

Identification of Leptosphaeria biglobosa ‘canadensis’ on Brassica juncea stubble from northern New South Wales, Australia
Angela P. Van de Wouw A E , Vicki L. Thomas B , Anton J. Cozijnsen A , Stephen J. Marcroft C , Phillip A. Salisbury B D and Barbara J. Howlett AA School of Botany, The University of Melbourne, Vic. 3010, Australia.
B Faculty of Land and Food Resources, The University of Melbourne, Vic. 3010, Australia.
C Marcroft Grains Pathology P/L, Grains Innovation Park, Horsham, Vic. 3400, Australia.
D Department of Primary Industries, VABC, Bundoora, Vic. 3083, Australia.
E Corresponding author. Email: apvdw2@unimelb.edu.au
Australasian Plant Disease Notes 3(1) 124-128 https://doi.org/10.1071/DN08049
Submitted: 13 June 2008 Accepted: 3 September 2008 Published: 18 September 2008
Abstract
Leptosphaeria biglobosa ‘canadensis’ is reported for the first time in Australia. All 88 Leptosphaeria isolates cultured from Brassica juncea stubble from northern NSW were L. biglobosa ‘canadensis’ whilst all 55 isolates cultured from Victorian stubble of the same B. juncea lines were L. maculans. Both L. biglobosa ‘canadensis’ and L. maculans formed similar sized lesions on B. juncea cotyledons after 14 days. However, L. biglobosa ‘canadensis’ isolates colonised stems less effectively than L. maculans and consequently caused less crown cankering.
The two Dothideomycetes, Leptosphaeria maculans and L. biglobosa, comprise a species complex associated with disease of crucifers including Brassica napus (canola, oilseed rape). Through the use of molecular techniques, L. biglobosa has been divided into six subclades, namely ‘canadensis’, ‘occiaustralensis’, ‘brassicae’, ‘australiensis’, ‘erysimii’ and ‘thlaspii’. Leptosphaeria maculans has been divided into two subclades, ‘brassicae’ and ‘lepidii’ (Mendes-Pereira et al. 2003; Voigt et al. 2005; Vincenot et al. 2008). In this paper we refer to L. maculans ‘brassicae’ as L. maculans.
During infection of B. napus, L. maculans produces grey/green large leaf lesions, followed by crown stem cankers. This disease, known as blackleg or Phoma stem canker, results in significant yield losses, particularly in Australia (Howlett 2004; Fitt et al. 2006a). In contrast, none of the subclades of L. biglobosa are reported to cause crown stem cankers. Leptosphaeria biglobosa ‘brassicae’ is the most common subclade. It is found in most canola growing areas with the exception of central Canada and Australia (Fitt et al. 2006a). These isolates cause small dark leaf lesions followed by pale brown stem lesions with a dark margin on the upper stem (known as Phoma or upper stem lesions) (West et al. 2002). In Australia, L. biglobosa ‘australiensis’ (cultured from B. napus) and more recently, L. biglobosa ‘occiaustralensis’ (cultured from B. napus and Raphanus raphanistrum (wild radish)) have been described in eastern and western Australia, respectively (Plummer et al. 1994; Vincenot et al. 2008).
Blackleg is primarily controlled by breeding varieties with resistance to L. maculans (Delourme et al. 2006). Brassica juncea (Indian mustard) is generally more resistant to L. maculans than B. napus (Purwantara et al. 1998; Li et al. 2008). Additionally, B. juncea is more drought-tolerant and can be grown in regions with short, warm to hot growing seasons in which soil water supply is unreliable (Oram et al. 2005). In Canada, B. juncea has been grown as a condiment mustard crop for 20 years, specifically in the hotter, drier areas of western Canada. In 2002, B. juncea varieties were released that produced canola quality oils (Burton et al. 2004). Over the past 20 years in Australia, small acreages in regions including northern New South Wales have produced condiment mustard. Brassica juncea varieties that produce canola-quality oils were commercially released in 2007 (Burton et al. 2008).
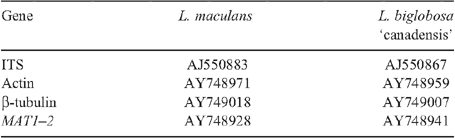
Since blackleg disease is a threat to the Australian oilseed industry, fungal populations are monitored for virulence changes in relation to both B. juncea and B. napus varieties released. During 2006 and 2007, 152 Leptosphaeria ascospore isolates were cultured from B. juncea stubble collected from trial sites in Victoria (Horsham and Beulah) and northern NSW (Tamworth, Burren Junction and Rowena). Fifty-five of these isolates were subjected to Polymerase Chain Reaction analysis and the resultant DNA fragments of four regions of the genome were sequenced. These genomic regions were the internal transcribed spacer (ITS) of rDNA (amplified with primers 5′-CCGTTGGTGAACCAGCGGAGGGATC-3′ and 5′-TCCGCTTATTGATATGCTTAAG-3′, Mendes-Pereira et al. 2003), actin (primers 5′-GAGCAGGAGATCCAGACTGC-3′ and 5′-TTCGAGATCCACATCTGCTG-3′), β-tubulin (primers 5′-GTCGAGAACTCCGACGAGAC-3′ and 5′-ATCTGGTCCTCGACCTCCTT-3′) and the mating type allele, MAT1–2 (primers 5′-GATGCCATGCACAAGAAGCTC-3′ and 5′-GCTTGGCCTTGCGCGACTGGC-3′). These latter three sets of primers were designed by aligning the sequences of L. biglobosa and L. maculans isolates (Voigt et al. 2005; Vincenot et al. 2008) and choosing a nucleotide region common to all isolates.
Sequences of the ITS, actin, β-tubulin and MAT1–2 from isolates cultured from B. juncea stubble were compared with previously published sequences from L. maculans and L. biglobosa isolates (Table 1). On this basis, all 25 isolates from Victoria were identified as L. maculans (displaying 100% sequence similarity with previously published L. maculans sequences for each genomic region analysed) whilst all 30 northern NSW isolates were L. biglobosa ‘canadensis’ (displaying 100% sequence similarity with previously published L. biglobosa ‘canadensis’ sequences for each genomic region analysed). This is the first report of L. biglobosa ‘canadensis’ in Australia. Representative sequences from L. maculans and L. biglobosa ‘canadensis’ isolates have been deposited into GenBank (Accession numbers FJ172238, FJ172239, FJ172240, FJ172241, FJ172242, FJ172243 and FJ172244).
|
As well as by DNA sequencing, which is expensive and time-consuming, L. maculans and L. biglobosa ‘canadensis’ isolates can be distinguished by the size of the ITS fragments separated on a 2% agarose gel (580 and 555 bp, respectively) (Mendes-Pereira et al. 2003). On this basis, an additional 39 Victorian isolates cultured from B. juncea were classified as L. maculans and 58 isolates from northern NSW were classified as L. biglobosa ‘canadensis’. Two L. biglobosa ‘canadensis’ isolates have been deposited into the New South Wales Plant Pathology Herbarium (DAR79245 and DAR79246).
Ascospore isolates (250) were also cultured from B. napus stubble collected from field trials in Victoria (Horsham, Geelong and Lake Bolac), southern NSW (Wagga Wagga and Illabo) and South Australia (Moyhall, Bordertown and Yeelena) between 2005 and 2007. All of these isolates were identified as L. maculans. Surprisingly, no L. biglobosa or L. maculans isolates were cultured from B. napus stubble collected from the same sites as the B. juncea stubble in northern NSW. This stubble contained isolates belonging to the genera Alternaria, Pleospora and Embellisia.
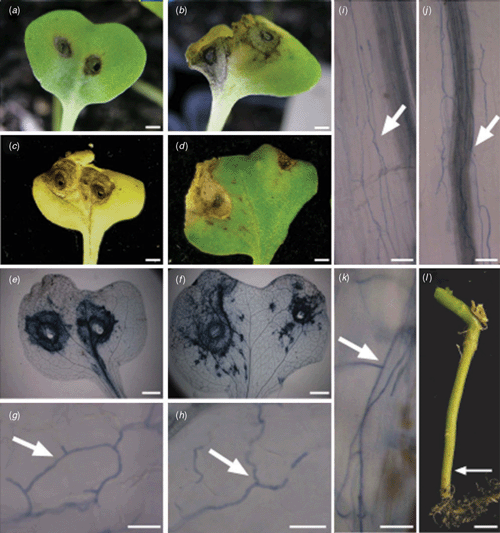
The infection pathway of the L. maculans and L. biglobosa ‘canadensis’ isolates cultured from B. juncea stubble was determined on three B. juncea advanced breeding lines, one B. juncea variety and three B. napus varieties. The B. juncea lines and variety were JC05002, JC05006, JC05007 and Dune, respectively. The resistance genes in these B. juncea lines and variety are uncharacterised. The B. napus varieties were Q2 (susceptible to most Australian isolates), Beacon (polygenic resistance) and Surpass 400 (B. rapa ssp. ‘sylvestris’-derived resistance) (Crouch et al. 1994). Cotyledons were wounded and inoculated with conidia (105) as described by Purwantara et al. (1998). Three L. maculans (06VTJ085, 06VTJ095 and 06VTJ112) and two L. biglobosa ‘canadensis’ (06VTJ140 and 06VTJ154) isolates were individually tested. Four cotyledons of each variety, inoculated with either L. maculans or L. biglobosa ‘canadensis’ were examined. After 8 days, small lesions (1–2 mm in diameter) formed on cotyledons of all plants inoculated with L. maculans isolates, whilst large necrotic lesions (5–7 mm in diameter) were observed on cotyledons inoculated with L. biglobosa ‘canadensis’ isolates (Fig. 1a, b). After 14 days, lesions of similar size were observed on all lines and varieties inoculated with either L. maculans or L. biglobosa ‘canadensis’ (Fig. 1c, d). Tissue was stained with lactophenol trypan-blue and mounted in glycerol (Keogh et al. 1980). The lesions caused by L. maculans isolates radiated symmetrically from the inoculation site whilst lesions caused by L. biglobosa ‘canadensis’ were spread asymmetrically across the cotyledon (Fig. 1e, f). Despite the differences in lesion appearance, hyphae of all isolates grew intercellularly within the cotyledons (Fig. 1 g, h) and petioles (Fig. 1i, j). A similar infection pattern was seen in the B. napus varieties inoculated with these isolates (data not shown).
|
Ten weeks post-inoculation, two stems of each variety inoculated with either L. maculans or L. biglobosa ‘canadensis’ were stained with trypan blue. In both B. juncea and B. napus branched hyphae of L. maculans were growing intercellularly at the base of the stem (Fig. 1k, l). In contrast, no hyphae of L. biglobosa ‘canadensis’ were observed (data not shown). This is the first report of disease progression for L. biglobosa ‘canadensis’ on both B. juncea and B. napus.
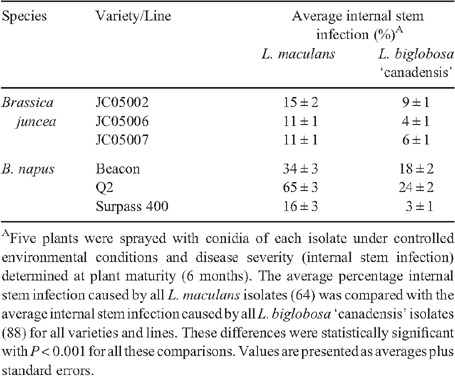
To determine whether these isolates could form crown stem cankers after inoculation by a non-wounding method, plants were sprayed with conidia of individual L. maculans (64) and L. biglobosa ‘canadensis’ (88) isolates. Five plants of each variety or line were sprayed at the first leaf stage with conidia (107) suspended in 0.05% Tween 20. Internal infection was determined at maturity (6 months) by cross-sectioning stems at the crown and visually assessing the percentage of blackening (Marcroft et al. 2004). The internal stem infection data were averaged for all L. maculans isolates and for all L. biglobosa ‘canadensis’ isolates for each Brassica variety or line. Differences in averages were statistically analysed using one-way ANOVA. Leptosphaeria biglobosa ‘canadensis’ isolates caused significantly less internal stem infection at the crown of the stem than L. maculans isolates on all B. juncea lines and B. napus varieties tested (P < 0.001) (Table 2). As expected, less internal stem infection was observed in B. juncea lines compared with B. napus varieties when infected with L. maculans or L. biglobosa ‘canadensis’ isolates. Eighteen of 64 L. maculans isolates caused more than 30% internal stem infection in B. juncea lines, compared with only 7 of 88 L. biglobosa ‘canadensis’ isolates. For B. napus varieties, 49 of 64 L. maculans isolates caused more than 50% internal stem infection compared with 13 of 88 L. biglobosa ‘canadensis’ isolates.
|
Similar to our findings with L. biglobosa ‘canadensis’, L. biglobosa ‘brassicae’ has been shown to develop cotyledonary lesions more quickly on B. napus than L. maculans (Fitt et al. 2006b). In the present study, despite hyphae of L. biglobosa ‘canadensis’ and L. maculans growing intercellularly within the cotyledon and petiole, no L. biglobosa ‘canadensis’ hyphae were observed in the eight stems of infected plants examined after 10 weeks. However, internal stem infection was evident at plant maturity, albeit significantly less than in plants inoculated with L. maculans. This reduced efficiency of L. biglobosa ‘canadensis’ to colonise the stem could be due to several factors including the inability to produce the toxin, sirodesmin PL, which significantly contributes to the virulence of L. maculans in B. napus stems (Elliott et al. 2007).
Previously in Australia two L. biglobosa ‘australiensis’ and 19 L. biglobosa ‘occiaustralensis’ isolates were identified amongst populations of L. maculans isolates (Plummer et al. 1994; Vincenot et al. 2008). In the present study, all 88 isolates identified on B. juncea stubble from northern NSW were L. biglobosa ‘canadensis’. It is curious that these isolates were found in high frequency in this region and not in any other Australian region tested. One possible explanation is that northern NSW is a summer dominant rainfall region, which may influence pseudothecia development and affect timing of ascospore release resulting in more favourable conditions for L. biglobosa ‘canadensis’. The epidemiology of this fungus is unknown.
Due to its ability to undergo sexual recombination, populations of L. maculans can rapidly adapt to selection pressures including exposure to major gene resistance (Sprague et al. 2006). All L. biglobosa ‘canadensis’ isolates cultured in this study were from ascospores, demonstrating that L. biglobosa ‘canadensis’ is undergoing sexual recombination in the field. Whether populations of L. biglobosa ‘canadensis’ will adapt to selection pressures and develop the ability to invade stems of B. juncea and B. napus is unknown.
Acknowledgements
We thank the Grains Research and Development Corporation, Australia for funding. We also thank Paul Parker and Rod Bambach from the New South Wales Department of Primary Industries for the collection of stubble.
Burton WA,
Ripley VL,
Potts DA, Salisbury PA
(2004) Assessment of genetic diversity in selected breeding lines and cultivars of canola-quality Brassica juncea and their implications for canola breeding. Euphytica 136, 181–192.
| Crossref | GoogleScholarGoogle Scholar |

Burton WA,
Flood RF,
Norton RM,
Field B,
Potts DA,
Robertson MJ, Salisbury PA
(2008) Identification of variability in phenological responses in canola-quality Brassica juncea for utilisation in Australian breeding programs. Australian Journal of Agricultural Research 59, 874–881.
| Crossref |
Crouch JH,
Lewis BG, Mithen RF
(1994) The effect of A genome substitution on the resistance of Brassica napus to infection by Leptosphaeria maculans. Plant Breeding 112, 265–278.
| Crossref | GoogleScholarGoogle Scholar |
Delourme R,
Chevre AM,
Brun H,
Rouxel T,
Balesdent MH,
Dias JS,
Salisbury P,
Renard M, Rimmer SR
(2006) Major gene and polygenic resistance to Leptosphaeria maculans in oilseed rape (Brassica napus). European Journal of Plant Pathology 114, 41–52.
| Crossref | GoogleScholarGoogle Scholar |
Elliott CE,
Gardiner DM,
Thomas G,
Cozijnsen AJ,
Van de Wouw A, Howlett BJ
(2007) Production of the toxin sirodesmin PL by Leptosphaeria maculans during infection of Brassica napus. Molecular Plant Pathology 8, 791–802.
| Crossref | GoogleScholarGoogle Scholar |
Fitt BDL,
Brun H,
Barbetti MJ, Rimmer SR
(2006a) World-wide importance of phoma stem canker (Leptosphaeria maculans and L. biglobosa) on oilseed rape (Brassica napus). European Journal of Plant Pathology 114, 3–15.
| Crossref | GoogleScholarGoogle Scholar |
Fitt BDL,
Huang YJ,
van den Bosch F, West J
(2006b) Coexistence of related pathogen species on arable crops in space and time. Annual Review of Phytopathology 44, 163–182.
| Crossref | GoogleScholarGoogle Scholar | PubMed |
Howlett BJ
(2004) Current knowledge of the interaction between Brassica napus and Leptosphaeria maculans. Canadian Journal of Plant Pathology 24, 245–252.
Keogh RC,
Deverall BJ, McLeod S
(1980) Comparison of histological and physiological-responses to Phakopsora pachyrhizi in resistant and susceptible soybean. Transactions of the British Mycological Society 74, 329–333.
Li CX,
Wratten N,
Salisbury PA,
Burton WA, Potter TD ,
et al
.
(2008) Response of Brassica napus and B. juncea germplasm from Australia, China and India to Australian populations of Leptosphaeria maculans. Australasian Plant Pathology 37, 162–170.
| Crossref | GoogleScholarGoogle Scholar |
Marcroft SJ,
Sprague SJ,
Pymer SJ,
Salisbury PA, Howlett BJ
(2004) Crop isolation, not extended rotation length, reduces blackleg (Leptosphaeria maculans) severity of canola (Brassica napus) in south-eastern Australia. Australian Journal of Experimental Agriculture 44, 601–606.
| Crossref | GoogleScholarGoogle Scholar |
Mendes-Pereira E,
Balesdent MH,
Brun H, Rouxel T
(2003) Molecular phylogeny of the Leptosphaeria maculans-L. biglobosa species complex. Mycological Research 107, 1287–1304.
| Crossref | GoogleScholarGoogle Scholar | PubMed |
Oram RN,
Kirk JTO,
Veness PE,
Hurlstone CJ,
Edlington JP, Halsall DM
(2005) Breeding Indian mustard (Brassica juncea) for cold-pressed, edible oil production – a review. Australian Journal of Agricultural Research 56, 581–596.
| Crossref | GoogleScholarGoogle Scholar |
Plummer KM,
Dunse K, Howlett BJ
(1994) Nonaggressive strains of the blackleg fungus, Leptosphaeria maculans, are present in Australia and can be distinguished from aggressive strains by molecular analysis. Australian Journal of Botany 42, 1–8.
| Crossref | GoogleScholarGoogle Scholar |
Purwantara A,
Burton WA,
Salisbury PA, Howlett BJ
(1998) Reaction of Brassica juncea (Indian mustard) lines to Australian isolates of Leptosphaeria maculans under glasshouse and field conditions. European Journal of Plant Pathology 104, 895–902.
| Crossref | GoogleScholarGoogle Scholar |
Sprague SJ,
Hayden HL,
Brun H,
Marcroft SJ,
Pinochet X,
Rouxel T,
Balesdent MH, Howlett BJ
(2006) Major gene resistance in Brassica napus (oilseed rape) is overcome by changes in virulence of populations of Leptosphaeria maculans in France and Australia. European Journal of Plant Pathology 114, 33–40.
| Crossref | GoogleScholarGoogle Scholar |
Vincenot L,
Balesdent MH,
Li H,
Barbetti MJ,
Sivasithamparam K,
Gout L, Rouxel T
(2008) Occurrence of a new subclade of Leptosphaeria biglobosa in Western Australia. Phytopathology 98, 321–329.
| Crossref | GoogleScholarGoogle Scholar |
Voigt K,
Cozijnsen AJ,
Kroymann J,
Poggeler S, Howlett BJ
(2005) Phylogenetic relationships between members of the crucifer pathogenic Leptosphaeria maculans species complex as shown by mating type (MAT1–2), actin, and β-tubulin sequences. Molecular Phylogenetics and Evolution 37, 541–557.
| Crossref | GoogleScholarGoogle Scholar | PubMed |
West JS,
Balesdent MH,
Rouxel T,
Narcy JP,
Huang YJ,
Roux J,
Steed JM,
Fitt BDL, Schmit J
(2002) Colonization of winter oilseed rape tissues by A/Tox+ and B/Tox0 Leptosphaeria maculans (phoma stem canker) in France and England. Plant Pathology 51, 311–321.
| Crossref | GoogleScholarGoogle Scholar |