

Influence of land development on home range use dynamics of female elk
S. L. Webb A B , M. R. Dzialak A , S. M. Harju A , L. D. Hayden-Wing A and J. B. Winstead AA Hayden-Wing Associates, LLC, 2308 South 8th St, Laramie, WY 82070, USA.
B Corresponding author. Email: stephen@haydenwing.com
Wildlife Research 38(2) 163-167 https://doi.org/10.1071/WR10101
Submitted: 19 January 2010 Accepted: 25 February 2011 Published: 20 April 2011
Journal Compilation © CSIRO Publishing 2011 Open Access CC BY-NC-ND
Abstract
Context: Many ungulate species exhibit strong site fidelity to previously established areas, particularly females. However, development of the landscape may cause animals to shift their distribution to more secure areas.
Aims: To determine range use dynamics (i.e. size and overlap of core areas and home ranges) of female elk (Cervus elaphus) relative to land development features (e.g. roads, well pads, buildings, developments, towns, etc.) after controlling for environmental features (i.e. forest cover).
Methods: During the four-year study, we fitted elk (n = 165) with GPS collars annually and programmed collars to attempt one location fix every 3 h (eight locations per day) for one year. Females (n = 18) were subsequently recaptured and refitted with GPS collars to provide range use dynamics of individual elk over two to three years. We calculated sizes of core areas and home ranges using adaptive kernel estimators, overlap between annual ranges, and establishment of ranges relative to land development.
Key results: Overlap of annual core areas (48.6%) and home ranges (67.9%) was high despite annual increases in land development. Sizes of core areas and home ranges and annual overlap (i.e. site fidelity) were negatively influenced by land development after controlling for forest cover.
Conclusions: These data reveal that female elk show high levels of site fidelity even in the presence of increasing annual land development. Females did not appear to abandon previously established areas, but used ranges in a manner that minimised interaction with development within these areas based on reductions in range use size and fidelity as land development increased.
Implications: To help mitigate impacts on elk, land development should be minimised and large areas of forest protected so elk can avoid areas associated with human activity.
Additional keywords: Cervus elaphus, Colorado, core area, fidelity, forest cover, home range, land development, roads.
Introduction
Development of land can affect home range dynamics and space use patterns of ungulates (Van Dyke and Klein 1996). Therefore, understanding effects of land development and environmental features responsible for variation in range use and fidelity becomes increasingly important as landscapes are fragmented due to human development (e.g. buildings, houses, roads) and land-use practices (e.g. timber harvest, energy development). Home range size and fidelity have important implications for management and conservation of populations (McCoy et al. 2005; Felix et al. 2007; Webb et al. 2007; Walter et al. 2009; Tracz et al. 2010) because of shifting distributions of animals within populations.
Forest cover, land development and human activity may influence where animals settle, and thus, have implications on population size, persistence and cohesion of animals in the wild. For instance, the home range size of female elk (Cervus elaphus) is influenced positively by the amount of forest cover in areas where animals are exposed to predation risk (Anderson et al. 2005). In areas of human activity, forest cover mitigates some of the impacts that human activity have on animal behaviour (Edge et al. 1985; Kuck et al. 1985; Unsworth et al. 1998; Rumble et al. 2005). These examples illustrate the importance of forest cover for reducing predation pressure or human disturbance and for shaping space use patterns of animals. Even in the presence of forest cover, land development is likely to influence space use patterns, in which year-to-year fidelity of ranges may be reduced and animals may use small areas further from development more intensively.
In the Rocky Mountain region (including Colorado, New Mexico and Wyoming), coal-bed natural gas (CBNG) production has been increasing since 1989 (Bureau of Land Management 2005). Because development for CBNG is expanding (Bureau of Land Management 2005), we considered changes in annual space use patterns of female elk as a means to quantify the effects of increasing levels of development (3894 ha during year 1 to 4433 ha during year 4) on populations. The objectives of this study were to: (1) document sizes and overlap of annual core areas and home ranges, (2) determine how land development influences size and overlap of annual core areas and home ranges, and (3) determine the relationship between development and forest cover within core areas and home ranges.
Materials and methods
Study area
The 4082-km2 study area was located in the northern portion of the Raton Basin in Costilla, Las Animas and Huerfano counties of south-central Colorado, and Colfax and Taos counties in northern New Mexico, USA. In these regions, land ownership is predominantly private, comprising ~89% of the area (Vitt 2007). Ranching, hunting, energy development and residential home development are the primary land use practices. The interior of the study area (1370 km2) encompassed pre-existing and expanding CBNG development, which contained 2421 well pads (1.77 well pads per km2) and 2933 wells (2.14 wells per km2) in 2009. Due to rapidly increasing development for energy, most animal capture was concentrated within the interior of the study area, which allowed annual changes in site fidelity to be assessed relative to development.
Topography ranged from rolling ridges and valleys to steep alpine slopes and cliffs (Vitt 2007) with elevation from 1800 to 4300 m. Mean annual precipitation ranged from 150 mm at lower elevations to 510 mm at higher elevations (Vitt 2007). We obtained site-specific temperature data from seven weather stations located across the study area at elevations ranging from 1983 to 2841 m. At the highest elevation weather station, minimum and maximum January and July temperatures were −26.3, 12.4, 3.2 and 26.4°C, respectively. At the lowest elevation weather station, minimum and maximum January and July temperatures were −25.2, 20.9, 7.1 and 33.8°C, respectively.
Capture and handling
We captured adult (≥2.5 years of age) female elk using a helicopter and either a dart gun or net gun annually during February and March 2006–09. Female elk (n = 165) were fitted with GPS collars programmed to collect one location every 3 h (eight locations per day) for one year. Eighteen elk were recaptured and refitted with GPS collars for one to two more years, giving us records of individual space use patterns over multiple years. Thus, the final sample size for this analysis was determined by survival (Webb et al. 2011) and recapture success. We estimated age of elk using tooth eruption, replacement and wear techniques (Quimby and Gaab 1957). The average age of females at the time of capture was 5.9 years (±0.4 s.e.). Animals captured using the net gun were manually restrained with hobbles and fitted with blindfolds to reduce stress. Darted elk were anaesthetised using either carfentanil or thiafentanil (A-3080), restrained with hobbles and fitted with blindfolds. Elk were fitted with a GPS collar (TGW-3590, Telonics, Inc., Mesa, AZ) and released at the site of capture; anaesthetised elk were released after reversal with naltrexone. Animal capture and handling protocols were approved by the Colorado Division of Wildlife (Permit No. 06TR1083, 07TR1083, 08TR1083 and 09TR1083A001).
Development and vegetation cover
We determined year-specific changes in the area of land surface developed from annual high-resolution aerial photography (0.3 m), and updated images of development as annual aerial photography became available. Each home range was attributed with development known to be present during that year. We delineated the following land developments: oil and gas well pads and ancillary facilities, residences, buildings, industries, ranching activities, roads, railroads and pipelines. We measured the width (w) for a subsample of linear features (i.e. roads, railroads and pipelines) to determine surface area developed. Roads were divided into five classes (1–5) to determine average width (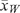 ) of development associated with each (Webb et al. 2011). Buffers were equal to 1/2 (). We set our buffer distances by rounding to the nearest 10 cm. Buffer distances (m) were 5.2, 4.0, 2.6, 1.8, 1.7 and 2.1 for Class 1, 2, 3, 4, and 5 roads and railroad tracks, respectively (Webb et al. 2011). Average sizes (ha ± s.e.) of land developments on the study area were 1.5 (±0.15) for ranching, 3.2 (±0.73) for industrial, 17.2 (±13.63) for community, 0.3 (±0.02) for residences and 0.5 (±0.01) for well pads. Land development increased by 10.6% from year 1 (3893.9 ha) to year 2 (4308.4 ha), 2.9% from year 2 to year 3 (4432.9 ha), and remained virtually unchanged from year 3 to year 4. Land development features were interpreted, digitised, and attributed based on annual aerial photography (2005–09). We used heads-up digitising of all visible polygon and linear surface features within our study area and performed all spatial analyses using ArcGIS® 9.3 software (ESRI, Inc., Redlands, CA). All land development features were merged into a single feature layer using the Union Overlay method in ArcGIS® 9.3.
) of development associated with each (Webb et al. 2011). Buffers were equal to 1/2 (). We set our buffer distances by rounding to the nearest 10 cm. Buffer distances (m) were 5.2, 4.0, 2.6, 1.8, 1.7 and 2.1 for Class 1, 2, 3, 4, and 5 roads and railroad tracks, respectively (Webb et al. 2011). Average sizes (ha ± s.e.) of land developments on the study area were 1.5 (±0.15) for ranching, 3.2 (±0.73) for industrial, 17.2 (±13.63) for community, 0.3 (±0.02) for residences and 0.5 (±0.01) for well pads. Land development increased by 10.6% from year 1 (3893.9 ha) to year 2 (4308.4 ha), 2.9% from year 2 to year 3 (4432.9 ha), and remained virtually unchanged from year 3 to year 4. Land development features were interpreted, digitised, and attributed based on annual aerial photography (2005–09). We used heads-up digitising of all visible polygon and linear surface features within our study area and performed all spatial analyses using ArcGIS® 9.3 software (ESRI, Inc., Redlands, CA). All land development features were merged into a single feature layer using the Union Overlay method in ArcGIS® 9.3.
Because forest cover is an important habitat requirement of ungulates, we developed a vegetation cover-type map to control for the effects of cover on range use size and fidelity relative to land development. We used high-resolution (0.3 m) true-colour and colour-infrared (CIR) aerial photography and Feature Analyst® 4.2 (FA; Visual Learning Systems, Inc., Missoula, MT) for ArcGIS® 9.3 (Visual Learning Systems, Inc. 2008). We conducted a supervised classification using delineated polygons of known vegetation type for use with object-based feature extraction algorithms. The true-colour and CIR bands were combined using FA, which resulted in four spectral bands (i.e. red, green, blue and near-infrared). We also specified that the green spectral band be used to develop a texture band. We used digital elevation models to develop an elevation band, which finally resulted in six bands (i.e. four spectral bands, one texture band and one elevation band). Last, we varied our resolution, or pixel, classifier pattern and size combinations based on vegetation type. Prior to running classifiers, we resampled vegetation cover types that occurred over extensive areas (i.e. dense forest, open forest, oak-dominated shrubland, alpine and grassland) to 3 m resolution and vegetation cover types that were more restricted or linear (i.e. riparian) to 1.5 m resolution. We used the Manhattan classifier pattern and a width of seven pixels to classify extensive vegetation types. For more restricted vegetation types we used the Bull’s Eye 2 classifier and a width of 15 pixels. For these analyses, we reclassified our six vegetation classes into either ‘cover’ or ‘non-cover’ habitat. We considered all classes as cover except alpine and grassland habitats.
Area of use metrics
We calculated the annual size of 50% core areas and 95% home ranges using adaptive-kernel estimators (Worton 1989) in Home Range Tools (Rodgers et al. 2005) for ArcGIS®. Annual core areas and home ranges were calculated from 16 March 2006 to 28 February 2007; 1 March 2007 to 28 February 2008; 1 March 2008 to 28 February 2009; and 1 March 2009 to 31 December 2009. We used unit-variance standardisations and the biased cross validation smoothing parameter when calculating volume-probability polygons. To assess annual fidelity of core areas and home ranges, we calculated annual overlap in core areas and home ranges, reported here as percentage of overlap. Overlap area of two successive annual core areas or home ranges was calculated, divided by the average size of the two core areas or home ranges, and multiplied by 100 to determine percentage of overlap. We used estimates of annual ranges in lieu of seasonal ranges for several reasons. First, we observed that most elk used for these analyses did not make long distance movements between disjunct summer and winter ranges, rather animals gradually moved between higher and lower elevations. Second, the general patterns of space use captured by annual ranges also included both summer and winter range space use. Last, we were not interested in testing hypotheses related to seasonal space use patterns because we were only interested in broad temporal scales and patterns associated with landscape-level development.
Data analysis
To assess how development influences range use dynamics, we calculated the area (ha) of cover and development within each core area and home range using Hawth’s Tools and the Polygon in Polygon Analysis (PIPA). Some home ranges extended beyond the extent of our cover (n = 4) and land development (n = 5) maps; thus requiring that we account for this disparity in our analyses. We subtracted the amount of home range area outside the extent of our cover or development map from the original home range size and used the amount of cover within the newly calculated home range area for statistical analyses. However, we used actual home ranges when reporting sizes and overlap. Less than 3% of the home range area was outside the extent of our cover and development map.
Statistical analysis
We used linear regression (LR) models to determine whether the proportion of development influenced core area and home range size. We also used LR to assess how changes in proportion of development influenced core area and home range overlap. We calculated the change in proportion of development by determining the difference in proportions between successive year core areas and home ranges. Negative values represented a decrease in the amount of development in the second year, whereas positive values indicated greater development in the second year. We used an arcsine transformation because our dependent variable (i.e. overlap) was a proportion. For the aforementioned LR models, we controlled for the proportion of cover by incorporating cover as a covariate into our models. We used normal probability plots to ascertain whether assumptions of regression models were satisfied. All statistical analyses were conducted using SAS® 9.2 (SAS Institute Inc., Cary, NC).
Results
We analysed data on 18 female elk that were fitted with GPS collars for ≥2 years. Only one elk was fitted with a GPS collar for three years. Thus, we were able to analyse core area and home range overlap for 19 between-year comparisons using 37 annual core areas and home ranges. On average, GPS collars recorded 2321 (±471 s.d.) locations per elk per year.
Home range dynamics
Average annual home range size was 249.5 km2 (±27.8 s.e.) and between year home range overlap was 67.9% (±4.4 s.e.). Only one female relocated her home range from one year to the next, resulting in <1% overlap of the two annual home ranges. Home range overlap averaged 71.6% (±2.4 s.e.) when this particular female was excluded from the analysis. On average, 87% (±1.0 s.e.; range: 78.1–95.8%) of the home range area comprised cover but only 2.8% (±0.2 s.e.; range: 1.2–4.8%) of the total home range area comprised land development.
We further analysed how the proportion of development within home ranges influenced home range size and overlap. Development negatively influenced home range size (R2 = 0.524; t33 = −4.98; P < 0.001) and overlap (R2 = 0.534; t15 = −2.69; P = 0.016). Home range size (β = −11 285.1 ± 2 264.6 s.e.) and overlap (β = −15.39 ± 5.71 s.e.) decreased as the proportion of development increased.
Core area dynamics
Annual core area size and overlap were calculated for the same 18 individual elk. Size of core areas averaged 39.0 km2 (±4.5 s.e.) and overlap between years was 48.6% (±4.0 s.e.). Core areas did not overlap for one female because this particular female relocated her home range during the second year. When this female was excluded from analysis, core area overlap averaged 51.3% (±3.1 s.e.). The percentage of cover and development in core areas was 82.5% (±2.1 s.e.; range: 51.9–97.6%) and 2.8% (±0.3 s.e.; range: 0.5–10.1%), respectively.
Similarly to home ranges, core area size was negatively influenced (β = −79 309 ± 24 812 s.e.) by the proportion of development (R2 = 0.369; t33 = −3.2; P = 0.003). Proportion of development also negatively influenced overlap of core areas (β = −0.29 ± 2.89 s.e.), although not significantly (R2 = 0.12; t15 = −0.1; P = 0.923).
Discussion
Female elk exhibited high levels of site fidelity (i.e. core area and home range) even with increasing levels of development. Strong seasonal and annual site fidelity among female elk is common (Rudd et al. 1983; Edge et al. 1985, 1986; Irwin 2002; Raedeke et al. 2002) because fidelity likely carries advantages including knowledge of forage resources and security cover (Edge et al. 1985; Tracz et al. 2010). Although site fidelity was high in this population of elk, it showed a tendency to decrease as development increased. This finding is similar to previous studies on ungulates that found site fidelity can remain high but is reduced as development increases. For example, site fidelity was reduced by development of a hydro-electric dam (Andersen 1991), military manoeuvres (Andersen et al. 1996; Stephenson et al. 1996), logging (Edge et al. 1985; Linnell and Andersen 1995), ranching operations (Hood and Inglis 1974), simulated mining (Kuck et al. 1985), and installation of oil wells (Van Dyke and Klein 1996). However, boreal woodland caribou (Rangifer tarandus caribou) showed strong fidelity to home ranges even with increasing petroleum development (Tracz et al. 2010).
Increasing levels of development may cause animals to abandon traditional home ranges (Peek et al. 1982). However, two factors may alleviate the necessity to abandon established ranges. First, animals may not abandon home ranges if they are able to exploit undeveloped areas of their home range (Hershey and Leege 1982). Second, forest cover mitigates some of the effects development and human activity have on animal behaviour (Edge et al. 1985) because animals are able to retreat to safe environments. The aforementioned factors were supported by our data; elk established areas with high proportions of cover (87% within home ranges and 82.5% within core areas) and shifted areas of concentrated use within their home ranges (i.e. overlap areas) to areas with minimal development. However, some areas with cover (e.g. open-canopy forest) may not have functioned solely as security cover, but could have simultaneously provided foraging resources.
Based on these data and data from previous studies, ungulates appear to respond spatially to development in two different ways. First, animals use larger areas in relation to development, which allows them to seek new areas with less development and less human activity. For example, home range size increased for elk in relation to increased logging (Pedersen et al. 1980 cited in Andersen 1991), moose (Alces alces) in relation to military disturbance (Andersen et al. 1996), elk calves that were disturbed by simulated mining (Kuck et al. 1985), and mule deer (Odocoileus hemionus) in response to military manoeuvres (Stephenson et al. 1996). Second, animals use small areas further from development more intensively. Our data support the second response strategy because development negatively influenced core area and home range size. Edge et al. (1985) observed a similar response in elk; home range size decreased as development associated with logging increased. Thus, elk may minimise the extent of their movements when development is present to minimise contact with human activity. Companion studies (Dzialak et al. 2011; Harju et al. 2011) identified similar responses of elk using resource selection functions; elk selected for cover but tended to avoid areas of development. Thus, all available data suggest that, although development is present in areas used by elk, they prefer to use areas of cover away from development more intensively.
Female elk showed strong fidelity to annual ranges even in the face of increasing development and human activity. Tolerating development below a critical threshold (Linnell and Andersen 1995) likely carries advantages to individuals exhibiting strong site fidelity (Edge et al. 1985; Tracz et al. 2010). Despite relatively high levels of site fidelity, females did redistribute their home ranges to areas with fewer developments from one year to the next, but still in the same general vicinity as the previous year. This may be one adaptation of elk to maintain fidelity to annual or seasonal ranges when faced with increasing levels of development. Other patterns of adaptation also have arisen in this same segment of the population. For example, in the presence of human activity, female elk seek areas of forest cover away from human developments during the day, but at night exploit areas nearer to developments because human activity is reduced (Dzialak et al. 2011).
It is apparent from these data that female elk in this population are unlikely to abandon established home range areas as long as development remains below tolerance thresholds. However, elk may continue to alter their home range use patterns relative to development within the areas they previously inhabited. The population of elk observed in this study may be less sensitive to development because multiple forms of development have been occurring since the mid-1980s; thus, animals may have had time to adapt to these developments. Measures should be taken to minimise the amount of development and human activity during sensitive seasons (e.g. calving and winter) and within areas used by a large percentage of the population. To help mitigate the effects of development on elk, large areas of forest should be maintained and managed because elk use areas with greater forest cover as development increased. Further, it may be beneficial to consider topography when planning placement of roads, buildings, well pads or other infrastructure to reduce line-of-sight visibility between wildlife and development (Edge and Marcum 1991). Strong fidelity to traditional ranges, coupled with the observation that females shifted home ranges to minimise interaction with human activity, suggest that established home ranges represent critical habitat within which land development should be constrained to the greatest extent feasible.
Acknowledgements
We thank Pioneer Natural Resources Co. for funding our research; Colorado Division of Wildlife for project support; landowners in the Raton Basin for participation and access to land; R. G. Osborn for project assistance; J. Mudd for assistance using Feature Analyst; and A. M. Foley, C. Hedley, and several anonymous reviewers for helpful comments on earlier drafts.
References
Andersen, R. (1991). Habitat changes in moose ranges: effects on migratory behavior, site fidelity and size of summer home-range. Alces (Thunder Bay, Ont.) 27, 85–92.Andersen, R., Linnell, J. D. C., and Langvatn, R. (1996). Short term behavioural and physiological response of moose Alces alces to military disturbance in Norway. Biological Conservation 77, 169–176.
| Short term behavioural and physiological response of moose Alces alces to military disturbance in Norway.Crossref | GoogleScholarGoogle Scholar |
Anderson, D. P., Forester, J. D., Turner, M. G., Frair, J. L., Merrill, E. H., Fortin, D., Mao, J. S., and Boyce, M. S. (2005). Factors influencing female home range sizes in elk (Cervus elaphus) in North American landscapes. Landscape Ecology 20, 257–271.
| Factors influencing female home range sizes in elk (Cervus elaphus) in North American landscapes.Crossref | GoogleScholarGoogle Scholar |
Bureau of Land Management (2005). ‘Oil and Gas Activity on Public Lands.’ (U.S. Department of the Interior, Bureau of Land Management: Cheyenne, WY.)
Dzialak, M. R., Harju, S. M., Osborn, R. G., Wondzell, J. J., Hayden-Wing, L. D., Winstead, J. B., and Webb, S. L. (2011). Prioritizing conservation of ungulate calving resources in multiple-use landscapes. PLoS ONE 6, e14597.
| Prioritizing conservation of ungulate calving resources in multiple-use landscapes.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3MXhvFahu7s%3D&md5=ea07d2bad56b13766c82a8068d2d781aCAS | 21297866PubMed |
Edge, W. D., and Marcum, C. L. (1991). Topography ameliorates the effects of roads and human disturbance on elk. In ‘Proceedings of a Symposium on Elk Vulnerability’. (Eds A. G. Christensen, L. J. Lyon and T. N. Lonner.) pp. 132–137. (Montana State University: Bozeman, MT.)
Edge, W. D., Marcum, C. L., and Olson, S. L. (1985). Effects of logging activities on home-range fidelity of elk. The Journal of Wildlife Management 49, 741–744.
| Effects of logging activities on home-range fidelity of elk.Crossref | GoogleScholarGoogle Scholar |
Edge, W. D., Marcum, C. L., Olson, S. L., and Lehmkuhl, J. F. (1986). Nonmigratory cow elk herd ranges as management units. The Journal of Wildlife Management 50, 660–663.
| Nonmigratory cow elk herd ranges as management units.Crossref | GoogleScholarGoogle Scholar |
Felix, A. B., Walsh, D. P., Hughey, B. D., Campa, H., and Winterstein, S. R. (2007). Applying landscape-scale habitat-potential models to understand deer spatial structure and movement patterns. The Journal of Wildlife Management 71, 804–810.
| Applying landscape-scale habitat-potential models to understand deer spatial structure and movement patterns.Crossref | GoogleScholarGoogle Scholar |
Harju, S. M., Dzialak, M. R., Osborn, R. G., Hayden-Wing, L. D., and Winstead, J. B. (2011). Conservation planning using resource selection models: altered selection in the presence of human activity changes spatial prediction of resource use. Animal Conservation 14, .
Hershey, T. J., and Leege, T. A. (1982). ‘Elk Movements and Habitat Use on a Managed Forest in North-central Idaho.’ Wildlife Bulletin 10. (Idaho Department of Fish and Game: Boise, ID.)
Hood, R. E., and Inglis, J. M. (1974). Behavioral responses of white-tailed deer to intensive ranching operations. The Journal of Wildlife Management 38, 488–498.
| Behavioral responses of white-tailed deer to intensive ranching operations.Crossref | GoogleScholarGoogle Scholar |
Irwin, L. L. (2002). Migration. In ‘North American Elk: Ecology and Management’. (Eds D. E. Toweill and J. W. Thomas.) pp. 493–513. (Smithsonian Institution Press: Washington DC, WA.)
Kuck, L., Hompland, G. L., and Merrill, E. H. (1985). Elk calf response to simulated mine disturbance in southeast Idaho. The Journal of Wildlife Management 49, 751–757.
| Elk calf response to simulated mine disturbance in southeast Idaho.Crossref | GoogleScholarGoogle Scholar |
Linnell, J. D. C., and Andersen, R. (1995). Site tenacity in roe deer: short-term effects of logging. Wildlife Society Bulletin 23, 31–35.
McCoy, J. E., Hewitt, D. G., and Bryant, F. C. (2005). Dispersal by yearling male white-tailed deer and implications for management. The Journal of Wildlife Management 69, 366–376.
| Dispersal by yearling male white-tailed deer and implications for management.Crossref | GoogleScholarGoogle Scholar |
Pedersen, R. J., Adams, A. W., and Skovlin, J. M. (1980). ‘Elk Habitat Use in an Unlogged and Logged Forest Environment.’ Wildlife Research Report 9. (Oregon Department of Fish and Wildlife: Portland, OR.)
Peek, J. M., Scott, M. D., Nelson, L. J., Pierce, D. J., and Irwin, L. L. (1982). Role of cover in habitat management for big game in northwestern United States. Transactions of the North American Wildlife and Natural Resources Conference 47, 363–373.
Quimby, D. C., and Gaab, J. E. (1957). Mandibular dentition as an age indicator in Rocky Mountain elk. The Journal of Wildlife Management 21, 435–451.
| Mandibular dentition as an age indicator in Rocky Mountain elk.Crossref | GoogleScholarGoogle Scholar |
Raedeke, K. J., Millspaugh, J. J., and Clark, P. E. (2002). Population characteristics. In ‘North American Elk: Ecology and Management’. (Eds D. E. Toweill and J. W. Thomas.) pp. 449–491. (Smithsonian Institution Press: Washington DC, WA.)
Rodgers, A. R., Carr, A. P., Smith, L., and Kie, J. G. (2005). ‘HRT: Home Range Tools for ArcGIS.’ (Ontario Ministry of Natural Resources, Centre for Northern Forest Ecosystem Research: Thunder Bay, Ontario.)
Rudd, W. J., Ward, A. L., and Irwin, L. L. (1983). Do split hunting seasons influence elk migrations from Yellowstone National Park? Wildlife Society Bulletin 11, 328–331.
Rumble, M. A., Benkobi, L., and Gamo, R. S. (2005). Elk responses to humans in a densely roaded area. Intermountain Journal of Sciences 11, 10–24.
Stephenson, T. R., Vaughan, M. R., and Andersen, D. E. (1996). Mule deer movements in response to military activity in southeast Colorado. The Journal of Wildlife Management 60, 777–787.
| Mule deer movements in response to military activity in southeast Colorado.Crossref | GoogleScholarGoogle Scholar |
Tracz, B. V., LaMontagne, J. M., Bayne, E. M., and Boutin, S. (2010). Annual and monthly range fidelity of female boreal woodland caribou in response to petroleum development. Rangifer 30, 31–44.
Unsworth, J. W., Kuck, L., Garton, E. O., and Butterfield, B. R. (1998). Elk habitat selection on the Clearwater National Forest, Idaho. The Journal of Wildlife Management 62, 1255–1263.
| Elk habitat selection on the Clearwater National Forest, Idaho.Crossref | GoogleScholarGoogle Scholar |
Van Dyke, F., and Klein, W. C. (1996). Response of elk to installation of oil wells. Journal of Mammalogy 77, 1028–1041.
| Response of elk to installation of oil wells.Crossref | GoogleScholarGoogle Scholar |
Visual Learning Systems, Inc. (2008). ‘Feature Analyst® 4.2 for ArcGIS® Reference Manual.’ (Visual Learning Systems, Inc.: Missoula, MT.)
Vitt, A. (2007). ‘Trinchera Data Analysis Unit E-33. Game Management Units 83, 85, 140, 851. Elk Management Plan.’ (Colorado Division of Wildlife: Pueblo, CO.)
Walter, W. D., VerCauteren, K. C., Campa, H., Clark, W. R., Fischer, J. W., Hygnstrom, S. E., Mathews, N. E., Nielson, C. K., Schauber, E. M., Van Deelen, T. R., and Winterstein, S. R. (2009). Regional assessment on influence of landscape configuration and connectivity on range size of white-tailed deer. Landscape Ecology 24, 1405–1420.
| Regional assessment on influence of landscape configuration and connectivity on range size of white-tailed deer.Crossref | GoogleScholarGoogle Scholar |
Webb, S. L., Hewitt, D. G., and Hellickson, M. W. (2007). Scale of management for mature male white-tailed deer as influenced by home range and movements. The Journal of Wildlife Management 71, 1507–1512.
| Scale of management for mature male white-tailed deer as influenced by home range and movements.Crossref | GoogleScholarGoogle Scholar |
Webb, S. L., Dzialak, M. R., Wondzell, J. J., Harju, S. M., Hayden-Wing, L. D., and Winstead, J. B. (2011). Survival and cause-specific mortality of female Rocky Mountain elk exposed to human activity. Population Ecology 53, .
| Survival and cause-specific mortality of female Rocky Mountain elk exposed to human activity.Crossref | GoogleScholarGoogle Scholar |
Worton, B. J. (1989). Kernel methods for estimating the utilization distribution in home-range studies. Ecology 70, 164–168.
| Kernel methods for estimating the utilization distribution in home-range studies.Crossref | GoogleScholarGoogle Scholar |