

The effect of water level on lateral movements of fish between river and off-channel habitats and implications for management
Jarod Lyon A B D , Ivor Stuart A C , David Ramsey A and Justin O’Mahony AA Arthur Rylah Institute for Environmental Research, Department of Sustainability and Environment Victoria, 123 Brown Street, Heidelberg, Vic. 3084, Australia.
B The Environment Institute and School of Earth and Environmental Sciences, University of Adelaide, Adelaide, South Australia 5005, Australia.
C Present address: Kingfisher Research, 20 Chapman Street, Diamond Creek, Vic. 3089, Australia.
D Corrresponding author. Email: jarod.lyon@dse.vic.gov.au
Marine and Freshwater Research 61(3) 271-278 https://doi.org/10.1071/MF08246
Submitted: 28 August 2008 Accepted: 26 June 2009 Published: 29 March 2010
Abstract
Off-channel habitats, such as wetlands and backwaters, are important for the productivity of river systems and for many species of native fish. This study aimed to investigate the fish community, timing and cues that stimulated movement to and from off-channel habitats in the highly regulated Lake Hume to Lake Mulwala reach of the Murray River, south-eastern Australia. In 2004–05, 193 712 fish were collected moving bi-directionally between a 50-km section of the Murray River and several off-channel habitats. Lateral fish movements approximated water level fluctuations. Generally as water levels rose, fish left the main river channel and moved into newly flooded off-channel habitats; there was bi-directional movement as water levels peaked; on falling levels fish moved back to the permanent riverine habitats. Fish previously classified as ‘wetland specialists’, such as carp gudgeons (Hypseleotris spp.), have a more flexible movement and life-history strategy including riverine habitation. The high degree of lateral movement indicates the importance of habitat connectivity for the small-bodied fish community. Wetlands adjacent to the Murray River are becoming increasingly regulated by small weirs and ensuring lateral fish movement will be important in maintaining riverine-wetland biodiversity.
Additional keywords: Australia, carp gudgeon, floodplain, Murray River.
Introduction
The regulation of rivers and their floodplains by dams and weirs has caused major changes in freshwater systems around the world (Walker et al. 1978; Nicola et al. 1996; Williams 1998). These impacts include altered flow regimes (including un-seasonal flooding and drying cycles), barriers to fish migration, thermal pollution, proliferation of invasive species, loss of in-stream habitat, loss of flow variability and decreases in primary productivity (Ricciardi and Rasmussen 1998; Nicol et al. 2004; Sherman et al. 2007). Over the past 20 years in Australia, there has been a shift away from studying the effect of in-stream barriers towards management and on-ground restoration techniques that may help ameliorate the problems. With most of continental Australia, in particular the south-east region, still subjected to the longest drought on record (2001–2010), rivers and their biota are showing signs of increasing stress (Bond et al. 2008). Consequently, river restoration is firmly entrenched on the social and political agenda with several major environmental rehabilitation initiatives underway.
For fish, many of the restoration programs concentrate on the main river channel (e.g. new fishways, re-introduction of large woody debris; Nicol et al. 2004; Barrett and Mallen-Cooper 2006). Off-channel habitats are, however, also important for fish populations because of the increased habitat diversity offered by floodplains, with heightened survival, feeding and reproduction opportunities (Junk et al. 1989; Zeug and Winemiller 2008). Improving lateral connectivity is also important for fish populations as floodplains provide feeding and nursery zones (Copp 1997; Castello 2008), and fish community structure, functioning and subsequent fishery production can relate to river–floodplain connectivity (Junk et al. 1989).
To date, in temperate Australia, the movements of small-bodied (<100 mm long) fish species between the main river channel and off-channel habitats, such as shallow wetlands, has received relatively little attention. The terminology used in some of the management literature describes small-bodied fish as ‘wetland specialists’ or ‘generalists’, whereas some large-bodied fish are ‘main channel specialists’ based on their early life-history (McCarthy et al. 2006). However, it is likely that many small-bodied fish regularly move between off-channel habitats and the main river channel, and small-bodied fish can numerically dominate floodplain and riverine fish communities. The degree to which fish move between these separate habitats is of particular importance in managed river systems where water-saving initiatives can involve construction of wetland regulators that may impede fish movements (Jungwirth 1998; Jones and Stuart 2008).
The aims of this study were to investigate the river water level cues that stimulated fish movement to and from off-channel habitats. We hypothesised that river water levels influence the abundance and community composition of small-bodied native and non-native fish moving between the main river and off-channel habitats. We also predicted more fish to move at night rather than day to avoid predation.
Materials and methods
Study area
The study area was in south-eastern Australia, on the upper Murray River between Lake Hume and Lake Mulwala (150 river-km). This reach is one of the most highly degraded sections of the entire 2200-km long Murray River due to the large amounts of irrigation water delivered between the two anthropogenic storage lakes. Hence, the Murray River between Lake Hume and Lake Mulwala essentially acts as a major irrigation conduit for water between August and April each year, and as such, has an inverted flow regime (high in summer, low in winter), which provides unseasonal connections between the river and its floodplain. With an annual irrigation release, the highly regulated reach of Murray River examined in the present study provides annual opportunities for fish to access adjacent anabranches, billabongs and other floodplain habitats.
A total of seven sites were sampled with four of the study sites (Dead River, Doolans Lagoon, Red Cliffs and S-lagoon) in the vicinity of the township of Howlong (35°58′36.123 S, 146°37′26.836 E). The remaining three sites (Lumbys Lagoon, Golf Course Wetland and Corowa Alcove) were near the township of Corowa (35°59′40.847 S, 146°23′27.780 E) (Fig. 1) ~30 river-km downstream. All sites had a direct association either with the Murray River or a major anabranch via a connecting channel.

|
Each site was chosen to represent the off-channel habitats in the study area (usually backwaters or wetlands) and were predominately shallow (<1.0 m depth) or occasionally deep (up to 2.4 m depth), productive wetlands and differ from main river habitats with their relatively shallow well-vegetated margins and slightly warmer and more turbid waters. The substrate of the wetlands was dominated by clay and sand. The type of off-channel habitat (backwater or wetland) and their relative importance as fish habitat was not specifically compared. The wetland-river connections were usually narrow (1–2 m wide) channels that commenced to flow following an increase in river level.
The Murray River in the study area is ~100 m wide and up to 5 m deep with a highly regulated discharge regime from the deepwater outlets on Hume Dam (3.06 GL), ~60 km upstream (Fig. 1). The average daily flows in this river reach can fluctuate between 1400 and 27 000 ML day–1 depending largely on the needs of downstream irrigators. The irrigation season runs primarily between August and May, which leads to high summer discharges and depressed winter discharges. This variability in river discharge causes the adjacent wetlands to connect and disconnect multiple times within an average irrigation season. There is also considerable thermal pollution in this reach with the water temperature depressed by 1–5°C in spring and summer (September to March; Sherman et al. 2007).
Sampling
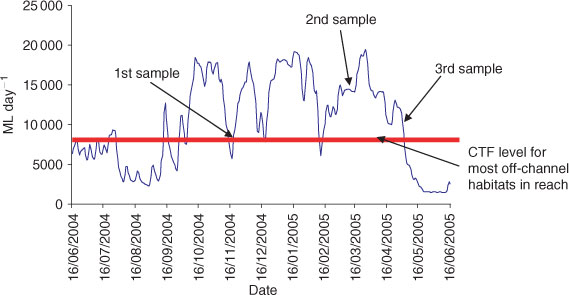
Sampling was undertaken between November 2004 and May 2005 to encompass a variety of water level conditions (Fig. 2). Each site was sampled during three different water levels to establish the use of each off-channel habitat by resident fishes and to determine any emigration/immigration. Due to the high variation (depending on irrigation requirements) in river flows in the study reach (between Lake Hume and Lake Mulwala), not all water level types were surveyed at all sites. The three surveyed water level types were: (i) rising water level (beginning of irrigation season and thereafter opportunistically); (ii) high water level (during irrigation season, water steady, i.e. neither rising or falling); and (iii) falling water level (end of irrigation season and opportunistically). These water levels were considered a surrogate for river discharge (ML day–1) and other inherent change in river conditions (e.g. water velocity).
|
Our assumption during sampling was that fish collected during the three sampling periods (rising, high and falling) were representative of those instantaneous river conditions and not those immediately antecedent. For example, in this heavily regulated river reach, water levels could fall between ‘rising’ and ‘high’ sampling events but fish collected were still assumed to reflect the capture conditions of the day. Our observations of fish led us to believe that these assumptions were appropriate as fish abundance appeared to quickly respond to new flow conditions. In addition, we did not assume that fish collected moving into the wetland during a rising flow were the same ones that moved out during a falling flow. River flows also provide artificial access for fish to floodplains, hence our observations might represent adaptations to local regulated conditions rather than natural seasonal patterns or movement behaviours in other parts of the Murray River.
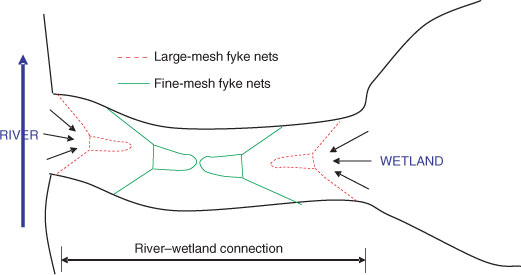
Two sets of double-wing fyke nets were set within the river–wetland connection of each off-channel habitat, facing bi-directionally, to catch fish moving in and out of the off-channel habitats. Large-mesh fyke nets (20-mm stretched mesh) were placed on the outsides (both river and wetland sides), with fine mesh fyke nets (0.5-mm stretched mesh) on the insides (Fig. 3). The large-mesh nets were set to catch large-bodied fish, while smaller species passed through these and were captured in the fine-mesh nets, thus the nets might collect the majority of fish sizes and species. Nets were checked as close as possible to dawn and dusk at each site to give an indication of diurnal patterns (Balcombe and Closs 2004; Baumgartner et al. 2008). Each site was sampled for 3–4 days (i.e. six to eight net checks) at each water level type.
|
All fish collected were individually weighed (nearest g) and measured (nearest mm) (fork length (FL) or total length (TL) depending on tail morphology), and were released well outside the net fleet in the direction they were originally going. In the case of large catches of small-bodied fish, a random sub-sample of 50 individuals of each species was weighed and measured individually, after which the remainder was weighed in bulk (by species). Some catches were so large that weighing the fish in bulk by species was not possible (i.e. more than 4000 fish) – in these cases a random sub-sample (100 fish) of the catch was taken to determine species composition (including weights and lengths of each individual in the sub-sample). The remainder of the sample was then weighed in bulk.
Statistical analyses
To test our two hypotheses, the effect of water level type (three levels – rising, high, falling), direction (two levels – moving into the off-channel habitats, moving out of the off-channel habitats) and time (two levels – day, night) on the number (count) of fish caught at a particular site were analysed using Poisson regression. The sampling effort at each site (amount of time each net was set) was fitted in the model as an offset, after taking natural logs. An offset is a term added to the model, with a known rather than an estimated coefficient. Including the sampling effort as an offset means that predictions from the model are equivalent to catch per unit effort (CPUE). An over-dispersion parameter was also included in the models to account for extra Poisson variation. We were not interested in differences in the mean responses among sites per se; rather, we were interested in inferences across all sites. Hence, site was fitted as a random effect in the model by fitting a random intercept for each site with the fixed effects being water level type, direction and time. All models were fitted in the software package R ver. 2.7.0 (R Development Core Team 2007) using the package lme4 (ver. 0.99875–9) (Bates 2007).
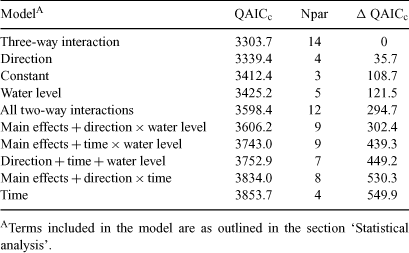
We fitted 10 models to the fish catch data with each model specifying a different combination of terms for the fixed effects. The most complex model included all main effects and interactions including the three-way interaction. The relative support for each of these models was assessed by calculating Akaike’s Information Criterion (AIC), corrected for small sample size and over-dispersion (QAICc) (Burnham and Anderson 1998). QAICc values were rescaled as differences between the model and the model with the lowest QAICc value (ΔQAICc). For the model with the most support, credible intervals were calculated by sampling from the posterior distribution of the parameters of the fitted model using Markov Chain Monte Carlo (MCMC) sampling (Gelman and Hill 2006). The 95% credible intervals were calculated from the posterior distribution containing 10 000 samples, after discarding a burn-in of 1000 samples.
Results
A total of 193 712 fish from 13 species were captured during the sampling period. The dominant species were carp gudgeons (Hypseleotris spp. (90.7%)), with 175 654 individuals collected (Table 1). In general, more fish were captured moving in and out of shallow wetlands than were caught moving in and out of backwaters or the deep wetland surveyed. The greatest abundance of fish were collected from one of the shallow wetlands, Lumbys Lagoon, with 54 027 native fish and 1221 non-native fish captured (Table 1). Sites at Doolans Lagoon and Dead River also revealed large numbers of fish with more than 35 000 native fish captured at each site, including eight large-bodied golden perch (Macquaria ambigua) at each. At Corowa Alcove, 29 258 native fish were captured over the sampling period, while at Red Cliffs only 1034 native fish were captured. However, Red Cliffs was only sampled on a rising water level.

|
Total catch model
To test the first hypothesis that river height influences the abundance index (CPUE) of small-bodied native and non-native fish moving between the main river and off-channel habitats, models were fitted to the total catch and also separately to catches of the three most abundant species, carp gudgeons, Australian smelt (Retropinna semoni) and Eastern Gambusia (Gambusia holbrooki). The other fish species had insufficient data for estimation. For the total catch, model selection indicated that the model including all possible interactions between water level type, direction and time had much higher support than any other model, with a difference in QAICc of 35.7 with the next best model (Table 2). Differences in QAICc of more than 10 indicate almost no support for the next best model (Burnham and Anderson 1998). Hence, this model indicated that the total CPUE differed depending on the direction (moving in or moving out of the off-channel habitats). However, the number of fish caught going in each direction was, in turn, dependent on river height (rising, steady, falling) and time (day or night) and our original hypothesis was supported.
|
Our second hypothesis that more fish move between the main river and off-channel habitats at night than at day, was not supported, because the highest CPUE occurred during the day compared with night. When the water level type was rising or steady, a higher CPUE was recorded for fish moving into the off-channel habitats compared with the CPUE for fish moving out. Conversely, when the water level type was falling, a higher CPUE was recorded for fish moving out of the off-channel habitats compared with fish moving in (Fig. 4).

|
Species models
For models fitted to the three fish species with sufficient data, model selection indicated that only the main effects of water level type, direction and time had the most support for both R. semoni and G. holbooki (ΔQAICc of 25.1 and 47.6 with the next best model, respectively). For Hypseleotris spp., the model including all possible interactions between water level type, direction and time was the most supported (ΔQAICc of 31.9 with the next best model). For R. semoni, the CPUE was highest during the day and when the water level type was rising, whereas for G. holbrooki, the CPUE was also highest during the day but when the water level type was falling (Fig. 4). For Hypseleotris spp., the patterns of CPUE were the same as those for the total catch.
Discussion
Lateral fish movement
In the present study of the upper Murray River, large numbers of fish were captured moving bi-directionally between the main-river and off-channel habitats. The highly regulated river reach provides annual opportunities for fish to access adjacent anabranches, billabongs and other floodplain habitats due to the managed irrigation season. The ecological purposes of the lateral fish migrations were not clarified but are probably related to the increased habitat diversity offered by floodplains with heightened survival, feeding and reproduction opportunities (Balcombe and Humphries 2006).
River height and lateral fish movement
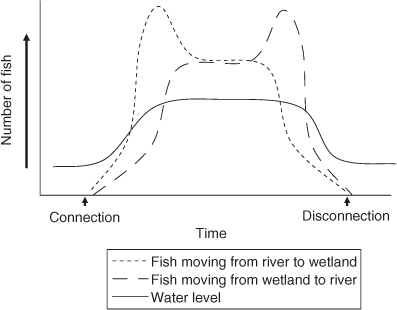
In the upper Murray River, lateral fish movement approximated water level fluctuations and generally as the Murray River rose, fish left the main river channel and moved into newly flooded off-channel habitats. However, there were also bi-directional fish movements as water levels peaked, and on falling water levels, movement back to the permanent riverine habitats occurred (Fig. 5). Hence, although water levels (as a surrogate measure of river flow) strongly influenced fish movement, the sampled populations still displayed a diversity of responses with some fish moving back to the Murray River during high river levels. The Murray River appears to act as more than a refuge from off-stream stranding but also as a regular habitation area.
|
Lateral movement of fish, in the Murray River, is dependent on the connecting channels between riverine and off-channel habitats in which movements of fish have occasionally been shown or hypothesised (Jones and Stuart 2008). Nevertheless, in many studies of fish populations in wetlands of south-eastern Australia, fish fauna are sampled either exclusively within the river or exclusively within the wetland. This historical sampling regime has inadvertently led to some of the management literature classifying fish as ‘wetland specialists’ (e.g. carp gudgeons and Australian smelt) or ‘generalist’ (e.g. Australian smelt and flat-headed gudgeons) species (McCarthy et al. 2006). Other management literature has highlighted the lack of a specialist riverine or wetland fish fauna and speculated that this probably reflects the naturally unpredictable nature of Australia’s floodplain systems, dry climate and recent marine origins of the native fish fauna (Darlington 1957; Gehrke and Harris 2000). Our results, for the species sampled, also suggest little evidence for distinct river- or wetland-only habitat preferences but emphasise the importance of wetland and main-channel interactions.
Typically, the wetland- and river-dwelling fish collected in this study also showed strong associations in the river–wetland connection, indicating large exchanges of biomass with the main channel (shown diagrammatically in Fig. 5). This observation is supported by longitudinal movement of thousands of Australian smelt, carp gudgeons and unspecked hardyhead in new fishways on the Murray River (Stuart et al. 2008). These fish species, and others, have occasionally been managed as ‘wetland specialists’ but actually appear to have a more flexible movement and life-history strategy including riverine habitation.
Similar trends have been observed in rivers and streams on other continents. Roach et al. (2009), using stable isotope analysis, found that lateral movement of fishes between channel and floodplain habitats in Venezuela was an important factor in the net transfer of organic carbon. Similarly, Lasne et al. (2007) conclude that there is a need for a high level of connectivity between the main channel and the floodplain in the Loire River to conserve native fish diversity. In particular, they indicated that the number of native species increased with increasing connectivity, while the number of exotic species present increased with increasing isolation from the main channel. In North America, Sullivan and Watzin (2009) describe that intact channel floodplain connections are critical for providing persistent floodplain and main channel for fish communities.
Many small-bodied fish appeared to return to the Murray River, particularly on a falling river, while others remained in the off-channel habitats where in the semi-permanent billabongs they might have high survival while exploiting the floodplain resources (Balcombe and Humphries 2006). Under natural flow conditions, opportunities to access off-channel habitats might have been less frequent but the time taken before disconnection would have been longer, allowing fish more time to escape stranding. For small-bodied fish, there is a trade-off between re-entering the permanent refuge of the main river channel and remaining in billabongs, which occasionally dry (Suarez et al. 2004). The current drought (8+ years) has resulted in the complete drying of many off-channel habitats and lateral re-population of billabongs from the Murray River is likely to be an important aspect of drought recovery. Hence, the exchange of fish between river and floodplain is likely important in maintaining biodiversity.
Some small-bodied native fish appear to recruit more strongly following flooding, (i.e. the flood pulse concept; Junk et al. 1989). In contrast to this was the high abundance of short-lived (<4 years), small-bodied fish collected in the Murray River during the present drought (Stuart et al. 2008). Hence, to maintain local populations, movement and recruitment must also necessarily occur during the frequent drought events (recently up to 8 years) or during the annual within-channel spring irrigation flows. For short-lived fishes, annual access to key low-lying wetlands is still likely to be important in maximising recruitment and population recovery following drought. For this reason, annual irrigation flows that are highly managed might still provide considerable benefits for small-bodied fish populations.
The narrow floodplain–river connecting channels were important for fish moving between these separate riverine and floodplain habitats. In the Murray–Darling Basin, flow through these channels has been historically controlled by large numbers of earthen levees to control flooding (Dexter et al. 1986). More recently, the drought has caused river managers to disconnect some wetlands from the main river to limit evaporative water loss. For some wetlands, new floodplain regulators are proposed and these would be used to cause a managed inundation event, primarily for riparian tree health. In these regulated systems, fish can be adversely impacted by floodplain regulation and maintaining lateral connectivity is likely to be important for river–floodplain fish assemblage biodiversity (Jones and Stuart 2008).
Diel and water-quality cues
Several fish species showed strong diel behaviour and this appears to be one of the underlying factors triggering bi-directional movement to and from off-channel habitats. When the diel patterns of individual fish link with population level responses, there can be changes to whole fish communities over the daily cycle and this information might be important in the management of rivers and their wetlands (Mallen-Cooper 1999; Baumgartner et al. 2008). For example, drawdown of rivers and closing of floodplain regulators to dry off-channel habitats should be managed operationally within the appropriate diel context to minimise stranding of fish (Hohausova et al. 2003; Jones and Stuart 2008).
Movement of fish into the off-channel habitats continued as the floodplain and river levels were declining, and this suggests other movement cues. The occasional temperature depression in the main river, caused by a nearby large dam, might have cued fish to enter the warmer floodplain environments. Alternatively, the plume of nutrient- and food-rich floodplain waters entering the river might also have stimulated fish to move off-stream. Such lateral movements of small-bodied fish may be in response to a combination of abiotic water quality parameters and their specific life-history stage needs (feeding, spawning, refuge/dispersal) (Roach et al. 2009; Sullivan and Watzin 2009).
Several non-native fishes also moved from the Murray River into the off-channel habitats, which suggests that during seasonal low flows these species also utilise permanent refuges in the main river. On rising flows, Gambusia left the river channel and entered the floodplain where, in the warm shallow floodplains, they breed and can displace and impact upon native fishes (Wilson 2005). These observations are important for controlling pest fishes as the river–floodplain connecting channels are natural movement bottlenecks where there are predictable fish movements, opportunities to manipulate discharge and constricted sites for targeted control efforts (Jones and Stuart 2008).
Restoring connectivity for fish is also a significant aspect of drought recovery for uncommon or threatened species (e.g. olive perchlet, southern pygmy perch and Murray jollytail) that might occasionally disperse. Providing greater passage for small-bodied migratory fish also achieves a more holistic goal of restoring passage for all aquatic migratory biota (Northcote 1998) and increases available resources for larger-bodied species.
There are often restrictive longitudinal or lateral perceptions of fish demographics but our results again confirm that these gradients are intimately linked (Jungwirth et al. 2002). With floodplains becoming increasingly disconnected from their rivers, there remains a great challenge in maintaining riverine–wetland biodiversity. Meeting this goal might readily be achieved by restoring lateral river–floodplain connections and enhancing the natural exchange of fish. Managing valuable floodplain habitats as an integral and connected part of whole river systems will likely have great biodiversity benefits.
Acknowledgements
The authors thank Kris Pittman and Karl Pomorin for help with the surveys. Zeb Tonkin, Justin O’Connor and John Koehn (Arthur Rylah Institute), Martin Mallen-Cooper and two anonymous referees improved an earlier draft of the manuscript. This work was carried out under NSW DPI collecting permit F93/158. This project was funded under the Murray–Darling Basin Commission’s Living Murray Program. This project was carried out under Department of Sustainability and Environment ethics permit AEC 04/003 and under Adelaide University Animal Ethics approval number 0000009058.
Balcombe, S. R. , and Closs, G. P. (2004). Variation in carp gudgeon (Hypseleotris spp.) abundance in dense macrophytes. Journal of Freshwater Ecology 15, 389–395.
Baumgartner, L. , Stuart, I. , and Zampatti, B. (2008). Diel variation in accumulations of fish downstream of three weirs in a regulated lowland river. Journal of Fish Biology 72, 218–232.
Castello, L. (2008). Lateral migration of Arapaima gigas in floodplains of the Amazon. Ecology of Freshwater Fish 17, 38–46.
| Crossref | GoogleScholarGoogle Scholar |
Dexter, D. B. , Rose, H. J. , and Davies, N. (1986). River regulation and associated forest management problems in the River Murray red gum forests. Australian Forestry 49, 16–27.
Hohausova, E. , Copp, G. H. , and Jankovsky, P. (2003). Movement of fish between a river and its backwater: diel activity and relation to environmental gradients. Ecology of Freshwater Fish 12, 107–117.
| Crossref | GoogleScholarGoogle Scholar |
Jungwirth, M. , Muhar, S. , and Schmutz, S. (2002). Re-establishing and assessing ecological integrity in riverine landscapes. Freshwater Biology 47, 867–887.
| Crossref | GoogleScholarGoogle Scholar |
Nicol, S. J. , Lieschke, J. A. , Lyon, J. P. , and Koehn, J. D. (2004). Observations on the distribution and abundance of carp and native fish, and their responses to a habitat restoration trial in the Murray River, Australia. New Zealand Journal of Marine and Freshwater Research 38, 541–551.
Ricciardi, A. , and Rasmussen, J. B. (1998). Predicting the identity and impact of future biological invaders: a priority for aquatic resource management. Canadian Journal of Fisheries and Aquatic Sciences 55, 1759–1765.
| Crossref | GoogleScholarGoogle Scholar |
Zeug, S. C. , and Winemiller, K. O. (2008). Relationships between hydrology, spatial heterogeneity, and fish recruitment dynamics in a temperate floodplain river. River Research and Applications 24, 90–102.
| Crossref | GoogleScholarGoogle Scholar |
