

Wet-season effects on the distribution of juvenile pigeye sharks, Carcharhinus amboinensis, in tropical nearshore waters
Danielle M. Knip A C , Michelle R. Heupel B , Colin A. Simpfendorfer A , Andrew J. Tobin A and James Moloney BA Fishing and Fisheries Research Centre, School of Earth and Environmental Sciences, James Cook University, Townsville, Qld 4811, Australia.
B School of Earth and Environmental Sciences, James Cook University, Townsville, Qld 4811, Australia.
C Corresponding author. Email: danielle.knip@my.jcu.edu.au
Marine and Freshwater Research 62(6) 658-667 https://doi.org/10.1071/MF10136
Submitted: 15 June 2009 Accepted: 4 October 2010 Published: 24 June 2011
Journal Compilation © CSIRO Publishing 2011 Open Access CC BY-NC-ND
Abstract
Tropical nearshore environments are highly dynamic systems owing to extreme freshwater flow and flooding episodes that occur in wet-season months. We hypothesised that juvenile sharks in tropical nearshore waters respond to seasonal freshwater inflow by moving away from areas of strong flow. An array of fifty-eight acoustic receivers deployed in Cleveland Bay, north Queensland, Australia, passively tracked thirty-two juvenile pigeye sharks, Carcharhinus amboinensis, throughout two wet seasons from 2008 to 2010. Influences associated with wet seasons appeared to play a role in habitat use by juvenile C. amboinensis in this region. Home ranges and distribution of individuals showed distinct changes, with individuals moving north away from sources of freshwater inflow during high flows. The location of individuals within the bay was strongly influenced by freshwater inflow in both years. Although juvenile C. amboinensis moved in response to freshwater inflow, home-range sizes remained stable, and the amount of space individuals used did not change in relation to freshwater inflow. By defining the response of juvenile sharks to highly variable freshwater flow events, this research provides useful information for understanding species behaviour in a dynamic and changing climate, and contributes towards effective management of tropical river systems.
Additional keywords: environmental effects, passive acoustic monitoring.
Introduction
Freshwater flow and flooding has a major impact on nearshore regions (e.g. bays, lagoons and estuaries) and can cause large fluctuations in the physical characteristics of an environment (Finlayson and McMahon 1988; Fraser 1997). Tropical regions are susceptible to increased amounts of freshwater impinging on these systems as a result of extreme rainfall events occurring in wet-season months (Balek 1983). Rainfall in the tropics is generally around 2000 mm year–1, with some regions experiencing up to 10 000 mm year–1 (Latrubesse et al. 2005). Such a significant input of freshwater causes high variability in the flows of tropical river systems, especially rivers in northern Australia, which are affected by annual monsoonal events (Balek 1983; Finlayson and McMahon 1988). Understanding the effects of extreme annual flow and flooding episodes on nearshore inhabitants is important, as freshwater flow influences species differently (Ter Morshuizen et al. 1996; Flannery et al. 2002), and may even cause mortality in some species (Whitfield and Paterson 1995).
Mobile fish species can modify their behaviour or physiology to cope with fluctuating environmental conditions, or may leave a region when conditions extend beyond their physiological limitations. For example, some teleost species move from deep to shallow water in response to low dissolved-oxygen concentrations (Pihl et al. 1991). Similar results have been found with sharks; for example, leopard sharks (Triakis semifasciata) leave nearshore regions during anoxic periods (Carlisle and Starr 2009) and bonnethead sharks (Sphyrna tiburo) leave estuaries when salinity declines (Ubeda et al. 2009). Few studies have, however, investigated the specific response of individual animals (e.g. movement and distribution) to seasonal variation (e.g. freshwater inflow) in tropical nearshore regions. Studies have found a general decrease in the abundance and diversity of fish species in tropical nearshore regions during wet-season events (Cyrus and Blaber 1992; Fraser 1997). One study specifically examined the movement and distribution of juvenile bull sharks (Carcharhinus leucas) in the estuarine portion of a Florida river and determined that individuals moved downriver with decreasing salinity and increasing freshwater flow rate (Heupel and Simpfendorfer 2008).
The pigeye shark, Carcharhinus amboinensis, inhabits tropical nearshore regions in the Indo–West Pacific, and in Australia it ranges throughout northern waters from Brisbane (Queensland) to Carnarvon (Western Australia) (Compagno 1984; Last and Stevens 2009). Size at birth is 60–65 cm, maximum size is ~280 cm and diet mainly consists of teleost fish, crustaceans, cephalopods and molluscs (Compagno 1984; Last and Stevens 2009). Unlike its close relative, the widely-distributed C. leucas, little is known about the movement of C. amboinensis and how it uses nearshore habitats. In northern Australia, C. amboinensis was reported to have relatively localised movements (Stevens et al. 2000; Last and Stevens 2009). In addition, C. amboinensis is thought to occasionally enter brackish water, and in South Africa individuals were reported using warm turbid waters close to large estuaries and river mouths (Cliff and Dudley 1991; Last and Stevens 2009). Tropical nearshore regions provide habitat for early life-stages of many populations of sharks (Heupel et al. 2007). Similar to other large-bodied shark species, juvenile C. amboinensis may use nearshore regions as nursery areas. Thus, understanding how this predator uses nearshore regions and what factors affect its movement and distribution is crucial to defining its early life-history and ecology within nearshore systems.
The purpose of this study was to determine how freshwater flows during the wet season affect movement and distribution of juvenile C. amboinensis in a tropical nearshore environment. Individual sharks were tracked over two consecutive years (two wet seasons) using passive acoustic monitoring. Home-range size, home-range location and distribution of individuals were examined in relation to freshwater inflow across seasons. With altered climate scenarios an increasing concern for tropical nearshore regions, a better understanding of species responses to environmental and seasonal variability is needed. The data from this study were used to test the hypothesis that juvenile C. amboinensis respond to increased freshwater inflow by shifting their distribution away from sources of strong flows.
Materials and methods
Study site
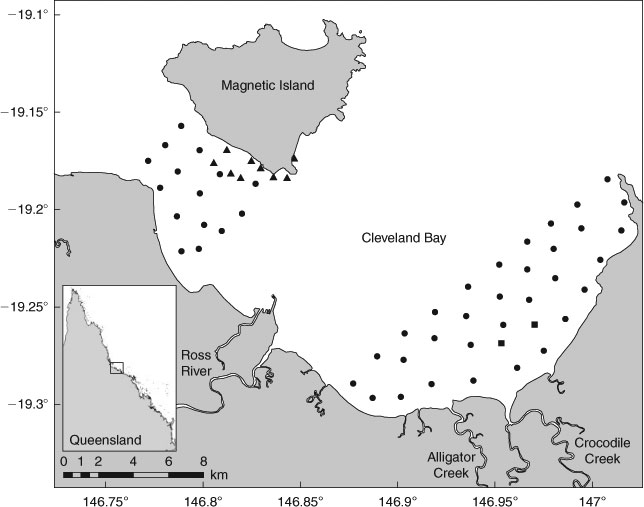
Cleveland Bay is a shallow and dynamic coastal habitat on the north-east coast of Australia adjacent to Townsville (Fig. 1). Most of the bay is <10 m deep and maximum tidal range reaches 4.2 m. The bay covers ~225 km2 and supports a diverse range of habitats. The main substrate is soft mud, but there are also small patches of coastal reefs, areas of seagrass (Cymodocea serrulata, Halodule uninervis and Halophila spp.) and the southern shore is lined with mangroves. Ross River, Crocodile Creek and Alligator Creek are the main river systems that run into the bay, and increased freshwater flow during the wet season causes strong currents and large decreases in salinity in the south-east portion of the bay.
|
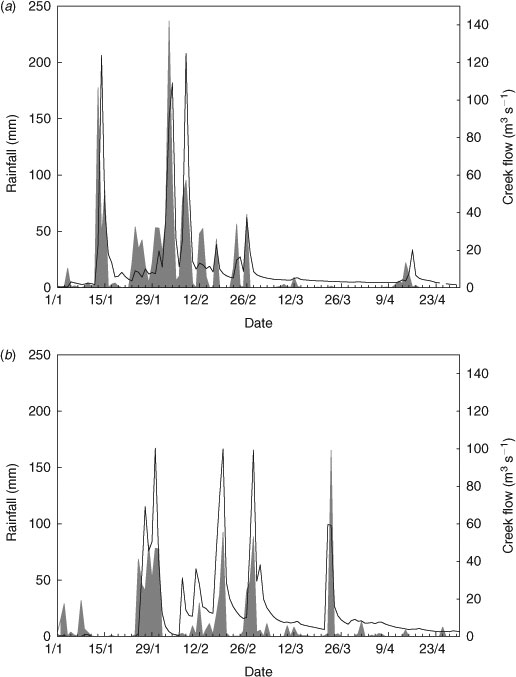
The wet season in northern Queensland is variable. Average annual rainfall in Townsville is around 1200 mm but has been higher over the past two years; Townsville received ~1830 mm of rainfall from December 2008 to April 2009, and 1460 mm of rainfall from December 2009 to April 2010 (Fig. 2). In 2009, 88% of total wet-season rainfall occurred in February and March, whereas in 2010, only 64% of wet-season rainfall occurred in these two months (Fig. 2). Freshwater inflow from creeks adjacent to Cleveland Bay is directly influenced by rainfall, with the largest flow rates occurring immediately after rainfall events (Fig. 2). Owing to the variability in rainfall between the two years of this study, the amount and pattern of freshwater inflow also varied. A major consequence of increased freshwater inflow during the wet season is decreased salinity in Cleveland Bay, and the lowest salinity levels have been recorded at times of highest freshwater inflow (Walker 1981).
|
Field methods
Monitoring of target species within the study site was conducted using passive acoustic monitoring. Forty-seven VR2 or VR2W acoustic receivers (Vemco Ltd, Canada) were deployed in Cleveland Bay in November 2008 (Fig. 1). To cover additional area and habitats, nine receivers were added to the array in August 2009, and two were added in March 2010 (Fig. 1). The detection range of V16 acoustic transmitters within the study site was ~900 m. The array included all representative habitats within the bay: reef, seagrass, sand, mud and river mouths. Coverage allowed target species to be monitored throughout all available habitat types. Downloading of data from receivers occurred every 6–8 weeks.
Sharks were captured with baited hooks on short long-lines. Long-lines were 500-m bottom-set mainlines (8-mm nylon rope) anchored at both ends and soaked for 1 h. Gangions consisted of 1 m of 5-mm nylon cord and 1 m of wire leader. Size 14/0 Mustad tuna circle hooks were used and baited with frozen butterfly bream (Nemipterus sp.), mullet (Mugil cephalus), blue threadfin (Eleutheronema tetradactylum) or fresh trevally (Caranx sp.). All captured sharks were identified, measured to the nearest mm, sexed and tagged with a rototag in the first dorsal fin and a single-barb dart tag in the dorsal musculature for identification before release. Juvenile C. amboinensis were also fitted with a V16 acoustic transmitter (Vemco Ltd, Canada), which was surgically implanted into the body cavity to ensure long-term retention. A small incision (3–4 cm) was made in the abdomen and the transmitter inserted. The wound was closed by suturing both the muscle and skin layers to ensure adequate wound closure and healing. Each transmitter had a unique code and emitted a pulse series at 69 kHz to identify each individual shark tagged. Transmitters pulsed on a random repeat interval of 45–75 s with a battery life of ~18 months.
Data analysis
Data collected from acoustic receivers included date, time and identity of individuals detected. Only data from acoustic receivers on the eastern side of the array were included in this analysis for two reasons: first, detections of C. amboinensis on the western array of receivers were rare, and second, the western array did not cover areas of high freshwater inflow. Locations for sharks in the bay were estimated every 30 min using a mean-position algorithm that provided an individual’s centre of activity (COA) (Simpfendorfer et al. 2002). COA locations represent an individual’s mean position for the set time-step. For small sharks, the error of COA estimates in relation to real-time locations has been calculated to be ~225 m (Simpfendorfer et al. 2002).
Home ranges of individual C. amboinensis were calculated based on COA estimates using 95 and 50% kernel utilisation distributions (95 and 50% KUD) with the adehabitat package in R (Calenge 2006). Home ranges were calculated at weekly and monthly intervals to examine changes in distribution and habitat use over time. Home ranges were plotted in ArcView 3.3 (Esri Products, Redlands, CA, USA) to show spatial and temporal distribution patterns of individual sharks.
To examine the effects of a tropical wet season on the movement of juvenile C. amboinensis and to remain consistent between years, data were analysed from December to April for two years (2008–2009 and 2009–2010). Freshwater inflows from creeks into Cleveland Bay are of similar magnitude, and data from Alligator Creek were used as flow data available at this site were continuous. The latitudes of COA locations of juvenile C. amboinensis were used to represent the distribution of animals in the bay relative to the mouths of the main sources of freshwater inflow, which were all on the southern boundary of the bay. Mean weekly latitude locations and home-range sizes of C. amboinensis were compared and correlated against creek flow.
The normality of the data (both latitude locations and home ranges) was checked by examination of quantile–quantile plots, and in all cases the data satisfied the assumptions for parametric testing. Tests of regression analysis demonstrated that there was no significant difference between fitting linear, polynomial, or segmented models to the data. The strength of each model was the same, and a linear model was used for further statistical testing of the data. Analysis of covariance (ANCOVA) was used to test for differences in latitude location and home-range size of C. amboinensis between age cohorts and years. Analysis of variance (ANOVA) was used to test for significance of creek flow on latitude location and home-range size of C. amboinensis. ANOVA was also used to test for differences in latitude location of C. amboinensis across dry season weeks. All data analyses were conducted in R (R Development Core Team 2009).
Results
Thirty-two juvenile C. amboinensis were released with acoustic transmitters in Cleveland Bay from 2008 to 2010 (Table 1). Young-of-the-year individuals were released in February 2009 (n = 15; 5 female, 10 male) and February–March 2010 (n = 13; 5 female, 8 male). One-year-old individuals were released in December 2008 (n = 2; 2 male) and December 2009 (n = 2; 2 male). Two young-of-the-year individuals released in 2009 were present in the bay the following year and were therefore included in the one-year-old cohort for the second year of the study. No effort was made to have an equal ratio of males and females and transmitters were fitted to animals regardless of sex. The size range of young-of-the-year individuals was 63–82 cm stretch total length (STL) and the size range of one-year-old individuals was 92.5–100 cm STL. All C. amboinensis individuals were sexually immature, and young-of-the-year individuals were identified by an umbilical scar (either open or closed).

|
Distribution and freshwater inflow
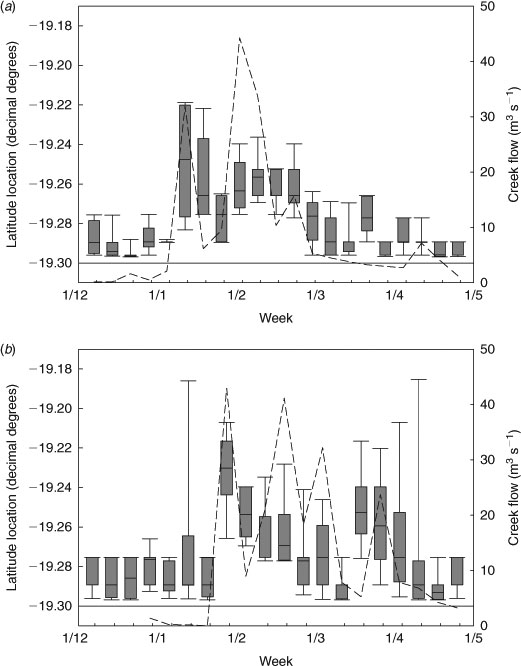
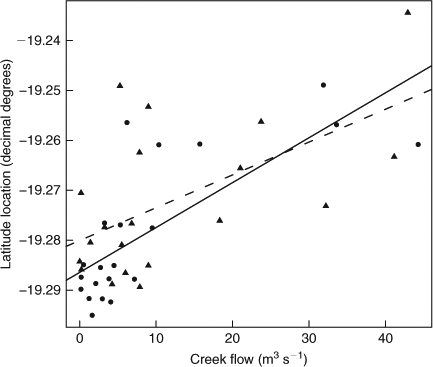
Distributional shifts of juvenile C. amboinensis during wet-season months occurred at times of greatest freshwater inflow from Alligator Creek (Fig. 3). During periods of high freshwater inflow, juvenile C. amboinensis responded by moving north away from the creek mouths (Fig. 3). As freshwater inflow decreased following the wet season, juvenile C. amboinensis responded by returning to the southern portion of the bay adjacent to the creek mouths (Fig. 3). Regression analysis showed that creek flow strongly influenced latitude location of sharks in the bay (r2 = 0.441, P < 0.001) (Fig. 4). Although the residuals appeared non-uniform with regression analysis, the assumption of linearity was met (F3,58 = 14.530, P < 0.001). There was no significant difference in mean weekly latitude location between cohorts in a year (2008–2009: F1,28 = 0.040, P = 0.843; 2009–2010: F1,26 = 0.135, P = 0.717) and freshwater inflow did not affect mean weekly latitude location of cohorts differently (2008–2009: F1,28 = 3.569, P = 0.069; 2009–2010: F1,26 = 0.689, P = 0.414), indicating that freshwater inflow affected young-of-the-year and one-year-old individuals in the same manner. Although the relationship between freshwater inflow and latitude location was stronger in 2008–2009, freshwater inflow was a significant factor influencing the location of juvenile C. amboinensis in Cleveland Bay in both years (2008–2009: F1,19 = 26.190, P < 0.001; 2009–2010: F1,18 =9.194, P = 0.007). There was no significant difference in the effect of freshwater inflow on latitude location of juvenile C. amboinensis between years (F1,37 = 1.178, P = 0.285).
|
|
Home range
Home ranges of juvenile C. amboinensis typically encompassed areas in the southern portion of Cleveland Bay adjacent to the creek mouths (Fig. 5). During February (the peak of the wet season), however, core home ranges (50% KUDs) shifted away from the creek mouths, and total home ranges (95% KUDs) included more area in the northern portion of the bay (Fig. 5). After the wet season (April), home-range analysis indicated that juvenile C. amboinensis remained in the southern portion of the bay (Fig. 5). Small portions of 95% KUDs overlapped with land in some individual cases (Fig. 5), suggesting some slight overestimations of the amount of space used. Home-range location did not change outside wet-season weeks. Juvenile C. amboinensis remained in the southern portion of the bay and there was no significant difference in latitude location between dry season weeks (F9,9 = 1.057, P = 0.468).
|
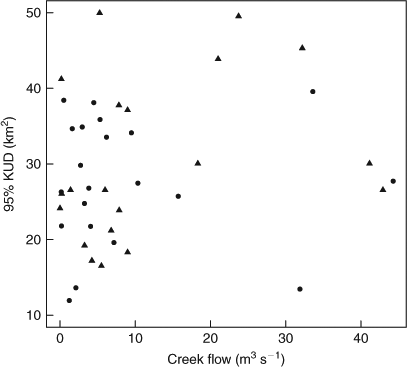
Although juvenile C. amboinensis shifted location in response to freshwater inflow, total weekly home-range size remained stable (mean: 29.05 km2). Regression analysis revealed no relationship between mean weekly home-range size and creek flow (r2 = 0.009, P = 0.249) (Fig. 6). There was no significant difference in mean weekly home-range size between cohorts in a year (2008–2009: F1,28 = 0.126, P = 0.726; 2009–2010: F1,26 = 1.149, P = 0.294), young-of-the-year and one-year-old individuals used the same amount of space, and freshwater inflow did not affect home-range size of cohorts differently (2008–2009: F1,28 = 2.108, P = 0.158; 2009–2010: F1,26 = 1.383, P = 0.250). Freshwater inflow did not significantly influence home-range size of juvenile C. amboinensis in Cleveland Bay, with mean total weekly home-range size in the wet season (31.56 km2) similar to that in the dry season (26.15 km2) (2008–2009: F1,19 = 0.391, P = 0.539; 2009–2010: F1,18 = 3.434, P = 0.080), thus indicating that these animals used the same amount of space regardless of freshwater inflow.

|
Discussion
Effects of freshwater inflow
The results of this study demonstrate that juvenile C. amboinensis responded to wet-season freshwater flows by changing the way they used Cleveland Bay. Although the same amount of space was used, their location within this nearshore habitat showed a distinct seasonal change. This response to freshwater inflow is similar to that reported for other shark species, such as S. tiburo in central Florida (Ubeda et al. 2009). The response of sharks to these changing conditions suggests a lack of tolerance for lower salinity levels, a preference for alternative conditions, or some other ecological factor.
Increased freshwater inflow may cause decreased salinity levels in nearshore environments and in Cleveland Bay a strong inverse relationship has been reported between freshwater inflow and salinity (Walker 1981). During this study, salinity levels in Cleveland Bay generally remained between 30 and 35, but in the wet season salinity dropped as low as 8 (surface) and 15.5 (bottom) near the creek mouths (D. Knip, unpubl. data). Thus, it is possible that the distribution of juvenile C. amboinensis was in part influenced by the low salinity levels that occurred during wet-season months. However, the closely related C. leucas is a euryhaline species, with juveniles showing an affinity for low salinities ranging from 7 to 17 within estuarine environments (Simpfendorfer et al. 2005; Heupel and Simpfendorfer 2008). Being close relatives, it could be presumed that C. amboinensis has similar physiological capabilities and behavioural characteristics to C. leucas. Although previous studies have reported C. amboinensis using brackish water and turbid areas adjacent to creek and river mouths (Cliff and Dudley 1991), this species has not been found to penetrate freshwater. Also unlike C. leucas, juvenile C. amboinensis appeared to avoid creeks and rivers during times of high freshwater inflow. For example, juvenile C. leucas moved down river in a Florida estuary with increasing freshwater flow, but individuals continued using estuarine habitat even at times of high flow (>113 m3 s–1) (Heupel and Simpfendorfer 2008). At similar flow rates, juvenile C. amboinensis moved away from creek mouths and used deeper marine areas. Further, in February 2009 when juvenile C. amboinensis were spatially displaced within Cleveland Bay, young-of-the-year C. leucas were captured in fishing nets in both Alligator and Crocodile Creeks (A. Tobin, unpubl. data). Therefore, the results of this study suggest that C. amboinensis is less tolerant of high freshwater inflow than C. leucas, and provide evidence that low salinity levels (e.g. 7–17) may be beyond the physiological limits of C. amboinensis.
Variation in rainfall during this study resulted in different freshwater inflow patterns between years. The total amount of freshwater inflow was similar in both years, with maximum flow occurring in February, but in 2010, high freshwater inflow continued throughout March. The movement response from juvenile C. amboinensis may not only depend on the total amount of freshwater inflow, but the rate of inflow as well. A large volume of freshwater inflow concentrated in a shorter time period (i.e. 2009) resulted in a stronger movement response from juvenile C. amboinensis. In 2009, there were only two large peaks of freshwater inflow in a short period of time and the response of juvenile C. amboinensis was strong, with individuals moving away from the southern portion of the bay until high freshwater inflow ceased. Because there were, however, multiple peaks of freshwater inflow spread out over a longer period of time in 2010, the relationship between flow and distribution of juvenile C. amboinensis was weaker. Individuals still moved away from creek mouths during times of highest freshwater inflow, but distribution was more spread out over time and some individuals continued using northern portions of the bay into the month of April. In both years, however, individuals displayed a strong association with creek mouth habitat, consistently returning to and remaining in southern portions of the bay after times of high freshwater inflow.
Moving in response to strong currents and flows has been reported in other shark species. For example, juvenile sandbar sharks (Carcharhinus plumbeus) inhabiting estuaries in the north-west Atlantic moved with tidal flow and showed greatest straight-line movement when currents were strongest (Medved and Marshall 1983; Wetherbee and Rechisky 1999). In a south-west Florida estuary, juvenile blacktip sharks (Carcharhinus limbatus) also moved with tidal flow (Steiner and Michel 2007), and was considered a possible strategy to minimise energetic costs associated with swimming in strong currents. It is possible that juvenile C. amboinensis moved away from creek mouths during times of high freshwater inflow to avoid strong currents and preserve energy. Unlike C. amboinensis and these other examples, juvenile C. leucas were not found to swim with tidal flow and movements of young individuals were random and not influenced by strong currents (Steiner and Michel 2007). These examples suggest that the behaviour of juvenile C. amboinensis is more similar to other young carcharhinid sharks than to the closely related C. leucas.
The greatest northward movement of juvenile C. amboinensis away from creek mouth habitat in both years occurred during the first peak of freshwater inflow. In tropical environments, the wet season occurs during summer, which is when water temperature is highest. Increased water temperature results in increased biological productivity (i.e. algal blooms), which in turn decreases dissolved oxygen concentrations and may even cause anoxic conditions in creeks, rivers and estuaries (NOAA 1998; Perna and Burrows 2005). It is likely that the first flooding event of a wet season pushes low quality water with low dissolved oxygen concentrations into nearshore regions. Thus, the first freshwater flow probably had the largest impact on the physical environment of Cleveland Bay, explaining why juvenile C. amboinensis had the greatest movement response with the first peak of freshwater inflow. Dissolved oxygen has been found to influence the movement and distribution of other shark species. For example, juvenile C. leucas using an estuarine habitat were caught in higher abundances in areas of high dissolved oxygen (Heithaus et al. 2009) and low dissolved oxygen caused the exodus of T. semifasciata from shallow nearshore environments (Carlisle and Starr 2009). It is possible that lower dissolved oxygen concentrations during times of increased freshwater inflow may in part influence the movement of juvenile C. amboinensis away from shallow nearshore waters adjacent to creek habitats.
Displacement and space use
High freshwater flow rates can be a disturbance for some inhabitants of rivers, creeks and estuaries. Mobile fish species may respond by leaving areas at times of high flow, and it is common for the community structure of these nearshore environments to vary seasonally (Rayner et al. 2008). Fish generally leave an area of high flow to seek refuge from high flow rates or to target prey items that have also left the high flow environment (Winemiller and Jepsen 1998). Thus, it is possible that the increased freshwater inflow that occurred during this study moved fish species from adjacent creeks into Cleveland Bay. Juvenile C. amboinensis may have moved away from the creeks at times of high freshwater inflow to follow target prey species that also left the creek habitat at that time. Changes in the movement of predators in relation to prey distribution as a result of wet-season events have been documented with other tropical species, such as the water python (Liasus fuscus) (Madsen and Shine 1996). Thus, predator species may change their behaviour in relation to prey distribution, and sharks have been reported to show an association with habitats where prey is most abundant (Heithaus et al. 2002).
Although juvenile C. amboinensis moved in response to increased freshwater inflow and used areas outside of creek mouth habitat during the wet season, the amount of space individuals used remained consistent. Some predatory fish species have been documented to move greater distances and use more space at times of high freshwater flow (de Morais and Raffray 1999; Scruton et al. 2005). Increased movement and home-range expansion during the wet season was attributed to greater amounts of water available, and to individuals needing to move greater distances to locate adequate prey. The fact that habitat is similar throughout the eastern side of Cleveland Bay, and space use of juvenile C. amboinensis did not differ between wet and dry seasons, suggests that this region provides adequate food resources for this species in both seasons, even when individuals are using areas outside of creek mouth habitat.
Conclusions
This study is the first to document fine-scale movements of C. amboinensis and determine environmental variables that influence its distribution. It appears that juvenile C. amboinensis individuals associate with shallow creek mouth habitats in tropical nearshore regions, but are highly influenced by freshwater inflow and move away from creeks in response to increased flow rates during the wet season. Whether the main driver of this movement response is freshwater inflow itself or a result of freshwater inflow (e.g. changes in salinity, dissolved oxygen or prey distribution) is yet to be determined. Unexpectedly, the behaviour of C. amboinensis was unlike that of its close relative C. leucas, with juvenile C. amboinensis appearing to display a low tolerance for high-flow and low-salinity environments. This suggests that the physiological capabilities of C. amboinensis are different to that of C. leucas. Further studies analysing movement and habitat use of C. amboinensis in relation to different environmental factors (e.g. salinity and dissolved oxygen) are needed to determine the specific requirements and physiological limitations of this species. Additional future work should also include biological studies, such as defining how foraging behaviour or diet may shift with seasonal changes.
Tropical nearshore regions are highly dynamic environments with fluctuating conditions, and are predicted to experience further changes and become more variable owing to factors associated with climate change. In northern Australia, nearshore regions are most vulnerable to changes in rainfall patterns and freshwater inflow, which will alter the habitats, productivity and physical characteristics of these environments (Chin et al. 2010). The results of this study indicate that C. amboinensis responds strongly to changes in freshwater inflow, so is likely to be highly exposed to the effects of altered rainfall and changing salinity levels. Specifically, intensified rainfall and periods of flooding in tropical nearshore regions may result in this species having more sporadic and extreme movements (Chin et al. 2010). Uncertainty in future projections of rainfall in tropical nearshore regions presents a concern for species vulnerable to the effects of a changing climate. However, by defining the response of juvenile C. amboinensis to highly variable freshwater flow events, this research provides a better understanding of species behaviour and responses within a dynamic and changing environment. With an increased risk of changing conditions in nearshore regions, knowledge of species behaviours and capabilities is needed to implement effective management initiatives. Thus, the results of this study provide information that will be useful for both river regulation and control in tropical regions and predicting responses in species behaviour as a result of changing climate scenarios.
Acknowledgements
We thank the staff and students of the Fishing and Fisheries Research Centre, including A. Mapleston, A. Chin, J. White, O. Li, A. Harry and numerous other volunteers for providing assistance with this project; and the editors and two anonymous referees for their comments on the manuscript. Funding for this research was provided by the Australian Research Council and Great Barrier Reef Marine Park Authority (GBRMPA) awarded to M. R. Heupel and C. A. Simpfendorfer. Additional research funding was granted to D. M. Knip from the GBRMPA and James Cook University (JCU) School of Earth and Environmental Sciences (SEES). D. M. Knip was also supported by a JCU Postgraduate Research Scholarship co-funded by the SEES. Flow data from Alligator Creek was provided by the Department of Environment and Resource Management (Queensland, Australia) and Townsville rainfall data was obtained from the Australian Bureau of Meteorology. All research activities were conducted under GBRMPA permit #G09/29895.1 and Queensland Department of Primary Industries and Fisheries permit #90911. Treatment of all animals was conducted under ethical guidelines approved by JCU animal ethics #A1214.
References
Balek, J. (1983). ‘Hydrology and Water Resources in Tropical Regions.’ (Elsevier: Amsterdam.)Calenge, C. (2006). The package adehabitat for the R software: a tool for the analysis of space and habitat use by animals. Ecological Modelling 197, 516–519.
| The package adehabitat for the R software: a tool for the analysis of space and habitat use by animals.Crossref | GoogleScholarGoogle Scholar |
Carlisle, A. B., and Starr, R. M. (2009). Habitat use, residency, and seasonal distribution of female leopard sharks Triakis semifasciata in Elkhorn Slough, California. Marine Ecology Progress Series 380, 213–228.
| Habitat use, residency, and seasonal distribution of female leopard sharks Triakis semifasciata in Elkhorn Slough, California.Crossref | GoogleScholarGoogle Scholar |
Chin, A., Kyne, P. M., Walker, T. I., and McAuley, R. B. (2010). An integrated risk assessment for climate change: analysing the vulnerability of sharks and rays on Australia’s Great Barrier Reef. Global Change Biology 16, 1936–1953.
| An integrated risk assessment for climate change: analysing the vulnerability of sharks and rays on Australia’s Great Barrier Reef.Crossref | GoogleScholarGoogle Scholar |
Cliff, G., and Dudley, S. F. J. (1991). Sharks caught in the protective gill nets off Natal, South Africa. 5. The java shark Carcharhinus amboinensis (Muller and Henle). South African Journal of Marine Science 11, 443–453.
Compagno, L. J. V. (1984). FAO species catalogue, Volume 4. Sharks of the world; an annotated and illustrated catalogue of shark species known to date, Part 2. Carcharhiniformes. FAO Fisheries Synopsis 125, 461–463.
Cyrus, D. P., and Blaber, S. J. M. (1992). Turbidity and salinity in a tropical northern Australian estuary and their influence on fish distribution. Estuarine, Coastal and Shelf Science 35, 545–563.
| Turbidity and salinity in a tropical northern Australian estuary and their influence on fish distribution.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaK3sXis1Smsbg%3D&md5=816c2d3bea8ffc470160df0e0b797bafCAS |
de Morais, L. T., and Raffray, J. (1999). Movements of Hoplias aimara during the filling phase of the Petit-Saut dam, French Guyana. Journal of Fish Biology 54, 627–635.
Finlayson, B. L., and McMahon, T. A. (1988). Australia vs the world: a comparative analysis of streamflow characteristics. In ‘Fluvial Geomorphology of Australia’. (Ed. R. F. Warner.) pp. 17–40. (Academic Press: Sydney.)
Flannery, M. S., Peebles, E. B., and Montgomery, R. T. (2002). A percent-of-flow approach for managing reductions of freshwater inflows from unimpounded rivers to Southwest Florida estuaries. Estuaries 25, 1318–1332.
| A percent-of-flow approach for managing reductions of freshwater inflows from unimpounded rivers to Southwest Florida estuaries.Crossref | GoogleScholarGoogle Scholar |
Fraser, T. H. (1997). Abundance, seasonality, community indices, trends and relationships with physicochemical factors of trawled fish in upper Charlotte Harbor, Florida. Bulletin of Marine Science 60, 739–763.
Heithaus, M. R., Dill, L. M., Marshall, G. J., and Buhleier, B. M. (2002). Habitat use and foraging behaviour of tiger sharks (Galeocerdo cuvier) in a seagrass ecosystem. Marine Biology 140, 237–248.
| Habitat use and foraging behaviour of tiger sharks (Galeocerdo cuvier) in a seagrass ecosystem.Crossref | GoogleScholarGoogle Scholar |
Heithaus, M. R., Delius, B. K., Wirsing, A. J., and Dunphy-Daly, M. M. (2009). Physical factors influencing the distribution of a top predator in a subtropical oligotrophic estuary. Limnology and Oceanography 54, 472–482.
| 1:CAS:528:DC%2BD1MXhsVCrtrzN&md5=9eda3392ab64119db2216a2567ddd650CAS |
Heupel, M. R., and Simpfendorfer, C. A. (2008). Movement and distribution of young bull sharks Carcharhinus leucas in a variable estuarine environment. Aquatic Biology 1, 277–289.
| Movement and distribution of young bull sharks Carcharhinus leucas in a variable estuarine environment.Crossref | GoogleScholarGoogle Scholar |
Heupel, M. R., Carlson, J. K., and Simpfendorfer, C. A. (2007). Shark nursery areas: concepts, definition, characterization and assumptions. Marine Ecology Progress Series 337, 287–297.
| Shark nursery areas: concepts, definition, characterization and assumptions.Crossref | GoogleScholarGoogle Scholar |
Last, P. R., and Stevens, J. D. (2009). ‘Sharks and Rays of Australia.’ 2nd edn. (CSIRO Publishing: Melbourne.)
Latrubesse, E. M., Stevaux, J. C., and Sinha, R. (2005). Tropical rivers. Geomorphology 70, 187–206.
| Tropical rivers.Crossref | GoogleScholarGoogle Scholar |
Madsen, T., and Shine, R. (1996). Seasonal migration of predators and prey – a study of pythons and rats in tropical Australia. Ecology 77, 149–156.
| Seasonal migration of predators and prey – a study of pythons and rats in tropical Australia.Crossref | GoogleScholarGoogle Scholar |
Medved, R. J., and Marshall, J. A. (1983). Short term movements of young sandbar sharks, Carcharhinus plumbeus (Pisces, Carcharhinidae). Bulletin of Marine Science 33, 87–93.
National Oceanic and Atmospheric Administration (NOAA) (1998). Oxygen depletion in coastal waters. In ‘NOAA’s State of the Coast Report’. (Ed. N. N. Rabalais.) pp. 1–16. (NOAA: Silver Spring, MD.) Available at http://oceanservice.noaa.gov/websites/retiredsites/sotc_pdf/HYP.PDF[Verified 4 November 2010].
Perna, C., and Burrows, D. (2005). Improved dissolved oxygen status following removal of exotic weed mats in important fish habitat lagoons of the tropical Burdekin River floodplain, Australia. Marine Pollution Bulletin 51, 138–148.
| Improved dissolved oxygen status following removal of exotic weed mats in important fish habitat lagoons of the tropical Burdekin River floodplain, Australia.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2MXitF2gtLw%3D&md5=37a12d545d686b17707d58c1007e2cf6CAS | 15757716PubMed |
Pihl, L., Baden, S. P., and Diaz, R. J. (1991). Effects of periodic hypoxia on distribution of demersal fish and crustaceans. Marine Biology 108, 349–360.
| Effects of periodic hypoxia on distribution of demersal fish and crustaceans.Crossref | GoogleScholarGoogle Scholar |
R Development Core Team (2009). ‘R: A Language and Environment for Statistical Computing.’ (R Foundation for Statistical Computing: Vienna.)
Rayner, T. S., Pusey, B. J., and Pearson, R. G. (2008). Seasonal flooding, instream habitat structure and fish assemblages in the Mulgrave River, northeast Queensland: towards a new conceptual framework for understanding fish–habitat dynamics in small tropical rivers. Marine and Freshwater Research 59, 97–116.
| Seasonal flooding, instream habitat structure and fish assemblages in the Mulgrave River, northeast Queensland: towards a new conceptual framework for understanding fish–habitat dynamics in small tropical rivers.Crossref | GoogleScholarGoogle Scholar |
Scruton, D. A., Pennell, C. J., Robertson, M. J., Ollerhead, L. M. N., and Clarke, K. D. (2005). Seasonal response of juvenile Atlantic salmon to experimental hydropeaking power generation in Newfoundland, Canada. North American Journal of Fisheries Management 25, 964–974.
| Seasonal response of juvenile Atlantic salmon to experimental hydropeaking power generation in Newfoundland, Canada.Crossref | GoogleScholarGoogle Scholar |
Simpfendorfer, C. A., Heupel, M. R., and Hueter, R. E. (2002). Estimation of short-term centers of activity from an array of omnidirectional hydrophones and its use in studying animal movements. Canadian Journal of Fisheries and Aquatic Sciences 59, 23–32.
| Estimation of short-term centers of activity from an array of omnidirectional hydrophones and its use in studying animal movements.Crossref | GoogleScholarGoogle Scholar |
Simpfendorfer, C. A., Freitas, G. F., Wiley, T. R., and Heupel, M. R. (2005). Distribution and habitat partitioning of immature bull sharks (Carcharhinus leucas) in a Southwest Florida Estuary. Estuaries 28, 78–85.
| Distribution and habitat partitioning of immature bull sharks (Carcharhinus leucas) in a Southwest Florida Estuary.Crossref | GoogleScholarGoogle Scholar |
Steiner, P. A., and Michel, M. (2007). Effects of tidal current on the movement patterns of juvenile bull sharks and blacktip sharks. In ‘Shark Nursery Grounds of the Gulf of Mexico and the East Coast Waters of the United States’. (Eds C. T. McCandless, N. E. Kohler and H. L. Pratt Jr.) pp. 251–264. (American Fisheries Society: Bethesda, MD.)
Stevens, J. D., West, G. J., and McLoughlin, K. J. (2000). Movements, recapture patterns, and factors affecting the return rate of Carcharhinid and other sharks tagged off Northern Australia. Marine and Freshwater Research 51, 127–141.
| Movements, recapture patterns, and factors affecting the return rate of Carcharhinid and other sharks tagged off Northern Australia.Crossref | GoogleScholarGoogle Scholar |
Ter Morshuizen, L. D., Whitfield, A. K., and Paterson, W. (1996). Influence of freshwater flow regime on fish assemblages in the Great Fish River and estuary. Southern African Journal of Aquatic Sciences 22, 52–61.
Ubeda, A. J., Simpfendorfer, C. A., and Heupel, M. R. (2009). Movements of bonnetheads, Sphyrna tiburo, as a response to salinity change in a Florida estuary. Environmental Biology of Fishes 84, 293–303.
| Movements of bonnetheads, Sphyrna tiburo, as a response to salinity change in a Florida estuary.Crossref | GoogleScholarGoogle Scholar |
Walker, T. (1981). Seasonal salinity variations in Cleveland Bay, northern Queensland. Australian Journal of Marine and Freshwater Research 32, 143–149.
| Seasonal salinity variations in Cleveland Bay, northern Queensland.Crossref | GoogleScholarGoogle Scholar |
Wetherbee, B. M., and Rechisky, E. L. (1999). Movement patterns of juvenile sandbar sharks on their nursery grounds in Delaware Bay. In ‘Biotelemetry 15: Proceedings of the 15th International Symposium on Biotelemetry’. (Eds J. H. Eiler, D. J. Alcorn and M. R. Neuman.) pp. 91–98. (International Society on Biotelemetry: Wageningen, The Netherlands.)
Whitfield, A. K., and Paterson, A. W. (1995). Flood-associated mass mortality of fishes in the Sundays Estuary. Water S.A. 21, 385–389.
Winemiller, K. O., and Jepsen, D. B. (1998). Effects of seasonality and fish movement on tropical river food webs. Journal of Fish Biology 53, 267–296.
| Effects of seasonality and fish movement on tropical river food webs.Crossref | GoogleScholarGoogle Scholar |