

Distribution and abundance of roadkill on Tasmanian highways: human management options
Alistair J. Hobday A B D and Melinda L. Minstrell CA School of Zoology, Private Bag 5, University of Tasmania, Hobart, Tasmania 7001, Australia.
B CSIRO Marine and Atmospheric Research, GPO Box 1535, Hobart, Tasmania 7001, Australia.
C Department of Rural Health, Faculty of Health Science, University of Tasmania, Hobart, Tasmania 7001, Australia.
D Corresponding author. Email: alistair.hobday@csiro.au
Wildlife Research 35(7) 712-726 https://doi.org/10.1071/WR08067
Submitted: 9 May 2008 Accepted: 26 September 2008 Published: 17 November 2008
Abstract
An obvious sign of potential human impact on animal populations is roadkill. In Tasmania, this impact is perceived as relatively greater than in other Australian states, and is often noted by visitors and locals alike, such that calls for management action are common in the popular press. The goal of this three-year study was to assess the frequency and distribution of species killed on Tasmanian roads. Seasonal surveys were completed along five major routes, for a total of 154 trips. Over 15 000 km of road were surveyed and 5691 individuals in 54 taxa were recorded for an average roadkill density of 0.372 km−1. Over 50% of encountered roadkill could be identified to species, with common brushtail possums (Trichosurus vulpecula) and Tasmanian pademelon (Thylogale billardierii) the most common species identified, both in overall numbers and frequency of trips encountered. The 10 most common taxa accounted for 99% of the items observed. The seasonal occurrence, relationship with vehicle speed, and clustering in local hotspots for particular taxa all suggest that mitigation measures, such as vehicle speed reduction in specific areas, may be effective in reducing the number of animals killed. Mitigation measures, however, will not apply equally to all species and, in particular, success will depend on changing human behaviours.
Additional keywords: hotspots, vehicle speed, road mortality, traffic safety.
Introduction
Human impact on faunal populations is evident on many roads throughout the world, with carcasses from a wide range of taxa encountered in urban, country and wild regions (Dickerson 1939; Forman and Alexander 1998; Seiler and Helldin 2006). These carcasses, which mostly result from a lethal interaction with vehicles, constitute so-called roadkill. The impact of roadkill on animals can be direct (e.g. mortality) or indirect (e.g. population subdivision) (Forman and Alexander 1998; Trombulak and Frissell 2000). The impact on humans can likewise be direct (e.g. vehicle accidents) or indirect (e.g. decrease in enjoyment of a tourist holiday due to frequent roadkill encounters: Magnus et al. 2004).
Mortality due to roadkill can be significant, particularly for small populations. For example, 50% of the known deaths of the endangered Florida panther (Felis concolor) is due to roadkill (Harris and Gallagher 1989), while roadkill is the largest mortality factor for English populations of European badgers (Meles meles) (Gallagher and Nelson 1979). In fact, roadkill has been implicated in population declines of a range of iconic and/or threatened species, including moose (Alces alces) (Bangs et al. 1989), deer (Odocoileus virginianus) (Sarbello and Jackson 1985), koala (Phascolarctos cinereus) (Dique et al. 2003), wolf (Canis lupus) (Fuller 1989), eastern quoll (Dasyurus viverrinus) (Jones 2000), and amphibians (van Gelder 1973; Fahrig et al. 1995). Mortality varies among species; for example, 30% of common toads (Bufo bufo) crossing roads are killed by vehicles (van Gelder 1973), while for more ‘road-wise’ species, crossing roads is less problematic (Jones 2000).
Beyond an association with direct mortality, roads also have indirect impacts on animal populations due to habitat fragmentation and reducing movement of animals (Forman and Alexander 1998; Trombulak and Frissell 2000; Jaeger and Fahrig 2004). In turn, this is predicted to lead to restricted gene flow, although this has mostly been demonstrated in taxa with limited dispersal such as amphibians (Reh and Seitz 1990). Some animals may be attracted to roadsides for feeding opportunities, and foraging on roadkill may also have a direct benefit for scavenging species, provided they can avoid collisions (Forman and Alexander 1998).
Roadkill also has a direct impact on the human population, via accidents and insurance claims for wildlife–vehicle collisions (Seiler 2005). In Tasmania, Australia, in 2007 one insurer received 527 such claims, averaging A$1414 (R. Doedens, RACT, pers. comm.). Also in Tasmania, accidents as a result of encounters with animals have been linked to three human fatalities in the period 1993–2003, and an average of 7.2 injury-causing accidents per year were reported to police over the same period (Magnus 2006). A larger number of single-vehicle lethal accidents may be related to animal interactions – the animal escaped but the humans did not. The experience of tourists is also negatively affected by encountering high levels of roadkill when they visit Tasmania (Magnus et al. 2004).
Concern regarding the level of roadkill from the general population in Tasmania and elsewhere is obvious, and expressed through media interest and complaints in letters to the editors of the local papers (Hobday, personal collection; Magnus et al. 2004). Management responses include road signage, information to tourists, clean-up by councils, and speed limits (Dique et al. 2003; Magnus 2006). However, these responses tend to be reactionary, rather than targeted. In Tasmania there have been calls for management intervention to reduce the incidence of roadkill, but data to guide management decisions are often lacking (see also Dickerson 1939).
Thus, the motivation for this study was to collect baseline data needed to inform potential mitigation options. The study was designed to record the species seen as roadkill by the ‘typical driver’ and also the location, in order to assess the frequency of roadkill, describe the spatial and temporal patterns of roadkill distribution, identify whether roadkill hotspots occur at fine scales, and inform development of effective roadkill mitigation (Magnus 2006). Successful management intervention can typically only succeed following an understanding and awareness of roadkill patterns and causes (Seiler 2005). Responses can include attempts to mitigate via human behaviour (such as speed limits or road-signs), or animal behaviour (e.g. vegetation management, ramps, reflectors, culverts, whistles mounted on vehicles) (Magnus 2006; Seiler and Helldin 2006). For example, previous research in Tasmania showed that increased mortality and population decline of eastern quolls (Dasyurus viverrinus) and Tasmanian devils (Sarcophilus harrisii) was related to increased vehicle speed, and a subsequent reduction in speed led to population recovery (Jones 2000).
Media correspondents often argue that Tasmania may have a large roadkill because of the high abundance of many wildlife species (Short and Smith 1994). Thus, high levels of roadkill may be a consequence of high faunal density, with only minimal population impacts (Dickerson 1939; Seiler and Helldin 2006). While this untested claim is feasible at the statewide scale, it may not hold at smaller scales. Thus we test this hypothesis at smaller scales by relating fine-scale estimates of roadkill density to estimates of local faunal density from spotlight surveys. Analysis of seasonal differences in roadkill abundance, perhaps due to seasonal human or animal behaviours, were investigated as these may allow targeted mitigation at particular times of the year. To assess the need for speed-based mitigation options, we investigated relationships between (vehicle) speed and roadkill, to determine whether particular taxa were killed at high or low speeds, assuming that the survey vehicle speed is an indicator of the speed of the vehicle that killed the individual animal. Finally, cumulative roadkill as a function of speed was used to derive a mitigation curve to estimate the expected reduction in roadkill if speed of vehicles is reduced.
Methods
Data collection
Data were collected by an observer in a moving vehicle, using a GPS (Garmin GPS 36) and laptop computer. The survey vehicle was a Toyota Landcruiser with high passenger elevation and viewing windows. A location and speed from the GPS was recorded every second to generate a route profile. Accuracy of the GPS was to within 1 s and 10 m at 100 km h−1. Each roadkill item was manually logged using dedicated software on the laptop computer as the vehicle passed and then a species identity entered for that record. Vehicle speed was not modified to log or better identify the roadkill item. Thus, roadkill identification took place at the particular vehicle speed at the time of detection. If identification was not possible (sighting conditions, vehicle hazards, carcass degradation), then it was recorded as unidentified mammal, bird or snake (identification to this taxonomic resolution was always possible). Roadkill was not removed as this was impractical for the scale of the survey, and dangerous in many places. Thus, double-counting is possible on repeats of the same road within short periods, although density estimates will be unaffected. Both authors had extensive experience in roadkill identification before the implementation of the GPS system (AH: 15 years experience; MM: 2 years). Validation occurred via independent identification made by the driver and recorder, and via occasional roadside inspection of carcasses, revealing over 95% accuracy. In cases of uncertainty, the general taxonomic categories were used. There was no validation of detection efficiency, such as walking the surveyed route, or planting a known set of carcasses and then surveying (e.g. Taylor and Goldingay 2004). Surveying from a moving vehicle means that total roadkill may be underestimated as only animals above 500 g, and birds larger than 130 mm, could be reliably detected in our survey.
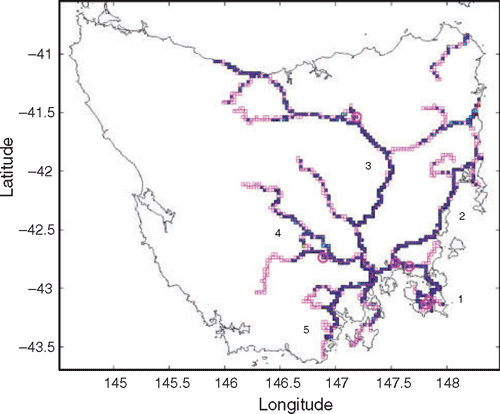
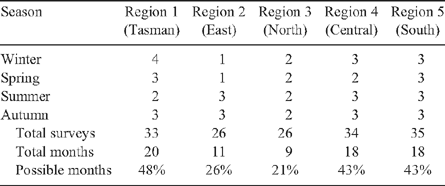
Survey routes covered between June 2001 and November 2004 were divided into five general regions radiating out from the city of Hobart (Fig. 1). The roads surveyed included major state and federal highways (often single-lane), and secondary sealed roads. Limited coverage of unsealed roads occurred, but these data were combined with those from sealed roads for analysis. Surrounding vegetation included native and regenerated forest, woodland, grasslands and farming regions, although this was not recorded during the surveys. An attempt was made to cover each region at least once per season (summer [December–February], autumn [March–May], winter [June–August] and spring [September–November]). This coverage was often exceeded, although exactly the same route was not covered in each replicate. Although persistence of roadkill in the environment is not well known, this and other surveys suggest maximum timescales on the order of weeks (e.g. Taylor and Goldingay 2004), and thus we assumed our seasonal surveys were independent.
|
In 16 of the 154 surveys, species-occurrence data could only be recorded using the vehicle speedometer, due to exhaustion of the GPS battery. These data are included only in summary analyses, while analyses requiring exact position used only the GPS-complete survey data.
Data analyses
Analyses at three spatial scales were conducted: statewide, regional, and fine-scale. Each division of the data reduced the numbers in any category, limiting the statistical treatment of the data. ANOVA and linear regression were used to test hypotheses, and an α of 0.05 was used to indicate significant differences. Analyses and statistical tests were conducted using customised software written in Matlab (v7 The Mathworks).
Data were collected from a moving vehicle that attempted to maintain the posted or suitable road speed during daylight hours; as a result, several biases are possible. For example, observer-related improvements over time in species identification or detection ability may introduce spurious temporal trends, while a detection size-bias, with small roadkill detected only at low speeds, may lead to underestimates of the smaller species. These potential biases are examined as a precursor to more detailed analyses using regression analyses. Adult weight averaged over the sexes was used as a proxy for size in mammals (Strahan 1983), while length of birds was taken from Reader’s Digest (1988). Seasonal patterns in overall roadkill density and for each taxon were tested with ANOVA.
For assessment of annual and seasonal effects on roadkill abundance, years were defined from June to May because surveys began in June 2001, at the beginning of a winter. Data from June to November 2004 were excluded from annual analyses as they represented only a partial fourth year.
Fine-scale distribution – roadkill hotspots
To investigate the fine-scale spatial distribution of roadkill, all surveyed roads were divided into a grid of 0.025-degree boxes, hereafter called route-boxes. At the latitude of Tasmania (~42°S), these correspond to boxes of ~2.7 × 2.1 km in dimension. The density of roadkill within each of the route-boxes was analysed. Use of larger and smaller route-box scales (0.01–0.1 degree) did not markedly change the results.
Roadkill hotspots were investigated using the subset of route-boxes (1) with more than five visits and a total distance surveyed of more than 20 km, and (2) where more than 10 roadkill items were recorded. This dual threshold was employed to minimise spurious identification of hotspots. Thus, hotspots were areas with high roadkill detected on multiple occasions with sufficient sample effort. The locations of highest roadkill density for all taxa combined, and for selected taxa, were identified from this hotspot analysis.
Roadkill and speed
This analysis uses the speed at which the roadkill was logged as a proxy for the vehicle speed at which the animal was killed. The overall relationship between vehicle speed and roadkill was analysed with cumulative frequency plots and mitigation curves. If speed is not related to roadkill, then a null model would suggest that each speed class should contain the same relative frequency as roadkill. That is, if 10% of the roads were surveyed at a speed of 90–95 km h−1, the null model would suggest that this speed interval should contain 10% of the roadkill. Vehicle speeds recorded every second from the GPS were used to generate the speed frequency data. The ratio of observed to expected roadkill (based on relative frequency of the speed interval) can be used to correct a mitigation curve. The mitigation curve is the cumulative percentage of roadkill observed as a function of speed (corrected or uncorrected) and is used to illustrate mitigation benefits.
Given that vehicle speed varies between urban and country areas, and that animal abundance is higher in country areas, a potential relationship between speed and roadkill density may be spurious. To examine this possible bias, analysis of speed at the scale of the route-box was undertaken. Both speed limit and driving conditions vary within individual route-boxes, in both country and urban areas. If roadkill are associated with the fastest portion of each route-box, then the role of speed can be isolated. Route-box speed is defined as the mean speed of the vehicle travelling along roads in the route-box (recorded every second).
Roadkill relationship to faunal density
The relationship between roadkill and faunal density was examined by analysis of the frequency of live animals from spotlight faunal surveys conducted at over 30 locations around Tasmania, for the period 1995–2003. These data were collected annually for one to seven 10-km replicates within each spotlight location (Driessen et al. 1996; Greg Hocking, DPIW, Tasmania, unpubl. data). Abundance of 16 common mammal species was recorded in the spotlight surveys, of which all but fallow deer (Dama dama) and forester kangaroo (Macropus giganteus) were recorded in this study (see Table 3).
To address the hypothesis that roadkill density is related to faunal density, each route-box was matched to the closest spotlight location. Selected route-boxes had to be within 20 km of a spotlight survey location and have had greater than 5 km of survey effort. Spotlight data were converted to sightings per kilometre (density). Correlation analyses, using all species in each dataset and only the species in common, were undertaken.
Results
Abundance of roadkill
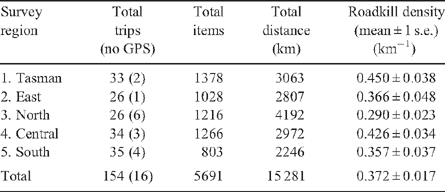
In total, 154 surveys (138 using GPS) were completed over the 42 months of the survey period, covering 15 281 km (Table 1, Fig. 1). In all, 5691 roadkill items in 54 taxa, comprising 22 mammal, 32 bird and 2 reptile species, were recorded (Table 1). Of these items, 57% could be identified to a specific taxon, while 43% could be classified only as unidentified mammal, bird or snake. The overall density of roadkill detected was 0.372 km−1 (one roadkill item every 2.7 km) (Table 1). The average survey covered 99.2 km (range 16–324 km) and recorded 37 (range 3–125) roadkill items in 8.3 (range 2–15) taxa.
|
Each region was surveyed between 26 and 35 times, with more than 2000 km total effort (Table 1), over 1–4 seasons (Table 2). Each region was surveyed in 9–20 of the 42 months of the study (21–48% of possible months) (Table 2).
|

|
The 10 most abundant taxa accounted for 99% of the roadkill recorded, and there were many species with fewer than 10 individuals recorded (Table 3). The most frequent item was unidentified mammals, at 38% of all items detected. The most frequent species identified was the common brushtail possum (BTP), accounting for 27% of all carcasses, and 48% of all identified carcasses (Table 3). BTP were recorded in 97% of the surveys (Table 3). Other taxa comprising the top 10 were Tasmanian pademelon, rabbit, unidentified birds, Bennetts’ wallaby, silver gull (Larus novaehollandiae), masked lapwing (Vanellus miles), forest raven (Corvus tasmanicus), and Tasmanian devil. A total of 49 Tasmanian devils was recorded, which is less than 1% of the total carcasses (1.5% of identified taxa). However, they were widespread and encountered in 20% of the trips (Table 3).
There was no significant trend in the detection of roadkill items or taxa over time (regression tests: items: F1,153 = 0.142, P = 0.707; taxa: F1,153 = 0.817, P = 0.367). The percentage of unidentified roadkill (mammal, bird or snake) also remained constant over time, at ~43%. Extremes in unidentified roadkill (0–70%) occurred by chance in smaller surveys, where less roadkill was detected.
The smallest mammal detected in the surveys was the eastern barred bandicoot (640 g), and the smallest bird, the European goldfinch (Carduelis carduelis) (130 mm). While a detection bias probably exists for a wider range of animal sizes (e.g. frogs and small lizards were not detected in this study), there was no significant relationship between mean detection speed and body size (weight) for mammals (taxa with more than 10 observations: regression F1,9 = 0.952, one-tailed P = 0.18). For the seven bird species with more than 10 observations, body size (length) of recorded individuals did increase with mean speed (regression F1,5 = 5.21, one-tailed P = 0.035). Overall, however, the survey method was not sensitive to the potential biases examined, and the dataset is appropriate for more detailed analyses.
Seasonal and interannual patterns
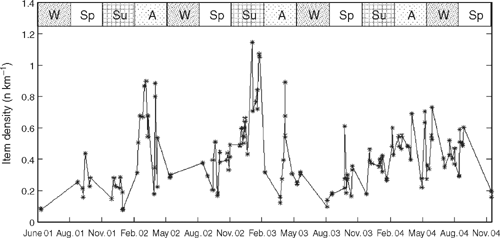
There was both interannual and seasonal variation in the density of roadkill over the course of this study (Fig. 2). Mean roadkill density varied between years, with 2003–04 the lowest (0.25 km−1, 51 surveys) and 2002–03, the highest (0.49 km−1, 46 surveys). In the first year of the study, 2001–02, there was an intermediate density (0.36 km−1, 36 surveys). Density of roadkill was lowest in winter and highest in late summer and autumn, when over 1 carcass km−1 was observed in some surveys (Fig. 2).
|
Overall, there was a significant difference in density between seasons (ANOVA: F3,134 = 6.516, P < 0.0004) (Fig. 3), although at the regional level the difference was significant only for Region 4 (ANOVA: F3,27 = 3.739, P < 0.023). Each region had a slightly different peak in roadkill, either summer or autumn (Fig. 3). A total of 12 individual taxa demonstrated a significant seasonal difference in density (Table 3). An example is shown for common brushtail possum (BTP) (Fig. 4). For all regions combined there was a significant seasonal difference, with BTP density highest in autumn (ANOVA: F3,134 = 6.003, P < 0.0008). At the regional scale, there was a significant seasonal BTP density difference only in Region 4. There was no significant trend in roadkill density for the combined dataset over the 42 months for any species.
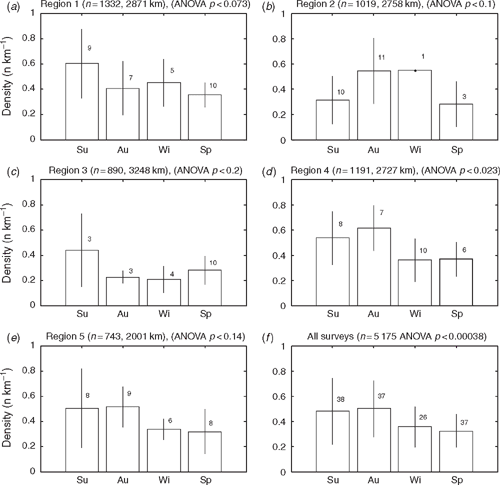
|
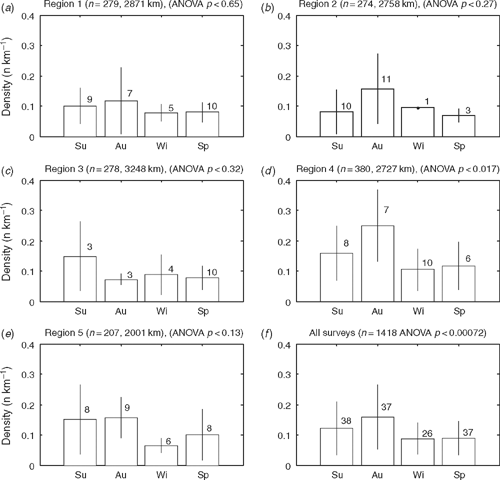
|
Hotspots – statewide and regional patterns
At the statewide scale, there were 6858 total route-box visits, and some route-boxes had up to 60 visits. The mean distance travelled in each box visit was up to 6 km (for extremely curved roads), and some boxes had over 100 km of total survey effort. Maximum roadkill density in individual route-boxes without considering minimum survey effort was over 6 km−1. However, among the identified ‘hotspots’ (see Methods), average roadkill densities in the six highest-density ones exceeded 1.3 items km−1 (maximum 2.32 km−1). Hotspots were found around the state, with four of the highest-density hotspots in Region 1 (Tasman), one in Region 3 (North), and one in Region 4 (Central) (Fig. 1).
Within each of the five regions, the range of density for the six highest-density hotpots was: Region 1: 1.08–2.32 km−1; Region 2: 0.67–2.25 km−1; Region 3: 0.75–1.39 km−1; Region 4: 0.87–2.04 km−1; and Region 5: 0.64–0.97 km−1. The locations of the six highest-density route-boxes in Region 1 provide examples of regional patterns (Fig. 5). Four are clumped in the southern portion of the Tasman Peninsula where native forest occurs, while two are in farming and woodland regions. Distribution of roadkill within the route-box hotspots at a scale of less than 2 km varies from apparently random, continuous, or clumped (Fig. 6).
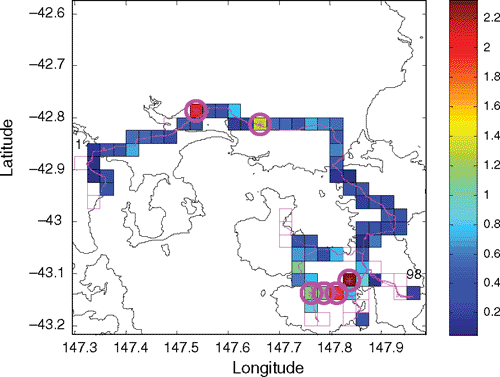
|
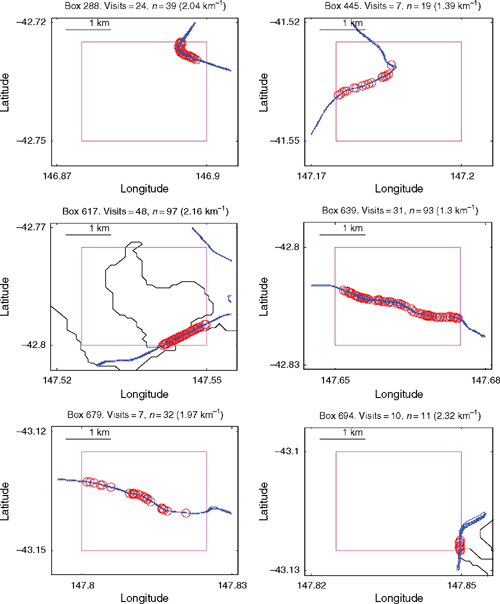
|
Together, these results demonstrate that roadkill is not distributed evenly along the surveyed roads, and when route-boxes are ordered from highest to lowest this becomes even more apparent (Fig. 7). Ranking the cumulative abundance of roadkill demonstrates that 70% of the roadkill occurs in 20–45% of the road in each region (Fig. 7b). Similarly, 70% of the roadkill density for a trip occurs in 20–30% of the road, depending on the region (Fig. 7d). Overall, for 30% of the route, abundance/density of roadkill is negligible or zero.
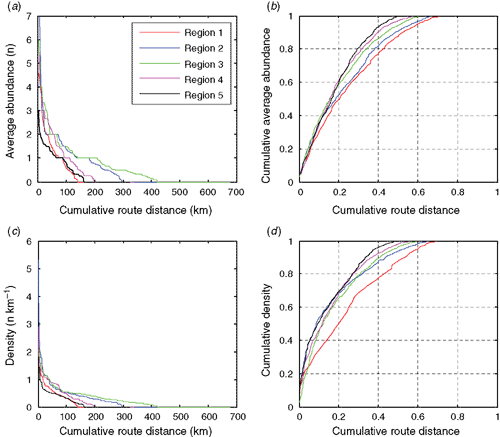
|
Associations between speed and roadkill
Overall, relatively more roadkill was observed at higher speeds, with 50% of roadkill detected when the survey vehicle speed was greater than 80 km h−1 (Fig. 8a). If roadkill is randomly distributed with regard to vehicle speed, then the ratio of observed to expected roadkill in a particular speed interval should equal 1.0. However, at low speeds relatively few roadkill were observed, compared with high speeds (Fig. 8b). Using a maximum speed of 100 km h−1 for calculational convenience, the mitigation curve shows that a reduction in speed of 20% would be expected to result in a reduction in roadkill of ~50% (Fig. 8c). Speed was related to roadkill density even within small regions, where faunal density might be expected to be similar (mean route-box transit distance is ~2 km). In all regions, roadkill was observed at locations in each route-box where the survey vehicle travelled fastest (Table 4).
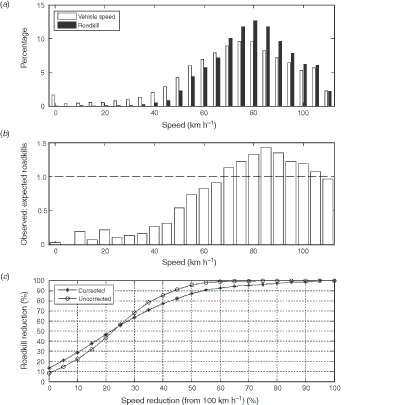
|
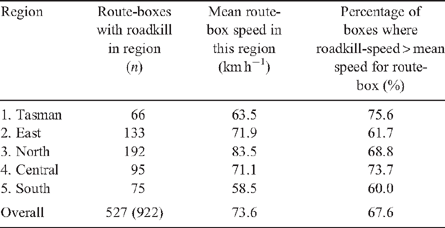
|
The mean speed at which each taxon was observed as roadkill was significantly different (ANOVA: F20,5044 = 9.305, P < 0.001) (Fig. 9). Some taxa, such as echidna (ECH) were seen at high average speeds, while others were seen at lower average speeds (e.g. eastern barred bandicoot, BAN). For some species, lower average speeds may indicate habitat distribution; for example, house sparrow (Passer domesticus, HSP) and blackbird (Turdus merula, BBI) are predominately found in urban areas where speeds are lower.
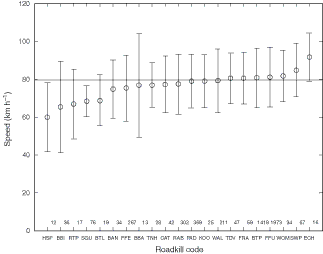
|
Is faunal density related to roadkill density?
Faunal spotlight densities vary widely across Tasmania, suggesting different local population sizes of particular species, but there were no significant trends in density of live taxa over time recorded in these spotlight surveys. Correlation analyses using the 256 roadkill route-boxes within 20 km of a spotlight survey location showed that the density of roadkill was not related to the density of live animals (regression test: F1,254 = 0.241, P = 0.624). The spotlight survey included a limited set of animals (16 taxa), and no birds. However, even when the analysis included only the density of the shared and abundant taxa (BTP, PAD, WAL: Table 3), there was no significant relationship between live density and roadkill at this scale.
Discussion
This study is one of the most extensive conducted in Australia, and possibly worldwide, in terms of time-span (>3 years), geographic range (>200 km), species coverage (>50) and survey effort (>15 000 km) (see also Smith-Patten and Patten 2008). As in other studies, a few taxa dominated the roadkill fauna (e.g. Taylor and Goldingay 2004; Smith-Patten and Patten 2008). Analyses at three spatial scales (statewide, regional, and fine-scale) showed large spatial variation in roadkill density. At the largest scale, the density of roadkill in Tasmania (0.372 km−1) was higher than estimates from surveys of similar effort in Australia and elsewhere (Table 5). This overall high roadkill density is probably due to the high density of wildlife in Tasmania, which has been attributed to less habitat disturbance, and, until recently, an absence of foxes (Short and Smith 1994; Dennis 2002). High-density locations were distributed around the state, indicating that mitigation could be locally targeted, rather than applied statewide. Seasonal peaks in roadkill differed between regions, also indicating that mitigation measures could be applied during specific seasons, rather than at all times of the year, and still reduce roadkill.

|
Roadkill density was highest along roads surveyed in the Tasman Peninsula region (0.450 ± 0.038 km−1, mean ± 1 s.e.) and lowest along roads in the Northern region (0.290 ± 0.028 km−1). Within each of these regions, however, high roadkill densities occurred in local hotspots. Detailed maps like those in Fig. 6, for any route-box and any taxa, demonstrate the fine-scale distribution of roadkill. It is at this scale that the appropriate mitigation measures can be selected and targeted. For example, animal escape ramps would be appropriate if roadkill is clumped, road fencing for shorter segments of road with high roadkill, and speed restrictions or signage warning of the high chance of roadkill for longer segments with more continuous roadkill. The maximum densities of roadkill in hotspots in each region exceeded 2 km−1. If less stringent criteria were used to identify hotspots (repeated surveys and minimum roadkill encountered), even higher densities could be claimed. Hotspots identified from multiple surveys are most likely to be persistent, rather than transitory, and so are suitable candidate areas for mitigation efforts if a reduction in roadkill is sought (e.g. Magnus 2006).
The most common roadkill species (for example the common brushtail possum, Tasmanian pademelon and Bennetts’ wallaby) were generally not of high conservation concern. Thus, while unsightly, impacts may not be sufficient to lead to population declines, unless populations are under stress (e.g. Tasmanian devil facial tumour disease: Hawkins et al. 2006), or small (Jones 2000), at which time roadkill mortality may be significant (Seiler and Helldin 2006). Other common taxa were non-native (e.g. feral cat), and it is possible that such roadkill may even be a benefit to native wildlife.
Estimated abundance of roadkill in Tasmania
Preliminary estimates of the total mortality due to roadkill can be made using the data collected in this study, which focussed on state and federal highway almost exclusively. The surveys covered a total of 922 route-boxes (0.025 degree), with an average road length of 2.03 km, giving a total estimate of 1872 unique kilometres of roads. The average roadkill density was 0.372 km−1, leading to an estimated abundance of 696 roadkill over the survey area at any given time. The 1872 km surveyed here represents ~53% of the 3507 km of state and federal roads, leading to a standing stock estimate of 1305 roadkill on these roads at any given time. Roadkill does not persist indefinitely on the road: it is removed, washed away, flattened beyond recognition and scavenged. If the persistence is two weeks (Taylor and Goldingay 2004) (turnover 26 times per year), then the total estimated roadkill is 33 930 per year on these roads. Given a total of 23 380 km of roads in Tasmania (Doug Ling, RACT, pers. comm.), and assuming a similar density of roadkill on all roads, an estimated 226 131 roadkills occur per year. Turnover times of four weeks (low estimate) or one week (high estimate) lead to estimates of 113 000 and 450 000 roadkill, respectively. It has also been estimated that up to 30% of animals struck by cars die some distance from the road, where they would not be counted in our survey (this estimate is from roadside searches on foot compared with vehicle searches and immediate attempts to find individual animals heavily struck in collisions: N. Mooney, DPIW, pers. comm.). Using this factor to further extrapolate our figures, Tasmanian roadkill of animals larger than bandicoots and small birds is thus in the range of 377 000–1 500 000. If the mid-estimate of 294 000 animals is used after correction for mortality away from the road (226 000 × 130%), then mortality for individual species based on the percentage occurrence in the observed sample can be estimated. For example, the common brushtail possum represented 48% of the identified taxa, leading to an annual mortality estimate as roadkill of 108 543 individuals. By way of context, the legal Tasmanian harvest of common brushtail possum for meat in 2000 was 41 000 (DPIW). Total Bennetts’ wallaby mortality (7% of identified roadkill) is estimated as 15 829 year−1, while Tasmanian devils (1.5% of identified roadkill) is 3392 year−1. At the time of this survey, the total population of Tasmanian devils was estimated to be 60 000–90 000 (C. Hawkins, DPIW, pers comm.). Thus, roadkill mortality represented ~3.8–5.7% of the estimated Tasmanian devil population at that time. This mortality on roads is likely to be significant in the context of the recent impact of disease on Tasmanian devils (Hawkins et al. 2006).
Factors influencing roadkill
The goal of this study was to document the patterns in roadkill around Tasmania in order to inform effective mitigation. A framework for understanding the detection of roadkill patterns is presented here to stimulate additional research, and some of the factors relevant to the present study are briefly discussed (Table 6). Obviously, roadkill events result from the interaction of an animal and a human-operated vehicle. The natural contributing factors include, for example, animal density (no relationship detected in this study), and foraging behaviours that attract animals to roads (Table 6; see also Seiler and Helldin 2006). Human contributions can include the speed of the vehicle, and the time of day at which driving occurs. Once the mortality event has occurred, persistence of the carcass on the road is again subject to natural and human influences, which will determine whether the carcass is subsequently detected in a survey. Finally, if the carcass persists on the road, detection will also be influenced by natural factors, such as the amount of vegetation that may screen the carcass at the side of the road, and human factors such as detection abilities due to sun glare or approaching traffic. This framework will be important in consideration of mitigation options as variation in any of these factors can obscure the true pattern of mortality.

|
Significant differences in mean annual roadkill density were found among the three years of this study. A variety of explanations are possible, including variation in environmental conditions, such as rainfall, which may interact with the behaviour of animals such that fewer/more encounter roads (Table 6). Changes in animal abundance can also lead to changes in roadkill. However, no significant changes in faunal density estimated from roadkill or spotlighting surveys occurred over the course of this study. Although non-significant, there was a decline in roadkill densities of Tasmanian devils (regression: F1,136 = 2.398, P = 0.123), in line with its documented population decline due to disease over the same period (Hawkins et al. 2006).
Significant seasonal variation was observed in the most abundant taxa only, showing that even with this high level of sampling effort, differences can be hard to detect, assuming they exist. Seasonal differences within regions for individual taxa, such as the brushtail possum, may allow selection of mitigation measures that focus on species of conservation interest in a particular region for a portion of the year. Those observed here may be due to changes in human and/or animal behaviours (Table 6). For example, vehicle traffic may increase during summer-holiday periods, and/or animal movements may increase during periods of juvenile dispersal or breeding, or in response to seasonally changing resource availability (Smith-Patten and Patten 2008). However, roadkill abundance in this study peaked between late summer and autumn, depending on year, rather than in early summer, when juveniles of the three most common roadkill taxa (BTP, PAD and WAL) disperse (Strahan 1983). This may suggest that seasonal patterns are determined by an environmental factor that is more likely to change annually and with local conditions, such as resource availability. The analyses of seasonal density changes conducted here used calendar months as a proxy for seasons. However, seasons defined on the basis of rainfall (which has interannual variation) may also be appropriate. This remains an area for future investigation with these data, although caution should be exercised in a posteriori searching of large datasets until significant differences are found.
At the finest spatial scale investigated in this study there was also variation in the distribution of roadkill. This is the most interesting scale with regard to mitigation, as it is the scale at which individual mortality events occur. Human and natural factors can be manipulated at this scale to reduce roadkill.
Management implications – mitigating roadkill
The analyses presented here showed that localised high-density roadkill areas, or ‘hotspots’, exist on Tasmanian roads. Consideration of the statewide data identified the hotspots with most roadkill, while the regional analyses identified locations that may be of interest to managers charged with implementing mitigation measures (Magnus 2006). The discovery of these roadkill hotspots, and the finding that even within route-boxes roadkill can be clumped (Fig. 6), has important implications for those who seek to mitigate roadkill (e.g. Magnus 2006). Localised hotspots allow local (~1-km scale) and seasonal mitigation strategies and may inform design of new roads that minimise impacts on wildlife (e.g. Taylor and Goldingay 2003; Seiler 2005; Magnus 2006; Grilo et al. 2008). For existing roads, or where engineering modifications are not cost-effective, mitigation can be targeted at animal or human behaviour.
Mitigation via changes to animal behaviour
Mitigation of roadkill via changing animal behaviour has been attempted for a variety of measures, including ultrasonic whistles, overpasses, escape routes, roadside lighting or reflectors, reduction of roadside grass and water (Magnus 2006). In general, when tested in appropriate experiments, there has been a lack of success with interventions aimed at changing animal behaviour, or animal discouragement (Reeve and Anderson 1993). For example, in a single-blind experiment testing the efficacy of ultrasonic whistles mounted on vehicles, there was no observable difference in behaviour of animals when the whistles were activated and not activated (Z. Magnus, unpublished). Of 1460 animals observed, 2.7% (n = 39) were hit by the vehicle from which the trials were being conducted (the driver was independent of the researcher). There was no significant difference between the number of animals hit when the whistles were activated (n = 18) compared with not activated (n = 21) (Magnus et al. 2004).
Not all animals that encounter roads are killed and many complete crossings between road-separated habitats (e.g. Seiler and Helldin 2006). Preventing animal access to roads has been used in some areas where contact between animal and vehicle is potentially lethal to humans (e.g. moose in northern Canada). However, complete habitat division may be an inappropriate and costly mitigation (Seiler and Helldin 2006). Mitigation that completely prevents animals from crossing roads may lead to more population fragmentation, smaller population sizes, and reduced genetic diversity (Forman and Alexander 1998).
Mitigation via changes to human behaviour
It may be more effective to instead modify human behaviour (but see Walker et al. 2006) at times and locations where roadkill occurs. The human mitigation strategy used most widely is adjustment of vehicle speed, providing both animals and humans with a greater time for avoidance (Seiler and Helldin 2006). However, speed limits, as such, may not be legally enforceable, and their efficacy is unknown. The most common attempt to achieve speed reduction is via installation of road signs warning of animal presence, but their effectiveness has been questioned (Dique et al. 2003). In one study, rates of grey kangaroo roadkill before and after warning road signage was installed did not differ (Coulson 1982). Recent signage installed in Tasmania has included spatial and temporal information (e.g.‘dusk till dawn, next 2 km’: Magnus et al. 2004). Lack of observed wildlife or roadkill in such signed areas has been cited as a reason that drivers do not adjust speed. The obvious alternative, leaving large carcasses in obvious places, may be unsafe for vehicles, as well as conflict with tourist experiences (Magnus et al. 2004). In limited locations, where speed can be reduced via obstacles, roadkill has been significantly reduced (Jones 2000), while in other locations, the speed limits have become legally enforceable (e.g. koala zones in south-east Queensland: Dique et al. 2003). Speed reduction as a roadkill mitigation approach is not likely to be equally effective for all species, however, as the mean speed at which species occurred as roadkill in this study varied. Animals killed at low speeds are likely to be those that have accidental encounters with vehicles (for example, as happens commonly with wallabies dashing from the scrub) rather than those that cannot avoid the vehicle (or vice versa) (for example, spotted-tailed quolls, which are slow to react as a speeding vehicle approaches). Reduction of speed should decrease the incidence of roadkill for species killed at high speeds (e.g. echidna), but obviously will be less effective for species killed at lower speeds.
The period of time that humans have to reduce collisions with animals can also be increased by increasing road visibility (Seiler 2005), for example by roadside mowing (e.g. Magnus 2006). This has the added advantage of removing cover for animals, such that roadsides may become less favoured. Of course, mowing may also promote growth of fresh grasses, which may be favoured as forage by some species. A detailed consideration of the negative effects of a seemingly positive mitigation strategy should be undertaken as part of a management response plan (Magnus 2006).
Together, analyses presented here showed that roadkill was not distributed evenly in time or space, and local hotspots exist. There was an association between vehicle speed and roadkill density at small scales. The implication of this finding is that human behaviour, via driving speed, could be modified in a limited portion of the road for some portion of the year. As an example, consider a journey over a distance of 200 km. At a speed of 100 km h−1, this journey would take 2 h. If vehicle speed was reduced by 20% (to 80 km h−1) in only 10% of the road (20 km), travel time for this trip would increase by 3 min (2.5% longer), and according to our results, potentially decrease overall roadkill by up to 50%. Overall, reduction of speed is likely to be an effective solution for humans seeking to reduce roadkill in Tasmania and elsewhere.
Acknowledgements
We appreciate the contribution of the following people for assisting with data recording: Alastair Walsh, Dan Ricard, Peter Hobday, Simon Talbot, Bruce Deagle, Scott Ling, Adam Stephens, Paige Eveson, George Watters, Brian Cox, Liesel Fitzgerald, Sarah Metcalf, and Cam Jones. Supplementary data were provided by Doug Ling (RACT) and Greg Hocking (Department of Primary Industries and Water), and we are grateful to them. Suggestions and comments from Zoe Magnus, Greg Hocking and Nick Mooney improved the discussion and implications identified in this paper. Reviews by associate editor Andrea Taylor and two anonymous referees helped improve the focus of this paper considerably, and are gratefully appreciated.
Bangs, E. E. , Bailey, T. N. , and Porter, M. F. (1989). Survival rates of adult female moose on the Kenai Peninsula, Alaska. Journal of Wildlife Management 53, 557–563.
| Crossref | GoogleScholarGoogle Scholar |
Hawkins, C. E. , Baars, C. , Hesterman, H. , Hocking, G. J. , and Jones, M. E. , et al. (2006). Emerging disease and population decline of an island endemic, the Tasmanian devil Sarcophilus harrisii. Biological Conservation 131, 307–324.
| Crossref | GoogleScholarGoogle Scholar |
Reeve, A. F. , and Anderson, S. H. (1993). Ineffectiveness of Swareflex reflectors at reducing deer–vehicle collisions. Wildlife Society Bulletin 21, 127–132.
Short, J. , and Smith, A. (1994). Mammal decline and recovery in Australia. Journal of Mammalogy 75, 288–297.
| Crossref | GoogleScholarGoogle Scholar |
Taylor, B. D. , and Goldingay, R. L. (2003). Cutting the carnage: wildlife usage of road culverts in north-eastern New South Wales. Wildlife Research 30, 529–537.
| Crossref | GoogleScholarGoogle Scholar |

Taylor, B. D. , and Goldingay, R. L. (2004). Wildlife road-kills on three major roads in north-eastern New South Wales. Wildlife Research 31, 83–91.
| Crossref | GoogleScholarGoogle Scholar |
Trombulak, S. C. , and Frissell, C. A. (2000). Review of ecological effects of roads on terrestrial and aquatic communities. Conservation Biology 14, 18–30.
| Crossref | GoogleScholarGoogle Scholar |
van Gelder, J. J. (1973). A quantitative approach to the mortality resulting from traffic in a population of Bufo bufo L. Oecologia 13, 93–95.
| Crossref | GoogleScholarGoogle Scholar |
Walker, L. , Williams, J. , and Jamrozik, K. (2006). Unsafe driving behaviour and four wheel drive vehicles: observational study. British Medical Journal ,
| Crossref | GoogleScholarGoogle Scholar | PubMed |