

Relationships between annual plant productivity, nitrogen deposition and fire size in low-elevation California desert scrub
Leela E. Rao A E , John R. Matchett B , Matthew L. Brooks B , Robert F. Johnson A , Richard A. Minnich C and Edith B. Allen A DA University of California, Center for Conservation Biology, 900 University Avenue, Riverside, CA 92521, USA.
B US Geological Survey, Western Ecological Research Center, Yosemite Field Station, 40298 Junction Drive – Suite A, Oakhurst CA 93644, USA.
C University of California, Department of Earth Sciences, 900 University Avenue, Riverside, CA 92521, USA.
D University of California, Department of Botany and Plant Sciences, 900 University Avenue, Riverside, CA 92521, USA.
E Corresponding author. Email: lrao@arb.ca.gov
International Journal of Wildland Fire 24(1) 48-58 https://doi.org/10.1071/WF13152
Submitted: 21 December 2012 Accepted: 23 July 2014 Published: 10 November 2014
Journal Compilation © IAWF 2015
Abstract
Although precipitation is correlated with fire size in desert ecosystems and is typically used as an indirect surrogate for fine fuel load, a direct link between fine fuel biomass and fire size has not been established. In addition, nitrogen (N) deposition can affect fire risk through its fertilisation effect on fine fuel production. In this study, we examine the relationships between fire size and precipitation, N deposition and biomass with emphasis on identifying biomass and N deposition thresholds associated with fire spreading across the landscape. We used a 28-year fire record of 582 burns from low-elevation desert scrub to evaluate the relationship of precipitation, N deposition and biomass with the distribution of fire sizes using quantile regression. We found that models using annual biomass have similar predictive ability to those using precipitation and N deposition at the lower to intermediate portions of the fire size distribution. No distinct biomass threshold was found, although within the 99th percentile of the distribution fire size increased with greater than 125 g m–2 of winter fine fuel production. The study did not produce an N deposition threshold, but did validate the value of 125 g m–2 of fine fuel for spread of fires.
Additional keywords: biomass, fine fuel, Mojave, Sonoran.
Introduction
The frequency and size of fires in desert regions of California is positively related to precipitation (Brooks and Matchett 2006). This relationship is especially strong at lower elevations where sparse vegetation is typically insufficient to carry fire except following periods of high precipitation, when annual plants fill the spaces between perennial plants creating continuous fuelbeds that promote fire spread (Brooks 1999; Brooks and Minnich 2006). Productivity of non-native annual grasses can be especially prolific in response to high precipitation (Gutiérrez 1992; Rao and Allen 2010). Their biomass tends to persist longer as standing fuel than does that of annual forbs that tend to disarticulate and disperse at a faster rate (Brooks 1999). As a result, non-native annual grasses are often considered to be transformers of desert ecosystems through their effects on fuelbeds, fire regimes and other ecosystem properties (D'Antonio and Vitousek 1992; Brooks et al. 2004; Brooks and Minnich 2006; Steers and Allen 2011; Balch et al. 2013).
Although it is clear that non-native annual grasses are associated with increased fire risk, it remains unknown how much biomass is required to allow fire to spread. This has not been evaluated largely because identification of annual plant fuel load thresholds necessary for fire spread in deserts requires data on biomass that is seldom known before a fire burns. Precipitation is used as a surrogate for fine fuel biomass in models that correlate fire with winter precipitation (e.g. Brooks and Matchett 2006). Currently, biomass thresholds can only be estimated from general fuel models typically developed for non-desert ecosystems (e.g. Anderson 1982). For instance, management burns are not recommended unless there is at least 100 g m–2 of fine fuel in grasslands, but threshold values are not quantified for wildfires. Direct quantification of thresholds is necessary to predict when and where fires may be most likely to occur and to develop fire hazard assessments for land managers.
Because nitrogen (N) deposition also contributes to fuel productivity (Rao and Allen 2010; Rao et al. 2010) there is a need to evaluate how much N deposition from atmospheric pollution contributes to fire size above and beyond the influence of precipitation. This is important because of recent and projected future increases of N deposition in the California deserts (Fenn et al. 2010; Pardo et al. 2011) and the potential need to manage point and non-point sources of N pollution from a fire management standpoint. N deposition levels in California deserts can be as high as 16 kg N ha–1 year–1 downwind of populous coastal cities (Fenn et al. 2010). Most occurs as dry deposition in oxidised form from combustion, although reduced N forms are also deposited in the desert (Fenn et al. 2003a; Fenn et al. 2003b). Low background levels of soil N in deserts limit plant growth even with increased precipitation (Rao and Allen 2010), and when amounts of N are increased non-native species often utilise it more readily than native species (Brooks 2003; Allen et al. 2009). In contrast to precipitation, the link between N deposition and fire size is not well established (but see Rao and Allen 2010; Rao et al. 2010). The threshold, or critical load, for N deposition that promotes sufficient fine fuel to increase fire risk based on standard fuel models was modelled as 3 kg N ha–1 year–1, with an upper stabilisation level of 9 kg N ha–1 year–1, at which precipitation becomes the main control (Rao et al. 2010). A more ecoregion-specific empirical threshold could be determined for the California desert by examining biomass values co-occurring with historic fire datasets in relation to N deposition and precipitation.
Given the need to determine thresholds of N deposition and biomass production for fire management purposes, the primary objectives of this study are to:
-
Compare the ability of N deposition and precipitation with that of herbaceous fine fuel biomass (modelled) to determine fire size.
-
Determine the relationship between herbaceous fuel biomass and fire size, focusing on the identification of a biomass threshold.
-
Describe the range of N deposition and precipitation values necessary to produce the biomass threshold.
These objectives were met by examining a 28-year fire record for low-elevation desert scrub across California and coupling the fire record with modelled precipitation, N deposition and biomass information.
Methods
Study area
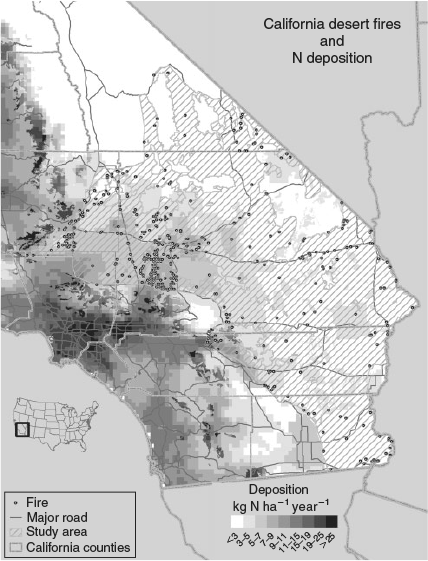
The study area was 6 209 206 ha in size (Fig. 1), and included portions of the Mojave and Colorado Deserts within California at elevations less than 1000 m. We limited the analysis to desert wash, desert scrub and desert succulent scrub California Gap Analysis Program landcover types (Davis et al. 1998). These lower-elevation vegetation types contain native perennial fuels that are too sparse to carry fire without non-native annual plants filling the interspaces between them and have historically been subject to lower fire frequency (Brooks and Matchett 2006; Balch et al. 2013). Because the exact fire perimeters were unknown, we used the point of origin of each fire to determine if it occurred below 1000 m. We excluded fires with a point of origin in riparian, agricultural or other developed areas, which were especially prevalent along the south-west and south-east margins of the study region.
|
Data sources
Fire data
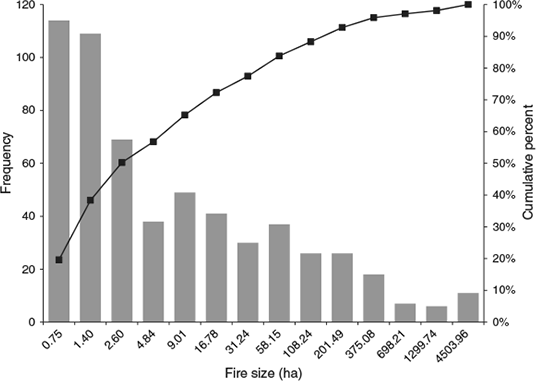
Fire data were obtained from the US Geological Survey Wildfire database (US DOI 2009). The database included the federal fire history reports from 1980 to 2008. Data were processed to remove duplicate fires, fires with invalid dates and fires with a point of origin outside the study area. We selected only fires greater than 0.4 ha in size because smaller fire areas represent events that did not result in fire spread (e.g. roadside automobile fires). Because the primary intent of this study was to examine the relationship between biomass growth and fire size, we only used fires that occurred during the summer fire season, defined as 1 May–31 October. Winter fires were excluded because of uncertainty regarding when the biomass fuelling the fire was generated, which resulted in uncertainty regarding whether summer precipitation, prior year winter precipitation or current year winter precipitation should be used to model fine fuel biomass. Of the 548 543 national records, 582 were used for this study with temporal period of May 1980–September 2008 from fires located on both private and public lands. Cumulative and frequency distributions of fire size are presented in Fig. 2.
|
The fire data were point based with a total area burned associated with each point. We converted each fire data point into a circle with the same area as the reported size, with the centre of the circle being the point of origin of the fire. No attempt was made to verify the accuracy of the locations of the fires in the fire database, but the database in this study is generally the same as the database used in previous analyses (Brooks and Matchett 2006) with the addition of data from 2005 to 2008. Any inaccuracies in the fire database would affect the results to a greater or lesser degree depending on fire size and the homogeneity or heterogeneity of land cover within each quarter section-based location, with possible accuracy problems having a greater effect for smaller fires and less so for larger fires. For each fire we calculated the distance to the nearest road from the point of origin, area-weighted precipitation in the year of the fire and the year preceding the fire, and area-weighted total annual N deposition within the fire area during 2002.
Distance to road
The distance to a road has been shown to have significant explanatory power for fire size due to its correlation with human influence, particularly in the urban–wildland interface (Syphard et al. 2007; Holden et al. 2009; Brunelle et al. 2010). Because of the importance of human influence on fire size, we included distance to road (roadDist) in our models as a controlling variable to enable us to detect the potentially smaller effects of precipitation and N deposition. We defined roadDist as the distance from the point of fire origin, in metres, to the nearest major road (annual average daily traffic >8000 vehicles) or national highway (RITA 2012).
Precipitation
We utilised PRISM weather data at a 4-km grid resolution to calculate the area-weighted seasonal precipitation for each fire (www.prism.oregonstate.edu, verified 10 August 2010). The winter season was defined as 1 November–30 April, and summer as 1 May–31 October. Four precipitation variables were developed: precipitation during the first full winter before the fire (pptWin1), during the second full winter before the fire (pptWin2), during the first full summer before the fire (pptSmr1) and during the second full summer before the fire (pptSmr2). Because the fires were limited to those occurring during the summer season, pptSmr1 is actually the summer precipitation that occurred the year before the fire. Summer precipitation from the year of the fire was not included because most rainfall occurs middle to late summer (Hereford et al. 2006) whereas most fires occur in the early to middle summer (Brooks and Esque 2002), before the onset of significant rainfall. We included precipitation from 2 years before the fire in our analysis because this has been correlated with fire frequency in previous studies (Rogers and Vint 1987; Margolis and Balmat 2009).
Nitrogen deposition
The US Environmental Protection Agency’s Community Multiscale Air Quality (CMAQ), a photochemical grid model, was used to model N deposition (Ndep) over the study area. CMAQ uses meteorological and emissions inputs to model the atmospheric photochemistry, transport and deposition of pollutants. Meteorological data were produced by operating the National Center for Atmospheric Research (NCAR)/Penn State Mesoscale Model (MM5) for the year 2002 on a 4-km (2.5-mile) resolution grid. Emissions inventories were derived from previous oxidised and reduced aerosol N, ozone and particulate matter modelling studies for California (Tonnesen et al. 2003) and for the western US (Tonnesen et al. 2002). A full description of the model can be found in Tonnesen et al. (2007), and the dataset and metadata are available at http://ccb.ucr.edu/biocomplexityfiles/data/California%202002.zip (verified 14 August 2013). N deposition values used in our analyses are a mix of 4- and 36-km resolution model results (Fig. 1), with higher-resolution modelling in the Los Angeles airshed (Tonnesen et al. 2007; Fenn et al. 2010). There are 138 points in the low-resolution area, and because this is also the low-deposition area, we believe that the resolution is sufficient for our purposes. N deposition was area-weighted within each of the scaled fire circles to give a mean deposition value for the area burned for each fire event. Although our fire record dates back to 1980, we used the earliest available 2002 CMAQ data to provide a relative ranking of the N deposition that would be observed over our study area. CMAQ N deposition modelled data are correlated with measurements from the National Atmospheric Deposition Program (NADP) (Clark et al. 2013), but this has relatively few measurement sites in the west both at present and historically. Further, dry deposition, which forms most of the deposition in arid climates, is not measured by NADP and must be inferred. To determine precise critical N deposition loads, deposition data from earlier in the fire period would be preferred but were not available.
Biomass
We calculated fine fuel biomass for each fire using pptWin1 and Ndep. A regression to calculate biomass from winter precipitation and N deposition was developed using the DayCent model and 25 years of modelled biomass for four sites in Joshua Tree National Park (JTNP). The DayCent model is a biogeochemical model that predicts annual production based on inputs of climate, soil profile and plant-specific properties (e.g. percentage tissue carbon (C), N and lignin). The calibration and validation of biomass modelled for the four JTNP sites are described in detail in Rao et al. (2010). Briefly, two model parameterisations were developed; one for native winter annuals mixed with the exotic annual grasses Schismus barbatus and S. arabicus, and one for native winter annuals mixed with the exotic annual grass Bromus rubens. These exotic annual grasses occur throughout the study area, depending on precipitation and elevation. The model was parameterised for one site and validated with a second site using 5 years of field data that spanned some of the driest and wettest years on record (2003–2007). The parameterised models were then run for 25 years (1983–2008) to generate predicted biomass based on the precipitation record from four nearby weather stations, two sites with representative Schismus spp. communities and two sites with representative B. rubens communities. The biomass estimates were then combined and a single regression performed to develop a simplified ‘winter annual’ biomass model based on precipitation and N deposition.
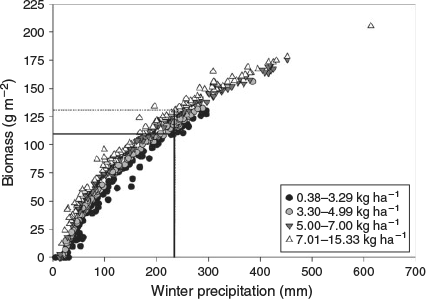
As discussed in Rao et al. (2010), the DayCent model over-predicts biomass production in very dry years when there is insufficient precipitation to result in germination (<2 mm). Therefore, any modelled biomass amounts from years with less than 2 mm of precipitation were set to zero before developing the simple regression model. The resultant model was: biomasspred = 50.9(pptWin1)1/3 + 23.6(Ndep)1/3 – 96.6 (n = 98, R2 = 0.70). The regression model was validated using 28 data points from 5 years of actual biomass data from the four sites at JTNP as well as several other sites outside the Park. Although the validation data were limited to data collected at or near JTNP, the biomass was collected over a period of 5 years that were some of the driest and wettest years on record, with a range of 48 to 440 mm annual precipitation, which effectively brackets the range of precipitation from our study area based on 30-year normal PRISM precipitation data (48–341 mm annual precipitation with a mean of 124 ± 33 mm). In addition, the model was developed at the ecotone between the Mojave and Colorado deserts of California – the same two desert regions that define the study area in our analysis – and encompassed the range of elevations, temperature regimes and seasonality of precipitation that would affect herbaceous plant growth in the study area (Hereford et al. 2006).
The validation results indicated that the model performed well at low biomass (intercept near zero), but at high actual biomass, modelled biomass was under-predicted (biomasspred*1.396–3.840, R2 = 0.54). As a result, a correction factor was applied to the modelled regression to adjust the slope of the model. The final corrected validation model had a slope of one and an intercept of zero. Using the corrected regression equation, annual winter biomass produced 1 year before the fire (bioWin1) was calculated using the modelled 2002 N deposition values for each fire and the winter precipitation 1 year before the fire (pptWin1). Any negative calculated biomass amounts were treated as zero.
Statistical analyses
The distribution of wildfire sizes used in this study – and of wildfires in general – departs strongly from normal, with smaller fires much more frequent than larger ones (Fig. 2). This presents a challenge when using parametric multiple regression models that attempt to quantify how the expected mean changes in response to some set of predictor variables. For example, the mean may be significantly influenced by a small number of very large fires (i.e. outliers), and it is difficult for such models to meet their parametric assumptions, even when severely transforming the response using, for example, a log-scale transformation. Outliers could be trimmed before analysis; however, in our dataset this likely removes from consideration those fires having the greatest ecological and economic effects because they cumulatively burn a much greater area than all smaller fires combined. In order to get a more complete assessment of how fire size responded to our predictor variables of interest, we used a modelling technique called quantile regression to assess the relationships between fire size distribution and the precipitation, N deposition and biomass variables. Quantile regression (Koenker and Bassett 1978) estimates the effects of explanatory variables for different portions of the distribution of a response variable, rather than just modelling the mean response, and has been shown to be a useful technique for analysing a variety of ecological datasets (Cade and Noon 2003), including identifying relationships between wildfire size and climate variables (Slocum et al. 2010). A series of modelling functions is estimated at different levels of τ, with τ representing the fractions of expected response values (e.g. fire size) below the estimated function and 1 – τ, the expected values above it, conditional upon a set of predictor variables (e.g. N deposition, precipitation, biomass). For example, a τ = 0.5 represents the expected median of a distribution, so quantile regression can model how the median changes in response to differing values of the various predictor variables. Functions can be estimated for a range of τ between zero and one, resulting in multiple rates of change (slopes) between an explanatory variable and the response variable, therefore providing a more complete picture of how the distribution of the response is changing given changes in a set of predictor variables. Quantile regression is a semi-parametric modelling technique because no parametric distributional form is assumed for the random error part of the model, whereas a parametric form is assumed for the deterministic part (Cade and Noon 2003). Graphical visualisations of two model variables – fire size and distance to road – suggested that they should be log-transformed before analysis to meet this assumption. For our analyses, we used the ‘quantreg’ package (Koenker 2010) for R statistical software system (R Development Team 2010) to run quantile regression models.
In addition to modelling a constant, linear (at least on a log scale) response of fire sizes to fine fuel biomass at various levels of τ, we were interested in finding evidence of a distinct threshold in biomass – a level of fine fuel biomass below which there is little change in fire size distribution patterns and above which there is a dramatic change. To evaluate this possibility with quantile regression, we ran models having a single ‘knot’ at various levels of winter biomass, specifically from 25 to 150 g m–2 in 25 g m–2 increments. This range in knots brackets the range of fine fuel thresholds necessary for carrying fire reported in the grassland literature (Anderson 1982; Scifres and Hamilton 1993; Fenn et al. 2003a). Because knot models can only be run at discrete levels chosen a priori, we also ran a model with winter biomass as a second-order polynomial to evaluate if a model without a specified threshold would fit the data better than the models with and without the knots.
Numerous unique models can be formed using various combinations of predictor variables in order to test which ones best explain the distribution of fire sizes; however, it is not appropriate to combine certain predictor variables in the same model. For example, winter biomass, winter precipitation and N deposition should not be combined in the same model since winter biomass is simply derived from winter precipitation and N deposition via a predictive model. Likewise, including winter biomass knotted at 25 and 150 g m–2 in the same model would not make sense because the model would no longer be identifying a single threshold. We ultimately specified nine distinct models (Table 1) to address the key objectives of our analysis, primarily to determine (1) if there is any benefit in using fine fuel biomass (modelled from precipitation and N deposition) v. simply using precipitation for predicting fire size, and (2) if there is evidence of a threshold response in fire size to fine fuel biomass. The other variables, such as distance to road and summer precipitation, which may have explanatory power but are not central to our analytical objectives, were included in all models. The nine models in the set were compared to each other using an information theoretic approach, specifically by calculating Akaike’s Information Criteria (AIC) values. In standard multiple regression, each model has a single AIC value, but in quantile regression an AIC value can be calculated for each level of τ for each model. The trends in AIC values across τ levels between models can be used to assess which models are best predicting changes in fire size distribution.

|
For the N deposition threshold analysis, the quantile regression results were used to determine if a biomass threshold occurred from which an N deposition threshold could be derived. We also evaluated the N deposition threshold by examining the relationship between N deposition and area burned. We weighted each burn under a given N deposition level by the total study area under a given N deposition interval because a majority of the study area was under low to moderate levels of N deposition, with only a few regions experiencing high N deposition (Fig. 1). N deposition intervals were analysed every 1 kg N ha–1. The area-weighted burned value was compared to a null hypothesis of equal weighted area burned using a chi-square test.
Results
Model comparison
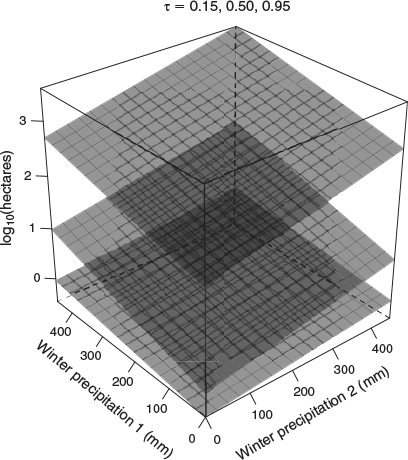
No one model performed best across the entire range of τ values (Table 2). Model 2, which contained modelled herbaceous biomass in place of winter precipitation and N deposition, generally had the lowest AIC values for τ levels at or below 0.6, but most AIC – AICmin values were less than 3, indicating that all the models were essentially equivalent. Model 1, which contained the combination of precipitation and N deposition, was a significantly better predictor of fire size distribution at τ values between 0.70 and 0.90. The coefficients for winter precipitation in the year before the fire and 2 years before the fire for Model 1 had a quadratic response, with the greatest effect of winter precipitation occurring at 0.50 < τ < 0.99 (Fig. 3). The overlapping peaks of the two winter precipitation coefficients indicate a strongly additive effect of 2 years of high winter precipitation on intermediate percentiles of fire size (Fig. 3), although some amount of additiveness affects most of the fire size distribution (Fig. 4).

|

|
|
There does not appear to be an effect of summer precipitation due to the large amount of uncertainty in the summer precipitation coefficients, as indicated by the bootstrapped 95% confidence intervals that bracket the zero line throughout the entire τ range (Fig. 3). There was generally no influence of N deposition on fire size distribution, except for a slight negative relationship at τ = 0.80. There was a positive, gradually increasing effect of distance to road across the fire size distribution, indicating that the infrequent larger fires were the farthest from roads.
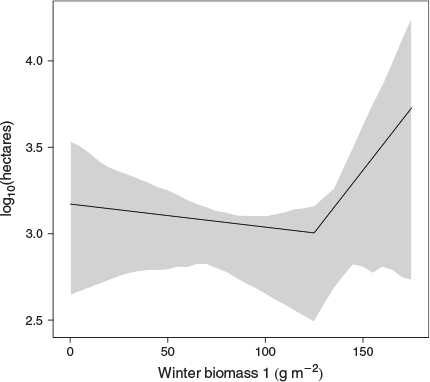
The best predictor of the 99th percentile of the fire size distribution was Model 7, which had a ‘knot’ at 125 g m–2 of fine winter biomass. The shape of the relationship between winter biomass and fire size at τ = 0.99 indicates the effect of winter biomass is level, or decreases slightly up to the knot, after which there is a strong, positive relationship between annual winter biomass and the fire size representing the largest 1% of fires, conditional upon values of the other predictor variables (Fig. 5).
|
N deposition threshold
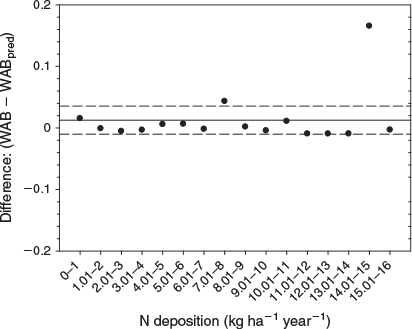
We were not able to discern a fine fuel threshold for the majority of fires in our dataset, and as such could not derive an N deposition threshold for most fires based on biomass. At the 99th percentile of the fire size distribution, there was a biomass threshold of 125 g m–2, although no N deposition threshold at this percentile was apparent. In addition, because precipitation was the predominant driver of biomass (Fig. 6), it was difficult to derive a statistically meaningful N deposition threshold from the biomass threshold. We also evaluated the relationship between N deposition and area burned using a weighted area calculation of area burned. Subtracting the actual weighted area burned at each N deposition interval from the expected weighted area burned assuming equal probability of burning at each interval, gave a mean difference close to zero (Fig. 7; mean = 0.013). A chi-square analysis indicated that overall the actual differences were not significantly different from the expectation of an equal proportion of area burned at each weighted N deposition level (P = 0.45). At both 7 and 14 kg N ha–1 the expected weighted area burned was greater than the 95% confidence interval, but not significantly so.
|
|
Discussion
Fine fuel biomass and fire size distribution
The quantile regression results indicated that under most levels of τ, there was no real difference between the models containing precipitation and biomass and those containing the combination of precipitation and N deposition for the prediction of fire size distribution in our study area (Table 2). This result is likely due to the fact that winter annual biomass is primarily driven by precipitation (Noy-Meir 1973; Beatley 1974), with N deposition having a much smaller effect (Fig. 6; Rao and Allen 2010), especially given the small range of N deposition values across the study area (Fig. 1). Additionally, in most cases the knot models did not perform better than those without a knot, and as such we were unable to identify a fine fuel threshold for the majority (τ < 0.99) of fires. There are several possible reasons as to why biomass was not a stronger predictor of fire size distribution in this study. One likely explanation is that the biomass model was too simplistic. The biomass model used only winter precipitation and N deposition, and was created using data from the DayCent model (Rao et al. 2010). The DayCent model could not be used to estimate biomass for each fire directly because there were insufficient data, namely soil profile information, to run the model for each fire location. There were also not enough field samples of biomass to develop a robust empirical biomass model, which is why we relied on modelled data. More research is needed into the development of robust fine fuel biomass models using field observations.
Other possible explanations for why biomass did not have superior predictive performance compared to precipitation and N deposition are that the dataset was not sufficiently large, or the data sufficiently precise or accurate. Additionally, the spatial and temporal variability may have been too large, making it difficult to tease out the drivers behind variability in fire size. We were only able to include one controlling variable, distance from road, in our analysis. Numerous other variables (such as fuel moisture, relative humidity, wind speed and ground slope) can have an important influence on fire spread (Keeley and Fotheringham 2001; Crimmins and Comrie 2004), and other controlling variables may have helped in reducing the variability and increasing the chance of detecting the effect of biomass. Additional information, particularly fuel moisture content, also would have enabled different types of analyses such as structural equation modelling to be conducted.
Although predicted biomass was not found to be a better determinant of fire size distribution, we did find that winter precipitation strongly influenced the distribution (Fig. 3), with 2 years of high winter precipitation resulting in a substantial increase in the size of the largest fires (Fig. 4). Several studies support the link between increased precipitation and fire size in low-elevation deserts (Brown and Minnich 1986; Rogers and Vint 1987; Crimmins and Comrie 2004; Brooks and Matchett 2006). Several other studies from the arid south-western US have shown a link between a period of drought and increased fire size, possibly with increased precipitation 2 years before the fire, but those patterns are driven by higher-elevation, fire-prone shrubland, woodland and forest landscapes (Swetnam and Betancourt 1998; Westerling et al. 2003; Littell et al. 2009). Drought does not appear to result in a distribution pattern that contains large fires, likely due to the timing of precipitation in the Mojave and Colorado deserts of California, which primarily occurs between September and May. Because dry weather conditions are common during the summer, the limiting factor for fire is usually the amount and connectivity of fine fuels that were generated during the winter growing season, not fuel moisture content (Brooks and Minnich 2006; Krawchuk and Moritz 2011). The cover of non-native grasses relative to that of native forbs is also important since non-native annual grass fuels are more persistent throughout the summer, whereas native forbs disarticulate more quickly (Brooks 1999). Thus, our data support the hypothesis that 2 years of above-average winter precipitation generate significant fine fuel biomass because the first wet winter will result in both higher production of annuals and the addition of large numbers of seeds to the soil, and a second wet winter and increased seed reserves will result in production of sufficient annuals to sustain fire (McLaughlin and Bowers 1982). Additional studies are needed to determine both the fine fuel threshold needed to carry small to intermediate-sized fires in this ecosystem, as well as the conditions necessary to generate sufficient biomass.
Our analysis revealed a fine fuel threshold of 125 g m–2 influencing the fire size at the 99th percentile of the fire size distribution. Before the model knot at 125 g m–2, the fire size of the 99th percentile was relatively flat, but after the knot the fire size increased with increasing biomass production (Fig. 5). This biomass threshold is at the higher end of the range of fine fuel thresholds of 70–150 g m–2 reported as being able to carry fire in grass-dominated systems (Anderson 1982; Scifres and Hamilton 1993; Fenn et al. 2003a). For reasons already discussed, no biomass threshold appeared to influence most of the fire size distribution, although the change in fire size at the 99th percentile of the fire size distribution when fine fuel loads exceeded 125 g m–2 does support the hypothesis that annual biomass production above a certain threshold creates a continuous fine fuel load that facilitates fire spread and is necessary for the occurrence of large fires in a low-elevation desert ecosystem that would otherwise be fuel limited (Brown and Minnich 1986; Brooks and Matchett 2006; Meyn et al. 2007).
One caveat to the biomass threshold estimate is that the biomass data are calculated for peak biomass or the amount of total green biomass (dry mass) at the end of the growing season. Thus, these are overestimates of the biomass present at the time of the fire unless the fire occurred in the late spring, since some of the dry biomass disarticulates during the dry season, especially if forbs rather than grasses are dominant. Additionally, these data do not take into consideration the biomass that may have been generated as a result of summer precipitation in the same year as the fire. Care must also be taken when interpreting the results at the 99th percentile, as there is substantial uncertainty about this estimate, which can be seen in the large 95% confidence intervals in Fig. 5. However, despite these limitations and those of the dataset as previously described, the biomass threshold found for large fires in our dataset falls within the range of thresholds reported previously in the literature for fine fuels, lending credence to the use of a fine fuel threshold of ~100 g m–2 in fire planning and risk assessment in desert scrub ecosystems.
Thresholds of N deposition
Based on the best-performing model for intermediate-sized fires, Model 1, there was no strong effect of N deposition when it was included as a distinct predictor variable (Fig. 3). The fact that larger fires were associated with lower N deposition levels may be related to remoteness, which limits the level and speed of firefighting activities. This hypothesis is supported by the positive relationship between distance to road and fire size (Fig. 3) and a weakly negative correlation between distance to road and N deposition (r = –0.43). Conversely, because areas of high N deposition are generally closer to urban and agricultural areas, increased firefighting capabilities in these areas may have somewhat limited fire spread, thus further limiting our ability to detect an N deposition effect. Additionally, even areas with low N deposition are capable of producing high fine fuel loads when precipitation is sufficient.
Analysis of the area burned under each N deposition level in our study area indicated that under 7 and 14 kg ha–1 year–1 of N deposition more area burned than would be expected, although these results were not statistically significant (Fig. 7). Studies evaluating the critical loads of N deposition in desert scrub have identified 3.2–9.3 kg N ha–1 year–1 as being sufficient to cause changes in the community composition or increase biomass above a 100 g m–2 fine fuel threshold (Fenn et al. in press; Fenn et al. 2010; Rao et al. 2010). Whether a particular area falls within that range is dependent upon both its soils and precipitation regime (Rao et al. 2010).
In addition to the limitations of our dataset previously described, our inability to discern an N threshold for the majority of fires is likely related to the fact that most of the variation in annual biomass productivity is driven by inter-annual precipitation rather than differences in the spatial variation in N deposition (Fig. 6). Although field studies have demonstrated increases in annual biomass with N fertilisation in desert scrub (Gutiérrez 1992; Brooks 2003; Rao and Allen 2010), the reality is that the difference in biomass production between low and high N deposition is a very small range to detect within the linear trend of fire size in our dataset given that precipitation is the dominant forcing factor. However, because N additions can promote annual biomass production overall and non-native invasive grasses in particular (Brooks 2003; DeFalco et al. 2003; Salo et al. 2005; Rao and Allen 2010), it remains important to evaluate N deposition in relation to fire size and fire risk. This is particularly important in areas of known or suspected elevated N deposition given that N deposition could be sufficient to increase the annual biomass produced to above the fire-carrying threshold compared to background N deposition levels (Fig. 6). Additional studies in regions with more widespread increased levels of N deposition may also improve the ability to detect the influence of N deposition on fire size and assist in the development of an N deposition threshold.
Management implications
In the future, the arid regions of southern California will be under increasing pressure from climate change, which is predicted to increase temperatures and the severity of extreme weather events, particularly drought-like La Niña conditions (Beuhler 2003; Weiss and Overpeck 2005; Seager et al. 2007). Because of the link between fine fuel production and fire size in low-elevation desert scrub, this change in climate will affect fire risk. The ability to assess fire risk before the start of the summer fire season can assist not only with budgeting and fire season readiness, but also with protection of threatened and endangered species. Specifically, this region is home to the desert tortoise (Gopherus agassizii (Cooper)), a nationally threatened species that is harmed by wildfire through direct mortality and loss of habitat (Brooks and Esque 2002; Esque et al. 2003).
The results from our study indicate that the fire size distribution in our study area can be predicted using the precipitation received during that winter rainy season and the prior year’s winter precipitation, and that not much additional predictive power is gained by using annual plant biomass modelled from winter precipitation and N deposition. Precipitation data are generally readily available to land managers, with no need to survey biomass. However, the fire size representing the 99th percentile of the fire size distribution did have a relationship with predicted biomass, and there did appear to be a threshold of 125 g m–2, suggesting that, in order to be prepared for the largest, most ecologically and economically destructive fires, it is prudent that land managers sample peak annual biomass. Since assessment of peak biomass is a relatively simple measure and one that is often conducted for other land management purposes, having a fine fuel threshold associated with increased fire risk can assist land managers in deciding where to put limited resources. Although our study did not identify a single N deposition threshold, because of the demonstrated association between winter annual biomass production and N fertilisation (Brooks 2003; Rao and Allen 2010), care should be taken when estimating fire risk in high-deposition areas using precipitation alone, as fire risk will likely be underestimated (Rao et al. 2010).
Acknowledgements
Data analysis and writing of this manuscript were supported with funds from NSF DEB 04–21530, National Park Service PMIS No. 110325, the US Geological Survey Invasive Species Program, the US Geological Survey John Wesley Powell Center for Analysis and Synthesis and the California Air Resources Board. Any use of trade, firm or product names is for descriptive purposes only and does not imply endorsement by the US Government.
References
Allen EB, Rao LE, Steers RJ, Bytnerowicz A, Fenn ME (2009) Impacts of atmospheric nitrogen deposition on vegetation and soils in Joshua Tree National Park. In ‘The Mojave Desert: ecosystem processes and sustainability’. (Eds RH Webb, LF Fenstermaker, JS Heaton, DL Hughson, EV McDonald, DM Miller) pp. 78–100. (University of Nevada Press: Las Vegas)Anderson HE (1982) Aids to determining fuel models for estimating fire behavior. USDA Forest Service, Intermountain Forest and Range Experiment Station, General Technical Report INT-122. (Ogden, UT)
Balch JK, Bradley BA, D’Antonio CM, Gómez-Dans J (2013) Introduced annual grass increases regional fire activity across the arid western USA (1980–2009). Global Change Biology 19, 173–183.
| Introduced annual grass increases regional fire activity across the arid western USA (1980–2009).Crossref | GoogleScholarGoogle Scholar | 23504729PubMed |
Beatley JC (1974) Phenological events and their environmental triggers in Mojave Desert ecosystems. Ecology 55, 856–863.
| Phenological events and their environmental triggers in Mojave Desert ecosystems.Crossref | GoogleScholarGoogle Scholar |
Beuhler M (2003) Potential impacts of global warming on water resources in southern California. Water Science and Technology 47, 165–168.
Brooks ML (1999) Alien annual grasses and fire in the Mojave Desert. Madrono 46, 13–19.
Brooks ML (2003) Effects of increased soil nitrogen on the dominance of alien annual plants in the Mojave Desert. Journal of Applied Ecology 40, 344–353.
| Effects of increased soil nitrogen on the dominance of alien annual plants in the Mojave Desert.Crossref | GoogleScholarGoogle Scholar |
Brooks ML, Esque TC (2002) Alien plants and fire in desert tortoise (Gopherus agassizii) habitat of the Mojave and Colorado deserts. Chelonian Conservation and Biology 4, 330–340.
Brooks ML, Matchett JR (2006) Spatial and temporal patterns of wildfires in the Mojave Desert, 1980–2004. Journal of Arid Environments 67, 148–164.
| Spatial and temporal patterns of wildfires in the Mojave Desert, 1980–2004.Crossref | GoogleScholarGoogle Scholar |
Brooks ML, Minnich RA (2006) Southeastern deserts bioregion. In ‘Fire in California’s ecosystems’. (Eds NG Sugihara, JWV Wagtendonk, KE Shaffer, J Fites-Kaufman, AE Thode) pp. 391–414. (University of California Press: Berkeley, CA)
Brooks ML, D’Antonio CM, Richardson DM, Grace JB, Keeley JE, DiTomaso JM, Hobbs RJ, Pellant M, Pyke D (2004) Effects of invasive alien plants on fire regimes. Bioscience 54, 677–688.
| Effects of invasive alien plants on fire regimes.Crossref | GoogleScholarGoogle Scholar |
Brown DE, Minnich RA (1986) Fire and changes in creosote bush scrub of the western Sonoran Desert, California. American Midland Naturalist 116, 411–422.
| Fire and changes in creosote bush scrub of the western Sonoran Desert, California.Crossref | GoogleScholarGoogle Scholar |
Brunelle A, Minckley TA, Blissett S, Cobabe SK, Guzman BL (2010) A nearly 8000 year fire history from an Arizona/Sonora borderland cienega. Journal of Arid Environments 74, 475–481.
| A nearly 8000 year fire history from an Arizona/Sonora borderland cienega.Crossref | GoogleScholarGoogle Scholar |
Cade BS, Noon BR (2003) A gentle introduction to quantile regression for ecologists. Frontiers in Ecology and the Environment 1, 412–420.
| A gentle introduction to quantile regression for ecologists.Crossref | GoogleScholarGoogle Scholar |
Clark CM, Morefield PE, Gilliam FS, Pardo LH (2013) Estimated losses of plant biodiversity in the United States from historical N deposition (1985–2010). Ecology 94, 1441–1448.
| Estimated losses of plant biodiversity in the United States from historical N deposition (1985–2010).Crossref | GoogleScholarGoogle Scholar | 23951703PubMed |
Crimmins MA, Comrie AC (2004) Interactions between antecedent climate and wildfire variability across south-eastern Arizona. International Journal of Wildland Fire 13, 455–466.
| Interactions between antecedent climate and wildfire variability across south-eastern Arizona.Crossref | GoogleScholarGoogle Scholar |
D'Antonio CM, Vitousek PM (1992) Biological invasions by exotic grasses, the grass fire cycle, and global change. Annual Review of Ecology and Systematics 23, 63–87.
Davis FW, Stoms DM, Hollander AD, Thomas KA, Stine PA, Odion D, Borchert MI, Thorne JH, Gray MV, Walker RE, Warner K, Graae J (1998) The California Gap Analysis Project–Final Report. University of California, Santa Barbara, CA. Available at http://legacy.biogeog.ucsb.edu/projects/gap/gap_rep.htm [Verified 3 January 2014]
DeFalco LA, Bryla DR, Smith-Longozo V, Nowak RS (2003) Are Mojave Desert annual species equal? Resource acquisition and allocation for the invasive grass Bromus madritensis subsp rubens (Poaceae) and two native species. American Journal of Botany 90, 1045–1053.
| Are Mojave Desert annual species equal? Resource acquisition and allocation for the invasive grass Bromus madritensis subsp rubens (Poaceae) and two native species.Crossref | GoogleScholarGoogle Scholar | 21659204PubMed |
Esque TC, Schwalbe CR, DeFalco LA, Duncan RB, Hughes TJ (2003) Effects of desert wildfires on desert tortoise (Gopherus agassizii) and other small vertebrates. The Southwestern Naturalist 48, 103–111.
| Effects of desert wildfires on desert tortoise (Gopherus agassizii) and other small vertebrates.Crossref | GoogleScholarGoogle Scholar |
Fenn ME, Baron JS, Allen EB, Rueth HM, Nydick KR, Geiser L, Bowman WD, Sickman JO, Meixner T, Johnson DW, Neitlich P (2003a) Ecological effects of nitrogen deposition in the western United States. Bioscience 53, 404–420.
| Ecological effects of nitrogen deposition in the western United States.Crossref | GoogleScholarGoogle Scholar |
Fenn ME, Haeuber R, Tonnesen GS, Baron JS, Grossman-Clarke S, Hope D, Jaffe DA, Copeland S, Geiser L, Rueth HM (2003b) Nitrogen emissions, deposition, and monitoring in the western United States. Bioscience 53, 391–403.
| Nitrogen emissions, deposition, and monitoring in the western United States.Crossref | GoogleScholarGoogle Scholar |
Fenn ME, Allen EB, Weiss SB, Jovan S, Geiser LH, Tonnesen GS, Johnson RF, Rao LE, Gimeno BS, Yuan F, Meixner T, Bytnerowicz A (2010) Nitrogen critical loads and management alternatives for N-impacted ecosystems in California. Journal of Environmental Management 91, 2404–2423.
| Nitrogen critical loads and management alternatives for N-impacted ecosystems in California.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3cXhtFyiu7nE&md5=c478e58a0cdbbb91b04389146e290977CAS | 20705383PubMed |
Fenn ME, Driscoll CT, Zhou Q, Rao LE, Meixner T, Allen EB, Yuan F, Sullivan TJ (in press) Use of a combined biogeochemical model approach and empirical data to assess critical nitrogen loads. In ‘critical loads and dynamic risk assessment of nitrogen, acidity and metals for terrestrial and aquatic ecosystems’. (Eds W de Vries, JP Hettelingh) (Springer: Dordrecht, Netherlands)
Gutiérrez JR (1992) Effects of low water supplementation and nutrient addition on the aboveground biomass production of annual plants in a Chilean coastal desert site. Oecologia 90, 556–559.
| Effects of low water supplementation and nutrient addition on the aboveground biomass production of annual plants in a Chilean coastal desert site.Crossref | GoogleScholarGoogle Scholar |
Hereford R, Webb RH, Longpré CI (2006) Precipitation history and ecosystem response to multidecadal precipitation variability in the Mojave Desert region, 1893–2001. Journal of Arid Environments 67, 13–34.
| Precipitation history and ecosystem response to multidecadal precipitation variability in the Mojave Desert region, 1893–2001.Crossref | GoogleScholarGoogle Scholar |
Holden ZA, Morgan P, Evans JS (2009) A predictive model of burn severity based on 20-year satellite-inferred burn severity data in a large southwestern US wilderness area. Forest Ecology and Management 258, 2399–2406.
| A predictive model of burn severity based on 20-year satellite-inferred burn severity data in a large southwestern US wilderness area.Crossref | GoogleScholarGoogle Scholar |
Keeley JE, Fotheringham CJ (2001) Historic fire regime in southern California shrublands. Conservation Biology 15, 1536–1548.
| Historic fire regime in southern California shrublands.Crossref | GoogleScholarGoogle Scholar |
Koenker R (2010) quantreg: quantile regression. (R Package version 4.53) Available at http://CRAN.R-project.org/package=quantreg [Verified 27 June 2012]
Koenker R, Bassett G (1978) Regression quantiles. Econometrica 46, 33–50.
| Regression quantiles.Crossref | GoogleScholarGoogle Scholar |
Krawchuk MA, Moritz MA (2011) Constraints on global fire activity vary across a resource gradient. Ecology 92, 121–132.
| Constraints on global fire activity vary across a resource gradient.Crossref | GoogleScholarGoogle Scholar | 21560682PubMed |
Littell JS, McKenzie D, Peterson DL, Westerling AL (2009) Climate and wildfire area burned in western U. S. ecoprovinces, 1916–2003. Ecological Applications 19, 1003–1021.
| Climate and wildfire area burned in western U. S. ecoprovinces, 1916–2003.Crossref | GoogleScholarGoogle Scholar | 19544740PubMed |
Margolis EQ, Balmat J (2009) Fire history and fire–climate relationships along a fire regime gradient in the Santa Fe Municipal Watershed, NM, USA. Forest Ecology and Management 258, 2416–2430.
| Fire history and fire–climate relationships along a fire regime gradient in the Santa Fe Municipal Watershed, NM, USA.Crossref | GoogleScholarGoogle Scholar |
McLaughlin SP, Bowers JE (1982) Effects of wildfire on a Sonoran Desert plant community. Ecology 63, 246–248.
| Effects of wildfire on a Sonoran Desert plant community.Crossref | GoogleScholarGoogle Scholar |
Meyn A, White PS, Buhk C, Jentsch A (2007) Environmental drivers of large, infrequent wildfires: the emerging conceptual model. Progress in Physical Geography 31, 287–312.
| Environmental drivers of large, infrequent wildfires: the emerging conceptual model.Crossref | GoogleScholarGoogle Scholar |
Noy-Meir I (1973) Desert ecosystems: environment and producers. Annual Review of Ecology and Systematics 4, 25–51.
| Desert ecosystems: environment and producers.Crossref | GoogleScholarGoogle Scholar |
Pardo LH, Robin-Abbott MJ, Driscoll CT (2011) Assessment of nitrogen deposition effects and empirical critical loads of nitrogen for ecoregions of the United States. USDA Forest Service, Northern Research Station General Technical Report NRS-80. (Newton Square, PA)
R Development Team (2010) R: a language and environment for statistical computing. (R Foundation for Statistical Computing: Vienna, Austria)
Rao LE, Allen EB (2010) Combined effects of precipitation and nitrogen deposition on native and invasive winter annual production in California deserts. Oecologia 162, 1035–1046.
| Combined effects of precipitation and nitrogen deposition on native and invasive winter annual production in California deserts.Crossref | GoogleScholarGoogle Scholar | 19967416PubMed |
Rao LE, Allen EB, Meixner T (2010) Risk-based determination of critical nitrogen deposition loads for fire spread in southern California deserts. Ecological Applications 20, 1320–1335.
| Risk-based determination of critical nitrogen deposition loads for fire spread in southern California deserts.Crossref | GoogleScholarGoogle Scholar | 20666252PubMed |
Research and Innovative Technology Administration (RITA) (2012) National transportation atlas database. Bureau of Transportation Statistics. Available at http://www.bts.gov/publications/national_transportation_atlas_database/ [Verified 9 December 2009]
Rogers GF, Vint MK (1987) Winter precipitation and fire in the Sonoran Desert. Journal of Arid Environments 13, 47–52.
Salo LF, McPherson GR, Williams DG (2005) Sonoran desert winter annuals affected by density of red brome and soil nitrogen. American Midland Naturalist 153, 95–109.
| Sonoran desert winter annuals affected by density of red brome and soil nitrogen.Crossref | GoogleScholarGoogle Scholar |
Scifres CJ, Hamilton WT (1993) Prescribed burning for brushland management: the south Texas example. (Texas A & M University Press: College Station, TX)
Seager R, Ting M, Held I, Kushnir Y, Lu J, Vecchi G, Huang H, Harnik N, Leetmaa A, Lau N, Li C, Velez J, Naomi N (2007) Model projections of an imminent transition to a more arid climate in southwestern North America. Science 316, 1181–1184.
| Model projections of an imminent transition to a more arid climate in southwestern North America.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2sXls1Kisb8%3D&md5=b87835429aba96d91bfb63b6e4bc98a0CAS | 17412920PubMed |
Slocum MG, Beckage B, Platt WJ, Orzell SL, Taylor W (2010) Effect of climate on wildfire size: a cross-scale analysis. Ecosystems 13, 828–840.
| Effect of climate on wildfire size: a cross-scale analysis.Crossref | GoogleScholarGoogle Scholar |
Steers RJ, Allen EB (2011) Fire effects on perennial vegetation in the western Colorado Desert, USA. Fire Ecology 7, 59–74.
| Fire effects on perennial vegetation in the western Colorado Desert, USA.Crossref | GoogleScholarGoogle Scholar |
Swetnam TW, Betancourt JL (1998) Mesoscale disturbance and ecological response to decadal climatic variability in the American Southwest. Journal of Climate 11, 3128–3147.
| Mesoscale disturbance and ecological response to decadal climatic variability in the American Southwest.Crossref | GoogleScholarGoogle Scholar |
Syphard AD, Radeloff VC, Keeley JE, Hawbaker TJ, Clayton MK, Stewart SI, Hammer RB (2007) Human influence on California fire regimes. Ecological Applications 17, 1388–1402.
| Human influence on California fire regimes.Crossref | GoogleScholarGoogle Scholar | 17708216PubMed |
Tonnesen GS, Wang ZS, Omary M, Chien CJ, Wang B, Morris R, Houyoux M, Adelman Z, Shankar U (2002) WRAP Regional Haze CMAQ 1996 Model Performance Evaluation. Presentation at the ‘WRAP Section 309 SIP Coordination Meeting’, 10 July 2002, Denver, CO.
Tonnesen GS, Wang ZS, Omary M, Chien CJ, Wang B (2003) Central California Ozone Study (CCOS) 2000 Model Intercomparison for SAQM, CMAQ and CAMx with CB4 and SAPRC99. Presentation to the California Air Resources Board, Sacramento, CA, 27 January 2003.
Tonnesen GS, Wang Z, Omary M, Chien CJ (2007) Assessment of nitrogen deposition: modeling and habitat assessment, PIER Energy-Related Environmental Research, CEC-500–2006–032. (California Energy Commission: Sacramento, CA)
US DOI (2009) WILDFIRE: wildland fire information. (US Department of Interior) Available at http://wildfire.cr.usgs.gov/firehistory/data.html [Verified 8 July 2009]
Weiss JL, Overpeck JT (2005) Is the Sonoran Desert losing its cool? Global Change Biology 11, 2065–2077.
| Is the Sonoran Desert losing its cool?Crossref | GoogleScholarGoogle Scholar |
Westerling AL, Gershunov A, Brown TJ, Cayan DR, Dettinger MD (2003) Climate and wildfire in the western United States. Bulletin of the American Meteorological Society 84, 595–604.
| Climate and wildfire in the western United States.Crossref | GoogleScholarGoogle Scholar |