

Effects of mowing regimes and climate variability on hay production of Leymus chinensis (Trin.) Tzvelev grassland in northern China
Taogetao Baoyin A , Frank Yonghong Li B C , Qinghai Bao A , Hugjiltu Minggagud A and Yankai Zhong AA Ecology, College of Life Sciences, Inner Mongolia University, Huhhot, Inner Mongolia 100013, China.
B AgResearch Grasslands Research Centre, Tennent Drive, Private Bag 11008, Palmerston North 4442, New Zealand.
C Corresponding author. Email: lifyhong@126.com
The Rangeland Journal 36(6) 593-600 https://doi.org/10.1071/RJ13088
Submitted: 30 August 2013 Accepted: 7 April 2014 Published: 17 June 2014
Journal Compilation © Australian Rangeland Society 2014
Abstract
Leymus chinensis (Trin.) Tzvelev grassland is the most widely distributed native steppe vegetation type suitable for haymaking in the semiarid pastoral regions of north-east Asia. The long-term effects of four mowing regimes (mowing once a year, twice a year, once every 2 years and twice every 3 years), and climatic variability on the hay production from L. chinensis grassland were investigated using permanent plots over 27 years. The results showed that (i) the overall cumulative annual herbage production over the 27 years was the highest under ‘mowing once a year’, and ‘mowing twice a year’ did not produce any significant change in the amount of harvested herbage; (ii) mowing induced a reduction in herbage production mostly in the first couple of years, and the reduction was larger under frequent than less frequent mowing. The annual herbage production in harvest years was higher under less frequent mowing (once every 2 years, or twice every 3 years) than that under annual mowing, but the higher herbage production in harvest years under less frequent mowing could not compensate for the herbage not made into hay in the years without harvest; (iii) annual herbage production generally increased with annual precipitation, but the response of annual herbage production to precipitation was best described by quadratic (instead of linear or logarithmic) equations, which suggested a decline in annual herbage production under the highest annual precipitation in the region, and the relationships stood under different mowing regimes; and (iv) an analysis of hay yield and quality, and costs of the mowing operation, showed that the haymaking from L. chinensis grassland was financially viable as long as the herbage production was not extremely low under drought. It is concluded that the best practice for haymaking from L. chinensis grassland should be based on the rule of ‘mowing once a year’ in high-production years and grazing in low-production years; and a light grazing early in the season may increase hay quality mowed in the autumn if the grassland grows well early in the season in high-production years.
Additional keywords: annual herbage production, climatic variation, long-term effects, net herbage accumulation, mowing frequency, semiarid steppe.
Introduction
Haymaking from natural steppe grassland is an important practice for preparing winter feed for livestock production in the semiarid region in north-east Asia, including northern China, Mongolia and East Russia (Suttie 2000). Leymus chinensis (Trin.) Tzvelev grassland is one of the most widely distributed steppe vegetation types in northern China, and has been traditionally used as rangeland for both livestock grazing and haymaking (Wu 1980). As a consequence of the shift in social regimes from nomadic to sedentary during the past several decades (Thwaites et al. 1998) and a recent change in the ‘land-use right’ from common to household in northern China (Li and Huntsinger 2011), it is becoming increasingly critical for pastoral farmers to prepare sufficient winter feed for stable livestock production. Leymus chinensis grassland is recognised as one of the best types of vegetation for haymaking (Wu 1980).
The L. chinensis grassland occurs on a range of soils along a gradient of climatic aridity, on the eastern Mongolian plateau and north-east China plain. The L. chinensis grassland communities, although dominated primarily by L. chinensis, vary significantly in species composition, and are categorised into two vegetation types, i.e. the ‘typical steppe’ grassland in the typical dry steppe environments, with Stipa grandis P. Smirn., Agropyron michnoi Roshev. and Cleistogenes squarrosa (Trin.) Keng as other major species, and the ‘meadow steppe’ grassland, which contain quite different other major species, such as Filifolium sibiricum (L.) Kitam., Stipa baicalensis Roshev. and Carex pediformis C. A. Meyer on relatively humid soils, or Suaeda glauca (Bunge) Bunge on alkaline or saline soils (Wu 1980; Wang and Ba 2008).
Mowing grassland from year to year without any fertiliser application is the normal practice in the semiarid region in northern China. A reduction in forage production as a result of mowing of native grasslands has been increasingly reported since the 1980s (Zhong and Piao 1988; Zhong 1991). To develop an appropriate mowing regime for the sustainable use of this important natural resource, a long-term experiment was conducted, starting in 1982, to investigate the effects of four mowing regimes (frequency) on the forage production and species composition of a L. chinensis ‘typical steppe’ grassland using permanent plots (Zhong 1991). The early results from this experiment have been documented and analysed in several papers. However, these early reports used only a part of the long-term dataset, either the data from the first several years (Zhong and Piao 1988; Zhong 1991) or used the data for a longer period (1982−97) but examined only the effects of one or two mowing regimes (Bao et al. 2004, 2005a, 2005b). In addition, these early reports focussed on the effects of mowing on species composition, diversity and herbage production. The net forage accumulation under different mowing regimes has not yet been fully analysed. Meanwhile, the effects of highly variable climate (especially precipitation) on forage production have also not yet been analysed in combination with the mowing regimes.
The effects of climate on the herbage production from ungrazed L. chinensis grasslands have been reported in several papers (Jiang 1988; Xiao et al. 1996; Bai et al. 2004; Wang and Zhou 2004; Wang et al. 2005) as well as under livestock grazing (Li et al. 2008; Wan et al. 2011), but not under mowing except for the early reports from this study (Zhong and Piao 1988; Bao et al. 2005a). All of these production studies (Jiang 1988; Xiao et al. 1996; Bai et al. 2004; Wang et al. 2005; Li et al. 2008; Wan et al. 2011) focussed on herbage production, which was always greater than the harvestable herbage from haymaking. The average residual height after mowing in the L. chinensis grassland was 6 cm in the region (Zhong 1991).
The objective of the present study was to use the full dataset (1982–2008) to extract the long-term effects of different mowing regimes and climatic variation on forage production from the L. chinensis grassland, and to recommend the best management practices for haymaking that are adapted to the highly variable semiarid climate. To use the long-term observations was necessary to capture the mowing-induced soil changes and its feedback on grassland production, and to capture the interactions between mowing and the large inter-annual variation in climate.
Materials and methods
Experimental site and grassland
The mowing experiment was established in a permanent 24-ha exclosure of natural L. chinensis grassland. The exclosure was established in 1978 by the Inner Mongolia Grassland Ecosystem Research Station of the Chinese Academy of Sciences, located in the Xilingol region of Inner Mongolia (latitude 43°269′N–44°089′N and longitude 116°049′E–117°059′E) (Jiang 1988). The region experiences a temperate semiarid climate, with an annual mean temperature of ~0.5°C and annual average precipitation of ~350 mm, 60–80% of which falls during the summer season from June to August. The precipitation has a large inter-annual variation, ranging from 180 to 550 mm. The L. chinensis steppe grassland is the major vegetation and is on a sandy-loam dark chestnut soil (or Calcic-orthic Aridisol in the US soil taxonomy classification system). The soil has a humus layer of 20–30 cm and a calcic layer at ~50 cm in depth (Jiang 1988; Wang and Cai 1988). Prior to exclosure, the grassland had been used as mowing land for haymaking but had also been subjected to grazing by sheep and cattle at a low stocking rate (Jiang 1988). Leymus chinensis is the dominant grass comprising 55 ± 15% (mean ± standard deviation of mean) of total forage production over the years. Other major species include tall bunchgrasses (mostly Stipa grandis and Agropyron michnoi) comprising 25 ± 12%, short bunchgrasses [Cleistogenes squarrosa and Koeleria cristata (L.) Schrad] and sedge (Carex korshingski Kom.) comprising 5 ± 4%, forbs comprising 11 ± 6% and legumes comprising 5 ± 4%.
Mowing experiment and measurements
An area of 24 × 20 m was chosen and marked using permanent plots for the mowing experiment (Fig. 1). The area is on a gentle slope (~3°) in the 24-ha permanent exclosure and livestock grazing was excluded. The six replicates of each of the four mowing treatments (mowing frequencies) were randomly assigned to six plots. The four treatments in the order of increasing cutting frequency were: (i) ‘1/2-cut’ – herbage harvested once every second year; (ii) ‘2/3-cut’ – herbage harvested twice every 3 years (one harvest per year for 2 successive years and no harvest for the third year); (iii) ‘1-cut’ – herbage harvested once every year; and (iv) ‘2-cut’ – herbage harvested twice every year. Each plot was 2 × 2 m, and 24 plots were used. The grasses were cut to 6 cm aboveground level over the whole plot at each harvest time, but only the herbage from the 1 × 1 m in the centre of each plot was collected and sorted to determine the dry matter (DM) weight and species composition. In addition, the control plots (six quadrats of 1 × 1 m) were harvested randomly around the permanent plots within a distance of 2–4 m to the boundary (once a year at the same time of harvesting as the ‘1-cut’ treatment), trying to avoid the previously harvested areas, to represent the grassland under no mowing (without effects of previous mowing) (Zhong 1991). The original design included more than 24 plots to investigate the effects of different cutting heights on grassland (the unlabelled plots in Fig. 1). These cutting height treatments ceased after the initial stage of the experiment. These unused plots were also cut to a height of 6 cm when the adjacent experimental plots were cut.

|
All live and standing dead herbage harvested from the 1 × 1-m quadrat in the centre of each plot was separated by species, oven-dried at 60°C and weighed. The herbage mass included the standing dead from the current year’s plant growth but not the standing dead from the previous year’s growth; this standing dead was easily identified (grey and partly rotten) and was excluded. The other 3 m2 surrounding the 1 × 1-m quadrat in each plot was also cut to the same residual height at the same time. The herbage production was harvested each year on 16 August (this might be 1–3 days later if the weather did not allow the harvest), except for the ‘2-cut’ treatment, which was harvested on both 15 June and 15 September. Mid August is the time when the herbage mass of grassland reaches its peak value (Xiao et al. 1996; Ma et al. 2010), and when the farmers start to mow grassland for hay in the region (Zhong and Piao 1988).
Data analysis
First, we examined the effects of mowing frequency on annual herbage production (AHP) and net herbage accumulation under the four mowing treatments over the 27-year period. There were 12, 16, 24, 24 and 24 harvest years, respectively, for treatment 1/2-cut, 2/3-cut, 1-cut, 2-cut and control. For AHP, the difference under any two treatments was tested using a paired t-test based on the AHP pairs in the years when both treatments have a harvest, with P < 0.05 indicting a significant difference. The trends in AHP across years over the whole experimental period were also examined. To separate the effects of mowing on the AHP from that of climate, the relative AHP under different mowing regimes compared with that of the no-mowing control was calculated [relative AHP = 100 × AHP under mowing/AHP under control (%)] and analysed.
Second, we explored the relationship between the variation in AHP and climatic factors, to see if the relationship was affected by the different mowing regimes. This was done by fitting different curve types to describe the AHP response to climatic variables (precipitation and temperature). Three curve types were tested: (i) linear – indicating a linear response of AHP to changes in climatic factors, (ii) logarithmic – indicating the change of AHP with a changing climatic factor being towards a maximum/minimum; and (iii) quadratic – indicating the highest AHP occurred at neither the low nor the high end of a climatic gradient. All analyses were conducted using SigmaPlot version 12.5 from Systat Software, Inc. (San Jose, CA, USA, SigmaPlot 2014). The goodness of the curve fit was judged by the coefficients of determination r2 (after adjusting the observations by subtracting their mean) and the significant levels P (ANOVA). A greater r2 and smaller P means a better fit, and a curve fit is significant when P < 0.05.
Results
Variation and trends in climate
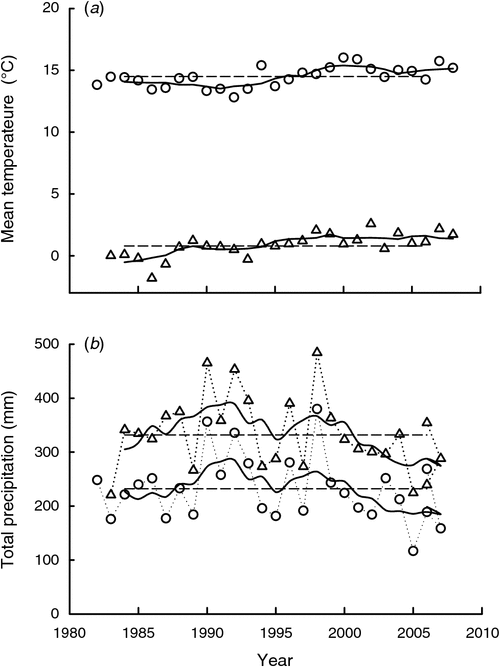
The annual mean temperature (mean of the daily mean temperature within a year, and the daily mean temperature is the mean of the eight observations 3 h apart within each 24-h period) was 0.8°C and had a significant increase (r2 = 0.51, P < 0.001) of 2.3°C over the 27-year period. The mean daily temperature during the plant growing season (1 April–20 August) was 14.5°C, and also showed a significant increase (r2 = 0.35, P = 0.001) of 1.8°C over the whole 27-year period (Fig. 2a). Annual precipitation averaged 332 mm during the 27-year period, 69% which fell during the growing season. Precipitation showed a large inter-annual variation, and was higher in the middle than in the early and late stages of the study. Precipitation in the growing season fitted a significant quadratic curve (r2 = 0.36 and P = 0.004) over the whole experimental period, though the annual precipitation showed only a similar trend (r2 = 0.19 and P = 0.095) (Fig. 2b).
|
Effects of mowing frequency on herbage production
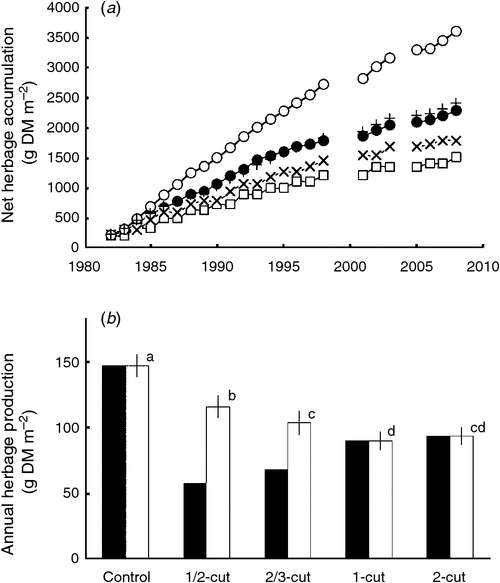
The total net herbage accumulation over the 27-year period was very similar between two treatments that harvested herbage every year (treatments 1-cut and 2-cut), and was higher under more frequent harvests, i.e. the total net herbage accumulation was in the order (2-cut = 1-cut) >2/3-cut >1/2-cut (Fig. 3a).
|
The AHP in a harvest year was higher under the low than the high cutting frequency. That is, the order of AHP averaged over the harvest years was 1/2-cut >2/3-cut > (2-cut = 1-cut) (Fig. 3b). The AHP in the control treatment was much higher (P < 0.001, paired t-test) than under any of the cutting treatments (Fig. 3b), and reflected an increasing difference between the herbage produced each year in the control plots compared with all the other treatments (Fig. 3a). However, since the AHP under the control was harvested from different areas in different years, the average AHP under the control over the experimental period does not represent a realisable herbage production from the grassland.
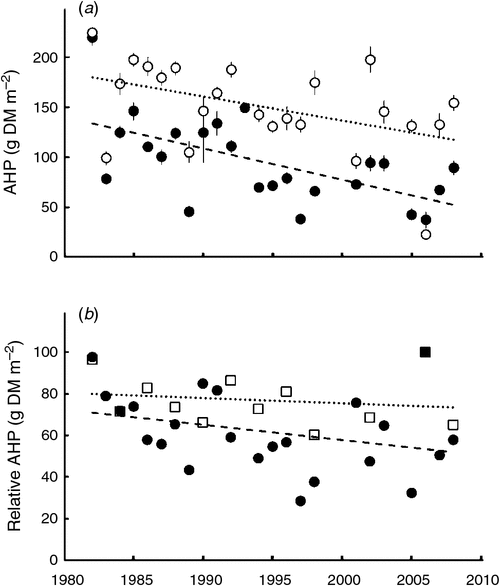
The AHP in harvest years showed a significant decline (P < 0.05) over the 27 years under treatment 1-cut and 2-cut (2-cut data not shown), but the decline was not significant under the other two mowing treatments or the control (P > 0.05) (Fig. 4a). The relative AHP under mowing against the control treatment showed no significant decline (P > 0.10) under all the mowing treatments (Fig. 4b), although a marked reduction in relative AHP appeared to have occurred in the first 1 or 2 years of the experimental period under all the mowing treatments.
|
Effects of climatic variation on herbage production
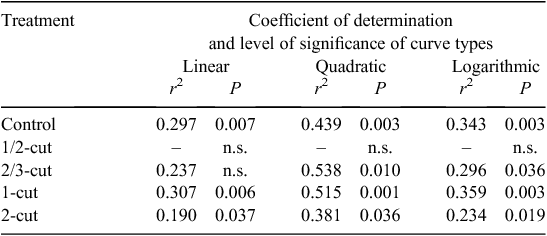
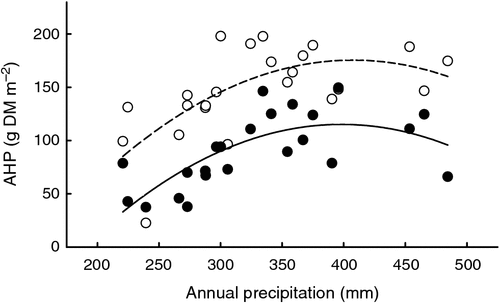
The AHP of the L. chinensis grassland showed a significant positive linear correlation with the precipitation accumulated over a 1-year period before harvest (annual r = precipitation from mid August to mid August) under all except the ‘1/2–cut’ treatment (Table 1). The increase of the AHP with annual precipitation was fitted with linear, logarithmic or quadratic equations, with the best fit from using quadratic equations, i.e. with the greatest coefficients of determination (r2) and significance levels (P) (Table 1 and Fig. 5). The shape of the response curves was similar for all treatments, with the maximum AHP being reached between 380 and 420 mm of precipitation (Fig. 5) The correlation of the AHP with precipitation (R) changed little between using the annual R and using the plant-growing-season R (accumulated from 1 April to harvest in mid August).
|
|
The AHP also showed a significant negative correlation with annual mean temperature under the 1-cut treatment (r2 = 0.241, P = 0.017), and a negative trend under 2/3-cut treatment (r2 = 0.326, P = 0061). No other significant relationships (linear, quadratic or logarithmic) were found between AHP and the annual or plant-growing-season mean temperature.
Discussion
Herbage production and mowing frequency
Our results show that harvesting more than once per year does not significantly increase the AHP from the L. chinensis grassland (Fig. 3a), and it also increases the operation cost of haymaking. This result is different from that obtained in temperate grasslands under more humid environments, where more herbage production is normally expected under multiple than single harvests in each year (Turner et al. 1993). The result in the present experiment probably means that the rate of growth of the herbage after mowing in mid June is similar to that of the plots that had not been harvested at this time. Plant canopy is still expanding in mid June (Xiao et al. 1996); removal of the herbage at this time does not lead to improved light and soil temperature conditions, or less plant senescence and littering, which stimulate plant growth and herbage accumulation. Our long-term observations do not support the results obtained in a 1-year cutting experiment on the typical steppe grassland in the same region by Schiborra et al. (2006) who found that multiple cuts lead to a higher herbage yield. A 1-year experiment cannot capture the feedback of the mowing-induced plant and soil changes (e.g. in root : shoot ratio and soil organic matter) to grassland production.
Our results also show that harvesting less than once a year (i.e. no harvest in some years) will inevitably result in a significant reduction in overall herbage production. That is, the greater herbage production in a harvest year from less frequently mowed grassland (e.g. 1/2-cut and 2/3-cut treatments) cannot compensate for the herbage not made into hay in the years without harvest.
The harvestable AHP across the experimental period showed a significant decrease (P < 0.05) under frequent mowing (1-cut and 2-cut treatments), and showed a non-significant decline under less frequent mowing (1/2-cut and 2/3-cut treatments) or no-mowing control (P > 0.05) (Fig. 4a). These declines are not only the consequences of long-term mowing but are also related to the increasing aridity during the experimental period across the years. The continuous drought during the last 8 years (less precipitation and high temperature during 2001–08, Fig. 2) might partly contribute to the general trend of a decline in grassland production over the experimental period. When the effects of inter-annual variation in climatic factors are removed, i.e. when the relative AHP under a mowing against control is considered, the relative AHP over the experimental period showed no significant decline under all the four mowing treatments (P > 0.10) (Fig. 4b). The marked reduction in relative AHP in the first 1 or 2 years (e.g. by 20 and 30% under the 1/2-cut and 1-cut treatments in Fig. 4b) suggests an immediate effect of mowing on grassland production. Mowing in 1 year will directly affect the herbage production in the following year. In other words, the grassland may reach a new equilibrium under a certain mowing regime within couple of years. This change in herbage production after mowing is most likely attributable to mowing resulting in lower nutrient and energy storage in belowground biomass (Turner et al. 1993) and to the changes in microclimate; significant higher soil temperature and lower soil moisture levels, which have been observed in mown compared with unmown grasslands (Shao et al. 2012). The nutrient removal by haymaking over the long-term period is another major factor attributable to the reduction in herbage production under mowing. Zhong et al. (2012) reported a significant decline in soil organic matter and mineral nitrogen content under mowing when compared with the control (measured in 1992, after mowing for 11 years). The decline is more pronounced in the 20–40-cm than the 0–20-cm soil layers, possibly reflecting the return of plant litter in upper soil layers while plant nutrient uptake may be from deeper in the soil profile.
Response in herbage production to variation in climate
In semiarid grasslands, precipitation plays a critical role in grassland production (Le Houerou et al. 1988; Knapp et al. 2006; Sala et al. 2012). That the best fit of the grassland production (AHP) to annual precipitation was a quadratic rather than a linear or logarithmic relationship (Table 1) indicates that plant production generally increases with the annual precipitation when the precipitation does not cover the high end of its variation range in the region. Plant water-use efficiency (DM production per mm precipitation) decreases as the precipitation increases. The result is a decline in plant production when the annual precipitation approaches the high end of its variation range in the region. While most of the previous research has examined the plant production-to-precipitation response using linear correlations in semiarid grasslands (Xiao et al. 1996; Wang et al. 2005; Knapp et al. 2006; Bai et al. 2008; Sala et al. 2012), our results, using the permanent plots, suggest that the relationship can be better described using quadratic instead of linear relationships. This non-linear relationship is supported by another independent dataset on the aboveground production of L. chinensis monitored in the same exclosure (Wang and Zhou 2004). This decline in herbage production under the highest precipitation in the region might be attributed to the lower solar radiation and temperature associated with high precipitation. The precipitation and mean daily temperature during the growing season (1 April–20 August) were negatively correlated (r2 = 0.183, P = 0.026) over the experimental period. It may also be due to an acceleration of plant tissue turnover with more-than-usual water supply (Ren et al. 2011), which may lead to a less herbage mass at harvest.
The relationship between AHP and precipitation at one site across years is different from their spatial relationship along the precipitation gradient. While our results suggest a non-linear temporal relationship between AHP and precipitation at the one site, it does not imply that a similar relationship exists for different parts of the steppe grassland with different total annual rainfall. The relationship between rainfall and herbage production across different parts of the grassland steppe has been reported as being linear (Li et al. 1994; Bai et al. 2008). Changes in the composition of plant species along such spatial gradients occurred (Li et al. 1994; Bai et al. 2008), whereas it was relatively stable in the studied permanent plots. No shift in species was observed in the studied grassland though the proportion of species in the herbage varied during the 27 years (Bao et al. 2004). The effects of precipitation on grassland production are not only determined by the amount of annual precipitation or precipitation in the growing season, but to a large extent also by the seasonal pattern (Knapp et al. 2006) and by the legacy of the precipitation and plant community structure from previous years (Sala et al. 2012). The extremely high precipitation found in 1990, 1992 and 1998 (Fig. 2), all had high rainfall events occurring in July and early August, before the harvest of herbage. The relationships between grassland herbage production and precipitation warrant further detailed studies.
The AHP shows a significant negative correlation with annual mean temperature under the 1-cut treatment (P = 0.017), and a non-significant negative trend under the 2/3-cut treatment (P = 0.061), which is most likely related with the negative correlation between temperature and precipitation in the region; a high temperature means less precipitation and more evaporation. The increase in temperature in the growing season (by 1.8°C over 27 years, Fig. 2) may also mean an increase in the length of the growing season, but its effect on the time of the maximum AHP, and thus the best time for haymaking, cannot be established. The seasonal pattern of herbage mass of the studied grassland has been monitored over a 22-year period (from 1982 to 2003; Ma et al. 2010); the data show that the maximum herbage mass appears mostly in August (the number of years with the maximum herbage mass in July, August and September are respectively 5, 15 and 2 years out of the 22 years). The maximum herbage mass in good rainfall years (1990, 1992 and 1998) are all in August. An increase in the herbage mass harvested in May, and a decline in the herbage mass harvested in September, has been detected over the 22-year period; the spring temperature increase and summer drought perhaps contributing to the increase in herbage mass in spring and the decline in the autumn (Ma et al. 2010).
Management implications
Mowing once a year would seem to be the most appropriate for the L. chinensis grasslands in the region. Less-frequent mowing (or no mowing in some years) allows the grassland to rest and increases AHP but this leads to a lower cumulative harvest of herbage from the grassland. Multiple mowing within a 1-year period does not produce more hay. These findings from a long-term experiment are very valuable for designing the best mowing regimes for the use of L. chinensis grasslands. However, non-yield factors that affect haymaking need to be considered when recommending the best practice. These factors include the heterogeneity of L. chinensis grasslands, the nutritive value of the hay under different mowing regimes and the costs of mowing and these are considered below.
-
The heterogeneity of L. chinensis grasslands. The L. chinensis grasslands vary significantly in species composition (Wu 1980). The production response of L. chinensis to mowing and to climate variation may be similar in different grassland communities, but the production response of these heterogeneous L. chinensis communities may also vary. The results from this long-term but one-site study are most applicable to the L. chinensis ‘typical steppe’ grasslands. Caution is necessary when applying the findings of this study to the L. chinensis ‘meadow steppe’ grasslands in other regions, although a mowing experiment on the L. chinensis meadow steppe grassland on the north-east China plain also recommended ‘mowing once a year’ as the appropriate practice (Zhang 1993).
-
The nutritive value of hay under different mowing regimes. The nutritive value of the hay in this experiment was not monitored over the whole experimental period but was determined during a 3-year period (Zhong et al. 2012). The results show that the hay has a higher crude protein content under mowing twice (134 g kg–1 DM) than mowing once a year (116 g kg–1 DM), which is in agreement with the results of Duo and Zhao (2001) and Schiborra et al. (2006) that the nutritive value of herbage of L. chinensis grasslands increases with cutting frequency. The crude protein content under the three mowing regimes when the herbage was harvested in August (1-cut, 2/3-cut, 1/2-cut) is very similar (107–116 g kg–1 DM), and the slight difference among them is mainly from that the different proportions of standing dead material in the herbage harvested (Zhong et al. 2012). When considering both the hay yield and nutritive value, it is obvious that any mowing regimes with grassland unused in any year (i.e. 1/2-cut or 2/3-cut) will lead to a waste of herbage production, so is not a choice in practice. Compared with mowing once a year, mowing twice a year (2-cut) increases the nutritive value of the hay (Zhong et al. 2012), though not yield, which appears to suggest mowing twice a year being a better choice than once a year. However, mowing twice a year will nearly double the cost of the mowing operation but result in little increase in profit, because of the low increase in yield. Another reason that local herders do not harvest hay in late June is to avoid the possible wet weather that may reduce the quality of the mown hay before it can be baled.
-
The cost-benefit of mowing. The current (2013) costs of haymaking from 1 ha of grassland consist of the cost of mowing (90 yuan ha–1) and baling (1.5 yuan per bale of 15 kg) (1 US$ = 6.2 yuan). For the average AHP of the studied grassland (1500 kg DM ha–1), the cost of haymaking will be 240 yuan ha–1. The price of the hay is ~0.9 yuan kg–1, which implies the value of harvestable hay is 1350 yuan ha–1. However, during the drought years, mowing hay from the short grassland might not be justified. For example, in the driest year over the experimental period, the grassland produced only 378 kg DM ha–1 harvestable herbage. Although the cost of haymaking (128 yuan ha–1) is less than the value of the hay made (340 yuan ha–1), it is obviously more justified to leave the grassland for grazing and so save the unnecessary cost of the mowing operation. Wan et al. (2011) and Schönbach et al. (2011) tested a mixed land-management system in the study region in which grasslands were alternatively grazed and mowed from year to year, and suggested it was a better system than the traditional continuous grazing system. Our results support the mixed grassland-management system but suggest that a more adaptive mowing strategy should be used as the basis of forage production, that is, to mow the grassland in high-production years and graze in low-production years. Our results also imply that, if the grassland grows well early in the season, to have a light grazing may increase the nutritive value of hay harvested in the autumn without increasing the operational cost.
Conclusions
Herbage production for haymaking from the L. chinensis typical steppe grassland is the highest under ‘mowing once a year’. More frequent ‘mowing twice a year’ does not increase the herbage production, while less frequent mowing (i.e. no mowing in some years) lead to a lower cumulative harvest of herbage. The AHP from the grassland generally increases with annual precipitation, but the production-to-precipitation response is best described by a quadratic relationship, which suggests a decline in herbage production under the highest annual precipitation in the region. The best practice for haymaking from the L. chinensis grassland needs to adapt to the climate-induced inter-annual variation in herbage production, and to consider both the hay yield and nutritive value, and the costs of mowing. We suggest that the best practice for haymaking should be based on the rule of ‘mowing once a year’ in high-production years and grazing in low-production years and a light grazing in the early season may also be used to increase the nutritive value of hay mowed in the autumn if the grassland grows well early in the season.
Acknowledgements
The long-term project site was maintained by the Inner Mongolia Grassland Ecosystem Research Station of the Chinese Academy of Sciences. This research was supported by the Special Fund for Agricultural Science Research in the Public Interest (201303060), the Ministry of Science and Technology of China for Restoration and Sustainable Use of the Steppe Grasslands in Semiarid Regions (2011BAC07B01), and the Inner Mongolia Fundamental Research Fund for Optimisation of Steppe Ecosystems Management (20110904). The authors thank R. D. B. Whalley and two anonymous referees for comments on an early version of the manuscript.
References
Bai, Y., Han, X., Wu, J., Chen, Z., and Li, L. (2004). Ecosystem stability and compensatory effects in the Inner Mongolia grassland. Nature 431, 181–184.| Ecosystem stability and compensatory effects in the Inner Mongolia grassland.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2cXntlGks7w%3D&md5=4521d03d8ef508af376edf636aff5f41CAS | 15356630PubMed |
Bai, Y., Wu, J., Xing, Q., Pan, Q., Huang, J., Yang, D., and Han, X. (2008). Primary production and rain-use efficiency across a precipitation gradient on the Mongolia Plateau. Ecology 89, 2140–2153.
| Primary production and rain-use efficiency across a precipitation gradient on the Mongolia Plateau.Crossref | GoogleScholarGoogle Scholar | 18724724PubMed |
Bao, Y., Li, Z., and Zhong, Y. (2004). Compositional dynamics of plant functional groups and their effects on stability of community ANPP during 17 years of mowing succession on Leymus chinensis steppe of Inner Mongolia, China. Acta Botanica Sinica 46, 1155–1162.
Bao, Q., Baoyin, T., Zhong, Y., Sun, W., and Liu, M. (2005a). Effects of different cutting systems on main typical steppe populations in Inner Mongolia. Chinese Journal of Applied Ecology 16, 2333–2338.
| 16515182PubMed |
Bao, Y., Li, Z., Zhong, Y., and Yang, C. (2005b). Effects of mowing frequency on the energy standing crops of major populations in a Leymus chinensis steppe of Inner Mongolia. Chinese Bulletin of Botany 22, 153–162.
Baoyin, T., and Li, F. Y. (2009). Can shallow ploughing and harrowing facilitate restoration of Leymus chinensis grassland? Results from a 24-year monitoring program. Rangeland Ecology and Management 62, 314–320.
| Can shallow ploughing and harrowing facilitate restoration of Leymus chinensis grassland? Results from a 24-year monitoring program.Crossref | GoogleScholarGoogle Scholar |
Duo, L., and Zhao, S. (2001). Effects on mowing grasslands of Leymus chinensis by the same stress in growing season. Acta Agresta Sinica 9, 25–33.
Jiang, S. (1988). Setting up the experimental sites for grassland ecosystem research and vegetation status. Research on Grassland Ecosystems 3, 1–12.
Knapp, A. K., Burns, C. E., Fynn, R. W. S., Kirkman, K. P., Morris, C. D., and Smith, M. D. (2006). Convergence and contingency in production–precipitation relationships in North American and South African C4 grasslands. Oecologia 149, 456–464.
| Convergence and contingency in production–precipitation relationships in North American and South African C4 grasslands.Crossref | GoogleScholarGoogle Scholar | 16821014PubMed |
Le Houerou, H. N., Bingham, R. L., and Skerbek, W. (1988). Relationship between the variability of primary production and the variability of annual precipitation in world arid lands. Journal of Arid Environments 15, 1–18.
Li, W., and Huntsinger, L. (2011). China’s grassland contract policy and its impacts on herder ability to benefit in Inner Mongolia: tragic feedbacks. Ecology and Society 16, .
Li, Y., Mo, W., Yang, C., and Ye, B. (1994). Aerial biomass and carrying capacity of steppe vegetation in Inner Mongolia and their relations with climate. Journal of Arid Land Resources and Environment 8, 43–50.
Li, Y., Wang, W., Liu, Z., and Jiang, S. (2008). Grazing gradient versus restoration succession of Leymus chinensis (Trin.) Tzvel. grassland in Inner Mongolia. Restoration Ecology 16, 572–583.
| Grazing gradient versus restoration succession of Leymus chinensis (Trin.) Tzvel. grassland in Inner Mongolia.Crossref | GoogleScholarGoogle Scholar |
Ma, W., Liu, Z., Wang, Z., Wang, W., Liang, C., Tang, Y., He, J.-S., and Fang, J. (2010). Climate change alters inter-annual variation of grassland above-ground productivity: evidence from a 22-year measurement series in the Inner Mongolian grassland Journal of Plant Research 123, 509–517.
| Climate change alters inter-annual variation of grassland above-ground productivity: evidence from a 22-year measurement series in the Inner Mongolian grasslandCrossref | GoogleScholarGoogle Scholar | 20130954PubMed |
Ren, H., Xu, Z., Huang, J., Clark, C., Chen, S., and Han, X. (2011). Nitrogen and water addition reduce leaf longevity of steppe species. Annals of Botany 107, 145–155.
| Nitrogen and water addition reduce leaf longevity of steppe species.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3cXhsF2rsr3F&md5=305511581c263c454e9b9f61aed47a82CAS | 21084404PubMed |
Sala, O. E., Gherardi, L. A., Reichmann, L., Jobbagy, E., and Peters, D. (2012). Legacies of precipitation fluctuations on primary production: theory and data synthesis. Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences 367, 3135–3144.
| Legacies of precipitation fluctuations on primary production: theory and data synthesis.Crossref | GoogleScholarGoogle Scholar | 23045711PubMed |
Schiborra, A., Gierus, M., Wan, H. W., Bai, Y. F., and Taube, F. (2006). Herbage yield and quality as influenced by cutting frequency of natural grassland of Inner Mongolia. Grassland Science in Europe 11, 592–594.
Schönbach, P., Wan, H., Gierus, M., Bai, Y., Müller, K., Lin, L., Susenbeth, A., and Taube, F. (2011). Grassland responses to grazing: effects of grazing intensity and management system in the Inner Mongolia steppe. Plant and Soil 340, 103–115.
| Grassland responses to grazing: effects of grazing intensity and management system in the Inner Mongolia steppe.Crossref | GoogleScholarGoogle Scholar |
Shao, C., Chen, J., Li, L., and Zhang, L. (2012). Ecosystem responses to mowing manipulations in an arid Inner Mongolia steppe: an energy perspective. Journal of Arid Environments 82, 1–10.
| Ecosystem responses to mowing manipulations in an arid Inner Mongolia steppe: an energy perspective.Crossref | GoogleScholarGoogle Scholar |
SigmaPlot (2014). Available at: www.sigmaplot.com (accessed 5 February 2014).
Suttie, J. M. (2000). ‘Hay and Straw Conservation.’ FAO Plant Production and Protection Series No. 29. (FAO: Rome, Italy.) Available at: www.fao.org/docrep/005/x7660e/x7660e00.htm (accessed 22 April 2014).
Thwaites, R., de Lacy, T., Li, Y. H., and Liu, X. H. (1998). Property rights, social change, and grassland degradation in Xilingol Biosphere Reserve, Inner Mongolia, China. Society & Natural Resources 11, 319–338.
| Property rights, social change, and grassland degradation in Xilingol Biosphere Reserve, Inner Mongolia, China.Crossref | GoogleScholarGoogle Scholar |
Turner, C. L., Seastedt, T. R., and Dyer, M. I. (1993). Maximization of above-ground grassland production: the role of defoliation frequency, intensity, and history. Ecological Applications 3, 175–186.
| Maximization of above-ground grassland production: the role of defoliation frequency, intensity, and history.Crossref | GoogleScholarGoogle Scholar |
Wan, H., Bai, Y., Schönbach, P., Gierus, M., and Taube, F. (2011). Effects of grazing management system on plant community structure and functioning in a semi-arid steppe: scaling from species to community. Plant and Soil 340, 215–226.
| Effects of grazing management system on plant community structure and functioning in a semi-arid steppe: scaling from species to community.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3MXitFGkt74%3D&md5=0c6cb8ae94f9a812170c690881cae9a8CAS |
Wang, D., and Ba, L. (2008). Ecology of meadow steppe in north-east China. The Rangeland Journal 30, 247–254.
| Ecology of meadow steppe in north-east China.Crossref | GoogleScholarGoogle Scholar |
Wang, J., and Cai, W. (1988). Studies on genesis, types and characteristics of the soils in the Xilin River Basin, Inner Mongolia. Research on Grassland Ecosystem 3, 23–83.
Wang, Y., and Zhou, G. (2004). Responses of temporal dynamics of above-ground net primary productivity of Leymus chinensis community to precipitation fluctuation in Inner Mongolia. Acta Ecologica Sinica 24, 1140–1145.
Wang, S., Niu, H., Cui, X., Jiang, S., Li, Y., Xiao, X., Wang, G., Huang, D., Qi, Q., and Yang, Z. (2005). Plant communities: ecosystem stability in Inner Mongolia. Nature 435, E6.
| Plant communities: ecosystem stability in Inner Mongolia.Crossref | GoogleScholarGoogle Scholar |
Wu, Z. (1980). ‘Vegetation of China.’ (Science Press: Beijing, China.) [In Chinese]
Xiao, X., Jiang, S., Wang, Y., Ojima, D. S., and Bonham, C. D. (1996). Temporal variation in above-ground biomass of Leymus chinensis steppe from species to community levels in the Xilin River Basin, Inner Mongolia, China. Vegetatio 123, 1–12.
| Temporal variation in above-ground biomass of Leymus chinensis steppe from species to community levels in the Xilin River Basin, Inner Mongolia, China.Crossref | GoogleScholarGoogle Scholar |
Zhang, H. Y. (1993). Discussion on the optimal cutting time of Leymus chinensis grasslands. Grassland of China 5, 64–66.
Zhong, Y. (1991). A study on the reasonable mowing system of natural mowing steppe in Bayinxile farm, Inner Mongolia. Acta Ecologica Sinica 11, 242–249.
Zhong, Y., and Piao, S. (1988). The analysis of the experimental results on mowing succession of Leymus chinensis steppe. Research on Grassland Ecosystems 2, 158–171.
Zhong, Y., Bao, Q., and Sun, W. (2012). ‘Experimental Research on Mowing Grassland Dynamics in Inner Mongolia.’ (Inner Mongolia University Press: Huhhot.) [In Chinese]