for (irrigated) intensive cropping systems in the North China Plain.Crossref | GoogleScholarGoogle Scholar |
Lipschultz F, Zafiriou OC, Wofsy SC, McElroy MB, Valois FW, Watson SW (1981) Production of NO and N2O by soil nitrifying bacteria. Nature 294, 641–643.
| Production of NO and N2O by soil nitrifying bacteria.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaL38XhsVKmsbY%3D&md5=b1a4906e5cc3667b9a9c651b38e8bed0CAS |
Maag M, Vinther FP (1996) Nitrous oxide emission by nitrifiation and denitrification in different soil types and at different soil moisture contents and temperatures. Applied Soil Ecology 4, 5–14.
| Nitrous oxide emission by nitrifiation and denitrification in different soil types and at different soil moisture contents and temperatures.Crossref | GoogleScholarGoogle Scholar |
Martikainen PJ (1985) Nitrous oxide emission associated with autotrophic ammonium oxidation in acid coniferous forest soil. Applied and Environmental Microbiology 50, 1519–1525.
Martikainen PJ, de Boer W (1993) Nitrous oxide production and nitrification in acidic soil from a dutch coniferous forest. Soil Biology & Biochemistry 25, 343–347.
| Nitrous oxide production and nitrification in acidic soil from a dutch coniferous forest.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaK3sXitlaqs7s%3D&md5=3a7032c960326cda502003f57a96861eCAS |
Mathieu O, Hénault C, Leveque J, Baujard E, Milloux MJ, Andreux F (2006) Quantifying the contributions of nitrification and denitrification to the nitrous oxide flux using 15N tracers. Environmental Pollution 144, 933–940.
| Quantifying the contributions of nitrification and denitrification to the nitrous oxide flux using 15N tracers.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD28XpslSlsbc%3D&md5=79ca4c57b20b2f40c745a4a04fdf1e48CAS | 16569469PubMed |
Mørkved PT, Dörsch P, Henriksen TM, Bakken LR (2006) N2O emissions and product ratios of nitrification and denitrification as affected by freezing and thawing. Soil Biology & Biochemistry 38, 3411–3420.
| N2O emissions and product ratios of nitrification and denitrification as affected by freezing and thawing.Crossref | GoogleScholarGoogle Scholar |
Mørkved PT, Dörsch P, Bakken LR (2007) The N2O product ratio of nitrification and its dependence on long-term changes in soil pH. Soil Biology & Biochemistry 39, 2048–2057.
| The N2O product ratio of nitrification and its dependence on long-term changes in soil pH.Crossref | GoogleScholarGoogle Scholar |
Nielsen TH, Revsbech NP (1998) Nitrification, denitrification, and N-liberation associated with two types of organic hotspots in soil. Soil Biology & Biochemistry 30, 611–619.
| Nitrification, denitrification, and N-liberation associated with two types of organic hotspots in soil.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaK1cXjtl2jsrc%3D&md5=4a505d71bf2aa1215b045e254ec35dd1CAS |
Parkin TB (1987) Soil microsites as a source of denitrification variability. Soil Science Society of America Journal 51, 1194–1199.
| Soil microsites as a source of denitrification variability.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaL2sXmtlGnsbo%3D&md5=61775361fb9516a9522105fa32e3094bCAS |
Parton WJ, Hartman M, Ojima D, Schimel D (1998) DAYCENT and its land surface submodel: description and testing. Global and Planetary Change 19, 35–48.
| DAYCENT and its land surface submodel: description and testing.Crossref | GoogleScholarGoogle Scholar |
Poth M, Focht DD (1985) 15N kinetic analysis of N2O production by Nitrosomonas europaea: an examination of nitrifier denitrification. Applied and Environmental Microbiology 49, 1134–1141.
Rayment GE, Lyons DJ (2011) ‘Soil chemical methods: Australasia.’ (CSIRO Publishing: Melbourne)
Remde A, Conrad R (1990) Production of nitric oxide in Nitrosomonas europaea by reduction of nitrite. Archives of Microbiology 154, 187–191.
| Production of nitric oxide in Nitrosomonas europaea by reduction of nitrite.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaK3cXlsVylt7s%3D&md5=44e4696ebcd351c2b064bbff0b896e99CAS |
Shaw LJ, Nicol GW, Smith Z, Fear J, Prosser JI, Baggs EM (2006) Nitrosospira spp. can produce nitrous oxide via a nitrifier denitrification pathway. Environmental Microbiology 8, 214–222.
| Nitrosospira spp. can produce nitrous oxide via a nitrifier denitrification pathway.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD28XisFWrsrw%3D&md5=477dcb88e5abbb13bf0e4ac04fb3544bCAS | 16423010PubMed |
Strong DT, Sale PWG, Helyar KR (1997) Initial soil pH affects the pH at which nitrification ceases due to self-induced acidification of microbial microsites. Australian Journal of Soil Research 35, 565–570.
| Initial soil pH affects the pH at which nitrification ceases due to self-induced acidification of microbial microsites.Crossref | GoogleScholarGoogle Scholar |
Thorburn PJ, Biggs JS, Collins K, Probert ME (2010) Using the APSIM model to estimate nitrous oxide emissions from diverse Australian sugarcane production systems. Agriculture, Ecosystems & Environment 136, 343–350.
| Using the APSIM model to estimate nitrous oxide emissions from diverse Australian sugarcane production systems.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3cXisFenurg%3D&md5=94287091d4343e47b5666b535f246724CAS |
Tortoso AC, Hutchinson GL (1990) Contributions of autotrophic and heterotrophic nitrifiers to soil NO and N2O emissions. Applied and Environmental Microbiology 56, 1799–1805.
Turner DA, Chen D, Galbally IE, Leuning R, Edis RB, Li Y, Kelly K, Phillips F (2008) Spatial variability of nitrous oxide emissions from an Australian irrigated dairy pasture. Plant and Soil 309, 77–88.
| Spatial variability of nitrous oxide emissions from an Australian irrigated dairy pasture.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD1cXosVyhsrs%3D&md5=dcf89a214c05b543b47afd88ab85abb8CAS |
van der Weerden TJ, Kelliher FM, de Klein CAM (2012) Influence of pore size distribution and soil water content on nitrous oxide emissions. Soil Research 50, 125–135.
Venterea RT, Clough TJ, Coulter JA, Breuillin-Sessoms F (2015) Ammonium sorption and ammonia inhibition of nitrite-oxidizing bacteria explain contrasting soil N2O production. Scientific Reports 5, 12153
| Ammonium sorption and ammonia inhibition of nitrite-oxidizing bacteria explain contrasting soil N2O production.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC2MXhtlCjsbvN&md5=00ee5db3b02de9fa1bfc5431b867a273CAS | 26179972PubMed |
Vorenhout M, van der Geest HG, van Marum D, Wattel K, Eijsackers HJP (2004) Automated and continuous redox potential measurements in soil. Journal of Environmental Quality 33, 1562–1567.
| Automated and continuous redox potential measurements in soil.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2cXmtFKhtLc%3D&md5=c50a428fbf3d8eab0f7e1818df13417eCAS | 15254139PubMed |
Walter HM, Keeney DR, Fillery IR (1979) Inhibition of nitrification by acetylene. Soil Science Society of America Journal 43, 195–196.
| Inhibition of nitrification by acetylene.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaE1MXitVaksrw%3D&md5=168011623024104a0a08c9ee894f9663CAS |
Yoshida T, Alexander M (1970) Nitrous Oxide Formation by Nitrosomonas Europaea and Heterotrophie Microorganisms. Soil Science Society of America Journal 34, 880–882.
| Nitrous Oxide Formation by Nitrosomonas Europaea and Heterotrophie Microorganisms.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaE3MXitVKntw%3D%3D&md5=93223ca75c42597dac6b6cde7365798bCAS |
Yoshinari T, Knowles R (1976) Acetylene inhibition of nitrous oxide reduction by denitrifying bacteria. Biochemical and Biophysical Research Communications 69, 705–710.
| Acetylene inhibition of nitrous oxide reduction by denitrifying bacteria.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaE28XhslKnurs%3D&md5=4b1ef1238dcb3972cd1dbca974aee2eeCAS | 817722PubMed |
Yu K, Patrick WH (2003) Redox range with minimum nitrous oxide and methane production in a rice soil under different pH. Soil Science Society of America Journal 67, 1952–1958.
| Redox range with minimum nitrous oxide and methane production in a rice soil under different pH.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD3sXovFCkt7o%3D&md5=4186a72461c97b67e2a43f9932cdd044CAS |
Yu R, Kampschreur MJ, van Loosdrecht MCM, Chandran K (2010) Mechanisms and specific directionality of autotrophic nitrous oxide and nitric oxide generation during transient anoxia. Environmental Science & Technology 44, 1313–1319.
| Mechanisms and specific directionality of autotrophic nitrous oxide and nitric oxide generation during transient anoxia.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3cXhtFamsLk%3D&md5=3b680a24bc2dd70b4b456d91bfa4dbc1CAS |
Zhu X, Burger M, Doane TA, Horwath WR (2013a) Ammonia oxidation pathways and nitrifier denitrification are significant sources of N2O and NO under low oxygen availability. Proceedings of the National Academy of Sciences of the United States of America 110, 6328–6333.
| Ammonia oxidation pathways and nitrifier denitrification are significant sources of N2O and NO under low oxygen availability.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3sXnvVOltbk%3D&md5=e6595e76a6f8266c666327a9d5dbe70cCAS | 23576736PubMed |
Zhu X, Silva LCR, Doane TA, Horwath WR (2013b) Iron: the forgotten driver of nitrous oxide production in agricultural soil. PLoS One 8, e60146.
| Iron: the forgotten driver of nitrous oxide production in agricultural soil.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3sXmtVemtb8%3D&md5=a203e23872b272fbe9db928cee92351aCAS | 23555906PubMed |