

Effects of climate change on fish reproduction and early life history stages
Ned W. Pankhurst A C and Philip L. Munday BA Australian Rivers Institute, Griffith University, Gold Coast, Qld 4222, Australia.
B ARC Centre of Excellence for Coral Reef Studies, and School of Marine and Tropical Biology, James Cook University, Townsville, Qld 4811, Australia.
C Corresponding author. Email: n.pankhurst@griffith.edu.au
Marine and Freshwater Research 62(9) 1015-1026 https://doi.org/10.1071/MF10269
Submitted: 29 October 2010 Accepted: 1 March 2011 Published: 21 September 2011
Journal Compilation © CSIRO Publishing 2011 Open Access CC BY-NC-ND
Abstract
Seasonal change in temperature has a profound effect on reproduction in fish. Increasing temperatures cue reproductive development in spring-spawning species, and falling temperatures stimulate reproduction in autumn-spawners. Elevated temperatures truncate spring spawning, and delay autumn spawning. Temperature increases will affect reproduction, but the nature of these effects will depend on the period and amplitude of the increase and range from phase-shifting of spawning to complete inhibition of reproduction. This latter effect will be most marked in species that are constrained in their capacity to shift geographic range. Studies from a range of taxa, habitats and temperature ranges all show inhibitory effects of elevated temperature albeit about different environmental set points. The effects are generated through the endocrine system, particularly through the inhibition of ovarian oestrogen production. Larval fishes are usually more sensitive than adults to environmental fluctuations, and might be especially vulnerable to climate change. In addition to direct effects on embryonic duration and egg survival, temperature also influences size at hatching, developmental rate, pelagic larval duration and survival. A companion effect of marine climate change is ocean acidification, which may pose a significant threat through its capacity to alter larval behaviour and impair sensory capabilities. This in turn impacts on population replenishment and connectivity patterns of marine fishes.
Additional keywords: acidification, estrogens, larval behaviour, spawning, temperature.
Introduction
Temperature is a fundamental physical regulatory factor in the lives of fishes and this effect is expressed particularly strongly in the control of all reproductive processes from gamete development and maturation, ovulation and spermiation, spawning, embryogenesis and hatching, to larval and juvenile development and survival. In reproductively mature adults, temperature is generally considered to be a secondary cue to photoperiod in phasing reproductive seasonality but it has a major role in synchronising the final stages of reproductive maturity, and also in truncating reproductive episodes (reviewed in Pankhurst and Porter 2003). The effects of temperature can be differentially expressed depending on when in the annual thermal cycle spawning normally occurs, with increasing spring temperatures being required to cue maturation in spring and early summer spawners (e.g. Stacey 1984; Scott and Pankhurst 1992; Shimizu 2003), but elevated temperatures delaying the onset of maturation and ovulation in autumn-spawning species (reviewed in Pankhurst and King 2010). Temperature has a similarly important role in the modulation of post-fertilisation processes both through its rate-determining effects on embryogenesis and hatching (Pauly and Pullin 1988) and subsequent larval development (Howell et al. 1998), growth (Jobling 1997) and survival (Sponaugle and Cowen 1996).
Complications in the assessment of the effects of temperature in the natural environment arise from the fact that much of our current understanding is generated from controlled laboratory experiments that typically test thermal tolerances rather than behavioural preferences. For example, studies on salmonids show that there is often a wide gap between tolerance and preference profiles. Atlantic salmon Salmo salar have upper thermal tolerances in the 22–24°C range (Barton 1996) but ocean-range preferences for temperatures at 4–10°C (Reddin et al. 2000) and similar effects are reported for other salmonids (reviewed in Pankhurst and King 2010). This means that temperatures that might not be high enough to elicit tolerance-related responses in captivity can induce significant preference-related effects in the natural environment. Single-domain temperature experiments also generally ignore the synergistic interaction of temperature with other physical and biotic variables. Temperature effects on reproduction can be differentially expressed under different photoperiod regimes (Pankhurst and Porter 2003) and recent experiments on tropical damselfish show that temperature effects are modulated by nutritional status (Donelson et al. 2010). Finally, some species exhibit variation in thermal reaction norms across their geographic range that indicates some capacity for acclimation and adaptation to temperature gradients (Angilletta 2009; Gardiner et al. 2010), and potentially to a changing climate.
The effects of climate change on aquatic species will vary with latitude, habitat, water column characteristics, and in riverine systems, flow regimes. However, some general expectations of climatic change are for possible phase-shifting of seasonal temperature profiles, elevated seasonal maxima and minima, and as a consequence of increases in atmospheric CO2, increasing ocean acidification. Riverine habitats are likely to experience elevated temperatures in association with decreased flow rates and increasing incidence of hypoxic conditions, but with quite marked regional differences in effect. For example, parts of tropical Australia are predicted to experience increased rainfall and flow regimes whereas both are likely to be markedly reduced in south-western Australia (Lough et al. 2011). A reasonable prediction is that the effects of temperature change on reproduction will be most widespread in marine systems, but probably most extreme in riverine systems. Lacustrine habitats will experience a range of effects depending on lake inflows, water column structure and lake basin topography (Wahl and Löffler 2009). In addition to increases in temperature, elevations in atmospheric CO2 mean that marine systems will also be affected by ocean acidification, which has the potential to directly influence reproduction and early life stages of marine species (Ishimatsu et al. 2008), and to interact with the effects of temperature (Pörtner and Farrell 2008).
With the caveats noted above, the implication is that any changes in thermal regime occasioned by climate change will have major consequences for fish reproduction, and that these effects will be exercised across all stages of the reproductive process (Graham and Harrod 2009; Jonsson and Jonsson 2009; Pankhurst and King 2010). The extent of these effects will be determined by a range of factors including specific physiological tolerances, capacity for acclimation and adaptation, scope for behavioural avoidance, capacity to extend or shift ranges, and the timing of thermal challenges with respect to the reproductive cycle. The possible outcomes range from extremes of complete reproductive and recruitment failure to changes in seasonal phasing of reproduction and possible increases in species range (Munday et al. 2008a). This review examines the basis for some of these effects at the level of the individual and the possible consequences for population-level events in terms of adult reproduction, and subsequent egg, larval and juvenile development and survival.
Reproduction
Endocrine control of reproduction
Changes in environmental variables are transduced into effects on reproductive processes through the hypothalamo–pituitary–gonadal (HPG) axis. This begins with hypothalamic synthesis and synaptic release of peptide gonadotropin-releasing hormones (GnRH) onto the gonadotropic cells of the pituitary gland, where they stimulate the synthesis and release of the protein hormones, follicle-stimulating hormone (FSH) and luteinising hormone (LH) (reviewed in Planas and Swanson 2008; Levavi-Sivan et al. 2010; Zohar et al. 2010). In many species, there is also inhibitory tone imposed by the action of dopamine (DA)-secreting neurones, with the release of FSH and LH being dependent on the balance between GnRH stimulation and DA inhibition (reviewed in Dufour et al. 2010). The GnRH–pituitary interaction is further modulated by melatonin produced by the light-sensitive pineal gland by as-yet poorly understood mechanisms (reviewed in Migaud et al. 2010), by the kisspeptin system through direct effects on the activity of GnRH-producing neurones (reviewed in Akazome et al. 2010) and by the feedback effects of gonadal steroids (reviewed in Pankhurst 2008).
FSH is involved in stimulating the early stages of gamete development and LH in the control of maturational events, with both exercising their effects at the gonadal level through binding with G protein-coupled membrane-bound receptors in the ovary and the testis, giving rise to an increase in intracellular cAMP levels (Planas and Swanson 2008). This in turn results in the activation of protein kinases and the susbesequent de novo synthesis and releases of gonadal steroids through the sequential cleavage of the base molecule cholesterol. In males, the predominant steroids produced are the androgens, testosterone (T) and its more biologically active metabolite 11-ketotestosterone (11KT); females also produce T but this is further metabolised to the oestrogen 17β-oestradiol (E2). Both sexes also produce a progesterone-like maturational steroid (typically 17,20β-dihydroxy-4-pregnen-3-one, 17,20βP or 17,20β,21-trihydroxy-4-pregnen-3-one, 20β-S) in response to gonadal stimulation by LH (Pankhurst 2008). The actions of gonadal steroids include the stimulation of gametogenesis in both sexes, spermiation in males and vitellogenesis (yolk accumulation) and oocyte maturation in females, as well as regulating secondary sexual characters and a suite of sexual behaviours (Pankhurst 1998, 2008). A key step in the reproductive development of females is the stimulation by E2 of hepatic synthesis of the egg yolk precursor vitellogenin (Vtg) which is released into the plasma from where it is taken up by the developing oocytes. E2 also stimulates the ovarian and hepatic synthesis of zona pellucida proteins (ZP) that will ultimately form the chorion (egg shell) of the ovulated ovum (Tyler et al. 2000; Babin et al. 2007; Modig et al. 2007).
Temperature and the HPG axis
Temperature change has the capacity to affect the HPG axis at multiple sites through its reaction-rate-determining effects on hormone synthesis and action, and its effects on hormone structure. This is reflected in a minimum temperature threshold for most endocrine events, increasing hormone synthesis, activity and metabolism across the physiological tolerance range and decreasing activity at the top end of that range (Pankhurst and King 2010). Inhibitory effects at higher temperature may arise from conformational changes in proteins (e.g. FSH, LH and their receptors, steroid-synthesising enzymes), and also the increasing tendency for steroid hormones to form water-soluble conjugates at high temperatures (reviewed in Van Der Kraak and Pankhurst 1997). Steroid conjugates (usually sulfates or glucuronides) suffer the dual fate of no longer being soluble in (and able to pass through) cell membranes to gain access to their intracellular receptors, and of being more available for kidney filtration and excretion in the urine, significantly reducing their plasma residence times.
Irrespective of the mechanisms involved, it is clear that thermal inhibition of reproduction is present across a wide spread of taxa, habitats and temperature ranges with the main difference between species being the absolute temperature at which the suppressive effects occur (Table 1). In cold temperate and sub-Arctic species, the inhibitory effects typically appear at temperatures of 11–12°C, among cold-temperate species at around 18°C, temperate species at ~24°C and tropical species at 30°C and above (Table 1). This supports the view that all species are likely to show similar responses to rising temperatures, but that the thresholds for these effects will vary in relation to specific thermal tolerance ranges. There is also some evidence that the ranges over which normal function can be maintained may be broader in cool-water and temperate species than among tropical species (Nilsson et al. 2009; Donelson et al. 2010).

|
More detailed understanding of the mechanisms involved in the thermal inhibition of reproductive processes is derived mainly from studies on temperate species, with most evidence from studies on salmonids. Arctic charr, Salvelinus alpinus, held at 10°C (a temperature that suppresses normal ovulation) show greater responsiveness (LH secretion and subsequent ovulation) to synthetic analogues of GnRH (GnRHa) combined with the dopamine antagonist pimozide than GnRHa alone, suggesting that part of the inhibitory mechanism involves increases in dopamine inhibition at the level of the pituitary (Gillet and Breton 2009). In Atlantic salmon, Salmo salar, elevated summer and autumn temperatures inhibit the activity of the steroid-converting enzyme P450 aromatase (arom) and the subsequent ovarian synthesis of E2 from androgen precursors. This in turn results in reductions in the hepatic synthesis of Vtg and subsequent reductions in final egg size, fertility and survival. Exposure to elevated temperature during vitellogenesis also results in reduced affinity of hepatic E2-receptors. Exposure to elevated temperatures at the completion of vitellogenesis (late autumn) inhibits the synthesis of the maturational steroid 17,20βP and subsequent progression of oocytes through final oocyte maturation (the resumption of meiosis) and ovulation (reviewed in Pankhurst and King 2010).
Similar effects are seen in non-salmonids, with wolffish, Anarhichas lupus, showing reduced plasma levels of T and E2, retarded ovulation, and reduced fertility and survival (Tveiten and Johnsen 2001; Tveiten et al. 2001), red seabream, Pagrus major, showing inhibition of expression of genes coding for arom and 11β-hydroxylase (which mediates the conversion of T to 11KT) (Lim et al. 2003), and freshwater pejerrey, Odontesthes bonariensis, showing reduced plasma levels of E2 in females and T in males (Soria et al. 2008). At higher levels in the HPG axis, red seabream exposed to typical summer temperatures (that truncate normal spawning) showed reduced central expression of genes for GnRH and its pituitary receptor, and reduced pituitary expression of the β-subunit gene for LH (LHβ) (Okuzawa et al. 2003). Similarly, pejerrey displayed reduced expression of LHβ and the receptor for FSH in females exposed to moderate elevations in temperature (Soria et al. 2008). There is also some evidence that high temperature may interfere with the transduction of photoperiod signals to the reproductive axis, although as noted earlier, the mechanism in fish is still not well understood. The amplitude of the nocturnal surge in melatonin in fish is greater at higher temperature and if, as in some birds, melatonin exerts its role through stimulation of factors that inhibit GnRH synthesis (reviewed in Migaud et al. 2010), then temperature-regulated increases in melatonin secretion could also result in an inhibitory effect at the pituitary level. With the caveat that the understanding of all of the possible effects of temperature on reproductive physiology is not complete, it is clear that temperature regulates reproduction at multiple sites in the reproductive pathway (Fig. 1).

|
Stress and reproduction
An alternative, non-exclusive explanation for the inhibitory effects of elevated temperature is the possibility that thermal inhibition occurs through activation of the hormonally-mediated stress response. In turn, stress is known to have marked inhibitory effects on reproduction in fish (Pankhurst and Van Der Kraak 1997; Leatherland et al. 2010; Schreck 2010). Stress stimulates activation of an acute catecholamine-mediated response that has the primary effect of rapidly increasing energy availability and the delivery of O2 to the tissues, followed by a longer and more sustained activation of the hypothalamic–pituitary–interrenal (HPI) axis, resulting in plasma elevations of the steroid cortisol in teleosts and chondrosteans, and 1α-hydroxycorticosterone in elasmobranchs (reviewed in Pankhurst 2011). Short-term increases in corticosteroids increase the availability of a variety of energy substrates, but longer-term exposure to elevated cortisol results in suppressive effects on a range of functions including reproduction, growth and immune function (Pankhurst 2011). Some of the longer-term effects can be explained by the largely catabolic effects of cortisol, but stress is also capable of suppressing plasma levels of T and E2 in as little as 15–30 min after the imposition of stress (Haddy and Pankhurst 1999), and there is equivocal evidence that this rapid effect is the direct result of cortisol action (Pankhurst et al. 1995).
A consideration for the role of stress in temperature-mediated suppression of reproduction is whether environmental temperature change actually stimulates a stress response. Increases in temperature can be an effective stressor in the laboratory context (Pankhurst 2011) but elevations in temperature that inhibit reproduction in broodstock Atlantic salmon do not result in significant elevations of plasma cortisol (Pankhurst et al. 2011), strongly suggesting that the marked inhibitory effects observed in salmonids are not being exercised through the HPI axis. There is also the consideration as to whether environmental conditions are routinely stressful for fish in the natural environment, and the conclusion is that there is not strong evidence for initiation of stress responses under quite wide ranges of environmental conditions (Schreck 2010; Pankhurst 2011). The caveat here is that events that do stimulate stress responses in free-ranging vertebrates from other classes are typically associated with extreme weather events. For fish populations, this is likely to coincide with storms and floods, when sampling from wild populations is generally precluded (Pankhurst 2011). However, environmental changes predicted for riverine environments such as increasing temperatures, decreased flow rates and O2 saturation (Morrongiello et al. in press) may well generate conditions that do stimulate activation of the HPI axis. A reasonable prediction is that this will contribute to reproductive suppression in these environments, separately or additively to direct thermal effects on reproductive endocrine processes.
Additional considerations
Despite the general consistency of the effects of elevated temperature on reproduction described above, data from an increasing number of studies suggest that there will be subtleties in the way that these effects are likely to be expressed. This in turn is going to make the precise nature of effects arising from climate change harder to predict. A first consideration will be when in the reproductive cycle thermal challenge occurs. Studies on austral stocks of Atlantic salmon have shown that females are reasonably robust in terms of their response to elevated temperature for much of the summer and autumn, but highly sensitive to thermal disruption in February during a period of substantial oocyte growth and gonadal reorganisation (King et al. 2007). The implication is that elevated temperature may have more, or less, benign effects depending on whether or not it consistently occurs across this period of heightened sensitivity.
Temperature change may also have differential impacts on sympatric species with different seasonal spawning patterns. For example, spiny damselfish, Acanthochromis polyacanthus, spawn in spring and early summer on Australia’s Great Barrier Reef, and show temperature inhibition of ovarian steroidogenesis above 30°C (Donelson et al. 2010; N. W. Pankhurst, P. M. Pankhurst and L. Gonzalez-Reynoso, unpubl. data). In the same regions, humbug dascyllus, Dascyllus aruanus, spawn later in the summer when temperatures are typically higher, and do not show evidence of inhibition of steroidogenesis until temperatures reach 33°C (N. W. Pankhurst, P. M. Pankhurst and L. Gonzalez-Reynoso, unpubl. data). This suggests that as in temperate species (Pankhurst and Porter 2003), increasing temperature plays a normal role in truncating spawning episodes in spring spawners, and cueing the onset of spawning in summer spawners. Under this scenario, the effects of any increase in temperature will depend on the timing of this increase with respect to the reproductive cycle and the natural seasonal timing of spawning. For example, a higher summer maximum may have negligible effects on spiny damselfish reproduction if it occurs during a period when spawning is normally suppressed, but might reduce a spring-spawning window if the rate of the spring increase in temperature became steeper, (i.e. the effects will be strongly dependent not only on the absolute temperature but also the annual pattern of thermal change).
The response to thermal stress may also be variable within the same population. Recent work with Atlantic salmon shows that the effect of summer and autumn exposure to high temperature (22°C) is partially determined by the age class of broodstock. Both first-spawning-season (maiden) and second-spawning-season (repeat) females show depression of plasma E2 and Vtg levels, reduced expression of Vtg and ZP genes, delay in maturation and ovulation and reduced fertility and egg survival (Pankhurst et al. 2011). However, repeat females show higher fertility and survival than maidens for reasons that are not clear but may relate to overall larger egg size and maternal endowment with vitellogenin. A similar effect operating in natural populations would generate differential impacts depending on the age structure of the population, and one prediction may be that heavily fished populations dominated by younger fish might have heightened susceptibility to thermal disruption of reproduction.
Within-population variability is also likely to occur as a function of individual nutritional status. It is well known that there is a tightly coupled relationship between nutritional status and reproductive output (Lambert et al. 2000); however, experiments with spiny damselfish have shown that high nutritional status confers some protection from the inhibitory effects of exposure to high temperature (Donelson et al. 2010). The basis for the effect is not known but may be related to the permissive effects that several nutritionally regulated endocrine factors (e.g. thyroid hormones, insulin-like growth factor I) exert on the HPG axis (reviewed in Pankhurst et al. 2008). On this basis, another prediction may be that temperature effects on reproduction of natural populations will be exacerbated if there is a concurrent negative impact on food availability.
An observation based on the consistent effect of increasing temperature in inhibiting gonadal arom activity in fishes is that arom suppression will have at least two additional potential effects on reproduction based on the roles of arom in sex determination, and sex inversion, respectively. Phenotypic sex in fish can be quite labile during the period of primary sex determination, such that female phenotypes emerge in the presence of brain arom-dependent oestrogen synthesis, and males in the absence of arom activity (reviewed in Devlin and Nagahama 2002). Higher temperatures inhibit arom activity and drive sex determination towards the male phenotype (reviewed by Guiguen et al. 2010). A prediction here is that increasing sea temperatures will increase the proportion of fish developing as male, even within the thermal range over which reproductive performance can be maintained. The possible impacts of this on population resilience are not known. Similarly, many teleost species undergo sex inversion (Frisch 2004), with the presence of gonadal arom activity maintaining the ovarian condition in protogynous species, and being requisite for the transition from male to female in protandrous species. The primary control mechanism appears to be mainly exercised by social context (e.g. Munday et al. 2006) but the underlying physiological mechanism is strongly dependent on the subsequent activation or inhibition of arom activity. Thermal inhibition of arom activity may generate premature transition from female to male in protogynous species and inhibition of transition to female in protandrous species, even in the presence of the appropriate social context. Here also, the possible impacts at population level are not known.
Ocean acidification and reproduction
Uptake of additional CO2 at the ocean surface, owing to increasing concentrations of CO2 in the atmosphere, is causing ocean pH to decline and reducing the carbonate ion concentration of the shallow ocean. This process, known as ocean acidification, is considered to be a serious threat to marine species, especially for calcifying species that require carbonate ions to form their shells and skeletons (Hoegh-Guldberg et al. 2007; Fabry et al. 2008; Smith 2009). Elevated pCO2 can also have a direct physiological effect on aquatic species through disruption of acid–base balance and limiting oxygen supply (Pörtner et al. 2004; Pörtner and Farrell 2008). The effects of increasing pCO2 in water is probably of greater concern than reducing pH per se, because of the high permeability to biological tissue of gaseous CO2 relative to hydrogen ions (Brauner 2009). Experiments with red seabream (Pagrus major) demonstrate that larval fish are more sensitive to the effects of acidification with CO2 than to the same pH achieved with mineral acids (Kikkawa et al. 2004). Increased pCO2 in tissue causes acidosis (lowering of pH and accumulation of bicarbonate), which can be detrimental to many cellular processes, including protein synthesis, enzymatic function and oxygen transport (Pörtner et al. 2004). Fish compensate for acidosis by acid–base equivalent ion transport from the body to the environment, mostly across the branchial epithelium, and to a lesser extent, via the kidneys and intestine (Claiborne et al. 2002).
In general, fishes appear to be more tolerant to increases in ambient CO2 than many invertebrates (Ishimatsu et al. 2008; Widdicombe and Spicer 2008), possibly because of their well-developed mechanisms for acid–base regulation (Pörtner et al. 2004; Melzner et al. 2009). However, very little is known about the effects that chronic exposure to levels of pCO2 predicted to occur over the next 50–100 years (up to ~1000 ppm CO2) might have on fish reproduction. The few preliminary studies available suggest the impacts might not be substantial. Sperm motility of the flounder, Limanda yokohamae, is arrested by mild increases in pCO2 (Inaba et al. 2003), but similar effects were not observed in 10 other species from a range of families (Inaba et al. 2003) or in the Baltic cod, Gadus morhua (Frommel et al. 2010). Sensitivity of fish eggs to elevated CO2 varies markedly between species, but species tested to date typically have 24-h LC50 (lethal concentration resulting in 50% mortality) values well above 10 000 ppm CO2 (Ishimatsu et al. 2008). Furthermore, Munday et al. (2009a) did not detect any effect of exposure to 1000 ppm CO2 on the embryonic duration or survival of clownfish (Amphiprion percula) eggs. Eggs of pelagic spawners might be more sensitive to CO2 stress than the eggs of benthic spawners such as clownfishes, because pelagic eggs probably experience less fluctuation in environmental pCO2 than benthic eggs, but this hypothesis has not been adequately tested.
One potential concern is that higher pCO2 may limit the scope for aerobic performance in adults (Pörtner and Farrell 2008), which could affect reproductive output. Aerobic scope of two tropical cardinalfishes, Ostorhinchus doederleini and O. cyanosoma, declined by 33% and 47%, respectively, when they were exposed to ~1000 ppm CO2 at the average summer temperature (29°C) for the study population and at temperatures up to 3°C above average (Munday et al. 2009b). Whether such a loss in aerobic capacity has an effect on reproduction is unknown, but it is reasonable to suspect that it will. For example, collapse of aerobic scope in association with anomalously high water temperature has been linked to failed migration (and thus spawning) in sockeye salmon, Oncorhynchus nerka (Farrell et al. 2008). The possible effects of elevated CO2 on endocrine pathways that mediate reproduction in fishes are currently unknown.
Early life history stages
Effects on egg incubation
Eggs are one of the most thermally sensitive life stages in fishes and tolerance limits appear to be within ± 6°C of the spawning temperature for many species (Rombough 1997). Small increases in temperature can dramatically increase egg mortality, especially in tropical species (Gagliano et al. 2007). Consequently, survivorship to hatching could decline as oceans and rivers warm, unless species adjust the timing of spawning to suit the optimal temperature for embryo development. Such shifts appear likely because gametogenesis is highly temperature-sensitive in many fish species (discussed above) and breeding may cease before critical thermal limits for egg survival are reached. For example, the critical temperature for gametogenesis of brook trout, Salvelinus fontinalis, is ~2°C lower that the thermal limit for normal development of fertilised eggs (Rombough 1997). Nevertheless, some species spawn at suboptimal temperatures and may suffer reduced embryonic survival as a result, both because increased temperature during ovulation can reduce gamete viability (Van Der Kraak and Pankhurst 1997) and because increased temperature during embryogenesis increases mortality (Gillet et al. 1996; Pankhurst and Thomas 1998; Janhunen et al. 2010).
Temperature also has a highly significant effect on the rate of embryonic development. For many species, the rate of embryonic development more than triples for each 10°C increase in temperature (i.e. Q10 > 3) (Rombough 1997). An increased developmental rate means that the incubation period declines as average water temperature increases. Incubation period is also dependent on egg size, with larger eggs taking longer to develop than small eggs (Pauly and Pullin 1988). Consequently, increased temperature may advance hatching by minutes to hours in small eggs, and by hours to days in large eggs, with the effects being most marked in cold-water species with long incubation periods (Rombough 1997). Whether shorter incubation periods affect individual fitness may depend on the potential for a mismatch between the timing of hatching and favourable conditions for larval survival. For example, hatching of benthic eggs often occurs at night when larvae are less susceptible to visual predators (Robertson 1991; Michael 2008). At least in some species, hatching can be cued by environmental factors that are not temperature-dependent, such a diurnal light cycles, which may help ensure larvae hatch at the appropriate time even if they are competent to hatch earlier.
Effects on larvae
Temperature affects metabolism, growth, developmental rate and stage duration of larval fishes (reviewed by Houde 1989; Blaxter 1991; Benoît et al. 2000). Metabolic rate, measured as mass-specific oxygen consumption, increases sharply with increasing temperature in larval fishes, although responses vary considerably among species (Houde 1989; Rombough 1997). Higher metabolic rates mean that fish have higher basal energy demands at higher temperatures. Larval growth rates also increase with temperature for both temperate (Blaxter 1991; Benoît et al. 2000) and tropical species (McCormick and Molony 1995; Meekan et al. 2003; Green and Fisher 2004), with temperature explaining up to 89% of variation in growth rates among cohorts of some species (Sponaugle and Cowen 1996). Thermal reaction norms of growth in larval fish tend to be approximately linear until the lethal upper thermal limit is reached, at least in the majority of species studied to date (Sponaugle and Cowen 1996; Rombough 1997). This contrasts with thermal reaction norms of growth in juveniles and adults, which usually decline well before the lethal thermal limit is reached (e.g. Munday et al. 2008b). As a result, we might expect that larval growth rates will tend to be maintained as water temperatures increase as a result of global warming, even if temperatures exceed optimum conditions for some other life processes.
In warmer water, developmental rates increase and therefore stage durations are shorter. The time until yolk absorption, metamorphosis and pelagic larval duration (PLD) are all negatively correlated with temperature, both within species (Rombough 1997; Fuiman et al. 1998; Green and Fisher 2004) and among species (Houde 1989; Benoît et al. 2000). In one of the clearest examples of this effect, PLD of reef fishes is closely correlated with temperature in a range of species (McCormick and Molony 1995; Sponaugle and Cowen 1996; Green and Fisher 2004). There are also strong correlations between PLD and growth rates, with fast-growing larvae often exhibiting shorter larval durations (Houde 1989; McCormick and Molony 1995; Sponaugle and Cowen 1996; Benoît et al. 2000; Green and Fisher 2004). The relationship between growth rate and PLD will influence size at settlement, which is often variable between cohorts (Sponaugle and Cowen 1996; Fuiman et al. 1998) because these rates are not perfectly reciprocal and because growth also depends on a range of other factors such as food supply. Larger size at settlement may offer some survival advantages (Sogard 1997); however, small individuals settling in warmer conditions may grow faster after settlement and quickly reach equivalent sizes to fish that settle in cooler conditions (Sponaugle and Cowen 1996).
As mortality rates are usually very high during the larval phase, faster growth and reduced PLD at higher temperatures might increase larval survivorship (Houde 1989; Bergenius et al. 2002; O’Connor et al. 2007). Across a broad range of species, the slope of the relationship between PLD and temperature is steepest in cool-water species (O’Connor et al. 2007), and small increases in water temperature might be expected to have the greatest effect on high-latitude species. However, even among tropical fishes, PLD can decline by 4–8% per °C (Munday et al. 2009c), which could have a significant effect on survivorship. Within the temperature range currently experienced by reef fishes, warmer years generally appear to favour good recruitment events for a variety of species (Meekan et al. 2001; Wilson and Meekan 2002; Cheal et al. 2007), which is consistent with the hypothesis that reduced PLD tends to increase larval survivorship. Recruitment of the bluehead wrasse, Thalassoma bifasciatum, in the Florida Keys increases with temperature, but also becomes more variable at high temperatures (Sponaugle and Cowen 1996), possibly because of the increased risk of starvation in warm-water cohorts. Although the growth rate of marine fish larvae increases with temperature, growth efficiency does not (Houde 1989; Rombough 1997). Furthermore, larvae require more food at higher temperatures to sustain higher metabolic rates. As a result, larvae are more susceptible to starvation at higher temperatures and this may explain observations of more variable recruitment and episodes of recruitment failure during periods of anomalously high water temperature. For example, recruitment of most reef fish species failed in French Polynesia during a warm El Niño period (Lo-Yat et al. 2011), suggesting that either reproduction or larval survival of a broad range of species was dramatically reduced at high summer temperatures.
Together, these observations demonstrate that increased temperatures will affect the life history and demography of larval fish, possibly affecting recruitment success and population dynamics. Small increases in temperature might tend to favour recruitment in some species, especially at higher latitudes. Larger temperature increases could lead to recruitment failures, especially at low latitudes, and at times or places where food supply is limited (Munday et al. 2008a). Larval success and patterns of recruitment will also be strongly affected by the effects of temperature on reproduction, which is likely to be one of the first processes impacted by increasing water temperature. A summary of the demonstrated and possible effects of increased temperature and pCO2 on early life history traits is shown in Fig. 2.
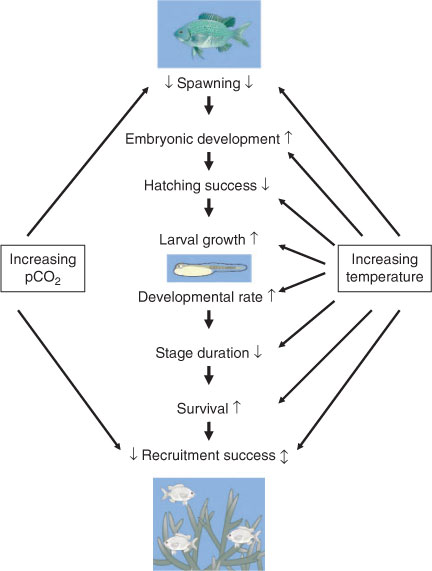
|
Ocean acidification and early life history stages
Larval stages are predicted to be more sensitive to elevated pCO2 than adults because they have a larger surface area-to-volume ratio, and thus are more susceptible to changes in ambient conditions, and because they might have less-developed mechanisms for acid–base balance compensation (Fabry et al. 2008; Ishimatsu et al. 2008; Melzner et al. 2009). Acid–base balance in adult fish is mostly maintained by ion transport across the gills (Claiborne et al. 2002). Although very little is known about the mechanisms and pathways of acid–base regulation in larval fishes, it is clear that they must be capable of acid–base homeostasis (Brauner 2009). Indeed, the early ontogenetic development of gills in larval fish may be more important in ionoregulation and maintaining acid–base balance than for oxygen delivery (Fu et al. 2010). Similar to the egg stage, the 24-h LC50 for larval fish is generally above 10 000 ppm CO2 for the few species tested to date (Kikkawa et al. 2003; Ishimatsu et al. 2008). Furthermore, two recent studies have not detected significant negative effects on life history traits of larval reef fishes reared in conditions simulating near-future CO2 in the ocean. Munday et al. (2009a) did not detect negative effects on the size at hatching, growth rate, size at settlement, or critical swimming speed of clownfish, Amphiprion percula, larvae reared from hatching in seawater aerated with up to 1000 ppm CO2. Similarly, juvenile spiny damselfish, Acanthochromis polyacanthus, reared for 3 weeks at up to 850 ppm CO2 were the same size as fish reared in current-day CO2 controls, did not suffer higher mortality, and exhibited no differences in skeletal or otolith development compared with controls (Munday et al. 2011). Although more studies are required, the limited evidence available to date suggests that pCO2 levels predicted to occur in the ocean during the next 50–100 years might not have a serious effect on the growth and development of larval fishes.
A potentially serious consequence of rising pCO2 is that it can affect the olfactory system of some marine fishes, rendering them unable to distinguish between ecologically important chemical cues (Munday et al. 2009d). Clownfish larvae reared at 1000 ppm CO2 exhibited a broad attraction to any chemical cue presented in binary-choice flume trials and became attracted to chemical cues that they avoided when reared in control seawater. At 1000 ppm CO2, they became attracted to the smell of inappropriate habitats (Munday et al. 2009d), could no longer discriminate the smell of kin versus non-kin (Munday et al. 2009d), and became attracted to the smell of predators (Dixson et al. 2010). At even higher pCO2 (~1700 ppm), they failed to respond to any chemical cues, indicating complete impairment of the olfactory system. In subsequent experiments, Munday et al. (2010) showed that: (1) olfactory impairment of larval clownfishes and damselfishes (Pomacentrus wardi) occurred at 700–850 ppm CO2, (2) behaviour of larval damselfish was altered in their natural coral reef habitat, and (3) that this caused a 5- to 9-fold increase in mortality from predation compared with controls. These results suggest that increased pCO2 in the ocean could have a significant effect on the successful replenishment of reef fish populations, although the effects on predators and the outcome of predator–prey interactions when both partners have been exposed to elevated CO2 have not yet been tested. Ecological connectivity between fish populations could also be significantly affected if exposure to elevated CO2 impairs navigation and homing abilities of larval fishes, as suggested by the original studies (Munday et al. 2009c).
The physiological mechanisms responsible for impairment of olfactory sensitivity and changes in behaviour of larval fish exposed to elevated CO2 are currently unknown, but do not appear to be related to changes in the chemical stimuli or abnormal developmental of the sensory system. Neither control nor CO2-treated fish altered their responses to chemical cues when presented in CO2-acidified versus control water (Munday et al. 2010; Dixson et al. 2010), indicating that elevated CO2 affects the fish, not the chemical cue itself. No modification of the sensory epithelium was apparent in CO2-treated fish (Munday et al. 2009d) and these fish regained their normal sensory ability after two days in control water (Munday et al. 2010). It is possibly that the effects on behaviour and olfactory ability be related to incomplete acid–base compensation or some other effect of elevated CO2 on the nervous system, but this has yet to be examined.
Acclimation and adaptation
Many fish species have geographic ranges spanning a considerable gradient in average, maximum and minimum temperature, suggesting some capacity for acclimation or adaptation to temperature change (Munday et al. 2008a). There is evidence that thermal exposure history can moderate subsequent responses to increasing temperature. Populations of bullhead, Cottus gobio, harvested from a stream system which had a less variable annual thermal range (4.5–11.5°C) than another (0.5–19.2°C) were found to be less robust in resisting the inhibitory effects of high temperature (Reyjol et al. 2009). Similarly, populations of four species of reef fish from two families (Apogonidae and Pomacentridae) on the northern Great Barrier Reef (GBR) exhibited poorer performance at high temperatures compared with fish from the southern GBR, which experience a more variable thermal range (Gardiner et al. 2010). This indicates that local populations of these species are either acclimated or adapted to the local thermal environment, and that populations from more variable environments might have enhanced capacity to cope with future thermal stress.
Although acute thermal stress often depresses reproduction, long-term exposure to higher temperatures can confer some improvement through acclimatory processes. For example, laboratory experiments on spiny damselfish show that fish reared at higher temperature since birth have a greater capacity to maintain reproduction at temperatures over 30°C compared with individuals that have only a few weeks thermal acclimation (J. Donelson et al. 2011). Furthermore, the thermal history of the parents is important, with embryos and larvae from parents reared at high temperatures exhibiting greater thermal tolerance or improved performance at high temperatures in several species (Rombough 1997).
In addition to acclimatory responses owing to thermal exposure history, individuals within a population can exhibit different thermal preferences (i.e. individual genetic variability), and different populations can be locally adapted to different thermal regimes (Crawshaw and O’Connor 1997; Farrell et al. 2008; Munday et al. 2008a). This variation provides the raw material for selection of genotypes that are more tolerant. The potential for genetic adaptation to rapid climate change depends on a range of factors, including the amount of adaptive genetic variation present, effective population sizes, generation time, and connectivity between populations that can aid in the spread of tolerant genotypes (Skelly et al. 2007; Munday et al. 2008a, 2009c). Unfortunately, much of this information is unknown for both marine and freshwater fishes and should be a priority area for future research.
Conclusion
Climate change will, or is already, affecting reproductive and early life history events of most fishes. This is occurring at a variety of levels and through a range of mechanisms which as our understanding develops are emerging as increasingly complex. These include the interplay of changes in physical variables with habitat, when in the reproductive cycle the thermal challenge occurs, the timing of spawning, whether events are extreme enough to initiate a physiological stress response, the energy status and reproductive age of the fish, and the thermal exposure history and adaptive capacity of the individual or the population. There is also the very strong suspicion that we are substantially under-informed to make useful predictions about likely effects beyond general assumptions, except for the relatively few species that have received the bulk of research attention. This in turn limits our capacity to develop specific options as management strategies. Temperature is one of the most fundamental variables affecting the lives of fishes but we still know discouragingly little about its effects.
Acknowledgements
We thank the anonymous referees and the Guest Editors of this volume for their helpful comments on an earlier version of the manuscript.
References
Akazome, Y., Kanda, S., Okubo, K., and Oka, Y. (2010). Functional and evolutionary insights into vertebrate kisspeptin systems from studies of fish brain. Journal of Fish Biology 76, 161–182.| Functional and evolutionary insights into vertebrate kisspeptin systems from studies of fish brain.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3cXkt1agsb8%3D&md5=ef98653f76647550df958a4f0993a2b7CAS |
Angilletta, M. J. (2009). ‘Thermal Adaptation: A Theoretical and Empirical Synthesis.’ (Oxford University Press: New York.)
Babin, P. J., Carnevali, O., Lubzens, E., and Schneider, W. J. (2007). Molecular aspects of oocyte vitellogenesis in fish. In ‘The Fish Oocyte: From Basic Studies to Biotechnical Applications.’ (Eds P. J. Babin, J. Cerdá and E. Lubzens.) pp. 39–76. (Springer: Dordrecht.)
Barton, B. A. (1996). General biology of salmonids. In ‘Principles of Salmonid Aquaculture.’ (Eds W. Pennell and B. A. Barton.) pp. 29–95. (Elsevier: Amsterdam.)
Benoît, H. P., Pepin, P., and Brown, J. A. (2000). Patterns of metamorphic age and length in marine fishes, from individuals to taxa. Canadian Journal of Fisheries and Aquatic Sciences 57, 856–869.
| Patterns of metamorphic age and length in marine fishes, from individuals to taxa.Crossref | GoogleScholarGoogle Scholar |
Bergenius, M. A. J., Meekan, M. G., Robertson, D. R., and McCormick, M. I. (2002). Larval growth predicts the recruitment success of a coral reef fish. Oecologia 131, 521–525.
| Larval growth predicts the recruitment success of a coral reef fish.Crossref | GoogleScholarGoogle Scholar |
Blaxter, J. H. S. (1991). The effects of temperature on larval fishes. Netherlands Journal of Zoology 42, 336–357.
| The effects of temperature on larval fishes.Crossref | GoogleScholarGoogle Scholar |
Brauner, C. J. (2009). Acid–base balance. In ‘Fish Larval Physiology’. (Eds R. N. Finn and B. G. Kapoor.) pp. 185–198. (Science Publishers: Enfield.)
Cheal, A. J., Delean, S., Sweatman, H., and Thompson, A. A. (2007). Spatial synchrony in coral reef fish populations and the influence of climate. Ecology 88, 158–169.
| Spatial synchrony in coral reef fish populations and the influence of climate.Crossref | GoogleScholarGoogle Scholar | 1:STN:280:DC%2BD2s3ot1Khug%3D%3D&md5=40a3de0c2cf25573c55ad52f8e779decCAS |
Claiborne, J. B., Edwards, S. L., and Morrison-Shetlar, A. I. (2002). Acid–base regulation in fishes: cellular and molecular mechanisms. The Journal of Experimental Zoology 293, 302–319.
| Acid–base regulation in fishes: cellular and molecular mechanisms.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD38XlvVKktr0%3D&md5=ad26d46df5d54a0862d7bdc3e5e341d7CAS |
Crawshaw, L. I., and O’Connor, C. S. (1997). Behavioural compensation for long-term thermal change. In ‘Global Warming: Implications for Freshwater and Marine Fish’. (Eds C. M. Wood and D. G. McDonald.) pp. 351–376. (Cambridge University Press: Cambridge.)
Devlin, R. H., and Nagahama, Y. (2002). Sex determination and sex differentiation in fish: an overview of genetic, physiological, and environmental influences. Aquaculture 208, 191–364.
| Sex determination and sex differentiation in fish: an overview of genetic, physiological, and environmental influences.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD38XktFKjsbY%3D&md5=01b7fecd33f7675f212072da2280172cCAS |
Dixson, D. L., Munday, P. L., and Jones, G. P. (2010). Ocean acidification disrupts the innate ability of fish to detect predator olfactory cues. Ecology Letters 13, 68–75.
| Ocean acidification disrupts the innate ability of fish to detect predator olfactory cues.Crossref | GoogleScholarGoogle Scholar |
Donelson, J. M., Munday, P. L., McCormick, M. I., Pankhurst, N. W., and Pankhurst, P. M. (2010). Effects of elevated water temperature and food availability on the reproductive performance of a coral reef fish. Marine Ecology Progress Series 401, 233–243.
| Effects of elevated water temperature and food availability on the reproductive performance of a coral reef fish.Crossref | GoogleScholarGoogle Scholar |
Donelson, J. M., Munday, P. L., McCormick, M. I., and Nilsson, G. E. (2011). Acclimation to predicted ocean warming through developmental plasticity in a tropical reef fish. Global Change Biology 17, 1712–1719.
| Acclimation to predicted ocean warming through developmental plasticity in a tropical reef fish.Crossref | GoogleScholarGoogle Scholar |
Dufour, S., Sebert, M.-E., Weltzein, F.-A., Rousseau, K., and Pasqualini, C. (2010). Neuroendocrine control by dopamine of teleost reproduction. Journal of Fish Biology 76, 129–160.
| Neuroendocrine control by dopamine of teleost reproduction.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3cXkt1agsb4%3D&md5=bea3c1be994c2a5e255e623e5bf98b7fCAS |
Fabry, V. J., Seibel, B. A., Feely, R. A., and Orr, J. C. (2008). Impacts of ocean acidification on marine fauna and ecosystem processes. ICES Journal of Marine Science 65, 414–432.
| Impacts of ocean acidification on marine fauna and ecosystem processes.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD1cXntFegtL4%3D&md5=25f0681d20c358a38b757d741fc1cf94CAS |
Farrell, A. P., Hinch, S. G., Cooke, S. J., Patterson, D. A., Crossin, G. T., et al. (2008). Pacific salmon in hot water: applying aerobic scope models and biotelemetry to predict the success of spawning migrations. Physiological and Biochemical Zoology 81, 697–709.
| Pacific salmon in hot water: applying aerobic scope models and biotelemetry to predict the success of spawning migrations.Crossref | GoogleScholarGoogle Scholar | 1:STN:280:DC%2BD1cjhtlWitQ%3D%3D&md5=4997e7a8bf95e78c6c51729853573e09CAS |
Frisch, A. (2004). Sex-change and gonadal steroids in sequentially-hermaphroditic teleost fish. Reviews in Fish Biology and Fisheries 14, 481–499.
| Sex-change and gonadal steroids in sequentially-hermaphroditic teleost fish.Crossref | GoogleScholarGoogle Scholar |
Frommel, A. Y., Stiebens, V., Clemmesen, C., and Havenhand, J. (2010). Effect of ocean acidification on marine fish sperm (Baltic cod: Gadus morhua). Biogeosciences Discussion 7, 5859–5872.
| Effect of ocean acidification on marine fish sperm (Baltic cod: Gadus morhua).Crossref | GoogleScholarGoogle Scholar |
Fu, C., Wilson, J. M., Rombough, P. J., and Brauner, C. J. (2010). Ions first: Na+ uptake shifts from the skin to the gills before O2 uptake in developing rainbow trout, Oncorhynchus mykiss. Proceedings of the Royal Society Series B, Biological Sciences 277, 1553–1560.
| Ions first: Na+ uptake shifts from the skin to the gills before O2 uptake in developing rainbow trout, Oncorhynchus mykiss.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3cXhtVehtbnK&md5=8d822fb4930205d8ace322639fab9e8aCAS |
Fuiman, L. A., Poling, K. R., and Higgs, D. M. (1998). Quantifying developmental progress from comparative studies of larval fishes. Copeia 1998, 602–611.
| Quantifying developmental progress from comparative studies of larval fishes.Crossref | GoogleScholarGoogle Scholar |
Gagliano, M., McCormick, M. I., and Meekan, M. G. (2007). Temperature-induced shifts in selective pressure at a critical developmental transition. Oecologia 152, 219–225.
| Temperature-induced shifts in selective pressure at a critical developmental transition.Crossref | GoogleScholarGoogle Scholar |
Gardiner, N. M., Munday, P. L., and Nilsson, G. E. (2010). Counter-gradient variation in respiratory performance of coral reef fishes at elevated temperatures. PLoS ONE 5, e13299.
| Counter-gradient variation in respiratory performance of coral reef fishes at elevated temperatures.Crossref | GoogleScholarGoogle Scholar |
Gillet, C. (1991). Egg production in an Arctic charr (Salvelinus alpinus L.) brood stock: effects of temperature on the timing of spawning and the quality of eggs. Aquatic Living Resources 4, 109–116.
| Egg production in an Arctic charr (Salvelinus alpinus L.) brood stock: effects of temperature on the timing of spawning and the quality of eggs.Crossref | GoogleScholarGoogle Scholar |
Gillet, C., and Breton, B. (2009). LH secretion and ovulation following exposure of Arctic charr to different temperature and photoperiod regimes: Responsiveness of females to a gonadotropin-releasing hormone analogue and a dopamine antagonist. General and Comparative Endocrinology 162, 210–218.
| LH secretion and ovulation following exposure of Arctic charr to different temperature and photoperiod regimes: Responsiveness of females to a gonadotropin-releasing hormone analogue and a dopamine antagonist.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD1MXlslWhu7w%3D&md5=759da0068c7233ef4855bdd99b99f542CAS |
Gillet, C., Breton, B., and Mikolajczyk, T. (1996). Effects of GnRHa and pimozide treatments on the timing of ovulation and egg quality in Arctic charr (Salvelinus alpinus) at 5 and 10°C. Aquatic Living Resources 9, 257–263.
| Effects of GnRHa and pimozide treatments on the timing of ovulation and egg quality in Arctic charr (Salvelinus alpinus) at 5 and 10°C.Crossref | GoogleScholarGoogle Scholar |
Graham, C. T., and Harrod, C. (2009). Implications of climate change for the fishes of the British Isles. Journal of Fish Biology 74, 1143–1205.
| Implications of climate change for the fishes of the British Isles.Crossref | GoogleScholarGoogle Scholar | 1:STN:280:DC%2BC3cjnt1ygug%3D%3D&md5=c45aed507470998e7e370afdb13d5565CAS |
Green, B. S., and Fisher, R. (2004). Temperature influences swimming speed, growth and larval duration in coral reef fish larvae. Journal of Experimental Marine Biology and Ecology 299, 115–132.
| Temperature influences swimming speed, growth and larval duration in coral reef fish larvae.Crossref | GoogleScholarGoogle Scholar |
Guiguen, Y., Fostier, A., Piferrer, F., and Chang, C.-F. (2010). Ovarian aromatase and estrogens: a pivotal role for gonadal sex differentiation and sex change in fish. General and Comparative Endocrinology 165, 352–366.
| Ovarian aromatase and estrogens: a pivotal role for gonadal sex differentiation and sex change in fish.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3cXktVegtQ%3D%3D&md5=f0e2a5989788b735776ba61c3c6dffafCAS |
Haddy, J. A., and Pankhurst, N. W. (1999). Stress-induced changes in concentrations of plasma sex steroids in black bream. Journal of Fish Biology 55, 1304–1316.
| Stress-induced changes in concentrations of plasma sex steroids in black bream.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD3cXht1CmsQ%3D%3D&md5=22424bb4c4bc6952e920d9e571f30832CAS |
Hoegh-Guldberg, O., Mumby, P. J., Hooten, A. J., Steneck, R. S., Greenfield, P., et al. (2007). Coral reefs under rapid climate change and ocean acidification. Science 318, 1737–1742.
| Coral reefs under rapid climate change and ocean acidification.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2sXhsVWhu7fN&md5=b074c829a513c1a5f7a4dd1596f39ea3CAS |
Houde, E. D. (1989). Comparative growth, mortality, and energetics of marine fish larvae – temperature and implied latitudinal effects. Fishery Bulletin 87, 471–495.
Howell, B. R., Day, O. J., Ellis, T., and Baynes, S. M. (1998). Early life stages of farmed fish. In ‘Biology of Farmed Fish’. (Eds K. D. Black and A. D. Pickering.) pp. 27–66. (Sheffield Academic Press: Sheffield.)
Inaba, K., Dreanno, C., and Cosson, J. (2003). Control of flatfish sperm motility by CO2 and carbonic anhydrase. Cell Motility and the Cytoskeleton 55, 174–187.
| Control of flatfish sperm motility by CO2 and carbonic anhydrase.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD3sXmtFWjtLg%3D&md5=9ce0cce0d6c1f64065abfe2dea09560dCAS |
Ishimatsu, A., Hayashi, M., and Kikkawa, T. (2008). Fishes in high-CO2, acidified oceans. Marine Ecology Progress Series 373, 295–302.
| Fishes in high-CO2, acidified oceans.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD1MXisVCrsLo%3D&md5=4929319c5a0d6651489b1368b768a496CAS |
Janhunen, M., Piironen, J., and Peuhkuri, N. (2010). Parental effects on embryonic viability and growth in Arctic charr Salvelinus alpinus at two incubation temperatures. Journal of Fish Biology 76, 2558–2570.
| Parental effects on embryonic viability and growth in Arctic charr Salvelinus alpinus at two incubation temperatures.Crossref | GoogleScholarGoogle Scholar | 1:STN:280:DC%2BC3crhtlaqsw%3D%3D&md5=6c61de340b635518e42fb6f6bc4bce52CAS |
Jobling, M. (1997). Temperature and growth: modulation of growth rate via temperature change. In ‘Global Warming: Implications for Freshwater and Marine Fish’. (Eds C. M. Wood and D. G. McDonald.) pp. 225–253. (Cambridge University Press: Cambridge.)
Jobling, M., Johnsen, H. K., Pettersen, G. W., and Henderson, R. J. (1995). Effect of temperature on reproductive development in Arctic charr, Salvelinus alpinus (L.). Journal of Thermal Biology 20, 157–165.
| Effect of temperature on reproductive development in Arctic charr, Salvelinus alpinus (L.).Crossref | GoogleScholarGoogle Scholar |
Jonsson, B., and Jonsson, N. (2009). A review of the likely effects of climate change on anadromous Atlantic salmon Salmo salar and brown trout Salmo trutta with particular reference to water temperature and flow. Journal of Fish Biology 75, 2381–2447.
| A review of the likely effects of climate change on anadromous Atlantic salmon Salmo salar and brown trout Salmo trutta with particular reference to water temperature and flow.Crossref | GoogleScholarGoogle Scholar | 1:STN:280:DC%2BC3cjnsVOqug%3D%3D&md5=eb886b896e7dc76914ee588406a3fdf9CAS |
Kikkawa, T., Ishimatsu, A., and Kita, J. (2003). Acute CO2 tolerance during the early developmental stages of four marine teleosts. Environmental Toxicology 18, 375–382.
| Acute CO2 tolerance during the early developmental stages of four marine teleosts.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD3sXpsVCmtrk%3D&md5=30e23985093239d95b1814d2fb5170faCAS |
Kikkawa, T., Kita, J., and Ishimatsu, A. (2004). Comparison of the lethal effect of CO2 and acidification on red sea bream (Pagrus major) during the early developmental stages. Marine Pollution Bulletin 48, 108–110.
| Comparison of the lethal effect of CO2 and acidification on red sea bream (Pagrus major) during the early developmental stages.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2cXisFKhtQ%3D%3D&md5=6a6fb26dd672421bdf430907ce030cdcCAS |
King, H. R., Pankhurst, N. W., and Watts, M. (2007). Reproductive sensitivity to elevated water temperatures in female Atlantic salmon is heightened at certain stages of vitellogenesis. Journal of Fish Biology 70, 190–205.
| Reproductive sensitivity to elevated water temperatures in female Atlantic salmon is heightened at certain stages of vitellogenesis.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2sXis1Sjsr8%3D&md5=0d8ebdc8f30c79e1c8e493f6fb3a9ac9CAS |
Lambert, Y., Dutil, J.-D., and Ouellet, P. (2000). Nutritional condition and reproductive success in wild fish populations. In ‘Proceedings of the 6th International Symposium on the Reproductive Physiology of Fish’. (Eds B. Norberg, O. S. Kjesbu, G-L. Taranger, E. Andersson and S. O. Stefansson.) pp. 77–84. (John Grieg A/S: Bergen.)
Leatherland, J. F., Li, M., and Barkataki, S. (2010). Stressors, glucocorticoids and ovarian function in teleosts. Journal of Fish Biology 76, 86–111.
| Stressors, glucocorticoids and ovarian function in teleosts.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3cXkt1agsLY%3D&md5=31b1499b5214c412597eef87d4ed6f49CAS |
Levavi-Sivan, B., Bogerd, J., Mañanós, E. L., Gómez, A., and Lareyre, J. J. (2010). Perspectives on fish gonadotropins and their receptors. General and Comparative Endocrinology 165, 412–437.
| Perspectives on fish gonadotropins and their receptors.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3cXktVensg%3D%3D&md5=9099d6245c87141d4429d17b917d1ed2CAS |
Lim, B.-S., Kagawa, H., Gen, K., and Okuzawa, K. (2003). Effects of water temperature on the gonadal development and expression of steroidogenic enzymes in the gonad of juvenile red seabream, Pagrus major. Fish Physiology and Biochemistry 28, 161–162.
| Effects of water temperature on the gonadal development and expression of steroidogenic enzymes in the gonad of juvenile red seabream, Pagrus major.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2cXks1Wqtro%3D&md5=b04a01b9b85ff4ced5ef6f9d368fa5adCAS |
Lo-Yat, A., Meekan, M. G., Lecchini, D., Martinez, E., Galzin, R., et al. (2011). Extreme climatic events reduce ocean productivity and larval supply in a tropical reef ecosystem. Global Change Biology 17, 1695–1702.
| Extreme climatic events reduce ocean productivity and larval supply in a tropical reef ecosystem.Crossref | GoogleScholarGoogle Scholar |
Lough, J. M., and Hobday, A. J. (2011). Observed climate change in Australian marine and freshwater environments. Marine and Freshwater Research 62, 984–999.
| Observed climate change in Australian marine and freshwater environments.Crossref | GoogleScholarGoogle Scholar |
McCormick, M. I., and Molony, B. W. (1995). Influence of water temperature during the larval stage on size, age and body condition of a tropical reef fish at settlement. Marine Ecology Progress Series 118, 59–68.
| Influence of water temperature during the larval stage on size, age and body condition of a tropical reef fish at settlement.Crossref | GoogleScholarGoogle Scholar |
Meekan, M. G., Ackerman, J. L., and Wellington, G. M. (2001). Demography and age structures of coral reef damselfishes in the tropical eastern Pacific Ocean. Marine Ecology Progress Series 212, 223–232.
| Demography and age structures of coral reef damselfishes in the tropical eastern Pacific Ocean.Crossref | GoogleScholarGoogle Scholar |
Meekan, M. G., Carleton, J. H., McKinnon, A. D., Flynn, K., and Furnas, M. (2003). What determines the growth of tropical reef fish larvae in the plankton: food or temperature? Marine Ecology Progress Series 256, 193–204.
| What determines the growth of tropical reef fish larvae in the plankton: food or temperature?Crossref | GoogleScholarGoogle Scholar |
Melzner, F., Gutowska, M. A., Langenbuch, M., Dupont, S., Lucassen, M., et al. (2009). Physiological basis for high CO2 tolerance in marine ectothermic animals: pre-adaptation through lifestyle and ontogeny? Biogeosciences 6, 2313–2331.
| Physiological basis for high CO2 tolerance in marine ectothermic animals: pre-adaptation through lifestyle and ontogeny?Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3cXltVCqsA%3D%3D&md5=6caa9b72c3c5ff2219c0190e3914df39CAS |
Michael, S. W. (2008). ‘Damselfishes and Anemonefishes: the Complete Illustrated Guide to Their Identification, Behaviours, and Captive Care.’ (TFH: New York.)
Migaud, H., Davie, A., and Taylor, J. F. (2010). Current knowledge on the photoneuroendocrine regulation of reproduction in temperate fish species. Journal of Fish Biology 76, 27–68.
| Current knowledge on the photoneuroendocrine regulation of reproduction in temperate fish species.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3cXkt1agsLs%3D&md5=5e46db9c129c7d4d0c712cde225f0de1CAS |
Modig, C., Westerlund, L., and Olsson, P.-E. (2007). Oocyte zona pellucida proteins. In ‘The Fish Oocyte; From Basic Studies to Biotechnical Applications’. (Eds P. J. Babin, J. Cerdá and E. Lubzens.) pp. 113–139. (Springer: Dordrecht.)
Morrongiello, J. R., Beatty, S. J., Bennett, J. C., Crook, D. A., Ikedife, D. N. E. N., Kennard, M. J., Kerezsy, A., Lintermans, M., McNeil, D. G., Pusey, B. J., and Rayner, T. (2011). Climate change and its implications for Australia's freshwater fish. Marine and Freshwater Research 62, 1082–1098.
| Climate change and its implications for Australia's freshwater fish.Crossref | GoogleScholarGoogle Scholar |
Munday, P. L., White, J. W., and Warner, R. R. (2006). A social basis for the development of primary males in a sex-changing fish. Proceedings of the Royal Society Series B, Biological Sciences 273, 2845–2851.
| A social basis for the development of primary males in a sex-changing fish.Crossref | GoogleScholarGoogle Scholar |
Munday, P. L., Jones, G. P., Pratchett, M. S., and Williams, A. J. (2008a). Climate change and the future for coral reef fishes. Fish and Fisheries 9, 261–285.
| Climate change and the future for coral reef fishes.Crossref | GoogleScholarGoogle Scholar |
Munday, P. L., Kingsford, M. J., O’Callaghan, M., and Donelson, J. M. (2008b). Elevated temperature restricts growth potential of the coral reef fish Acanthochromis polyacanthus. Coral Reefs 27, 927–931.
| Elevated temperature restricts growth potential of the coral reef fish Acanthochromis polyacanthus.Crossref | GoogleScholarGoogle Scholar |
Munday, P. L., Donelson, J. M., Dixson, D. L., and Endo, G. G. K. (2009a). Effects of ocean acidification on the early life history of a tropical marine fish. Proceedings of the Royal Society Series B, Biological Sciences 276, 3275–3283.
| Effects of ocean acidification on the early life history of a tropical marine fish.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD1MXhtFCgsLrJ&md5=39f3ac5437e108caa78e0f735a3df105CAS |
Munday, P. L., Crawley, N. E., and Nilsson, G. E. (2009b). Interacting effects of elevated temperature and ocean acidification on the aerobic performance of coral reef fishes. Marine Ecology Progress Series 388, 235–242.
| Interacting effects of elevated temperature and ocean acidification on the aerobic performance of coral reef fishes.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD1MXht1SrtrnM&md5=d4ff6d30a9d56ac2d9fc60193d0456efCAS |
Munday, P. L., Leis, J. M., Lough, J. M., Paris, C. B., Kingsford, M. J., et al. (2009c). Climate change and coral reef connectivity. Coral Reefs 28, 379–395.
| Climate change and coral reef connectivity.Crossref | GoogleScholarGoogle Scholar |
Munday, P. L., Dixson, D. L., Donelson, J. M., Jones, G. P., Pratchett, M. S., et al. (2009d). Ocean acidification impairs olfactory discrimination and homing ability of a marine fish. Proceedings of the National Academy of Sciences of the United States of America 106, 1848–1852.
| Ocean acidification impairs olfactory discrimination and homing ability of a marine fish.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD1MXitV2isrg%3D&md5=b393a4ea34b06c3afc4a75c959e7efeeCAS |
Munday, P. L., Dixson, D. L., McCormick, M. I., Meekan, M., Ferrari, M. C. O., et al. (2010). Replenishment of fish populations is threatened by ocean acidification. Proceedings of the National Academy of Sciences of the United States of America 107, 12930–12934.
| Replenishment of fish populations is threatened by ocean acidification.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3cXpsVagur4%3D&md5=29915a1998e7190bf98e919f51aaec55CAS |
Munday, P. L., Gagliano, M., Donelson, J. M., Dixson, D. L., and Thorrold, S. R. (2011). Ocean acidification does not affect the early life history development of a tropical marine fish. Marine Ecology Progress Series 423, 211–221.
| Ocean acidification does not affect the early life history development of a tropical marine fish.Crossref | GoogleScholarGoogle Scholar |
Nilsson, G. E., Crawley, N., Lunde, I. G., and Munday, P. L. (2009). Elevated temperature reduces the respiratory scope of coral reef fishes. Global Change Biology 15, 1405–1412.
| Elevated temperature reduces the respiratory scope of coral reef fishes.Crossref | GoogleScholarGoogle Scholar |
O’Connor, M. I., Bruno, J. F., Gaines, S. D., Halpern, B. S., Lester, S. E., et al. (2007). Temperature control of larval dispersal and the implications for marine ecology, evolution, and conservation. Proceedings of the National Academy of Sciences of the United States of America 104, 1266–1271.
| Temperature control of larval dispersal and the implications for marine ecology, evolution, and conservation.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2sXht12jtL4%3D&md5=6c1c390793bbdd6527a6927e7658fc2eCAS |
Okuzawa, K., Kumakura, N., Gen, K., Yamaguchi, S., Lim, B.-S., et al. (2003). Effect of high water temperature on brain–pituitary–gonad axis of the red seabream during its spawning season. Fish Physiology and Biochemistry 28, 439–440.
| Effect of high water temperature on brain–pituitary–gonad axis of the red seabream during its spawning season.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2cXks1ajtrc%3D&md5=3872b6e95f409a7d497e07f406ba82b5CAS |
Pankhurst, N. W. (1998). Reproduction. In ‘Biology of Farmed Fish’. (Eds K. D. Black and A. D. Pickering.) pp. 1–26. (Sheffield Academic Press: Sheffield.)
Pankhurst, N. W. (2008). Gonadal steroids: Functions and patterns of change. In ‘Fish Reproduction’. (Eds M. J. Rocha, A. Arukwe and B. G. Kapoor.) pp. 67–111. (Science Publishers: Enfield, NH, USA.)
Pankhurst, N. W. (2011). The endocrinology of stress in fish: an environmental perspective. General and Comparative Endocrinology 170, 265–275.
| The endocrinology of stress in fish: an environmental perspective.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3MXht1Kntw%3D%3D&md5=1ca92080125b5483a9399c419ac10d27CAS |
Pankhurst, N. W., and King, H. R. (2010). Temperature and salmonid reproduction: implications for aquaculture. Journal of Fish Biology 76, 69–85.
| Temperature and salmonid reproduction: implications for aquaculture.Crossref | GoogleScholarGoogle Scholar | 1:STN:280:DC%2BC3cjnsV2qtQ%3D%3D&md5=d0d625b57b8a103bea2b8bc79dc91636CAS |
Pankhurst, N. W., and Porter, M. J. R. (2003). Cold and dark or warm and light: variations on the theme of environmental control of reproduction. Fish Physiology and Biochemistry 28, 385–389.
| Cold and dark or warm and light: variations on the theme of environmental control of reproduction.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2cXks1ajsb4%3D&md5=66f0f55fda1023fd073162ad0fc9b422CAS |
Pankhurst, N. W., and Thomas, P. M. (1998). Maintenance at elevated temperature delays the steroidogenic and ovulatory responsiveness of rainbow trout Oncorhynchus mykiss to luteinizing hormone releasing hormone analogue. Aquaculture 166, 163–177.
| Maintenance at elevated temperature delays the steroidogenic and ovulatory responsiveness of rainbow trout Oncorhynchus mykiss to luteinizing hormone releasing hormone analogue.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaK1cXkslyksLc%3D&md5=ee436eeeda946f3ba1847a7928fa3b7eCAS |
Pankhurst, N. W., and Van Der Kraak, G. (1997). Effects of stress on growth and reproduction. In ‘Fish Stress and Health in Aquaculture’. (Eds G. K. Iwama, A. D. Pickering, J. P. Sumpter and C. B. Schreck.) pp. 73–93. (Cambridge University Press: Cambridge.)
Pankhurst, N. W., Van Der Kraak, G., and Peter, R. E. (1995). Evidence that the inhibitory effects of stress on reproduction in teleost fish are not mediated by the action of cortisol on ovarian steroidogenesis. General and Comparative Endocrinology 99, 249–257.
| Evidence that the inhibitory effects of stress on reproduction in teleost fish are not mediated by the action of cortisol on ovarian steroidogenesis.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaK2MXnvVeks7w%3D&md5=c4a43608087ce79066348e4b1e961479CAS |
Pankhurst, N. W., Purser, G. J., Van Der Kraak, G., Thomas, P. M., and Forteath, G. N. R. (1996). Effect of holding temperature on ovulation, egg fertility, plasma levels of reproductive hormones and in vitro ovarian steroidogenesis in the rainbow trout Oncorhynchus mykiss. Aquaculture. 146, 277–290.
| Effect of holding temperature on ovulation, egg fertility, plasma levels of reproductive hormones and in vitro ovarian steroidogenesis in the rainbow trout Oncorhynchus mykiss.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaK2sXktlKrtw%3D%3D&md5=cedebe59a024370d4cf58f1f552634a1CAS |
Pankhurst, N. W., Fitzgibbon, Q. P., Pankhurst, P. M., and King, H. R. (2008). Habitat-related variation in reproductive endocrine condition in the coral reef damselfish Acanthochromis polyacanthus. General and Comparative Endocrinology 155, 386–397.
| Habitat-related variation in reproductive endocrine condition in the coral reef damselfish Acanthochromis polyacanthus.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD1cXislCktw%3D%3D&md5=715e0493f7cf08014b13360aaba84e53CAS |
Pankhurst, N. W., King, H. R., Anderson, K., Elizur, A., Pankhurst, P. M., et al. (2011). Thermal impairment of reproduction is differentially expressed in maiden and repeat spawning Atlantic salmon. Aquaculture 316, 77–87.
| Thermal impairment of reproduction is differentially expressed in maiden and repeat spawning Atlantic salmon.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3MXlsFGlt7g%3D&md5=8c8980c3ee9729dc43eb8157d323970dCAS |
Pauly, D., and Pullin, R. S. V. (1988). Hatching time in spherical pelagic marine eggs in response to temperature and egg size. Environmental Biology of Fishes 22, 261–271.
| Hatching time in spherical pelagic marine eggs in response to temperature and egg size.Crossref | GoogleScholarGoogle Scholar |
Planas, J. V., and Swanson, P. (2008). Physiological function of gonadotropins in fish. In ‘Fish Reproduction’. (Eds M. J. Rocha, A. Arukwe and B. G. Kapoor.) pp. 37–66. (Science Publishers: Enfield, NH, USA.)
Pörtner, H. O., and Farrell, A. P. (2008). Physiology and climate change. Science 322, 690–692.
| Physiology and climate change.Crossref | GoogleScholarGoogle Scholar |
Pörtner, H. O., Langenbuch, M., and Reipschlager, A. (2004). Biological impact of elevated ocean CO2 concentrations: lessons from animal physiology and earth history. Journal of Oceanography 60, 705–718.
| Biological impact of elevated ocean CO2 concentrations: lessons from animal physiology and earth history.Crossref | GoogleScholarGoogle Scholar |
Reddin, D. G., Helbig, J., Thomas, A., Whitehouse, B. G., and Friedland, K. D. (2000). Survival of Atlantic salmon (Salmo salar L.) related to marine climate. In ‘The Ocean Life of Atlantic Salmon’. (Ed. D. Mills.) pp. 88–91. (Blackwell Science: Oxford.)
Reyjol, Y., Léna, J.-P., Hervant, F., and Pont, D. (2009). Effects of temperature on biological and biochemical indicators of the life-history strategy of bullhead Cottus gobio. Journal of Fish Biology 75, 1427–1445.
| Effects of temperature on biological and biochemical indicators of the life-history strategy of bullhead Cottus gobio.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD1MXhs1Slur7M&md5=1c7f6d6959886e8a4a7ac101a88abce8CAS |
Robertson, D. R. (1991). The role of adult biology in the timing of spawning of tropical reef fishes. In ‘The Ecology of Fishes on Coral Reefs’. (Ed. P. F. Sale.) pp. 357–386. (Academic Press: San Diego.)
Rombough, P. J. (1997). The effects of temperature on embryonic and larval development. In ‘Global Warming: Implications for Freshwater and Marine Fish’. (Eds C. M. Wood and D. G. McDonald.) pp. 177–223. (Cambridge University Press: Cambridge.)
Schreck, C. B. (2010). Stress and fish reproduction: the roles of allostasis and hormesis. General and Comparative Endocrinology 165, 549–556.
| Stress and fish reproduction: the roles of allostasis and hormesis.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3cXktVentQ%3D%3D&md5=026f2f41d60ecb00cb956263e9cec28cCAS |
Scott, S. G., and Pankhurst, N. W. (1992). Interannual variation in the reproductive cycle of the New Zealand snapper Pagrus auratus (Bloch & Schneider) (Sparidae). Journal of Fish Biology 41, 685–696.
| Interannual variation in the reproductive cycle of the New Zealand snapper Pagrus auratus (Bloch & Schneider) (Sparidae).Crossref | GoogleScholarGoogle Scholar |
Shimizu, A. (2003). Effect of photoperiod and temperature on gonadal activity and plasma steroid levels in a reared strain of the mummichog (Fundulus heteroclitus) during different phases of its annual reproductive cycle. General and Comparative Endocrinology 131, 310–324.
| Effect of photoperiod and temperature on gonadal activity and plasma steroid levels in a reared strain of the mummichog (Fundulus heteroclitus) during different phases of its annual reproductive cycle.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD3sXjtFWmtLs%3D&md5=65d8a1703e4b97a574552ece56328be2CAS |
Skelly, D. K., Joseph, L. N., Possingham, H. P., Freidenburg, L. K., Farrugia, T. J., et al. (2007). Evolutionary responses to climate change. Conservation Biology 21, 1353–1355.
| Evolutionary responses to climate change.Crossref | GoogleScholarGoogle Scholar |
Smith, A. M. (2009). Bryozoans as southern sentinels of ocean acidification: a major role for a minor phylum. Marine and Freshwater Research 60, 475–482.
| Bryozoans as southern sentinels of ocean acidification: a major role for a minor phylum.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD1MXmtlOms70%3D&md5=3a4d5f476c9a2aa59bf018271f974778CAS |
Sogard, S. M. (1997). Size-selective mortality in the juvenile stage of teleost fishes: a review. Bulletin of Marine Science 60, 1129–1157.
Soria, F. N., Strüssman, C. A., and Miranda, L. A. (2008). High water temperatures impair the reproductive ability of the pejerrey fish Odontesthes bonariensis: effects on the hypophyseal–gonadal axis. Physiological and Biochemical Zoology 81, 898–905.
| High water temperatures impair the reproductive ability of the pejerrey fish Odontesthes bonariensis: effects on the hypophyseal–gonadal axis.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD1cXhsFansbzF&md5=323e292c47a097e96b1ec577a68a516fCAS |
Sponaugle, S., and Cowen, R. K. (1996). Larval supply and patterns of recruitment for two Caribbean reef fishes, Stegastes partitus and Acanthurus bahianus. Marine and Freshwater Research 47, 433–447.
| Larval supply and patterns of recruitment for two Caribbean reef fishes, Stegastes partitus and Acanthurus bahianus.Crossref | GoogleScholarGoogle Scholar |
Stacey, N. E. (1984). Control of the timing of ovulation by exogenous and endogenous factors. In ‘Fish Reproduction: Strategies and Tactics’. (Eds G. W. Potts and R. J. Wootton.) pp. 207–222. (Academic Press: London.)
Tveiten, H., and Johnsen, H. K. (2001). Thermal influences on temporal changes in plasma testosterone and oestradiol-17β concentrations during gonadal recrudescence in female common wolffish. Journal of Fish Biology 59, 175–178.
| 1:CAS:528:DC%2BD3MXksF2mtLk%3D&md5=95b8301bc218710a1c6d1c2103b88da2CAS |
Tveiten, H., Solevåg, S. E., and Johnsen, H. K. (2001). Holding temperature during the breeding season influences final maturation and egg quality in common wolffish. Journal of Fish Biology 58, 374–385.
| Holding temperature during the breeding season influences final maturation and egg quality in common wolffish.Crossref | GoogleScholarGoogle Scholar |
Tyler, C. R., Santos, E. M., and Prat, F. (2000). Unscrambling the egg – cellular, biochemical, molecular and endocrine advances in oogenesis. In ‘Proceedings of the 6th International Symposium on the Reproductive Physiology of Fish’. (Eds B. Norberg, O. S. Kjesbu, G-L. Taranger, E. Andersson and S. O. Stefansson.) pp. 273–280. (John Grieg A/S: Bergen.)
Van Der Kraak, G., and Pankhurst, N. W. (1997). Temperature effects on the reproductive performance of fish. In ‘Global Warming: Implications for Freshwater and Marine Fish’. (Eds C. M. Wood and D. G. McDonald.) pp. 159–176. (Cambridge University Press: Cambridge.)
Wahl, B., and Löffler, H. (2009). Influences on the natural reproduction of whitefish (Coregonus lavaretus) in Lake Constance. Canadian Journal of Fisheries and Aquatic Sciences 66, 547–556.
| Influences on the natural reproduction of whitefish (Coregonus lavaretus) in Lake Constance.Crossref | GoogleScholarGoogle Scholar |
Widdicombe, S., and Spicer, J. I. (2008). Predicting the impact of ocean acidification on benthic biodiversity: what can animal physiology tell us? Journal of Experimental Marine Biology and Ecology 366, 187–197.
| Predicting the impact of ocean acidification on benthic biodiversity: what can animal physiology tell us?Crossref | GoogleScholarGoogle Scholar |
Wilson, D. T., and Meekan, M. G. (2002). Growth-related advantages for survival to the point of replenishment in the coral reef fish Stegastes partitus (Pomacentridae). Marine Ecology Progress Series 231, 247–260.
| Growth-related advantages for survival to the point of replenishment in the coral reef fish Stegastes partitus (Pomacentridae).Crossref | GoogleScholarGoogle Scholar |
Zohar, Y., Muñoz-Cueto, J. A., Elizur, A., and Kah, O. (2010). Neuroendocrinology of reproduction in teleost fish. General and Comparative Endocrinology 165, 438–455.
| Neuroendocrinology of reproduction in teleost fish.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3cXktVensw%3D%3D&md5=7946053c849bf4e3daaba0bd213dfcf2CAS |