

Twenty-five-year longevity of European hake (Merluccius merluccius) from novel use of bomb radiocarbon dating in the Mediterranean Sea
Sergio Vitale A C , Allen H. Andrews B , Pietro Rizzo A , Salvatore Gancitano A and Fabio Fiorentino AA National Research Council, Institute for Coastal Marine Environment, Via L. Vaccara, 61, I-91026 Mazara del Vallo, TP, Italy.
B NOAA Fisheries, Pacific Islands Fisheries Science Center, 1845 Wasp Boulevard, Honolulu, HI 96818, USA.
C Corresponding author: Email: sergio.vitale@cnr.it
Marine and Freshwater Research 67(7) 1077-1080 https://doi.org/10.1071/MF15376
Submitted: 9 March 2015 Accepted: 25 January 2016 Published: 24 May 2016
Journal Compilation © CSIRO Publishing 2016 Open Access CC BY-NC-ND
Abstract
The high variability of growth and longevity estimates for European hake (Merluccius merluccius) reflects the existence of two opposing hypotheses on growth rates that differ by a factor of 2: (1) a fast-growing hypothesis (FGH) with a maximum age near 15 years; and (2) a slow-growing hypothesis (SGH) with a maximum age near 30 years. A recently established regional radiocarbon (14C) reference led to a first-time application of bomb 14C dating in the Mediterranean Sea to three of the largest-sized and potentially oldest-catch female European hake. Because age reading of otoliths is very subjective and poorly defined, these fish were aged blind with bomb radiocarbon (14C) dating as an independent estimate of validated age. The validated ages were compared with the theoretical maximum ages from the most reliable FGH and SGH von Bertalanffy growth functions. Among the three bomb 14C ages, the most diagnostic length-at-age was an alignment with the bomb 14C rise period for two of the three fish, providing validated ages of 22 years (74.5-cm total length) and 25 years (88-cm total length). The results provide estimates of length-at-age that are in agreement with the SGH and cannot be accounted for by the FGH.
Additional keywords: age validation, carbon-14, growth, lifespan, Merlucciidae.
Introduction
The potential longevity of commercial species is relevant to fisheries management because of its relationship to key parameters in stock assessment models, such as growth rates and estimates of mortality (Tyler et al. 1989). This is particularly true in the case of the European hake (Merluccius merluccius L., 1758) because it is the dominant commercial fish species in demersal fisheries, both in the north-eastern Atlantic and Mediterranean Sea (Alheit and Pitcher 2012). Age investigations for this species have focussed primarily on interpretation of larval, juvenile and adult fish otoliths, with little concern for the accuracy of longevity estimates. Uncertainties on the validity of current age estimation methods, which vary considerably in age reading protocol, are widely reported and controversial (e.g. Orsi-Relini et al. 1992; Morales-Nin et al. 1998; García‐Rodríguez and Esteban 2002; de Pontual et al. 2006; Mellon-Duval et al. 2010).
Based on the use of contradictory age-reading approaches in estimating European hake growth (e.g. Ragonese et al. 2012), two opposing hypotheses are under consideration in the present study: (1) a fast-growing hypothesis (FGH), with a maximum age near 15 years (e.g. García‐Rodríguez and Esteban 2002; Mellon-Duval et al. 2010); and (2) a slow-growing hypothesis (SGH), with a maximum age near 30 years (e.g. Aldebert and Carries 1989; Levi et al. 1994). Thus, European hake longevity differs by a factor of 2 and is still a challenge for stock assessments that are regularly conducted by the General Fisheries Commission for the Mediterranean (GFCM).
A recently established regional radiocarbon (14C) reference in the Ligurian Sea from a 50-year-old coral led to an opportunity to apply bomb 14C dating in the Mediterranean Sea. The aims of the present study were to: (1) independently estimate the age of three of the largest female European hake using bomb 14C dating (e.g. Andrews et al. 2012); and (2) compare these validated age estimates with the maximum theoretical ages derived from contrasting von Bertalanffy growth functions for European hake of the Mediterranean Sea.
Material and methods
The available otoliths of large female European hake caught off the South of Sicily (Geographical Sub-Area (GSA) 16) in the late 1980s–90s were chosen from the archive of the Institute for Marine and Coastal Environment unit of Mazara del Vallo, Italy (IAMC-CNR; Table 1). Because age reading of otoliths is very subjective and poorly defined, bomb 14C dating was used as an independent estimate of age. The effectiveness of the bomb 14C assays was based on possible birth years of either 1958–1966 (SGH) or 1967–1983 (FGH). This determination would be diagnostic in terms of validated age because of the well-differentiated 14C levels defined by a regional coral reference and would be independent of age-reading protocol.

|
Radiocarbon analyses were performed by Beta Analytic Radiocarbon Dating (www.radiocarbon.com, accessed 21 April 2016) on ~9 mg samples of otolith core material. The extractions were made by hand grinding the external layers away from the core of the otolith and likely represented a few months of growth based on 1-year-old otoliths that weighed ~35 mg. Core extraction was guided by observations of a well-defined translucent band that is formed at settlement from the pelagic phase (Morales-Nin et al.1998). In addition, the concentric nature of the earliest layers provided a well-centred extraction. Otolith calcium carbonate was processed for 14C content using standard accelerator mass spectrometry procedures. The measured 14C values, reported as Δ14C based on a deviation from a pre-nuclear standard (Stuiver and Polach 1977), were used to calculate an approximate year-of-formation (birth year) from an alignment with a regional Δ14C reference series. Calculated fish age was based on the difference from collection year to the Δ14C reference alignment. The Δ14C reference was a recently validated time series from a 50-year-old coral (Cladocora caespitosa) from the Ligurian Sea (Tisnérat-Laborde et al. 2013). This Δ14C record is appropriate for direct comparison with the juvenile portion of otoliths (core material) because each formed in the mixed layer of the central Mediterranean where ocean circulation patterns and mixed layer stratification is well documented (Tisnérat-Laborde et al. 2013).
Two von Bertalanffy growth functions (VBGF) were chosen as a representation of each growth scenario (FGH and SGH; Table 2). In particular, the VBGF obtained by Mellon-Duval et al. (2010) and García‐Rodríguez and Esteban (2002) were considered to be the best representation of the FGH. Those produced by Aldebert and Carries (1989) and Levi et al. (1994) were chosen as the best representation of the SGH. The theoretical lifespan of European hake for each growth scenario, assuming longevity (tmax) is the time required to attain 95% of asymptotic length (L∞) (Froese and Binohlan 2000), was determined by replacing length at age t (lt) with 0.95 L∞ and solving for tmax as follows:
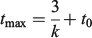
where k is the growth coefficient defined in Table 2.

|
The bomb 14C ages were ultimately compared with the theoretical tmax to evaluate its consistency with the longevity estimates from each VBGF scenario.
Results and discussion
Each otolith sample provided a robust 14C measurement that could be compared with the recently defined Δ14C reference series from the Ligurian Sea in the first application of bomb 14C dating of a fish species in the Mediterranean Sea (Table 1). The most diagnostic Δ14C measurement was from specimen EH-2 at 45.0‰ because of its well-defined relationship to the Δ14C rise period (before 1967). This Δ14C value was ~20‰ lower than the peak and post-peak plateau of the bomb 14C signal defined by the coral for the region (Fig. 1). Hence, the calibrated birth year of 1963 is well defined with a corresponding validated age of 22 years. The oldest fish was specimen EH-1, where the Δ14C value of 59.9‰ was calibrated to a birth year of 1966. This value is just below the beginning of the peak period, which approaches a low value of 65‰ in 1969, but is below the peak plateau (Fig. 1). Assuming there are no major fluctuations in the reference record plateau, the birth year of this fish was most likely 1966 for an age of 25 years. This finding is further supported by the more massive whole otolith of EH-1 relative to EH-2 (1.042 v. 0.705 g); given the mass–growth of European hake otoliths is a reasonable proxy for age. As expected, the least diagnostic Δ14C value was from specimen EH-3, the smallest fish used in the present study, at 66.3‰. In this case, the Δ14C value was within the range of Δ14C values for the peak plateau of the coral Δ14C record (Fig. 1). Hence, this fish could have had a birth year any time more recently than 1967; however, it can be clearly stated that this fish was not older than 18 years of age because of the temporal limitations of the known bomb 14C rise period throughout the marine environment (Grottoli and Eakin 2007). Similarly, specimens EH-1 and EH-2 could not have been older than 27 and 24 years of age respectively. This is assuming a 2-year margin of uncertainty in the coral reference record, similar to what has been observed elsewhere with other coral records (Andrews et al. 2012, 2013, 2015).

|
Otoliths have been the hard part of choice for European hake age reading in the Mediterranean Sea since the mid-1960s, but the feasibility debate continues (e.g. Piñeiro and Saínza 2003; Courbin et al. 2007). The findings of the present study provide a baseline from which this controversy can begin to be resolved. Use of the VBGF parameters from the FGH and SGH scenarios provided a theoretical range for maximum age (tmax) of 13–14 and 27–32 years respectively (Table 2). Because of the diagnostic alignment of the Δ14C values measured in the otolith cores of the two largest fish, it can be concluded that the FGH underestimates maximum age and is not a valid growth scenario for the entire lifespan of European hake (Tables 1, 2). Furthermore, it must be considered that Mellon-Duval et al. (2010) have validated by tag–recapture a high growth rate for 96 juvenile European hake females (<24-cm total length). At this time, the validated longevity of large adult European hake obtained in the present study and the tag–recapture validated early growth represent the only reliable length-at-age information for the Mediterranean. This indicates the full life history of European hake could be a combination of rapid early growth followed by a prolonged lifespan. In this regard, it is worth noting that the longevity reported herein refers to fish born in the 1960s and the increase of fishing effort over the past 50 years could have affected biological traits (Hidalgo et al. 2009). Moreover, throughout the Mediterranean, this species inhabits heterogeneous habitats characterised by a wide range of abiotic and biotic factors that might induce adaptive changes in growth. Despite the low sample size because of limited availability of European hake at the largest sizes and potentially oldest catch, the goal of estimating longevity (independent of age reading protocol) was achieved with bomb 14C dating.
References
Aldebert, Y., and Carries, J. C. (1989). La croissance du merlu dans le Golfe du Lion. Méthodologie. Problèmes posés. In ‘Report of the Age-Reading Workshop on Mediterranean Hake and Sardine’, 10–15 April 1989, Palma de Mallorca, Espana. (Eds P. Oliver, F. Alvarez, and B. Morales-Nin) Rapport sur les Peches 447, pp. 66–70. (FAO: Rome.)Alheit, J., and Pitcher, T. J. (Eds) (2012). ‘Hake: Biology, Fisheries and Markets.’ (Chapman and Hall: London.)
Andrews, A. H., DeMartini, E. E., Brodziak, J., Nichols, J. S., and Humphreys, R. L. (2012). A long-lived life history for a tropical, deep-water snapper (Pristipomoides filamentosus): bomb radiocarbon and lead–radium dating as extensions of daily increment analyses in otoliths. Canadian Journal of Fisheries and Aquatic Sciences 69, 1850–1869.
| A long-lived life history for a tropical, deep-water snapper (Pristipomoides filamentosus): bomb radiocarbon and lead–radium dating as extensions of daily increment analyses in otoliths.Crossref | GoogleScholarGoogle Scholar |
Andrews, A. H., Barnett, B. K., Allman, R. J., Moyer, R. P., and Trowbridge, H. D. (2013). Great longevity of speckled hind (Epinephelus drummondhayi), a deep-water grouper, with novel use of post-bomb radiocarbon dating in the Gulf of Mexico. Canadian Journal of Fisheries and Aquatic Sciences 70, 1131–1140.
| Great longevity of speckled hind (Epinephelus drummondhayi), a deep-water grouper, with novel use of post-bomb radiocarbon dating in the Gulf of Mexico.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3sXhtVehsbvO&md5=0dd4691aea55b33a6cb92fbc7f802c11CAS |
Andrews, A. H., Choat, J. H., Hamilton, R. J., and DeMartini, E. E. (2015). Refined bomb radiocarbon dating of two iconic fishes of the Great Barrier Reef. Marine and Freshwater Research 66, 305–316.
| Refined bomb radiocarbon dating of two iconic fishes of the Great Barrier Reef.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC2MXltVWnurY%3D&md5=b461360a5cedd669ad82563cb8a41c8cCAS |
Courbin, N., Fablet, R., Mellon, C., and De Pontual, H. (2007). Are hake otolith macrostructures randomly deposited? Insights from an unsupervised statistical and quantitative approach applied to Mediterranean hake otoliths. ICES Journal of Marine Science 64, 1191–1201.
de Pontual, H., Groison, A., Piñeiro, C., and Bertignac, M. (2006). Evidence of underestimation of European hake growth in the Bay of Biscay, and its relationship with bias in the agreed method of age estimation. ICES Journal of Marine Science 63, 1674–1681.
| Evidence of underestimation of European hake growth in the Bay of Biscay, and its relationship with bias in the agreed method of age estimation.Crossref | GoogleScholarGoogle Scholar |
Froese, R., and Binohlan, C. (2000). Empirical relationships to estimate asymptotic length, length at first maturity and length at maximum yield per recruit in fishes, with a simple method to evaluate length frequency data. Journal of Fish Biology 56, 758–773.
| Empirical relationships to estimate asymptotic length, length at first maturity and length at maximum yield per recruit in fishes, with a simple method to evaluate length frequency data.Crossref | GoogleScholarGoogle Scholar |
García‐Rodríguez, M., and Esteban, A. (2002). How fast does hake grow? A study on the Mediterranean hake (Merluccius merluccius L.) comparing whole otoliths readings and length frequency distributions data. Scientia Marina 66, 145–156.
Grottoli, G. G., and Eakin, C. M. (2007). A review of modern coral δ18O and Δ14C proxy records. Earth-Science Reviews 81, 67–91.
| A review of modern coral δ18O and Δ14C proxy records.Crossref | GoogleScholarGoogle Scholar |
Hidalgo, M., Massuti, E., Guijarro, B., Moranta, J., Ciannelli, L., Lloret, J., Oliver, P., and Stenseth, N. C. (2009). Population effects and changes in life history traits in relation to phase transitions induced by long-term fishery harvesting: European hake (Merluccius merluccius) off the Balearic Islands. Canadian Journal of Fisheries and Aquatic Sciences 66, 1355–1370.
| Population effects and changes in life history traits in relation to phase transitions induced by long-term fishery harvesting: European hake (Merluccius merluccius) off the Balearic Islands.Crossref | GoogleScholarGoogle Scholar |
Levi, D., Cannizzaro, L., Garofalo, G., Andreoli, M. G., Norrito, G., and Rizzo, P. (1994). An essay on hake gerontocracy, i.e. what to do with the elderly members of a populations? In ‘Estado actual de los conoeimientos de las poblaciones de merluza que habitan la plataforma continental atlàntica y mediterranea de la Union Europea con especial atencion a la Peninsula Ibérica’. (Eds A. Gonzáles-Garcés and F. J. Pereiro.) pp. 125–153. (Instituto Español de Oceanografía: Vigo, Spain)
Mellon-Duval, C., de Pontual, H., Metral, L., and Quemener, L. (2010). Growth of European hake (Merluccius merluccius) in the Gulf of Lions based on conventional tagging. ICES Journal of Marine Science 67, 62–70.
| Growth of European hake (Merluccius merluccius) in the Gulf of Lions based on conventional tagging.Crossref | GoogleScholarGoogle Scholar |
Morales-Nin, B., Torres, G. J., Lombarte, A., and Recasens, L. (1998). Otolith growth and age estimation in the European hake. Journal of Fish Biology 53, 1155–1168.
| Otolith growth and age estimation in the European hake.Crossref | GoogleScholarGoogle Scholar |
Orsi-Relini, L., Fiorentino, F., and Zamboni, A. (1992). Growth of the Mediterranean hake. Experiences gained in the Lugurian Sea. In ‘Proceedings of the 25th European Marine Biology Symposium’, 10–15 September 1990, Ferrara, Italy. (Eds G. Golombo, G. Ferreri, V. U. Ceccherelli, and R. Rossi.) pp. 307–315. (Olsen and Olsen, Printers and Publishers: Fredensborg, Denmark.)
Piñeiro, C., and Saínza, M. (2003). Age estimation, growth and maturity of the European hake, Merluccius merluccius (Linnaeus, 1758) from Iberian Atlantic waters. ICES Journal of Marine Science 60, 1086–1102.
| Age estimation, growth and maturity of the European hake, Merluccius merluccius (Linnaeus, 1758) from Iberian Atlantic waters.Crossref | GoogleScholarGoogle Scholar |
Ragonese, S., Vitale, S., Mazzola, S., Pagliarino, E., and Bianchini, M. L. (2012). Behavior of some growth performance indexes for exploited Mediterranean hake. Acta Adriatica 53, 105–122.
Stuiver, M., and Polach, H. A. (1977). Discussion: reporting of 14C data. Radiocarbon 19, 355–363.
Tisnérat-Laborde, N., Montagna, P., McCulloch, M., Siani, G., Silenzi, S., and Frank, N. (2013). A high-resolution coral-based Δ14C record of surface water processes in the western Mediterranean Sea. Radiocarbon 55, 1617–1630.
| A high-resolution coral-based Δ14C record of surface water processes in the western Mediterranean Sea.Crossref | GoogleScholarGoogle Scholar |
Tyler, A. V., Beamish, R. J., and McFarlane, G. A. (1989). Implications of age determination. Errors to yield estimates. In ‘Effects of Ocean Variability on Recruitment and an Evaluation of Parameters Used in Stock Assessment Models’. (Eds R. J. Beamish and G. A. Mc Farlane.) pp. 27–35. (Canadian Special Publication Fisheries and Aquatic Sciences: Vancouver, BC.)