

Molecular evidence for the association of Cucumber mosaic virus causing mosaic and leaf deformation of Pisum sativum in Western Uttar Pradesh, India
A. A. Khan A , A. Rafiq A , S. Kumar B , D. Pratap B , D. K. Verma B and Q. A. Naqvi A CA Plant Virology Lab, Aligarh Muslim University (AMU), Aligarh 202 002, India.
B Molecular Virology Lab, National Botanical Research Institute, Lucknow 226 001, India.
C Corresponding author. Email: qmnaqvi@rediffmail.com
Australasian Plant Disease Notes 3(1) 42-43 https://doi.org/10.1071/DN08016
Submitted: 7 February 2008 Accepted: 30 March 2008 Published: 9 April 2008
Abstract
Cucumber mosaic virus associated with the mosaic, leaf deformation and stunting of Pisum sativum was identified as a member of subgroup IB on the basis of the capsid protein gene sequence.
Sweet pea (Pisum sativum L., family Leguminaceae) is a major consumable food crop in the tropical and subtropical regions of the world including India. Pea crop growing in the experimental plots, gardens and fields around Aligarh Muslim University, Aligarh (Western Uttar Pradesh) India, were found naturally infected, exhibiting mosaic, leaf deformation and stunting of the entire plant. The disease affected fruit yield and was prevalent. The incidence varied in different experimental fields from 22–37%. Infected plants were collected from three field locations in Aligarh and were maintained in an insect proof glasshouse.
Leaf dip preparations of the infected pea leaves revealed the presence of 28 nm isometric particles, typical of Cucumber mosaic virus (CMV). Symptomatic leaf tissue was ground in 0.1 M phosphate buffer (1 : 1, w/v pH 7.0) and squeezed through double layered muslin cloth. The filtrate was mechanically inoculated to several indicator species. Necrotic lesions developed on Gomphrena globosa, Chenopodium amaranticolor, C. murale and Spinacia oleracea within 7–10 days post inoculation (dpi). Systemic mosaic symptoms developed (20–25 dpi) on Nicotiana tabacum cv. White Burley, N. tabacum cv. Samsun-NN, N. glutinosa, N. benthamiana, N. rustica and Cucumis sativus. The pea isolate was vector-transmitted using healthy Myzus persicae and Aphis gossypii in a non-persistent manner to N. tabacum cv. White Burley, N. glutinosa, N. benthamiana and Cucumis sativus following the method of Noordam (1973). The presence of the virus in inoculated hosts was confirmed by enzyme linked immunosorbent assay (ELISA) (Verma et al. 2005) using CMV specific antiserum (PVAS242a, American Type Culture Collection, USA) and alkaline phosphatase linked secondary antibodies (DSMZ, Germany). The mean absorbance at 405 nm for negative and positive controls were 0.023 ± 0.003 and 0.438 ± 0.002, respectively, whereas in infected samples absorbance ranged between 0.346–0.433 ± 0.002.
For molecular identification of CMV, total RNA was isolated from three naturally infected pea plants each collected from three different location and healthy leaves of pea using RNAqueous (Ambion, USA). First strand cDNA (cDNA) was synthesised using total RNA as template, CMV-CP specific reverse primer 5′-GCATGGTACCTCAAACTGGGAGCAC-3′ and RTase (200 U, MBI Fermentas, USA) following the manufacturer’s procedure. Complementary DNA was amplified in GeneAmp PCR9700 system thermal cycler (Applied Biosystems, USA) in 50 µL reaction mixture containing 10 ng cDNA as template, 0.2 µM forward (5′-GCATTCTAGATGGACAAATCTG AATC-3′) and aforesaid reverse primer and 1.5 U Taq DNA polymerase (Invitrogen, USA). Following the initial 5 min denaturation, 30 cycles of amplification were performed with denaturation at 94°C for 1 min, annealing at 45°C for 1 min and elongation at 72°C for 1 min followed by a final elongation of 10 min at 72°C. Electrophoresis in 0.8% agarose gel at 5 V/cm revealed a ~650 bp product from each of the infected pea plants collected from the three different locations.
The amplified product from one location was gel purified (Au-Prep Sigma gel extraction kit) and cloned into pGEM-T easy vector system (Promega, USA). Randomly, 10 ampicillin positive clones were screened through restriction digestion and two clones were sequenced in an automated sequencer (ABI Prism 310). For sequencing SP6 and T7 primers were mixed with purified plasmid DNA and the reaction was carried out using Big Dye Terminator Sequencing V3 sequencing kit, Applied Biosystem, USA. The sequence data obtained was submitted to EMBL Database (Accession EU140547).
Maximum (93–94%) amino acid identity was obtained with CMV subgroup IB when analysed with BLAST in NCBI GenBank. Lower (89–90%) identity was with subgroup IA and 74–77% identity was with subgroup II (Table 1). Multiple nucleotide and deduced amino acid alignments with the available sequence were carried out using CLUSTAL-W program v. 1.82 (Higgins et al. 1994) and the aligned files were bootstrapped 100 times generating a neighbour-joining phylogenetic tree using Tree-Explorer. The pea isolate grouped with a clade of Indian CMV strains of subgroup IB. Our biological, serological and molecular studies confirmed the pea virus to be CMV of subgroup IB (Fig. 1).

|
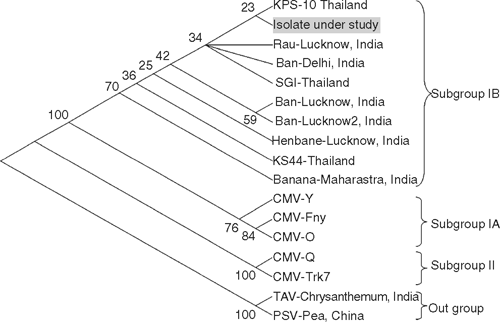
|
In India, pea has been reported to be infected by Pea common mosaic virus (Sreenivasan and Nariani 1966), Pea top necrosis virus (Roy and Gupta 1977), Pea necrotic mosaic virus (Sharma and Gupta 1978), Tomato spotted wilt virus and Bean yellow mosaic virus (Rishi and Hampton 1987). A literature survey has also revealed the occurrence of CMV on pea causing mosaic disease, which has been characterised only at the biological and serological level (Rao et al. 1995). To the best of our knowledge this is the first report characterising the CMV pea isolate at the molecular level.
Acknowledgement
The authors are thankful to the Chairman, Department of Botany for providing A.A. Khan with a place to work, the Department of Science and Technology, New Delhi for providing financial assistance under SERC Fast Track Young Scientist Scheme.
Higgins D,
Thompson J,
Gibson T,
Thompson JD,
Higgins DG, Gibson TJ
(1994) CLUSTAL W: improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matri choice. Nucleic Acids Research 22, 4673–4680.
| Crossref | GoogleScholarGoogle Scholar | PubMed |

Rao GP,
Singh M,
Shukla K, Gupta SN
(1995) A new strain of Cucumber mosaic virus causing mosaic disease of pea in India. Indian Journal of Virology 2, 43–48.
Rishi N, Hampton RO
(1987) Update Pisum germplasm and pea seed-borne mosaic virus in North India. Pisum Newsletter 19, 55–58.
Roy AN, Gupta MN
(1977) Top necrosis virus disease of pea: a new record. Current Science 46, 464.
Sharma RK, Gupta MN
(1978) Studies on a necrosis mosaic disease of pea: a new report from India. Acta Botanica Indica 6, 60–67.
Sreenivasan TN, Nariani TK
(1966) Studies on a mosaic disease on pea. Indian Phytopathology 19, 189–193.
Verma N,
Singh L,
Singh AK,
Kulshreshtha S,
Raikhy G,
Hallan V,
Ram R, Zaidi AA
(2005) Ornithogalum: a new host of Cucumber mosaic virus (CMV) from India. Plant Pathology 54, 256.
| Crossref | GoogleScholarGoogle Scholar |