

Challenges of feeding dairy cows in Australia and New Zealand
W. J. Wales A C and E. S. Kolver BA Department of Economic Development, Jobs, Transport and Resources, Ellinbank, Vic. 3821, Australia.
B DairyNZ, Private Bag 3221, Hamilton 3240, New Zealand.
C Corresponding author. Email: bill.wales@ecodev.vic.gov.au
Animal Production Science 57(7) 1366-1383 https://doi.org/10.1071/AN16828
Submitted: 21 December 2016 Accepted: 5 April 2017 Published: 19 May 2017
Journal Compilation © CSIRO Publishing 2017 Open Access CC BY-NC-ND
Abstract
There is a continuing evolution of feeding systems in both Australian and New Zealand dairy industries and this presents challenges for the future. Since the turn of the century, the two countries have diverged in industry growth characteristics, with Australian dairying having contracted, with 10% less milk being produced because of 20% fewer cows producing 15% more per cow, whereas New Zealand dairying has expanded, producing 83% more milk driven by a 54% increase in cow numbers and a 31% increase in milk production per cow. Solutions to optimise feed efficiency included the common themes of (1) growing more forage on farm, (2) increasing its utilisation and (3) more efficient use of supplements resulting in increases in DM intake, and they remain relevant. In New Zealand, many of the recent research activities have aimed at improving feed supply while limiting environmental impacts driven by increasing societal concern surrounding the environmental footprint of a growing and intensifying agricultural sector. In Australia, many of the recent research activities have aimed at improving feed efficiency, with a focus on understanding situations where partial mixed ration feeding systems (Australian Farm Systems 3 and 4) are sustainable. Simply growing more feed on farm can no longer be a sole objective; farms must be operated with a view to reduce the environmental footprint, with New Zealand dairy farmers increasingly needing to farm within nitrogen limits. The present review revisits and reinforces many of the concepts developed in previous reviews, but also examines the evolution of feeding systems in both countries and opportunities to improve feed efficiency and profit, while satisfying public expectations around environmental stewardship. We also identify some of the gaps in the current knowledge that warrant further research.
Additional keywords: feeding systems, grazing, intake, milk response.
Introduction
There is a continuing evolution of feeding systems in both Australia and New Zealand dairy industries and this presents challenges for the future. Several reviews have explored these challenges. For example, Thorrold and Doyle (2007) presented a snapshot of the current and future state of dairy farming in New Zealand and Australia. This was followed up by Clark et al. (2013), who focussed on drivers of dairy production in both countries. The present review revisits and reinforces many of the concepts developed in the previous work, but also looks at the evolution in feeding systems in both countries and opportunities to improve feed efficiency and profit, while satisfying public expectations around environmental stewardship. We also identify some of the gaps in the current knowledge that warrant further research.
Evolution of farm and feeding systems since the 1980s
National
Dairying in Australia and New Zealand has undergone a significant structural change over the past three decades, summarised by declines in the number of farms and an increase in total milk produced (Table 1). However, since the turn of the century, the two countries have diverged in industry growth characteristics. Australian dairying has contracted, with 10% less milk being produced because of 20% fewer cows producing 15% more per cow (L/cow; Dairy Australia 2015a). Over a similar period, New Zealand dairying has dramatically expanded, producing 83% more milk, driven by a 54% increase in cow numbers and a 31% increase in milk production per cow (kg fat+protein)/cow; DairyNZ 2015). Both countries have seen the development of fewer, larger herds, although the effect has arguably been greater in Australia where there were fewer than half the number of dairy farms operating in 2014–2015 than there were 15 years earlier. In New Zealand, the expansion of dairying into Canterbury and Southland has accelerated the increase in cow numbers and herd size.
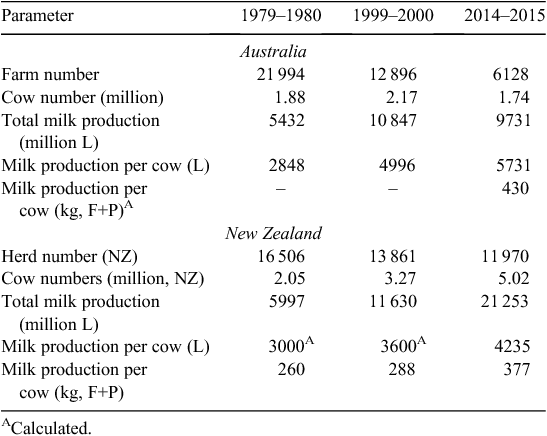
|
Farm systems
During the past 35 years, farming systems in both countries have intensified in terms of stocking rate and the proportion of imported supplementary feed used on-farm, and a range of farming systems now exists in Australia and New Zealand. Both countries have attempted to describe the characteristics of these farm systems on the basis of differences in key feed inputs, although it is noted that these categories are somewhat arbitrary as there is a continuum of feed combinations in practice (Dairy Australia 2015b; DairyNZ 2016a). The five farm-system categories that have been developed in each country define the timing, purpose and amount of grazing and supplementary feed use, ranging from grazing only and little use of supplementary feed to zero grazing and a high reliance on purchased feeds. The range in systems in Australia is broader than that in New Zealand, with New Zealand categories restricted to approximately the first three systems defined for Australia. The five Australian feeding systems are as follows:
-
System 1: low bail system, including grazed pasture + other forages + up to 1.0 t/cow.year of concentrates fed in the dairy shed during milking;
-
System 2: moderate-high bail system, including grazed pasture + other forages + more than 1 t/cow.year concentrates fed in the dairy shed during milking;
-
System 3: partial mixed ration (PMR) system, including pasture grazed for all year + PMR on feed pad ± concentrates fed in the dairy shed;
-
System 4: hybrid system, including pasture grazed for a period of less than 9 months per year + PMR on feed pad ± concentrates fed in the dairy shed;
-
System 5: total mixed ration (TMR) system, i.e. zero grazing, where cows are housed and fed a total mixed ration.
The five New Zealand feeding systems are as follows:
-
System 1: all grass self-contained, all stock on the dairy platform;
-
System 2: ~4–14% of total feed is imported, either supplement or grazing off, and fed to dry cows;
-
System 3: ~10–20% of total feed is imported to extend lactation (typically autumn feed) and for dry cows;
-
System 4: ~20–30% of total feed is imported and used at both ends of lactation and for dry cows;
-
System 5: at least 30% of total feed is imported and used all year, throughout lactation and for dry cows.
In the most recent survey conducted during the 2014–2015 season, Dairy Australia (2015b) reported that 23% of farms across Australia employed feeding systems where less than 1 t grain/cow per year was fed (Australian System 1), representing a 5% unit decline from the 2010–2011 survey (Dairy Australia 2011). In the current survey, 64% of dairy farmers fed moderate to high amounts of concentrate in the dairy (>1 t/cow per year; Australian System 2); an increase of 10% units compared with the results from the 2010–2011 survey. Farms where grazed pasture was supplemented with PMR offered on a feed pad (Australian Systems 3 and 4) comprised 12% of dairy farms and 1% of farms employed systems characterised as Australian System 5 (TMR).
Farms identified as being in Australian Systems 1 and 2 have increased from 82% to 87% nationally in the 4 years up until 2014–2015, with a concomitant reduction from 18% to 13% in farms employing Systems 3–5. This reflects improvements in home-grown forage supply due to better seasonal conditions, meaning that costs associated with feeding supplements with a mixer wagon could be reduced or avoided.
At a regional level in Australia, ~10% of dairy farms in Tasmania and Gippsland in Victoria do not feed any grain, which contrasts with the remaining regions where this number is less than 4%. Approximately 20% of dairy farms in each of the regions of Queensland, New South Wales, Western Australia and the Murray region of northern Victoria–southern New South Wales use PMR and hybrid systems compared with the other regions, where this percentage is less than 10% (Dairy Australia 2015b). Much of the recent nutrition research has focussed on these regions that use more PMR.
In New Zealand, over the same period (2010–2011 to 2014–2015), the proportion of herds in low-input systems (Systems 1 and 2) reduced from 42% to 32%, was maintained at 39% in medium-input systems (System 3), and increased from 19% to 29% in high-input systems (Systems 4 and 5; DairyNZ 2016a). New Zealand farm systems are now broadly represented across low-, medium- and high-input classifications. Farmers have intensified their systems through increased supplement use and higher stocking rates in response to high milk and land prices, dry seasonal conditions, increased feeding infrastructure on farms, and because of dairy expansion in the South Island, which requires off-farm winter grazing. Conversely, many farmers during the 2015–2016 and 2016–2017 season reviewed their farm systems and adjusted to the low milk prices by reducing stocking rates, keeping more young stock and wintered cows on the milking platform as well as reducing expenditure on harvested and imported supplementary feeds. It is unclear at this stage, whether these adjustments will be sustained in the future or whether this is a short-term adjustment. Also driving the growth of higher-input systems has been the expansion of dairying in the South Island, which requires more off-farm winter grazing.
At a national level, this intensification has resulted in broadly similar or improved profitability. The average financial performance of these low-, medium- and high-input systems over the past 10 years (2005–2006 to 2014–2015) shows that profitability for low, medium, high systems has been $NZ1823/ha (range $NZ871–2905/ha), $NZ2004/ha ($NZ814–3363/ha) and $NZ2202/ha ($NZ228–3797/ha) respectively; Return on assets has been 6.6% (–4.9–14.8%), 6.6% (–7.0–16.2%), and 7.4% (–7.8–18.7%) respectively; equity growth has been 6.4% (–12.9–17.7%), 6.9% (–14.4–19.3%) and 8.5% (–15.0–25.2%) per annum; and closing term liabilities (as a measure of risk) have been $NZ19.4/kg milk fat + protein (F+P; $NZ12.9–23.2/kg F+P), $NZ18.8/kg F+P ($NZ13.5–21.5/kg F+P), and $NZ18.8/kg F+P ($NZ14.5–21.3/kg F+P) (from DairyNZ 2008, 2010, 2012, 2014, 2016a; note return on assets and equity growth not reported for 2005–2006). At a national level, this implies that a profit response to the additional feed eaten has resulted from the on-farm changes that have occurred during the past 10 years; however, there is a wide range around these average 10-year returns. In particular, returns from the more intensive systems have been more volatile than those produced from lower-input systems. The milk price environment within which these returns were achieved was representative of the range of expected milk prices, averaging $NZ6.09/kg F+P ($NZ4.14–7.69/kg F+P).
Feeds
The evolution in feeding systems from farms dominated by homegrown forage only, to those relying on more concentrates and imported feeds, has been greatest in Australian systems, which now average 1.6 t DM/cow annually, having increased from 0.7 t DM/cow 25 years ago (Dharma et al. 2012). These changes have occurred due to the relatively low cost of cereal grain compared with other energy-based supplements and motivation to increase production per cow and per hectare (Jacobs 2014).
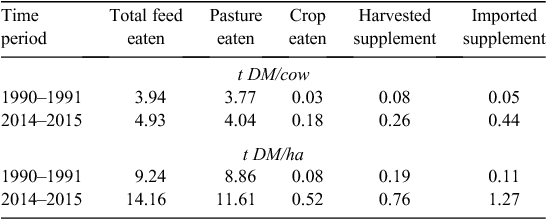
In New Zealand, despite the increased use of supplementary feed, pasture continues to be the dominant feed offered to cows (82% of the diet). The total amount of feed eaten by dairy cows at a national level has increased 161% over the past 25 years; a growth rate of 3.8% (compound annual growth rate, CAGR; DairyNZ 2016b; Table 2). This increase has occurred primarily (80%) through more cows (5.02 vs 2.4M cows), from both new dairy land and increased stocking rates. On a per cow basis, feed eaten has increased 1 t (0.8% CAGR) to 4.93 t DM/cow. Imported supplements, including palm-kernel extract (Elaeis guineensis), increased 7.7% CAGR, harvested supplement including maize (Zea mays) silage and barley (Hordeum vulgare) increased 6.2% CAGR, while grown crops including fodder beet (Beta vulgaris), kale (Brassica oleracea) and swedes (Brassica napobrassica) increased 4.9% CAGR between 1990–1991 and 2014–2015.
|
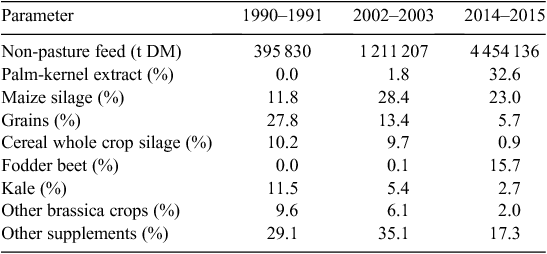
Non-pasture feeds consumed by dairy cows increased from 0.16 t DM/cow in 1990–1991 to 0.89 t DM/cow in 2014–2015. As a proportion of a cow’s total diet, non-pasture feeds increased from 4.2% in 1990–1991 to 18.0% in 2014–2015, being made up of harvested crop, imported supplements and grazed crop. The amount of pasture eaten per cow increased in the 1990s, but has eased during the 2000s. However, pasture eaten per hectare has increased at a rate of 1.7% per year to 2006–2007, but growth has slowed to a rate of 0.7% since.
Feed eaten per hectare has increased 53% (+1.7% CAGR) or 4.9 t DM/ha from 1990–1991 to 2014–2015. This is a faster increase than a per cow basis as it includes an increase in stocking rate. Pasture eaten accounted for just over half the increase in feed eaten per hectare, while imported supplements (+1.2 t DM/ha), harvested supplement (+0.6 t DM/ha) and grown crops (+0.4 t DM/ha) have all increased over the past 25 years.
Compared with Australia, greater use has been made in New Zealand of palm-kernel extract, maize silage and homegrown crops as grain and concentrate supplements are comparatively more expensive. The increased use of maize silage since the 1990s has driven the growth in non-pasture feed, as has palm-kernel extract since the drought of 2008 (Table 3). More recently, the rapid increase in fodder beet use in winter, and increasingly in summer, has displaced kale and other brassica crops.
|
Feed conversion efficiency
The combined effects of cow genetic gain over the past 25 years, improved farm management, longer lactations, improved pastures and increased use of supplementary feed have increased the feed conversion efficiency of the New Zealand national herd. The national herd in 2014–2015 produced more milk F+P as a percentage of liveweight (82% vs 52% of liveweight), produced more milk fat+protein from each tonne of DM (76 vs. 62 kg MS/t DM eaten) and required less feed to produce a kilogram of milk fat+protein (13 vs 16 kg DM/kg MS) than it did in 1990–1991 (DairyNZ 2016b).
The differences between Australian and New Zealand dairying systems reflect slightly different economic drivers. As an export nation subject to large year-to-year commodity-price fluctuations, New Zealand has evolved low to moderate production cost systems where pasture makes up the majority (82%) of the diet. Although subject to similar commodity volatility, tempered somewhat by a larger domestic dairy market, Australia has evolved a wider range of farm systems that include higher feed-input systems, which have more of a focus on per cow production and efficiency. This has been a result of the domestic production of low-cost cereal grains along with volatility of climate and availability of water for forage production. These differences have meant that, in recent years, the focus of feeding research in New Zealand has largely been on growing and utilising homegrown forage within anticipated future environmental constraints, while Australia has focussed on improving the efficiency of responses to feeds imported into the farm system.
Profitability in dairy production systems in both countries has in the past, and is still, dependent on efficient use of the farm resources that include cows, land and water, labour, capital and knowledge (Beever and Doyle 2007). As milk production has increased through improvements in genetic merit, cows require more digestible nutrients to avoid excessive body condition loss and ensure appropriate reproductive performance and longevity. This can be achieved through the provision of a well formulated TMR (Kolver and Muller 1998; Bargo et al. 2002); however, the challenge remains of how to increase nutrient intake in systems where pasture makes up a large part of the diet. It is well established that grazing cows consume less than their potential intake, due to factors listed under sward, animal and management factors (Wales et al. 2005; Beever and Doyle 2007). Overcoming the seasonality of pasture growth and of nutritive characteristics also continues to be a challenge for both countries (Chapman et al. 2009).
Limitations within current feeding systems
In 2005, Garcia and Fulkerson reviewed the opportunities for improvement in the Australian dairy industry, following 10 years of only modest increases in total factor productivity. Their analysis identified below-optimal feed efficiency as a key issue affecting profitability of dairy farmers into the future. The solutions to optimise feed efficiency included the common themes of (1) growing more forage on farm, (2) increasing its utilisation and (3) more efficient use of supplements. These solutions result in increases in DM intake and remain relevant to both Australia and New Zealand.
Dairy cows approach their genetic potential for DM intake and milk production on TMR feeding systems. For example, Kolver and Muller (1998) reported that cows in early lactation consuming TMRs produce 44 kg milk/day, compared with 30 kg milk/day by similar cows grazing pasture without supplements. This increase in milk production was a result of nutrient intake approaching a maximum through a combination of greater DM intake and a ration formulated to provide a more optimal mix of nutrients to meet requirements. This raises the question as to what factors are limiting milk production in our grazing systems and what can be done to remove these limitations?.
Milk production of cows relying solely on grazed herbage is constrained to less than 30 kg/day (~2.3 kg milk fat+protein/day) by seasonal variations in pasture growth and nutritive characteristics (Doyle et al. 2001). A key limitation is the quantity (both under and over supply) of nutrients supplied from grazed pasture, particularly in the more variable Australian environment. Although temperate pasture can provide a high-quality, balanced feed for moderate-yielding cows (Hills et al. 2015), supplementary feeds are used in grazing systems as a grazing management tool, when the supply of pasture is insufficient for herd demand, or when pasture does not supply adequate nutrition to support high production, as is the case on many Australian farms.
A key factor influencing the nutrient supply is amount of pasture available for grazing in each season. The seasonal growth or net pasture accumulation rate of pasture varies considerably in all dairy regions in both countries, with no 2 years following the same pattern of DM supply (Chapman et al. 2009). Figure 1, adapted from Cullen et al. (2008), highlights this seasonal variation and is a result of variations in local climatic conditions, nutrient supply, pasture species, access to irrigation water and management practices. Overcoming this seasonal variation in nutrient supply from pasture remains an important challenge. In addition to variability in DM supply, the nutritive characteristics of the component of the sward consumed by grazing cows is also affected by climatic, soil and plant factors due to such things as adequacy of fertiliser and water, and species and growth stage of the plant. Typically, the metabolisable energy (ME) and crude protein concentrations of the sward decline through spring, reaches nadir during summer and increases in autumn. There are also regional variations in the nutritive characteristics (Doyle et al. 2001).

|
Conceptually, the ideal sward would have characteristics, nutrient profile and physical attributes similar to a TMR formulated to provide nutrients in relation to requirements, while having physical characteristics necessary to maintain a stable rumen environment. However, grazed pasture never provides the mix of nutrients provided in a well formulated TMR. For example, the nutritive characteristics of a mixed grass–clover sward compared with a TMR of similar digestibility, had lower concentrations of non-structural carbohydrates and higher concentrations of neutral detergent fibre, in addition to the physical differences of longer particle sizes and lower % DM (Kolver and Muller 1998). Cows grazing this type of sward also consumed less DM, starch and ME, and more crude protein and water, due to the low DM concentration of the herbage. In comparison with TMR, high-producing cows consuming only grazed pasture had lower DM intake, milk production, milk protein and fat concentrations, lost more body condition and had lower liveweight (Kolver and Muller 1998; Bargo et al. 2002). Differences in milk production between pasture and TMR was predominantly driven by the lower DM intake of pasture, rather than differences in the proportion or digestibility of individual nutrients. At grazing, DM intake would need to approach 5% of a cow’s bodyweight to achieve intakes similar to those observed for cows consuming the TMR. However, Kolver and Muller (1998) concluded that current pasture species are unlikely to provide nutrients in sufficient quantities to achieve milk yields similar to those in cows consuming TMR. As forages are unlikely to provide the ideal mix of nutrients, identifying profitable supplementation strategies will continue to be important.
Factors limiting intake of grazing cows
In all grazing systems, herbage allowance has the biggest effect on DM intake with characteristics of the sward, such as pasture mass, spatial distribution of plants and their nutritive characteristics also being important (Wales et al. 2005). Additional nutrients are commonly provided by cereal supplements around milking time, with further supplements of pasture silages and hays and maize silage/palm-kernel extract. These types of feeding systems have intake limited to ~4% of liveweight, equivalent to 18 and 24 kg DM for a 450- and 600-kg cow respectively (Doyle et al. 2001). The genetic potential for intake under ideal feeding management may be 5% of liveweight, equivalent to intakes of 23 and 30 kg DM for a 450- and 600-kg cow respectively. Several factors are purported to limit intake in grazing systems and overcoming these limitations remains a key challenge for our industries.
The concept of rumen fill is often brought up as limiting intake, due to breakdown of structural carbohydrates, with neutral detergent fibre (NDF) being rate limiting (Mertens 1987). This concept has been useful when applied to TMR fed to appetite; however, the regulation of intake in grazing systems is more complex because of interactions between signals generated by supply of energy and essential nutrients from the digestive tract and intermediary metabolism (Weston 1982). Rumen fill appears to have a role in intake regulation when digestibility of the diet is less than 75% (Dove 1996) and Ketelaars et al. (1992) suggested that the intake of highly digestible feeds (i.e. >75% DM digestibility) is physiologically determined and physical restrictions to intake are less important. For example, rumen fill was lower on Persian clover (Trifolium resupinatum) than on perennial ryegrass (Lolium perenne) of similar digestibility but higher NDF (Williams et al. 2005), and in grazing cows consuming different amounts of Persian clover there were no differences in average rumen fill, DM or NDF loads over a range in intake from 5.6 to 20.4 kg DM/cow.day (Williams et al. 2014).
At grazing, factors such as the time available for grazing and rumination (Rook 2000; Gregorini 2012), dietary preferences (Provenza 1995), sensory factors such as palatability (Weston 1982) and management strategies (Gregorini 2012) also play roles in intake regulation. The complexities influencing intake mean that predicting intake from simple relationships based on single factors, such as digestibility or NDF concentration, will not be universally applicable across the extremes of grazing to TMR feeding systems. Therefore, when attempting to increase intake above current barriers, it is necessary to consider sward and plant characteristics and the complex signals between the digestive tract and digestive process, intermediary metabolism and sufficiency of energy and essential nutrients, and behavioural signals.
Another factor, often overlooked, is the depressive effects on intake of diets that are low in DM. For dairy cows, the critical water content was estimated to be 82% (18% DM), with a depression of 0.34 kg DM intake for each percentage increase in water content above this amount (Verite and Journet 1970). When water was added to the rumen per fistulum, Cabrera Estrada et al. (2004) showed that intake and eating rate were restricted by internal water of grass, but not by external water, suggesting that the effects of water content may be associated with the large volumes of fresh herbage that need to be processed during digestion. Water content of highly digestible pastures can be as high as 88% water, generally during winter and spring, which would restrict the intake by ~2 kg DM/cow.day.
To maximise intake from pasture, cows must consume plants that degrade quickly and are highly digestible, leading to fast rates of passage through the rumen, with intake determined by the rate of eating (bite mass by bite rate) multiplied by time spent eating (meal duration by number of meals; Allden and Whittaker 1970; Rook 2000). Grazing ruminants vary bite mass, bite rate and grazing time in response to changes in sward conditions (Hodgson 1981; Milne et al. 1982; Penning et al. 1991; Gibb et al. 1997). Bite mass, for example, increases with increased sward height and bulk density, ranging from 0.25 to 4 g DM in cattle offered micro-swards of lucerne (Medicago sativa) and paspalum (Paspalum dilatatum) (Ungar 1996) and the rate of biting is related to the ease of prehensions, herbage shear force and bite mass, as smaller mouthfuls often lead to an increase in bite rate.
Sward height and sward density also influence hourly intake rate via effects on bite mass (Mayne et al. 1997). Intake of fasted cows in short-term grazing experiments was maximised at between 3.5 and 4 kg DM/h when sward heights were greater than 18 cm (measured with a sward stick). However, intake rate was also high at 3 kg DM/h, when sward height was 15 cm. With shorter swards, bulk density becomes important, with intake rates varying from 1 to 2.5 kg DM/h for swards varying in bulk density from 1.7 to 3.1 kg/m3. Thus, high intake rates may be achieved by grazing dairy cows offered swards taller than 15 cm or dense swards below 15 cm. However, these high, short-term intake rates are difficult to translate into grazing systems as swards change during grazing events where grazing management and optimum allowance of pasture on offer are important.
Recent research activities aimed at improving feed supply, while limiting environmental impacts
Recent feedbase and nutrition research in New Zealand has focussed on the first two themes described by García and Fulkerson (2005) of growing more forage on farm and increasing its utilisation, while also limiting environmental impacts. These solutions result in increases in DM intake and remain relevant to both Australian and New Zealand. In New Zealand, there has been increasing societal concern surrounding the environmental impact of a growing and intensifying agricultural sector (Ministry for the Environment and Statistics New Zealand 2015). In various regions, the dairy industry is facing water-quality regulations, with a strong focus on nitrogen (N) leaching. Consequently, the dairy industry faces increasing pressure to improve efficiency and productivity as well as environmental performance (Monaghan and de Klein 2014). Simply growing more feed on farm can no longer be a sole objective; it must be done with a view to reducing the environmental footprint, with New Zealand farmers increasingly needing to farm within N limits.
Forages for reduced nitrate leaching
The Forages for Reduced Nitrate Leaching program is a cross-sector approach to developing profitable farming systems that enhances N-use efficiency, and reduce environmental impact for arable, dairy, sheep and beef farms. It has explored optimised pasture mixtures and cropping sequences to meet a research target of 20% reduction in nitrates leached per hectare.
Perennial ryegrass and white clover (Trifolium repens) are the most widely used species in New Zealand grazing swards. A promising novel strategy to reduce N leaching is the inclusion of more plant species to these traditional swards (Pembleton et al. 2015). Species such as chicory (Cichorium intybus), plantain (Plantago major) and lucerne are among the key species being investigated. Such diverse sward mixtures can reduce N leaching by reducing the amount and concentration of N excreted in the urine (Pembleton et al. 2015). This can arise from several mechanisms, including the lower N content of diverse swards, an improved ratio between water-soluble carbohydrates and protein, or the presence of plant secondary compounds that may increase water intake and urination volumes (Totty et al. 2013). In addition, chicory and plantain tend to have higher water contents than do grasses and may act as diuretics, increasing the total volume of urine and, hence, diluting the N concentration of urine patches (Pembleton et al. 2015). Consequently, the inclusion of diverse swards in the diet of dairy cattle has been shown to achieve reductions in the excretion of total urinary N in the order of 20–50% (Woodward et al. 2012). This, together with observed increases in urine volume, has led to 20–60% reductions in N concentrations observed in the urine of cattle grazing diverse swards (Woodward et al. 2012; Totty et al. 2013; Edwards et al. 2015). Plantain appears to be the component of a diverse pasture that drives the reduced urinary N concentrations (Edwards et al. 2015).
In line with these results, system modelling has indicated that reductions in N leaching of 11–19% can be achieved at the farm scale, if 20–50% of the milking platform is allocated to diverse swards (Beukes et al. 2014). Milk production was maintained or increased slightly in this study, and is reflective of forage yields achieved from diverse pastures similar to those from traditional pastures, with diverse-pasture production typically being stronger during summer in non-irrigated areas (Woodward et al. 2013).
Outstanding issues with diverse pastures include sward persistency beyond 3–4 years, and development of optimised management, and the resultant profitability when used by farms that have regulatory N limits.
Other crops and forages have also been shown to be useful in providing good yields and reduced N leaching. Low-N crops such as fodder beet, which can grow more than 24 t DM/ha (Chakwizira et al. 2016), have the potential to reduce urinary N (Edwards et al. 2014). Italian ryegrass (Lolium multiflorum), which has high winter growth rates, can also reduce N leached by more than 25% as a result of increased daily N uptake during winter (Malcolm et al. 2014).
The grazing of any winter crop contributes a large proportion of total N leached from the dairy operation. This is due to high crop yields leading to higher stocking densities at a time of year when leaching risk is greater. Following the winter feeding of the forage crop, the land is typically left fallow until a new crop is sown in mid-spring or returned to new pasture. Catch crops, such as oats (Avena sativa), can be used to cover fallow ground and take up urinary N deposited during winter grazing. This practice can reduce nitrate leaching by 30%, while boosting annual crop yield by 6–12 t DM/ha (Inside Dairy 2016).
Pastoral 21
A large national set of farm-system trials, Pastoral 21 Phase 2 (P21), was initiated over a 5-year period (2011–2016) in New Zealand, with the aim of addressing these challenges by developing industry accessible, adoptable, system-level solutions for profitably increasing production, while reducing environmental footprint. A key outcome was to field test the proposed solutions for demonstrable efficacy and value (Chapman et al. 2012).
Results from Waikato and Canterbury region multi-year farm-scale studies showed that large reductions in N leached, in the order of 30–50%, can be achieved with minimal impact on production and profitability (within 5%; Macdonald 2014). In the Waikato, the 4-year farm study of a combination of reduced N fertiliser (46 vs 137 kg N/ha), use of standoff pads at strategic times, limited amounts of a low protein feed (267 vs 0 kg grain/cow), a lower stocking rate (2.6 vs 3.2 cows/ha) and improved cow genetics (199 vs 129 breeding worth, as at 2014) showed that there are feed and management options available to reduce N and P losses. The P21 program is now investigating how to boost profits while retaining the N leaching gains that have been made. Urine spreading and diluting technologies are being tested (Selbie et al. 2015), and may provide a more cost-effective mitigation option.
Similar results (30% reduction in N leached) were achieved over 3 years at a farm scale in Canterbury, with a combination of a lower stocking rate (3.5 vs 5.0 cows/ha), reduced N fertiliser (158 vs 311 kg N/ha), less brought in supplement (0.34 vs 4.95 t DM/ha) and the use of diverse pastures (ryegrass and white clover with chicory, plantain, and red clover (Trifolium pratense)) and a catch crop following the winter kale crop (Macdonald 2014). These options are currently being tested on commercial farms.
Chicory and plantain
Pasture-based dairy farms often face feed shortages during periods of summer–autumn soil moisture deficits, and may require supplementary feed to maintain milk production. Chicory and plantain can produce more DM in summer with superior nutritive characteristics compared with perennial grasses (Li and Kemp 2005; Moorhead and Piggot 2009). Chicory and plantain can also increase milk production (Waugh et al. 1998; Chapman et al. 2008; Pembleton et al. 2016). DairyNZ developed the chicory and plantain research program, which ran from 2010 to 2013, to determine the milk production response to increasing amounts of chicory and plantain in the diet, to develop grazing management guidelines and to understand the impact of chicory and plantain on farm profitability.
In two summer experiments, dairy cows were offered perennial ryegrass-based pasture or ryegrass-based pasture plus either chicory or plantain at 20%, 40% or 60% of the diet (Minnee et al. 2012). When the ME of ryegrass was moderate (10.5 MJ/kg DM), the milk fat+protein yields were similar for all treatments. However, when the ME of the ryegrass dropped to 9.6 MJ /kg DM, cows fed herbs ate ~1 kg DM more per day and milk fat+protein production was 17% higher than for cows fed only ryegrass-based pasture. A third summer experiment included maize silage in the diet with herbs and pasture. At similar feed allowances and intakes, cows fed pasture plus chicory or plantain produced 13% more milk fat+protein than did cows fed only pasture. When maize silage was added to the pasture plus herb diet, milk fat+protein production was increased by 7% (compared with pasture only) but the cows consumed on average 2 kg DM/day more than did cows on the other diets. This implied that the conversion of DM into milk fat+protein was less efficient when maize silage comprised part of the diet, with insufficient intake of crude protein implicated; however, a longer-term study is needed to determine whether this additional feed was allocated to body condition rather than milk.
Grazing management guidelines have been developed from research that indicates that grazing frequency has the greatest impact on yield (Lee et al. 2012). In the first year after spring sowing, grazing chicory and plantain at 25–35 cm height increased herbage production, while in the second year, both species should be grazed at 25 cm height to limit reproductive stem growth and maintain nutritive characteristics.
The value of chicory crops to perennial ryegrass-based dairy farms in the Waikato was assessed using non-linear optimisation models of three farms differing in the degree to which imported supplement was used (Romera et al. 2015). Results indicated that at a milk price of $NZ7/kg fat+protein, farmers were unlikely to receive adequate reward for the additional complexity arising from the utilisation of chicory crops, especially when imported supplement provides more flexibility.
Future feed system
Predicting future production systems and feeding trends is impossible and the most that can be achieved is a presentation of alternative futures in light of such things as volatile seasonal weather conditions and environmental pressures to reduce nutrient losses and different options farmers may take to achieve targets.
An attempt at what the typical New Zealand dairy farm may look like in 2030–2031 was made by forecasting two future farming scenarios in 2030–2031, on the basis of current trends in types of feed used on-farm, stocking rates, farm growth and milk fat+protein production (DairyNZ 2016b); in the modest milk fat+protein growth scenario, milk production increased by 0.8% per year, and the high-growth scenario assumed milk production would increase at twice this rate, namely 1.7% per year. There were two pathways to the modest-growth scenario, either through holding stocking rates constant and a modest increase in milk fat+protein per cow or by reducing stocking rates and increasing milk fat+protein per cow more to achieve the same overall production. The high-growth forecast scenario also assumed that stocking rates would increase from 2.8 to 3.0 cows/ha.
Annual per cow feed demand was derived on the basis of estimated milk fat+protein per cow and constant average cow liveweights. Two pathways were developed for the high-growth scenario, with either pasture and crop or imported supplementary feed being the drivers of the additional feed consumed, while for the modest-growth scenario, it was assumed that the growth would be driven from increased pasture and crop eaten per cow.
A summary of the key feed use per hectare in 2030–2031 is provided in Table 4. The modest-growth scenario with constant stocking rates shows feed eaten increasing to 14.4 t DM/ha, with the increase occurring from more pasture and fodder beet consumed, while palm-kernel extract imports and maize silage decline. The second modest-growth scenario with reduced stocking rates shows that feed eaten per hectare reaches only 14.1 t DM/ha, with the difference being lower pasture consumed per hectare.

|
In the fast-growth supplements scenario, total feed eaten increases to 15.7 t DM/ha due to increased milk fat+protein production. Most of the additional feed consumed will be from increased palm-kernel extract, maize silage and barley. In the fast-growth pasture and crop scenario, the increase in feed eaten will be driven by twice as much fodder beet, while the volumes of imported supplements were capped at current levels. Pasture eaten per hectare is 1 t higher than the modest-growth constant stocking-rate scenario.
The modest-growth scenario is the most likely dairy industry pathway into the future, although if more productivity improvements occur over the next decade, the fast growth pasture and crop scenario is considered more likely. Other factors not considered in the analysis, but which will clearly influence the production systems, include such things as consumer acceptance, social licence to operate, natural capital and compliance for market access, emissions, sustainability-framework targets, competing demands on resources and emergence of transformative technologies.
Recent research activities aimed at improving feed efficiency
In Australia, much of the recent funding from the dairy industry and state governments has supported research focusing on understanding when PMR feeding systems (Australian Systems 3 and 4) are profitable, and fit into the third theme of more efficient use of supplements, as described by García and Fulkerson (2005). This commenced immediately following 10 years of below-average rainfall across Australia, characterised by reduced availability of irrigation water and reduced pasture DM production (Dairy Australia 2011), with research and development supported in Victoria, Queensland and Western Australia. This drought led to an increased reliance on purchased forage and concentrates on at least 18% of Australian dairy farms. In addition, a wider range of supplements was used, including an array of by-products that created challenges in providing a consistent diet with optimised nutrient concentrations.
Feeding systems based on the provision of mixed rations to dairy cows grazing perennial pastures (termed PMR systems) have presented the dairy industry with opportunities for improved intake, production and feed efficiency, but also significant challenges (Wales et al. 2013). There is an expectation that milk production per cow will increase as systems use higher amounts of imported feed. However, results from a survey conducted in 2010 (Dairy Australia 2011) showed that PMR systems were not performing demonstrably better than were those using large amounts of grain offered in the dairy at milking time, despite having opportunities to feed formulated rations. In 2010, the advantage of PMR over high-grain feeding in the bail was 480 versus 471 kg fat+protein/cow.year (~6400 versus 6200 kg milk/cow.year). This lack of a benefit provided the impetus for research to quantify the benefits of PMR systems. To this end, improvements in PMR systems have remained modest, with the latest survey statistics (2014–2015) showing about the same advantage of ~200 kg in average milk production from cows on PMR systems that produced a total of 6600 kg/year.
In Victoria, a series of 11 short-term grazing experiments and six digestibility experiments were conducted in a temperate, high-rainfall environment. The objectives were to define the marginal milk production responses to PMR consumed with limited amounts of pasture, and to quantify the contribution to increased milk production from increased intake and DM digestibility, and hence ME supply. To achieve the objectives, the following four main research questions were addressed: (1) can milk responses be improved by feeding PMR based on cereal and maize grain compared with feeding equivalent ME as cereal grain fed in the dairy and forage in the paddock; (2) can replacement of some wheat with canola meal (Brassica napus) in the PMR increase intake and milk production further; (3) can DM intake and milk responses be increased through increasing the herbage allowance; and (4) are there negative impacts on health and welfare of cows in PMR systems compared with systems where supplements are consumed in the dairy?
Milk responses can be improved by feeding PMR
The PMR component of the diets was formulated to be isoenergetic, with supplements fed in the dairy on the basis of NRC (2001). Overall, 10 kg DM of total supplement (comprising ~70% concentrate and ~30% forage) was required for responses of increased DM intake and increased production of energy-corrected milk (ECM) for cows consuming cereal/maize-based +/− canola meal+forage PMR compared with cereal grain fed in the dairy and forage in the paddock (Auldist et al. 2013, 2014, 2016; Golder et al. 2014). Also, milk production reached a maximum at higher amounts of total supplement intake. For example, Auldist et al. (2013) reported that maximum milk production occurred when cows were consuming 1.5 kg more of supplement DM as a PMR than when they consumed cereal grain in the dairy. These conclusions hold for situations where PMR is offered as two equal amounts after milking, but not for situations where it is offered as a single feed once per day (Wright et al. 2014).
The improved ECM-production responses of PMR were often associated with milk fat concentration not declining to the same extent with an increasing supplement intake, as it did when feeding cereal supplements in the dairy (Auldist et al. 2013, 2014, 2016; Golder et al. 2014). Diet is clearly a major factor in this response (Shingfield et al. 2010; Bauman and Griinari 2003). Diets with low fibre and high starch and/or lipids rich in polyunsaturated fatty acids are often associated with low milk fat concentration (Stockdale et al. 1987). Differences could also be expected in the site, and end products, of digestion in the PMR diets where approximately half of the starch from wheat (Triticum aestivum) was replaced with maize grain starch (Khorasani et al. 2001). The transferability of these results to New Zealand systems may be limited as grain supplementation rarely approaches the amounts from which benefits of using a PMR are observed.
Replacing wheat with canola meal further improves milk responses
Protein supplements have been associated with increased DM intake and milk production (Butler 1998; Broderick 2003; Ipharraguerre and Clark 2005; Olmos Colmenero and Broderick 2006). A series of experiments showed that, under some circumstances, replacing ~20% of the wheat DM with canola meal in a PMR ration fed twice daily to grazing cows can increase intake and milk production. The canola meal provided more CP in the PMR, while maintaining the ME concentration. Cows consuming PMR with canola meal compared with a PMR without canola meal increased DM intake from both pasture (lower residuals or post-grazing biomass) and supplement (less refusals) by 0.5–1.9 kg DM/cow.day (Auldist et al. 2014; Golder et al. 2014). In these spring and autumn experiments (early lactation and late lactation respectively), cows offered the PMR with canola meal produced more ECM (ranging from 1.7 to 4.0 kg ECM/cow.day) than did cows offered the PMR diet without canola meal (Auldist et al. 2014; Golder et al. 2014). These differences occurred when supplement intake was at least 12.0 kg DM supplement/cow.day.
Increasing pasture allowance further improves DM intake and milk responses
Cows consuming PMRs graze further into swards, resulting in higher DM intakes despite very low pasture allowances (14–15 kg DM/cow.day, measured to ground level) and an equal opportunity to graze compared with cows consuming cereal grain in the dairy (Auldist et al. 2014, 2016). Three experiments were undertaken to quantify the additional DM intake from providing cows with higher allowances of pasture. Increasing the herbage allowance from 15 to 40 kg DM/cow.day (measured to ground level) increased pasture intake by up to 5.4 kg DM/cow.day, when 12 kg DM of supplement PMR was also on offer (Auldist et al. 2016, 2017), with an increase of up to 5 kg ECM/cow.day. The additional pasture intake is a result of increased bite mass and bite rate, with no differences in time spent grazing per day (Wright et al. 2016).
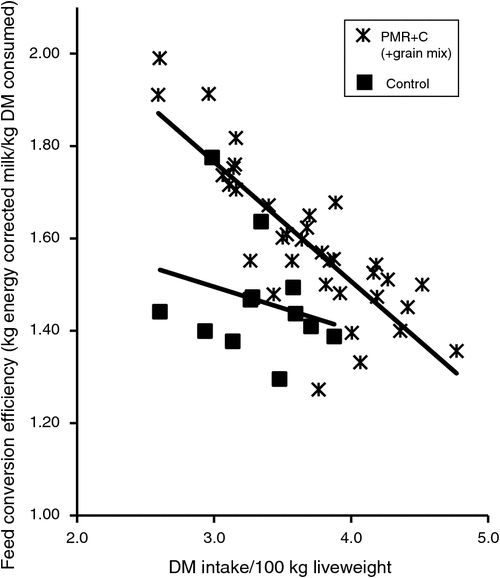
Data from the PMR experiments conducted during early lactation (Auldist et al. 2013, 2016) have been combined to explore relationships between intake and feed efficiency (Fig. 2). Data from experiments where cereal grain was fed in the dairy illustrate the negative relationship between intake and feed efficiency; that is, as intake increases (expressed on % liveweight basis), feed conversion efficiency decreases. Analysis of the series of PMR+canola meal experiments in early lactation showed higher feed efficiencies, such that at equivalent feed conversion efficiency cows consuming the well formulated PMR consumed more DM, and produced more ECM. This demonstrates that the PMR feeding system represents a more optimised diet. These relationships also point out the danger in attempting to optimise a partial measure of productivity, such as feed conversion efficiency. Without context, feed conversion efficiencies of 2 kg ECM/kg intake (% liveweight) appear desirable, but, in this situation, intake was very restricted at 2.6% of bodyweight (equivalent to ~16 kg DM for a 600-kg cow), and body reserves were likely to be supporting a significant amount of milk production, which, over the longer term, is unsustainable and is also associated with negative effects on fertility, health and milk production in the following lactation.
|
There are no negative impacts on health and welfare of cows in PMR systems
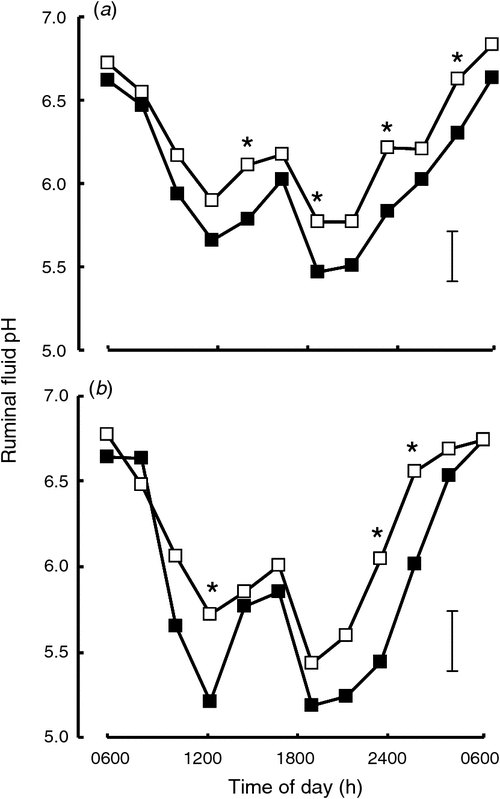
The final objective was to determine whether there were any negative health and welfare outcomes in cows on PMR systems, with the focus on risk of ruminal acidosis, hoof health and behaviour of cows while on feedpad and at grazing. Auldist et al. (2013) speculated that a well formulated PMR, with slower-degradable starch sources, such as maize grain, could lead to more stable ruminal fermentation than with grain fed in the dairy. Ruminal pH is used as an indicator of acidosis risk. Two metabolism experiments confirmed that ruminal pH was consistently higher in samples from cows consuming pasture and PMR diets than in those from cows consuming pasture and grain diets (Greenwood et al. 2014; see Fig. 3). Similarly, Golder et al. (2014) identified acidosis risk for a range of diets and showed that PMR diets had significantly lower risks of ruminal acidosis than did diets where wheat was fed in the dairy at a range of intakes (Fig. 4). It was further speculated that the reduced ruminal-fluid pH in the cows receiving high amounts of wheat grain in the dairy during milking would compromise the digestion on the diet overall, since ruminal-fluid pH below ~6.0 is known to compromise fibre digestion (Leddin et al. 2010). However, Greenwood et al. (2014) found no difference in whole-tract digestibility of DM, N, starch or NDF between the wheat grain in the dairy and PMR diets, despite the differences in ruminal pH, as described above. Foot health was not affected, with experiments indicating that there was no effect of feeding system on the presence of foot lesions associated with white line disease, paintbrush haemorrhage and traumatic bruising in early lactation (Coombe et al. 2013) or on the concentrations of serum haptoglobin, as an indicator of subacute ruminal acidosis (Coombe et al. 2015). In terms of feeding and social behaviour, feeding supplements as a PMR for ~1.5 h in the experiment described by Auldist et al. (2013) resulted in a small reduction in grazing time, while rumination and lying time and number of lying bouts were unchanged and, hence, there was no indication of negative consequences for cow welfare (Hetti Arachchige et al. 2013). Aggressive interactions decreased on the feedpad when feeding space was increased from 0.6 to 0.75 to 1.0 m and feeding behaviour improved, with the effects being greatest for subordinate cows (Hetti Arachchige et al. 2014).
|

|
The Queensland subtropical dairy industry produces 5% of the national milk from ~500 dairy farms (Dairy Australia 2015b). These farms are in two regions in the south, one in central and one in northern Queensland, and feeding systems have evolved to exploit the natural advantages of each region. For example, TMR systems dominate in the Darling Downs where concentrates are cheap and grazing and PMR systems are concentrated in the South East Coast region where a more reliable rainfall allows pasture to be grown and grazed, albeit of lower digestibility than that in temperate dairying regions.
A key nutritional challenge has been how to profitably feed dairy cows in environments where home-grown forage supply is highly variable and of lower digestibility than temperate dairying regions. The autumn period of the year is when pasture grazing options are most limited and milk production declines, which is significant in these year-round calving systems. Higher-cost purchased energy and protein concentrate supplements are typically fed to help overcome the nutrient deficit representing ~40% of the diet DM. In a recent survey, average feed-related costs ranged from 29 cents/L for PMR systems to 38 cents/L in TMR systems and margin over feed costs from 25 to17 cents/L (Murphy and Simpson 2014). Reducing costs of production is a key strategy to improve profitability and feed costs make up the largest category of costs.
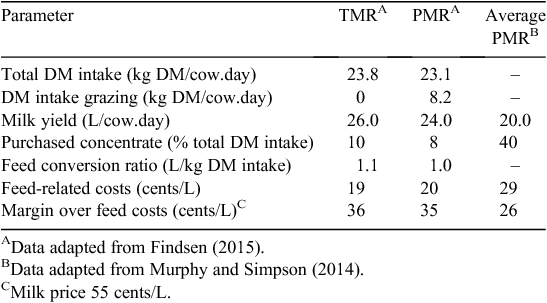
The dairy industry in Queensland is focussed on development of strategies that reduce the cost of rations, which increases the margin over feed costs through incorporating more highly digestible/high-protein home-grown forages (Findsen 2015). Their approach has been to assemble diets based on grazing and feeding of forages to reduce reliance on purchased concentrate supplements, within the commonly used TMR and PMR feeding systems in the subtropics. The forage options include silages produced from white and forage sorghums (Sorghum spp.), maize (high cut), lucerne, soybean (Glycine max), lablab (Lablab purpureus) and lucerne hay. Well made, these forages provide high-energy and -protein options for incorporation into rations and have reduced the reliance on purchased concentrates to 10% of the total ration (Table 5). To overcome variability in rainfall and forage supply, sufficient silages are produced to provide buffers in line with the risk profile of the farmers. Although this strategy introduces more complexity to the feeding system, the margin over feed costs can potentially be increased by 10 cents/L, due to more optimised supply of nutrients produced on farm, leading to increased milk production and high DM intakes.
|
Dairy production in Western Australia is conducted in a region with a Mediterranean-type environment, where annual rainfall is ~800 mm, falling in the cool months from May to November when cows graze, followed by a hot, dry period from December to April when pasture growth is minimal and grazing is limited (Bolland and Russell 2010). The predominant feeding strategy is to feed concentrates during milking, and graze during the cooler months and offer conserved forage in the paddock during the hotter months (Australian System 4). An alternative strategy that is being tried on farm is feeding a portion of the concentrate with the forage as part of a mixed ration during the summer period, with the aim of increasing feed efficiency and milk production. A comparison of these two strategies has recently been reported as part of a survey of the physical performance of 10% of dairy farms in Western Australia, conducted over a 15-month period (McDonnell et al. 2016). Their hypothesis was that incorporating concentrate as part of a ration would produce more milk and increase annual feed conversion efficiency because rumen fluctuations would be reduced, in comparison with ‘slug feeding’ of concentrates in the dairy. Results showed significant variation among farms within each system. Average milk production and annualised feed conversion efficiency were similar (7680 kg/cow and 1.37 kg ECM/kg total intake respectively), and McDonnell et al. (2016) concluded that the alternative feeding systems are currently not optimised. Further research to optimise the alternative strategy is exploring the role of canola meal and slower-degradable starch sources on a commercial farm and preliminary results suggest that milk responses in the order of 3 kg/cow.day are possible through fine tuning proportions of the components of the ration (R. McDonnell, pers. comm.).
Gaps in knowledge and fertile areas for further feedbase and nutrition research
Incorporating high-yielding crops and diverse/alternative forages into systems
The rapid growth of fodder-beet use in New Zealand has demonstrated potentially high yields, and farmers and researchers are understanding the best way to integrate fodder beet into systems, with transition programs being important to maintain animal health. However, for a feed that is currently having such a large impact on the New Zealand feed supply, it has exposed gaps in our understanding, particularly around the health of animals with long-term exposure to fodder beet. An area of concern from an environmental and perceived animal-welfare perspective is winter grazing of crops and exposure of animals to mud.
The development of diverse pastures will likely require further iterations of optimisation before they are proven technologies for farmers from both countries to use with confidence. Most promising is plantain as it is easily incorporated into existing systems, has a reasonable persistency, and has potentially significant environmental-footprint attributes, which could mean that farms can grow productivity and profitability under an imposed N limit.
Capturing forage-yield benefits from genetic gain
As far as improving the performance of the base pasture, several technologies are in development that could prove to be game-changing over the next 10 years. The challenge will be to demonstrate capture of these benefits at a farm-system level.
The forage-value index (FVI), which combines the performance value and economic value of a range of yields, and potentially nutritive and persistency traits, will help farmers in both countries select pasture cultivars that are expected to deliver greater farm profits. In New Zealand, the difference between the highest-ranked and the lowest-ranked perennial ryegrass cultivar ranges from $556 profit/ha to $863/ha, depending on the region. Since the 1960s, pasture selection has delivered $12–$18/ha.year in extra farm profits (Chapman et al. 2017). The immediate research challenge is to evaluate the FVI in a farm-system context with support from modelling, and to extend the FVI beyond yield evaluation to nutritive and persistence traits of the cultivars.
Currently, there are several exciting genetic options in development that will require evaluation at a farm scale. Genomic selection can be used to predict performance, shorten the breeding cycle and speed up genetic gain, as has been recently demonstrated in dairy cows (Barrett et al. 2015). New DNA tools are allowing identification of ryegrass strains that may be hybridised, with Italian ryegrass demonstrating more than a 20% yield improvement with hybrid vigour (DairyBio 2016). A real step change in Australasian pasture productivity will be achieved if the barriers of photosynthesis and energy levels in pastures can be overcome. AgResearch have recently developed a GM technology to enhance photosynthesis in C3 plants (Barrett et al. 2015; Bryan and Roberts 2015). The high-ME technology has boosted lipid concentrations in ryegrass from 3.5% to 8%, lifting ME levels by 10%, but the real breakthrough has been the 25–40% faster growth rates in glasshouse trials, which are the result of a more efficient photosynthesis. Encouragingly, modelling and glasshouse work conducted so far includes a 12% increase in milk fat+protein production, improvements in animal reproduction, possible increases in animal liveweight, 17% decrease in N2O emissions, 15–30% decrease in methane emissions, a potentially enhanced water use efficiency and drought tolerance, and reduced nitrate leaching (Bryan and Roberts 2015). Field and animal-nutrition experiments are necessary to confirm the value of these novel forages for Australasia’s pastoral industry, leading to recommendations of any change to current supplementary feeding or agronomic or grazing management practices. Current work in New Zealand on condensed tannins in white clover and novel endophytes, and on high-fructan grasses in Australia, will also provide homegrown, grazeable forage options in the future (Barrett et al. 2015).
Potential of feeding grain mixes to increase milk fat+protein production
The shift towards more complicated feeding systems is at odds with desire from farmers from both countries to manage simple feeding systems that are easy to implement. For example, PMR systems require more investment and expertise to realise the benefits. However, the benefits of PMR over traditional supplementary-feeding systems raised the possibility that feeding the grain portion of the PMR to cows using existing grain-feeding infrastructure in most modern dairies could capture some or all of the milk-production benefit observed with PMR systems, without the need for farmers to purchase a mixer wagon and build a feed pad. Feed companies currently formulate mixes of concentrates and implementing improved formulations would be relatively simple. Feeding a mix of concentrates from a PMR was tested in an experiment with cows grazing a restricted pasture allowance and fed a mix of maize grain, wheat grain and canola meal in the dairy at milking times, with lucerne hay fed in the paddock (Auldist et al. 2016). Milk production was compared with that from a PMR containing the four components fed on a feed pad after milking and wheat grain fed in the dairy (and lucerne in the paddock). Results showed that most of the advantage of the PMR over wheat fed in the dairy was achieved by feeding the grain mix. This indicates that the benefits of the PMR are related to the composition of the ration rather than the method of feeding. This was also supported by a lack of milk response from a simple PMR mix of cereal grain and forage fed to cows on a feed pad (Auldist et al. 2013).
Much of the increased milk response in cows fed a formulated grain mix and PMR strategies was associated with the arrested decline in milk fat concentrations as supplement intake increased, compared with the marked decline in milk fat concentrations observed in the cows consuming equivalent amounts of wheat grain in the dairy. The formulated grain-mix strategy could, therefore, be a much cheaper option than PMR, which would be relevant to a much wider cross-section of pasture-based dairy industries in Australia and New Zealand. Environmental benefits are also possible through the use of supplementary feeding to dilute N intake and increase milk production, which increases the efficiency of N use and potentially reduces N leaching. Beukes et al. (2014) suggested that N-use efficiency of standard pasture only was ~14% and we calculated that diets based on formulated grain mixes can improve the conversion of dietary crude protein intake to milk protein to 26%, on the basis of data presented in Auldist et al. (2016).
Although the recent research has highlighted the potential of using formulated grain mixes, the underlying mechanisms behind the observed milk responses are not fully understood. It is important to understand the reasons for the responses to allow the benefits to be reliably captured in other environments/dairying regions, which may have a different choice of ingredients available. These include the reasons for the consistent milk fat percentage responses and for the increased motivation of cows to graze pasture when canola meal has replaced wheat. Research is needed to quantify the site and the extent of digestion of carbohydrates and protein and the supply of metabolisable protein and adequacy of amino acids, for example, which would provide more precise information on carbohydrate and protein degradation of temperate pastures and local supplements.
Mechanistic nutrition models to determine complementary supplement mixes in grazing systems
Predicting milk production from information about the diet in grazing systems is difficult due to the complex interactions among the requirement for nutrients by the cow, the characteristics of different feeds and the contributions from body reserves towards the animal’s requirements for the milk produced. Historically, we have formulated rations to meet requirements for estimated ME and crude protein, while ensuring that NDF from long forage sources falls within certain bands to ensure that sufficient rumination is occurring. However, there are significant limitations to this approach, particularly with higher-producing dairy cows, where ME intake is often not the limiting nutrient for production. As pasture nutritive characteristics change throughout the growing season, decisions on which and how much supplement (or supplement mixes) to feed are further complicated.
Mechanistic nutrition models, such as the Cornell Net Carbohydrate and Protein System (CNCPS 2000), have been designed to improve the prediction of milk responses. The complex mathematical construct is based on up-to-date literature, and is most applicable to cows in a housed environment consuming TMR to appetite (Fox et al. 2004). Compared with TMR, modelling the performance of dairy cows grazing pasture introduces additional complexity because of the greater variation in the nutritive characteristics of the forage, and, hence, greater variability in nutrient intake among cows.
Further advances in our understanding of nutrient supply from pasture and supplements through targeted research to describe a full suite of nutrients at different times of the year, and the refinement of nutritional models such as the CNCPS, make it possible to improve precision in formulating supplements to complement nutrients from pasture. A good example of this is in the recent research where the formulation of complicated mixes of grain supplements led to increased milk production compared with other dietary treatments of similar ME concentrations (Golder et al. 2014).
Individualised or differential feeding of concentrates to increase herd milk production
The increased availability of computerised feeding and milk-monitoring technologies has led to farmer interest in individually feeding concentrates to cows during milking on the basis of factors such as milk yield, days in lactation and parity. However, there is currently little evidence that individualised or differential feeding of concentrate supplements to dairy cows increases milk production, particularly in systems where forage is available ad libitum, such as those systems where housed cows are provided with silage to appetite and different amounts of the same supplement are offered to different cows (Hills et al. 2015). In these situations, no additional nutrients are brought into the system and the available nutrients are just distributed differently among cows.
The exception may be in systems where cows graze as a herd and receive supplements in the dairy where competition for a restricted pasture resource may limit nutrient intake in cows of higher genetic merit. In these types of systems, there is significant variation in pasture and total DM and nutrient intake of cows, which suggests that individualised feeding could lead to improved marginal milk-production responses. The difference between previous research with housed cows and grazing systems is that nutrient intake can be highly variable in grazing cows because of the effect of the decline in nutrient characteristics with pasture depletion within a grazing event. Hills et al. (2015) concluded that further research must be conducted to better understand the relationship between factors such as grazing and other activity-based behaviours and pasture intake, substitution rates and milk production. To this end, recent research from New Zealand (Dela Rue 2015) concluded that individualised feeding did not increase milk fat+protein production, where more of the same supplement was fed to higher-producing cows and less to lower-producing cows than with flat-rate feeding during early lactation. However, what has not been explored in experiments with grazing cows is identifying the limiting nutrients from grazed pasture and providing these nutrients as supplements. For example, a more energy-dense supplement may be appropriate for a lower-producing cow, while additional protein may be required for cows of higher genetic merit.
Economic frameworks to reliably assess tactical feeding management options
Research into milk responses to feed in combination with prices and costs has been limited, despite an increased need due to changes to more diverse pasture bases and supplementary feeding across both countries since the 1980s. Milk-response functions to supplementary feeding and production economics can be used to help farmers determine the most profitable amount of an input to use, given input cost and output price. The theory and principles are well established, with Heady (1952) conducting the seminal work in agricultural production economics in the 1950s, particularly in the economics of milk-response functions, with virtually all studies in agricultural production economics since then citing the principles established by Heady.
The experiments used by Heady remain relevant to the USA, but the principles developed are more widely applicable in the analyses of input–output relationships in dairy production in Australia and New Zealand. Some more recent international applications of production economics include those of André et al. (2010) who examined the profit-maximising amount of concentrate to feed cows in confined systems and those of Adduci et al. (2015) who developed a model to evaluate the efficiency of dairy cow rations. A limitation to applying the production-economics approach in grazing systems has been the lack of milk-response functions. Recent Victorian research developed response functions of milk production to cereal-based concentrates (Heard et al. 2017) and this was used in production-economics analyses to find the profit-maximising amount of the supplement (Ho et al. 2017), given a particular situation on-farm and a set of prices and costs.
Dairy farmers often consider supplementary feed, fertiliser and water as substitutes when making decisions about how to feed their cows, and response functions are needed for non-cereal-based supplements and water to understand the trade-offs between such inputs in making short-term feeding decisions. Some of this work is underway, but there are still gaps in other areas. Work is currently being conducted to develop milk-production responses to supplementary forages (C. Ho, pers. comm.). Data about milk-production responses to PMR feeding under a range of conditions are available, which could potentially be used to develop generalised responses to PMR feeding. The milk-production response to non-cereal-based supplements such as protein supplements is lacking, despite their being common supplements to grazing cows. Until research is conducted in this area, economic benefits from addition of protein-based supplements will remain unclear.
Current and previous work investigating the response to supplementary feed has focussed on milk production, but other benefits have not been quantified, for example, the potential benefits to increasing body condition. There is a need to develop analogous response functions, to change in supplement feed intake, for these other benefits. An obvious benefit of better information about response functions is improving the biological relationships used in modelling.
Recent studies by Rogers et al. (2016) defined the relationship between decreased water use and the survival and yield of lucerne and perennial pasture under different strategies. However, the economic costs and benefits, and the subsequent impact on farm profit have not been assessed. A generalised framework is also needed to enable the cost–benefit analysis of other crops and pastures as the water-response functions become available.
The aim is to help farmers make more informed tactical decisions by developing a framework incorporating a range of response functions enabling the exploration of the trade-off among fertiliser, supplementary feed and water. This would allow more informed decisions on how much extra of these inputs could be used at different times of the year, under price and cost combinations, to increase farm profit.
Integrating new technologies to optimise nutrient intake in individual cows in real time
Decisions on what type, and how much, supplement to offer are often based on some estimate of pasture DM intake by the herd and of the average nutrient concentration of the pasture consumed, to achieve a desired total DM intake and milk production, within environmental constraints. This approach cannot optimise milk production because of seasonal changes in the amount and nutrient density of grazed forage, and variation in nutrient intake among cows from both the grazed forage and offered supplements. This contrasts with cows consuming total mixed rations where, in theory, each mouthful contains the optimised mix of nutrients. A better solution would be to optimise nutrient intake in real time at the individual-cow level, which could be achieved by knowledge of the nutrient intake of grazed forage by each cow and a supplement being tailored for each cow to achieve a profitable milk response.
Technology exists to enable some of this to occur, but the greatest challenge will be integrating these elements into a feeding management decision support framework that removes much of the herd manager’s input. These technologies include near-infrared sensors that remotely measure canopy spectral reflectance to determine mass and nutrient density of swards (e.g. Starks et al. 2006). Real-time estimation of pasture intake by individual cows in a herd is also now possible with sensor and wireless-sensor networks that relate specific behaviour to intake of pasture. For example, Greenwood et al. (2017) developed reliable algorithms that predict intake of pasture when forage is not limiting, but concluded that further refinement of the algorithms is needed to account for variation within and among pastures and use of different sensor types that enable more specific classification of ingestive behaviours such as bite size, eating time, number of chews and eating rate. With real-time information on nutrient intake, mechanistic nutrition models may provide a supplementary-feed solution allowing optimal milk production that has a reduced impact on the environment. The challenge is to automate the model for each cow in the herd, on the basis of real-time estimates of nutrient intake from grazed forage. The next challenge is to use wireless-sensor networks to inform the feeding systems in the dairy to deliver the supplementary feed to each cow. In theory, the nutrient intake of each cow would approach that of a cow consuming a TMR.
References
Adduci F, Labella C, Musto M, D’Adamo C, Freschi P (2015) Use of technical and economic parameters for evaluating cow ration efficiency. Italian Journal of Agronomy 10, 202–207.| Use of technical and economic parameters for evaluating cow ration efficiency.Crossref | GoogleScholarGoogle Scholar |
Allden WG, Whittaker IAM (1970) The determinants of herbage intake by grazing sheep: the interrelationship of factors influencing herbage intake and availability. Australian Journal of Agricultural Research 21, 755–766.
| The determinants of herbage intake by grazing sheep: the interrelationship of factors influencing herbage intake and availability.Crossref | GoogleScholarGoogle Scholar |
André G, Berentsen PBM, Van Duinkerken G, Engel B, Oude Lansink AGJM (2010) Economic potential of individual variation in milk yield response to concentrate intake of dairy cows. The Journal of Agricultural Science 148, 263–276.
| Economic potential of individual variation in milk yield response to concentrate intake of dairy cows.Crossref | GoogleScholarGoogle Scholar |
Auldist MJ, Marett LC, Greenwood JS, Hannah M, Jacobs JL, Wales WJ (2013) Effects of different strategies for feeding supplements on milk production responses in cows grazing a restricted pasture allowance. Journal of Dairy Science 96, 1218–1231.
| Effects of different strategies for feeding supplements on milk production responses in cows grazing a restricted pasture allowance.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC38Xhsl2isrrP&md5=bb23cee3d355f8e22393efd7c924cc90CAS |
Auldist MJ, Marett LC, Greenwood JS, Wright MM, Hannah M, Jacobs JL, Wales WJ (2014) Replacing wheat with canola meal in a partial mixed ration increases the milk production of cows grazing at a restricted pasture allowance in spring. Animal Production Science 54, 869–878.
| Replacing wheat with canola meal in a partial mixed ration increases the milk production of cows grazing at a restricted pasture allowance in spring.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC2cXpt12mt7Y%3D&md5=95bcd71bfc8610f11297ff8fc792aeafCAS |
Auldist MJ, Marett LC, Greenwood JS, Wright MM, Hannah M, Jacobs JL, Wales WJ (2016) Milk production responses to different strategies for feeding supplements to grazing dairy cows. Journal of Dairy Science 99, 657–671.
| Milk production responses to different strategies for feeding supplements to grazing dairy cows.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC2MXhvVCitbnE&md5=6e7da1dc21f54c3989463b9129a68193CAS |
Auldist MJ, Wright MM, Marett LC, Jacobs JL, Wales WJ (2017) Effects of pasture allowance on milk production of dairy cows offered increasing amounts of partial mixed rations in spring. Animal Production Science 57, 1305–1310.
| Effects of pasture allowance on milk production of dairy cows offered increasing amounts of partial mixed rations in spring.Crossref | GoogleScholarGoogle Scholar |
Bargo F, Muller LD, Delahoy JE, Cassidy TW (2002) Performance of high producing dairy cows with three different feeding systems combining pasture and total mixed rations. Journal of Dairy Science 85, 2948–2963.
| Performance of high producing dairy cows with three different feeding systems combining pasture and total mixed rations.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD38Xptlagt74%3D&md5=7b7b0c7ffc458966176a8eb9ce3b4236CAS |
Barrett BA, Faville MJ, Nichols SN, Simpson WR, Bryan GT, Conner AJ (2015) Breaking through the feed barrier: options for improving forage genetics. Animal Production Science 55, 883–892.
| Breaking through the feed barrier: options for improving forage genetics.Crossref | GoogleScholarGoogle Scholar |
Bauman DE, Griinari JM (2003) Nutritional regulation of milk fat synthesis. Annual Review of Nutrition 23, 203–227.
| Nutritional regulation of milk fat synthesis.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD3sXntFSjurc%3D&md5=0bc6e7831734e144bb490b1ee4e04b6bCAS |
Beever DE, Doyle PT (2007) Feed conversion efficiency as a key determinant of dairy herd performance: a review. Australian Journal of Experimental Agriculture 47, 645–657.
| Feed conversion efficiency as a key determinant of dairy herd performance: a review.Crossref | GoogleScholarGoogle Scholar |
Beukes PC, Gregorini P, Romera AJ, Woodward SL, Khaembah EN, Chapman DF, Nobilly F, Bryant RH, Edwards GR, Clark DA (2014) The potential of diverse pastures to reduce nitrogen leaching on New Zealand dairy farms. Animal Production Science 54, 1971–1979.
| The potential of diverse pastures to reduce nitrogen leaching on New Zealand dairy farms.Crossref | GoogleScholarGoogle Scholar |
Bolland M, Russell W (2010) Changes in chemical properties of 48 intensively grazed, rain-fed dairy paddocks on sandy soils over 11 years of liming in south-western Australia. Australian Journal of Soil Research 48, 682–692.
| Changes in chemical properties of 48 intensively grazed, rain-fed dairy paddocks on sandy soils over 11 years of liming in south-western Australia.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3cXhsVCjsbnJ&md5=a7a0e487e2d7220b77e1dc239cb6e1aaCAS |
Broderick GA (2003) Effects of varying dietary protein and energy levels on the production of lactating dairy cows. Journal of Dairy Science 86, 1370–1381.
| Effects of varying dietary protein and energy levels on the production of lactating dairy cows.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD3sXjtVajtLc%3D&md5=5ba38e3b924f384c5a4a4e7020dd3799CAS |
Bryan G, Roberts N (2015) ‘Superior forages could result from GM technology.’ Technical series online, issue 28, December 2015. pp. 1–5. (DairyNZ: Hamilton, New Zealand) Available at https://www.dairynz.co.nz/media/4439072/technical-series-june-2016-forages.pdf [Verified 15 November 2016]
Butler WR (1998) Effect of protein nutrition on ovarian and uterine physiology in dairy cattle. Journal of Dairy Science 81, 2533–2539.
| Effect of protein nutrition on ovarian and uterine physiology in dairy cattle.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaK1cXmsVCrt70%3D&md5=492d10fcc86cffcf08c545e53e638c05CAS |
Cabrera Estrada JI, Delagarde P, Faverdin P, Peyraud JL (2004) Dry matter intake and eating rate of grass by dairy cows is restricted by internal, but not external water. Animal Feed Science and Technology 114, 59–74.
| Dry matter intake and eating rate of grass by dairy cows is restricted by internal, but not external water.Crossref | GoogleScholarGoogle Scholar |
Chakwizira E, de Ruiter JM, Malcolm B, Maley S (2016) Winter growth and quality of fodder beet. Plant & Food Research Report for Seed Force (PFR SPTS No. 12866) pp. 1–13. (The New Zealand Institute for Plant & Food Research Limited: Auckland, New Zealand)
Chapman DF, Tharmaraj J, Nie ZN (2008) Milk-production potential of different sward types in a temperate southern Australian environment. Grass and Forage Science 63, 221–233.
| Milk-production potential of different sward types in a temperate southern Australian environment.Crossref | GoogleScholarGoogle Scholar |
Chapman DF, Cullen BR, Johnson IR, Beca D (2009) Intrannual variation in pasture growth rate in Australian and New Zealand dairy regions and its consequences for system management. Animal Production Science 49, 1071–1079.
| Intrannual variation in pasture growth rate in Australian and New Zealand dairy regions and its consequences for system management.Crossref | GoogleScholarGoogle Scholar |
Chapman D, Clark D, Beukes P, Edwards G, Dalley D, Pinxterhuis I, Romera A, Shepherd M, Cameron K, Di H, Woodward S, Glassey C, Roach C, Curtis J, Lynch B (2012) Strategies for increasing dairy system productivity while controlling environmental footprint: a common systems research approach in two regions of New Zealand. In ‘Proceedings of the 5th Australasian dairy science symposium’. (Ed. JL Jacobs). pp. 417–421 (Australasian Dairy Science Symposium: Melbourne, Australia)
Chapman DF, Bryant JR, Olayemi ME, Edwards GR, Thorrold BS, McMillan WH, Kerr GA, Judson G, Cookson T, Moorhead A, Norriss M (2017) An economically based evaluation index for perennial and short-term ryegrass in New Zealand dairy farm systems. Grass and Forage Science 72, 1–21.
| An economically based evaluation index for perennial and short-term ryegrass in New Zealand dairy farm systems.Crossref | GoogleScholarGoogle Scholar |
Clark D, Malcolm B, Jacobs J (2013) Dairying in the Antipodes: recent past, near prospects. Animal Production Science 53, 882–893.
| Dairying in the Antipodes: recent past, near prospects.Crossref | GoogleScholarGoogle Scholar |
CNCPS (2000) ‘Cornell Net Carbohydrate and Protein System. [4.0].’ Computer program. (Cornell University: Ithaca, NY)
Coombe JE, Pyman MF, Mansell PD, Auldist MJ, Anderson GA, Wales WJ, Malmo J, Conley MJ, Fisher AD (2013) The effects on claw health of supplement feeding grazing dairy cows on feed pads. The Veterinary Journal 198, 672–677.
| The effects on claw health of supplement feeding grazing dairy cows on feed pads.Crossref | GoogleScholarGoogle Scholar |
Coombe JE, Pyman MF, Mansell PD, Auldist MJ, Anderson GA, Wales WJ, Conley MJ, Manos S, Hannah M, Fisher AD (2015) The effects on ruminal pH and serum haptoglobin after feeding a grain-based supplement to grazing dairy cows as a partial mixed ration or during milking. Veterinary Journal 204, 105–109.
| The effects on ruminal pH and serum haptoglobin after feeding a grain-based supplement to grazing dairy cows as a partial mixed ration or during milking.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC2MXivFGrtrg%3D&md5=eb814df3d36bd375f33a72d1921f7e7aCAS |
Cullen BR, Eckard RJ, Callow MN, Johnson IR, Chapman DF, Rawnsley RP, Garcia SC, White T, Snow VO (2008) Simulating pasture growth rates in Australian and New Zealand grazing systems. Australian Journal of Agricultural Research 59, 761–768.
| Simulating pasture growth rates in Australian and New Zealand grazing systems.Crossref | GoogleScholarGoogle Scholar |
Dairy Australia (2011) ‘Situation and outlook.’ Available at http://www.dairyaustralia.com.au/Industry-information/About-Dairy-Australia/~/media/Documents/Stats%20and%20markets/S%20and%20O/S%20and%20O%20-%20May%202011/Dairy%20Situation%20and%20Outlook%20Report%20May%202011%20V4.pdf [Verified 15 November 2016]
Dairy Australia (2015a) ‘Australian dairy industry in focus 2015.’ (Dairy Australia: Melbourne) Available at http://www.dairyaustralia.com.au/~/media/Documents/Stats%20and%20markets/Australian%20Dairy%20Industry%20In%20Focus/Australian%20Dairy%20Industry%20In%20Focus%202015.pdf [Verified 15 November 2016]
Dairy Australia (2015b) ‘Dairy feeding update: briefing notes 2015.’ (Dairy Australia: Melbourne) Available at http://www.dairyaustralia.com.au/Industry-information/About-Dairy-Australia/~/media/Documents/Pastures%20and%20feeding/Feed%20management/Dairy%20Feeding%20Update%20-%20Briefing%20notes.pdf [Verified 15 November 2016]
DairyBio (2016) ‘Better ryegrass through hybrid breeding.’ Available at http://dairybio.com.au/designer-forages/pasture-breeding-tools/ [Verified 10 November 2016]
DairyNZ (2008) ‘DairyNZ economic survey 2006–07.’ (DairyNZ: Hamilton, New Zealand) Available at www.dairynz/publications/ [Verified 10 November 2016]
DairyNZ (2010) ‘DairyNZ economic survey 2008–09.’ (DairyNZ: Hamilton, New Zealand) Available at www.dairynz/publications/ [Verified 10 November 2016]
DairyNZ (2012) ‘DairyNZ economic survey 2010–11.’ (DairyNZ: Hamilton, New Zealand) Available at www.dairynz/publications/ [Verified 10 November 2016]
DairyNZ (2014) ‘DairyNZ economic survey 2012–13.’ (DairyNZ: Hamilton, New Zealand) Available at www.dairynz/publications/ [Verified 10 November 2016]
DairyNZ (2015) ‘New Zealand dairy statistics 2014–15.’ (DairyNZ: Hamilton, New Zealand) Available at www.dairynz/publications/ [Verified 10 November 2016]
DairyNZ (2016a) ‘DairyNZ economic survey 2014–15.’ (DairyNZ: Hamilton, New Zealand) Available at www.dairynz/publications/ [Verified 10 November 2016]
DairyNZ (2016b) Feed use in the NZ dairy industry. Report to Ministry for Primary Industries. DairyNZ, Hamilton, New Zealand.
Dela Rue B (2015) ‘Individualised supplementation of grazing cows: how does it stack up?’ Technical Series in Brief, Issue 28, December 2015. Available at www.dairynz/techseriesdec15 [Verified 10 November 2016]
Dharma S, Shafron W, Oliver M (2012) ‘Australian dairy: farm technology and management practices 2010–11.’ (Australian Bureau of Agricultural and Resource Economics and Sciences: Canberra)
Dove H (1996) Constraints to the modelling of diet selection and intake in the grazing ruminant. Australian Journal of Agricultural Research 47, 257–275.
Doyle PT, Stockdale CR, Wales WJ, Walker GP, Heard JW (2001) Limits to optimising milk production and composition from pastures. In ‘Recent advances in animal nutrition in Australia 2001’. (Ed. P Cronje). pp. 9–17 (Animal Science University of New England: Armidale, NSW)
Edwards GR, de Ruiter JM, Dalley DE, Pinxterhuis JB, Cameron KC, Bryant RH, Di HJ, Malcolm BJ, Chapman DF (2014) Urinary nitrogen concentration of cows grazing fodder beet, kale and kale–oat forage systems in winter. In ‘Proceedings of the 6th Australasian dairy science symposium’. (Ed. JR Roche) pp. 144–147. (Australasian Dairy Science Symposium: Hamilton, New Zealand)
Edwards G, Bryant R, Smith N, Hague H, Taylor S, Ferris A, Farrell L (2015) Milk production and urination behavior of dairy cows grazing diverse and simple pastures. In ‘Proceedings of the New Zealand Society of Animal Production’. (Ed. J McEwan) pp. 79–83. (New Zealand Society of Animal Production: Dunedin, New Zealand)
Findsen C (2015) C4Milk progress report 4. Dairy Australia. Publisher Animal Science, Department of Agriculture and Fisheries, State of Queensland, Gatton, Australia. Available at www.dairyaustralia.com.au [Verified 1 October 2016]
Fox DG, Tedeschi LO, Tylutki TP, Russell JB, Van Amburgh ME, Chase LE, Pell AN, Overton TR (2004) The Cornell Net Carbohydrate and Protein System model for evaluating herd nutrition and nutrient excretion. Animal Feed Science and Technology 112, 29–78.
| The Cornell Net Carbohydrate and Protein System model for evaluating herd nutrition and nutrient excretion.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2cXmvFymsQ%3D%3D&md5=d548ba44c4e36faaa3a8a6114dc557e4CAS |
García SC, Fulkerson WJ (2005) Opportunities for future Australian dairy systems: a review. Australian Journal of Experimental Agriculture 45, 1041–1055.
| Opportunities for future Australian dairy systems: a review.Crossref | GoogleScholarGoogle Scholar |
Gibb MJ, Huckle CA, Nuthal R, Rook AJ (1997) Effect of sward surface height on intake and grazing behaviour by lactating Holstein Friesian cows. Grass and Forage Science 52, 309–321.
| Effect of sward surface height on intake and grazing behaviour by lactating Holstein Friesian cows.Crossref | GoogleScholarGoogle Scholar |
Golder HE, Denman SE, McSweeney C, Wales WJ, Auldist MJ, Wright MM, Marett LC, Greenwood JS, Hannah MC, Celi P, Bramley E, Lean IJ (2014) Effects of partial mixed rations and supplement amounts on milk production and composition, ruminal fermentation, bacterial communities, and ruminal acidosis. Journal of Dairy Science 97, 5763–5785.
| Effects of partial mixed rations and supplement amounts on milk production and composition, ruminal fermentation, bacterial communities, and ruminal acidosis.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC2cXhtFShu7zE&md5=660789ea648535e4eedc8bd282481723CAS |
Greenwood JS, Auldist MJ, Marett LC, Hannah MC, Jacobs JL, Wales WJ (2014) Ruminal pH and whole-tract digestibility in dairy cows consuming fresh cut herbage plus concentrates and conserved forage fed either separately or as a partial mixed ration. Animal Production Science 54, 1056–1063.
| Ruminal pH and whole-tract digestibility in dairy cows consuming fresh cut herbage plus concentrates and conserved forage fed either separately or as a partial mixed ration.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC2cXhtlaks7%2FK&md5=2cb75b7916f0e7ea2de4fe81af44a155CAS |
Greenwood PL, Paull DR, McNally J, Kalinowski T, Ebert D, Little B, Smith DV, Rahman A, Valencia P, Ingham AB, Bishop-Hurley GJ (2017) Use of sensor-determined behaviours to develop algorithms for pasture intake by individual grazing cattle. Crop and Pasture Science
| Use of sensor-determined behaviours to develop algorithms for pasture intake by individual grazing cattle.Crossref | GoogleScholarGoogle Scholar |
Gregorini P (2012) Diurnal grazing pattern: its physiological basis and strategic management. Animal Production Science 52, 416–430.
| Diurnal grazing pattern: its physiological basis and strategic management.Crossref | GoogleScholarGoogle Scholar |
Heady EO (1952) ‘Economics of agricultural production and resource use.’ (Prentice Hall: Upper Saddle River, NJ)
Heard JW, Hannah M, Ho CKM, Kennedy E, Doyle PT, Jacobs JL, Wales WJ (2017) Predicting milk responses to cereal-based supplements in grazing dairy cows. Animal Production Science 57, 746–759.
| Predicting milk responses to cereal-based supplements in grazing dairy cows.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC2sXjs1Srtr0%3D&md5=a80de69a7bd451851b872a5e15659697CAS |
Hetti Arachchige A, Fisher AD, Auldist MJ, Wales WJ, Jongman EC (2013) Effects of different systems of feeding supplements on time budgets of cows grazing restricted pasture allowances. Applied Animal Behaviour Science 148, 13–20.
| Effects of different systems of feeding supplements on time budgets of cows grazing restricted pasture allowances.Crossref | GoogleScholarGoogle Scholar |
Hetti Arachchige AD, Fisher AD, Wales WJ, Auldist MJ, Hannah MC, Jongman EC (2014) Space allowance and barriers influence cow competition for mixed rations fed on a feed-pad between bouts of grazing. Journal of Dairy Science 97, 3578–3588.
| Space allowance and barriers influence cow competition for mixed rations fed on a feed-pad between bouts of grazing.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC2cXmtlCktbw%3D&md5=3e546c2850402ba399d82c9ecd126e76CAS |
Hills JL, Wales WJ, Dunshea FR, Garcia SC, Roche JR (2015) Invited review: an evaluation of the likely effects of individualized feeding of concentrate supplements to pasture-based dairy cows. Journal of Dairy Science 98, 1363–1401.
| Invited review: an evaluation of the likely effects of individualized feeding of concentrate supplements to pasture-based dairy cows.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC2MXnslyhtg%3D%3D&md5=43099e0679335245d595eb1af1fd2c4fCAS |
Ho CKM, Heard JW, Wales WJ, Jacobs JL, Doyle PT, Malcolm B (2017) Evaluating the economics of concentrate feeding decisions in grazing dairy cows. Animal Production Science
| Evaluating the economics of concentrate feeding decisions in grazing dairy cows.Crossref | GoogleScholarGoogle Scholar |
Hodgson J (1981) Variation in the surface characteristics of the sward and the short-term rate of herbage intake by calves and lambs. Grass and Forage Science 36, 49–57.
| Variation in the surface characteristics of the sward and the short-term rate of herbage intake by calves and lambs.Crossref | GoogleScholarGoogle Scholar |
Inside Dairy (2016) ‘Two wins from using a catch crop.’ (DairyNZ: Hamilton, New Zealand) Available at www.dairynz/publications/ [Verified 10 November 2016]
Ipharraguerre IR, Clark JH (2005) Varying protein and starch in the diet of dairy cows. II. Effects on performance and nitrogen utilization for milk production. Journal of Dairy Science 88, 2556–2570.
| Varying protein and starch in the diet of dairy cows. II. Effects on performance and nitrogen utilization for milk production.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2MXlsFCmsbY%3D&md5=580aceb4e485c2725202e5c0cd0ed4edCAS |
Jacobs JL (2014) Challenges in ration formulation in pasture-based milk production systems. Animal Production Science 54, 1130–1140.
Ketelaars JJ, Tolkamp MH, Tolkamp BJ (1992) Toward a new theory of feed intake regulation in ruminants 1. Causes of differences in voluntary feed intake: critique of different views. Livestock Production Science 30, 269–296.
| Toward a new theory of feed intake regulation in ruminants 1. Causes of differences in voluntary feed intake: critique of different views.Crossref | GoogleScholarGoogle Scholar |
Khorasani GR, Okine EK, Kennelly JJ (2001) Effects of substituting barley grain with corn on ruminal fermentation characteristics, milk yield, and milk composition of Holstein cows. Journal of Dairy Science 84, 2760–2769.
| Effects of substituting barley grain with corn on ruminal fermentation characteristics, milk yield, and milk composition of Holstein cows.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD38Xosl2rsQ%3D%3D&md5=5059df3b25b21aa8b5ab26945389b7a6CAS |
Kolver ES, Muller LD (1998) Performance and nutrient intake of high producing Holstein cows consuming pasture or a total mixed ration. Journal of Dairy Science 81, 1403–1411.
| Performance and nutrient intake of high producing Holstein cows consuming pasture or a total mixed ration.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaK1cXjsVyjsrs%3D&md5=a05525dca4e64aec72ee27a23e2e103dCAS |
Leddin CM, Stockdale CR, Hill J, Heard JW, Doyle PT (2010) Increasing amounts of crushed wheat fed with Persian clover herbage reduced ruminal pH and dietary fibre digestibility in lactating dairy cows. Animal Production Science 50, 837–846.
| Increasing amounts of crushed wheat fed with Persian clover herbage reduced ruminal pH and dietary fibre digestibility in lactating dairy cows.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3cXht1ajt7%2FP&md5=fb6274e4fad60a465d2083ecb85e4e03CAS |
Lee JM, Hemmingson NR, Minnee EMK, Clark CEF (2012) Chicory and plantain management strategies to increase herbage dry matter yield, nutritive value and plant survival. In ‘Proceedings of the 5th Australasian dairy science symposium’. (Ed. JL Jacobs). pp. 393–397. (Australasian Dairy Science Symposium: Melbourne)
Li G, Kemp PD (2005) Forage chicory (Cichorium intybus L.): a review of its agronomy and animal production. Advances in Agronomy 88, 187–222.
| Forage chicory (Cichorium intybus L.): a review of its agronomy and animal production.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD1cXitlKisr0%3D&md5=883568b32e4b4d7aad175c282dd4722fCAS |
Macdonald K (2014) Driving production, profit and environment in all-grazed systems. In ‘Inside Dairy Technical Series (October) Issue 24’. pp. 1–4. (DairyNZ: Hamilton New Zealand) [Verified 11 October 2016]
Malcolm BJ, Cameron KC, Di HJ, Edwards GR, Moir JL (2014) The effect of four different pasture species compositions on nitrate leaching losses under high N loading. Soil Use and Management 30, 58–68.
| The effect of four different pasture species compositions on nitrate leaching losses under high N loading.Crossref | GoogleScholarGoogle Scholar |
Mayne CS, McGilloway DA, Cushnahan A, Laidlaw AS (1997) The effect of sward height and bulk density of herbage intake and grazing behavior of dairy cows. In ‘Proceedings of the XVIII international grassland congress’. Vol. 1, pp. 2-15–2-16. (Winnepeg, Manitoba, Canada)
McDonnell RP, Staines MvH, Edmunds BE, Morris R (2016) Feeding management, production and performance of 13 pasture-based dairy farms in a Mediterranean environment. Animal Production Science
| Feeding management, production and performance of 13 pasture-based dairy farms in a Mediterranean environment.Crossref | GoogleScholarGoogle Scholar |
Mertens DR (1987) Predicting intake and digestibility using mathematical models of rumen function. Journal of Animal Science 64, 1548–1558.
| Predicting intake and digestibility using mathematical models of rumen function.Crossref | GoogleScholarGoogle Scholar | 1:STN:280:DyaL2s3itF2kuw%3D%3D&md5=c470065bde74208601413935bae5414dCAS |
Milne JA, Hodgson J, Thompson R, Souter WG, Barthram GT (1982) The diet ingested by sheep grazing swards differing in white clover and perennial ryegrass content. Grass and Forage Science 37, 209–218.
| The diet ingested by sheep grazing swards differing in white clover and perennial ryegrass content.Crossref | GoogleScholarGoogle Scholar |
Ministry for the Environment and Statistics New Zealand (2015) ‘Environment Aotearoa 2015.’ Available at http://www.mfe.govt.nz/publications/environmental-reporting/environment-aotearoa-2015 [Verified 11 October 2016]
Minnee EMK, Clark CEF, McAllister TB, Hutchinson KJ, Lee JM (2012) Chicory and plantain as feeds for dairy cows in late lactation. In ‘Proceedings of the 5th Australasian dairy science symposium’, Melbourne, Australia. pp. 426–428.
Monaghan RM, de Klein CAM (2014) Integration of measures to mitigate reactive nitrogen losses to the environment from grazed pastoral dairy systems. The Journal of Agricultural Science 152, 45–56.
| Integration of measures to mitigate reactive nitrogen losses to the environment from grazed pastoral dairy systems.Crossref | GoogleScholarGoogle Scholar |
Moorhead AJE, Piggot GJ (2009) The performance of pasture mixtures containing ‘ceres tonic’ plantain (Plantago lanceolata) in Northland. In ‘Proceedings of the New Zealand Grassland Association’, Waitangi, New Zealand. pp. 195–199.
Murphy R, Simpson G (2014) ‘Balancing dairy production and profits in northern Australia, Queensland Dairy Accounting Scheme – 2014.’ (Animal Science, Department of Agriculture, Fisheries and Forestry: Gatton, Qld) Available at www.dairyaustralia.com.au [Verified 11 October 2016]
National Research Council (2001) ‘Nutrient requirements of dairy cattle.’ (National Academy Press: Washington, DC)
Olmos Colmenero JJ, Broderick GA (2006) Effect of dietary crude protein concentration on milk production and nitrogen utilization in lactating dairy cows. Journal of Dairy Science 89, 1704–1712.
| Effect of dietary crude protein concentration on milk production and nitrogen utilization in lactating dairy cows.Crossref | GoogleScholarGoogle Scholar |
Pembleton KG, Tozer KN, Edwards GR, Jacobs JL, Turner LR (2015) Simple versus diverse pastures: opportunities and challenges in dairy systems. Animal Production Science 55, 893–901.
| Simple versus diverse pastures: opportunities and challenges in dairy systems.Crossref | GoogleScholarGoogle Scholar |
Pembleton KG, Hills JL, Freeman MJ, McLaren DK, French M, Rawnsley RP (2016) More milk from forage: milk production, blood metabolites, and forage intake of dairy cows grazing pasture mixtures and spatially adjacent monocultures. Journal of Dairy Science 99, 3512–3528.
| More milk from forage: milk production, blood metabolites, and forage intake of dairy cows grazing pasture mixtures and spatially adjacent monocultures.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC28XjsVams7k%3D&md5=9a8dbf669c4d35316dd4c4116bf2c8f8CAS |
Penning PD, Parsons AJ, Orr RJ, Treacher TT (1991) Intake and behaviour responses by sheep to changes in sward characteristics under continuous stocking. Grass and Forage Science 46, 15–28.
| Intake and behaviour responses by sheep to changes in sward characteristics under continuous stocking.Crossref | GoogleScholarGoogle Scholar |
Provenza FD (1995) Postingestive feedback as an elementary determinant of food preference and intake in ruminants. Journal of Range Management 48, 2–17.
| Postingestive feedback as an elementary determinant of food preference and intake in ruminants.Crossref | GoogleScholarGoogle Scholar |
Rogers ME, Lawson AR, Kelly KB (2016) Lucerne yield, water productivity and persistence under variable and restricted irrigation strategies. Crop and Pasture Science 67, 563–573.
| Lucerne yield, water productivity and persistence under variable and restricted irrigation strategies.Crossref | GoogleScholarGoogle Scholar |
Romera AJ, Doole GJ, Khaembah EN (2015) Value of chicory crops to pasture-based dairy farms in the Waikato region of New Zealand. The Journal of Agricultural Science 153, 124–137.
| Value of chicory crops to pasture-based dairy farms in the Waikato region of New Zealand.Crossref | GoogleScholarGoogle Scholar |
Rook AJ (2000) Principles of foraging and grazing behavior. In ‘Grass, its production and utilization’. (Ed. A Hopkins) pp. 229–246. (Blackwell Science: Oxford, UK)
Selbie D, Buckthought LE, Shepherd MA (2015) The challenge of the urine patch for managing nitrogen in grazed pasture systems. Advances in Agronomy 129, 229–292.
| The challenge of the urine patch for managing nitrogen in grazed pasture systems.Crossref | GoogleScholarGoogle Scholar |
Shingfield KJ, Bernard L, Leroux C, Chilliard Y (2010) Role of trans fatty acids in the nutritional regulation of mammary lipogenesis in ruminants. Animal 4, 1140–1166.
| Role of trans fatty acids in the nutritional regulation of mammary lipogenesis in ruminants.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3cXmvVajtrw%3D&md5=f1ad0c248764b0c3e07bdadac8a4fc8bCAS |
Starks PJ, Zhao D, Phillips WA, Coleman SW (2006) Herbage mass, nutritive value and canopy spectral reflectance of bermudagrass pastures. Grass and Forage Science 61, 101–111.
| Herbage mass, nutritive value and canopy spectral reflectance of bermudagrass pastures.Crossref | GoogleScholarGoogle Scholar |
Stockdale CR, Callaghan A, Trigg TE (1987) Feeding high energy supplements to pasture-fed dairy cows, effects of stage of lactation and level of supplement. Australian Journal of Agricultural Research 38, 927–940.
| Feeding high energy supplements to pasture-fed dairy cows, effects of stage of lactation and level of supplement.Crossref | GoogleScholarGoogle Scholar |
Thorrold B, Doyle PT (2007) Nature or nurture: forces shaping the current and future state of dairy farming in New Zealand and Australia. In ‘Proceedings of the 3rd Australasian dairy science symposium’, Melbourne, Australia. pp. 450–460.
Totty V, Greenwood S, Bryant R, Edwards G (2013) Nitrogen partitioning and milk production of dairy cows grazing simple and diverse pastures. Journal of Dairy Science 96, 141–149.
| Nitrogen partitioning and milk production of dairy cows grazing simple and diverse pastures.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC38XhvV2rsb3M&md5=b430041cb3af1fc45d37f2dcf13b2b2fCAS |
Ungar ED (1996) Ingestive behavior. In ‘The ecology and management of grazing systems’. (Eds J Hodgson, AW Illius) pp. 185–218. (CAB International: Wallingford, UK)
Verite R, Journet M (1970) Influence de la teneur en eau et la deshydration de l’herbe sur la valeur alimentaire por les vaches laitiers. Annales de Zootechnie 19, 255–268.
| Influence de la teneur en eau et la deshydration de l’herbe sur la valeur alimentaire por les vaches laitiers.Crossref | GoogleScholarGoogle Scholar |
Wales WJ, Stockdale CR, Doyle PT (2005) Plant and sward characteristics to achieve high intake in ruminants: In ‘Utilisation of grazed grass in temperate animal systems. Proceedings of a satellite workshop of the XXth international grassland congress’, July 2005, Cork, Ireland. pp. 37–47.
Wales WJ, Marett LC, Greenwood JS, Wright MM, Thornhill JB, Jacobs JL, Ho CKM, Auldist MJ (2013) Use of partial mixed rations in pasture-based dairying in temperate regions of Australia. Animal Production Science 53, 1167–1178.
| Use of partial mixed rations in pasture-based dairying in temperate regions of Australia.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3sXhsFGgsrvK&md5=8d84f026e062c3b20e320a93d4becdf8CAS |
Waugh CD, Clark DA, Harris SL, Thom ER, Copeman PJA, Napper AR (1998) Chicory for milk production. In ‘Proceedings of the New Zealand Grassland Association’, Nelson, New Zealand. pp. 33–37.
Weston RH (1982) Animal factors affecting feed intake. In ‘Nutritional limits to animal production from pastures’. (Ed. JB Hacker) pp. 183–198. (Commonwealth Agricultural Bureaux: Farnham Royal, UK)
Williams YJ, Walker GP, Doyle PT, Egan AR, Stockdale CR (2005) Rumen fermentation characteristics of dairy cows grazing different allowances of Persian clover- or perennial ryegrass-dominant swards in spring. Australian Journal of Experimental Agriculture 45, 665–675.
| Rumen fermentation characteristics of dairy cows grazing different allowances of Persian clover- or perennial ryegrass-dominant swards in spring.Crossref | GoogleScholarGoogle Scholar |
Williams YJ, Doyle PT, Egan AR (2014) Diurnal variation in rumen fill of dairy cows grazing Persian clover at different pasture allowances. Animal Production Science 54, 1388–1393.
Woodward SL, Waghorn GC, Bryant MA, Benton A (2012) Can diverse pasture mixtures reduce nitrogen losses? In ‘Proceedings of the 5th Australasian dairy science symposium’, Melbourne, Australia. pp. 463–464.
Woodward S, Waugh C, Roach C, Fynn D, Phillips J (2013) Are diverse species mixtures better pastures for dairy farming? In ‘Proceedings of the New Zealand Grassland Association’, Tauranga, New Zealand. pp. 79–84.
Wright MM, Marett LC, Greenwood JS, Hannah M, Jacobs JL, Wales WJ, Auldist MJ (2014) Feeding a partial mixed ration once a day did not increase milk production compared with feeding grain in the dairy and forage in the paddock. Animal Production Science 54, 1405–1411.
| Feeding a partial mixed ration once a day did not increase milk production compared with feeding grain in the dairy and forage in the paddock.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC2cXhtlaktLnE&md5=499703477e918283379991f8256599f3CAS |
Wright MM, Auldist MJ, Kennedy E, Dunshea FR, Hannah M, Wales WJ (2016) Variation in feeding behavior and milk production among dairy cows when supplemented with 2 amounts of mixed ration in combination with 2 amounts of pasture. Journal of Dairy Science 99, 6507–6518.
| Variation in feeding behavior and milk production among dairy cows when supplemented with 2 amounts of mixed ration in combination with 2 amounts of pasture.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC28Xpt1emsLo%3D&md5=debaf5b47d0f1ff7cf4062e393179168CAS |