

New handbook for standardised measurement of plant functional traits worldwide
N. Pérez-Harguindeguy A Y , S. Díaz A , E. Garnier B , S. Lavorel C , H. Poorter D , P. Jaureguiberry A , M. S. Bret-Harte E , W. K. Cornwell F , J. M. Craine G , D. E. Gurvich A , C. Urcelay A , E. J. Veneklaas H , P. B. Reich I , L. Poorter J , I. J. Wright K , P. Ray L , L. Enrico A , J. G. Pausas M , A. C. de Vos F , N. Buchmann N , G. Funes A , F. Quétier A C , J. G. Hodgson O , K. Thompson P , H. D. Morgan Q , H. ter Steege R , M. G. A. van der Heijden S , L. Sack T , B. Blonder U , P. Poschlod V , M. V. Vaieretti A , G. Conti A , A. C. Staver W , S. Aquino X and J. H. C. Cornelissen FA Instituto Multidisciplinario de Biología Vegetal (CONICET-UNC) and FCEFyN, Universidad Nacional de Córdoba, CC 495, 5000 Córdoba, Argentina.
B CNRS, Centre d’Ecologie Fonctionnelle et Evolutive (UMR 5175), 1919, Route de Mende, 34293 Montpellier Cedex 5, France.
C Laboratoire d’Ecologie Alpine, UMR 5553 du CNRS, Université Joseph Fourier, BP 53, 38041 Grenoble Cedex 9, France.
D Plant Sciences (IBG2), Forschungszentrum Jülich, D-52425 Jülich, Germany.
E Institute of Arctic Biology, 311 Irving I, University of Alaska Fairbanks, Fairbanks, AK 99775-7000, USA.
F Systems Ecology, Faculty of Earth and Life Sciences, Department of Ecological Science, VU University, De Boelelaan 1085, 1081 HV Amsterdam, The Netherlands.
G Division of Biology, Kansas State University, Manhtattan, KS 66506, USA.
H Faculty of Natural and Agricultural Sciences, School of Plant Biology, The University of Western Australia, 35 Stirling Highway, Crawley, WA 6009, Australia.
I Department of Forest Resources, University of Minnesota, 1530 N Cleveland Avenue, St Paul, MN 55108, USA and Hawkesbury Institute for the Environment, University of Western Sydney, Locked Bag 1797, Penrith, NSW 2751, Australia.
J Centre for Ecosystems, Forest Ecology and Forest Management Group, Wageningen University, PO Box 47, 6700 AA Wageningen, The Netherlands.
K Department of Biological Sciences, Macquarie University, Sydney, NSW 2109, Australia.
L Department of Biological Sciences, Stanford University, Stanford, CA, USA.
M Centro de Investigaciones sobre Desertificación (CIDE-CSIC), IVIA Campus, Carretera Nàquera km 4.5, 46113 Montcada, Valencia, Spain.
N Institute of Agricultural Sciences, ETH Zurich, Universitätstrasse 2, LFW C56, CH-8092 Zürich, Switzerland.
O Peak Science and Environment, Station House, Leadmill, Hathersage, Hope Valley S32 1BA, UK.
P Department of Animal and Plant Sciences, The University of Sheffield, Sheffield S10 2TN, UK.
Q NSW Department of Primary Industries, Forest Resources Research Beecroft, NSW 2119, Australia.
R Naturalis Biodiversity Center, Leiden, and Institute of Environmental Biology, Ecology and Biodiversity Group, Utrecht University, Utrecht, The Netherlands.
S Ecological Farming Systems, Agroscope Reckenholz Tänikon, Research Station ART, Reckenholzstrasse 191, 8046 Zurich, Switzerland and Plant-Microbe Interactions, Institute of Environmental Biology, Faculty of Science, Utrecht University, Utrecht, The Netherlands.
T Department of Ecology and Evolutionary Biology, University of California, Los Angeles, 621 Charles E. Young Drive South, Los Angeles, CA 90095-1606, USA.
U Department of Ecology and Evolutionary Biology, University of Arizona, Tucson, AZ, USA.
V Institute of Botany, Faculty of Biology and Preclinical Medicine, University of Regensburg, D-93040, Regensburg, Germany.
W Department of Ecology and Evolutionary Biology, Princeton University, Princeton, NJ 08544, USA.
X Centro Agronómico Tropical de Investigación y Enseñanza, CATIE 7170, Cartago, Turrialba 30501, Costa Rica.
Y Corresponding author. Email: nperez@com.uncor.edu
Australian Journal of Botany 61(3) 167-234 https://doi.org/10.1071/BT12225
Submitted: 23 November 2011 Accepted: 29 January 2013 Published: 26 April 2013
Abstract
Plant functional traits are the features (morphological, physiological, phenological) that represent ecological strategies and determine how plants respond to environmental factors, affect other trophic levels and influence ecosystem properties. Variation in plant functional traits, and trait syndromes, has proven useful for tackling many important ecological questions at a range of scales, giving rise to a demand for standardised ways to measure ecologically meaningful plant traits. This line of research has been among the most fruitful avenues for understanding ecological and evolutionary patterns and processes. It also has the potential both to build a predictive set of local, regional and global relationships between plants and environment and to quantify a wide range of natural and human-driven processes, including changes in biodiversity, the impacts of species invasions, alterations in biogeochemical processes and vegetation–atmosphere interactions. The importance of these topics dictates the urgent need for more and better data, and increases the value of standardised protocols for quantifying trait variation of different species, in particular for traits with power to predict plant- and ecosystem-level processes, and for traits that can be measured relatively easily. Updated and expanded from the widely used previous version, this handbook retains the focus on clearly presented, widely applicable, step-by-step recipes, with a minimum of text on theory, and not only includes updated methods for the traits previously covered, but also introduces many new protocols for further traits. This new handbook has a better balance between whole-plant traits, leaf traits, root and stem traits and regenerative traits, and puts particular emphasis on traits important for predicting species’ effects on key ecosystem properties. We hope this new handbook becomes a standard companion in local and global efforts to learn about the responses and impacts of different plant species with respect to environmental changes in the present, past and future.
Additional keywords: biodiversity, ecophysiology, ecosystem dynamics, ecosystem functions, environmental change, plant morphology.
Contents
-
Introduction and discussion...................................169
-
1 Selection of species and individuals...................................170
-
1.1 Selection of species...................................170
-
1.2 Selection of individuals within a species...................................171
-
1.3 Replicate measurements...................................172
-
2 Whole-plant traits...................................172
-
2.1 Life history and maximum plant lifespan...................................172
-
2.2 Life form...................................173
-
2.3 Growth form...................................173
-
2.4 Plant height...................................175
-
2.5 Clonality, bud banks and below-ground storage organs...................................176
-
2.6 Spinescence...................................177
-
2.7 Branching architecture...................................178
-
2.8 Leaf area : sapwood area ratio...................................178
-
2.9 Root-mass fraction...................................179
-
2.10 Salt resistance...................................179
-
2.11 Relative growth rate and its components...................................181
-
2.12 Plant flammability...................................182
-
2.13 Water-flux traits...................................184
-
3 Leaf traits...................................186
-
3.1 Specific leaf area...................................186
-
3.2 Area of a leaf...................................189
-
3.3 Leaf dry-matter content...................................190
-
3.4 Leaf thickness...................................190
-
3.5 pH of green leaves or leaf litter...................................191
-
3.6 Leaf nitrogen (N) concentration and leaf phosphorous (P) concentration...................................192
-
3.7 Physical strength of leaves...................................193
-
3.8 Leaf lifespan and duration of green foliage...................................195
-
3.9 Vein density...................................197
-
3.10 Light-saturated photosynthetic rate...................................198
-
3.11 Leaf dark respiration...................................198
-
3.12 Photosynthetic pathway...................................199
-
3.13 C-isotope composition as a measure of intrinsic water-use efficiency...................................200
-
3.14 Electrolyte leakage as an indicator of frost sensitivity...................................201
-
3.15 Leaf water potential as a measure of water status...................................202
-
3.16 Leaf palatability as indicated by preference by model herbivores...................................203
-
3.17 Litter decomposability...................................205
-
4 Stem traits...................................207
-
4.1 Stem-specific density...................................207
-
4.2 Twig dry-matter content and twig drying time...................................209
-
4.3 Bark thickness (and bark quality)...................................209
-
4.4 Xylem conductivity...................................210
-
4.5 Vulnerability to embolism...................................211
-
5 Below-ground traits...................................212
-
5.1 Specific root length...................................212
-
5.2 Root-system morphology...................................214
-
5.3 Nutrient-uptake strategy...................................214
-
6 Regenerative traits...................................215
-
6.1 Dispersal syndrome...................................215
-
6.2 Dispersule size and shape...................................216
-
6.3 Dispersal potential...................................216
-
6.4 Seed mass...................................217
-
6.5 Seedling functional morphology...................................218
-
6.6 Resprouting capacity after major disturbance...................................218
-
Acknowledgements...................................220
-
References...................................220
Introduction and discussion
Environmental changes such as those on climate, atmospheric composition, land use and biotic exchanges are triggering unprecedented ecosystem changes. The need to understand and predict them has given new stimulus to a long tradition of study of the plant features (traits) that reflect species ecological strategies and determine how plants respond to environmental factors, affect other trophic levels and influence ecosystem properties (Kattge et al. 2011). There is mounting evidence that variation in plant traits, and trait syndromes (i.e. recurrent associations of plant traits), within and among species, is associated with many important ecological processes at a range of scales. This has resulted in strong demand for standardised ways to measure ecologically meaningful plant traits. The predecessor of the present handbook (Cornelissen et al. 2003) was written to address that need, by providing standardised, easily implemented trait-measurement recipes for researchers worldwide. This updated version is an extension of that global collective initiative, with an even broader scope.
The identification of general plant trait trade-offs associated with strategies and trait syndromes across floras, taxa and ecosystems has been a long-standing focus in plant ecology, and has attracted increasing interest in recent decades (e.g. Chapin et al. 1993; Grime et al. 1997; Reich et al. 1997; Cornelissen et al. 1999; Aerts and Chapin 1999; Westoby et al. 2002; Díaz et al. 2004; Wright et al. 2004; Cornwell et al. 2008; Baraloto et al. 2010a; Freschet et al. 2010; Ordoñez et al. 2010; Kattge et al. 2011). Ample evidence indicates that plant traits and trait syndromes significantly affect ecosystem processes and services (for overviews, see Lavorel and Garnier 2002; Díaz et al. 2007; Chapin et al. 2008; De Bello et al. 2010; Cardinale et al. 2012). As a consequence, trait-based approaches are currently also gaining momentum in the fields of agronomy and forestry (e.g. Brussaard et al. 2010; Garnier and Navas 2012), conservation (e.g. Mace et al. 2010), archaeobotany (e.g. Jones et al. 2010), and at the interface between the evolution and ecology in communities and ecosystems (e.g. Edwards et al. 2007; Cavender-Bares et al. 2009; Faith et al. 2010; Srivastava et al. 2012).
The quantification of vegetation changes in the face of modifications in climate at the global scale has been significantly improved with the use of dynamic global vegetation models (DGVMs) (Cramer et al. 2001; Arneth et al. 2010). However, current-generation DGVMs do not yet incorporate continuous variation in plant traits among plant species or types (Cornwell et al. 2009). Next-generation models could benefit from the incorporation of functional traits and syndromes that are simple and general enough to be assessed at the regional and global scales, and yet informative enough to relate to biogeochemical dynamics, dispersal and large-scale disturbance (Ollinger et al. 2008; Stich et al. 2008; Doherty et al. 2010; Harrison et al. 2010; Ma et al. 2011).
As a consequence of this surge of theoretical and practical interest, there has been a rapid expansion of large regional and global trait databases (e.g. Díaz et al. 2004; Wright et al. 2004; Kleyer et al. 2008; Cornwell et al. 2008; Chave et al. 2009; Paula et al. 2009; Baraloto et al. 2010a; Zanne et al. 2010; Fortunel et al. 2012; Patiño et al. 2012). The TRY Initiative (Kattge et al. 2011; see Box 1) is compiling a communal worldwide database of plant traits, an unprecedented step in improving the capacity of the scientific community to access and utilise plant-trait information. In this context, standardisation of protocols applicable under a wide range of situations and geographical contexts becomes even more important.
| Box 1. Useful links for plant functional-trait workers |
| To find on-line protocols and updates related to this handbook: Nucleo Diversus/Tools (http://www.nucleodiversus.org). |
| To submit corrections, additions and comments to improve this handbook: traitshandbook@gmail.com. |
| Various complementary protocols for specific plant (eco-)physiological as well as environmental measurements not covered in this handbook can be accessed through the fellow project: Prometheus Wiki (Sack et al. 2010; http://prometheuswiki.publish.csiro.au/tiki-index.php). |
| To share plant functional-trait data with other researchers (both as a provider and as a recipient): TRY Worldwide Initiative and Database (Kattge et al. 2011; www.try-db.org). |
| To calculate functional diversity metrics and indices with your trait data: FDiversity Free Software Package (Casanoves et al. 2011; www.fdiversity.nucleodiversus.org). |
In this manual, we consider plant functional traits to be any morphological, physiological or phenological feature, measurable for individual plants, at the cell to the whole-organism level, which potentially affects its fitness (cf. McGill et al. 2006; Lavorel et al. 2007; Violle et al. 2007) or its environment (Lavorel and Garnier 2002). As proposed by Lavorel et al. (2007), we will call the particular value or modality taken by the trait at any place and time an ‘attribute’. Functional traits addressed in the present handbook range from simple indicators of plant function (e.g. leaf nutrient concentrations as an indicator of both potential rates of metabolism and of quality as food for herbivores) to plant functions themselves (e.g. palatability, decomposability, capacity to resprout after a fire), always measured at the species level. The traits contained in the handbook represent a set of functional traits of vascular plants that (1) can together represent key plant responses to the environment as well as key plant effects on ecosystem processes and services at various scales from local plots to landscapes to biomes, (2) can help answer questions of ecological and evolutionary theory as well as practical ones related to nature conservation and land management (see Box 2 for a Discussion) and (3) are in most cases candidates for relatively easy, inexpensive and standardised measurement in many biomes and regions.
| Box 2. Why measure plant traits and which traits to measure? |
| Plant functional traits give better insight into the constraints and opportunities faced by plants in different habitats than does taxonomic identity alone (Southwood 1977; Grime 1979). They also provide understanding of how functional diversity in the broad sense underpins ecosystem processes and the benefits that people derive from them (Chapin et al. 2000; Díaz et al. 2007), and offer the possibility of comparing distant ecosystems with very little taxonomic overlap (Reich et al. 1997; Díaz et al. 2004; Cornwell et al. 2008). The plant-trait approach often provides unique mechanistic insights into several theoretical and practical questions, although it is not necessarily less laborious or less expensive than other methods. |
| Which traits to measure to answer which questions? |
No methods handbook can answer the question of what are the best traits to measure, because this strongly depends on the questions at hand, the ecological characteristics and scale of the study area, and on practical circumstances. For instance, there is not much point in comparing multiple species for succulence within wet environments or for flammability within areas that burn only very rarely, whereas such data might be useful as a reference in larger-scale studies. In addition, rather than setting limits to researchers’ curiosity, this trait handbook aims at inspiring others to come up with and measure traits not covered here, including ‘new’ traits, to help answer exciting novel questions. Some examples of additional interesting traits not covered here are in the introductory text of Cornelissen et al. (2003). The first and foremost criterion in deciding what traits to aim for is the process of interest. Is the intended study about fundamental plant or organ design in response to environmental variation in the present or about the evolution that gave rise to today’s spectrum of designs? Is it about plant growth, reproduction or dispersal over the landscape? Does it involve plant survival in response to resources or disturbance? Is the main question about response to or effects on water, soil nutrient or fire regimes? Is it about vegetation feedbacks to atmosphere and climate? Does it involve the juvenile stage, the persistence of adults? Does it involve pollinators, dispersers or herbivores? Does the target process occur above or below ground? Is the focus on coarse differences across or among regions or continents or on subtle differences among individuals of two slightly different local populations? Are specific ecosystem services to people assessed or predicted? All these and further types of questions will have a direct impact on the selection of traits. |
Although there is no limit to the number of relevant traits in different research contexts, a small number of traits have been considered relevant almost universally, because they are at the core of the plant life cycle (Grime et al. 1997; Westoby 1998). These are plant size (usually expressed as height), seed size (usually expressed as seed mass) and the structure of leaf tissue (often expressed as specific leaf area or leaf dry-matter content). Beyond this, there are some ‘core lists’ of plant traits that are considered important for plant resource use, regeneration, dispersal and response to widespread disturbances (e.g. Hodgson et al. 1999; McIntyre et al. 1999; Weiher et al. 1999; Lavorel and Garnier 2002; Knevel et al. 2003). A discussion of these is beyond the scope of the present manual, and readers are referred to these papers for a first introduction. For a particular question, the brief ecological background, and especially the list of references provided for each trait, should help identify the most appropriate traits to measure. Logistic and financial considerations are equally relevant. For example, if resources are limited for measuring relative growth rate on hundreds of species representing a large gradient of productivity, the specific leaf areas and stem-specific densities of these species might serve as less precise but still useful proxies for broad patterns of variation in growth and vegetation productivity. Similarly, the choice of traits would be slightly different if the limiting factor is labour force or access to sophisticated analytical laboratories, or if the project involves an intensive one-off measurement campaign carried out by highly trained specialists or recurrent measurements by third parties. The recipes provided here, including the sections on Special cases or extras, should assist in making those decisions. |
This is a recipe book to be used in the field and in the laboratory, and contains comprehensive, detailed, step-by-step recipes for direct and, as far as possible, unambiguous use in any terrestrial biome. To that end, we have had to make hard choices. We did not intend to provide a comprehensive list of all traits that could potentially be measured nor a thorough description of the theory behind each trait. Rather, the present handbook contains consensus traits and methods that researchers have identified as being useful, reliable and feasible to be applied in large-scale comparative efforts. Some of them are well known and widely used, whereas for others, relatively novel methods are described. Particular emphasis is given to recipes appropriate for areas with high species richness, incompletely known floras and modest research budgets. We give only brief ecological background for each trait, with a short list of references with further details on significance, methodology and existing large datasets. The main section of each recipe contains a brief, standardised protocol, and under the heading Special cases or extras, we give pointers to interesting additional methods and parameters. Readers can find complementary methods and additional discussions and comments in specific associated web pages (see Box 1). Specific citations have not been included in the recipe descriptions. We hope that the authors of relevant publications (most of them cited at the end of each recipe) will understand this choice, made for clarity and brevity, and in full recognition of the important contribution that each of them and many additional studies have made to the theory and measurement procedure for each trait.
This new handbook both updates theory, methods and databases covered by its predecessor (Cornelissen et al. 2003), and provides protocols for several additional plant functional traits, especially for organs other than the leaf. It has better coverage of (1) measurements important in less studied biomes and ecosystems, (2) floras with special adaptations and (3) plant functions related to carbon and nutrient cycling, herbivory, water dynamics and fire. We hope that the focus on practical techniques and streamlined trait recipes will help this handbook become a useful reference in laboratories and in the field for studies around the world. We strongly invite users to share their experiences with us about both general issues and specific details of these protocols (see Box 1), so that the next edition will be an even better bed-side table companion.
1 Selection of species and individuals
This section presents guidelines for selecting species and individuals within species for trait measurement, as well as general considerations of the necessary number of replicates. In addition, suggested numbers of replicates for all traits are given in Appendix 1.
1.1 Selection of species
Study objectives will always determine which species are selected for trait measurement. For species-level analyses of trait variation, and for identifying general strategies or syndromes of resource use, or trade-offs at the local, regional or global scale (e.g. Reich et al. 1997; Westoby et al. 1998; Díaz et al. 2004; Wright et al. 2004; Gubsch et al. 2011), species or populations from a broad range of environments and phylogenetic groups should be selected. For questions about evolution, the choice of species may be based on the inclusion of representatives of different enough phylogenetic groups, or on other phylogenetically relevant criteria (such as being members of particular clades), with little consideration about their abundance in situ. In contrast, when trying to understand how environmental variables shape vegetation characteristics, or how vegetation characteristics affect local flows of matter and energy (e.g. primary and secondary production, carbon, water and mineral nutrient cycling), the main criterion for species selection should be local abundance. In those cases, species should be selected that collectively make up for ~80% of cumulative relative abundance, following Garnier et al. (2004) and Pakeman and Quested (2007) (see specifics for abundance measurements below). Exceptions may be made if this criterion would imply measurements for an impracticably large numbers of species, e.g. communities with unusually high species richness per unit area, especially combined with a very high evenness. Examples are tropical rainforests and fynbos vegetation, in which well over 100 species per plot may be needed to reach the 80% biomass threshold.
In forests and other predominantly woody vegetation, the most abundant species of the understorey may also be included (e.g. when the research question relates to the whole-community or ecosystem level), even if their biomass is much lower than that of the overstorey woody species. In predominantly herbaceous communities, species contribution to a particular community may vary with time during a growing season. As a first step, we suggest that the relative abundance and the traits should be measured at the time of peak standing biomass of the community. This does not always apply to reproductive structures, which obviously have to be measured when they are present and fully developed, which sometimes does not coincide with the time of maximum vegetative growth.
For comparing sites or for monitoring trends in ecosystem-level properties across environmental conditions (e.g. pollution, or different regional climate or fertility levels), indicator species can be selected on the basis of the sensitivity of their trait values to the environmental factor of interest, and their importance locally and regionally, as well as for the ease with which they can be found and identified in the field (independent of their relative abundance) (Ansquer et al. 2009; De Bello et al. 2011). In this sense, it may be useful to distinguish ‘variable’ traits from more ‘stable’ traits (Garnier et al. 2007). Although most traits show some variation within species along environmental gradients, or in response to specific environmental changes, the intraspecific variation of so-called ‘stable traits’ is low compared with their interspecific variation. The reverse is the case for so-called ‘variable traits’, which implies that they should preferably be measured in more than one site or condition across the habitat range (Garnier et al. 2007). By contrast, ‘stable traits’ can be measured for any representative population from the entire gradient. Traits known to often be ‘variable’ include vegetative and reproductive plant height, mineral nutrient concentration in leaves, onset of flowering, branching architecture and spinescence. Traits that are relatively ‘stable’ include categorical traits, such as life form, clonality, dispersal and pollination modes, and to a lesser degree photosynthetic type (C3 or C4). Some quantitative traits such as leaf and stem dry matter content, or leaf toughness can be ‘stable’ along certain gradients, e.g. of nutrients or disturbance, but not along others, e.g. a light gradient (cf. Poorter et al. 2009). Species may therefore vary in which quantitative traits are stable across given gradients, so tests should be made before a trait is taken to be stable for a given species (Albert et al. 2010, 2012; Hulshof and Swenson 2010; Messier et al. 2010; Moreira et al. 2012).
Appendix 1 gives a rough indication of the within-species variability (coefficients of variation; i.e. standard deviation divided by the mean, hereafter CV) for some of the quantitative traits described in the present handbook, along with the more frequently used units and the range of values that can be expected. Appendix 1 summarises field data collected in several studies for a wide range of species coming from different environments. Because of the low number of replicates generally used, each of the individual estimates bears an uncertainty (and CV will likely increase as scale increases); however, by looking at the range of CVs calculated across a wide range of species, a reasonable estimate of the typical within-species-at-a-site variability can be obtained. We, therefore, present in Appendix 1 the 20th and 80th percentiles of the CV distribution.
How species abundance should be measured to determine the species making up 80% of cumulative abundance (e.g. whether to lay out transects, select points or quadrats at random or systematically, or to follow a different method) is beyond the scope of the present handbook and is extensively covered in plant-ecology and vegetation-science textbooks. However, it should be noted that different methods are relevant to different ecological questions and associated traits (Lavorel et al. 2008; see also Baraloto et al. 2010b, specifically for tropical forest). Taxon-free approaches that do not require species identification offer an alternative to estimates of relative abundance, and effectively capture the contribution of more abundant species. These include measuring traits regardless of species identity, along a transect (‘trait-transect’ method, Gaucherand and Lavorel 2007), or for individuals rooted nearest to random sampling points, as long as the canopy structure is quite simple (‘trait-random’ method – Lavorel et al. 2008). Methods of taxon-free sampling have also been applied to tropical forests, being, in this case, strongly based on the frequency or basal area of individual trees (Baraloto et al. 2010b). Trait values obtained through these methods can differ from those obtained using the standard approach of selecting robust, ‘healthy-looking’ plants for trait measurement (see Section 1.2).
1.2 Selection of individuals within a species
For robust comparisons across species, traits should be generally measured on reproductively mature, healthy-looking individuals, unless specific goals suggest otherwise. To avoid interaction with the light environment, which may strongly depend on neighbouring vegetation, often plants located in well lit environments, preferably totally unshaded, should be selected. This is particularly important for some leaf traits (see Section 3.1). This criterion creates sampling problems for true shade species found, e.g. in the understorey of closed forests, or very close to the ground in multilayered grasslands. Leaves of these species could be collected from the least shady places in which they still look healthy and not discoloured (see Section 3.1). Plants severely affected by herbivores or pathogens should be excluded. If feasible, for consistency among sets of measurements, use the same individual to measure as many different traits as possible.
Defining ‘individuals’ reliably may be difficult for clonal species (see Section 2.5), so the fundamental unit on which measurements are taken should be the ramet, defined here as a recognisably separately rooted, above-ground shoot. This choice is both pragmatic and ecologically sound, because genets are often difficult to identify in the field and, in any case, the ramet is likely to be the unit of most interest for most functional, trait-related questions (however, be aware that sampling of neighbouring ramets may not provide biologically independent replicates for species-level statistics). Individuals for measurement should be selected at random from the population of appropriate plants, or by using a systematic transect or quadrat method.
1.3 Replicate measurements
Trait values are often used comparatively, to classify species into different functional groups or to analyse variation across species within or between ecosystems or geographical regions. This type of research almost inevitably implies a conflict between scale and precision; given constraints of time and labour, the greater the number of species covered, the fewer replicate measurements can be made for each species. The number of individuals (replicates) selected for measurement should depend on the natural within-species variability in the trait of interest (see Section 1.1 for a discussion on within-species variability), as well as on the number or range of species to be sampled. Appendix 1 shows the minimum and preferred number of replicates for different traits, mainly based on common practice. The most appropriate sample size depends on the purpose and scope of the study. Ideally, researchers should check within-species CV at their site before deciding this. In broad-scale interspecific studies, one may sample relatively few plants of any given species, whereas when the study concerns just a small number of species or a modest local gradient, one may need to sample more heavily within each species. It is highly recommended to quantify the relative contributions of intra- v. interspecific variation. A formal analysis of statistical power based on an assumed or known variance among individuals, compared with that among species means, can be used. Commonly used statistical packages generally include routines for power analysis, as well as for variance component analyses (used to partition variance among different levels, e.g. species v. individuals). Other more powerful techniques can also be used, such as mixed models (Albert et al. 2010; Messier et al. 2010; Moreira et al. 2012).
2 Whole-plant traits
2.1 Life history and maximum plant lifespan
Plant lifespan (usually measured in years) is defined as the time period from establishment until no live part remains of the respective individual. Maximum plant lifespan is an indicator of population persistence and is therefore strongly related to land use and climate change. Lifespan is limited in non-clonal plants but may be apparently nearly unlimited in clonal plants. Maximum lifespan is strongly positively associated with environmental stress regimes, e.g. low temperatures and low nutrient availability. The relationship with disturbance frequency is mostly negative, although long-lived (resprouting) clonal plants may also tolerate frequent disturbance. There may be a trade-off between maximum lifespan and dispersal in time and space. Long-lived species often exhibit a short-lived seed bank and produce seeds or fruits with low dispersal potential, in contrast to short-lived species, which often have a very long-lived seed bank and/or high dispersal potential.
How to assess?
-
Life history
This simple classification distinguishes among the common types of timing and duration of survival behaviour of individual plants in the absence of disturbances or catastrophes.
-
Annual. Plant senesces and dies at the end of its first growing season (from seed), after producing seed, which may propagate a new plant in the future (a winter annual germinates in late summer or autumn, and so has two seasons, although the first may be very short).
-
Biennial. Plant grows vegetatively the first season, then flowers in the second to produce seed, followed by senescence and death of the shoot and root system.
-
Perennial. The individual survives for at least three growing seasons.
-
-
Monocarpic perennials. After several to many seasons of vegetative growth, the plant produces seeds, then senesces and dies.
-
Polycarpic perennials. All or much of the stem and root system normally survives the harsh or dormant period between growing seasons; stem has lateral thickening over the years. (i) Herbaceous perennial. Aerial shoots (and sometimes roots) die off as growing season ends; in the next season, new shoots grow from a perennating organ such as a bulb, corm, rhizome or ‘root crown’ (bud-bearing stem base or hemicryptophytes) near or below ground surface. (ii) Woody perennial retains, from one growing season into the next, some living, leaf-bearing shoots, which die by the end of their third season or later.
-
Qualitative distinction between life-history classes
A plant with any perennating organ other than the seed is either a perennial or a biennial (the latter only by a storage taproot). If biennial, there should be individuals with a storage root but not an inflorescence, and others with both. A plant that lacks specialised perennating organs may still be perennial, by resprouting from its root-crown. If so, the crown will normally carry wrinkles or scars from bud outgrowth in previous seasons, and can eventually become quite thick and even woody (a caudex); in contrast, the root of an annual is usually relatively soft and smooth, its thickness extending continuously into the stem. A perennial in its first year of growth may resemble an annual in these respects, except that perennial wild plants usually do not flower in their first year, whereas an annual always does (many horticultural perennials, however, have been selected to do so).
-
Maximum plant lifespan quantitative assessment
In gymnosperms and angiosperms, even in some non-woody ones, species maximum lifespan can be estimated by counting the number of annual rings representing annual tissue increments. Recently, a study on 900 temperate herbaceous species revealed annual rings in perennating structures in more than 80% of the species. However, the formation of annual rings can depend on habitat conditions. Annual rings will be found in vegetation zones with clear seasonality (cold (winter) or drought seasons) such as the polar, boreal or austral, temperate and even in Mediterranean-type zones. In the two latter climate zones, annual rings may sometimes be absent. In some cases, annual rings may even be found in tropical species, especially in regions with a distinct dry and wet season. Maximum lifespan within a population is studied in the largest and/or thickest individuals. Data are collected from a minimum of 10, preferably 20 individuals (replicates). In woody species (trees, shrubs, dwarf shrubs), annual rings are determined either by cutting out a whole cross-section or a ‘pie slice’ of the main stem (trunk), or by taking a core with a pole-testing drill (tree corer). It is important to obtain a rather smooth surface for clear observation. The annual rings can usually be counted under a dissection microscope. Often a cross-section of a shoot does not represent the maximum age as precisely as the root collar (root-stem transition zone of primary roots), which is especially true for most shrubs where single shoots have a limited age. We, therefore, recommend digging out woody plants a bit and taking (additional) samples from the root collar. In herbaceous species, annual rings are mostly found at the shoot base or in the root collar, and also in rhizomes. Here, microscopic cross-sections are essential and have to be treated first by ‘eau de javelle’ to remove the cytoplasm and then stained (fuchsin, chryosidine, astrablue (FCA); alternatively, astrablue and safranin) to make the annual rings visible. In some cases, polarised light has proven to be useful to identify the annual rings. Maximum lifespan of a species or population is defined as the largest number of annual rings counted among all samples (although the mean lifespan of all individuals may be informative too).
Special cases or extras
-
In clonal plants, the identification of (maximum) lifespan is more complicated. If a ramet never becomes independent from the genet and will never be released from the mother plant, annual rings in the tap root (e.g. Armeria maritima, Silene acaulis) or annual morphological markers along the rhizome or stolon (e.g. Lycopodium annotinum, Dictamnus albus) are also a suitable tool to identify maximum lifespan of a genet. In the latter case, maximum lifespan can be higher if part of the rhizome or stolon is already decomposed. However, in clonal plants where the genet consists of more or less independent ramets, genet age can be estimated only indirectly by means of size or diameter of a genet in relation to mean annual size increment.
-
Geophyte species, especially monocotyledons, may disappear above ground for up to several years before reappearing. In such cases, only permanent-plot research with individually marked individuals will give an idea about the maximum lifespan of those species.
-
Cold-climate dwarf shrubs. In some of these species, e.g. the heather Cassiope tetragona, lateral annual rings are often very hard to discern, whereas annual shoot-length increments of woody stems can be distinguished under a microscope through the winter-mark septa separating them and through the annual sequence of distances between leaf scars.
-
Life history and location. Life history varies with location and should preferably be assessed in the field rather than by reference to floras. In particular, many short-lived, faster-growing species fall into different life-history categories in different regions and a few differ among habitats, even within the same region.
References on theory and significance: Rabotnov (1950); Schweingruber (1996); Fischer and Stöcklin (1997); Larson (2001); Schweingruber and Poschlod (2005); De Witte and Stöcklin (2010).
More on methods: Tamm (1972); Gatsuk et al. (1980); Cherubini et al. (2003); Rozema et al. (2009).
2.2 Life form
Plant life-form classification sensu Raunkiaer (1934) is a simple but still a useful way of functionally classifying plants. More information is given in Material S1, available as Supplementary Material for this paper.
2.3 Growth form
Growth form is mainly determined by the direction and extent of growth, and any branching of the main-shoot axis or axes. These affect canopy structure, including its height, and both the vertical and horizontal distribution of leaves. Growth form may be associated with ecophysiological adaptation in many ways, including maximising photosynthetic production, sheltering from severe climatic conditions, or optimising the height and positioning of the foliage to avoid or resist grazing by particular herbivores, with rosettes and prostrate growth forms being associated with high grazing pressure by mammals.
How to record?
Growth form is a hierarchical trait assessed through field observation or descriptions or figures or photographs in the literature. Because we are classifying types along a continuum, intermediate forms, between the categories recognised here, may be encountered, as well as occasional unique forms lying outside any of these categories.
-
Terrestrial, mechanically and nutritionally self-supporting plants
-
-
Herbaceous plants have either no or at most modest secondary growth, with stem and root tissues that are rather soft compared with typical wood.
-
Rosette plant. Leaves concentrated on a short, condensed section of stem or rhizome (see Category C under Section 2.5 for a definition of rhizome), at or very close to the soil surface; with an inflorescence (or single-flower peduncle) bearing either no or reduced leaves (bracts) produced from the rosette axis, above ground level. Graminoids whose principal photosynthetic leaves are attached to the base of their aerial stems (e.g. ‘bunch grasses’) fall in this category.
Elongated, leaf-bearing rhizomatous. The permanent axis is an elongated rhizome that directly bears photosynthetic leaves that extend individually up into the light. The rhizome can be located either at or below ground level (e.g. Pteridium aquilinum (bracken fern), Viola spp., Iris spp.), or (epiphytes) on an above-ground support such as a tree branch. Aerial inflorescences (or single-flower peduncles) with either reduced leaves (bracts), or none, may grow out from the rhizome.
Cushion plant (pulvinate form). Tightly packed foliage held close to soil surface, with relatively even and rounded canopy form (many alpine plants have this form).
Extensive-stemmed herb develops elongated aerial stem(s) whose nodes bear photosynthetic leaves that are distributed nearly throughout the canopy of the plant, except when shed from its more basal parts during later growth, and lacking in distally developed inflorescences. Graminoids (rhizomatous or not) with leafy aerial stems fall here.
Tussock. Many individual shoots of a dense colony or clone grow upward, leaving behind a tough, mostly dead supporting column topped by living shoots with active leaves (e.g. the Arctic cotton grass, Eriophorum vaginatum).
-
Semi-woody plants. Stem without secondary growth but often toughened by sclerification (or, alternatively, with relatively feeble, soft or ‘anomalous’ secondary growth).
-
Palmoid. Bears a rosette-like canopy of typically large, often compound leaves atop a usually thick (‘pachycaulous’), columnar, unbranched or little-branched stem (e.g. palms (Pandanus), tree ferns). Certain tropical or alpine Asteraceae such as Espeletia spp., cycads, Dracaena, arborescent Yucca spp. and some Bombacaceae can be regarded as having this growth form, although their stems undergo more extensive secondary growth (see also ‘Corner model’ within the references below).
Bambusoid. An excurrently branched (cf. Point A.3.d.i in the present Section) trunk lacking or having only weak secondary growth is stiffened by sclerification to support a vertically extensive, sometimes tree-sized canopy (bamboos; various tall, herbaceous dicots such as Chenopodium, Amaranthus and Helianthus).
Stem succulent. A usually leafless photosynthetic stem with extensive, soft, water-storage tissue and only limited secondary growth (cacti, and cactoid plants of other families; most leaf succulents fall instead into one of the subclasses of Points A.1 or A.3 in the present Section).
-
Woody plants develop extensive, usually tough, secondary xylem and phloem from vascular cambium, and corky outer bark from cork cambium (woody vines are covered in Point B.3 of the present Section).
-
Prostrate subshrub. Long-lived woody stem growing horizontally at ground level (examples include many Arctic willows and ericoids).
Dwarf shrub, or subshrub, with usually multiple, ascending, woody stems less than 0.5 m tall.
Shrub. Woody plant between 0.5 m and ~5 m tall, with canopy typically carried by several trunks that are usually thinner and younger than typical mature tree trunks.
Tree. Woody plant usually >5 m tall, with main canopy elevated on a long-lived, substantial, usually single (but upwardly branching), trunk. (i) Excurrent. Single main axis (trunk) extends up to, or almost to, the top, with shorter, ascending or horizontal branches giving a conical or (in mature trees) columnar form to the crown. (ii) Deliquescent. Trunk divides, somewhere above its base, into two to several, more or less equal branches that continue branching upward to produce a wider, more flat-topped crown.
Dwarf tree. Morphology as in one of Types (i) or (ii) but substantially <5 m tall. Many forest understorey trees, but also in various climatically or nutritionally challenging, unshaded habitats, such as ‘pine barrens’, semi-deserts, certain tropical cloud forests, bogs and near-timberline vegetation.
-
-
Plants structurally or nutritionally supported by other plants or by special physical features
-
-
Epiphyte. Plant that grows attached to the trunk or branch of a shrub or tree (or to anthropogenic supports) by aerial roots, normally without contact with the ground (e.g. many tropical orchids and Bromeliaceae).
-
Lithophyte. Plant that grows in or on rocks (e.g. many species of ferns, species of Nepenthes, Utricularia forestii, Cymbalaria muralis).
-
Climber. Plant that roots in the soil but relies, at least initially, on external support for its upward growth and leaf positioning.
-
Herbaceous vine. Usually attaches to its support either by twining or by means of tendrils.
Woody vine, including liana. Often attaches to a support by means of aerial roots.
Scrambler. Grows up through a sufficiently dense canopy of other plants, without any means of attachment (e.g. Galium spp.).
Strangler. May start epiphytically (but become soil-rooted) or by climbing from ground level. However, by secondary growth, it later becomes self-supporting, and may eventually envelope the initially supporting stem (e.g. certain tropical Ficus spp.).
-
Submersed or floating hydrophyte. Herbaceous, aquatic plant that relies on surrounding water for physical support. (Emergent hydrophytes (‘helophytes’) mostly fall into one of the subgroups of Point A.1 in the present Section.)
-
Parasite or saprophyte obtains important nutritional needs directly or indirectly from other vascular plants (parasite) or from dead organic matter in the soil (saprophyte) (see Nutrient uptake in Material S2 where other more specific forms of parasitism are covered).
-
References on theory, significance and large datasets: Cain (1950); Ellenberg and Müller-Dombois (1967); Whittaker (1975); Barkman (1988, and references therein); Rundel (1991); Richter (1992); Box (1996); Ewel and Bigelow (1996); Cramer (1997); Lüttge (1997); Medina (1999); McIntyre and Lavorel (2001).
More on methods: Barkman (1988, and references therein).
2.4 Plant height
Plant height is the shortest distance between the upper boundary of the main photosynthetic tissues (excluding inflorescences) on a plant and the ground level, expressed in metres. Plant height, or maximum height (Hmax), is the maximum stature a typical mature individual of a species attains in a given habitat. Hmax is associated with growth form, position of the species in the vertical light gradient of the vegetation, competitive vigour, reproductive size, whole-plant fecundity, potential lifespan, and whether a species is able to establish and attain reproductive size between two disturbance events (such as e.g. fire, storm, ploughing, grazing).
What and how to measure?
Healthy plants should be sampled that have their foliage exposed to full sunlight (or otherwise plants with the strongest light exposure for that species). Because plant height is quite variable both within and across species, there are three ways to estimate Hmax, depending on species size and the number of plants and time available, including the following: (1) for short species, measurements are taken preferably on at least 25 mature individuals per species; (2) for tall tree species, height measurements are time-consuming, and for these, the height of the five tallest mature individuals can be measured; and (3) for trees, when more time is available, measure ~25 individuals that cover the entire range of their height and diameter. Use an asymptotic regression to relate height to diameter, and derive the asymptote from the regression coefficients, or use the formula to calculate the height of the thickest individual in the stand.
The height to be measured is the height of the foliage of the species, not the height of the inflorescence (or seeds, fruits), or the main stem if this projects above the foliage. For herbaceous species, this is preferably carried out towards the end of the growing season. The height recorded should correspond to the top of the general canopy of the plant, discounting any exceptional branches, leaves or photosynthetic portions of the inflorescence.
For estimating the height of tall trees, some options are
-
a telescopic stick with decimetre marks; and
-
trigonometric methods such as the measurement of the horizontal distance from the tree to the observation point (d) and, with a clinometer or laser, the angle between the horizontal plane and the tree top (α) and between the horizontal plane and the tree base (β); tree height (H) is then calculated as H = d × [tan(α) + tan(β)]; height estimates are most accurate if the measurement angle is between 30 degrees (easier to define the highest point in the crown) and 45 degrees (a smaller height error caused by inaccuracy in the readings); the horizontal distance between the observer and the stem should preferably equal 1–1.5 times the tree height.
Special cases or extras
-
Rosettes. For plants with major leaf rosettes and proportionally very little photosynthetic area higher up, plant height is based on the rosette leaves.
-
Herbaceous. For herbaceous species, vegetative plant height may be somewhat tricky to measure (if the plant bends, or if inflorescence has significant photosynthetic portions), whereas reproductive plant height can be ‘safer’ in this sense. Additionally, some authors have suggested that the projection of an inflorescence above the vegetative part of the plant may be a useful trait in responses to disturbance, so both of these heights should be useful to measure. Others, while recording maximum canopy height, arbitrarily use a leaf length of two-thirds of the largest leaf as the cut-off point to estimate the position of a transition between vegetative and reproductive growth.
-
Epiphytes. For epiphytes or certain hemi-parasites (which penetrate tree or shrub branches with their haustoria), height is defined as the shortest distance between the upper foliage boundary and the centre of their basal point of attachment.
-
Large spreading crowns. For trees with large spreading crowns, it is difficult to estimate the height above the tree stem. For such individuals, it is easier to measure (with an optical rangefinder or laser) the vertical height as the distance from eye to a location at the crown margin that is level with the tree top; multiply this by the sine of the sighting angle to the horizontal (as measured with a clinometer) and add the vertical height from eye level down to tree base (a subtraction if eye level is below tree base level).
-
Dense undergrowth. For vegetation types with dense undergrowth that makes the measurement of Hmax difficult, there are modified versions of the equation above; they involve the use of a pole of known height that must be placed vertically at the base of the tree.
References on theory, significance and large datasets: Gaudet and Keddy (1988); Niklas (1994); Hirose and Werger (1995); Thomas (1996); Westoby (1998); Kohyama et al. (2003); King et al. (2006); Poorter et al. (2006, 2008); Moles et al. (2009).
More on methods: Korning and Thomsen (1994); Thomas (1996); Westoby (1998); McIntyre et al. (1999); Weiher et al. (1999).
2.5 Clonality, bud banks and below-ground storage organs
Clonality is the ability of a plant species to reproduce or regenerate itself vegetatively, thereby producing new ‘ramets’ (above-ground units) and expanding horizontally. Clonality can give plants competitive vigour and the ability to exploit patches rich in key resources (e.g. nutrients, water, light). Clonal behaviour may be an effective means of short-distance migration under circumstances of poor seed dispersal or seedling recruitment. Clonality also gives a plant the ability to form a bud bank, which can be a very important determinant of recovery and persistence after environmental disturbances. The bud bank consists of all viable axillary and adventitious buds that are present on a plant and are at its disposal for branching, replacement of shoots, regrowth after severe seasons (winter, dry season, fire season), or for vegetative regeneration after injury (adventitious buds that arise after the injury, which are an important means of regeneration in some plants, apparently lie outside the ‘bud bank’ concept). Both the characteristics of the bud bank and the type of clonal growth exhibited by plants determine their ability to recover from disturbances (see Material S3 for a protocol for Characterisation of the bud bank, based on Klimeš and Klimešová 2005). Clonal organs, especially below-ground ones, also serve as storage and perennating organs; a sharp distinction between these functions is often impossible.
How to collect and classify?
For above-ground clonal structures, observe a minimum of five plants that are far enough apart to be unlikely to be interconnected, and that are well developed. For below-ground structures, dig up a minimum of five healthy-looking plants (Appendix 1). In some cases (large and heavy root systems), partial excavation may give sufficient evidence for classification. It is best to assess clonality and bud banks near the end of the growing season. Remove the soil and dead plant parts before counting buds or classifying the organs. The species is considered clonal if at least one plant clearly has one of the clonal organs listed below (see References below in the present Section for discussion).
Categories are then
-
clonal organs absent;
-
clonal organs present above ground, including the following:
-
-
stolons – specialised, often hyper-elongated horizontal stems whose axillary bud growth and nodal rooting yields ultimately independent plants (e.g. strawberry (Fragaria vesca), saxifrage (Saxifraga flagellaris));
-
bulbils – deciduous, rooting bulblets produced from axillary or what would otherwise be flower buds, or by adventitious bud growth on leaves (e.g. Cardamine pratensis, Bryophyllum); analogous vegetative propagules of bryophytes are termed gemmae; and
-
simple fragmentation of the vegetative plant body (mostly aquatic plants, and bryophytes);
-
-
clonal organs present below ground, including the following:
-
-
rhizomes – more or less horizontal, below-ground stems, usually bearing non-photosynthetic scale leaves (e.g. many grasses and sedges), and sometimes instead bearing photosynthetic leaves that emerge above ground (e.g. Iris, Viola, bracken fern (Pteridium)); aerial, vegetative and/or reproductive shoots grow up from axillary (or sometimes terminal) buds on the rhizome; most rhizomes can branch, after which decline and decay of the portion proximal to the branch point yields independent, clonally generated individuals;
-
tubers and turions – conspicuously thickened, below-ground stems or rhizomes, functioning as carbohydrate storage organs and bearing axillary buds, that can propagate the plant (e.g. potato Solanum tuberosum, Jerusalem artichoke (Helianthus tuberosus)); similar organs formed on aquatic plants are termed turions;
-
bulbs – relatively short, below-ground stems that bear concentrically nested, fleshy scale-leaves that act as storage organs, the whole globose structure serving to perennate the plant and, through growth of axillary buds within the bulb into daughter bulbs or ‘offsets’, to multiply it vegetatively (e.g. tulip (Tulipa), onion (Allium));
-
corms – vertically oriented, globosely thickened underground stems that serve as storage organs and bear either scale or foliage leaves; axillary or terminal buds on the corm function for perennation and to a limited extent for clonal reproduction (e.g. Dahlia);
-
tuberous roots – thickened roots that serve primarily for storage but can form adventitious buds that permit clonal propagation (e.g. sweet potato (Ipomoea batatas));
-
suckers – shoots developed from adventitious buds produced on ordinary, non-storage roots (e.g. aspen (Populus tremuloides), wild plum (Prunus spp.)); the sucker shoots can become independent plants once the root connection between them and the parent is severed or dies;
-
lignotuber – a massive, woody expansion just below the ground surface, produced by secondary growth of the ‘root crown’ in many shrubs in fire-prone vegetation; after a fire that kills the shrub’s aerial canopy, adventitious buds on the lignotuber grow out to regenerate the shrub’s canopy (see Section 6.6), normally not resulting in clonal multiplication; and
-
layering – ordinary vegetative shoots that lie on or bend down to the ground, there produce adventitious roots and continue apical growth, becoming independent plants when their connection with the parent is severed (e.g. blackberry and raspberry (Rubus), certain spp. of spruce (Picea) and hemlock (Tsuga)).
-
If a plant species has clonal growth (Categories B or C above in the present Section), classify it according to one or more of the following categories:
-
regenerative clonal growth, occurring after injury and normally not multiplying the number of individuals, as with resprouting from a lignotuber;
-
additive (also termed multiplicative) clonal growth, which can be either the plant’s normal mode of multiplication or can be induced by environmental conditions such as high nutrient availability, and serves to promote the spread of the plant;
-
necessary clonal growth is indicated when clonality is required for the year-to-year survival of the plant, as with many plants that perennate from rhizomes, bulbs, tubers or tuberous roots and have no, or weak, seed reproduction.
Clonal growth may fulfil more than one of these functions, in which case it may not be possible to distinguish between them. In some cases, the functional nature of clonal growth may be simply
-
unknown or not evident, in which case it may be recorded as such.
References on theory, significance and large datasets: De Kroon and Van Groenendael (1997); Klimeš et al. (1997); Van Groenendael et al. (1997); Klimeš and Klimešová (2005); Knevel et al. (2003); Klimešová and Klimeš (2007).
More on methods: Böhm (1979); Klimeš et al. (1997);Van Groenendael et al. (1997); Weiher et al. (1998); Klimeš and Klimešová (2005).
2.6 Spinescence
Spinescence refers to the degree to which a plant is defended by spines, thorns and/or prickles. Spines are sharp, modified leaves, leaf parts or stipules; they also occur sometimes on fruits. Thorns are sharp, modified twigs or branches. Prickles are modified epidermis or cork (e.g. rose-stem prickles). Because spinescence is clearly involved in anti-herbivore defence, especially against vertebrate herbivores, the following two separate issues are critical in considering spinescence: (1) the effectiveness of physical defences in preventing or mitigating damage from herbivores; and (2) the cost to the plant in producing these defences. Different types, sizes, angles and densities of spines, thorns and prickles may act against different herbivores. Although in many cases, characterisations of plant spinescence by measuring spines is sufficient, some researchers may decide that experiments with actual herbivores, which examine the effectiveness of anti-herbivore defences, are necessary, e.g. by offering whole shoots (with and without spines) to different animals and recording how much biomass is consumed per unit time (see Special cases or extras in Section 3.16).
Spines, thorns and prickles can be an induced response to herbivory, meaning that some plants invest in these defences only when they have already been browsed by herbivores. Other types of damage, including pruning and fire, can also induce increased levels of spinescence. In addition, spinescence traits can change drastically with the age of the plant or plant part, depending on its susceptibility to herbivory. For this reason, spinescence sometimes cannot be considered an innate plant trait, but rather a trait that reflects the actual herbivore pressure and investment in defence by plants. In other words, although there are species that always have spines, and species that never have them, the spinescence of an individual plant is not necessarily representative of the potential range of spinescence in the whole species (e.g. some members of Acacia and Prosopis show a striking range of spine lengths within the same species, depending on individuals, age and pruning history). Spines, thorns and prickles can sometimes play additional roles in reducing heat or drought stress, especially when they densely cover organs.
How to measure?
Spines, thorns and prickles – summarised below as ‘spines’ – can either be measured as a quantitative trait or reduced to a qualitative, categorical trait. Data on spinescence are preferably measured from specimens in the field, and can also be gathered from herbarium specimens or descriptions in the literature. Spine length is measured from the base of the spine to its tip. If a spine branches, as many do, its length would be to the tip of the longest branch. Spine width, measured at the base of the spine, is often more useful for assessing effectiveness against herbivores and more generalisable across types of spines. The number of branches, if any, should also be recorded because branches can increase significantly the dangerousness of spines to herbivores. Ratio of spine length to leaf length can also be a useful character because it gives an idea of how protected the lamina is by the spine closest to it.
Spine strength or toughness. Spines are ‘soft’ if, when mature, they can be bent easily by pressing sideways with a finger, and ‘tough’ if they cannot be thus bent. Spine density is the number of spines per unit length of twig or branch, or area of leaf.
Biomass allocation to spines is also an important parameter for some research questions. Its estimation is more work-intensive than those above, but still relatively simple. Cut a standard length of stem or branch, cut off all spines, oven-dry and weigh leaves, shoot and spines separately and estimate fractional allocation as the ratio of spine dry weight to shoot dry weight.
These quantitative trait measurements can be converted into a categorical estimate of spinescence by using the classification proposed in Box 3.
| Box 3. Categorical estimates of spinescence |
|
Finally, to simply record the presence or absence of spines is sufficient in some cases. Bear in mind that the size, structure and behaviour of herbivores vary enormously, so the degree of protection provided by spine mass, size and distribution can be determined only with reference to a particular kind of herbivore. When selecting the most meaningful measurement/s of spinescence, always consider what herbivores are relevant.
References on theory, significance and large datasets: Milton (1991); Grubb (1992); Cooper and Ginnett (1998); Pisani and Distel (1998); Olff et al. (1999); Hanley and Lamont (2002); Rebollo et al. (2002); Gowda and Palo (2003); Gowda and Raffaele (2004); Agrawal and Fishbein (2006).
2.7 Branching architecture
Branching architecture refers to how intensively a plant branches (number of living ramifications per unit of stem length). Highly branched plants can be better defended against vertebrate herbivores, primarily by making feeding less efficient, denying access by herbivores to plant organs, and ensuring that, if herbivores do remove growing tips, there remain enough for the plant to continue growing. Conversely, less branched plants can be adapted to environments where growing tall quickly is necessary, as in a fire-prone savannah or a forest undergoing the pioneer stages of secondary succession. Branching architecture can also be adaptive in forest systems, where species that utilise low light tend to be more branched for a given height than are species that utilise only bright light.
Although there are complicated and elegant methods for evaluating branching architecture, a simple characterisation such as the one described below is often sufficient for understanding the adaptive significance of this trait. Like spinescence, branching architecture is a plastic trait that can differ within a species on the basis of browsing history, fire history, access to light, plant vigour or disease and even water stress. Branching architecture is also variable depending on the age and life history of the plant (see Section 1.1 for recommendations related to variable traits).
How to measure?
To assure measuring a branch that best represents the branching architecture of a plant (a branch that reaches the outer part of the canopy), work backwards from a terminal, leaf-bearing branch until reaching the first branch that is now leafless at its base but bears secondary branches that have leaves. The base of this branch will be the starting point for measuring (1) the total length of the branch, which is the distance from the starting point to the tip of its longest-living terminal and (2) the number of ramification points that lead to living branches; from each ramification point, move towards the tip, always following the most important branch (the main branch is often the thickest living branch coming from a ramification point; see Fig. 1 for a graphic explanation). An indicator of branching architecture, called apical dominance index (ADI), is obtained by dividing the number of ramifications by the total length of the branch in metres. The value of ADI can vary between zero (no branching) to >100 m–1 (extremely ramified).

|
References on theory, significance and large datasets: Horn (1971); Pickett and Kempf (1980); Strauss and Agrawal (1999); Enquist (2002); Archibald and Bond (2003); Cooper et al. (2003); Staver et al. (2011).
More on methods: Fisher (1986).
2.8 Leaf area : sapwood area ratio
The amount of leaf area a species produces per unit cross-section of sapwood (the inverse of Huber value, expressed in mm2 mm–2) is crucial for both water transport (with related effects on photosynthetic rate) and mechanical strength.
What and how to collect
The ratio leaf area : sapwood area (LA : SA) depends strongly on leaf phenology. Furthermore, there is variation between wet and dry seasons, variation among populations of a given species along moisture gradients, ontogenetic trajectories for given individuals, and within trees along a branch from trunk to tip. Declining function of sapwood with age is one reason why LA : SA generally increases when one moves from larger (older) towards smaller branches. Unfortunately, the age-related decline in sapwood function is not always well understood, can be difficult to measure, and may vary among species. All this should be considered when designing a sampling methodology and interpreting this trait (see Point 3 of Special cases or extras in the present Section).
To make meaningful comparisons among species, we recommend sampling terminal, sun-exposed shoots from the outer canopy. This means sampling terminal shoots either of a certain standard length, or of a certain age (1–3 years) (for shoots in which terminal bud scars allow their age to be determined). This approach maximises the likelihood that all the sapwood in the branch is still functional. We recommend sampling at the peak of the growing season when leaf area is highest. At this time, LA : SA should be at a maximum for the year; this is similar to the efforts to measure maximum photosynthetic rate as a way of making meaningful comparisons across species. Care should be taken to select shoots that have not lost leaves or parts of leaves to mechanical damage, herbivory or early senescence and abscission.
Measuring
Leaf area : sapwood area ratio can be measured at different scales, namely, from whole plant to just terminal branches (and this should be taken into consideration when scaling up measurements). Total leaf area of leaves distal to the collection point is measured by the same method as the area of individual leaves (see Section 3.2). Sapwood area at the collection point is most precisely measured with digital micrographs and image-analysis software (see Section 3.1 for free software); however, a calliper should work for most species in most situations. In measuring sapwood area, care should be taken to exclude bark, phloem, heartwood and pith from the area measured.
Special cases or extras
-
For herbaceous species, similar methods can be applied; however, care must be taken to identify the parts of the stem that can conduct water; this distinction may not be as clear as it is within most woody species. It can be quantified with a dye-transport experiment (see Point 3 below in the present Section).
-
Seasonal changes. Because cambial growth in many trees continues well after spring flush of bud growth is completed and the final leaf area for the season is attained, the LA : SA ratio is best measured as late in the growing season as possible, when all the season’s newly produced leaves remain attached, but (for evergreens) before the seasonal abscission of older leaves has occurred.
-
In ring-porous trees, the effective conductivity of xylem drops precipitously in older sapwood, sometimes within a very few annual rings. For these species, the conductivity of the sapwood (and its decline with sapwood age) can be quantified by placing the cut end of the shoot into a fairly strong solution of a dye, such as eosin, and allowing the foliage to transpire in air, and after 10–20 min, cutting a cross-section of the stem a few centimetres above its cut end and measuring the dye-stained area.
References on theory, significance and large datasets: Chiba (1991); Eamus and Prior (2001); Maherali and DeLucia (2001); Mäkelä and Vanninen (2001); McDowell et al. (2002); Preston and Ackerly (2003); Addington et al. (2006); Buckley and Roberts (2006); Maseda and Fernández (2006); Wright et al. (2006); Cornwell et al. (2007); Litton et al. (2007).
References on meta-analysis: Mencuccini (2003).
2.9 Root-mass fraction
Theory predicts that plants from nutrient-poor sites should allocate a greater fraction of new biomass to roots and maintain a higher proportional distribution of biomass in roots than in shoots. Distribution of biomass to roots can be simply expressed as the root-mass fraction (RMF, synonymous to root-mass ratio, RMR), identically calculated as the proportion of plant dry mass in roots. Note that a true allocation measurement requires quantifying turnover rates as well as standing distributions, which is labour-intensive and rarely carried out. Allocation and distribution are often used synonymously, and whether this is appropriate or not, we follow this convention herein. The RMF is preferable to the often used root : shoot ratio (RSR), because the RFM is bounded between 0 and 1, and can be immediately interpreted and compared, whereas the RSR is unconstrained and can vary from a tiny to a very large number. Notably, root allocation can be highly plastic across light, nutrient and water supplies. Some patterns can be apparently contradictory, because root allocation can allow both greater foraging below ground, which would be an advantage especially when resources are low, and also greater competition below ground, being an advantage when resources are plentiful. In reviews of experimental studies, including those that take an allometric approach, RMF typically decreases with increasing nitrogen availability. However, other studies have reported that for field plants, fast-growing species adapted to nutrient-rich habitats showed higher allocation to roots than did slow-growing species from nutrient-poor sites. Similarly, seedlings showing plastic responses to low light typically decrease their RMF, whereas plants adapted to chronic deep shade in rainforests tend to have higher RMF, apparently to survive periods of low water and nutrient supply in competition with surrounding trees. Note that some reports of differences in RMF across resource gradients are potentially confounded by failure to account for allometry and size (see References on theory, significance and large databases below in the present Section). Additionally, RMF does not directly translate to a high soil resource-uptake rate. Lower allocation to roots may well be compensated by higher specific root length (see Section 5.1) and by higher uptake rate per allocation to root mass, length or surface area.
The RMF can best be used for comparative purposes if measured for plants of similar mass. Alternatively, if plants are harvested of a range of mass, allometries can be used to estimate RMF for plants of a given size.
Care should be taken to harvest all the roots (see Section 5), despite the difficulty of separating roots from soil, particularly fine roots. However, in field studies, sometimes RMF includes only a subset of all below-ground tissues; in such a case, the researcher should be clear about what is included and what is not.
Special cases or extras
-
Storage organs and root fractioning. RMF should in theory include everything that is plant-developed (so not including mycorrhizae!). However, particular studies can subdivide specific fractions for specific purposes (i.e. fine roots, coarse roots, crowns, rhizomes (for grasses), tap roots (in trees)) to evaluate the relative proportions of each in relation to each other and/or to above-ground biomass.
-
-
References on theory, significance and large databases: Evans (1972); Grime (1979); Aerts et al. (1991); Elberse and Berendse (1993); Veneklaas and Poorter (1998); Aerts and Chapin (1999); Reich (2002); Sack et al. (2003); Poorter et al. (2012).
-
2.10 Salt resistance
Many areas of the world, including coastal ones, those with poorly drained soils in arid climates, and those with poorly designed irrigation systems, feature high concentrations of salt (>100–200 mM sodium chloride, NaCl). Only salt-resistant species, which exhibit strategies to reduce or avoid damaging effects of excess salt in their tissues, are able to maintain viable populations in such areas. Plants specialised for inhabiting saline soils, and often restricted to these, are termed halophytes.
Among members of the at least 139 plant families that include halophytes, evolution has yielded multiple solutions to the problem of excess salt in the environment, involving different biochemical, physiological, structural and/or phenological traits. Therefore, rather than a single recipe for assessing salt resistance, we give several traits and measurements that together help identify a species as salt-resistant, especially if these are accompanied by data on species distribution in saline areas. However, to positively classify a species as salt-sensitive would be problematic from these traits alone. Experimental testing of plant survival and growth under saline conditions is necessary, which would by no means be quick and easy for screening multiple species. Thus, the traits described below allow a qualitative rather than quantitative assessment of salt resistance, and do not allow the clear separation of more or less salt-resistant species from true halophytes. Hopefully, this text will stimulate research into novel approaches and protocols for testing salt resistance more efficiently and comprehensively.
Here, we simplify more extensive, previous classifications of mechanisms by which plants deal with excess environmental NaCl, focusing on three common strategies. Some salt-resistant plant species can limit the uptake of potentially damaging Na+ by their roots (NaCl ‘excluders’). However, many salt-resistant species cannot avoid significant NaCl uptake. These plants can either actively excrete excess salt or can accumulate NaCl in cell vacuoles, so as to prevent toxicity to the cytosol. The latter (‘salt-tolerant’) species are often succulent, with many characteristics of drought-tolerant species. Many salt-resistant species possess biochemical mechanisms to reduce salt stress or damage in the tissues, by accumulating compatible solutes (including secondary metabolites) in the cytosol. The salt-resistance traits detailed below fall into the foregoing categories, except special biochemical adaptations that are not covered here.
What and how to measure?
Selective root cation uptake. Roots of many salt-resistant plant species (particularly monocots) can discriminate against Na+, while maintaining uptake of essential potassium (K+). This selectivity for K+ over Na+ increases the K+ : Na+ ratio in the cytosol compared with that in the rooting medium. Because these ratios may vary with several environmental factors, including precipitation and evapotranspiration, we suggest sampling leaves and soil on at least three different days, at intervals of 2 weeks or more during the growing season, but not for 5 days after particularly heavy or prolonged rain. Collect leaves from five separate plants (Appendix 1), and a soil sample from the main fine-root zone below each. The Na+ and K+ concentrations of each sample are to be determined in the laboratory by a standard assay. Popular and convenient methods include atomic emission spectrometry (EAS), also called flame photometry, and atomic absorption spectrometry (AAS). Leaf samples are to be ground in an equal mass of water, which is then extracted from the homogenate by filtration. For soil, add water to a dry soil until it becomes water-saturated and then extract the liquid by suction or vacuum filtration. Na+ and K+ assays can be performed either on the water phase, or after evaporating it, depending on the Na+ and K+ assay method.
Calculate, for each plant and associated soil sample, the K : Na selectivity (S) as S = ([K+]/[Na+])plant / ([K+]/[Na+])soil. A mean S value for a species is calculated from the mean of all replicate S values per sampling date, by taking the average of these over all sampling dates.
Salt excretion. Salt-excreting species eject NaCl through special glands or bladders on the (usually lower) surfaces of their photosynthetic organs (usually leaves, but in some cases stems). These glands are often visible (especially under a hand lens) as small, irregularly shaped white spots that are excreted salt crystals on the surface of the gland. A salty taste, on licking one of these, will confirm this. Some species excrete salt from their roots. Although this is more difficult to observe, one may check for similar salt excretions on the surfaces of any roots uncovered during soil sampling. Note that salt excretions on shoots or roots will wash off during wet weather, so are best sought after a dry period.
Salt compartmentalisation. Salt compartmentalisation is indicated by clear succulence of the leaves or photosynthetic stems. Succulent green stems can be treated and measured as if they were leaves (see Special cases or extras in Section 3.1). Succulence leads to high leaf water content (LWC) and leaf thickness (Lth), and may be quantified as the product of these parameters (succulence (mm) = Lth × LWC) (see Section 3.3). Values >800–1000 mm indicate significant succulence.
Strong salt-related succulence is found almost exclusively in dicotyledonous species, although certain salt-tolerant monocots can be somewhat succulent, e.g. Elytrigia juncea on beach dunes. Salt-tolerant succulents show a high NaCl level in their leaves, which can distinguish them from crassulacean acid metabolism (CAM) succulents (see Section 3.12; some salt-tolerant succulents are actually also CAM plants). This could be detected by the Na assay on leaf or stem extracts noted above, or would be revealed very easily by measuring the electrical conductivity of such extracts (see Electrolyte leakage in Section 3.14), which requires only a simple, widely available conductivity meter (NaCl in solution gives a high conductivity). Qualitative evidence for this can be a combination of juiciness and noticeably salty taste when chewing the tissue. This property has made some halophytes popular as human food, e.g. Salicornia spp.
Special cases and extras
-
Succulents and halophytes. Many salt-tolerant succulents are halophytes and occur only in saline environments; expression of the traits described above can depend on the actual salinity of the plants’ soil. We, therefore, suggest measuring soil salt concentrations (as described under Selective root cation uptake above, within the present Section) to accompany trait measurements. Several other salt-related habitat descriptors are also relevant, e.g. elevation and duration of daily marine inundation (if any) in salt marshes or on beaches, and location relative to the high tide mark visible as a litter belt, or white patches on the soil surface, indicating salt crystals in dry areas.
References on theory, significance and large datasets: Flowers et al. (1977, 1986); Yeo (1983); Rozema et al. (1985); Zhu (2001); Breckle (2002); Munns et al. (2002); Vendramini et al. (2002); Ashraf and Harris (2004); Flowers and Colmer (2008).
More on methods: Jennings (1976); Maas and Hoffman (1977); FAO (1999); Breckle (2002); Vendramini et al. (2002).
2.11 Relative growth rate and its components
Relative growth rate (RGR) is a prominent indicator of plant strategy with respect to productivity as related to environmental stress and disturbance regimes. RGR is the (exponential) increase in size relative to the size of the plant present at the start of a given time interval. Expressed in this way, growth rates can be compared among species and individuals that differ widely in size. By separate measurement of leaf, stem and root mass as well as LA, good insight into the components underlying growth variation can be obtained in a relatively simple way. These underlying parameters are related to allocation (leaf-mass fraction, the fraction of plant biomass allocated to leaf), leaf morphology (see Section 3.1), and physiology (unit leaf rate, the rate of increase in plant biomass per unit LA, a variable closely related to the daily rate of photosynthesis per unit LA; also known as net assimilation rate).
What and how to measure?
Ideally, RGR is measured on a dry-mass basis for the whole plant, including roots. Growth analysis requires the destructive harvest of two or more groups of plant individuals, grown either under controlled laboratory conditions or in the field. Individuals should be acclimated to the current growth conditions. At least one initial and one final harvest should be carried out. The actual number of plants to be harvested for a reliable estimate increases with the variability in the population. Size variability can be reduced by growing a larger number of plants and selecting a priori similarly-sized individuals for the experiment, discarding the small and large individuals. Alternatively, plants can be grouped by eye in even-sized categories, with the number of plants per category equal to the number of harvests. By harvesting one plant from each category at each harvest, each harvest should include a representative sample of the total population studied. The harvest intervals may vary from less than 1 week in the case of fast-growing herbaceous species, to more than 2 months or longer in the case of juvenile individuals of slow-growing woody species. As a rule of thumb, harvest intervals should be chosen such that plants have less than doubled mass during that interval.
At harvest, the whole root system is excavated and subsequently cleaned, gently washing away the soil (see details on procedure under Section 5). Plants are divided into three functional parts, including leaves (light interception and carbon (C) uptake), stem (support and transport) and roots (water and nutrient uptake, as well as storage). The petioles can either be included in the stem fraction (reflecting support; this is the preferred option), or combined with the leaf fraction (to which they belong morphologically), or they can be measured separately. LA is measured (for details, see Section 3.1) before the different plant parts are oven-dried for at least 48 h at 70°C and weighed.
Destructive harvests provide a wealth of information, but are extremely labour-intensive and, by their nature, destroy at least a subset of the materials being studied. Alternatively or additionally, growth can be followed non-destructively for several individuals (~10–15 per treatment), by non-destructively measuring an aspect of plant size at two or more moments in time. By repeatedly measuring the same individuals, a more accurate impression of RGR can be obtained. However, RGR cannot be factorised into its components then, and repeated handling may cause growth retardation. Ideally, the whole volume of stems (and branches) is determined in woody species (see Section 4.1), or the total area of leaves, in case of herbaceous plants. In the latter case, leaf length and width are measured, along with the number of leaves. To estimate LA, a separate sample of leaves (~20) has to be used to determine the linear-regression slope of leaf length × width.
How to calculate RGR?
From two consecutive harvests at times t1 and t2, yielding plant masses M1 and M2, the average RGR is calculated as
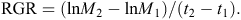
In the case of a well balanced design where plants are paired, it is probably simplest to calculate RGR and its growth parameters for each pair of plants, and then use the RGR values for each pair to average over the population. Otherwise, the average RGR over the whole group of plants is calculated from the same equation. In doing so, make sure to first ln-transform the total mass of each plant before the averaging. In the case of more than two harvests, average RGR can be derived from the linear-regression slope of ln(mass) over time.
The average unit leaf rate (ULR) over a given period is

where A1 and A2 represent the LA at t1 and t2, respectively. Again, the simplest option would be to calculate ULR from each pair of plants.
Average leaf mass fraction (LMF) during that period is the average of the values from the 1st and 2nd harvest, as follows:
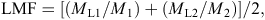
where ML1 and ML2 indicate the leaf mass at t1 and t2, respectively.
Similarly, average specific leaf area (SLA) is calculated as follows:
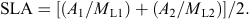
Special cases or extras
-
Confounding effect of seed size. Especially tree seedlings may draw on seed reserves for a long time after germination. Inclusion of the seed in the total plant mass underestimates the RGR of the new seedling, whereas exclusion of the seed mass will cause an overestimation of growth. For large, persistent seeds, the decrease in seed mass between t1 and t2 can therefore be added to M1 (excluding seed mass itself from M1 and M2).
-
Ontogenetic drift. As plants change over time, they readjust allocation, morphology and leaf physiology. Consequently, LMF, SLA and ULR may change with plant size, and RGR generally decreases over time, the more so in fast-growing species. This does not devalue the use of the RGR, as plant growth does not necessarily have to be strictly exponential. As long as plant growth is somehow proportional to the plant size already present, RGR is an appropriate parameter that encapsulates the average RGR over a given time period. However, ontogenetic drift is an important characteristic of plant growth, and a higher frequency of harvests may provide better insight into this phenomenon. In comparing species or treatments, it may be an option to compare plants at a given size or size interval, rather than over a given period of time.
-
Related to ontogenetic drift, shrubs and trees accumulate increasing amounts of xylem, of which large proportions may die depending on the species. This inert mass would greatly reduce RGR. Previous studies express RGR (‘relative production rate’) on the living parts of large woody plants by treating the biomass increment over Year 1 as M1 and the increment over Year 2 as M2 It could similarly be based on annual diameter (or volume) increments.
-
Smooth curves. In the case of frequent (small) harvests, a special technique can be applied, in which polynomial curves are fitted through the data. This is an art in itself!
References on theory, significance and large datasets: Evans (1972); Grime and Hunt (1975); Kitajima (1994); Cornelissen et al. (1996); Walters and Reich (1999); Poorter and Nagel (2000); Poorter and Garnier (2007); Rees et al. (2010).
More on methods: Evans (1972); Causton and Venus (1981); Hunt (1982); Poorter and Lewis (1986); Poorter and Welschen (1993); Cornelissen et al. (1996); Rees et al. (2010).
2.12 Plant flammability
Flammability-enhancing traits are important contributors to fire regimes in (periodically) dry regions and therefore they have important ecological impacts (particularly on ecosystem dynamics), as well as socioeconomic and climatic consequences. The intrinsic flammability of a plant depends on both its traits while alive and the effects of its leaves, branches, and stems after those organs’ deaths. The flammability of those organs (either living or dead) depends on (1) the type or quality of the tissue, and (2) the architecture and structure of the plant and its organs (which is mainly related to heat conductivity).
Note that the flammability of a given species can be overridden by the flammability of the entire plant community (e.g. amount of litter, community structure and continuity, organic matter content of the soil) and by the particular climatic conditions (e.g. after a long very dry period, many plants would burn quite independently of their flammability).
How to define and assess?
Flammability (broadly defined as the propensity to burn) is a compound plant functional trait. Its components vary among authors and disciplines. Most studies are based on flammability measurements of small plant fragments in chambers in the laboratory. Although this produces highly standardised results, it does not scale up well to whole-shoot flammability. We propose a standard method for measuring flammability in which the basic architectural arrangement of the measured shoots is preserved (see More on methods in the present Section for reference, and Fig. 2 for illustration) This involves a low-tech device in which shoots up to 70 cm long are placed, preheated and ignited in a standard way, then the following measurements are taken:

|
-
maximum temperature reached during burning (in °C, MT), measured with an infrared thermometer from a distance of 50 cm to the burning shoot;
-
burning rate (BR), a value obtained by dividing the length of the sample that was burnt by the burning time (in seconds); it gives an idea of how quickly flames can spread across the plant and to what extent the plant is able to carry fire; and
-
burnt biomass percentage (BB), consisting of a visual estimation of the burnt biomass (percentage intervals); the intervals are 1 = <1%, 2 = 1–10%, 3 = 11–25%, 4 = 26–50%, 5 = 51–75% and 6 = 76–100%.
For calculating (overall) flammability, the scores on each component must be transformed to a proportional scale, with the value 1 being assigned to a reference value. In the case of BB, the value 1 was assigned to the maximum possible value for this component (i.e. 6), whereas for the other components, the reference value was based on the literature and the results of our experiments. Reference values are: MT = 500°C; BR = 1 cm–1. Standardised MT, BR and BB scores are then added to obtain a compound value of flammability (rounded to two decimals) that could vary between 0 (no flammability) and ~3 (maximum flammability).
As an alternative to the direct measurement of flammability, an estimation may be obtained by measuring several plant attributes that are known to influence plant flammability. Five classes are defined for each of the attributes described below (stem and twig water content, canopy architecture, surface : volume ratios, standing litter, volatile oils, waxes and resins). Flammability is subsequently calculated as the average (rounded to one decimal) of the class scores for each of the following individual attributes (see Table 1 for ranges of values of each attribute within each class). We strongly recommend testing and calibrating this estimation against direct measurements of flammability as described above, or against direct measurements of ignitability and combustibility as described under Special cases or extras in the present Section.

|
-
Water content of branches, twigs and leaves. Flammability is expected to be greater in species with higher leaf dry-matter content (see Section 3.3 for protocol) and higher twig dry-matter content (see Section 4.2 for protocol) and it is probably also a function of drying rate (here, represented inversely by drying time from saturation to dry equilibrium).
-
Canopy architecture. Plants with complex architecture, i.e. extensive branching, tend to spread fire easily. The degree (number of orders) of ramification (branching) is used here as a close predictor of canopy architectural complexity, and ranges from zero (no branches) to 5 (four or more orders of ramification) (see Section 2.7).
-
Surface-to-volume ratios. Smaller twigs (i.e. twigs of smaller cross-sectional area) and smaller leaves should have a higher surface-to-volume ratio (and, thus, faster drying rate) and therefore be more flammable. Since twig and leaf size tend to be correlated in interspecific comparisons, according to allometric rules, we use leaf size here to represent both traits. A complication is that some species are leafless during the dry season; however, leaf litter is likely to still be around in the community and affect flammability during the dry season (see Section 3.2). However, for ground fires, substantial accumulation of litter of small leaves may pack densely, obstructing oxygen flow and actually strongly inhibiting fire to spread.
-
Standing litter. The relative amount of fine dead plant material (branches, leaves, inflorescences, bark) still attached to the plant during the dry season is critical, since litter tends to have very low water content and thus enhance plant flammability. ‘Fine’ litter means litter with diameter or thickness less than 6 mm. We define five subjective classes from no ‘fine standing litter’, via ‘substantial fine standing litter’ to ‘the entire above-ground shoot died back as one standing litter unit’.
-
Volatile oils, waxes and resins in various plant parts contribute to flammability. This is a subjective, categorical trait ranging from ‘none’ to ‘very high concentrations’. Check for aromatic (or strong, unpleasant) smells as well as sticky substances that are released on rubbing, breaking or cutting various plant parts. Scented flowers or fruits are not diagnostic for this trait.
Special cases or extras
-
Ignitability indicates how easily a plant ignites (i.e. starts to produce a flame). It can be measured directly by measuring the time required for a plant part to produce a flame when exposed to a given heat source located at a given distance. Ignitability experiments are usually performed several times (e.g. 50), and the different fuels are ranked by taking into account both the proportion of successful ignitions (ignition frequency) and the time required to produce flames (ignition delay). Tissues producing flames quickly in most of the trials are ranked as extremely ignitable, whereas tissues that rarely produce flames and/or take a long time to produce them are considered of very low ignitability. These experiments are run in the laboratory under controlled conditions (moisture and temperature) by locating a heat source (e.g. electric radiator, epiradiator, open flame) at a given distance (few centimetres) from the sample. The values used to rank species according to ignitability depend on the type and power of the heat source, on the distance of the heat source to the sample, on the shape and size of the samples and on the relative humidity of the environment in the days before the test; these experimental conditions should be kept constant for all trials and samples. We propose using an open flame at 420°C, placing the plant material at 4 cm from the flame. A standard quantity of 1 g of fresh material is used.
-
Plant tissue heat conductivity (combustibility) can be assessed by the heat content (calorific value, kJ g–1), which is a comprehensive measure of the potential thermal energy that can be released during the burning of the fuel. It is measured with an adiabatic bomb calorimeter using fuel pellets of ~1 g, and the relative humidity of the environment in the days before the test should be standardised as well. Evidence shows that heat content varies relatively little among species and is only a modest contributor to interspecific variation in flammability.
-
Combustibility and structural variables. In relation to the surface area-to-volume ratio, other structural variables have been used to characterise the combustibility, especially the proportion of biomass of different fuel classes (size distribution). Typically, the fuel classes used are the biomass fractions of (1) foliage, (2) live fine woody fuel (<6-mm diameter; sometimes subdivided in <2.5 and 2.5–6 mm), (3) dead fine woody fuel (<6 mm), and (4) coarse woody fuel (6–25, 26–75, >75 mm). The summed proportion of live and dead fine fuels (foliage and woody of <6 mm) may be the best correlate of overall surface area-to-volume ratio.
-
Fuel bulk density (= fuel weight/fuel volume) and canopy compactness (ratio of fuel volume to canopy volume) have also been used to characterise heat conductivity, mainly at the population and community levels. Furthermore, high litter fall and low decomposition rate will increase the combustibility of the community.
References on theory, significance and large datasets: Mutch (1970); Rothermel (1972); Bond and Midgley (1995); Bond and Van Wilgen (1996); Schwilk and Ackerly (2001); Gill and Zylstra (2005); Scarff and Westoby (2006); Cornwell et al. (2009); Pausas et al. (2012).
More on methods: Papió and Trabaud (1990); Stephens et al. (1994); Valette (1997); Dimitrakopoulos and Panov (2001); Etlinger and Beall (2004); Scarff and Westoby (2006); Van Altena et al. (2012); for flammability of whole shoots described above, see Jaureguiberry et al. (2011).
2.13 Water-flux traits
Plants play a key role in most hydrological fluxes in terrestrial ecosystems, including the capture of precipitation, retention and spatial (re)distribution of water above and below ground, and water loss through evaporation and transpiration. There are considerable differences among species in the extent and manner in which they influence water fluxes. These species-based differences affect not only their own growth and survival, but also influence key ecosystem processes such as growth and nutrient cycling through direct and indirect effects of moisture distribution. This section focuses on plant traits that affect hydrological fluxes external to the plants, and therefore excludes traits related to the flow of water through plants or the storage of water in plants (see Sections 3.3 and 4.4, among others). Differences in the impact on fluxes among species are mainly due to differences in plant architecture, morphology and surface features, which can all be quantified. The effects of litter and small plants on surface runoff and infiltration, and the effect of roots on below-ground movement of water through preferential pathways and altered soil hydraulics, can also be important; however, they are not further discussed here.
A large fraction of incident precipitation hits plant surfaces before reaching the ground. The fate of this water can be (1) free throughfall, (2) retention followed by evaporation or uptake into the leaf, (3) release as throughfall, as a consequence of drainage from saturated surfaces or (4) drainage via twigs and stems (stemflow). Precipitation reaches the ground as throughfall or stemflow. Rainfall interception regulates the amount of water reaching the soil (and plant roots) and may ensure a less erosive water supply during and after heavy showers. Rainfall interception is influenced by the density of the plant crown and its ability to retain water on its surfaces. Water retention on plant surfaces is greatest in low-intensity rainfall in the absence of wind. As vegetation surfaces approach saturation, further interception of precipitation leads to drainage of excess water (but see fog interception under Special cases and extras in the present Section). Plant traits that determine the actual water retention on plant surfaces in a given environment are surface angle and ‘wettability’. Hydrophobic surfaces have sharp contact angles and droplets tend to remain separate, whereas hydrophilic surfaces have small contact angles and water spreads over the surface as a film. The surface traits that most affect wettability are cuticular waxes and trichomes.
How to assess
-
Gap fraction. The openness or gap fraction of a plant’s crown is an important determinant of the free throughfall fraction (p) (as well as its light interception). For each species, we recommend taking measurements of individual plants, with minimal canopy overlap with neighbours and minimal neighbour foliage below the crown. Gap fraction can be estimated photographically. The lens has to be held neatly horizontally below the canopy and capture the entire crown but not much more. If digital pictures have enough contrast, imaging software can help calculate the fraction of sky (light) relative to foliage cover (dark) within the crown outline. If time or facilities are limited, the gap fraction will be a reasonable predictor of free throughfall. However, p is not necessarily equal to the gap fraction measured, because rain drops do not always fall strictly vertically. See Special cases and extras in the present Section for direct measurement of p.
-
Stemflow. Stemflow can be quantified by attaching collars or spirals around plant stems that channel the water running down the stems into a collector. Usually, a flexible gutter-shaped form (such as a hose cut in half) is fixed around the stem and sealed with silicone or other sealant. At the bottom of the gutter, a hose leads the captured water into a collector. Collars and collectors need to be able to handle large volumes of water because stemflow concentrates the water running off large areas of foliage. For a quantitative measure of the extent to which different plant species channel precipitation to their stems, it is important to do comparisons under similar rainfall conditions, or to use rainfall simulators. Stemflow varies with plant size and architecture, because these influence e.g. the capacity for water storage on stems, the inclination angles of stems and the extent of branching, among other things. From the perspective of plant traits, stemflow is best expressed as a percentage of the volume of rain falling on the plant, as follows:
-
Water retention on plant surfaces. This parameter can be estimated at the whole-plant level or for plant parts such as leaves and stems. The procedure involves (1) weighing the plant (or part) without surface water, (2) wetting of the plant (part) and (3) weighing the plant (part) with wet surfaces. The amount of water retained can be expressed in different ways, including water per unit plant surface area or per crown projected area. Wetting can be achieved by immersion or by simulated rainfall or fog. Immersion will tend to give maximum water retention, which is seldom reached in the field. The most realistic estimates are obtained using natural or simulated precipitation with plants in the field. Designs for rainfall or fog simulators can be found in the soil erosion and pesticide spray literature, respectively. Rainfall retention can also be estimated as total interception (rainfall minus throughfall and stemflow) of a discrete rain event that is just large enough to saturate the crown, in a situation of negligible evaporation. This provides estimates of retention where weighing is impractical or impossible.
-
Leaf wettability. How easily a leaf can get wet is determined by measuring contact angles between water droplets and the leaf surface. Droplets on water-repellent surfaces have greater contact angles and are more spherical. A droplet of standard volume (2–5 μL) is pipetted onto the leaf surface and observed from the side with a microscope. The microscope can be fitted with a goniometer for direct measurement of the angle, or images can be obtained with a camera. An alternative to the measurement of the contact angle is its calculation based on measurement of the contact area between droplet and leaf.
The sampling strategy depends on the research aims. Choose leaves randomly or select in a standardised way according to leaf position on the plant or leaf age. For most leaves, the upper (usually adaxial) surface is the logical surface for wettability measurements. For others, particularly isobilateral and needle-type leaves, as well as leaves that are often exposed to inclined rainfall, fog or mist, measurements on both sides of the leaf are recommended.
-
Droplet retention ability. How well water ‘sticks’ to a leaf can be measured by placing a droplet of water on a horizontal leaf surface and measuring the angle of inclination of the leaf at which the droplet first begins to move. It is useful to measure droplet-retention ability on the same leaves as those used for determination of leaf wettability.
Special cases or extras
-
Free throughfall (p). The parameter p can be estimated graphically as the slope of the regression line that describes the relationship between throughfall and rainfall, using precipitation events that are of insufficient size and intensity to cause drainage from these plants. If precipitation is recorded continuously, observations made immediately after commencement of large precipitation events can be used as well. For small-scale applications, rainfall simulators may be used for comparative studies. In most cases, free throughfall will be slightly overestimated because part of rainfall striking vegetation may not be retained because of the force of the impact or the movement of crowns. Methods for quantification of rainfall and throughfall are described in the hydrological literature. They typically include standard commercial rain gauges (pluviometers or pluviographs) or custom-made collectors such as funnels, troughs or large sheets. Continuous measurements of precipitation utilise devices such as tipping-bucket mechanisms that produce output for dataloggers. Variability of throughfall under plants is very large as a result of clustering of foliage and channelling towards the centre of the plant or towards the outer parts of the crown. It can be useful to take these patterns into account by sampling along radii of the plant. Representative (equal-area) sampling of concentric parts of the crown is achieved by following the rule rn = √n × r1, where rn is the distance between the nth collector and the centre of the plant.
-
Drip tips. These are morphological features that influence leaf wetness and interception by accelerating drainage from wet leaves. Water is channelled towards the long and narrow tip at the low end of a hanging leaf, which is unable to retain the accumulating water, thus reducing the duration of leaf wetness. Measurement of drip-tip length involves a decision about the position of the base of the drip tip. It is recommended that this is established by drawing the tip of a normally tapering (acute or obtuse angled) tip.
-
Water absorption by leaves. A certain fraction of water on leaf surfaces may be absorbed by the leaf. Typically, the water on wet leaves represents ~0.2-mm depth. Although most of this water will have evaporated before it can be taken up, water uptake into the leaf can be significant even in non-arid environments. Water uptake is particularly efficient in plants with specialised trichomes such as certain bromeliads. A simple method to determine rates and amounts of water uptake is by weighing.
-
Fog interception. Fog consists of small water droplets deposited on plant surfaces through air flow rather than gravity (‘horizontal precipitation’). Its interception can be a net gain because fog would not normally precipitate in the absence of vegetation that captures it. Fog interception can be particularly important to many epiphytes or to plants from dry substrates such as rocks and coastal deserts. The main plant traits affecting the rate of fog interception are the surface area in the direction of air flow (e.g. cross-sectional area of a tree crown) and the dimensions of canopy elements. Narrow structural elements with thinner boundary layers are relatively efficient in capturing fog. Trichomes (see Point 3 above in the present Section) and aerial roots, common for epiphytes, may further increase fog capture.
-
Epiphyte load, which in some cases can be species-specific but generally is not, can hugely influence water interception of rainfall and fog, and probably stemflow (increasing water retention on the host plant and modifying water fluxes). Wherever epiphyte load is important, it should be accounted for, e.g. as a covariate. Epiphyte load can be assessed as mass, as percentage cover or as counts (for large epiphytes). Then, their interception and retention properties can be quantified as described above.
References on theory, significance and large datasets: Skinner et al. (1989); Brewer et al. (1991); Puigdefábregas and Pugnaire (1999); David et al. (2005); Martorell and Ezcurra (2007).
More on methods: Aston (1979); Herwitz (1985); Veneklaas et al. (1990); Meyer (1994); Domingo et al. (1998); Brewer and Núñez (2007); Burd (2007).
3 Leaf traits
3.1 Specific leaf area
Specific leaf area (SLA) is the one-sided area of a fresh leaf, divided by its oven-dry mass. (Note that leaf mass per area (LMA), specific leaf mass (SLM) and specific leaf weight (SLW), are simply 1/SLA). SLA is frequently used in growth analysis because it is often positively related to potential RGR across species. SLA tends to scale positively with mass-based light-saturated photosynthetic rate and with leaf nitrogen (N) concentration, and negatively with leaf longevity and C investment in quantitatively important secondary compounds such as tannins or lignin. In general, species in permanently or temporarily (e.g. deserts after a rain event) resource-rich environments tend, on average, to have a higher SLA than do those in resource-poor environments, although there can also be considerable variation in SLA among co-occurring species.
Specific leaf area is a function of leaf dry-matter content (see Section 3.3), and Lth (see Section 3.4). Both components can contribute to SLA to different degrees, depending on the habitat and the plant group in question. In cool-temperate herbaceous datasets, low SLA of slow-growing species tends to be related to high leaf dry-matter content, more than to high leaf thickness. When woody perennials are dominant, Lth can be equally influential. Some species that normally grow in deeply shaded, and thus presumably resource-limited, micro-habitats (e.g. Oxalis) have a high SLA and low Lth. In areas with severe soil-nutrient limitations, slow-growing plants with sclerophyllous leaves (with thick epidermal walls and cuticle, abundant sclerification, high ratio of crude fibre to protein) are common. In these, low SLA is associated with high leaf dry-matter content more than with high Lth. In contrast, in the succulent plants that are common in some seasonally dry subtropical to tropical areas, low SLA is associated with low leaf dry-matter content and high Lth. As a consequence of these variations, SLA and its components are often, but not always, related to each other and to productivity gradients in a simple way. We then recommend additional measurement of the two component variables, namely, leaf dry-matter content and Lth, as well as SLA.
What and how to collect?
Select the relatively young (presumably more photosynthetically active) but fully expanded and hardened leaves from adult plants (unless the research is focussed on e.g. seedlings, or expanding or senescent leaves). Wherever possible, avoid leaves with obvious symptoms of pathogen or herbivore attack, or with a substantial cover of epiphylls. SLA is strongly affected by light intensity. Therefore, for many research questions it is best (giving the fairest comparison across individuals or species) to sample outer canopy leaves (also called ‘sun leaves’) from plants growing under relatively optimal conditions. For species that typically grow in the overstorey, take leaves from plant parts most exposed to direct sunlight. For true shade species (those that never grow in full sunlight), collect leaves from the least shaded parts found (but not from those that look light-stressed or bleached). Any rachis (stalk-like midrib of a compound leaf) and all veins are considered part of the leaf for a standardised SLA measurement (but see Special cases or extras below in the present Section, for a discussion on this and on whether petioles should be included in the measurement). We recommend collecting whole twig sections with the leaves still attached and not removing the leaves until just before processing.
Storing and processing
Plants in the field may be dehydrated to an unknown extent, which may lead to shrinkage of the leaves and therefore somewhat unreliable measurements of LA (see Special cases or extras below in the present Section). This is more of a problem for soft-textured high-SLA leaves than for low-SLA, sclerophyllous leaves. Ideally, the samples (twigs with leaves attached) should be cut and immediately placed into test tubes or flasks, with the cut end submerged in deionised water. If this is not feasible, wrap the samples in moist paper and put them in sealed plastic bags. In that case, breathe into the bag before closing it to enhance CO2 concentration and air humidity, which will minimise transpirational water loss, and store the bags in the dark. Tissues of some xerophytic species (e.g. bromeliads, cacti, some species with very small, highly resinous leaves) rot very quickly when moist and warm; therefore, they are better stored dry in paper bags. If in doubt (e.g. in mildly succulent species), and if recollecting would be difficult, try both moist and dry storage simultaneously and use the dry-stored leaves in the case of rotting of the moist-stored ones. Store the collected samples in a cool box or fridge (never in a freezer) in the dark, until further processing in the laboratory. If no cool box is available and temperatures are high, it is better to store the samples in plastic bags without any additional moisture. Measure as soon as possible after collecting and rehydrating, preferably within 24 h. If storage is to last for more than 24 h, low temperatures (2–6°C) are essential to avoid rotting.
Rehydration is preferable for most plants and essential for leaves subjected to dry storage or for ‘soft’ leaves, such as those with SLA values higher than 10–15 m2 kg–1. In situations where the rehydration procedure described above cannot be applied, storage in sealed, moist plastic bags (with or without addition of damp paper) for 12 h is an acceptable option, although generally yields approximately ~5% lower values than does the complete rehydration method. Xerophytic and especially succulent leaves should not be rehydrated for more than 6 h, whatever the storage or rehydration method used might be. If this process fails, we recommend collecting leaves of these species in the morning after a rain event, or a few hours after generous watering.
Measuring
Each leaf (including or excluding petiole, see Special cases and extras below in the present Section) is cut from the stem and gently patted dry before measurement. Projected area (as in a photograph) can be measured with specialised leaf-area meters such as those from Delta-T Devices (Cambridge, UK) or LI-COR (Lincoln, NE, USA). Always calibrate the area meter by using pieces of known area before measuring leaves and always check (e.g. on the monitor) that the whole leaf is positioned flat and completely within the scanning area. If you are to use a portable LA meter, make sure that the estimation error is not too high for your purposes, by running a preliminary check against LAs scanned in the laboratory, using a range of different leaf sizes. Images of leaves can also be electronically captured in the field or in the laboratory, and stored for later processing, e.g. with a digital camera. Leaves are pressed gently under a glass plate. Including a ruler or an object of known size in the image allows for size calibration. A camera mounted on a tripod, with two lamps lighting from different sides and no flash gives the best results. A third option is to determine LA with a flatbed scanner (with the advantage that many scanners can draw their power via USB, e.g. from a laptop). Scan in colour mode to obtain maximal information for the threshold level between what is to be considered leaf and background Coloured scans will also allow for post hoc measurement of other features of interest. In all cases, make sure the leaves are not curled-up or overlapping. Try to position the leaves as flat as possible in the position that gives the largest area, but without squashing them to the extent that the tissue might get damaged. Cutting leaves into smaller pieces may facilitate flattening. In both cases, LA can be measured with image analysis software. Freely downloadable programs are e.g. Leafarea (A. P. Askew, University of Sheffield, UK, downloadable from the Nucleo DiverSus toolbox, see Box 1) or, for more complex analyses including other plant organs, ImageJ (from the US National Institutes of Health; http://www.nih.gov/, accessed 22 February 2013) and GIMP (from the GNU Project; http://www.gnu.org/, accessed 22 February 2013). Transform images to the HSV colourspace for a better separation of leaf and background. Additional details on image processing can be found at Prometheus Wiki (see Box 1).
After the area measurement, put each leaf sample in the oven (petioles and laminae either separately or in the same envelope, according to the objective of the study, see Special cases or extras below in the present Section), ideally at 70°C for at least 72 h, or at 80°C for 48 h (avoid higher temperatures); then determine the dry mass. Be aware that, once taken from the oven, the samples will take up some moisture from the air. Put them therefore in a desiccator with silica gel until weighing, or else back in the oven to re-dry. Weighing several tiny leaves as if they were one and then dividing the weight by the number of leaves will generally improve the accuracy of the weighing. When converting SLA into LMA values or vice versa, always do so for each individual replicate, rather than for the average of several replicates.
Special cases or extras
-
Petioles. An important issue is whether or not petioles should be included in SLA measurements. The appropriate decision depends on the research question at hand. Some authors consider that the petiole is an integral part of the leaf because it is shed at abscission together with the leaf, and because it provides support and a vascular system without which the leaf cannot be displayed. Therefore, they include petioles in SLA measurements. Other authors consider that the petiole should not be included in the SLA because the main function of the petiole is the spatial positioning and hydraulic support of the leaf, thus resembling the function of the stem, whereas the main function of the leaf blade is light interception and C fixation. The fraction of leaf dry mass represented by the petiole varies from ~zero to almost 50%; therefore, inclusion of the petiole may reduce the calculated SLA drastically. Although inclusion or not of the petiole may sometimes not be crucial within a single study, it can be a source of considerable and systematic error when comparing different studies, or even in certain same-site comparisons of species with very different leaf structures. Therefore, the best (albeit more time-consuming) option is to measure leaf blade and petioles separately, so that SLA can be calculated in both ways, thereby facilitating comparisons with other studies. When using digital images, we suggest to scan or photograph petioles and the rest of the leaves of each replicate in the same image, but in clearly different sectors, so that they can be measured together or separately according to the objectives of the study; then oven-dry and weigh petioles belonging to a replicate separately from the rest of the leaves from the same replicate. In general, make the decision that best suits your study objectives; however, specify in your publication whether petioles are included or not.
-
Compound leaves. Sometimes species have a lower SLA because of their thick rachis. The decision of whether to include the rachis or not, and the practical steps in each case, are similar to those on petioles (see Point 1 above in the present Section). As a default option, we recommend considering the rachis as part of the leaf for the SLA calculation, and indicating this clearly when reporting the study. Be aware than in some plants, the rachis can be more than 10 times heavier that the sum of the leaflets. Another decision to make in the case of compound leaves is whether to measure the SLA of a typical individual leaflet or that of all leaflets taken together. As in the case of rachis and petiole, the decision largely depends on the objectives of the study. We recommend taking both measurements, and above all, we recommend to clearly specify in the publication whether the area reported is that of an individual leaflet or the whole leaf.
-
LA shrinkage. Leaves decrease in size when they desiccate. Shrinkage is defined as a percentage, calculated as 100 × (1 – dry area/saturated area). Shrinkage averages ~20% and can reach 80% for some species. The maximum area shrinkage reflects differences in leaf structure and is correlated with other leaf traits including cuticular conductance, pressure–volume parameters such as modulus of elasticity and turgor loss point, leaf dry-matter content (LDMC), and Lth, as well as plant growth form and deciduousness. LA shrinkage is both a potential problem (causing biases in leaf-area measurements for herbarium material and fossils) and an easily measured trait that reflects multiple structural and hydraulic properties of a leaf. To measure this trait, collect and measure leaves following the exact same protocol as for LA (Section 3.2) and LDMC (Section 3.3). Measure fresh projected LA. Cut leaf into pieces in the cases when the leaf is not flat. This step is crucial because some leaves may experience very little shrinkage in area (5%), so the accuracy of the initial measurement of the saturated area is crucial. Then oven-dry the leaf, pressed flat in envelopes or in a plant press. Measure projected LA on the dried leaf, taking care that the same surface is measured. This may be difficult or impossible for some needle-like leaves, and especially thick leaves may need to be broken into several smaller flat pieces. Calculate shrinkage using the formula given above.
-
Projected v. total LA. In ‘standard’ leaves, LA is measured as a one-sided projected LA. However, in non-flat leaves, the projected LA is smaller than the total one-sided LA. Projected LA is generally related to light interception, whereas total LA is related to the total amount of photosynthetically active tissue. There are, however, cases, such as leaf-rolling in some grass species, where both light interception and gas exchange are reduced. Clearly, the choice of measurement depends on the research question, although knowledge of both would provide the best insight.
-
Needle-like leaves. Needle-like leaves are a specific case where projected and total LAs are different. Projected LA could be measured following the standard routines; however, because the leaves are generally narrow, make sure that your equipment is sensitive enough to adequately measure such leaves. For a rough measurement, you can measure leaf length with a ruler and leaf width with a calliper and subsequently multiply 2 × length × width.
-
Tiny leaves. True leaves from some species (e.g. Callitris sp.) have very tiny scales closely appressed to fine soft twigs. In such cases, you might treat the terminal twiglets as a leaf analogue, because they are shed as a single unit.
-
Leaves of grasses and grass-like plants. Usually, only the lamina is considered, excluding the leaf sheath. However, as in the case of petioles (see Point 1 above in the present Section), the decision on which measurement to take depends on the research objectives. Several species have leaves that tend to curl, or even roll up. They are generally much easier managed by cutting leaves into shorter pieces of 5–10 cm.
-
Succulent and leafless plants. For plants whose main photosynthetic organs are not true leaves, take the plant part that is the functional analogue of a leaf and treat as above. For some species with photosynthetic spines or non-succulent stems (e.g. Ulex, Senna aphylla), this could mean taking the top 2 cm of a young twig. For cacti and other succulents, we recommend taking a whole leaf or equivalent (e.g. a cladode in Opuntia) whenever possible. This sometimes poses practical difficulties, e.g. a whole Agave leaf or a rib of a columnar cactus are often too big to process or even to collect. In such cases, we recommend taking several ‘pastry cutter’ portions (of known area) of young but fully hardened leaves (e.g. in Agave) or ‘ribs’ (in cacti), including epidermis and mesophyll on both sides, plus the internal succulent parenchyma. Although this internal parenchyma does not always contain chlorophyll and therefore some authors recommend not considering it in SLA measurements, it has an essential role in the CAM metabolism of succulent plants (see also Section 3.12). The younger stems of some rushes and sedges (Eleocharis, Juncus) and the ‘branches’ of horsetails (Equisetum) or similar green leafless shoots can be treated as leaves too. Because many of these species occur in a range of environments, it is important to specify the exact method used in each case.
-
Ferns. For ferns, only collect fronds (fern ‘leaves’) without the spore-producing sori, often seen as green or brown structures of various shapes at the lower side or margin of the frond.
-
Leaves of tall trees. Upper-canopy leaves of sun-exposed trees should be preferred. If these cannot be easily reached, some workers rely on professional climbers, slingshots or guns. In not extremely tall trees, an alternative could be to consider exposed leaves halfway the crown length, at the outer half of the crown (inner leaves are sometimes older), which could be accessed with a pruner on an extension pole.
-
Very large leaves. Once they have been placed in plastic bags, large leaves may be put in a hard-cover folder to avoid wrinkling and folding. If leaves are larger than the window of the area meter, cut the leaf up into smaller parts and measure the cumulative area of all parts. Leaves with very thick veins or rachis can cast a lateral shade on the LA meter, thus overestimating the LA. In the case of a thick central vein, remove with a scissor the protruding upper or lower part of the vein and scan the leaves without that removed part, but include it in the dry-mass measurement. In the case of a thick rachis, remove the rachis and measure its diameter and length halfway, and calculate the rachis area as the product of the two. Then scan the leaves without rachis but include the rachis in the dry mass. If you want to rely on subsamples of leaves, make sure that there is not too large of a variation in SLA over the leaf.
-
Heterophyllous plants. In the case of species with two or more types of leaves with contrasting shape and anatomy, such as e.g. plants with both rosette and stem leaves, collect leaves of both types in proportion to their estimated contribution to total LA of the plant, so as to obtain a representative SLA value of the individual.
-
Low-tech options for the measurement of SLA and LA in the field. There are situations in which taking fresh leaves to the laboratory for scanning is not feasible, or portable scanning devices cannot be transported to or powered in the field site. One solution in these cases is to use a digital camera (see Measuring above in the present Section). Another practical and inexpensive alternative is obtaining leaf fragments of known area (e.g. with a punch-borer), avoiding thick veins, and placing the fragments in an envelope for drying (take several punches per replicate, because they tend to weigh very little). This method is a quick and accurate way to compare leaf laminae. However, it overestimates SLA as compared with measurements of whole leaves, especially in the case of large leaves with thick veins, ribs and petioles. Therefore, SLA measurements obtained in this way should not be compared with or combined with those taken on whole leaves, at least not without a specific calibration. Another alternative is to obtain plastic or paper prints or cut-outs of the leaves in the field, and measure their area later. This method works well for medium- to large-sized leaves, entire leaves, and leaves that are not too narrow (e.g. xerophytic grasses).
References on theory, significance and large datasets: Reich et al. (1992, 1999); Garnier and Laurent (1994); Poorter and Garnier (1999); Wilson et al. (1999); Castro-Díez et al. (2000); Niinemets (2001); Westoby et al. (2002); Díaz et al. (2004); Paula and Pausas (2006); Wright et al. (2007); Poorter et al. (2009); Hodgson et al. (2011); Blonder et al. (2012); Juneau and Tarasoff (2012).
More on methods: Chen and Black (1992); Garnier et al. (2001a, 2001b); Vendramini et al. (2002); Vile et al. (2005); Niinemets et al. (2007).
3.2 Area of a leaf
The area of a leaf (also called leaf area, LA) is the most common metric for leaf size and is defined as the one-sided or projected area of an individual leaf, expressed in mm2 (see Section 3.1). Interspecific variation in LA has been variously related to climatic variation, geology, altitude and latitude, where heat stress, cold stress, drought stress, nutrient stress and high-radiation stress all tend to select for relatively small leaves. Within climatic zones, variation in the LA may also be linked to allometric factors (plant size, twig size, anatomy and architecture, leaf number, number of lateral buds produced) and ecological strategy with respect to environmental nutrient stress and disturbances, and phylogenetic factors can also play an important role.
What and how to collect?
For the leaf-collecting protocol, see under Section 3.1 LA is rather variable within plants and we recommend collecting a large number of replicates (i.e. close to the higher end of the number of replicates recommended in Appendix 1). For storing leaves, see Section 3.1.
Measuring
Measure the individual leaf lamina for species with simple leaves. For compound-leaved species, either the leaflet area or the whole LA can be measured, and the appropriate decision depends on the research question at hand. For the heat balance, the leaflet area is important, which is functionally analogous to a simple leaf. When analysing total light capture, the whole leaf should be measured. Ideally, determine for compound-leaved species both the leaflet area and whole LA, because this allows one to address more questions and to compare the results with other studies. Measure the laminae with or without petiole and rachis, according to the objectives of your study (see Section 3.1), and always report this in your publication. Note that this whole LA may be different from the area used to determine SLA.
Special cases or extras
-
Leafless plants. Because leaflessness is an important functional trait, record LA as zero for leafless species (not as a missing value). However, be aware that these zeros may need to be excluded from certain data analyses. Alternatively, sample leaf analogues (see Succulent and leafless plants in Section 3.1).
-
Heterophyllous plants. See Section 3.1.
-
Ferns. See Section 3.1.
-
Leaf width. This is measured as the maximum diameter of an imaginary circle that can be fitted anywhere within a leaf, and is an additional trait of ecological interest related to leaf size. Narrow leaves, or divided leaves with narrow lobes, tend to have a smaller boundary layer and a more effective heat loss than do broad leaves with the same area. This is considered adaptive in warm, sun-exposed environments. There is also emerging evidence that leaf width contributes more positively than does the area of the whole leaf to the expression of canopy dominance.
-
Leaf number per node. Leaf size is a compromise between functional and resource-use efficiency. Plants are modular in construction and, as a result, these functions can be partially uncoupled. Species with alternate, opposite and whorled leaves frequently co-exist and leaf dry mass or area multiplied by the number of leaves per node provides additionally a crude estimate of the size of each growth module. This may in extreme cases be 10 times the value of a single leaf.
References on theory, significance and large datasets: Raunkiaer (1934); Parkhurst and Loucks (1972); Givnish (1987); Cornelissen (1999); Ackerly et al. (2002); Westoby et al. (2002); Milla and Reich (2007); Niinemets et al. (2007); Niklas et al. (2007); Poorter and Rozendaal (2008); Royer et al. (2008).
More on methods: see references in More on methods of Section 3.1.
3.3 Leaf dry-matter content
Leaf dry-matter content (LDMC) is the oven-dry mass (mg) of a leaf, divided by its water-saturated fresh mass (g), expressed in mg g–1. Using these units, LWC is simply 1000-LDMC. LDMC is related to the average density (fresh mass per fresh volume) of the leaf tissues. In laminar leaves, it is related to SLA by a formal relationship involving Lth and the average density of the leaf (ρF), as follows: LDMC = 1/(ρF × SLA × Lth). Assuming that the fresh mass per volume of leaves is close to 1 g cm–3, the equation simplifies to LDMC ≈ 1/(SLA × Lth). LDMC, therefore, tends to be inversely related to SLA and Lth. LDMC has been shown to correlate negatively with potential RGR and positively with leaf lifespan; however, the strengths of these relationships are usually weaker than those involving SLA. Litter derived from leaves with high LDMC also tends to decompose more slowly than that from leaves with low LDMC. Leaves with high LDMC tend to be relatively tough (see Section 3.7), and are thus assumed to be more resistant to physical hazards (e.g. herbivory, wind, hail) than are leaves with low LDMC. Some aspects of leaf water relations and flammability (see Section 2.12) also depend on LDMC. Commonly, but not always, species with low LDMC tend to be associated with productive, often highly disturbed environments. In cases where SLA is difficult to measure (see Section 3.1), LDMC may give more meaningful results, although the two traits may not capture the same functions (this is particularly obvious in some groups; e.g. succulents have slow growth, low SLA and low LDMC; see also Section 3.1).
What and how to collect?
Follow exactly the same procedure as for Section 3.1 In many cases, the same leaves will be used for the determination of both SLA and LDMC. As is the case for SLA, LDMC may vary substantially during the day.
Storing and processing
Similarly as for SLA, except that any dry storage should be avoided (however, see the case of xerophytic species in Section 3.1), and that full rehydration before measurement is compulsory.
Measuring
Following the rehydration procedure, the leaves are cut from the stem and gently blotted dry with tissue paper to remove any surface water before measuring water-saturated fresh mass. Each leaf sample is then dried in an oven (see Section 3.1), and its dry mass subsequently determined.
Special cases or extras
Most comments for SLA also apply to LDMC.
References on theory, significance and large datasets: Eliáš (1985); Witkowski and Lamont (1991); Garnier and Laurent (1994); Hodgson et al. (1999, 2011); Wilson et al. (1999); Garnier et al. (2001a); Niinemets (2001); Vile et al. (2005); Kazakou et al. (2009); Poorter et al. (2009).
More on methods: Wilson et al. (1999); Garnier et al. (2001b); Vendramini et al. (2002); Vaieretti et al. (2007); Ryser et al. (2008).
3.4 Leaf thickness
Leaf thickness (Lth, µm or mm) is one of the key components of SLA (see Sections 3.1 and 3.3), because SLA ≈ 1/(tissue density × Lth) (where density = dry mass/volume ≈ LDMC; see Section 3.3). Lth also plays a key role in determining the physical strength of leaves (see Section 3.7). For example, leaf ‘work to shear’ is (by definition) the product of Lth and tissue toughness. Optimisation theory, balancing photosynthetic benefits against C costs of respiration and transpiration, predicts that Lth should be higher in sunnier, drier and less fertile habitats, as well as in longer-lived leaves. These patterns are indeed often observed, at least in interspecific studies. Within individuals, many studies have shown that outer-canopy ‘sun’ leaves tend to be thicker than those from more-shaded parts of the canopy. Both within and among species, the strongest anatomical driver of variation in Lth is the number and thickness of mesophyll layers. Consequently, Lth is a strong driver of leaf N per area. Although higher Lth should lead to faster photosynthetic rates per unit LA (via a higher N : area ratio), this relationship is often weak in interspecific studies, for a combination of reasons. First, because of covariance of SLA and %N, thicker leaves often have lower %N and longer leaf-lifespan (which are associated with lower photosynthetic rate per unit leaf mass). Second, thicker-leaved species may have slower CO2 diffusion (lower mesophyll conductance) via longer diffusion pathways, greater internal self-shading of chloroplasts, or higher optical reflectivity in combination with lower internal transmittance. Thick leaves are also a feature of succulents.
What and how to collect?
Follow similar procedures as for Section 3.1 In many cases, the same leaves will be used for the determination of SLA, Lth and LDMC (and perhaps Section 3.7). For recommended sample size, see Appendix 1.
Storing and processing
Similarly as for SLA. Lth is strongly affected by LWC; hence, some form of rehydration should be seriously considered, as described for SLA, particularly if using a digital micrometer, where any slight loss of turgor results in an underestimation.
Measuring
Thickness tends to vary over the surface of the leaf, generally being thickest at the midrib, primary veins, margins and leaf base. Depending on the research question, you may be interested in the average thickness across the leaf, or the thickness at special locations or of special tissues. Often one measurement per leaf, at a position as standard as possible within the lamina (e.g. at an intermediate position between the border and the midrib, and between the tip and the base of the leaf, avoiding important secondary veins) is acceptable for broad interspecific comparisons. When more precision is needed, the average of thickness measurements at several points in the lamina will be more appropriate. Another way to estimate the average thickness over the entire leaf surface is to back-calculate it from the leaf volume divided by LA; however, it is laborious to accurately measure leaf volume, e.g. with a pycnometer. A relatively fast approximation of whole-leaf average Lth can be obtained by dividing leaf fresh mass by LA (which is the same as calculating 1/SLA × LDMC), i.e. by assuming that leaf fresh mass and volume are tightly related. This approach does not take into account the higher density of dry material in the leaf, or the lower density as a result of intercellular spaces; however, as an approximation it works well.
Other approaches are needed if one wants to distinguish between thickness of midrib, margin and intercostal regions of the leaf, or to compare replicates at a given point on the leaf, e.g. half-way between the leaf base and the tip, as is commonly done. One method is to measure these quantities directly from leaf cross-sections (hand-sections), or to use image analysis (see Section 3.1 for free software) to calculate average Lth across the section, by dividing the total cross-sectional area by the section width. On the positive side, this method enables reasonably accurate measurements to be made. On the down side, soft tissue may distort when hand-sectioned, and the method is relatively slow (e.g. 15 min per measurement).
Probably the fastest approach is to measure Lth using a dial-gauge or a digital micrometer (or even a linear variable displacement transducer; LVDT). Multiple measurements can be made within quick succession and averaged to give an indicative value of Lth for the feature in question (such as e.g. midrib or lamina between the main veins) or region of interest (e.g. near midpoint of leaf). If necessary, we recommend replacing the original contact points on the micrometer with contacts 2–3 mm in diameter; i.e. narrow enough to fit between major veins, but sufficiently broad so as not to dent the leaf surface when making measurements. However, for soft-leaved species such as Arabidopsis, permanent deformation is difficult to avoid.
Special cases or extras
-
Needle leaves. For needle leaves that are circular in cross-section, average Lth can be quickly estimated as Diameter × π/4 (equivalent to cross-sectional area divided by cross-section width). Still, because needle leaves typically taper towards the leaf tip, several measurements would normally need to be made.
References on theory, significance and large datasets: Clements (1905); Givnish (1979); Parkhurst (1994); Enríquez et al. (1996); Knapp and Carter (1998); Smith et al. (1998); Wilson et al. (1999); Green and Kruger (2001); Niinemets (2001); Díaz et al. (2004).
More on methods: Witkowski and Lamont (1991); Garnier and Laurent (1994); Shipley (1995); Wright and Westoby (2002); Vile et al. (2005); Poorter et al. (2009); Hodgson et al. (2011).
3.5 pH of green leaves or leaf litter
The pH of green leaf tissue and of senesced leaves (leaf litter), measured by grinding up the tissue and extracting it with distilled water, varies substantially among species. The variation is at least partly intrinsic (presumably genetic) because this pH can differ greatly among different species growing in the same soil (and also during the day in CAM plants, see below under Special cases or extras in the present Section) and is robust to differences in soil chemistry (including pH). Ground leaf-tissue pH integrates the effects of many compounds and processes in the leaf that affect its exchange capacity for H+ ions. However, some substances are particularly strong determinants of leaf-tissue pH. High concentrations of metal cations (calcium, magnesium, potassium) will give high pH, whereas high concentrations of organic acids and of C-rich secondary metabolites (chemical-defence compounds) such as tannins, will tend to give a lower pH. The latter may explain why leaf-tissue pH tends to be correlated (negatively) with important biogeochemical traits such as C concentration or C : N ratio, and (positively) with SLA (Section 3.1). Green leaf-tissue pH correlates positively with digestibility, making it an important predictor of palatability to herbivores, which may actually ‘taste’ acidity. Differences in leaf-tissue pH among species tend to persist during senescence, making leaf-litter pH a worthwhile trait also. It can be a reasonable proxy for litter decomposability, because substrate pH can be important to decomposers. Shifts in species composition, among species that differ in leaf-litter pH, can drive changes in the pH of the litter layer and of the soil’s organic horizon. For example, planting pines on high-pH soils dominated by bacteria can, via the leaf litter, lead to more acidic soil organic matter within decades, and a decomposer community dominated by fungi. Whether green leaves or senesced leaves, or both, should be measured depends on the question to be addressed; however, green leaves should be used if in doubt and if decomposition is not the main issue.
Physiological caveat
The cells of any plant tissue consist of three or more membrane-delimited compartments that can, and usually do, have different internal pH values. Therefore, the pH of the type of crude, water-extracted homogenate or ground material used for this method, although it can be useful as an ecological trait, must not be mistaken for the actual pH of the leaf as a whole or any part thereof. It will, at best, be a weighted average of the pH values of the various compartments of the leaf, possibly modified by reactions that occur between components of different compartments, including vacuoles, when they are mixed together. If leaves are oven-dried before grinding, loss of compartmentalisation under conditions favouring pH-modifying reactions will have occurred long before the tissue is ground. Still, the pH measured this way has been shown to also represent the pH of green leaves. Because, in leaf litter, metabolic processes have already stopped, unlike in living leaves, litter pH values should reflect the actual situation in the litter at the time of sampling.
What and how to collect
For green leaves, see Section 3.1 for the collecting and storing procedure before processing. If leaves are small, make sure enough of them are collected so as to get enough material for the analysis. Initial leaf rehydration is not necessary.
Processing and storing
As a default option, any petiole or rachis should be removed before pH analysis; however, see Special cases or extras in Section 3.1 Fresh green leaves can simply be ground or chopped as noted below. Leaves collected for leaf SLA analysis can instead be used, after being oven-dried to obtain their biomass (see Section 3.1), and subsequently stored air-dry, because pH values obtained from oven-dried leaves are largely comparable to those from fresh leaves. Air-dried leaves or leaf litter are ground as for leaf N analysis (see Section 3.6) and stored air-dry until the analysis.
Measuring
Add distilled or de-mineralised water to the ground leaf sample, to give an 8 : 1 volume ratio of water to leaf sample. Shake the samples in a laboratory rotary shaker for 1 h, then centrifuge until there is a clear separation of the sediment and the supernatant. The supernatant can then be measured for pH by using any of a wide range of laboratory pH meters, as long as calibration is adequate (using buffer solutions of pH 4 and pH 7). If samples are small in volume, we recommend adding 1.2 mL of water to 0.15 mL of ground leaf material in a 2.5-mL Eppendorf tube, then following the above procedure. A thin SenTix 41 electrode connected to an Inolab level 2 pH meter (both WTW, Weilheim, Germany) fits nicely inside such Eppendorf tubes.
Special cases or extras
-
Additional measurements on fresh leaves. Although measurements on dried, ground leaves tend to match those on fresh leaves fairly well, for various specific purposes, it may be of interest to (additionally) measure fresh leaves, the cell contents of which have remained intact until shortly before measurement. For instance, this may be useful to follow diurnal changes in leaf pH to indicate possible CAM (see Section 3.12). Related to this, in comparisons of CAM plants with other plants for leaf pH, leaves of CAM plants are best collected in the afternoon, i.e. well after nocturnal acid accumulation. Grinding fresh leaves does not always work well because solids may stick to the surfaces or to the balls in a ball mill, making cleaning between samples laborious. Instead, we recommend chopping the leaf sample into ~1-mm-diameter pieces with a razor blade or an automatic chopper that gives comparable fragmentation. This should be carried out immediately before shaking and centrifuging.
References on theory, significance and large datasets: Zinke (1962); Marschner (2012); Finzi et al. (1998); Cornelissen et al. (2006, 2011); Freschet et al. (2010).
More on methods: Cornelissen et al. (2006, 2011).
3.6 Leaf nitrogen (N) concentration and leaf phosphorous (P) concentration
Leaf N concentration (LNC) and leaf P concentration (LPC) are the total amounts of N and P, respectively, per unit of dry leaf mass, expressed in mg g–1 (or sometimes as % dry-leaf mass). Interspecific rankings of LNC and LPC are often correlated. Across species, LNC tends to be closely correlated with mass-based maximum photosynthetic rate and with SLA. High LNC or LPC are generally associated with high nutritional quality to the consumers in food webs. However, LNC and LPC of a given species tend to vary significantly with the N and P availability in their environments. The LNC : LPC (N : P) ratio is sometimes used as a tool to assess whether the availability of N or P is more limiting for plant growth. Actively N-fixing species, e.g. many legumes, tend to have higher LNC : LPC ratios than other plants growing at the same site.
What and how to collect?
See Section 3.1 for the leaf-collecting procedures. Initial rehydration is not necessary. Petiole or rachis are often cut off before LNC and LPC analysis, but are included in some other cases. See under Section 3.1 for discussion of when and whether to consider them. Oven dry at 60−70°C for 72 h. Leaves used for LA or SLA analysis can be also used to measure LNC and LPC, provided drying temperature has not been higher than 70°C. For replication, see under Section 3.1, but make sure that enough total leaf material per replicate is collected, according to the analytical method and equipment to be applied (~2 g dry matter per replicate for N and 5 g for P in the case of acid digestion, 0.2 g for N in the case of combustion techniques, see below under the present Section).
Storing and processing
After oven-drying the leaves, store the material air-dry and in the dark until use, to a maximum of 1 year. Grind each replicate separately. Manual grinding with mortar and pestle is an option for small numbers of samples, but is not recommended for large ones (repetitive strain injury). Effective, inexpensive mechanical grinders are available. Samples may also be ground by shaking them with steel balls in individual plastic vials on a roller mill, which is an efficient way to grind many samples at once. Avoid inter-sample contamination by cleaning the grinder or steel balls carefully between samples. Use a ball mill for small samples. Dry the ground samples again for at least 12 h before analysis.
Measuring
Several techniques are available to measure LNC and LPC in ground plant material. Macro- or micro-Kjeldahl (acidic) digestion, followed by colorimetric (flow-injection) analysis (using different reagents for N and P), has been widely used. Wet acidic digestion, followed by formation of blue phosphomolybdenum complex from orthophosphate is a more precise method for measuring total P. Alternatively, you can measure P by inductively coupled plasma–optical emission spectroscopy (ICP–OES). Kjeldahl digestion for N analysis is increasingly being replaced by methods that employ a combination of combustion analysis, converting organic matter into N2 and CO2, followed by mass spectrometry or gas chromatography. These combustion techniques provide concentrations of both N and C in the leaf, and if carried out with automated N analysers, are generally less labour- and chemical-intensive than are Kjeldahl analyses. Combustion techniques also generally recover more N than do Kjeldahl analyses, because some N fractions (e.g. NO2–, NO3– and some cyclic N compounds) do not react in Kjeldahl analysis. However, we believe that all of these standard methods should give reasonably accurate LNC and LPC. We recommend running a standard reference material with known LNC and LPC along with the samples.
Special cases or extras
-
Leafless and heterophyllous plants. See Section 3.1.
References on theory, significance and large datasets: Chapin (1980); Field and Mooney (1986); Lambers and Poorter (1992); Aerts (1996); Cornelissen et al. (1997); Grime et al. (1997); Reich et al. (1997, 2010); Aerts and Chapin (1999); Wright et al. (2004).
More on methods: Allen (1989); Anderson and Ingram (1993); Horneck and Miller (1998); Temminghoff and Houba (2004).
3.7 Physical strength of leaves
Physically stronger leaves are better protected against abiotic (e.g. wind, hail) and biotic (e.g. herbivory, trampling) mechanical damage, contributing to longer leaf lifespans. Physical investment in leaf strength is a good indicator of C investment in structural protection of the photosynthetic tissues. It also tends to have afterlife effects in the form of poor litter quality for decomposition. Because leaves have different strength properties according to the direction in which the force is applied, the physical strength of the leaves can be defined and measured in different ways. The three most common measured properties are force to tear (Ft), work to shear (Ws) and force to punch (Fp). Ft is the force needed to tear a leaf or leaf fragment, divided by its width, expressed in N mm–1. Note that Ft has been previously referred to as ‘leaf tensile strength’. Ws, sometimes called ‘force of fracture’, reflects the mean force needed to cut a leaf or leaf fragment at a constant angle and speed, expressed in N, or its analogue, J m–1. Fp is the force needed to force a punch through a leaf or leaf fragment (expressed in N mm–1). Both Ws and Fp are strongly influenced by Lth (see Section 3.4).
What and how to collect?
For the selection and collecting procedure, see Section 3.1; for recommended sample size, see Appendix 1.
Storing and processing
Follow the procedure described within Section 3.1 and store leaves in a cool box or fridge. Rehydration is not indispensable, although it may be desirable for accurate measurement of Lth. Measure as soon as possible after collecting, certainly within 72 h for species with soft leaves. Tougher leaves tend to keep their strength for several days. If this is not possible (e.g. if samples have to be sent to distant locations), an alternative is to air-dry the samples, by putting them between sheets of absorbing paper in a plant press, immediately after collecting. In this case, rehydration is needed before measuring. Toughness of fresh and rehydrated leaves is well correlated for sclerophyllous leaves and grass leaves, in the cases of both Ft and Ws.
Measuring
For fresh samples, proceed to measurement straight away. For air-dried samples, first rehydrate by wrapping in moist paper and put in a sealed plastic bag in the fridge for 24 h (gentle spraying may be better for some xerophytic, rotting-sensitive species; see Section 3.1). Here, we describe three methods that have produced good results and for which purpose-built equipment is available. If you have the choice, we recommend measuring the property that is most closely related to the process of interest. In the case of herbivory by vertebrate grazers, Ft is likely to be the most meaningful property. However, if the focus is on chewing insects or trampling by mammals, work to shear tests would be the best approach (Table 2).

|
-
Tearing (tensile) tests
Force to tear (Ft) can be easily and inexpensively measured with a simple apparatus that includes a 0–3-kg-range dynamometer (Fig. 3a). To proceed, cut a leaf fragment from the central section of the leaf but away from the midrib (central vein), unless the latter is not obvious (e.g. some Poaceae or Liliaceae), or the leaf is too small for doing so without using a magnifying lens. The length of the fragment follows the longitudinal axis (direction of main veins). The width of the leaf or leaf fragment depends on the tensile strength and tends to vary between 1 mm (extremely tough species) and 10 mm (very tender species). Whenever possible, we recommend using fragments with a length : width ratio between 5 and 10, to make sure that force is applied along its main axis. Measure the exact width of the leaf sample before placing it in the apparatus. Place it perpendicular to the edges of the clamps. Then fix both ends of the sample with the clamps. Try to do this gently without damaging the tissues. A thin piece of rubber added to the edges of the clamps could help. Very small leaves, very tough and slippery leaves, and slightly succulent leaves may be clamped tightly without much tissue damage, by using strong double-sided tape. Then pull slowly, with increasing force, until the leaf tears. Watch the dynamometer to read the force at the moment of tearing. For unit conversion, remember that 1 kg = 9.81 N. Divide the total force by the width of the leaf fragment to obtain Ft. Express the result in N mm–1. There are some more sophisticated instruments to measure Ft, such as the 5542 (Instron, Canton, MA, USA) or (with adaptations) Mecmesin Ultra Test Tensiometer (Mecmesin, Slinfold, UK).

|
Leaves too tender to provide an actual measurement with the apparatus are assigned an arbitrary tensile strength of zero. For leaves too tough to be torn, first try a narrower sample (down to 1 mm if necessary and possible). If still too tough, then tensile strength equals the maximum possible value in apparatus (assuming sample width of 1 mm). In the case of highly succulent leaves (or modified stems), which would be squashed if clamped into the apparatus, carry out the measurements on epidermis fragments.
-
Shearing (cutting) tests
At least five instruments have been used to measure work to shear (Ws). They all measure how much work is required to cut a leaf, either with a single blade against an anvil, or with a pair of blades (‘instrumented scissors’). A portable and widely used device for measuring the average force needed to fracture a leaf at a constant shearing angle (20°) and speed is detailed in Fig. 3b. A non-portable apparatus for this measurement is showed in Fig. 3c. In both cases, the apparatus consists of a mechanical portion, a power source and a computer in which the output file is recorded. To proceed, be sure to complete the calibration procedure according to the apparatus you are using. Considering that this methodology is highly sensitive, the operator must avoid any possible source of external noise (e.g. vibrations, wind), and must frequently clean the blade(s) with an alcohol-soaked wipe. Measure the exact thickness of the leaf sample (lamina or midrib) with a calliper, before placing it in the apparatus. Then, place the sample in the anvil and fix it with the clamp. Leaves are cut at right angles to the midrib, at the widest point along the lamina (or halfway between the base and the tip, if this is difficult to determine). In some studies, the midrib may be removed so that only lamina tissue is tested (or lamina and midrib are tested separately). Alternatively (and less precisely), one can remove the portion of the data that represents the midrib being cut, and analyse these data separately. The procedure to calculate final values can also differ between apparati (for calculation process, refer to the specific user’s manuals).
A calibrated copy of the apparatus described in Fig. 3c is available for use at CNRS in Montpellier, France (contact Eric Garnier; email: garnier@cefe.cnrs-mop.fr).
-
Punch tests
Force to punch (Fp) is the resistance of the actual leaf tissues (particularly the epidermis) to rupture, excluding toughness provided by midribs and main veins. Different penetrometers (Fig. 3d) have been used in the past (there is no standard design), all of which have some kind of fine-needle or a flat-ended punch (diameter ~0.5–5.5 mm) attached to a spring-loaded balance or a counterweight (being a container gradually filled with water and weighed after penetration). The punch goes through a die with a hole in its centre. A clearance of 0.05–0.1 mm between the punch and the edge of the hole in the die is recommended to avoid error in measurements as a result of friction between the punch and the die. A sharp edge in the flat end of the punch will also avoid overestimating toughness values resulting from compression and tension rather than shearing. To standardise the force per unit fraction length, one has to divide the force by the circumference of the punch. The data are therefore expressed in N mm–1. Consistency across the leaf tends to be reasonable, as long as big veins are avoided. Three measurements per leaf are probably sufficient. This test does not work well for many grasses and other monocots. This method has been more widely used in the tropics. Some recent studies have added the punch and die to more sophisticated apparati designed for measuring properties of materials (such as 5542, Instron).
Special cases or extras
-
Leafless plants. See Section 3.1.
-
Leaf-tissue toughness or leaf-specific toughness. This interesting additional parameter of leaf strength can be obtained by dividing Ft, Ws or Fp by the (average) thickness of the leaf sample. Measurement of Lth (in mm) for this purpose is most practically carried out with a calliper, immediately after measuring the leaf-sample width and before placing it in the measuring apparatus.
References on theory, significance and large datasets: Coley (1988); Choong et al. (1992); Grubb (1992); Turner (1994); Wright and Vincent (1996); Cornelissen et al. (1999); Lucas et al. (2000); Díaz et al. (2004); Read and Stokes (2006); Kitajima and Poorter (2010); Onoda et al. (2011).
More on methods: Hendry and Grime (1993); Darvell et al. (1996); Aranwela et al. (1999); Wright and Cannon (2001).
3.8 Leaf lifespan and duration of green foliage
Leaf lifespan (longevity) is defined as the time period during which an individual leaf (or leaf analogue) or part of a leaf (see Monocotyledons below in the present Section) is alive and physiologically active. It is expressed in days, months or years. Leaf lifespan relates to the nutrient-use strategy of a plant, thus providing an indirect index of important plant traits such as potential growth rate, nutrient-use efficiency and decomposability of litter. Long leaf lifespan is often considered a mechanism to conserve nutrients and/or reduce respiratory costs in habitats with environmental stress or low resource supply. Species with longer-lived leaves tend to invest significant resources in leaf protection and (partly as a consequence) grow more slowly than do species with short-lived leaves; they also conserve internal nutrients longer. The litter of (previously) long-lived leaves tends to be relatively resistant to decomposition.
Leaf lifespan does not necessarily indicate the phenology of growth or the proportion of the year when a plant is able to perform photosynthesis. This is because leaves may senesce all together (as in deciduous species), or their senescence may be spread out over a much longer period, in which case a plant may maintain several cohorts of leaves of different ages simultaneously, as is the case in many temperate evergreen species. In addition, some tropical evergreen species maintain a continuous green leaf canopy with a very short leaf lifespan, because they have a high and continuous rate of leaf production. Thus, to understand the ability of a plant to exploit its seasonal light environment and the timing of its growth, it is also useful to measure the duration of green leaves, defined by the number of months per year that the leaf canopy (or analogous main photosynthetic unit) is green, This measure constitutes one important component of the timing of plant growth, or phenology. Certain groups of competition avoiders (including some gap colonisers) may have very short periods of foliar display outside the main foliage peak of the more competitive species; some spring geophytes manage important growth at the beginning of the favourable season, before the canopy of the seasonally green species closes; in contrast, many evergreen species have a year-round ability to photosynthesise. Detailed information on monitoring all the aspects of plant phenology can be obtained from several networks set up for this purpose, including the NPN (http://www.usanpn.org/, accessed 22 February 2013) and Project Budburst (http://www.neoninc.org/budburst/index.php, accessed 22 February 2013).
Measuring leaf lifespan
Different methods are required for taxa with different kinds of phenological patterns and leaf demographic patterns. In all cases where a general measure is desired, select individuals and leaves by using the same criteria as in Section 3.1.
-
Dicotyledons. Method 1 (below) is the best, although it is most labour-intensive and takes a longer time period. Methods 2–4 can replace Method 1, but only if the criteria are met.
-
-
Periodic census of tagged or mapped leaves. Tracking the birth and death of individual leaves over time at repeated census intervals is the best, although most labour-intensive method. Tag or map individual leaves (not leafy cotyledons) when they appear for the first time at a census event, and record periodically (at intervals of roughly 1/10 of the ‘guesstimated’ lifespan) whether they are alive or dead. Because the census interval length increases relative to the mean lifespan, the accuracy of any individual measure will decrease; however, with a large sample size, the estimate of the mean should remain accurate. The leaf-identification information needs to be recorded on the leaf itself (or nearby on the branch, although in an unambiguously systematic way), and often is done with a brief code (such as colour and/or symbols). Alternatively, a branch and leaf drawing, or map, can be made in which different colours are used at each census event. In this method, the position along the branch that separates leaves produced in consecutive census intervals must be marked. Leaves produced in given census intervals are drawn in such a way that their relative spatial positions are clear. At the first census when a leaf is no longer present (or is visibly mostly or completely dead), it is crossed out on the drawing with the colour of the present census event. For each individual plant, select two or more branches or shoots and sample all leaves on them. Note that a total of at least 40 and ideally 160 leaves per species is necessary (see Appendix 1). To achieve this, we recommend increasing the number of individuals, rather than the number of branches or shoots per individual. Calculate the lifespan for each individual leaf and take the average per individual plant. In addition to providing the most accurate estimate of the mean and median leaf lifespan, this method also provides a frequency distribution and enables estimates of the variance, which most other methods do not provide.
-
Count leaves produced and died over a time interval. Count (for each shoot or branch) the total number of leaves produced and died over a time interval that represents a period of apparent equilibrium for leaf production and mortality (see below within the present Section). We recommend about eight counts over this time interval, but a higher frequency may be better in some cases. Then estimate mean leaf lifespan as the mean distance in time between the accumulated leaf-production number and the accumulated leaf-mortality number (facilitated by plotting leaf production and leaf death against time). This is a good method if the census is long enough to cover seasonal periodicity (so typically it needs to be several months up to a year if seasonal periodicity occurs) and the branch or shoot is in quasi-equilibrium in terms of leaf production and mortality. This period can be much shorter for fast-growing plants such as tropical rain forest pioneers, woody pioneers in temperate zone or many herbs. This technique is useful for plants in their exponential growth phase, and for plants with very long leaf lifespan (because one gets data more quickly). Number of individuals and leaves are the same as in Method 1 above.
-
Observe a cohort of leaves until one-half have died. This method measures the median leaf lifespan. Count the number of leaves that appear between two census events. Periodically revisit and count the number of leaves remaining. This method is effective when many leaves are produced within a short time period. Care must be taken to make these measures for multiple consecutive cohorts, if seasonal or interannual variation is likely to cause shifts in the median lifespan of different cohorts. Number of individuals and leaves are the same as in Method 1 above.
-
Counting ‘cohorts’ for many conifers and only some woody angiosperms. For woody angiosperms, it is important to be very familiar with the species. This method is very easy and quick, but can be used only if the species is known to produce foliage at regular known intervals (most frequently once per year) and each successive cohort can be identified either by differences in foliage properties or by scars or other marks on the shoot or branch. In that case, it is simple to count, branch by branch, the number of cohorts with more than 50% of the original foliage remaining until one gets to the cohort with less than 50% of the original foliage remaining, and use that as the estimate of mean leaf lifespan. This works if there is little leaf mortality for younger cohorts, and most mortality occurs in the year of peak ‘turnover’. Many conifers, especially Pinus and Picea, show this pattern, although some Pinus species may flush more than once per year. This method gives a slight overestimate, because there is some mortality in younger cohorts, and usually no or very few survivors in the cohorts older than this ‘peak turnover’ one. This method can also work (1) if there is some mortality in younger cohorts and a roughly equal proportion of survivors in cohorts older than the first cohort with >50% mortality, or (2) if one estimates %mortality cohort by cohort. This can be tricky. For instance, some conifers may appear to be missing needles (judging from scars) that were never there in the first place because of reproductive structures. Be aware that in Mediterranean-type climates, some species experience two growing seasons each year. Count one shoot (preferably the leader or a dominant shoot in the upper canopy) from at least 10 individual plants.
-
Duration of green foliage for species that produce most of leaves in a single ‘cohort’ within a small time period and shed them all within a short time period (see Measuring duration of green foliage, below in the present Section). This method is likely to be the least reliable of the five described herein.
-
-
Monocotyledons. For some monocots species, the longevity of entire blades can be measured as described above. However, given the growth habits of many monocots (see below within the present Protocol), this may not provide an estimate of green-tissue longevity that is comparable to the measures above for dicots. In some grasses and related taxa, the blade continues to grow new tissue while old tissue becomes senescent over time, making the mean of the whole blade lifespan much longer than the lifespan of a particular section of the blade and, therefore, not particularly meaningful as a measure of tissue longevity. In such cases, the production and mortality of specific zones of the blade can be assessed to estimate the tissue longevity, with an adaptation of Method 2 above.
Measuring duration of green foliage
Observe the foliage of 5–10 individuals of a given species several times throughout the year. We recommend a census for all species in the survey at least once a month during the favourable season (preferably including a census shortly before and shortly after the favourable season) and, if possible, one during the middle of the unfavourable season. The months in which the plants are estimated to have at least 20% of their potential peak-season foliage area are interpreted as ‘green’ months. This census can be combined with assessment of Leaf lifespan. Most species with individual leaf lifespans >1 year will be green throughout the year. Note that in some evergreen species from the aseasonal tropics, individual leaf lifespans can be as short as a few months.
Special cases or extras
-
Leafless plants. If photosynthetic tissues do not die and fall off as separate units, follow Method 2 (above) for specific zones of the photosynthetic tissues, as specified above for monocotyledons.
References on theory, significance and large datasets: Chabot and Hicks (1982); Coley (1988); Reich et al. (1992, 1997, 2004); Aerts (1995); Westoby et al. (2000); Wright et al. (2002, 2004); Poorter and Bongers (2006).
More on methods: Jow et al. (1980); Diemer (1998); Craine et al. (1999); Wright et al. (2002); Reich et al. (2004).
3.9 Vein density
Vein networks constrain the transport of water, C and nutrients within the leaf. Vein density, the length of minor veins per unit LA (mm mm–2) can characterise the structure of these networks. Vein density is a structural determinant of hydraulic conductance and photosynthetic rate. Depending on the species set considered, the vein density may correlate with other leaf traits, such as Lth, stomatal density and maximum gas-exchange rates. This trait shows plasticity across environments and is highly variable among species, showing both broad phylogenetic trends and potential adaptation to resource gradients.
Fresh or dried leaves may be used. For the leaf collecting protocol, see Section 3.1 Sampling intensity should account for known trait variation between sun and shade leaves and across the lamina of given leaves. If possible, measurements should be made on sections of lamina containing no large veins. For smaller leaves, use the entire leaf; for larger leaves, a 1-cm2 section is sufficient.
Measuring
Soak the leaf in 5% w/v NaOH–H2O for 24–72 h, until it becomes transparent. If the NaOH solution turns an opaque brown colour during soaking, replace the soaking solution. Then rinse the leaf in H2O and transfer to a 2% w/v NaOCl–H2O solution for 5–30 min, until it becomes bleached.
Rinse the leaf in H2O and transfer via dehydration series to pure ethanol (e.g. 30%, 50%, 70%, 100% ethanol, or 50%, 100% ethanol, each step lasting 30 s, for tender leaves, and 5 min for tougher leaves). Stain the leaf for 15 min in 1% w/v safranin O in ethanol and/or other lignin stains. Destain in ethanol. The leaf can be mounted in water or glycerol on plastic transparency film, or permanently, after transfer to 100% toluene, in immersion oil or Permount (allow several days after mounting for toluene to fully evaporate). Veins will appear red. Take particular care to ensure that all veins in the network are visible. The most numerous and functionally important minor veins can be very hard to see, and often require the epidermis to be removed for accurate visualisation. Vein counts should be made using microscope objectives of ×4 for ferns and up to ×40 for tropical angiosperms. Photograph the venation network under a light microscope, ensuring a large enough field of view (e.g. 1–10 mm2). Then measure the total length of veins in the image and divide this number by the image area to obtain the vein density, using image analysis software (see Section 3.1). This image analysis process may be semi-automated with freely available software (see More on methods, below in the present Section), but the accuracy should be tested.
Special cases or extras
-
Leaf handling. Leaves become delicate during processing and should be moved carefully between solutions or solution should be vacuumed out of the dish, so that leaves will not puncture or tear. For best results in clearing leaves of given species, the details of the protocol can be modified. Warm NaOH may be used (although not boiling), and longer clearing times, or more concentrated NaOCl–H2O solution for a shorter time period. Small or thin leaves may require less soaking time in all solutions. Conversely, very thick or dense leaves may require several days in the NaOH solution before they become transparent. Hairy leaves may require removal of epidermis.
-
Sclereids. In some species, leaves may have sclereids that can be mistaken for veins, and also have an important hydraulic function. Additional venation traits can be measured for more detailed investigations of leaf structure and function (see references below in the present Section).
References on theory and significance: Uhl and Mosbrugger (1999); Roth-Nebelsick et al. (2001); Sack and Frole (2006); Sack and Holbrook (2006); Brodribb et al. (2007, 2010); Boyce et al. (2009).
More on methods: Dilcher (1974); Gardner (1975); Brodribb and Feild (2010); Price et al. (2011).
3.10 Light-saturated photosynthetic rate
The light-saturated photosynthetic rate (Amax) under typical field conditions, usually expressed in µmol m–2 s–1 or nmol g–1 s–1, or both, is a valuable metric as a measure (or at least as an index) of metabolic capacity and a factor determining average realised photosynthetic rate (for upper-canopy foliage). Amax scales with other structural, chemical and longevity aspects of the leaf economic spectrum and, along with those other variables, enables scaling to canopy processes of whole ecosystems. Simultaneous measures of leaf water-vapour conductance are typically made in concert with the photosynthetic measurements.
What, when and how to measure?
Sample young, fully expanded leaves (see Section 3.1). These should be from sunlit parts of the canopy, unless specifically focusing on the shaded taxa of understorey. Measure leaves only if they have been in sufficiently high light just before measurement (e.g. direct sun for 5–10 min) to minimise concerns about leaf induction status or stomatal closure as a result of shading (see Section 3.1 for discussion).
Because realised photosynthesis is less than maximal because of a host of factors, including low or high temperatures, limited soil moisture or air humidity, negative leaf water potential, and source–sink inhibition, among others, care must be taken in choosing the time of year, time of day, and general conditions under which measurements can be made. Some knowledge of gas-exchange responses of the taxa under study will be essential. Do not make measurements during or just following (days to weeks) periods of severe water deficit, or unusual temperatures. Do make measurements on days when soil moisture, plant water status, air humidity, irradiance and temperatures are near optimal for the taxa in question. Measurements in most ecosystems should be made at mid- to late morning (e.g. from 0800 hours to 1100 hours local time) under non-limiting vapour-pressure deficits or temperatures. This minimises the risk of sampling during midday and afternoon declines in gas-exchange rate as a result of stomatal closure, source–sink inhibition or other causes. If a given morning, or mornings in general, are cold relative to photosynthetic temperature optima, measurement can be made later in the day. Because most published measurements have been made under ambient CO2 concentrations, that would be recommended. If rates can also be measured under saturating CO2 concentrations, that is also useful.
Any reliable leaf gas-exchange system can be used, and conditions in the chamber can be either set at levels considered optimal or left to track the in situ conditions (which need to be near optimal). If possible, measure intact foliage or else leaves on branches cut and then re-cut underwater. In the latter case, check whether given individuals fail to stay hydrated. Conduct some test comparisons of gas exchange on intact and ‘re-cut’ branches, to ensure the technique works for your taxa and system. Measurements can also be made on detached foliage; however, this requires even greater attention. Leaves should be measured within seconds, to a few minutes at most, after detachment, and tests of intact v. detached foliage should be made for a subsample, to ensure similar rates are observed.
If possible, the leaf material inside the chamber should be collected (see Section 3.1), measured for LDMC and SLA, and stored for any subsequent chemical analyses.
References on theory, significance and large datasets: Reich et al. (1992, 1997, 1999); Wright et al. (2004).
More on methods: Wong et al. (1979, 1985a, 1985b, 1985c); Reich et al. (1991a, 1991b); Ellsworth and Reich (1992).
3.11 Leaf dark respiration
Characterising leaf dark respiration (Rleaf) in a fashion that enables comparison among species and especially among sites and times of year is challenging, given the sensitivity and acclimation of Rleaf to temperature. However Rleaf under typical field conditions is valuable because it is both a measure of basal metabolism and a rough correlate of average realised night-time respiratory C flux. Rleaf scales with other metabolic, structural, chemical and longevity aspects of the leaf economic spectrum and, along with those other variables, enables scaling to canopy processes of whole ecosystems.
What, when and how to measure?
Sample young to medium-aged fully expanded leaves (see Section 3.1) to ensure negligible respiration associated with biosynthesis (‘growth respiration’). Do not make measurements during or soon after atypical conditions (such as e.g. heat or cold stress, water stress), unless that is the focus of the research. Sample foliage from parts of the canopy sunlit during daytime, unless one is specifically focussed on the shaded understorey taxa. If possible, measure intact leaves at night. In any case, leaves must have been in the dark for ~30 min to minimise variation resulting from very recently fixed photosynthate or transient light-induced respiratory CO2 losses.
Any reliable leaf gas-exchange system that can control leaf temperature can be used. If possible, it is best to measure intact foliage. Detached leaves should be kept moist, cool (to minimise C and water loss), and in the dark until measurement. If possible, tests of intact v. detached foliage should be made for a subsample, to ensure that similar rates are observed. See under Section 3.10 for any subsequent leaf handling.
Leaf dark respiration (Rleaf) can be measured while measuring photosynthetic rate (see Section 3.10), merely by turning off, or shielding, the chamber completely from incident light. However, flux rates for Rleaf are roughly an order of magnitude lower than those for Amax and, therefore, the signal to noise ratio of the typical portable photosynthesis system may be suboptimal for taxa with lower flux rates. One can reduce flow rates and/or increase the amount of foliage into the chamber, to alleviate this problem; however, this is not always sufficient to obtain reliable measurements. In such cases, using specialised chambers (which may hold more foliage) and/or choosing a standardised temperature that is at the high rather than low end of the candidate range (next paragraph) can help ameliorate this problem by increasing the flux rate.
For comparative Rleaf measurements, one typically chooses a standardised temperature appropriate for the site conditions (e.g. 25°C in the tropics, 20°C in the temperate zone, 10°C or 15°C in cold boreal or summer tundra conditions). However, because cross-study comparisons are often made of taxa grown in and/or measured under different temperatures, instantaneous temperature response functions can help in calibrating respiration across temperature regimes. Where possible, measure (subsets of) leaves at appropriate contrasting measurement temperatures, with 10°C intervals, or ideally at least four different temperatures over a 15−35°C range.
References on theory, significance and large datasets: Reich et al. (1998, 2008); Tjoelker et al. (2001); Wright et al. (2004); Atkin et al. (2005); Rodriguez-Calcerrada et al. (2010).
3.12 Photosynthetic pathway
Three main photosynthetic pathways operate in terrestrial plants, each with their particular biochemistry, including C3, C4 and CAM. These pathways have important consequences for optimum temperatures of photosynthesis (higher in C4 than in C3 plants), water- and nutrient-use efficiencies, and responsiveness to elevated CO2 Compared with C3 plants, C4 plants tend to perform well in warm, sunny and relatively dry and/or salty environments (e.g. in tropical savannah-like ecosystems), whereas CAM plants are generally very conservative with water and occur predominantly in dry and warm ecosystems. Some submerged aquatic plants have CAM too. There are obligate CAM species and also facultative ones, which may switch between C3 and CAM, depending on environmental factors (e.g. epiphytic orchids in high-elevation Australian rainforests). Two main identification methods are available, namely C-isotope composition and anatomical observations. CAM can be inexpensively confirmed by verifying that stomata are open at night and closed during the day, or by measuring diurnal patterns of organic acids or leaf pH values. Which method to choose (a combination would be the most reliable) depends on facilities or funding, as well as on the aim of the work (e.g. to contrast C4 v. C3, or CAM v. C3). Although C-isotope composition can be affected by environmental factors, intraspecific genetic differences and/or phenological conditions, intraspecific variability is small enough not to interfere with the distinction between C4 and C3 photosynthetic pathways. In many plant families, only C3 metabolism has been found. It is useful to know in which families C4 and CAM have been found, so that species from those families can be screened systematically as potential candidates for these pathways (see Material S4; Table 1). Below we describe two methods which in combination provide good contrast between pathway types.
What and how to collect?
Collect the fully expanded leaves or analogous photosynthetic structures of adult, healthy plants growing in full sunlight or as close to full sunlight as possible. We recommend sampling at least three leaves from each of three individual plants. If conducting anatomical analysis (see under (B) Anatomical analysis in the present Section), store at least part of the samples fresh (see Section 3.1).
-
C-isotope analysis
Storing and processing
Dry the samples immediately after collecting. Once dry, the sample can be stored for long periods of time without affecting its isotope composition. If this is not possible, the sample should first be stored moist and cool (see under Section 3.1) or killed by using a micro-wave and then be dried as quickly as possible at 70–80°C, to avoid changes caused by loss of organic matter (through leaf respiration or microbial decomposition). Although not the preferred procedure, samples can also be collected from a portion of a herbarium specimen. Be aware that insecticides or other sprays that may have been used to preserve the specimen, can affect its isotope composition.
Bulk the replicate leaves or tissues for each plant, then grind the dried tissues thoroughly to pass through a 40-µm-mesh or finer screen. It is often easier with small samples to grind all of the material with mortar and pestle. Only small amounts of tissue are required for a C-isotope-ratio analysis. In most cases, less than 3 mg of dried organic material is used.
Measuring
Carbon isotope ratios of organic material (δ13Cleaf) are measured using an isotope ratio mass spectrometer (IRMS, precision between 0.03 ‰ and 0.3 ‰, dependent on the IRMS used) and are traditionally expressed relative to the Pee Dee Belemnite (PDB) standard as δ13C in units of per mil (‰), i.e. parts per thousand. After isotopic analysis, the photosynthetic pathway of the species can be determined on the basis of the following (see graphic explanation in Material S4; Fig. 1):
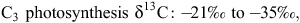
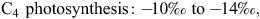
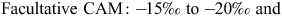
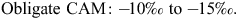
Separating C3 or C4 from CAM plants is difficult on the basis of δ13C alone (for facultative CAM plants, δ13C values have been found to range as widely as from –14‰ to –23‰). However, as a rule of thumb, if δ13C is between –10‰ and –23‰, and the photosynthetic tissue is succulent or organic acid concentrations are high during the night, but low during the day, then the plant is CAM. In such cases, anatomical observations and diurnal measurements of gas exchange or biochemical analysis would be decisive (see (B) Anatomical analysis in the present Section).
-
Anatomical analysis
C3 and C4 plants typically show consistent differences in leaf anatomy, best seen in a cross-section. Using a razor blade or microtome, make cross-sections of leaf blades of at least three plants per species, making sure to include some regular veins (particularly thick and protruding veins, including the midrib and major laterals, are not relevant). C3 plants have leaves in which all chloroplasts are essentially similar in appearance and spread over the entire mesophyll (photosynthetic tissues). The mesophyll cells are not concentrated around the veins and are usually organised into ‘palisade’ and ‘spongy’ layers parallel to, and respectively adjacent to, the upper to lower epidermis (see Material S4; Fig. 2) (vertically held C3 leaves often have a palisade layer adjacent to each epidermis and a spongy layer between the two palisades). The cells directly surrounding the veins (transport structures with thin-walled phloem and generally thicker-walled xylem cells), called bundle sheath cells, normally contain no chloroplasts. C4 plants, in contrast, typically exhibit ‘Kranz anatomy’, viz., the veins are surrounded by a distinct layer of bundle-sheath cells (Material S4; Fig. 2) that are often thick-walled, and possess abundant, often enlarged chloroplasts that contain large starch granules. The mesophyll cells are usually concentrated around the bundle-sheath cells, often as a single layer whose cells are radially oriented relative to the centre of the vein, and contain smaller chloroplasts with no starch grains. These differences can usually be identified easily under an ordinary light microscope. Many plant physiology and anatomy textbooks give further illustrations of Kranz v. typical C3 leaf anatomy (see More on methods below in the present Section).
If Kranz anatomy is observed, the species is C4 If not, it is likely to be C3 unless the plant is particularly succulent and belongs to one of the families with CAM occurrence. In the latter case, it could be classified as (possible) CAM. Many CAM leaves do not have typical C3 palisade or spongy mesophyll layers, but only a thin layer of more or less isodiametric, chloroplast-containing cells just under their epidermis, with the entire centre of the leaf consisting of large, thin-walled, colourless parenchyma cells that store water and organic acids. If living plants are within easy reach, an additional check could be to determine the pH of the liquid obtained by crushing fresh leaf samples in the afternoon (see Section 3.5), and again (with new, fresh samples from the same leaf population) at pre-dawn. Because in a CAM plant, organic (mostly malic) acids build up during the night, and are broken down during the day to supply CO2 for the photosynthesis in the leaf, CAM species show a distinctly lower pH after the night than they do in the afternoon. In addition, C-isotope ratios can provide further evidence to distinguish between CAM and C3 or C4 metabolism (see (A) C-isotope analysis above in the present Section).
Special cases or extras
-
Permanent slides or photographs and chloroplast visibility. A range of methods is available for making the microscope slides permanent; however, be aware that some may result in poorer visibility of the chloroplasts. One method for retaining the green colour of the chloroplasts is to soak the plant or leaves in a solution of 100 g CuSO4 in 25 mL of 40% formal alcohol (formaldehyde alcohol), 1000 mL distilled water and 0.3 mL 10% H2SO4 for 2 weeks, then in 4% formal alcohol for 1 week, subsequently rinse with tap water for 1–2 h and store in 4% formal alcohol until use. However, material thus treated can be sectioned only by using a microtome after embedding or freezing it, in contrast to many living, turgid leaves, which can be sectioned free-hand by using a suitable technique such as sectioning a rolled-up leaf or a stack of several leaves. Photomicrographs of freshly prepared sections are an alternative way to keep records for later assessment.
References on theory, significance and large datasets: O’Leary (1981); Farquhar et al. (1989); Earnshaw et al. (1990); Ehleringer et al. (1997); Lüttge (1997); Zotz and Ziegler (1997); Wand et al. (1999); Pyankov et al. (2000); Sage (2001); Hibberd and Quick (2002).
More on methods: Farquhar et al. (1989); Ehleringer (1991); Hattersley and Watson (1992); Mohr and Schopfer (1995); Belea et al. (1998); Pierce et al. (2002); Taiz and Zeiger (2010).
3.13 C-isotope composition as a measure of intrinsic water-use efficiency
Uptake of CO2 through stomata inevitably leads to loss of water vapour. The relative magnitude of photosynthesis and transpiration depends on several physiological, morphological and environmental factors, such that different species in different growing conditions can have widely different C gain per unit water loss. This quantity, the ratio of the rates of net photosynthesis and transpiration (= water use efficiency, WUE), is of great ecological interest and can be measured on short or long time scales.
On short time scales (= instantaneous), WUE is often measured with infrared gas analysis (see Section 3.10). However, instantaneous WUE changes dramatically for a given leaf over short time spans, e.g. because of variable light intensity and vapour pressure deficit. This makes separating species effects and environmental effects challenging. For comparative studies, we recommend taking into account the precautions outlined in Section 3.10, and to calculate ‘instantaneous intrinsic WUE’, the ratio of net photosynthesis to stomatal conductance. This excludes the effect of differences in vapour pressure on transpiration rates. As CO2 and water vapour share the same stomatal diffusion pathway, but with diffusion of water being 1.6 times faster than that of CO2, intrinsic WUE relates to the CO2 gradient as follows:
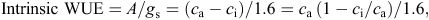
where A is net photosynthesis, gs is stomatal conductance, ca and ci are the mole fractions of CO2 in ambient air and in the substomatal cavity, respectively.
The C-isotope approach has proved extremely useful to study WUE over longer time scales. It relies on the fact that photosynthetic enzymes discriminate against the heavier stable isotope 13C (relative to 12C) during photosynthesis, so that C in leaves is always depleted in 13C compared with that in the atmosphere. The extent of the enzyme’s discrimination against 13C depends on ci. If ci is low relative to ca, then the air inside the leaf becomes enriched in 13C, and the ability of the enzyme to discriminate declines. As a result, the plant ends up fixing a greater proportion of 13C than a plant performing photosynthesis at a higher ci. In its simplest form, for C3 plants, Δ = a + (b – a)ci/ca, where Δ is photosynthetic 13C discrimination, a = 4.4‰ and b = 27‰. Therefore, Δ allows time-integrated estimates of ci : ca and intrinsic WUE. Note that Δ is calculated from δ13C (see Section 3.12), as follows: Δ = (δ13Cair – δ13Cplant)/(1 + δ13Cplant), which highlights the requirement for assumptions or measurements of the isotope composition of the air.
Because intrinsic WUE changes rapidly, the bulk leaf 13C : 12C ratio of fixed C correlates with the ci:ca ratio for the time period during which the C comprising the leaf was fixed weighted by the photosynthetic flux. In other words, the 13C : 12C represents a longer-term measure of ci : ca, especially reflecting ci : ca during favourable periods.
What and how to collect?
For intrinsic WUE assessment, δ13C is usually determined for leaves, but can be determined on any plant part, e.g. on tree rings for a historical record. Note that, in general, there is fractionation between leaves and stems, with all non-photosynthetic organs being more enriched in 13C than are leaves. This enables differentiation between growing conditions using tree rings of different ages, and also means that leaves that grew in different years or different seasons can have different Δ, which has implications for the sampling strategy. Leaves at different positions in a tree or in a canopy can vary in Δ as a result of differences in stomatal opening and photosynthetic capacity, and also because of differences in the isotope composition of the source air. To estimate Δ, the isotope composition of the air needs to be known. In freely circulating air such as at the top of a canopy, it is generally reasonable to assume that the isotope composition of air is constant and equal to that of the lower atmosphere (δ13Cair ≈ –8‰).
Storing and processing
Samples should be dried as soon as possible and finely ground. Grind the dried tissues thoroughly to pass through a 40-µm-mesh or finer screen. C-isotope ratio analysis requires only small samples (2–5 mg); however, it is recommended to sample and grind larger amounts of tissue to ensure representativeness.
Measuring
See Section 3.12 for measuring C-isotope concentrations.
Special cases or extras
-
Cellulose extracts. Isotopes are sometimes analysed using cellulose extracts to avoid variation introduced by the slightly different isotope composition of other C compounds. In most cases, however, the Δ values of the whole tissue and those of cellulose correlate very well. Shorter-term (typically at the scale of a day) studies of Δ have sampled recent assimilates rather than structural C, either by extracting non-structural carbohydrates from snap-frozen leaves, or by sampling phloem sap.
-
Assumptions. We reiterate here that the estimation of intrinsic WUE from C-isotope composition involves several assumptions, that intrinsic WUE does not necessarily correlate well with the actual WUE (photosynthesis to transpiration ratio), with mesophyll conductance being a particular complication, and that the equation for Δ given above is a simplification of the theory. It is also important to note that, because of their different biochemistry, the equation given for Δ does not apply to C4 or CAM plants, and in these groups, C-isotope composition is not useful for estimating intrinsic WUE.
References on theory, significance and large datasets: Farquhar et al. (1989); Cernusak et al. (2009).
More on methods: Ehleringer and Osmond (2000); Seibt et al. (2008); Diefendorf et al. (2010).
3.14 Electrolyte leakage as an indicator of frost sensitivity
Electrolyte leakage after freezing is an indicator of leaf frost sensitivity and is related to climate, season and plant geographical distribution. Leaves of species from warmer regions and/or growing at warmer sites along a steep regional climatic gradient have shown greater frost sensitivity than those of species from colder regions and/or growing at colder sites within a regional gradient. The technique described here is based on the idea that when a cell or tissue experiences an acute thermal stress, one of the first effects is disruption of membranes, eliminating a cell’s ability to retain solutes such as ions. Ion leakage from a tissue can be easily assessed by measuring changes in electrolyte conductivity of a solution bathing the tissue. The technique is suitable for a wide range of leaf types (from tender to sclerophyllous) and taxa (monocotyledons and dicotyledons), and is not affected by cuticle thickness.
What and how to collect?
Collect young, fully expanded sun leaves with no sign of herbivory or pathogen damage. Deciding when to collect is more complicated. The answer will depend on the question being asked, although in most cases, collection should be standardised across taxa. Depending on the contrast of interest, collect foliage during the peak growing season (see Section 3.1), or preferably near the end of the season (see Special cases and extras below in the present Section) or in winter (for winter evergreen species). If a species grows along a wide environmental gradient, and the objective is an interspecific comparison, collect the leaves from the point of the gradient where the species is most abundant. If many species are considered, try to collect them within the shortest possible time interval, to minimise differences resulting from acclimation to different temperatures in the field. Collect leaves from at least five randomly chosen adult individuals of each species.
Storing and processing
Store the leaf material in a cool container until processed in the laboratory (see Section 3.1). Process the leaves on the day of the harvest, so as to minimise natural senescence processes. For each plant, with a cork borer cut four circular 5-mm-diameter leaf disks (to provide for two treatments using two disks each, see below within the present Protocol), avoiding the main veins. For needle-like leaves, cut fragments of the photosynthetically active tissue that add up to a similar LA. Rinse the samples for 2 h in deionised water on a shaker, then blot dry and submerge two disks (or their equivalent in leaf fragments) in 1 mL of deionised water in each of two Eppendorf tubes. Complete submergence is important. For each treatment (see below within the present Protocol), prepare as many replicates (one replicate being two tubes, each containing two disks or equivalent leaf fragments) as the number of plants sampled.
Measurement
Apply the following two treatments, without any prior acclimation, to the two leaf disk/fragment samples in the respective tubes: (1) incubation at 20°C (or at ambient temperature, as stable as possible) for the control treatment, and (2) incubation at −8°C in a calibrated freezer, for the freezing treatment. Incubations should be for 14 h in complete darkness, to avoid light-induced reactions.
After applying the treatment, let the samples reach ambient temperature and then measure the conductivity of the solution. Do this by placing a sample of the solution from an Eppendorf tube into a standard previously calibrated conductivity meter (such as the Horiba C-172; Horiba, Kyoto, Japan) and by recording the conductivity. Then place the Eppendorf tube in a boiling water bath for 15 min to completely disrupt cell membranes, releasing all solutes into the external solution, then re-measure its conductivity. Prior to immersion, puncture the cap of each Eppendorf tube to allow relief of pressure during boiling.
Calculations
-
Percentage of electrolyte leakage (PEL) – separately, for the frost treatment and the control for each individual plant replication, as follows:
-
-
where es is the conductivity of a sample immediately after the treatment, and et is its conductivity after boiling. High values of PEL indicate significant disruption of membranes, and thus cell injury; the higher the PEL, the greater the frost sensitivity.
-
-
Corrected PEL – the PEL of the control treatment can vary among species because of intrinsic differences in membrane permeability, experimental manipulations and differences in injury when leaf disks or fragments are cut. To control for these and other sources of error, subtract the PEL of the control treatment of each replicate from that for the freezing treatment. Corrected PEL is thus
-
-
For calculating the mean, standard deviation or standard error for a species, the average corrected PEL for each individual plant replicate counts as one statistical observation.
-
Special cases and extras
-
Applicability to different plant functional types. The technique is not suitable for halophytes and succulents. It is not necessarily applicable to deciduous plants and hemicryptophytes, because their significant frost tolerance involves stems and buds rather than leaves. This tolerance could possibly be tested with sections cut from stems, although the reliability of this has not been investigated, to our knowledge, as it has been for leaves.
-
Season of collection. Because of the recognised wide occurrence of autumnal acclimation in frost tolerance, we recommend normally performing the procedure with leaves collected at or near the end of the growing season.
-
Incubation with dry ice. A different treatment, namely incubation at about −78°C (the temperature of dry ice), with the rest of the protocol the same as described above, can be used if a freezer whose temperature can be controlled at about −8°C is not available, or if one wishes to detect tolerance to the kind of severe frost that can occur at high latitudes or altitudes. It would not detect tolerance to merely mild frost.
-
Acclimation. The occurrence of acclimation to mild frost could be detected using a freezer at −8°C, on leaf samples collected on successive dates in summer and autumn.
-
Sensitivity to high temperatures. The same basic technique, with a modification in the treatment temperature, has been successfully applied to leaf sensitivity to unusually high temperatures (~40°C; see More on Methods below in the present Section).
-
Chilling sensitivity. is a physiological limitation that can be ecologically important in mountains at lower latitudes, and might be detected by this technique. It is usually tested for by incubation for 24 h or more at about +5°C, e.g. in an ordinary refrigerator. Alternatively, 0°C in a distilled water (or rain water) bath could be used, because this will not actually freeze plant tissue. A chilling-sensitive tissue would leak electrolytes after such incubation, whereas a chilling-tolerant tissue should not.
References on theory, significance and large datasets: Levitt (1980); Blum (1988); Earnshaw et al. (1990); Gurvich et al. (2002).
More on methods: Earnshaw et al. (1990); Gurvich et al. (2002).
3.15 Leaf water potential as a measure of water status
Species facing soil water shortage can avoid water stress to a degree by dropping leaves, or delay the development of water stress in their tissues by rooting deeply, or by shutting stomata and losing stored water only slowly through their cuticle. Alternatively, tissues may tolerate physiological desiccation. The bulk leaf water potential (ΨL; units MPa) is a simple indicator of leaf water status; the more negative the value, the more dehydrated the leaf.
When measured pre-dawn, the plant may have become equilibrated with the soil during the night, and the ΨL may thus represent the soil water potential in the ‘average’ root zone. However, recent work has shown instances of substantial disequilibrium between pre-dawn ΨL and soil water potential as a result of several mechanisms, including nocturnal transpiration, cavitation in the xylem and osmolyte accumulation in the cell walls. Thus, pre-dawn ΨL may be more negative than the soil water potential, and should be used only as a tentative index of soil water availability.
During the day, ΨL will decline below the soil water potential as a result of transpiration into the atmosphere. When measured in the dry season, the midday ΨL can provide a useful index of the degree of physiological drought experienced. Thus, the minimum value for ΨL that a plant reaches, usually at midday at the driest, hottest time of year, can be used as an index of the tolerance to water shortage that the species (or individuals and populations) demonstrate (assuming that the plants are still healthy and not drought-injured).
What and how to collect
Measurement of minimum values of ΨL is typically carried out at the end of the hot, dry season for evergreen species and in Mediterranean winter-rain ecosystems. However, in summer-rain ecosystems, the time of year at which drought stress is maximal may not be obvious. Repeated-measurements in different seasons can help find the real minimum ΨL for each species.
Depending on the type of pressure chamber used (see below within the present Protocol), either leaves or short, terminal, leafy twigs should be collected. Samples should be collected at midday and, as previously indicated (see Section 3.1), from shoots or individuals located in the sun. Leaves should have been exposed to direct sun for at least 30 min before collection (avoid cloudy days). We recommend measuring samples as soon as possible, or at least within half an hour of collecting all samples (with the number of samples depending on the number of pressure chambers available) over a period of no more than half an hour between the first and last measurement. Samples should be collected into sealable plastic bags, into which one has just exhaled to increase moisture and CO2 to try to minimise shoot transpiration within the bag. Samples sealed in plastic bags should be kept refrigerated and in darkness (e.g. in a refrigerated picnic fridge, or an insulated cooler box containing pre-frozen cooling bars or ice).
Measuring
The simplest way to measure leaf water potential is with a pressure chamber, or Scholander bomb (see diagram in Fig. 4). This consists of a pressure container into which the sample (leaf or terminal twig) is placed, a manometer or pressure gauge to measure the pressure inside the chamber, and as a pressure source, a pressure tank of liquid N, connected to the chamber through a needle valve and pressure-safe (normally copper) tubing. Many models with different characteristics are commercially available.

|
A leaf or shoot is placed inside the chamber, with its cut end projecting to the exterior through the sealing port. Pressure, from the N tank, is then gradually increased in the chamber. When a drop of water appears at the cut end of the specimen, the ‘balance pressure’ indicated by the gauge or manometer is recorded. Assuming that the xylem osmotic potential is very low, the balance pressure represents the equilibrium water potential of the plant material in the chamber, multiplied by –1. Leaf water potential is conventionally expressed in MPa. Minimum leaf water potentials usually vary from near 0 to −5 MPa, but can be lower in (semi-)arid ecosystems. Extreme care should be taken when pressure chambers are under high pressures.
References on theory, significance and large datasets: Hinckley et al. (1978); Ackerly (2004); Bucci et al. (2004); Bhaskar and Ackerly (2006); Lenz et al. (2006); Jacobsen et al. (2008); Bartlett et al. (2012).
References on methods: Scholander (1966); Turner (1988).
3.16 Leaf palatability as indicated by preference by model herbivores
Despite the vast diversity and complexity of herbivores, plants, and their interactions, a small number of components of leaf quality affect preference by generalist herbivores in a predictable way. Leaf palatability (as indicated by preference by herbivores) can be seen as an integrator of several underlying leaf-quality traits. Additionally, palatability tends to be correlated with litter decomposability among species because both are limited by similar constraints (e.g. low nutrient contents, high concentration of lignin and secondary metabolites).
One method for quantifying leaf palatability, as indicated by model herbivore preference, is a cafeteria assay in which generalist herbivores are allowed to feed selectively on leaf samples cut out from fresh leaves of a whole range of species distributed in random positions on a feeding arena. These experiments can provide useful information about herbivore preference for a broad range of plant species.
What and how to collect
Leaves are selected and collected as indicated in Specific leaf area in Section 3.1, preferably from at least 10 individuals per species. Be aware that all species must be collected within 2 days before the trial and stored in refrigerated and moist conditions. If different species grow in different seasons, at least two cafeteria trials should be carried out, making sure common species of contrasting quality are included in both for cross-calibration.
Storing and processing
All leaves are kept in sealed bags at 4–5°C until processed. Once all leaves have been collected, 10 1-cm2 samples (one from each individual) should be cut from each fresh leaf and randomly placed, securing with a thin needle, on a numbered grid cell on a polystyrene feeding arena (Fig. 5). The surface is covered by transparent plastic in the case of trials with snails. Large veins should be avoided unless leaves are too tiny. For narrow leaves, an equivalent area is reached by cutting an appropriate number of 10-mm lengths from the mid-leaf section and pinning these down together into a star shape. Tiny leaves may also need to be grouped like this (with minimal overlap) to make up to ~1 cm2 In the case of highly succulent and aphyllous species, a thin 1-cm2 fragment of epidermis and adjacent mesophyll (relatively young photosynthetic tissue) is used as a leaf analogue. While preparing all leaf samples be sure to keep the cut leaves in a water-saturated environment (e.g. plastic bags containing moistened paper towel) to preserve turgidity. Including additional samples from a known material, such as lettuce for human consumption, popular with some of the herbivores used for these cafeterias (snails and slugs), can be useful to test animal behaviour. If the animals do not eat from known preferred material, either the animals or the feeding conditions are probably not right.

|
Measuring
Once all samples have been pinned onto the arena, several herbivores (~10 per 500 leaf samples) are placed in a random position and the arena is closed with a plastic net (Fig. 5). Snails (e.g. Helix spp.) require a cool, dark and humid environment (spraying, dark plastic cover) that will stimulate consumption. Grasshoppers need a dry and light, and crickets a dry and dark environment. The arena should be covered with netting to avoid any escapes. After herbivores have been placed, consumption is measured by direct observation after 4, 8 and 12 h, and subsequently every 12 h for 3 days. The %LA consumed can be estimated by eye (with 2–10% accuracy per sample) on the basis of the original cut shape. Actual LA may also be measured accurately (see Section 3.1) before and after the trial, provided the samples do not deteriorate during this procedure. To be sure that the model herbivores do not have previous experience with the plants included in the trials, the herbivores may be bred, or collected when young and raised in captivity, without exposure to any of the plants included in the cafeteria experiments. This and a pre-trial 48-h starvation period (promoting consumption during the trial) are important to get unbiased results.
Special cases or extras
-
Independent feeding trials. It is recommended to assess leaf palatability in at least two independent feeding trials, using different model herbivores to cover a wider range of preferences by generalist herbivores. Snails are recommended for their generalist-feeding habits, but they consume few graminoid monocots; grasshoppers and crickets are better at discriminating between leaf qualities within graminoids. Instead of selecting one recording time, several consumption measurements can be compared with analyse-first choice and successive choices. Values of LA consumption can be transformed into values of leaf biomass if SLA is included in the calculations (see Section 3.1).
-
Palatability v. accessibility. Experiments can be designed to evaluate palatability v. accessibility, following the same theoretical background on which palatability tests are based. For example, by offering whole shoots with and without spines to different animals (in this case, model herbivores should be bigger than snails or grasshoppers) and recording how much biomass is consumed per unit time.
References on theory, significance and large datasets: Grime et al. (1970, 1996); Southwood et al. (1986); Coley (1987); Hartley and Jones (1997); Cornelissen et al. (1999); Singer (2000).
More on methods: Pérez-Harguindeguy et al. (2003).
3.17 Litter decomposability
Plant species exert strong control over decomposition rates through the ‘afterlife’ effects of attributes of living plant modules (leaves, stems and branches, roots) on the quality of their litter. Shifts in species composition, as a consequence of global changes, land use or natural succession are strong drivers of changing litter decomposition rates in various biomes worldwide, with feedbacks to climate through the release of CO2 To estimate the organ afterlife effects of different species on decomposition rates, a powerful method in recent years has been to compare mass losses of (leaf) litters of multiple species by incubating them simultaneously in semi-natural outdoor ‘common-garden experiments’, particularly in litter beds. Species-specific ‘decomposability’ thus derived integrates several structural and chemical traits of the leaf (or other plant part) because it is an expression of the quality of litter as a substrate for microorganisms. Decomposability is usually related to leaf dry-matter content, leaf toughness, as well as N and lignin content, in most studies; additionally, it has been found to be related to SLA, leaf pH, tannin, P or base-cation content in others.
What and how to collect
Here, we focus on leaf litter, although the protocol can also be applied to other plant parts with some adjustments (see under Special cases or extras in the present Section). Freshly senesced, undecomposed leaf litter should be collected from mature plants in the field. In species that shed their litter, it can be either collected from the ground within a few days after falling or by placing a clean cloth or net beneath the tree or branch and gently shaking it until senesced leaves fall. It is important to brush or rinse-off any soil before further processing. In species that retain dead leaves on plants or die back completely above ground, cut off leaves that are subjectively judged to have died very recently and are still undecomposed. Often such dead leaves are still shiny and, in deciduous woody species, may still show bright autumn colours. Petioles and rachides of compound leaves that are shed as an integral component of the leaf litter should be collected and processed as such (but see discussion under Section 3.1). In aphyllous species, where the entire shoot functions as the green photosynthetic unit analogous to leaves and also senesces as a unit, such units are collected as leaf litter. In leaves that gradually die from the tip down, as in many monocotyledons, collect those leaves in which the middle lamina section is at the right phase, i.e. complete senescence without any evidence of decay. For species in which leaves senesce sequentially in cohorts, we recommend collecting several times from the same plants, to get representative leaf litter.
Storing and processing
Litter collections should be air-dried in open paper bags until they reach their equilibrium moisture content. To preserve its characteristics, litter must never be dried at high temperatures. Once dry, the litter can be stored in the same bags for up to several months in a relatively dry environment away from sunlight. For each species, an air-dried subsample of leaf litter is weighed, and again after 48 h in an oven (60°C), so as to calculate initial oven-dry weights of the samples for incubation. There are no fixed rules regarding litterbag size, mesh type or mesh material, but litterbags should be made of non-degradable, inert but flexible materials. Typical litterbags measure from 10 × 10 cm to 20 × 20 cm and are made of 1-mm polyester or fibreglass mesh. Litterbag sizes may vary within one study, so as to standardise the ‘relative packing’ of litter inside the bags for morphologically contrasting species. Finer mesh size should be used when, among the litters to be compared, narrow-leaved species are included, whereas wider mesh sizes allow a more complete set of organisms to be involved in decomposition and have less effect on the litter microclimate. Comparing some of the same species at different mesh sizes will help calibrate among methods. A litterbag usually contains a standard small amount (e.g. 1 g) of dry litter; however, for some species, or in some environments, it may be necessary to increase or decrease the initial mass per litterbag. Very big leaves may have to be cut into sections, with representative proportions of midrib and petiole, whereas flexible long monocot leaves can sometimes be rolled up (if not, better cut them into segments). If there are large differences in initial mass among some species, we recommend incubating some reference species at two or three different initial masses, for calibration among species afterwards. It is important not to break the leaf litter during handling. Filling the bags using funnels or tubes inserted into the bags temporarily may help here. Litterbags can be sealed with polycarbonate glue, by sewing (with nylon thread), stapling (with non-rusting staples) or by using a laboratory heat-sealer. Make sure a resistant label (e.g. plastic, with code embossed) is firmly attached to the litterbag or sealed inside.
Measuring
Litterbags are incubated in a purpose-built outdoor litter bed. Litter beds can range from just a rather homogeneous square in the field, cleaned from vegetation and litter, with litterbags simply on the soil surface, to a much more elaborate bed, made by wooden or plastic squares (with natural drainage) filled with soil thoroughly mixed with a standard or combined litters, and covered with the same litter mixture (Fig. 6). Depending on the type of study, the composition of this mixture may be based on a particular community or several communities; however, a more standardised mixture not based on local communities may be used for particular purposes (see below within the present Protocol). In any case, the composition should be homogeneous across the bed, to prevent differences in decomposition rate caused by the microenvironment. To further account for environmental gradients within larger beds, it is recommended to divide the bed in equal statistical blocks, with one replicate of each species in a random position in each block. The litterbags are usually incubated 1–2 cm below the surface of this mixture, to reduce heterogeneity in moisture dynamics among the samples. Additional control samples of some of the species may be treated identically, and retrieved immediately after burial to control for loss or ingression of particles during burial. Spraying the samples with demineralised or rain water before incubation will get them to field capacity more quickly. Stretching 3–5-cm-mesh nylon net or a metal grid over the bed may protect the samples from the activities of mammals and birds. Additional litterbags filled with pieces of plastic broadly representing litter amount and shape can be incubated during the whole period to check for the contribution of exogeneous organic matter to the sample. Retrieval dates and sampling schedule may vary, first, according to the microclimatic conditions that will determine decomposition rates and, second, to the average quality of the litters within litterbags. In moist tropical environments, almost 90% mass loss can be reached in less than 2 months, given high litter quality, whereas in arid or cold environments, 2 years may be the minimum time needed for over 50% mass loss for the same litter quality. Two retrieval dates are usually enough to check for the consistency of decomposability ranking across species over time, whereas five or more may be needed if decomposition dynamics are to be analysed too. After retrieval, samples may either be cleaned up and processed directly, or first frozen. Adhering soil, soil fauna and other extraneous materials must be removed from the decomposed litter, to the degree possible using e.g. tweezers and small brushes (Fig. 6). Litter samples are then oven-dried for 48 h at 60°C, then weighed. Decomposition rate can be defined as the percentage of mass loss after the incubation period or as k (decomposition constant) derived from a presumed exponential mass-loss curve over time involving more than one harvest, as follows:

|
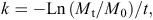
where k = decomposition rate constant (year–1), M0 = mass of litter at time 0, Mt = mass of litter at time t and t = duration of incubation (years).
Calibration among studies
A disadvantage of the litter-bed assay is that the incubation environments and periods vary greatly, so that mass loss or k values can be compared directly only within but not between studies or sites. We advocate the establishment of one or a few centralised litter beds, which could simultaneously host samples from multiple sites. It would be better still if such litter beds, or equivalent ‘common garden’ facilities, could provide close to standardised environmental conditions from year to year, e.g. in a controlled environment greenhouse. In the absence of such facility, we recommend (1) additional inclusion of a few species from each range of sites in a multispecies litter bed, so as to be able to calibrate mass-loss values across multiple sites, and thereby across multiple species from these sites or (2) inclusion of litter samples of a few reference species in multiple litter beds across sites.
Special cases or extras
-
Microcosms. Although most litter beds are situated under field conditions, litterbags can also be incubated in microcosms when more detailed effects of soil and litter fauna or leaching is studied, or when the researcher needs to standardise the incubation conditions for any other reason. As in common-garden experiments, microcosms can be sampled at certain time intervals during the process of decomposition, and the incubated material can be analysed for its mass loss, chemical changes and biological colonisation.
-
Contamination. When sand or clay contamination is high, even after removing obvious extraneous matters, the sample can be oven-dried and then sieved to remove bulk of the sand or clay fraction. If contamination is too high and difficult to eliminate, the incubated litter sample can be ashed for 4 h at 450°C, and the mass loss expressed on the basis of the ash-free mass (%) of the initial and final litter samples (this is not appropriate for soils high in organic matter).
-
Photodegradation. One should be aware that in some dry or windy environments with high irradiance, mass loss resulting from photodegradation or sand abrasion (in addition to microbial decomposition) may be substantial and this may affect the species rankings of mass loss in incubations under exposed conditions or below a litter layer. Species differences in photodegradation at a given exposure may themselves be of interest.
-
Litter decomposabilities of other organs (fine and coarse roots, fine and coarse stems), and the extent to which these are coordinated between organs across species, are also of great ecological relevance. A new method is available to include coarse woody debris together with fine litter in such comparative common-garden studies, taking into account differences in size and time scale of decomposition.
References on theory, significance and large datasets: Cornelissen (1996); Cadisch and Giller (1997); Cornelissen et al. (1999, 2007); Garnier et al. (2004); Austin and Vivanco (2006); Parton et al. (2007); Adair et al. (2008); Cornwell et al. (2008); Fortunel et al. (2009); Freschet et al. (2012).
More on methods: Taylor and Parkinson (1988); Cornelissen (1996); Robertson et al. (1999); Graça et al. (2005); Berg and Laskowski (2005); Freschet et al. (2012).
4 Stem traits
4.1 Stem-specific density
Stem-specific density (SSD, mg mm–3 or kg dm–3) is the oven-dry mass (at 70°C for 72 h, but see Special cases or extras in the present Section) of a section of the main stem of a plant divided by the volume of the same section, when still fresh. This trait is a synonym for ‘stem density’; we distinguish it from ‘wood density’ in that SSD can also be measured on herbaceous species and includes the stem bark (i.e. secondary phloem and cork if any), which in some cases accounts for a significant proportion of the overall stem structure (see Special cases and extras in the present Section). SSD is emerging as a core functional trait because of its importance for the stability, defence, architecture, hydraulics, C gain and growth potential of plants. Stem density partly underlies the growth-survival trade-off; a low stem density (with large vessels) leads to a fast growth, because of cheap volumetric construction costs and a large hydraulic capacity, whereas a high stem density (with small vessels) leads to a high survival, because of biomechanical and hydraulic safety, resistance against pathogens, herbivores or physical damage. In combination with plant size-related traits, it also plays an important global role in the storage of C.
What and how to collect?
Healthy adult plants should be selected according to previous suggestions. Depending on the objectives of the study, either stem density or wood density should be measured (see Special cases or extras below in the present Section). For stem density, remove only the loose bark that appears functionally detached from the stem (see Section 4.3). Stem density can increase, decrease or remain constant from pith to bark, so a representative sample must include a proportional representation of the complete stem. Stem density is higher at the insertion point of branches (because of tapering of vessels), and lower for branches and for outer bark (which includes more air spaces and cork). These density gradients have important implications for the sampling procedure. Collect either a whole stem cross-sectional slice or, for large trees, a triangular sector from the bark tapering into the centre (like a pizza slice) of cross-sectional area, ~1/8 of the total area. For herbaceous species or woody species with thin main stems (diameter <6 cm), cut with a knife or saw a ~10-cm-long section near the base of the stem (between 10- and 40-cm height). If possible, select a regular, branchless section, or else cut off the branches. For woody or thick succulent plants with stem diameters >6 cm, saw out a (pizza) slice from the trunk at ~1.3-m height; the slice should be 2–10 cm thick. Hard-wooded samples can be stored in a sealed plastic bag (preferably cool) until measurement. Soft-wooded or herbaceous samples are more vulnerable to shrinkage, so should be wrapped in moist tissue in plastic bags, and stored in a cool box or refrigerator until measurement.
Measuring
Stem volume can be determined in, at least, two different ways.
-
Water-displacement method. This procedure allows the volume of irregularly shaped samples, which cannot be properly measured with the dimensional method, to be measured easily. A graded test tube or beaker, large enough to hold the entire sample, is filled with distilled water (but not completely filled, to be sure that when the stem is placed within the water, the liquid will not escape from the test tube), placed on a balance, and tare the balance. The wood sample is then completely submerged under the water with the aid of a small-volume needle or tweezers, being careful to not touch the sides or bottom of the beaker which can cause variations in the weight being registered by the balance. When the wood sample is submerged, the increase on water level leads to an increase in the weight being registered by the balance (i.e. the weight of the displaced water), which equals the wood sample volume in cm3 (because water has a density of 1 g cm–3). The mass registered in the balance (i.e. the volume of the wood sample) is quickly read and registered. Tare again for each measurement, previously replacing water.
-
Measurement of dimensions (or dimensional method). The volume of a cylindrical sample can be determined simply by measuring its total length (L), and its diameter (D), on one or more places along the sample, using callipers. If the stem is very thin, determine the diameter on a cross-section of it under a microscope, using a calibrated ocular micrometer. Calculate the volume (V) of the cylinder as
In the case of hollow stems (e.g. young trees of Cecropia or some bamboo species), estimate the diameter of the hollow and subtract its cross-sectional area from the stem cross-sectional area before multiplying by L. This method can be applied to samples having different geometrical forms. After volume measurement (by any of the methods described above), the sample is dried in the oven. Besides free water, stems also contain bound water, which is removed only by drying at above 100°C. Samples should be dried in a well ventilated oven at 101–105°C for 24–72 h (small samples), until a constant weight is obtained. Large samples may need more drying time.
Additional useful methods from forestry
In forest ecological studies, samples are often taken with an increment borer, in which a wood core is cut from the bark inward, to just beyond the centre of the stem. Because a core does not taper inward towards the centre of the stem, such a sample may not be perfectly representative of the density of the stem as a whole; however, the difference from a density estimate using a sector or an entire stem section is usually probably small. Large-diameter corers (12 mm) are better because they cause less compaction. Samples with this method are usually taken at ~1.3 m above ground (‘breast height’). After the core is extracted, it can be stored in a plastic drinking straw, with the ends of the straw sealed.
In the timber industry, the ‘wood density’ is often measured at 12% moisture content, and density is reported as ‘air-dry weight’ (ADW) (a misnomer, because density is not weight, but weight/volume). SSD as described in the present protocol is called ‘oven dry weight’ (ODW). ADW can be transformed to ODW by using the formula ODW = 0.800 ADW + 0.0134 (R2 = 0.99) (this formula cannot be correct for ‘dry weights’, but only for densities). We suggest that data for ODW, directly measured or derived from ADW, can safely be used as SSD. This value ignores the contribution of the bark of a tree to its stem density; however, because bark usually makes up only a very small fraction of the mass of a large tree trunk, this error is probably unimportant, except as noted below.
Special cases or extras
-
Oven drying and wood-specific gravity. Because wood is mainly cellulose and lignin, containing substantial bound water and relatively small quantities of compounds of low molecular weight, many wood scientists and foresters oven-dry wood samples at 100–110°C, before determining both weight and volume. They then refer to the relationship between weight and volume as wood specific gravity.
-
Stems with holes. Very large holes in the stem are considered to be air or water spaces that do not belong to the stem tissue, whereas smaller spaces such as the lumens of xylem vessels, and intercellular spaces, are part of the stem tissue.
-
Wood density v. stem density. It might be worthwhile to make separate density estimates for wood and bark, because they have very different chemical and cellular compositions, physical properties and biological functions. Many trees and shrubs in savannah-type vegetation (and some Mediterranean and arid species) have, for example, a very thick corky bark (with very low density), and often the volume of the bark may be an important part (even 50%) of the stem volume. In these species, most of the structural support is given by the wood and wood is generally denser than bark. Thus, from the support viewpoint, wood density is the important parameter.
-
Xylem density. Some authors make a distinction between wood density (oven-dry wood density of the main trunk including sapwood and heartwood) from xylem density or sapwood density (measured on small, ~1.5-cm, terminal branches of trees). Xylem density has been proposed as a proxy for the tree hydraulic architecture, which, in turn, may limit tree performance in terms of transpiration, C exchange and growth.
-
Plants without a prominent above-ground stem (rosette plants, grasses and sedges). Try to isolate the short, condensed stem, near ground level, to which the leaves are attached, and obtain its density. In some plants, all the leaves are attached to an underground, often horizontal, modified stem called a rhizome (see Section 2.3), whose density can be determined, but which does not have the kind of mechanical leaf-supporting function that an above-ground stem has. Most rosette plants and basal-leaved graminoids produce aerial inflorescences, the density of whose stem can also be determined; however, this stem again usually has either no, or a reduced, function in supporting photosynthetic leaves, compared with that of an extensive-stemmed plant (see Section 2.3). If the plant has no recognisable above-ground leaf-supporting, stem, qualitatively recording it as ‘stemless’ is probably more convenient from the point of view of further analyses, than quantitatively recording its stem density as zero. If a plant branches from ground level (e.g. many shrubs), select the apparent main branch, or a random one if they are all similar.
-
Dense woods. When coring trees with very dense wood, a rope can be tied around the tree and fastened at the handle of the borer. When the handle is turned around during coring, the rope will wrap itself around the borer, increasing the tension on the rope, which helps push the borer bit into the tree.
-
Measuring other than main stem. If wood samples cannot be removed from a trunk or main stem, wood from branches 1–2 cm in diameter may be sampled. Main-stem wood density has been found on average to equal 1.411 × branch wood density, although this relationship can vary among species, so using it can sometimes introduce an error.
-
Components of stem volume (cell-wall material, water, gas). The volumetric fractions of cell-wall material, water and gas (‘air’) in the stem can be calculated as follows. The volume fraction of water is simply the decrease in weight (in g) on drying, divided by the original volume (in cm3) of the sample. The volume fraction that is cell-wall material equals the dry mass : fresh volume divided by the density of dry cell-wall material (1.53 g cm–3 for cell-wall material in dry wood); if an appreciable fraction of the stem volume is bark, using this number may involve an error because the density of bark cell-wall material is not necessarily the same as that for wood. The fraction of the original volume that was gas is simply 1 minus the foregoing two volume fractions.
References on theory, significance and large datasets: Putz et al. (1983); Loehle (1988); Reyes et al. (1992); Niklas (1994); Gartner (1995); Santiago et al. (2004); Van Gelder et al. (2006); Poorter et al. (2008); Chave et al. (2009); Patiño et al. (2009) Anten and Schieving (2010).
More on methods: Reyes et al. (1992); Brown (1997); Gartner et al. (2004); Chave et al. (2005); Swenson and Enquist (2008); Williamson and Wiemann (2010).
4.2 Twig dry-matter content and twig drying time
Twig dry-matter content (TDMC) is the oven-dry mass (mg) of a terminal twig, divided by its water-saturated fresh mass (g), expressed in mg g–1. Twig drying time is expressed in days (until equilibrium moisture). We consider TDMC to be a critical component of plant potential flammability, particularly fire conductivity after ignition (see Section 2.12). Twigs with high dry-matter content are expected to dry out relatively quickly during the dry season in fire-prone regions. TDMC should be positively correlated with specific density or dry-matter content of the main stem across woody species (see Section 4.1), and negatively correlated with potential RGR, although this has, to our knowledge, not been tested explicitly.
What and how to collect?
Collect one to three terminal (highest ramification-order; smallest diameter-class), sun-exposed twigs from a minimum of five plants. Twigs (or twig sections) should preferably be 20–30 cm long. If a plant has no branches or twigs, take the main stem; in that case, the procedure can be combined with that for SSD (see Section 4.1). For very fine, strongly ramifying terminal twigs, a ‘main twig’ with fine side twigs can be collected as one unit.
Storing and processing
Wrap the twigs (including any attached leaves) in moist paper and put them in sealed plastic bags. Store these in a cool box or fridge (never in a freezer!), until further processing in the laboratory. If no cool box is available in the field and temperatures are high, it is better to store the samples in plastic bags without any additional moisture; then follow the above procedure once back in the laboratory.
Measuring
Following the rehydration procedure (see Section 3.3), any leaves are removed and the twigs gently blotted dry with tissue paper, to remove any surface water before measuring water-saturated fresh mass. Each twig sample (consisting of 1–3 twigs) is then first dried in an oven or drying room at 40°C at 40% relative humidity of the outside air, or lower. Every 24 h, each sample is reweighed. Twig drying time is defined as the number of days it takes to reach 95% of the mass reduction of the sample as a result of drying (interpolating between weightings if necessary), where 100% is the final loss of mass when the weight of the sample ceases to decline further at the indicated temperature. Continue until you are certain that a steady dry weight has been reached. TDMC is defined (analogously to LDMC) as dry mass divided by saturated mass. The final dry weight obtained at 40°C is not the true dry mass of the twig, because some bound water will remain within the cell walls of the material (and probably also in its protein) at this temperature (see Section 4.1). However, the rather low drying temperature adopted here, compared with that in Section 4.1, is chosen so as to produce in twigs a dry condition relatively similar to that from air-drying outdoors, but which can be obtained relatively quickly.
Special cases or extras
-
Herbaceous plants. For herbaceous plants, the equivalent to TDMC is stem dry-matter content, which can be measured in exactly the same way as LDMC but using the main stem of forbs or the leaf sheaths of grasses.
References on theory and significance: Bond and Van Wilgen (1996); Lavorel and Garnier (2002); Shipley and Vu (2002).
More on methods: Garnier et al. (2001b).
4.3 Bark thickness (and bark quality)
Bark thickness is the thickness of the bark in mm, which is defined here as the part of the stem that is external to the wood or xylem – hence, it includes the vascular cambium. Thick bark insulates meristems and bud primordia from lethally high temperatures associated with fire, although the effectiveness depends on the intensity and duration of a fire, on the diameter of the trunk or branch, on the position of bud primordia within the bark or cambium and on bark quality and moisture. Thick bark may also provide protection of vital tissues against attack by pathogens, herbivores, frost or drought. In general, this trait has special relevance in trees or large shrubs subject to surface-fire regimes. Be aware that the structure and biochemistry of the bark (e.g. suberin in cork, lignin, tannins, other phenols, gums, resins) are often important components of bark defence as well.
What and how to collect?
Healthy, adult plants should be sampled as indicated above (see Section 1.1). Measure bark thickness on a minimum of five adult individuals, preferably (to minimise damage) on the same samples that are used for measurements of SSD (see Section 4.1). Measure this trait on main stem near the base between 10- and 40-cm height, because that is where surface fires occur (but see Special cases or extras in the present Section). If you do not use the same sample as for SSD, cut out a new piece of bark of at least a few centimetres wide and long. Avoid warts, thorns or other protuberances and remove any bark pieces that have mostly flaked off. The bark, as defined here, includes everything external to the wood (i.e. any vascular cambium, secondary phloem, phelloderm or secondary cortex, cork cambium or cork).
How to measure?
For each sample or tree, five random measurements of bark thickness are made with callipers (or special tools used in forestry), if possible to the nearest 0.1 mm. For species with fissured stems, see Special cases or extras in the present Section. In situ measurement with a purpose-designed forestry tool is an acceptable alternative. Take the average per sample. Bark thickness (mm) is the average of all sample means.
Special cases or extras
-
Bark quality. In addition to bark thickness, several structural or chemical components of bark quality may be of particular interest (see above within the present Protocol). An easy but possibly important one is the presence (1) v. absence (0) of visible (liquid or viscose) gums or resins in the bark.
-
Bark surface structure (texture) may determine the capture and/or storage of water, nutrients and organic matter. We suggest five broad (subjective) categories, including (1) smooth texture, (2) very slight texture (amplitudes of microrelief within 0.5 mm), (3) intermediate texture (amplitudes 0.5–2 mm), (4) strong texture (amplitudes 2–5 mm) and very coarse texture (amplitudes >0.5 mm). Bark textures may be measured separately for the trunk and smaller branches or twigs, because these may differ greatly and support different epiphyte communities.
-
Fissured stems. In each sample, take five random measurements of both the maximum (outside the fissure) and minimum (inside the fissure) bark thickness. Then calculate bark thickness as one-half the difference between them.
-
Alternative height for measurements. Typically, in forestry surveys, bark thickness is measured at breast height (as DBH). Measuring at the base of the tree, as suggested here, has advantages (more related to fire resistance) and problems (often the base of the tree is deformed). An alternative can be to make this measurement at ~50–60 cm (and in any case bark thickness at 50 cm is strongly related to bark thickness at breast height) or directly at DBH.
-
Decorticating bark. Decorticating bark is usually considered as standing litter, so it is not included in bark-thickness measurements (however, specific objectives may imply its measurement).
-
Bark investment. The complementary measurement of stem diameter can be useful to compare species for bark investments (by dividing the bark thickness by the stem radius).
References on theory and significance: Jackson et al. (1999); Brando et al. (2012).
4.4 Xylem conductivity
Water transport from soil to leaves is critical to land-plant activity. By replacing the water lost to the atmosphere via transpiration, water transport prevents highly negative and damaging leaf water potentials from developing, and permits continued photosynthesis. The efficiency of water transport is quantified as the stem-specific xylem hydraulic conductivity (KS), the rate of water flow per unit cross-sectional xylem area and per unit gradient of pressure (kg m–1 s–1 MPa–1). It can also be quantified as the LA-specific xylem hydraulic conductivity (KL), the rate of water flow per unit of supported LA per unit pressure gradient (kg m–1 s–1 MPa–1). KL expresses the conductivity of a branch, relative to the transpiration demand of the foliage that the branch services. Xylem refers here to the conducting tissue in the stem and, for trees, equates to sapwood. KL equals KS, divided by the ratio between LA and sapwood cross-sectional area (see Section 2.8). KS is a function of the numbers and conductivity of individual xylem conduits and their interconnections via pit membranes, per unit of xylem cross-sectional area. Conductivity of an individual conduit increases with the fourth power of its lumen diameter (as can be modelled using Poiseuille’s law of flow in cylindrical tubes).
What and how to collect
The measurement may conveniently be made indoors on stem samples brought in from the field, provided they are kept cool and with their ends in water. The tested branches should be longer than the length of the longest conducting element in the xylem, so that the measurement includes the resistivity of both the conduits and the inter-conduit connections. Conduit length varies greatly across species, gymnosperms having short tracheids (usually less than 1 cm long), whereas in some angiosperms, vessel length can reach more than a metre (although it is usually 10–30 cm, see Point 1 of Special cases or extras in the present Section). Segments ~30 cm long are commonly used. KS typically declines towards the leaves because of tapering of vessel numbers and sizes. In most cases, measurements are made on stems collected from the outer canopy, and so they can be considered the minimum conductivity for the stem part of the water-transport system.
Measuring
These methods have been developed largely for the stems of woody plants, for which the methods are simplest. Analogous methods have, however, been devised for herbaceous plants, leaves and roots. Relatively sophisticated types of apparatus for performing xylem-conductivity measurements have been described; however, in many cases, simple systems built from ordinary laboratory equipment can be used. In the simplest case, a known pressure head is applied to push a 10 mM KCl solution (use filtered, degassed water) through a stem with a known cross-sectional area and no (or with sealed-off) side branches. At the distal end of the segment, a collecting container catches emerging liquid, and after known time intervals, its volume is determined either directly or gravimetrically. Conductivity is then calculated as
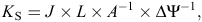
where J is the rate of water flow through the stem (kg s–1), L is the length of the segment (m), A is the mean cross-sectional area of the xylem of the stem (m2) (to a first approximation, the average of the areas at the two ends of the segment), and ΔΨ is the pressure difference (MPa) between the upstream and downstream ends of the segment. If the stem segment is held vertically during the measurement, its length (in m) divided by 10 should be added to the applied pressure (MPa) (but not if the segment is horizontal). KS is typically reported in kg m–1 s–1 MPa–1 With measurements on terminal or subterminal branch segments, one can usually assume that A is the entire xylem cross-sectional area (all of the xylem being conductive); however, with larger branches or tree trunks, A would have to be the conducting xylem area, at a maximum, the sapwood area; however, often an even smaller area is actually conductive, which is not easy to determine for routine measurements.
Xylem conductance refers to the capacity of a vascular system, with whatever length and cross-sectional area it happens to have, to transport water under a unit pressure difference. Conductance can be calculated from the above equation by simply omitting its A and L terms.
Special cases or extras
-
Length of vessels. This is needed to ensure that the length of stem segments used for conductance measurements exceeds that of their vessels. To determine the maximum length of the vessels, cut several stems in the field (see Fig. 7 for further details on the procedure). In the laboratory, from the upper end of one of these, remove a portion such that the segment that remains is likely to be a little longer than the length of its longest vessel. Proceed as indicated in Fig. 7.

|
References on theory, significance and large datasets: Zimmermann and Jeje (1981); Brodribb and Hill (2000); Meinzer et al. (2001); Zwieniecki et al. (2001); Tyree and Zimmermann (2002); Sperry (2003); Cavender-Bares et al. (2004); Maherali et al. (2004); Santiago et al. (2004); Holbrook and Zwieniecki (2005); Sperry et al. (2008a).
References on methods: Sperry et al. (1988); North and Noble (1992); Alder et al. (1996); Kocacinar and Sage (2003); Sack and Holbrook (2006).
4.5 Vulnerability to embolism
The xylem vulnerability to embolism indicates the risk of loss of water transport during drought. Vulnerability is expressed as the percentage of the water-saturated xylem conductance that is lost at given stem-water potentials. The water stream in the conduits is under tension, which can in some cases become as high as 100 times the atmospheric pressure. Any air entry into a water-conducting element will dissipate the tension in it and quickly expand (becoming an air embolism), effectively blocking water flow through that element. The more such emboli develop, the greater the loss of xylem conductance. The ability of a species to tolerate highly negative water potentials (= high tensions) without embolising varies greatly among species (cf. Section 3.15) and is an important aspect of drought tolerance.
What and how to collect
Follow the above instructions for collecting for measurement of xylem conductivity.
Measuring
Vulnerability to embolism is quantified by constructing a xylem vulnerability curve. This consists of plotting measured values of xylem conductance, on the y-axis, against the values of stem water potential (Ψ), on the x-axis, at which these conductance values (represented as a percentage of the maximum water-saturated conductance) were obtained. The shape of this curve is usually sigmoid. To characterise this curve by one number, the value of Ψ at its mid-point (50% loss of conductance) is commonly used.
Stem segments with different Ψ values can be obtained by any of three possible methods.
-
Evaporative dehydration. A large branch that includes some lateral secondary branches is cut from the plant and kept unwrapped during the day, or longer, for its xylem to partially dehydrate, and develop tension as a result of transpiration from its leaves. Ψ is determined periodically, by the pressure chamber method (cf. Section 3.15), on secondary branches that are removed at intervals for this purpose. When a target Ψ has been reached, conductance of a segment of the main axis near the last-removed lateral is measured. Water is then flushed through this segment briefly under a pressure high enough to displace all air embolisms from it (see references cited below), and its conductance is re-measured to obtain the maximum conductance of the segment. This affords a value for the percentage loss of conductance at that particular Ψ. Separate branches are similarly tested to obtain conductance-loss values for other values of Ψ.
-
Centrifugation. Attaching a short stem segment horizontally (and symmetrically) across the top of a centrifuge rotor and spinning it generates tension in the water within the xylem conduits. From the centrifugal force that was applied, a corresponding (negative) Ψ or P can be inferred. Advantages of the method are that stem segments can be brought to different effective water potentials very quickly. However, the technique is difficult for stem segments longer than ~20 cm, or ones that cannot be reliably attached to an available rotor, and it cannot be used for segments longer than the width of the rotor chamber. Details needed for actually employing this method are given in some of the cited references.
-
Air injection. This simple method is based on the principle of substituting internal tension by external positive air pressure, which is applied to a stem segment that is located within a pressure chamber. The procedure uses a special pressure chamber designed for a stem segment to pass completely through it, allowing a conductance measurement while the external pressure is applied. This type of chamber is commercially available from the PMS Instrument Co., Albany, Oregon, USA (http://pmsinstrument.com, accessed 15 February 2013).
References on theory, significance and large datasets: Tyree and Sperry (1989); Davis et al. (2002); Brodribb et al. (2003); Maherali et al. (2004); Holbrook and Zwieniecki (2005); Choat et al. (2007); Feild and Balun (2008); Sperry et al. (2008a).
References on methods: Cochard et al. (1992); Sperry et al. (1988, 2008b); Alder et al. (1997); Pammenter and Vander Willigen (1998).
5 Below-ground traits
Variation in root traits among species has large ramifications for their ecology. Fine roots are the primary organs for water and nutrient acquisition and they are also responsible for transferring resources between below-ground and above-ground parts. Fine roots can acquire resources directly or through symbionts. When accessing soil resources directly, fine roots selectively acquire mineral nutrients from a complex range of soil solutions and soil particles, while modifying soil chemistry via exudation of a range of compounds. Roots also must respond to heterogeneity of resource availability at multiple spatial and temporal scales, while resisting attacks from a wide range of organisms and environmental stresses.
There is no single evolutionary solution to all the variable challenges of acquiring soil resources in different ecosystems. Measuring several independent root traits in combination may help better understand the below-ground strategies of multiple species. Here, we describe three main sets of traits.
5.1 Specific root length
Specific root length (SRL), the ratio of root length to dry mass of fine roots, is the below-ground analogue of SLA (see Section 3.1), providing a ratio of a standard unit of acquisition (root length) to resource investment (mass). Plants with high SRL build more root length for a given dry-mass investment and are generally considered to have higher rates of nutrient and water uptake (per dry mass), shorter root lifespan and higher RGRs than for low-SRL plants. Yet, high SRL can result from having a low diameter or low tissue density, each of which is independently associated with different traits. For example, thin roots exert less penetrative force on soil and transport less water, whereas roots with low tissue density have lower longevity but greater rates of uptake under high nutrient conditions. As there is little operational difference in measuring just SRL or its two components, we recommend that both metrics be measured when measuring the functional traits of roots.
What and how to collect?
Roots are often measured in aggregate when comparing the live fine roots of plants, although individual fine roots or small numbers of fine roots can be enough to measure root diameter or branching order. Separating fine roots according to the timing or depth of sampling could be informative to answer particular questions. Roots measured in the field span a range of root age, whereas roots acquired from ingrowth cores or young plants would constrain age. The basis of comparison should be clear and root acquisition and preparation considered each time. Roots from the top 20 cm are the standard basis of comparison; however, the actual depth sampled should be allowed to be as varied as the height above ground from which to collect leaves.
In mixed-species assemblages, fine roots should be traced back to shoots for positive identification. This is not necessary in uniform stands and roots can often be distinguished among a small number of species. For small plants, it is often most feasible to excavate the entire plant to be washed out later, aiding in identification. A typical amount of root necessary for measurement generally fits in the palm of your hand. In general, it is better to have a small amount of root that is better prepared than a larger amount of less well prepared root. Preferably, atypically large or small individuals should be avoided.
Storing and processing
Unwashed roots can generally be stored under humid, chilled conditions for a week, with little degradation of structure. Washing techniques should be gentle for species with low-density roots, whereas more rigorous washing might be more suitable for high-density roots in soils with heavy clays or coarse organic matter that could compromise measurements. Washing roots from a sandy soil can require as little as 30 s under a hose, whereas clearing roots of organic matter from a tundra soil might require hours of painstaking plucking. In general, cleaning roots will require a combination of running water over a fine mesh sieve (0.2–1 mm) to remove fine heavy particles such as sand, rinsing in containers of water to remove coarser heavy particles such as pebbles, and plucking of debris with forceps to remove contaminants that are of a similar size and density as the roots of interest. Often roots have to be finger-massaged and individual roots separated to allow particles to be removed. If some fine particles such as clays are too difficult to remove, roots can be ashed at 650°C later and ash mass subtracted from gross root dry mass. Washed roots can be stored in a 50% ethanol solution for longer periods of time. A useful rule of thumb is to stop washing roots when it appears that you are losing as much of the fine roots as you are removing soil, or preferably slightly before.
Measuring
If necessary, under a dissecting microscope, sort apparently live, healthy roots from the recently washed sample. Live roots generally have a lighter, fully turgid appearance, compared with dead or dying roots of the same species which appear darker and floppy or deflated; however, note that live and dead roots may not be distinguishable by colour. It will help to observe a range of ages and colours of absorptive roots for each plant species before measurement, so as to properly identify healthy live roots. For woody species, roots are often divided by root (ramification) order, to better standardise comparisons across species.
Once roots have been obtained and prepared, determining SRL, diameter and tissue density requires digitising the roots and measuring their length and diameter. Digitisation can be carried out with almost any low-end flatbed scanner. A scanner with a resolution of 1600 dpi provides a resolution of 15 µm, which is still half the width of the finest roots of any plant. Nevertheless, a scanner with lower resolution may also work. A scanner that has a transparency adaptor that illuminates items on the scanner bed from above, is recommended to provide crisp root images. Roots are best imaged while submerged in a small amount of water, which also aids in teasing individual roots apart. A clear plastic tray works well. There should be no need to stain most roots to image them. After scanning, scanned roots should be dried (48 h at 60°C) and weighed. These root samples can also be ground and analysed for nutrient concentrations.
When roots have been scanned, units of root length need to be traced and their diameter determined. For a small number of roots, this can be carried out with image-analysis software (see Section 3.1 for free software). For a large number of samples or root length, the commercially available application, WinRhizo (Régent Instruments, Quebec, Canada), is recommended. The software will automatically determine the length, diameter and root volume distribution of a sample of root length, enabling easy calculations of SRL, average root diameter and root tissue density (root dry mass over volume, the latter being derived from length and radius). Although the software is expensive for occasional use, roots can be scanned independent of analysis software, saved in the JPEG format and analysed later by someone who owns the software. See under Special cases and extras in the present Section for manual methods when none of the above facilities is available.
Special cases or extras
-
Root diameter and tissue density. Not all roots of a given diameter and tissue density have similar cellular structure. Roots can vary in their relative proportions of cortex and stele (mainly phloem and xylem) as well as the construction of each. For these reasons, secondary to measuring gross morphology of fine roots, we also recommend cross-sectioning roots, so as to determine their cellular structure. For this purpose, multiple roots of each species are embedded in a polymer, cut on a microtome into 4-µm slices, stained with toluidine blue, which stains lignin blue–green and cellulose purple or red–violet, and then mounted on a glass slide. Digital images are made for each species using light microscopy at ×100 magnification and the cross-sectional areas of the root, stele, endodermis and large xylem elements are determined by tracing each portion of the root manually in image-analysis software. With these data, cell diameters and amounts of different tissues can be calculated relative to one another and to total cross-sectional area.
References on theory, significance and large datasets: Eissenstat and Yanai (1997); Wahl and Ryser (2000); Steudle (2001); Pregitzer et al. (2002); Roumet et al. (2006); Craine (2009); Paula and Pausas (2011).
More on methods: Newman (1966); Tennant (1975); Böhm (1979); Fitter (1996); Bouma et al. (2000); Craine et al. (2001); Craine (2009).
5.2 Root-system morphology
Characteristics of entire root systems can be independent of individual roots and need to be measured explicitly. There are three main traits of root systems that are best measured: depth, lateral extent, and intensity of exploration. For rooting depth, the simplest metric to determine is maximum rooting depth (maximum soil depth from which resources can be acquired, ranging from a few centimetres to tens of metres). The maximum lateral extent of roots defines the distance from the centre of the plant that roots can acquire resources from. It also determines the ability of plants to interact with spatial heterogeneity in soil resources. The amount of fine-root biomass or root length per unit soil volume indicate the intensity of soil exploration, and the ability of a species to compete for soil nutrients.
The depth distribution of roots combines depth and intensity of utilisation of soil. Depth distributions are a better indicator of the relative reliance of plants on different depths for soil resources and define their vertical distribution of influence on soil activity. In general, it is simpler to determine the root biomass with different depths, whereas understanding root length with depth is likely to be a better metric to understand the competitive ability of uptake capacity, for example. In general, biomass and length with depth would be strongly correlated if there were no change in SRL with depth.
Note that root tissue density and root diameter are positively related to longevity and negatively related to nutrient uptake. In addition, root tissue density is positively related to resistance to pathogens and drought.
Collection and analysis
Determining the maximum extent of roots depends on the species. Excavating entire plants is reasonable for some shallow-rooted species. For more deep-rooted species, a pit must be dug and a cross-section of the soil from a pit face excavated. In some extreme cases, roots have to be accessed from caves or boreholes.
Depth distributions can be determined by digging pits if a known cross-sectional area can be excavated with depth. With pits, a deep pit is dug and one pit face is smoothed vertically. Then a cross-section is removed with a flat shovel. Root systems can be removed in entirety or in sections. In other cases, an auger of 5–10-cm diameter should be used to remove biomass with depth. Typical depth distributions follow a somewhat exponential relationship. A standard set of depths would be 5, 10, 20, 40, 80, 120 and 200 cm for root systems largely confined to the top 2 m of soil. Incomplete root-depth distributions can be used to estimate maximum rooting depths; however, this will depend on the pattern of root biomass with depth. For some species, depth distributions can be determined randomly relative to the individual, or at a point that represents the midpoint of its lateral extent, whereas biomass will have to be determined directly below individuals for species with a tap root. To determine lateral extent of root systems, a horizontal strip of soil can be excavated, starting at the centre of the plant, so as to trace roots outward. In other cases, where individuals are bunched, the lateral extent of roots is likely to be equivalent to half the interplant distance, although this should be verified.
Once soils have been excavated, root storage and washing use the same protocols as described above. Intact root systems are best laid out on a large mesh screen, to be washed out with running water, and/or submerged in large tubs. Confirming the identity of species might require anatomical or molecular comparisons with other roots of that species; however, it is most easily carried out by tracing roots back to their above-ground parts or sampling in conspecific stands. If depth distributions are to be determined, fine (<2 mm) and coarse roots should be separated. Subsamples of cleaned fine roots can then be scanned, if desired, for diameter, length and volume analyses. Regardless, root biomass should be dried and weighed for biomass distributions.
Special cases or extras
-
Large shrubs and trees. When sampling larger shrubs and trees, the researcher will encounter thicker woody roots. The best way to deal with this is to use a specialised wood-cutting auger. Within the coarse root fraction, those root sections that are obviously particularly important for mechanical support or resource storage, usually exceeding 10 mm in diameter, are best kept separate from the relatively thin sections. They can still be combined for certain analyses later on.
-
Clayey soil. If the soil is particularly clayey, aggregated, or contains calcium carbonate, consider adding a dispersal agent (e.g. sodium hexametaphosphate) to the washing water. The best washing additive varies depending on the particular condition of the soil.
References on theory and significance: Adiku et al. (2000); Zwieniecki et al. (2002); Hodge (2004); Schenk and Jackson (2002); Dunbabin et al. (2004); Withington et al. (2006); Craine (2009); Lambers et al. (2011).
More on methods: Böhm (1979); Caldwell and Virginia (1989); Jackson (1999); Linder et al. (2000); Schenk and Jackson (2002).
5.3 Nutrient-uptake strategy
The performance of different species in nutrient-limited ecosystems is undoubtedly affected by their inherent interspecific differences in nutrient-uptake capacity. One factor affecting this is SRL (see Section 5.1); however, quantitative differences in the affinities of the ion-transport carrier and in the capacities of the roots among different species are superimposed on this. Furthermore, many plants exploit symbiotic associations with bacteria or fungi to enhance their nutrient-competitive ability, which is the subject of the present Section.
Symbioses with N2-fixing bacteria or mycorrhizal fungi have been found in many plant species. However, literature is strongly biased towards temperate species of Europe and North America. There is still much to be learned about nutrient-uptake strategies of species in less studied regions, as exemplified by the recent discovery of specialised N-foraging snow roots in the Caucasus. We provide specific protocols in Material S2 that deal with the following strategies:
-
N2-fixing bacteria – association with bacteria in nodules to fix atmospheric N2;
-
arbuscular mycorrhizae – symbiosis with arbuscular mycorrhizal fungi to aid in acquisition of nutrients and water;
-
ecto-mycorrhizae – symbiosis with ecto-mycorrhizal fungi to aid in uptake of inorganic nutrients and organic forms of N;
-
ericoid-mycorrhizae – symbiosis with ericoid mycorrhizal fungi to aid in uptake of organic forms of N;
-
orchids – symbiosis with orchid mycorrhizal fungi for acquiring nutrients from litter;
-
rarer types of mycorrhizae, e.g. arbutoid mycorrhizae (Arbutus, Arctostaphylos), ecto-endo-mycorrhizae (certain gymnosperms) and pyroloid mycorrhizae (Pyrolaceae);
-
myco-heterotrophic plants without chlorophyll that extract C and probably most nutrients from dead organic matter via mycorrhizal fungi;
-
root or stem-hemiparasitic green plants, such as mistletoes (Loranthaceae), that extract nutrients such as N and P from the roots or stems of a host plant;
-
holoparasitic plants without chlorophyll that extract C and nutrients directly from a host plant;
-
carnivorous plants that capture organic forms of N and P from animals;
-
hairy root clusters (proteoid roots), dauciform roots in sedges and capillaroid roots in rushes, that aid in P uptake;
-
other specialised strategies (mostly in epiphytes), including
-
-
tank plants (ponds) – nutrient and water capture and storage;
-
baskets – nutrient and water capture and storage;
-
ant nests – nutrient uptake and storage,
-
trichomes – nutrient and water capture through bromeliad leaves, and
-
root velamen radiculum – nutrient and water capture and storage; and
-
-
none – no obvious specialised N- ort P-uptake mechanism; uptake presumably directly through root hairs (or through leaves, e.g. in the case of certain ferns with very thin fronds).
Although most of the experimental work on the active uptake of nutrient ions by roots has been carried out on agricultural plants, some information on that of wild plants is available, but again relatively little for tropical and subtropical species. The radioisotope techniques for characterisation of ion-transport mechanisms are beyond the scope of the present handbook, but can be readily ascertained by consulting the ion-transport literature.
References on theory and significance: see Material S2; in addition, for snow roots, see Onipchenko et al. (2009).
6 Regenerative traits
6.1 Dispersal syndrome
The mode of dispersal of the ‘dispersule’ (or propagule = unit of seed, fruit or spore as it is dispersed) has obvious consequences for the distances it can cover, the routes it can travel and its final destination.
How to classify?
This is a categorical trait. Record all categories that are assumed to give significant potential dispersal (see Box 4), in order of decreasing importance. In the case of similar potential contributions, prioritise the one with the presumed longer-distance dispersal; e.g. wind dispersal takes priority over ant dispersal.
| Box 4. Dispersal syndromes |
|
It is important to realise that dispersules may (occasionally) get transported by one of the above modes even though they have no obvious adaptation for it. This is particularly true for endo-zoochory and exo-zoochory. Note that there is ample literature (e.g. in Floras) for dispersal mode of many plant taxa.
References on theory, significance and large datasets: Howe and Smallwood (1982); Van der Pijl (1982); Bakker et al. (1996); Howe and Westley (1997); Hulme (1998); Poschlod et al. (2000); McIntyre and Lavorel (2001); Tackenberg et al. (2003); Myers et al. (2004).
More on methods: Howe and Westley (1997); Forget and Wenny (2005); Pons and Pausas (2007).
6.2 Dispersule size and shape
Of interest is the entire reproductive dispersule (= dispersal structure or propagule) as it enters the soil. The dispersule may correspond with the seed; however, in many species, it constitutes the seed plus surrounding structures, e.g. the fruit. Dispersule size is its oven-dry mass. Dispersule shape is the variance of its three dimensions, i.e. the length, the width and the thickness (breadth) of the dispersule, after each of these values has been divided by the largest of the three values. Variances lie between 0 and 1 and are unitless. Small dispersules with low shape values (relatively spherical) tend to be buried deeper into the soil and live longer in the seed bank. Seed size and shape are then fundamental for seed persistence in the soil (seed-bank persistence).
What and how to collect?
The same type of individuals as for leaf traits and plant height should be sampled. Of interest is the unit that is likely to enter the soil. Therefore, only parts that fall off easily (e.g. pappus) are removed, whereas parts such as e.g. wings and awns remain attached. The flesh of fleshy fruits is removed too, because the seeds are usually the units to get buried in this case (certainly if they have been through an animal gut system first). The seeds (or dispersules) should be mature and alive. The dispersules can either be picked off the plant or be collected from the soil surface. In some parts of the world, e.g. in some tropical rain forest areas, it may be efficient to pay local people specialised in tree climbing (and identification) to help with the collecting.
Storing and processing
Store the dispersules in sealed plastic bags and keep in a cool box or fridge until measurement. Process and measure as soon as possible. For naturally dry dispersules, air-dry storage is also okay.
Measuring
Remove any fruit flesh, pappus or other loose parts (see above in the present Section). For the remaining dispersule, take the highest standardised value for each dimension (length, width and thickness) using callipers or a binocular microscope and calculate the variance. Then dry at 60°C for at least 72 h (or else at 80°C for 48 h) and weigh (= dispersule size).
Special cases or extras
We recommend complementing this trait with other direct or indirect assessment of banks of seeds or seedlings for future regeneration of a species. For seed-bank assessment, there are good methods to follow (see More on methods below in the present Section); however, (above-ground!) canopy seeds banks of serotinous species of fire-prone ecosystems (e.g. Pinus and Proteaceae such as Banksia, Hakea and Protea) and long-lived seedling banks of woody species in the shaded understorey of woodlands and forests may also make important contributions. Vivipary as in some mangroves could also be part included in such assessments.
References on theory, significance and large datasets: Hendry and Grime (1993); Thompson et al. (1993, 1997); Leishman and Westoby (1998); Funes et al. (1999); Weiher et al. (1999); Peco et al. (2003).
More on methods: Hendry and Grime (1993); Thompson et al. (1993, 1997); Weiher et al. (1999); Pons and Pausas (2007).
6.3 Dispersal potential
Dispersal potential is defined as the proportion of dispersules produced by one individual that travelz a certain distance, which can be chosen arbitrarily depending on the question. The dispersules may be seeds or fruits or vegetative propagules. In contrast to dispersal syndrome, dispersal potential allows the assessment of dispersability of a seed in relation to distance. It varies not only among species, but also strongly among species with the same dispersal syndrome. Therefore, it is a crucial variable when asking if dispersal is limiting the occurrence of a species in suitable habitats or species richness of plant communities, or if fragmentation is a threat to the survival of species or populations. The capacity to survive in disturbed habitats or in fragmented landscapes is often correlated with a high dispersal potential. Both seed production and also seed characters may be correlated with dispersal potential. The more seeds are produced, the higher the probability that one seed spans larger distances. The seed characters such as e.g. mass, form and structure of seed surface responsible for a high dispersal potential depend on the dispersal vector. There may be a trade off between dispersal potential (in space) and maximum plant lifespan as well as seed-bank persistence (dispersal in time). Long-lived species often exhibit a low dispersal potential, as do species with a long-term persistent seed bank.
How to record?
Dispersal potential is a continuous variable and may be recorded either by direct measurements in the field or can be identified by measurements of traits related to the dispersal potential, or by modelling approaches. Wind-dispersal potential is correlated with dispersule-releasing height and terminal velocity, dispersal potential by water to buoyancy of the dispersules and animal-dispersal potential to either attachment potential or survival after digestion. Dispersal by humans, machines or vehicles is very complex. Measuring dispersal potential, therefore, requires studies adapted to the specific question.
Measurements should be carried out on the intact dispersule, i.e. seed or fruit with all the structures, such as e.g. pappus and awns, that are still attached when it is released. Releasing height should be measured during dispersule release and is the difference between the highest elevation of the seed or fruit and the base of the plant. Terminal velocity is measured on freshly collected air-dry dispersules and, most simply, by the actual rate of fall in still air. Floating capacity (proportion of dispersules floating after a defined time) is measured by putting dispersules in glass beakers that are placed on a flask shaker moving with a frequency of 100 min–1. Attachment capacity (proportion of dispersules still attached after a defined time) is measured by putting seeds on the respective animal fur, which is then shaken by a shaking machine. Survival after digestion is measured either by digestion experiments with the respective animals or by simulating ingestion by a standardised mechanical treatment and digestion by a standardised chemical treatment, which have to be calibrated by digestion experiments.
To assess animal-dispersal potential, field studies should be added where possible, because the behaviour of animals (e.g. selection of species by grazing animals) strongly influences dispersal potential. Predicting animal-dispersal potential requires process-based models with the ability to predict over a range of scenarios.
Special cases or extras
-
For water plants, seed releasing height is the distance between the highest point of seeds or fruits and water surface.
-
Secondary process, e.g. dispersal by wind on the ground, may strongly affect dispersal potential. Such processes are often obvious only from field studies and may require the establishment of additional new methods.
References on theory and significance: Bruun and Poschlod (2006); Poschlod et al. (1998, 2005) Tackenberg (2003); Tackenberg et al. (2003); Schurr et al. (2005); Will and Tackenberg (2008); Cousens et al. (2010).
More on methods: Fischer et al. (1996); Römermann et al. (2005a, 2005b, 2005c).
6.4 Seed mass
Seed mass, also called seed size, is the oven-dry mass of an average seed of a species, expressed in mg. Stored resources in large seeds tend to help the young seedling to survive and establish in the face of environmental hazards (e.g. deep shade, drought, herbivory). Smaller seeds can be produced in larger numbers with the same reproductive effort. Smaller seeds also tend to be buried deeper in the soil, particularly if their shape is close to spherical, which aids their longevity in seed banks. Interspecific variation in seed mass also has an important taxonomic component, more closely related taxa being more likely to be similar in seed mass.
What and how to collect?
The same type of individuals as for leaf traits and plant height should be sampled, i.e. healthy adult plants that have their foliage exposed to full sunlight (or otherwise plants with the strongest light exposure for that species). The seeds should be mature and alive. If the shape of the dispersal unit (e.g. seed, fruit) is measured too (see Section 6.2 above), do not remove any parts until dispersule measurement is finished. We recommend collecting at least 10 seeds from each of 10 plants of a species, although more plants per species is preferred. Depending on the accuracy of the balance available, 100 or even 1000 seeds per plant may be needed for species with tiny seeds (e.g. orchids).
In some parts of the world, e.g. in some tropical rain forest areas, it may be efficient to work in collaboration with local people specialised in tree climbing to help with collecting (and identification).
Storing and processing
If dispersule shape is also measured, then store cool in sealed plastic bags, whether or not wrapped in moist paper (see Section 3.1), and process and measure as soon as possible. Otherwise air-dry storage is also appropriate.
Measuring
After measurements of dispersule shape (if applicable), remove any accessories (wings, comas, pappus, elaiosomes, fruit flesh), but make sure not to remove the testa in the process. In other words, first try to define clearly which parts belong to the fruit as a whole and which belong strictly to the seed. Only leave the fruit intact in cases where the testa and the surrounding fruit structure are virtually inseparable. Dry the seeds (or achenes, single-seeded fruits) at 80°C for at least 48 h (or until equilibrium mass in very large or hard-skinned seeds) and weigh. Be aware that, once taken from the oven, the samples will take up moisture from the air. If they cannot be weighed immediately after cooling down, put them in the desiccator until weighing, or else back in the oven to dry off again. Note that the average number of seeds from one plant (whether based on five or 1000 seeds) counts as one statistical observation for calculations of mean, standard deviation and standard error.
Special cases or extras
-
Within individual variation. Be aware that seed size may vary more within an individual than among individuals of the same species. Make sure to collect ‘average-sized’ seeds from each individual, and not the exceptionally small or large ones.
-
Available databases. Be aware that a considerable amount of published data are already available in the literature, and some of the large, unpublished databases may be accessible under certain conditions. Many of these data can probably be added to the database; however, make sure the methodology used is compatible.
-
Seed volume. There are also many large datasets for seed volume, often measured as π/6 × L1 × L2 × L3 (i.e. assuming an ellipsoidal shape). Most of these databases actually include both seed mass and volume. Using the appropriate calibration equations, those data can be also successfully used.
-
Additional measurements. For certain (e.g. allometric) questions, additional measurements of the mass of the dispersule unit or the entire infructescence (reproductive structure) may be of additional interest. Both dry and fresh mass may be useful in such cases.
References on theory, significance and large datasets: Mazer (1989); Seiwa and Kikuzawa (1996); Reich et al. (1998); Cornelissen (1999); Leishman et al. (2000); Westoby et al. (2002); Moles et al. (2005); Moles and Westoby (2006); Wright et al. (2007).
More on methods: Hendry and Grime (1993); Thompson et al. (1997); Westoby (1998); Weiher et al. (1999); Wright et al. (2007).
6.5 Seedling functional morphology
Seedling functional types refer to morphology of seedlings in relation to cotyledon function and position. It is a categorical trait that can be used to characterise plant regenerative strategies. The distribution of seedling traits across families is still rather poorly known, although the importance of seedling traits in systematics was recognised quite early. This trait has been created on the basis of woody species (trees and shrubs) and has been mainly used in tropical forests. Garwood (1996) established the following five seedling categories based on three cotyledon characters of presumed ecological significance (position, texture and exposure) (Fig. 8):

|
-
cryptocotylar hypogeal with reserve storage cotyledons (CHR),
-
cryptocotylar epigeal with reserve storage cotyledons (CER),
-
phanerocotylar epigeal with foliaceous cotyledons (PEF),
-
phanerocotylar epigeal with reserve storage cotyledons (PER) and
-
phanerocotylar hypogeal with reserve storage cotyledons (PHR).
These categories result from the combination of different possibilities in relation to cotyledon exposure (phanerocotylar or cryptocotylar), position (epigeal or hypogeal) and function (foliaceous or reserve storage, see below within the present Protocol). Although eight combinations are potentially possible, cryptocotylar foliaceous seedling types are biologically not possible, and phanerocotylar hypogeal foliaceous seedlings have not yet been reported. Seedling functional types are correlated with other plant traits such as seed size; e.g. large seed sizes are related to reserve storage seedling types, whereas small seed sizes are related to foliaceous and photosynthetic cotyledons. Because the above-mentioned types have been particularly identified for tropical forests, the occurrence and proportion of each type in other ecosystems should be tested.
What and how to collect?
Because this is not a plastic trait, we recommend germinating sufficient number of seeds to obtain about five seedlings per species. Seeds without evidence of, for example, pathogen damage and predation must be selected among the collected ones to run the experiments.
Measuring
Seedling morphology (cotyledon exposure, position and function) should be described when at least five individuals have developed at least three leaves each. Species are then assigned to the seedling morphological categories indicated above in the present Section (and in Fig. 8):
-
cotyledon exposure – phanerocotylar if the seed coat opens and two cotyledons emerge from seed, cryptocotylar if cotyledons remain within the seed coat;
-
cotyledon position – epigeal when hypocotyl develops at least 2 cm above soil surface, hypogeal when hypocotyl develops on the soil surface; or
-
cotyledon function – reserve cotyledon are fleshy, foliaceous cotyledons (also called paracotyledons) are primarily photosynthetic.
Special cases or extras
-
Chlorophyll in fleshy cotyledons. In many cases fleshy cotyledons do contain chlorophyll; however, they are still considered reserve organs (e.g. Aspidosperma spp.).
References on theory, significance and large datasets: Ng (1978); De Vogel (1980); Hladik and Miquel (1990); Garwood (1996); Kitajima (1996); Wright et al. (2000); Ibarra-Manriquez et al. (2001); Zanne et al. (2005); Leck et al. (2008).
6.6 Resprouting capacity after major disturbance
The capacity of a plant species to resprout after destruction of most of its above-ground biomass is an important attribute for its persistence in ecosystems where recurrent major disturbances are common. Fire (natural or anthropogenic), hurricane-force wind and logging are the most obvious and widespread major disturbances; however, extreme drought or frost events, severe grazing, browsing or trashing by large herbivores, landslides, flooding and other short-term large-scale erosion events also qualify. There appear to be ecological trade-offs between sprouters and non-sprouter plants. Compared with non-sprouters, sprouters tend to show a larger allocation of carbohydrates to below-ground organs (or storage organs at soil-surface level); however, their biomass growth tends to be slower, and their reproductive output lower. The contribution of sprouters to species composition tends to be associated with the likelihood of major biomass-destruction events, as well as to the degree of stress in terms of available resources.
How to assess?
Here, we define resprouting capacity as the relative ability of a plant species to form new shoots after destruction of most of its above-ground biomass, using reserves from basal or below-ground plant parts. The following method is a clear compromise between general applicability and rapid assessment on the one hand and precision on the other. It is particularly relevant for woody plants and graminoids, but may also be applied to forbs. Within the study site, search for spots with clear symptoms of a recent major disturbance event. In general, this event should have been within the same year. However, if only woody species are being considered, the assessment may be carried out up to 5 years after the disturbance (as long as shoots emerging from near the soil surface can still be identified unambiguously as sprouts following biomass destruction). For each species, try to find any number of adult plants between 5 and 50 individuals (depending on time available) from which as much as possible but at least 75% of the live above-ground biomass was destroyed, including the entire green canopy. This is to ensure that regrowth is only supported by reserves from basal or below-ground organs. Note that in the case of trunks and branches of woody plants, old, dead xylem (wood) is not considered as part of the live biomass. Thus, if a tree is still standing after a fire, but all its bark, cambium and young xylem have been killed, it should be recorded as destruction of 100% of the above-ground biomass.
Make sure that enough time has lapsed for possible resprouting. Estimate (crudely) the average percentage of above-ground biomass destroyed among these plants (a measure of disturbance severity) by comparing against average undamaged adult plants of the same species. Multiply this percentage by the percentage of the damaged plant population that has resprouted (i.e. formed new shoots emerging from basal or below-ground parts) and divide by 100 to obtain the ‘resprouting capacity’ (range 0–100, unitless). When data are available from more than one site, take the highest value as the species value, although this ignores the fact that great intraspecific variability in sprouting capacity may occur. In longer-term studies, resprouting may be investigated experimentally by clipping plants to simulate destruction of 75–100% of the above-ground biomass (in which case, the clipped parts can be used for other trait measurements as well). If fewer than five plants with ‘appropriate’ damage can be found, give the species a default value of 50 if any resprouting is observed (50 being halfway between ‘modest’ and ‘substantial’ resprouting, see below). In species where no resprouting is observed merely because no major biomass destruction can be found, it is important to consider this a missing value (not a value of zero).
Broad interspecific comparisons have to take into account an intraspecific error of up to 25 units as a result of the dependence of resprouting capacity on the severity of disturbance encountered for each species. However, within ecosystems where different species suffer the same disturbance regime, direct comparisons should be safe.
Special cases or extras
-
Data from literature. Useful and legitimate data may be obtained from the literature or by talking to local people (e.g. foresters, farmers, rangers). Make sure that the same conditions of major destruction of above-ground biomass have been met. In such cases, assign subjective numbers for resprouting capacity after major disturbance as follows: 0, never resprouting; 20, very poor resprouting; 40, moderate resprouting; 60, substantial resprouting; 80, abundant resprouting; and 100, very abundant resprouting. The same crude estimates may also be used for species for which the more quantitative assessment is not feasible, e.g. because the non-resprouting individuals are hard to find after disturbance, as is common in some herbs.
-
Strongly clonal plants. In the case of strongly clonal plants, it is important to assess whether damaged ramets can resprout from below-ground reserves and not from the foliage of a connected ramet. Therefore, in such species, resprouting should be recorded only if most above-ground biomass has been destroyed for all ramets in the vicinity.
-
Resprouting of young plants. Additional recording of resprouting ability of young plants may reveal important insights into population persistence, although this could also be seen as a component of recruitment. Thus, data on the age or size limits for resprouting ability may reveal important insights into population dynamics. It is known that some resprouting species cannot resprout before a certain age or size, and others may lose their resprouting capacity when they attain a certain age or size.
-
Resprouting after smaller biomass destruction. Additional recording of resprouting after less severe biomass destruction may provide useful insights into plant response to disturbances. For instance, Quercus suber and many Eucalyptus spp. can resprout from buds located in high positions along the stem, following a fire. Be aware that species highly adapted to fire (such as these examples) may give the false impression that an area has not been exposed to severe fires recently. Other species in the same area, or direct fire observations, should provide the evidence for that. The approach of recording resprouting after less severe biomass destruction could also be applied to the study of resprouting in the face of disturbances other than fire, such as herbivory or trashing by vertebrates.
References on theory, significance and large datasets: Noble and Slatyer (1980); Everham and Brokaw (1996); Pausas (1997); Kammesheidt (1999); Bellingham and Sparrow (2000); Bond and Midgley (2001); Higgins et al. (2000); Del Tredici (2001); Burrows (2002); Vesk and Westoby (2004); Pausas and Bradstock (2007); Poorter et al. (2010).
Acknowledgements
This contribution was funded, in part, by the Inter-American Institute for Global Change Research (IAI) CRN 2015 and SGP-CRA2015, which were supported by US National Science Foundation Grants GEO-0452325 and GEO-1138881 The work was also funded by FONCyT (PICT 20441 and 365), CONICET (PIP 2006-2011 11220080101532), SECYT-Universidad Nacional de Córdoba – Argentina, and the US NSF (DEB-0620652). The authors particularly thank Phil Grime and the UCPE–University of Sheffield, and Mark Westoby and the ARC–NZ Network of Vegetation Function–Macquarie University, for inspiring discussion and development of trait measurement. The authors also thank Marcelo Cabido, William Bond, Paula Tecco, Fernando Casanoves and Bryan Finegan for valuable discussions. The authors thank Diana Abal Solis and Juan Pablo Bellini for assistance with figures and diagrams and Valeria Falczuk for technical assistance in the laboratory. The authors appreciate helpful comments of Adrienne Nicotra, Hans Lambers and one anonymous reviewer, which greatly improved the quality of the manuscript. Many users of the previous handbook gave useful feedback on trait measurements. Finally, we thank plant species around the world for beginning to give away the secrets of their functioning and ecosystem-level impacts.
References
Ackerly DD (2004) Functional strategies of chaparral shrubs in relation to seasonal water deficit. Ecological Monographs 74, 25–44.| Functional strategies of chaparral shrubs in relation to seasonal water deficit.Crossref | GoogleScholarGoogle Scholar |
Ackerly DD, Knight CA, Weiss SB, Barton K, Starmer KP (2002) Leaf size, specific leaf area and microhabitat distribution of chaparral woody plants: contrasting patterns in species level and community level analyses. Oecologia 130, 449–457.
| Leaf size, specific leaf area and microhabitat distribution of chaparral woody plants: contrasting patterns in species level and community level analyses.Crossref | GoogleScholarGoogle Scholar |
Adair EC, Parton WJ, Del Grosso SJ, Silver WL, Harmon ME, Hall SA, Burke IC, Hart SC (2008) Simple three-pool model accurately describes patterns of long-term litter decomposition in diverse climates. Global Change Biology 14, 2636–2660.
Addington RN, Donovan LA, Mitchell RJ, Vose JM, Pecot SD, Jack SB, Hacke UG, Sperry JS, Oren R (2006) Adjustments in hydraulic architecture of Pinus palustris maintain similar stomatal conductance in xeric and mesic habitats. Plant, Cell & Environment 29, 535–545.
| Adjustments in hydraulic architecture of Pinus palustris maintain similar stomatal conductance in xeric and mesic habitats.Crossref | GoogleScholarGoogle Scholar |
Adiku SGK, Rose CW, Braddock RD, Ozier-Lafontaine H (2000) On the simulation of root water extraction: examination of a minimum energy hypothesis. Soil Science 165, 226–236.
| On the simulation of root water extraction: examination of a minimum energy hypothesis.Crossref | GoogleScholarGoogle Scholar |
Aerts R (1995) The advantages of being evergreen. Trends in Ecology & Evolution 10, 402–407.
| The advantages of being evergreen.Crossref | GoogleScholarGoogle Scholar |
Aerts R (1996) Nutrient resorption from senescing leaves of perennials are there general patterns? Journal of Ecology 84, 597–608.
| Nutrient resorption from senescing leaves of perennials are there general patterns?Crossref | GoogleScholarGoogle Scholar |
Aerts R, Chapin S (1999) The mineral nitrition of wild plants revisited: a re-evaluation of processes and patterns. Advances in Ecological Research 30, 1–67.
| The mineral nitrition of wild plants revisited: a re-evaluation of processes and patterns.Crossref | GoogleScholarGoogle Scholar |
Aerts R, Boot RGA, Van der Aart PJM (1991) The relation between above- and belowground biomass allocation patterns and competitive ability. Oecologia 87, 551–559.
| The relation between above- and belowground biomass allocation patterns and competitive ability.Crossref | GoogleScholarGoogle Scholar |
Agrawal AA, Fishbein M (2006) Plant defense syndromes. Ecology 87, S132–S149.
| Plant defense syndromes.Crossref | GoogleScholarGoogle Scholar | 16922309PubMed |
Albert CH, Thuiller W, Yoccoz NG, Douzet R, Aubert S, Lavorel S (2010) A multi-trait approach reveals the structure and the relative importance of intra- vs. interspecific variability in plant traits. Functional Ecology 24, 1192–1201.
| A multi-trait approach reveals the structure and the relative importance of intra- vs. interspecific variability in plant traits.Crossref | GoogleScholarGoogle Scholar |
Albert CH, De Bello F, Boulangeat I, Pellet G, Lavorel S, Thuiller W (2012) On the importance of intraspecific variability for the quantification of functional diversity. Oikos 121, 116–126.
| On the importance of intraspecific variability for the quantification of functional diversity.Crossref | GoogleScholarGoogle Scholar |
Alder NN, Sperry JS, Pockman WT (1996) Root and stem xylem embolism, stomatal conductance, and leaf turgor in Acer grandidentatum populations along a soil moisture gradient Oecologia 105, 293–301.
| Root and stem xylem embolism, stomatal conductance, and leaf turgor in Acer grandidentatum populations along a soil moisture gradientCrossref | GoogleScholarGoogle Scholar |
Alder NN, Pockman WT, Nuismer S (1997) Use of centrifugal force in the study of xylem cavitation. Journal of Experimental Botany 48, 665–674.
| Use of centrifugal force in the study of xylem cavitation.Crossref | GoogleScholarGoogle Scholar |
Allen SE (1989) ‘Chemical analysis of ecological material.’ 2nd edn. (Blackwell: Oxford, UK)
Anderson JM, Ingram JSI (1993) ‘Tropical soil biology and fertility: a handbook of methods.’ 2nd edn. (CAB International: Wallingford, UK)
Ansquer P, Duru M, Theau J-P, Cruz P (2009) Convergence in plant traits between species within grassland communities simplifies their monitoring. Ecological Indicators 9, 1020–1029.
| Convergence in plant traits between species within grassland communities simplifies their monitoring.Crossref | GoogleScholarGoogle Scholar |
Anten NPR, Schieving F (2010) The role of wood mass density and mechanical constraints in the economy of tree architecture. American Naturalist 175, 250–260.
| The role of wood mass density and mechanical constraints in the economy of tree architecture.Crossref | GoogleScholarGoogle Scholar |
Aranwela N, Sanson G, Read J (1999) Methods of assessing leaf-fracture properties. New Phytologist 144, 369–383.
| Methods of assessing leaf-fracture properties.Crossref | GoogleScholarGoogle Scholar |
Archibald S, Bond WJ (2003) Growing tall vs growing wide: tree architecture and allometry of Acacia karroo in forest, savanna, and arid environments. Oikos 102, 3–14.
| Growing tall vs growing wide: tree architecture and allometry of Acacia karroo in forest, savanna, and arid environments.Crossref | GoogleScholarGoogle Scholar |
Arneth A, Harrison SP, Zaehle S, Tsigaridis K, Menon S, Bartlein PJ, Feichter J, Korhola A, Kulmala M, O’Donnell D, Schurgers G, Sorvari S, Vesala T (2010) Terrestrial biogeochemical feedbacks in the climate system. Nature Geoscience 3, 525–532.
| Terrestrial biogeochemical feedbacks in the climate system.Crossref | GoogleScholarGoogle Scholar |
Ashraf M, Harris PJC (2004) Potential biochemical indicators of salinity tolerance in plants. Plant Science 166, 3–16.
| Potential biochemical indicators of salinity tolerance in plants.Crossref | GoogleScholarGoogle Scholar |
Aston AR (1979) Rainfall interception by eight small trees. Journal of Hydrology 42, 383–396.
| Rainfall interception by eight small trees.Crossref | GoogleScholarGoogle Scholar |
Atkin OK, Bruhn D, Hurry VM, Tjoelker MG (2005) The hot and the cold: unravelling the variable response of plant respiration to temperature. Functional Plant Biology 32, 87–105.
| The hot and the cold: unravelling the variable response of plant respiration to temperature.Crossref | GoogleScholarGoogle Scholar |
Austin AT, Vivanco L (2006) Plant litter decomposition in a semi-arid ecosystem controlled by photodegradation. Nature 442, 555–558.
| Plant litter decomposition in a semi-arid ecosystem controlled by photodegradation.Crossref | GoogleScholarGoogle Scholar | 16885982PubMed |
Bakker JP, Poschlod P, Strykstra RJ, Bekker RM, Thompson K (1996) Seed banks and seed dispersal: important topics in restoration ecology. Acta Botanica Neerlandica 45, 461–490.
Baraloto C, Paine CET, Poorter L, Beauchene J, Bonal D, Domenach A-M, Hérault B, Patiño S, Roggy J-C, Chave J (2010a) Decoupled leaf and stem economics in rain forest trees. Ecology Letters 13, 1338–1347.
| Decoupled leaf and stem economics in rain forest trees.Crossref | GoogleScholarGoogle Scholar | 20807232PubMed |
Baraloto C, Paine CET, Patiño S, Bonal D, Hérault B, Chave J (2010b) Functional trait variation and sampling strategies in species-rich plant communities. Functional Ecology 24, 208–216.
| Functional trait variation and sampling strategies in species-rich plant communities.Crossref | GoogleScholarGoogle Scholar |
Barkman J (1988) New systems of plant growth forms and phenological plant types. In ‘Plant form and vegetation structure’. (Eds MJA Werger, PJM Van der Aart, HJ During, JTA Verhoeven) pp. 9–44 (SPB Academic Publishers: The Hague, The Netherlands)
Bartlett M, Scoffoni C, Sack L (2012) The determinants of leaf turgor loss point and prediction of drought tolerance of species and biomes: a global meta-analysis. Ecology Letters 15, 393–405.
| The determinants of leaf turgor loss point and prediction of drought tolerance of species and biomes: a global meta-analysis.Crossref | GoogleScholarGoogle Scholar | 22435987PubMed |
Belea A, Kiss AS, Galbacs Z (1998) New methods for determination of C-3, C-4 and CAM-type plants. Cereal Research Communications 26, 413–418.
Bellingham PJ, Sparrow AD (2000) Resprouting as a life history strategy in woody plant communities. Oikos 89, 409–416.
| Resprouting as a life history strategy in woody plant communities.Crossref | GoogleScholarGoogle Scholar |
Berg B, Laskowski R (2005) Litter decomposition: a guide to carbon and nutrient turnover. Advances in Ecological Research 38, 1–17.
| Litter decomposition: a guide to carbon and nutrient turnover.Crossref | GoogleScholarGoogle Scholar |
Bhaskar R, Ackerly DD (2006) Ecological relevance of minimum seasonal water potentials. Physiologia Plantarum 127, 353–359.
| Ecological relevance of minimum seasonal water potentials.Crossref | GoogleScholarGoogle Scholar |
Blonder B, Buzzard V, Simova I, Sloat L, Boyle B, Lipson R, Aguilar-Beaucage B, Andrade A, Barber B, Barnes C, Bushey D, Cartagena P, Chaney M, Contreras K, Cox M, Cueto M, Curtis C, Fisher M, Furst L, Gellagos J, Hall R, Hauschild A, Jerez A, Jones N, Klucas A, Kono A, Lamb M, Ruiz Matthai JD, McIntyre C, McKenna J, Mosier N, Navabi M, Ochoa A, Pace L, Plassmann R, Richter R, Russakoff B, Aubyn HS, Stagg R, Sterner M, Stewart E, Thompson TT, Thornton J, Trujillo PJ, Volpe TJ, Enquist BJ (2012) The leaf area shrinkage effect can bias paleoclimate and ecology research. American Journal of Botany 99, 1756–1763.
| The leaf area shrinkage effect can bias paleoclimate and ecology research.Crossref | GoogleScholarGoogle Scholar | 23132615PubMed |
Blum A (1988) ‘Plant breeding for stress environments.’ (CRC Press: Boca Raton, FL)
Böhm W (1979) ‘Methods of studying root systems. Ecological studies 33.’ (Springer: Berlin)
Bond WJ, Midgley JJ (1995) Kill thy neighbour: an individualistic argument for the evolution of flammability. Oikos 73, 79–85.
| Kill thy neighbour: an individualistic argument for the evolution of flammability.Crossref | GoogleScholarGoogle Scholar |
Bond WJ, Midgley JJ (2001) Ecology of sprouting in woody plants: the persistence niche. Trends in Ecology & Evolution 16, 45–51.
| Ecology of sprouting in woody plants: the persistence niche.Crossref | GoogleScholarGoogle Scholar |
Bond WJ, Van Wilgen BW (1996) ‘Fire and plants.’ (Chapman and Hall: London)
Bouma TJ, Nielsen KL, Koutstaal B (2000) Sample preparation and scanning protocol for computerised analysis of root length and diameter. Plant and Soil 218, 185–196.
| Sample preparation and scanning protocol for computerised analysis of root length and diameter.Crossref | GoogleScholarGoogle Scholar |
Box EO (1996) Plant functional types and climate at the global scale. Journal of Vegetation Science 7, 309–320.
| Plant functional types and climate at the global scale.Crossref | GoogleScholarGoogle Scholar |
Boyce CK, Brodribb TJ, Feild TS, Zwieniecki MA (2009) Angiosperm leaf vein evolution was physiologically and environmentally transformative. Proceedings. Biological Sciences 276, 1771–1776.
| Angiosperm leaf vein evolution was physiologically and environmentally transformative.Crossref | GoogleScholarGoogle Scholar |
Brando PM, Nepstad DC, Balch JK, Bolker B, Christman MC, Coe M, Putz FE (2012) Fire-induced tree mortality in a neotropical forest: the roles of bark traits, tree size, wood density and fire behavior. Global Change Biology 18, 630–641.
| Fire-induced tree mortality in a neotropical forest: the roles of bark traits, tree size, wood density and fire behavior.Crossref | GoogleScholarGoogle Scholar |
Breckle SW (2002) Salinity, halophytes and salt affected natural ecosystems. In ‘Salinity: environment–plants–molecules’. (Eds A Läuchli, U Lüttge) pp. 53–77 (Kluwer Academic Publishers: London)
Brewer CA, Núñez CI (2007) Patterns of leaf wettability along extreme moisture gradient in western Patagonia, Argentina. International Journal of Plant Sciences 168, 555–562.
| Patterns of leaf wettability along extreme moisture gradient in western Patagonia, Argentina.Crossref | GoogleScholarGoogle Scholar |
Brewer CA, Smith WK, Vogelmann TC (1991) Functional interaction between leaf trichomes, leaf wettability and the optical properties of water droplets. Plant, Cell & Environment 14, 955–962.
| Functional interaction between leaf trichomes, leaf wettability and the optical properties of water droplets.Crossref | GoogleScholarGoogle Scholar |
Brodribb TJ, Feild TS (2010) Leaf hydraulic evolution led a surge in leaf photosynthetic capacity during early angiosperm diversification. Ecology Letters 13, 175–183.
| Leaf hydraulic evolution led a surge in leaf photosynthetic capacity during early angiosperm diversification.Crossref | GoogleScholarGoogle Scholar | 19968696PubMed |
Brodribb TJ, Hill RS (2000) Increases in water potential gradient reduce xylem conductivity in whole plants. Evidence from a low-pressure conductivity method. Plant Physiology 123, 1021–1028.
| Increases in water potential gradient reduce xylem conductivity in whole plants. Evidence from a low-pressure conductivity method.Crossref | GoogleScholarGoogle Scholar | 10889251PubMed |
Brodribb TJ, Holbrook NM, Edwards EJ, Gutiérrez MV (2003) Relations between stomatal closure, leaf turgor and xylem vulnerability in eight tropical dry forest trees. Plant, Cell & Environment 26, 443–450.
| Relations between stomatal closure, leaf turgor and xylem vulnerability in eight tropical dry forest trees.Crossref | GoogleScholarGoogle Scholar |
Brodribb TJ, Feild TS, Jordan GJ (2007) Leaf maximum photosynthetic rate and venation are linked by hydraulics. Plant Physiology 144, 1890–1898.
| Leaf maximum photosynthetic rate and venation are linked by hydraulics.Crossref | GoogleScholarGoogle Scholar | 17556506PubMed |
Brodribb TJ, Feild TS, Sack L (2010) Viewing leaf structure and evolution from a hydraulic perspective. Functional Plant Biology 37, 488–498.
| Viewing leaf structure and evolution from a hydraulic perspective.Crossref | GoogleScholarGoogle Scholar |
Brown S (1997) ‘Estimating biomass and biomass change of tropical forests. A primer.’ FAO Forestry Paper 134 (FAO: Rome)
Brussaard L, Caron P, Campbell B, Lipper L, Mainka S, Rabbinge R, Babin D, Pulleman M (2010) Reconciling biodiversity conservation and food security: scientific challenges for a new agriculture. Current Opinion in Environmental Sustainability 2, 34–42.
| Reconciling biodiversity conservation and food security: scientific challenges for a new agriculture.Crossref | GoogleScholarGoogle Scholar |
Bruun HH, Poschlod P (2006) Why are small seeds dispersed through animal guts: large numbers or seed size per se? Oikos 113, 402–411.
| Why are small seeds dispersed through animal guts: large numbers or seed size per se?Crossref | GoogleScholarGoogle Scholar |
Bucci SJ, Goldstein G, Meinzer FC, Scholz FG, Franco AC, Bustamante M (2004) Functional convergence in hydraulic architecture and water relations of tropical savanna trees: from leaf to whole plant. Tree Physiology 24, 891–899.
| Functional convergence in hydraulic architecture and water relations of tropical savanna trees: from leaf to whole plant.Crossref | GoogleScholarGoogle Scholar | 15172839PubMed |
Buckley TN, Roberts DW (2006) How should leaf area, sapwood area and stomatal conductance vary with tree height to maximize growth? Tree Physiology 26, 145–157.
| How should leaf area, sapwood area and stomatal conductance vary with tree height to maximize growth?Crossref | GoogleScholarGoogle Scholar | 16356911PubMed |
Burd M (2007) Adaptive function of drip tips: a test of the epiphyll hypothesis in Psychotria marginata and Faramea occidentalis (Rubiaceae). Journal of Tropical Ecology 23, 449–455.
| Adaptive function of drip tips: a test of the epiphyll hypothesis in Psychotria marginata and Faramea occidentalis (Rubiaceae).Crossref | GoogleScholarGoogle Scholar |
Burrows GE (2002) Epicormic strand structure in Angophora, Eucalyptus and Lophostemon (Myrtaceae) – implications for fire resistance and recovery. New Phytologist 153, 111–131.
| Epicormic strand structure in Angophora, Eucalyptus and Lophostemon (Myrtaceae) – implications for fire resistance and recovery.Crossref | GoogleScholarGoogle Scholar |
Cadisch G, Giller KE (1997) ‘Driven by Nature. Plant litter quality and decomposition.’ (CAB International: London)
Cain SA (1950) Life-forms and phytoclimate. Botanical Review 16, 1–32.
| Life-forms and phytoclimate.Crossref | GoogleScholarGoogle Scholar |
Caldwell MM, Virginia RA (1989) Root systems. In ‘Plant physiogical ecology: field methods and instrumentation’. (Ed. RW Pearcy) pp. 367–398 (Chapman and Hall: London)
Cardinale BJ, Duffy JE, Gonzalez A, Hooper DU, Perrings C, Venail P, Narwani A, Mace GM, Tilman D, Wardle DA, Kinzig AP, Daily GC, Loreau M, Grace JB, Larigauderie A, Srivastava DS, Naeem S (2012) Biodiversity loss and its impact on humanity. Nature 486, 59–67.
| Biodiversity loss and its impact on humanity.Crossref | GoogleScholarGoogle Scholar | 22678280PubMed |
Casanoves F, Pla L, Di Rienzo JA, Díaz S (2011) FDiversity: a software package for the integrated analysis of functional diversity. Methods in Ecology and Evolution 2, 233–237.
| FDiversity: a software package for the integrated analysis of functional diversity.Crossref | GoogleScholarGoogle Scholar |
Castro-Díez P, Puyravaud JP, Cornelissen JHC (2000) Leaf structure and anatomy as related to leaf mass per area variation in seedlings of a wide range of woody plant species and types. Oecologia 124, 476–486.
| Leaf structure and anatomy as related to leaf mass per area variation in seedlings of a wide range of woody plant species and types.Crossref | GoogleScholarGoogle Scholar |
Causton DR, Venus JC (1981) ‘The biometry of plant growth.’ (Edward Arnold: London)
Cavender-Bares J, Kitajima K, Bazzaz FA (2004) Multiple trait associations in relation to habitat differentiation among 17 floridian oak species. Ecological Monographs 74, 635–662.
| Multiple trait associations in relation to habitat differentiation among 17 floridian oak species.Crossref | GoogleScholarGoogle Scholar |
Cavender-Bares J, Kozak KH, Fine PVA, Kembel SW (2009) The merging of community ecology and phylogenetic biology. Ecology Letters 12, 693–715.
| The merging of community ecology and phylogenetic biology.Crossref | GoogleScholarGoogle Scholar | 19473217PubMed |
Cernusak LA, Tcherkez G, Keitel C, Cornwell WK, Santiago LS, Knohl A, Barbour MM, Williams DG, Reich PB, Ellsworth DS, Dawson TE, Griffiths HG, Farquhar GD, Wright IJ (2009) Why are non-photosynthetic tissues generally 13C enriched compared with leaves in C3 plants? Review and synthesis of current hypotheses. Functional Plant Biology 36, 199–213.
| Why are non-photosynthetic tissues generally 13C enriched compared with leaves in C3 plants? Review and synthesis of current hypotheses.Crossref | GoogleScholarGoogle Scholar |
Chabot BF, Hicks DJ (1982) The ecology of leaf life span. Annual Review of Ecology and Systematics 13, 229–259.
| The ecology of leaf life span.Crossref | GoogleScholarGoogle Scholar |
Chapin FS (1980) The mineral nutrition of wild plants. Annual Review of Ecology and Systematics 11, 233–260.
| The mineral nutrition of wild plants.Crossref | GoogleScholarGoogle Scholar |
Chapin FS, Autumn K, Pugnaire F (1993) Evolution of suites of traits in response to environmental stress. American Naturalist 142, S78–S92.
| Evolution of suites of traits in response to environmental stress.Crossref | GoogleScholarGoogle Scholar |
Chapin FS, Zaveleta ES, Eviner VT, Naylor RL, Vitousek PM, Lavorel S, Reynolds HL, Hooper DU, Sala OE, Hobbie SE, Mack MC, Diaz S (2000) Consequences of changing biotic diversity. Nature 405, 234–242.
| Consequences of changing biotic diversity.Crossref | GoogleScholarGoogle Scholar | 10821284PubMed |
Chapin FS, Trainor SF, Huntington O, Lovecraft AL, Zavaleta E, Natcher DC, McGuire D, Nelson JL, Ray L, Calef M, Fresco N, Huntington H, Rupp TS, DeWilde L, Naylor RL (2008) Increasing wildfire in Alaska’s boreal forest: pathways to potential solutions of a wicked problem. Bioscience 58, 531–540.
| Increasing wildfire in Alaska’s boreal forest: pathways to potential solutions of a wicked problem.Crossref | GoogleScholarGoogle Scholar |
Chave J, Andalo C, Brown S, Cairns MA, Chambers JQ, Eamus D, Fölster H, Fromard F, Higuchi N, Kira T, Lescure J-P, Nelson BW, Ogawa H, Puig H, Riéra B, Yamakura T (2005) Tree allometry and improved estimation of carbon stocks and balance in tropical forests. Oecologia 145, 87–99.
| Tree allometry and improved estimation of carbon stocks and balance in tropical forests.Crossref | GoogleScholarGoogle Scholar | 15971085PubMed |
Chave J, Coomes DA, Jansen S, Lewis SL, Swenson NG, Zanne AE (2009) Towards a worldwide wood economics spectrum. Ecology Letters 12, 351–366.
| Towards a worldwide wood economics spectrum.Crossref | GoogleScholarGoogle Scholar | 19243406PubMed |
Chen N, Black TA (1992) Defining leaf area index for non-flat leaves. Plant, Cell & Environment 15, 421–429.
| Defining leaf area index for non-flat leaves.Crossref | GoogleScholarGoogle Scholar |
Cherubini P, Gartner BL, Tognetti R, Bräker OU, Schoch W, Innes JL (2003) Identification, measurement and interpretation of tree rings in woody species from mediterranean climates. Biological Reviews of the Cambridge Philosophical Society 78, 119–148.
| Identification, measurement and interpretation of tree rings in woody species from mediterranean climates.Crossref | GoogleScholarGoogle Scholar | 12620063PubMed |
Chiba Y (1991) Plant form based on the pipe model theory II. Quantitative analysis of ramification in morphology. Ecological Research 6, 21–28.
| Plant form based on the pipe model theory II. Quantitative analysis of ramification in morphology.Crossref | GoogleScholarGoogle Scholar |
Choat B, Sack L, Holbrook NM (2007) Diversity of hydraulic traits in nine Cordia species growing in tropical forests with contrasting precipitation. New Phytologist 175, 686–698.
| Diversity of hydraulic traits in nine Cordia species growing in tropical forests with contrasting precipitation.Crossref | GoogleScholarGoogle Scholar | 17688584PubMed |
Choong MF, Lucas PW, Ong JSY, Pereira B, Tan HTW, Turner IM (1992) Leaf fracture toughness and sclerophylly: their correlations and ecological implications. New Phytologist 121, 597–610.
| Leaf fracture toughness and sclerophylly: their correlations and ecological implications.Crossref | GoogleScholarGoogle Scholar |
Clements ES (1905) The relation of leaf structure to physical factors. Transactions of the American Microscopical Society 26, 19–102.
| The relation of leaf structure to physical factors.Crossref | GoogleScholarGoogle Scholar |
Cochard H, Cruizat P, Tyree MT (1992) Use of positive pressures to establish vulnerability curves. Plant Physiology 100, 205–209.
| Use of positive pressures to establish vulnerability curves.Crossref | GoogleScholarGoogle Scholar | 16652947PubMed |
Coley PD (1987) Patrones en las defensas de las plantas: ¿por qué los herbívoros prefieren ciertas especies? Revista de Biologia Tropical 35, 151–164.
Coley PD (1988) Effects of plant growth rate and leaf lifetime on the amount and type of anti-herbivore defense. Oecologia 74, 531–536.
| Effects of plant growth rate and leaf lifetime on the amount and type of anti-herbivore defense.Crossref | GoogleScholarGoogle Scholar |
Cooper SM, Ginnett TF (1998) Spines protect plants against browsing by small climbing mammals. Oecologia 113, 219–221.
| Spines protect plants against browsing by small climbing mammals.Crossref | GoogleScholarGoogle Scholar |
Cooper DJ, D’Amico DR, Scott ML (2003) Physiological and morphological response patterns of Populus deltoides to alluvial groundwater pumping. Environmental Management 31, 215–226.
| Physiological and morphological response patterns of Populus deltoides to alluvial groundwater pumping.Crossref | GoogleScholarGoogle Scholar | 12520377PubMed |
Cornelissen JHC (1996) An experimental comparison of leaf decomposition rates in a wide range of temperate plant species and types. Journal of Ecology 84, 573–582.
| An experimental comparison of leaf decomposition rates in a wide range of temperate plant species and types.Crossref | GoogleScholarGoogle Scholar |
Cornelissen JHC (1999) A triangular relationship between leaf size and seed size among woody species: allometry, ontogeny, ecology and taxonomy. Oecologia 118, 248–255.
| A triangular relationship between leaf size and seed size among woody species: allometry, ontogeny, ecology and taxonomy.Crossref | GoogleScholarGoogle Scholar |
Cornelissen JHC, Castro-Díez P, Hunt R (1996) Seedling growth, allocation and leaf attributes in a wide range of woody plant species and types. Journal of Ecology 84, 755–765.
| Seedling growth, allocation and leaf attributes in a wide range of woody plant species and types.Crossref | GoogleScholarGoogle Scholar |
Cornelissen JHC, Werger MJA, Castro-Díez P, Van Rheenen JWA, Rowland AP (1997) Foliar nutrients in relation to growth, allocation and leaf traits in seedlings of a wide range of woody plant species and types. Oecologia 111, 460–469.
| Foliar nutrients in relation to growth, allocation and leaf traits in seedlings of a wide range of woody plant species and types.Crossref | GoogleScholarGoogle Scholar |
Cornelissen JHC, Pérez-Harguindeguy N, Díaz S, Grime JP, Marzano B, Cabido M, Vendramini F, Cerabolini B (1999) Leaf structure and defence control litter decomposition rate across species and life forms in regional floras on two continents. New Phytologist 143, 191–200.
| Leaf structure and defence control litter decomposition rate across species and life forms in regional floras on two continents.Crossref | GoogleScholarGoogle Scholar |
Cornelissen JHC, Lavorel S, Garnier E, Díaz S, Buchmann N, Gurvich DE, Reich PB, ter Steege H, Morgan HD, van der Heijden MGA, Pausas JG, Poorter H (2003) A handbook of protocols for standarised and easy measurement of plant functional traits worldwide. Australian Journal of Botany 51, 335–380.
| A handbook of protocols for standarised and easy measurement of plant functional traits worldwide.Crossref | GoogleScholarGoogle Scholar |
Cornelissen JHC, Quested H, Van Logtestijn RSP, Pérez-Harguindeguy N, Gwynn-Jones D, Díaz S, Callaghan TV, Press MC, Aerts R (2006) Foliar pH as a new plant trait: can it explain variation in foliar chemistry and carbon cycling processes among Subarctic plant species and types? Oecologia 147, 315–326.
| Foliar pH as a new plant trait: can it explain variation in foliar chemistry and carbon cycling processes among Subarctic plant species and types?Crossref | GoogleScholarGoogle Scholar |
Cornelissen JHC, Van Bodegom PM, Aerts R, Callaghan TV, Van Logtestijn RSP, Alatalo J, Stuart Chapin F, Gerdol R, Gudmundsson J, Gwynn-Jones D, Hartley AE, Hik DS, Hofgaard A, Jónsdóttir IS, Karlsson S, Klein JA, Laundre J, Magnusson B, Michelsen A, Molau U, Onipchenko VG, Quested HM, Sandvik SM, Schmidt IK, Shaver GR, Solheim B, Soudzilovskaia NA, Stenström A, Tolvanen A, Totland Ø, Wada N, Welker JM, Zhao X, MOL Team (2007) Global negative vegetation feedback to climate warming responses of leaf litter decomposition rates in cold biomes. Ecology Letters 10, 619–627.
| Global negative vegetation feedback to climate warming responses of leaf litter decomposition rates in cold biomes.Crossref | GoogleScholarGoogle Scholar |
Cornelissen JHC, Sibma F, Van Logtestijn RSP, Broekman R, Thompson K (2011) Leaf pH as a plant trait: species-driven rather than soil-driven variation. Functional Ecology 25, 449–455.
| Leaf pH as a plant trait: species-driven rather than soil-driven variation.Crossref | GoogleScholarGoogle Scholar |
Cornwell WK, Bhaskar R, Sack L, Cordell S, Lunch CK (2007) Adjustment of structure and function of Hawaiian Metrosideros polymorpha at high vs. low precipitation. Functional Ecology 21, 1063–1071.
| Adjustment of structure and function of Hawaiian Metrosideros polymorpha at high vs. low precipitation.Crossref | GoogleScholarGoogle Scholar |
Cornwell WK, Cornelissen JHC, Amatangelo K, Dorrepaal E, Eviner VT, Godoy O, Hobbie SE, Hoorens B, Kurokawa H, Pérez-Harguindeguy N, Quested HM, Santiago LS, Wardle DA, Wright IJ, Aerts R, Allison SD, Van Bodegom P, Brovkin V, Chatain A, Callaghan TV, Díaz S, Garnier E, Gurvich DE, Kazakou E, Klein JA, Read J, Reich PB, Soudzilovskaia NA, Vaieretti MV, Westoby M (2008) Plant species traits are the predominant control on litter decomposition rates within biomes worldwide. Ecology Letters 11, 1065–1071.
| Plant species traits are the predominant control on litter decomposition rates within biomes worldwide.Crossref | GoogleScholarGoogle Scholar | 18627410PubMed |
Cornwell WK, Cornelissen JHC, Allison SD, Bauhus J, Eggleton P, Preston CM, Scarff F, Weedon JT, Wirth C, Zanne AE (2009) Plant traits and wood fates across the globe: rotted, burned, or consumed? Global Change Biology 15, 2431–2449.
| Plant traits and wood fates across the globe: rotted, burned, or consumed?Crossref | GoogleScholarGoogle Scholar |
Cousens RD, Hill J, French K, Bishop ID (2010) Towards better prediction of seed dispersal by animals: conceptual frameworks and process-based models. Functional Ecology 24, 1163–1170.
| Towards better prediction of seed dispersal by animals: conceptual frameworks and process-based models.Crossref | GoogleScholarGoogle Scholar |
Craine JM (2009) ‘Resource strategies of wild plants.’ (Princeton University Press: Princeton, NJ)
Craine JM, Berin DM, Reich PB, Tilman D, Knops J (1999) Measurement of leaf longevity of 14 species of grasses and forbs using a novel approach. New Phytologist 142, 475–481.
| Measurement of leaf longevity of 14 species of grasses and forbs using a novel approach.Crossref | GoogleScholarGoogle Scholar |
Craine JM, Tilman DG, Wedin DA, Chapin S (2001) The relationships among root and leaf traits of 76 grassland species and relative abundance along fertility and disturbance gradients. Oikos 93, 274–285.
| The relationships among root and leaf traits of 76 grassland species and relative abundance along fertility and disturbance gradients.Crossref | GoogleScholarGoogle Scholar |
Cramer W (1997) Using plant functional types in a global vegetation model. In ‘Plant functional types’. (Eds TM Smith, HH. Shugart, FI Woodward) pp. 271–288 (Cambridge University Press: Cambridge, UK)
Cramer W, Bondeau A, Woodward FI, Prentice IC, Betts RA, Brovkin V, Cox PM, Fisher V, Foley JA, Friend AD, Kucharik C, Lomas MR, Ramankutty N, Sitch S, Smith B, White A, Young-Molling C (2001) Global response of terrestrial ecosystem structure and function to CO2 and climate change: results from six dynamic global vegetation models. Global Change Biology 7, 357–373.
| Global response of terrestrial ecosystem structure and function to CO2 and climate change: results from six dynamic global vegetation models.Crossref | GoogleScholarGoogle Scholar |
Darvell BW, Lee PKD, Yuen TDB, Lucas PW (1996) A portable fracture toughness tester for biological materials. Measurement Science & Technology 7, 954–962.
| A portable fracture toughness tester for biological materials.Crossref | GoogleScholarGoogle Scholar |
David JS, Valente F, Gash JHC (2005) Evaporation of intercepted rainfall. In ‘Encyclopedia of hydrological sciences’. (Ed. MG Anderson) pp. 627–634 (John Wiley and Sons: Chichester, UK)
Davis SD, Ewers FW, Sperry JS, Portwood KA, Crocker MC, Adams GC (2002) Shoot dieback during prolonged drought in Ceanothus (Rhamnaceae) chaparal of California: a possible case of hydraulic failure. American Journal of Botany 89, 820–828.
| Shoot dieback during prolonged drought in Ceanothus (Rhamnaceae) chaparal of California: a possible case of hydraulic failure.Crossref | GoogleScholarGoogle Scholar | 21665682PubMed |
De Bello F, Lavorel S, Gerhold P, Reier Ü, Pärtel M (2010) A biodiversity monitoring framework for practical conservation of grasslands and shrublands. Biological Conservation 143, 9–17.
| A biodiversity monitoring framework for practical conservation of grasslands and shrublands.Crossref | GoogleScholarGoogle Scholar |
De Bello F, Lavorel S, Albert CH, Thuiller W, Grigulis K, Dolezal J, Janecek S, Lêps J (2011) Quantifying the relevance of intraspecific trait variability for functional diversity. Methods in Ecology and Evolution 2, 163–174.
| Quantifying the relevance of intraspecific trait variability for functional diversity.Crossref | GoogleScholarGoogle Scholar |
De Kroon H, Van Groenendael JM (1997) ‘The ecology and evolution of clonal plants.’ (Backhuys Publishers: Leiden, The Netherlands)
De Vogel EF (1980) ‘Seedlings of dicotyledons.’ (Centre for Agricultural Publishing and Documentation: Wageningen, The Netherlands)
De Witte LC, Stöcklin J (2010) Longevity of clonal plants: why it matters and how to measure it. Annals of Botany 106, 859–870.
| Longevity of clonal plants: why it matters and how to measure it.Crossref | GoogleScholarGoogle Scholar | 20880935PubMed |
Del Tredici P (2001) Sprouting in temperate trees: a morphological and ecological review Botanical Review 67, 121–140.
| Sprouting in temperate trees: a morphological and ecological reviewCrossref | GoogleScholarGoogle Scholar |
Díaz S, Hodgson JG, Thompson K, Cabido M, Cornelissen JHC, Jalili A, Montserrat-Martí G, Grime JP, Zarrinkamar F, Asri Y, Band SR, Basconcelo S, Castro-Díez P, Funes G, Hamzehee B, Khoshnevi M, Pérez-Harguindeguy N, Pérez-Rontomé MC, Shirvany FA, Vendramini F, Yazdani S, Abbas-Azimi R, Bogaard A, Boustani S, Charles M, Dehghan M, de Torres-Espuny L, Falczuk V, Guerrero-Campo J, Hynd A, Jones G, Kowsary E, Kazemi-Saeed F, Maestro-Martínez M, Romo-Díez A, Shaw S, Siavash B, Villar-Salvador P, Zak MR (2004) The plant traits that drive ecosystems: evidence from three continents. Journal of Vegetation Science 15, 295–304.
Díaz S, Lavorel S, De Bello F, Quétier F, Grigulis K, Robson MT (2007) Incorporating plant functional diversity effects in ecosystem service assessments. Proceedings of the National Academy of Sciences, USA 104, 20684–20689.
| Incorporating plant functional diversity effects in ecosystem service assessments.Crossref | GoogleScholarGoogle Scholar |
Diefendorf AF, Mueller KE, Wing SL, Koch PL, Freeman KH (2010) Global patterns in leaf 13C discrimination and implications for studies of past and future climate. Proceedings of the National Academy of Sciences, USA 107, 5738–5743.
| Global patterns in leaf 13C discrimination and implications for studies of past and future climate.Crossref | GoogleScholarGoogle Scholar |
Diemer M (1998) Life span and dynamics of leaves of herbaceous perennials in high-elevation environments: ‘news from the elephant’s leg’. Functional Ecology 12, 413–425.
| Life span and dynamics of leaves of herbaceous perennials in high-elevation environments: ‘news from the elephant’s leg’.Crossref | GoogleScholarGoogle Scholar |
Dilcher DL (1974) Approaches to the identification of angiosperm leaf remains. Botanical Review 40, 1–157.
| Approaches to the identification of angiosperm leaf remains.Crossref | GoogleScholarGoogle Scholar |
Dimitrakopoulos AP, Panov PI (2001) Pyric properties of some dominant Mediterranean vegetation species. International Journal of Wildland Fire 10, 23–27.
| Pyric properties of some dominant Mediterranean vegetation species.Crossref | GoogleScholarGoogle Scholar |
Doherty RM, Sitch S, Smith B, Lewis SL, Thornton PK Doherty RM, Sitch S, Smith B, Lewis SL, Thornton PK (2010) Implications of future climate and atmospheric CO2 content for regional biogeochemistry, biogeography and ecosystem services across East Africa. Global Change Biology 16, 617–640.
| Implications of future climate and atmospheric CO2 content for regional biogeochemistry, biogeography and ecosystem services across East Africa.Crossref | GoogleScholarGoogle Scholar |
Domingo F, Sánchez G, Moro MJ, Brenner AJ, Puigdefábregas J (1998) Measurement and modelling of rainfall interception by three semi-arid canopies. Agricultural and Forest Meteorology 91, 275–292.
| Measurement and modelling of rainfall interception by three semi-arid canopies.Crossref | GoogleScholarGoogle Scholar |
Dunbabin V, Rengel Z, Diggle AJ (2004) Simulating form and function of root systems: efficiency of nitrate uptake is dependent on root system architecture and the spatial and temporal variability of nitrate supply. Functional Ecology 18, 204–211.
| Simulating form and function of root systems: efficiency of nitrate uptake is dependent on root system architecture and the spatial and temporal variability of nitrate supply.Crossref | GoogleScholarGoogle Scholar |
Eamus D, Prior L (2001) Ecophysiology of trees of seasonally dry tropics: comparisons among phenologies. Advances in Ecological Research 32, 113–197.
| Ecophysiology of trees of seasonally dry tropics: comparisons among phenologies.Crossref | GoogleScholarGoogle Scholar |
Earnshaw MJ, Carver KA, Gunn TC, Kerenga K, Harvey V, Griffiths H, Broadmeadow MSJ (1990) Photosynthetic pathway, chilling tolerance and cell sap osmotic potential values of grasses along an altitudinal gradient in Papua New Guinea. Oecologia 84, 280–288.
Edwards EJ, Still CJ, Donoghue MJ (2007) The relevance of phylogeny to studies of global change. Trends in Ecology & Evolution 22, 243–249.
| The relevance of phylogeny to studies of global change.Crossref | GoogleScholarGoogle Scholar |
Ehleringer JR (1991) 13C/12C fractionation and its utility in terrestrial plant studies. In ‘Carbon isotopes techniques’. (Eds DC Coleman, B Fry) pp. 187–200 (Academic Press: London)
Ehleringer JR, Osmond CB (2000) Stable isotopes. In ‘Plant physiological ecology: field methods and instrumentation’. (Eds RW Pearcy, J Ehleringer, HA Mooney, PW Rundel) pp. 281–300 (Kluwer Academic Publishers: Dordrecht, The Netherlands)
Ehleringer JR, Cerling TE, Helliker BR (1997) C4 photosynthesis, atmospheric CO2, and climate. Oecologia 112, 285–299.
| C4 photosynthesis, atmospheric CO2, and climate.Crossref | GoogleScholarGoogle Scholar |
Eissenstat DM, Yanai RD (1997) The ecology of root lifespan. Advances in Ecological Research 27, 1–60.
| The ecology of root lifespan.Crossref | GoogleScholarGoogle Scholar |
Elberse WTH, Berendse F (1993) A comparative study of the growth and morphology of eight grass species from habitats with different nutrient availabilities. Functional Ecology 7, 223–229.
| A comparative study of the growth and morphology of eight grass species from habitats with different nutrient availabilities.Crossref | GoogleScholarGoogle Scholar |
Eliáš P (1985) Leaf indices of woodland herbs as indicators of habitat conditions. Ekologia 4, 289–295.
Ellenberg H, Müller-Dombois D (1967) A key to Raunkiaer plant life forms with revised subdivisions. Berichte des geobotanischen Institutes der ETH. Stiftung Rübel 37, 56–73.
Ellsworth DS, Reich PB (1992) Water relations and gas exchange of Acer saccharum seedlings in contrasting natural light and water regimes. Tree Physiology 10, 1–20.
| Water relations and gas exchange of Acer saccharum seedlings in contrasting natural light and water regimes.Crossref | GoogleScholarGoogle Scholar |
Enquist BJ (2002) Universal scaling in tree and vascular plant allometry: toward a general quantitative theory linking plant form and function from cells to ecosystems. Tree Physiology 22, 1045–1064.
| Universal scaling in tree and vascular plant allometry: toward a general quantitative theory linking plant form and function from cells to ecosystems.Crossref | GoogleScholarGoogle Scholar |
Enríquez S, Duarte CM, Sand-Jensen K, Nielsen SL (1996) Broad-scale comparison of photosynthetic rates across phototrophic organisms. Oecologia 108, 197–206.
Etlinger MG, Beall FC (2004) Development of a laboratory protocol for fire performance of landscape plants. International Journal of Wildland Fire 13, 479–488.
| Development of a laboratory protocol for fire performance of landscape plants.Crossref | GoogleScholarGoogle Scholar |
Evans GC (1972) ‘The quantitative analysis of plant growth.’ (Blackwell Scientific Publications: Oxford, UK)
Everham EM, Brokaw NVL (1996) Forest damage and recovery from catastrophic wind. Botanical Review 62, 113–185.
| Forest damage and recovery from catastrophic wind.Crossref | GoogleScholarGoogle Scholar |
Ewel JJ, Bigelow SW (1996) Plant life forms and ecosystem functioning. In ‘Biodiversity and ecosystem processes in tropical forests’. (Eds GH Orians, R Dirzo, JH Cushman) pp. 101–126 (Springer: Berlin)
Faith DP, Magallón S, Hendry AP, Conti E, Yahara T, Donoghue MJ (2010) Evosystem services: an evolutionary perspective on the links between biodiversity and human well-being. Current Opinion in Environmental Sustainability 2, 66–74.
| Evosystem services: an evolutionary perspective on the links between biodiversity and human well-being.Crossref | GoogleScholarGoogle Scholar |
FAO (1999) ‘Soil salinity assessment. Methods and interpretation of electrical conductivity measurements.’ FAO irrigation and drainage paper 57 (FAO: Rome)
Farquhar GD, Ehleringer JR, Hubick KT (1989) Carbon isotope discrimination and photosynthesis. Annual Review of Plant Physiology and Plant Molecular Biology 40, 503–537.
| Carbon isotope discrimination and photosynthesis.Crossref | GoogleScholarGoogle Scholar |
Feild TS, Balun L (2008) Xylem hydraulic and photosynthetic function of Gnetum (Gnetales) species from Papua New Guinea. New Phytologist 177, 665–675.
| Xylem hydraulic and photosynthetic function of Gnetum (Gnetales) species from Papua New Guinea.Crossref | GoogleScholarGoogle Scholar |
Field C, Mooney HA (1986) The photosynthesis–nitrogen relationship in wild plants. In ‘On the economy of plant form and function’. (Ed. TJ Givnish) pp. 25–55 (Cambridge University Press: Cambridge, UK)
Finzi AC, Van Breemen N, Canham CD (1998) Canopy tree-soil interactions within temperate forests: species effects on soil carbon and nitrogen. Ecological Applications 8, 440–446.
Fischer M, Stöcklin J (1997) Local extinctions of plants in remnants of extensively used calcareous grasslands 1950–85 Conservation Biology 11, 727–737.
| Local extinctions of plants in remnants of extensively used calcareous grasslands 1950–85Crossref | GoogleScholarGoogle Scholar |
Fischer S, Poschlod P, Beinlich B (1996) Experimental studies on the dispersal of plants and animals on sheep in calcareous grasslands. Journal of Applied Ecology 33, 1206–1221.
| Experimental studies on the dispersal of plants and animals on sheep in calcareous grasslands.Crossref | GoogleScholarGoogle Scholar |
Fisher JB (1986) Branching patterns and angles in trees. In ‘On the economy of plant form and function’. (Ed. TJ Givnish) pp. 493–523 (Cambridge University Press: Cambridge, UK)
Fitter A (1996) Characteristics and functions of root systems. In ‘Plant roots: the hidden half’. 2nd edn. (Eds Y Waisel, A Eshel, U Kafkafi) pp. 1–20 (Marcel Dekker: New York)
Flowers TJ, Colmer TD (2008) Salinity tolerance in halophytes. New Phytologist 179, 945–963.
| Salinity tolerance in halophytes.Crossref | GoogleScholarGoogle Scholar |
Flowers TJ, Troke PF, Yeo AR (1977) The mechanism of salt tolerance in halophytes. Annual Review of Plant Physiology 28, 89–121.
| The mechanism of salt tolerance in halophytes.Crossref | GoogleScholarGoogle Scholar |
Flowers TJ, Hajibagheri MA, Clipson NJW (1986) Halophytes. The Quarterly Review of Biology 61, 313–337.
| Halophytes.Crossref | GoogleScholarGoogle Scholar |
Forget P, Wenny D (2005) How to elucidate seed fate? A review of methods used to study seed removal and secondary seed dispersal. In ‘Seed fate: predation, dispersal and seedling establishment’. (Eds P Forget, J Lambert, P Hulme, S Vander Wal) pp. 379–394 (CABI Publishing: Wallingford, UK)
Fortunel C, Garnier E, Joffre J, Kazakou E, Quested H, Grigulis K, Lavorel S, Ansquer P, Castro H, Cruz P, Doležal J, Eriksson O, Freitas H, Golodets C, Jouany C, Kigel J, Kleyer M, Lehsten V, Lepš J, Meier T, Pakeman R, Papadimitriou M, Papanastasis VP, Quétier F, Robson M, Sternberg M, Theau J-P, Thébault A, Zarovali M (2009) Leaf traits capture the effects of land use changes and climate on litter decomposability of grasslands across Europe. Ecology 90, 598–611.
| Leaf traits capture the effects of land use changes and climate on litter decomposability of grasslands across Europe.Crossref | GoogleScholarGoogle Scholar |
Fortunel C, Fine PVA, Baraloto C (2012) Leaf, stem and root tissue strategies across 758 Neotropical tree species. Functional Ecology 26, 1153–1161.
| Leaf, stem and root tissue strategies across 758 Neotropical tree species.Crossref | GoogleScholarGoogle Scholar |
Freschet GT, Cornelissen JHC, Van Logtestijn RSP, Aerts R (2010) Evidence of the ‘plant economics spectrum’ in a Subarctic flora. Journal of Ecology 98, 362–373.
| Evidence of the ‘plant economics spectrum’ in a Subarctic flora.Crossref | GoogleScholarGoogle Scholar |
Freschet GT, Aerts R, Cornelissen JHC (2012) A plant economics spectrum of litter decomposability. Functional Ecology 26, 56–65.
| A plant economics spectrum of litter decomposability.Crossref | GoogleScholarGoogle Scholar |
Funes G, Basconcelo S, Díaz S, Cabido M (1999) Seed bank dynamics of Lachemilla pinnata (Rosaceae) in different plant communities of mountain grassland in central Argentina. Annales Botanici Fennici 36, 109–114.
Gardner RO (1975) Overview of botanical clearing technique. Stain Technology 50, 99–105.
Garnier E, Laurent G (1994) Leaf anatomy, specific mass and water content in congeneric annual and perennial grass species. New Phytologist 128, 725–736.
| Leaf anatomy, specific mass and water content in congeneric annual and perennial grass species.Crossref | GoogleScholarGoogle Scholar |
Garnier E, Navas M-L (2012) A trait-based approach to comparative functional plant ecology: concepts, methods and applications for agroecology. Agronomy for Sustainable Development 32, 365–399.
| A trait-based approach to comparative functional plant ecology: concepts, methods and applications for agroecology.Crossref | GoogleScholarGoogle Scholar |
Garnier E, Shipley B, Roumet C, Laurent G (2001a) A standardized protocol for the determination of specific leaf area and leaf dry matter content. Functional Ecology 15, 688–695.
| A standardized protocol for the determination of specific leaf area and leaf dry matter content.Crossref | GoogleScholarGoogle Scholar |
Garnier E, Laurent G, Debain S, Berthelier P, Ducout B, Roumet C, Navas M-L (2001b) Consistency of species ranking based on functional leaf traits. New Phytologist 152, 69–83.
| Consistency of species ranking based on functional leaf traits.Crossref | GoogleScholarGoogle Scholar |
Garnier E, Cortez J, Billès G, Navas M-L, Roumet C, Debussche M, Laurent G, Blanchard A, Aubry D, Bellmann A, Neill C, Toussaint J-P (2004) Plant functional markers capture ecosystem properties during secondary succession. Ecology 85, 2630–2637.
| Plant functional markers capture ecosystem properties during secondary succession.Crossref | GoogleScholarGoogle Scholar |
Garnier E, Lavorel S, Ansquer P, Castro H, Cruz P, Dolezal J, Eriksson O, Fortunel C, Freitas H, Golodets C, Grigulis K, Jouany C, Kazakou E, Kigel J, Kleyer M, Lehsten V, Lepš J, Meier T, Pakeman R, Papadimitriou M, Papanastasis VP, Quested H, Quétier F, Robson M, Roumet C, Rusch G, Skarpe C, Sternberg M, Theau J-P, Vile D, Zarovali MP (2007) Assessing the effects of land-use change on plant traits, communities and ecosystem functioning in grasslands: a standardized methodology and lessons from an application to 11 European sites. Annals of Botany 99, 967–985.
| Assessing the effects of land-use change on plant traits, communities and ecosystem functioning in grasslands: a standardized methodology and lessons from an application to 11 European sites.Crossref | GoogleScholarGoogle Scholar |
Gartner BL (1995) ‘Plant stems: physiology and functional morphology.’ (Academic Press: San Diego, CA)
Gartner BL, Moore JC, Gardiner BA (2004) Gas in stems: abundance and potential consequences for tree biomechanics. Tree Physiology 24, 1239–1250.
| Gas in stems: abundance and potential consequences for tree biomechanics.Crossref | GoogleScholarGoogle Scholar |
Garwood NC (1996) Functional morphology of tropical tree seedlings. In ‘The ecology of tropical forest tree seedlings’. (Ed. MD Swaine) pp. 59–129 (Parthenon: New York)
Gatsuk LE, Smirnova OV, Vorontzova LI, Zaugolnova LB, Zhukova LA (1980) Age states of plants of various growth forms: a review. Journal of Ecology 68, 675–696.
| Age states of plants of various growth forms: a review.Crossref | GoogleScholarGoogle Scholar |
Gaucherand S, Lavorel S (2007) New method for rapid assessment of the functional composition of herbaceous plant communities. Austral Ecology 32, 927–936.
| New method for rapid assessment of the functional composition of herbaceous plant communities.Crossref | GoogleScholarGoogle Scholar |
Gaudet CL, Keddy PA (1988) A comparative approach to predicting competitive ability from plant traits. Nature 334, 242–243.
| A comparative approach to predicting competitive ability from plant traits.Crossref | GoogleScholarGoogle Scholar |
Gill AM, Zylstra P (2005) Flammability of Australian forests. Australian Forestry 68, 88–94.
Givnish TJ (1979) On the adaptive significance of leaf form. In ‘Topics in plant population biology’. (Ed. OT Solbrig, S Jain, GB Johnson, PH Raven) pp. 375–407 (Macmillan: London)
Givnish TJ (1987) Comparative studies of leaf form: assessing the relative roles of selective pressures and phylogenetic constarints. New Phytologist 106, 131–160.
| Comparative studies of leaf form: assessing the relative roles of selective pressures and phylogenetic constarints.Crossref | GoogleScholarGoogle Scholar |
Gowda JH, Palo RT (2003) Age-related changes in defensive traits of Acacia tortilis Hayne. African Journal of Ecology 41, 218–223.
| Age-related changes in defensive traits of Acacia tortilis Hayne.Crossref | GoogleScholarGoogle Scholar |
Gowda J, Raffaele E (2004) Spine production is induced by fire: a natural experiment with three Berberis species. Acta Oecologica 26, 239–245.
| Spine production is induced by fire: a natural experiment with three Berberis species.Crossref | GoogleScholarGoogle Scholar |
Graça MAS, Bärlocher F, Gessner MO (2005) ‘Methods to study litter decomposition.’ (Springer: Dordrecht, The Netherlands)
Green DS, Kruger EL (2001) Light-mediated constraints on leaf function correlate with leaf structure among deciduous and evergreen tree species. Tree Physiology 21, 1341–1346.
| Light-mediated constraints on leaf function correlate with leaf structure among deciduous and evergreen tree species.Crossref | GoogleScholarGoogle Scholar |
Grime JP (1979) Competition and the struggle for existence. In ‘Population dynamics’. (Eds RM Anderson, BD Turner, LR Taylor) pp. 123–139 (Blackwell Scientific Publications: Oxford, UK)
Grime JP, Hunt R (1975) Relative growth-rate: its range and adaptive significance in a local flora. Journal of Ecology 63, 393–422.
| Relative growth-rate: its range and adaptive significance in a local flora.Crossref | GoogleScholarGoogle Scholar |
Grime JP, Blythe GM, Thornton JD (1970) Food selection by the snail Cepaea nemoralis L. In ‘Animal populations in relation to their food resources’. (Ed. A Watson) pp. 73–99 (Blackwell Scientific Publications: Oxford, UK)
Grime JP, Cornelissen JHC, Thompson K, Hodgson JG (1996) Evidence of a causal connection between anti-herbivore defense and the decomposition rate of leaves. Oikos 77, 489–494.
| Evidence of a causal connection between anti-herbivore defense and the decomposition rate of leaves.Crossref | GoogleScholarGoogle Scholar |
Grime JP, Thompson K, Hunt R, Hodgson JG, Cornelissen JHC, Rorison IH, Hendry GAF, Ashenden TW, Askew AP, Band SR, Booth RE, Bossard CC, Campbell BD, Cooper JEL, Davison AW, Gupta PL, Hall W, Hand DW, Hannah MA, Hillier SH, Hodkinson DJ, Jalili A, Liu Z, Mackey JML, Matthews N, Mowforth MA, Neal AM, Reader RJ, Reiling K, Ross-Fraser W, Spencer RE, Sutton F, Tasker DE, Thorpe PC, Whitehouse J (1997) Integrated screening validates primary axes of specialisation in plants. Oikos 79, 259–281.
| Integrated screening validates primary axes of specialisation in plants.Crossref | GoogleScholarGoogle Scholar |
Grubb PJ (1992) A positive distrust in simplicity – lessons from plant defences and from competition among plants and among animal. Journal of Ecology 80, 585–610.
| A positive distrust in simplicity – lessons from plant defences and from competition among plants and among animal.Crossref | GoogleScholarGoogle Scholar |
Gubsch M, Buchmann N, Schmid B, Schulze ED, Lipowsky A, Roscher C (2011) Differential effects of plant diversity on functional trait variation of grass species. Annals of Botany 107, 157–169.
| Differential effects of plant diversity on functional trait variation of grass species.Crossref | GoogleScholarGoogle Scholar |
Gurvich DE, Díaz S, Falczuk V, Pérez-Harguindeguy N, Cabido M, Thorpe C (2002) Foliar resistence to simulated extreme temperature events in contrasting plant functional and chorological types. Global Change Biology 8, 1139–1145.
| Foliar resistence to simulated extreme temperature events in contrasting plant functional and chorological types.Crossref | GoogleScholarGoogle Scholar |
Hanley ME, Lamont BB (2002) Relationships between physical and chemical attributes of congeneric seedlings: how important is seedling defence? Functional Ecology 16, 216–222.
| Relationships between physical and chemical attributes of congeneric seedlings: how important is seedling defence?Crossref | GoogleScholarGoogle Scholar |
Harrison SP, Marlon JR, Bartlein PJ (2010) Fire in the Earth system. In ‘Changing climates, earth systems and society’. (Ed. J Dodson) pp. 21–48 (Springer: Dordrecht, The Netherlands)
Hartley SE, Jones CG (1997) Plant chemistry and herbivory, or why the world is green. In ‘Plant ecology’. (Ed. MJ Crawley) pp. 284–324 (Blackwell Science: Oxford, UK)
Hattersley PW, Watson L (1992) Diversification of photosynthesis. In ‘Grass evolution and domestication’. (Ed. GP Chapman) pp. 38–116 (Cambridge University Press: London)
Hendry GAF, Grime JP (1993) ‘Methods in comparative plant ecology. A laboratory manual.’ (Chapman and Hall: London)
Herwitz SR (1985) Interception storage capacities of tropical rainforest canopy trees. Journal of Hydrology 77, 237–252.
| Interception storage capacities of tropical rainforest canopy trees.Crossref | GoogleScholarGoogle Scholar |
Hibberd JM, Quick WP (2002) Characteristics of C4 photosynthesis in stems and petioles of C3 flowering plants. Nature 415, 451–454.
| Characteristics of C4 photosynthesis in stems and petioles of C3 flowering plants.Crossref | GoogleScholarGoogle Scholar |
Higgins SI, Bond WJ, Trolliope WSW (2000) Fire, resprouting and variability: a recipe for grass-tree coexistence in savanna. Journal of Ecology 88, 213–229.
| Fire, resprouting and variability: a recipe for grass-tree coexistence in savanna.Crossref | GoogleScholarGoogle Scholar |
Hinckley TM, Lassoie JP, Running SW (1978) Temporal and spatial variations in the water status of forest trees. Forest Science 20, a0001–z0001.
Hirose T, Werger MJA (1995) Canopy structure and shoton slux sartitioning smong species in a herbaceous plant community. Ecology 76, 466–474.
| Canopy structure and shoton slux sartitioning smong species in a herbaceous plant community.Crossref | GoogleScholarGoogle Scholar |
Hladik A, Miquel S (1990) Seedling types and plant establishment in an African rain forest. In ‘Reproductive ecology of tropical plants’. (Eds KS Bawa, M Hadley) pp. 261–282 (UNESCO/Parthenon: Carnforth, UK)
Hodge A (2004) The plastic plant: root responses to heterogeneous supplies of nutrients. New Phytologist 162, 9–24.
| The plastic plant: root responses to heterogeneous supplies of nutrients.Crossref | GoogleScholarGoogle Scholar |
Hodgson JG, Wilson PJ, Hunt R, Grime JP, Thompson K (1999) Allocating C-S-R plant functional types: a soft approach to a hard problem. Oikos 85, 282–294.
| Allocating C-S-R plant functional types: a soft approach to a hard problem.Crossref | GoogleScholarGoogle Scholar |
Hodgson JG, Montserrat-Martí G, Charles M, Jones G, Wilson P, Shipley B, Sharafi M, Cerabolini BEL, Cornelissen JHC, Band SR, Bogard A, Castro-Díez P, Guerrero-Campo J, Palmer C, Pérez-Rontomé MC, Carter G, Hynd A, Romo-Díez A, de Torres Espuny L, Royo Pla F (2011) Is leaf dry matter content a better predictor of soil fertility than specific leaf area? Annals of Botany 108, 1337–1345.
| Is leaf dry matter content a better predictor of soil fertility than specific leaf area?Crossref | GoogleScholarGoogle Scholar |
Holbrook NM, Zwieniecki MA (2005) ‘Vascular transport in plants.’ (Elsevier Academic Press: London)
Horn HS (1971) ‘The adaptive geometry of trees.’ (Princeton University Press: Princeton, NJ)
Horneck DA, Miller RO (1998) Determination of total nitrogen in plant tissue. In ‘Handbook and reference methods for plant analysis’. (Ed. YP Kalra) pp. 75–84 (CRC Press: New York)
Howe HF, Smallwood J (1982) Ecology of seed dispersal. Annual Review of Ecology and Systematics 13, 201–228.
| Ecology of seed dispersal.Crossref | GoogleScholarGoogle Scholar |
Howe HF, Westley LC (1997) Ecology of pollination and seed dispersal. In ‘Plant ecology’. (Ed. MJ Crawley) pp. 262–283 (Blackwell: Oxford, UK)
Hulme PE (1998) Post-dispersal seed predation: consequences for plant demography and evolution. Perspectives in Plant Ecology, Evolution and Systematics 1, 32–46.
| Post-dispersal seed predation: consequences for plant demography and evolution.Crossref | GoogleScholarGoogle Scholar |
Hulshof CM, Swenson NG (2010) Variation in leaf functional trait values within and across individuals and species: an example from a Costa Rican dry forest. Functional Ecology 24, 217–223.
| Variation in leaf functional trait values within and across individuals and species: an example from a Costa Rican dry forest.Crossref | GoogleScholarGoogle Scholar |
Hunt R (1982) ‘Plant growth curves. The functional approach to plant growth analysis.’ (Edward Arnold: London)
Ibarra-Manriquez G, Martínez Ramos M, Oyama K (2001) Seedling functional types in a lowland rain forest in México. American Journal of Botany 88, 1801–1812.
| Seedling functional types in a lowland rain forest in México.Crossref | GoogleScholarGoogle Scholar |
Jackson RB (1999) The importance of root distributions for hydrology, biogeochemistry and ecosystem function. In ‘Integrating hydrology, ecosystem dynamics and biogeochemistry in complex landscapes’. (Eds JD Tenhunen, P Kabat) pp. 219–240 (Wiley: Chichester, UK)
Jackson JF, Adams DC, Jackson UB (1999) Allometry of constitutive defense: a model and a comparative test with tree bark and fire regime. American Naturalist 153, 614–632.
| Allometry of constitutive defense: a model and a comparative test with tree bark and fire regime.Crossref | GoogleScholarGoogle Scholar |
Jacobsen AL, Pratt RB, Davis SD, Ewers FW (2008) Comparative community physiology: nonconvergence in water relations among three semi-arid shrub communities. New Phytologist 180, 100–113.
| Comparative community physiology: nonconvergence in water relations among three semi-arid shrub communities.Crossref | GoogleScholarGoogle Scholar |
Jaureguiberry P, Bertone GA, Díaz S (2011) Device for the standard measurement of shoot flammability in the field. Austral Ecology 36, 821–829.
| Device for the standard measurement of shoot flammability in the field.Crossref | GoogleScholarGoogle Scholar |
Jennings DH (1976) The effects of sodium chloride on higher plants. Biological Reviews of the Cambridge Philosophical Society 51, 453–486.
| The effects of sodium chloride on higher plants.Crossref | GoogleScholarGoogle Scholar |
Jones G, Charles M, Bogaard A, Hodgson J (2010) Crops and weeds: the role of weed functional ecology in the identification of crop husbandry methods. Journal of Archaeological Science 37, 70–77.
| Crops and weeds: the role of weed functional ecology in the identification of crop husbandry methods.Crossref | GoogleScholarGoogle Scholar |
Jow WM, Bullock SH, Kummerow J (1980) Leaf turnover rates of Adenostoma fasciculatum (Rosaceae). American Journal of Botany 67, 256–261.
| Leaf turnover rates of Adenostoma fasciculatum (Rosaceae).Crossref | GoogleScholarGoogle Scholar |
Juneau KJ, Tarasoff CS (2012) Leaf area and water content changes after permanent and temporary storage. PLoS ONE 7, e42604
| Leaf area and water content changes after permanent and temporary storage.Crossref | GoogleScholarGoogle Scholar |
Kammesheidt L (1999) Forest recovery by root suckers and aboveground sprouts after slash-and-burn agriculture, fire and logging in Paraguay and Venezuela. Journal of Tropical Ecology 15, 143–157.
| Forest recovery by root suckers and aboveground sprouts after slash-and-burn agriculture, fire and logging in Paraguay and Venezuela.Crossref | GoogleScholarGoogle Scholar |
Kattge J, Díaz S, Lavorel S, Prentice IC, Leadley P, Bönisch G, Garnier E, Westoby M, Reich PB, Wright IJ, Cornelissen JHC, Violle C, Harrison SP, Van Bodegom PM, Reichstein M, Enquist BJ, Soudzilovskaia NA, Ackerly DD, Anand M, Atkin O, Bahn M, Baker TR, Baldocchi D, Bekker R, Blanco CC, Blonder B, Bond WJ, Bradstock R, Bunker DE, Casanoves F, Cavender-Bares J, Chambers JQ, Chapin III FS, Chave J, Coomes D, Cornwell WK, Craine JM, Dobrin BH, Duarte L, Durka W, Elser J, Esser G, Estiarte M, Fagan WF, Fang J, Fernández-Méndez F, Fidelis A, Finegan B, Flores O, Ford H, Frank D, Freschet GT, Fyllas NM, Gallagher RV, Green WA, Gutierrez AG, Hickler T, Higgins SI, Hodgson JG, Jalili A, Jansen S, Joly CA, Kerkhoff AJ, Kirkup D, Kitajima K, Kleyer M, Klotz S, Knops JMH, Kramer K, Kühn I, Kurokawa H, Laughlin D, Lee TD, Leishman M, Lens F, Lenz T, Lewis SL, Lloyd J, Llusià J, Louault F, Ma S, Mahecha MD, Manning P, Massad T, Medlyn BE, Messie J, Moles AT, Müller SC, Nadrowski K, Naeem S, Niinemets Ü, Nöllert S, Nüske A, Ogaya R, Oleksyn J, Onipchenko VG, Onoda Y, Ordoñez J, Overbeck G, Ozinga WA, Patiño S, Paula S, Pausas JG, Peñuelas J, Phillips OL, Pillar V, Poorter H, Poorter L, Poschlod P, Prinzing A, Proulx R, Rammig A, Reinsch S, Reu B, Sack L, Salgado-Negret B, Sardans J, Shiodera S, Shipley B, Siefert A, Sosinski E, Soussana J-F, Swaine E, Swenson N, Thompson K, Thornton P, Waldram M, Weiher E, White M, White S, Wright SJ, Yguel B, Zaehle S, Zanne AE, Wirth C (2011) TRY – a global database of plant traits. Global Change Biology 17, 2905–2935.
| TRY – a global database of plant traits.Crossref | GoogleScholarGoogle Scholar |
Kazakou E, Violle C, Roumet C, Pintor C, Gimenez O, Garnier E (2009) Litter quality and decomposability of species from a Mediterranean succession depend on leaf traits but not on nitrogen supply. Annals of Botany 104, 1151–1161.
| Litter quality and decomposability of species from a Mediterranean succession depend on leaf traits but not on nitrogen supply.Crossref | GoogleScholarGoogle Scholar |
King DA, Davies SJ, Noor NSM (2006) Growth and mortality are related to adult tree size in a Malaysian mixed dipterocarp forest. Forest Ecology and Management 223, 152–158.
| Growth and mortality are related to adult tree size in a Malaysian mixed dipterocarp forest.Crossref | GoogleScholarGoogle Scholar |
Kitajima K (1994) Relative importance of photosynthetic traits and allocation patterns as correlates of seedling shade tolerance of 13 tropical trees. Oecologia 98, 419–428.
| Relative importance of photosynthetic traits and allocation patterns as correlates of seedling shade tolerance of 13 tropical trees.Crossref | GoogleScholarGoogle Scholar |
Kitajima K (1996) Cotyledon functional morphology, patterns of seed reserve utilization and regeneration niches of tropical tree seedlings. In ‘The ecology of tropical forest tree seedlings’. (Ed. MD Swaine) pp. 193–210 (UNESCO: Paris)
Kitajima K, Poorter L (2010) Tissue-level leaf toughness, but not lamina thickness, predicts sapling leaf lifespan and shade tolerance of tropical tree species. New Phytologist 186, 708–721.
| Tissue-level leaf toughness, but not lamina thickness, predicts sapling leaf lifespan and shade tolerance of tropical tree species.Crossref | GoogleScholarGoogle Scholar |
Kleyer M, Bekker RM, Knevel IC, Bakker JP, Thompson K, Sonnenschein M, Poschlod P, Van Groenendael JM, Klimeš L, Klimešová J, Klotz S, Rusch GM, Hermy M, Adriaens D, Boedeltje G, Bossuyt B, Dannemann A, Endels P, Götzenberger L, Hodgson JG, Jackel A-K, Kühn I, Kunzmann D, Ozinga WA, Römermann C, Stadler M, Schlegelmilch J, Steendam HJ, Tackenberg O, Wilmann B, Cornelissen JHC, Eriksson O, Garnier E, Peco B (2008) The LEDA traitbase: a database of life-history traits of the northwest European flora. Journal of Ecology 96, 1266–1274.
| The LEDA traitbase: a database of life-history traits of the northwest European flora.Crossref | GoogleScholarGoogle Scholar |
Klimeš L, Klimešová J (2005) Clonal traits. In ‘The LEDA traitbase. Collecting and measuring standards of life-history traits of the northwest European flora’. (Eds IC Knevel, RM Bekker, D Kunzmann, M Stadler, K Thompson) pp. 66–88 (University of Groningen: Groningen, The Netherlands)
Klimeš L, Klimešová J, Hendriks RJJ, Van Groenendael JM (1997) Clonal plant architecture: a comparative analysis of form and function. In ‘The ecology and evolution of clonal plants’. (Eds H De Kroon, JM Van Groenendael) pp. 1–29 (Backhuys Publishers: Leiden, The Netherlands)
Klimešová J, Klimeš L (2007) Bud banks and their role in vegetative regeneration – A literature review and proposal for simple classification and assessment. Perspectives in Plant Ecology, Evolution and Systematics 8, 115–129.
| Bud banks and their role in vegetative regeneration – A literature review and proposal for simple classification and assessment.Crossref | GoogleScholarGoogle Scholar |
Knapp AK, Carter GA (1998) Variability in leaf optical properties among 26 species from a broad range of habitats. American Journal of Botany 85, 940–946.
| Variability in leaf optical properties among 26 species from a broad range of habitats.Crossref | GoogleScholarGoogle Scholar |
Knevel IC, Bekker RM, Kleyer M (2003) Life-history traits of the northwest European flora: the LEDA database. Journal of Vegetation Science 14, 611–614.
| Life-history traits of the northwest European flora: the LEDA database.Crossref | GoogleScholarGoogle Scholar |
Kohyama T, Suzuki E, Partomihardjo T, Yamada T, Kubo T (2003) Tree species differentiation in growth, recruitment and allometry in relation to maximum height in a Bornean mixed dipterocarp forest. Journal of Ecology 91, 797–806.
| Tree species differentiation in growth, recruitment and allometry in relation to maximum height in a Bornean mixed dipterocarp forest.Crossref | GoogleScholarGoogle Scholar |
Kocacinar F, Sage RF (2003) Photosynthetic pathway alters xylem structure and hydraulic function in herbaceous plants. Plant, Cell & Environment 26, 2015–2026.
| Photosynthetic pathway alters xylem structure and hydraulic function in herbaceous plants.Crossref | GoogleScholarGoogle Scholar |
Korning J, Thomsen K (1994) A new method for measuring tree height in tropical rain forest. Journal of Vegetation Science 5, 139–140.
| A new method for measuring tree height in tropical rain forest.Crossref | GoogleScholarGoogle Scholar |
Lambers H, Poorter H (1992) Inherent variation in growth rate between higher plants: a search for physiological causes and ecological consequences. Advances in Ecological Research 23, 187–261.
| Inherent variation in growth rate between higher plants: a search for physiological causes and ecological consequences.Crossref | GoogleScholarGoogle Scholar |
Lambers H, Finnegan PM, Laliberté E, Pearse SJ, Ryan MH, Shane MW, Veneklaas EJ (2011) Phosphorus nutrition of proteaceae in severely phosphorus-impoverished soils: are there lessons to be learned for future crops? Plant Physiology 156, 1058–1066.
| Phosphorus nutrition of proteaceae in severely phosphorus-impoverished soils: are there lessons to be learned for future crops?Crossref | GoogleScholarGoogle Scholar |
Larson DW (2001) The paradox of great longevity in a short-lived tree species. Experimental Gerontology 36, 651–673.
| The paradox of great longevity in a short-lived tree species.Crossref | GoogleScholarGoogle Scholar |
Lavorel S, Garnier E (2002) Predicting changes in community composition and ecosystem functioning from plant traits: revisiting the Holy Grail. Functional Ecology 16, 545–556.
| Predicting changes in community composition and ecosystem functioning from plant traits: revisiting the Holy Grail.Crossref | GoogleScholarGoogle Scholar |
Lavorel S, Díaz S, Cornelissen JHC, Garnier E, Harrison SP, McIntyre S, Pausas JG, Pérez-Harguindeguy N, Roumet C, Urcelay C (2007) Plant functional types: are we getting any closer to the holy grail? In ‘Terrestrial ecosystems in a changing world’. (Ed. PD Canadell JG, Pitelka L) pp. 149–160 (Springer-Verlag: Berlin)
Lavorel S, Grigulis K, McIntyre S, Williams NSG, Garden D, Dorrough SB, Quétier F, Thébault A, Bonis A (2008) Assessing functional diversity in the field – methodology matters! Functional Ecology 22, 134–147.
Leck MA, Parker VT, Simpson RL (2008) ‘Seedling ecology and evolution.’ (Cambridge University Press: Cambridge, UK)
Leishman MR, Westoby M (1998) Seed size and shape are not related to persistence in soil in Australia in the same way as in Britain. Functional Ecology 12, 480–485.
| Seed size and shape are not related to persistence in soil in Australia in the same way as in Britain.Crossref | GoogleScholarGoogle Scholar |
Leishman M, Wright I, Moles A, Westoby M (2000) The evolutionary ecology of seed size. In ‘The ecology of regeneration in plant communities’. 2nd edn. (Ed. M Fenner) pp. 31–57 (CAB International: London)
Lenz TI, Wright IJ, Westoby M (2006) Interrelations among pressure-volume curve traits across species and water availability gradients. Physiologia Plantarum 127, 423–433.
| Interrelations among pressure-volume curve traits across species and water availability gradients.Crossref | GoogleScholarGoogle Scholar |
Levitt J (1980) ‘Responses of plants to environmental stresses.’ (Academic Press: New York)
Linder CR, Moore LA, Jackson RB (2000) A universal molecular method for identifying underground plant parts to species. Molecular Ecology 9, 1549–1559.
| A universal molecular method for identifying underground plant parts to species.Crossref | GoogleScholarGoogle Scholar |
Litton CM, Raich JW, Ryan MG (2007) Carbon allocation in forest ecosystems. Global Change Biology 13, 2089–2109.
| Carbon allocation in forest ecosystems.Crossref | GoogleScholarGoogle Scholar |
Loehle C (1988) Tree life histories: the role of defenses. Canadian Journal of Forest Research 18, 209–222.
| Tree life histories: the role of defenses.Crossref | GoogleScholarGoogle Scholar |
Lucas PW, Choong MF, Tan HTW, Turner IM, Berrick AJ (1991) The fracture toughness of the leaf of the dicotyledon Calophyllum inophyllum L (Guttiferae). Philosophical Transactions of the Royal Society of London. Series B. Biological Sciences 334, 95–106.
| The fracture toughness of the leaf of the dicotyledon Calophyllum inophyllum L (Guttiferae).Crossref | GoogleScholarGoogle Scholar |
Lucas PW, Turner IM, Dominy NJ, Yamashita N (2000) Mechanical defences to herbivory. Annals of Botany 86, 913–920.
| Mechanical defences to herbivory.Crossref | GoogleScholarGoogle Scholar |
Lüttge (1997) ‘Physiolgical ecology of tropical plants.’ (Springer-Verlag: Berlin)
Ma S, Baldocchi DD, Mambelli S, Dawson TE (2011) Are temporal variations of leaf traits responsible for seasonal and inter-annual variability in ecosystem CO2 exchange? Functional Ecology 25, 258–270.
| Are temporal variations of leaf traits responsible for seasonal and inter-annual variability in ecosystem CO2 exchange?Crossref | GoogleScholarGoogle Scholar |
Maas EV, Hoffman GJ (1977) Crop salt tolerance – current assessment. Journal of the Irrigation and Drainage Division 103, 115–134.
Mace GM, Cramer W, Díaz S, Faith DP, Larigauderie A, Le Prestre P, Palmer M, Perrings C, Scholes RJ, Walpole M, Walther BA, Watson JEM, Mooney HA (2010) Biodiversity targets after 2010 Current Opinion in Environmental Sustainability 2, 3–8.
| Biodiversity targets after 2010Crossref | GoogleScholarGoogle Scholar |
Maherali H, DeLucia EH (2001) Influence of climate-driven shifts in biomass allocation on water transport and storage in ponderosa pine. Oecologia 129, 481–491.
Maherali H, Pockman WT, Jackson RB (2004) Adaptive variation in the vulnerability of woody plants to xylem cavitation. Ecology 85, 2184–2199.
| Adaptive variation in the vulnerability of woody plants to xylem cavitation.Crossref | GoogleScholarGoogle Scholar |
Mäkelä A, Vanninen P (2001) Vertical structure of Scots pine crowns in different age and size classes. Trees 15, 385–392.
| Vertical structure of Scots pine crowns in different age and size classes.Crossref | GoogleScholarGoogle Scholar |
Marschner H (2012) ‘Mineral nutrition of higher plants.’ (Academic Press: London)
Martorell C, Ezcurra E (2007) The narrow-leaf syndrome: a functional and evolutionary approach to the form of fog-harvesting rosette plants. Oecologia 151, 561–573.
| The narrow-leaf syndrome: a functional and evolutionary approach to the form of fog-harvesting rosette plants.Crossref | GoogleScholarGoogle Scholar |
Maseda PH, Fernández RJ (2006) Stay wet or else: three ways in which plants can adjust hydraulically to their environment. Journal of Experimental Botany 57, 3963–3977.
| Stay wet or else: three ways in which plants can adjust hydraulically to their environment.Crossref | GoogleScholarGoogle Scholar |
Mazer J (1989) Ecological, taxonomic, and life history correlates of seed mass among Indiana dune angiosperms. Ecological Monographs 59, 153–175.
| Ecological, taxonomic, and life history correlates of seed mass among Indiana dune angiosperms.Crossref | GoogleScholarGoogle Scholar |
McDowell N, Barnard H, Bond B, Hinckley T, Hubbard R, Ishii H, Köstner B, Magnani F, Marshall J, Meinzer F, Phillips N, Ryan M, Whitehead D (2002) The relationship between tree height and leaf area: sapwood area ratio. Oecologia 132, 12–20.
| The relationship between tree height and leaf area: sapwood area ratio.Crossref | GoogleScholarGoogle Scholar |
McGill BJ, Enquist BJ, Weiher E, Westoby M (2006) Rebuilding community ecology from functional traits. Trends in Ecology & Evolution 21, 178–185.
| Rebuilding community ecology from functional traits.Crossref | GoogleScholarGoogle Scholar |
McIntyre S, Lavorel S (2001) Livestock grazing in subtropical pastures: steps in the analysis of attribute response and plant functional types. Journal of Ecology 89, 209–226.
| Livestock grazing in subtropical pastures: steps in the analysis of attribute response and plant functional types.Crossref | GoogleScholarGoogle Scholar |
McIntyre S, Lavorel S, Landsberg J, Forbes TDA (1999) Disturbance response in vebetation – towards a global perspective on functional traits. Journal of Vegetation Science 10, 621–630.
| Disturbance response in vebetation – towards a global perspective on functional traits.Crossref | GoogleScholarGoogle Scholar |
Medina E (1999) Tropical forests: diversity and function of dominant life-forms. In ‘Handbook of plant functional ecology’. (Eds FI Pugnaire, F Valladares) pp. 407–448 (Marcel Dekker: New York)
Meinzer FC, Clearwater MJ, Goldstein G (2001) Water transport in trees: current perspectives, new insights and some controversies. Environmental and Experimental Botany 45, 239–262.
| Water transport in trees: current perspectives, new insights and some controversies.Crossref | GoogleScholarGoogle Scholar |
Mencuccini M (2003) The ecological significance of long-distance water transport: short-term regulation, long-term acclimation and the hydraulic costs of stature across plant life forms. Plant, Cell & Environment 26, 163–182.
| The ecological significance of long-distance water transport: short-term regulation, long-term acclimation and the hydraulic costs of stature across plant life forms.Crossref | GoogleScholarGoogle Scholar |
Messier J, McGill BJ, Lechowicz MJ (2010) How do traits vary across ecological scales? A case for trait-based ecology. Ecology Letters 13, 838–848.
| How do traits vary across ecological scales? A case for trait-based ecology.Crossref | GoogleScholarGoogle Scholar |
Meyer LD (1994) Rainfall simulators for soil conservation research. In ‘Soil erosion research methods’. (Ed. R Lal) pp. 83–104 (St Lucie Press: Delray Beach, FL)
Milla R, Reich PB (2007) The scaling of leaf area and mass: the cost of light interception increases with leaf size. Proceedings. Biological Sciences 274, 2109–2115.
| The scaling of leaf area and mass: the cost of light interception increases with leaf size.Crossref | GoogleScholarGoogle Scholar |
Milton SJ (1991) Plant spinescence in arid southern Africa – does moisture mediate selection by mammals. Oecologia 87, 279–287.
| Plant spinescence in arid southern Africa – does moisture mediate selection by mammals.Crossref | GoogleScholarGoogle Scholar |
Mohr H, Schopfer P (1995) ‘Plant physiology.’ 4th edn. (Springer: Berlin)
Moles A, Westoby M (2006) Seed size and plant strategy across the whole life cycle Oikos 113, 91–105.
| Seed size and plant strategy across the whole life cycleCrossref | GoogleScholarGoogle Scholar |
Moles A, Ackerly D, Webb C, Tweddle J, Dickie J, Pitman A, Westoby M (2005) Factors that shape seed mass evolution. Proceedings of the National Academy of Sciences, USA 102, 10540–10544.
| Factors that shape seed mass evolution.Crossref | GoogleScholarGoogle Scholar |
Moles AT, Warton DI, Warman L, Swenson NG, Laffan SW, Zanne AE, Pitman A, Hemmings FA, Leishman MR (2009) Global patterns in plant height. Journal of Ecology 97, 923–932.
| Global patterns in plant height.Crossref | GoogleScholarGoogle Scholar |
Moreira B, Tavsanoglu Ç, Pausas JG (2012) Local versus regional intraspecific variability in regeneration traits. Oecologia 168, 671–677.
| Local versus regional intraspecific variability in regeneration traits.Crossref | GoogleScholarGoogle Scholar |
Munns R, Husain S, Rivelli AR, James RA, Condon AG, Lindsay MP, Lagudah ES, Schachtman DP, Hare RA (2002) Avenues for increasing salt tolerance of crops, and the role of physiologically based selection traits. Plant and Soil 247, 93–105.
| Avenues for increasing salt tolerance of crops, and the role of physiologically based selection traits.Crossref | GoogleScholarGoogle Scholar |
Mutch RW (1970) Wildland fires and ecosystems – A hypothesis. Ecology 51, 1046–1051.
| Wildland fires and ecosystems – A hypothesis.Crossref | GoogleScholarGoogle Scholar |
Myers J, Vellend M, Gardescu S, Marks P (2004) Seed dispersal by white-tailed deer: implications for long-distance dispersal, invasion, and migration of plants in eastern North America. Oecologia 139, 35–44.
| Seed dispersal by white-tailed deer: implications for long-distance dispersal, invasion, and migration of plants in eastern North America.Crossref | GoogleScholarGoogle Scholar |
Newman EI (1966) A method of estimating the total root length of a sample. Journal of Applied Ecology 3, 139–145.
| A method of estimating the total root length of a sample.Crossref | GoogleScholarGoogle Scholar |
Ng FSP (1978) Strategies of establishment in Malayan forest trees. In ‘Tropical trees as living systems’. (Eds PB Tomlinson, MH Zimmerman) pp. 129–162 (Cambridge University Press: Cambridge, UK)
Niinemets Ü (2001) Global-scale climatic controls of leaf dry mass per area,density and thickness in trees and shrubs. Ecology 82, 453–469.
| Global-scale climatic controls of leaf dry mass per area,density and thickness in trees and shrubs.Crossref | GoogleScholarGoogle Scholar |
Niinemets Ü, Portsmuth A, Tena D, Tobias M, Matesanz S, Valladares F (2007) Do we underestimate the importance of leaf size in plant economics? Disproportional scaling of support costs within the spectrum of leaf physiognomy. Annals of Botany 100, 283–303.
| Do we underestimate the importance of leaf size in plant economics? Disproportional scaling of support costs within the spectrum of leaf physiognomy.Crossref | GoogleScholarGoogle Scholar |
Niklas KJ (1994) ‘Plant allometry: the scaling of form and process.’ (The University of Chicago Press: Chicago, IL)
Niklas KJ, Cobb ED, Niinemets U, Reich PB, Sellin A, Shipley B, Wright IJ (2007) “Diminishing returns” in the scaling of functional leaf traits across and within species groups. Proceedings of the National Academy of Sciences, USA 104, 8891–8896.
| “Diminishing returns” in the scaling of functional leaf traits across and within species groups.Crossref | GoogleScholarGoogle Scholar |
Noble IR, Slatyer RO (1980) The use of vital attributes to predict successional changes in plant communities subject to recurrent disturbances. Vegetatio 43, 5–21.
| The use of vital attributes to predict successional changes in plant communities subject to recurrent disturbances.Crossref | GoogleScholarGoogle Scholar |
North GB, Noble PS (1992) Drought-induced changes in hydraulic conductivity and structure in roots of Ferocactus acanthodes and Opuntia ficus-indica. New Phytologist 120, 9–19.
| Drought-induced changes in hydraulic conductivity and structure in roots of Ferocactus acanthodes and Opuntia ficus-indica.Crossref | GoogleScholarGoogle Scholar |
O’Leary MH (1981) Carbon isotope fractionation in plants. Phytochemistry 20, 553–567.
| Carbon isotope fractionation in plants.Crossref | GoogleScholarGoogle Scholar |
Olff H, Vera FWM, Bokdam J, Bakker ES, Gleichman JM, de Maeyer K, Smit R (1999) Shifting mosaics in grazed woodlands driven by the alternation of plant facilitation and competition. Plant Biology 1, 127–137.
| Shifting mosaics in grazed woodlands driven by the alternation of plant facilitation and competition.Crossref | GoogleScholarGoogle Scholar |
Ollinger SV, Richardson AD, Martin ME, Hollinger DY, Frolking SE, Reich PB, Plourde LC, Katul GG, Munger JW, Oren R, Smith M-L, Pau U KT, Bolstad PV, Cook BD, Day MC, Martin TA, Monson RK, Schmid HP (2008) Canopy nitrogen, carbon assimilation, and albedo in temperate and boreal forests: functional relations and potential climate feedbacks. Proceedings of the National Academy of Sciences, USA 105, 19 336–19 341.
| Canopy nitrogen, carbon assimilation, and albedo in temperate and boreal forests: functional relations and potential climate feedbacks.Crossref | GoogleScholarGoogle Scholar |
Onipchenko VG, Makarov MI, Van Logtestijn RSP, Ivanov VB, Akhmetzanova AA, Teekev DK, Ermak AA, Salpargarova FS, Kozhevnikova AD, Cornelissen JHC (2009) New nitrogen uptake strategy: specialized snow roots. Ecology Letters 12, 758–764.
| New nitrogen uptake strategy: specialized snow roots.Crossref | GoogleScholarGoogle Scholar |
Onoda Y, Schieving F, Anten NPR (2008) Effects of light and nutrient availability on leaf mechanical properties of Plantago major: a conceptual approach. Annals of Botany 101, 727–736.
Onoda Y, Westoby M, Adler PB, Choong AMF, Clissold FJ, Cornelissen JHC, Díaz S, Dominy NJ, Elgart A, Enrico L, Fine PVA, Howard JJ, Jalili A, Kitajima K, Kurokawa H, McArthur C, Lucas PW, Markesteijn L, Pérez-Harguindeguy N, Poorter L, Richards L, Santiago LS, Sosinski EE, Van Bael SA, Warton DI, Wright IJ, Joseph Wright S, Yamashita N (2011) Global patterns of leaf mechanical properties. Ecology Letters 14, 301–312.
| Global patterns of leaf mechanical properties.Crossref | GoogleScholarGoogle Scholar |
Ordoñez JC, Van Bodegom PM, Witte J-PM, Bartholomeus RP, Van Hal JR, Aerts R (2010) Plant strategies in relation to resource supply in mesic to wet environments: does theory mirror nature? American Naturalist 175, 225–239.
| Plant strategies in relation to resource supply in mesic to wet environments: does theory mirror nature?Crossref | GoogleScholarGoogle Scholar |
Pakeman RJ, Quested HM (2007) Sampling plant functional traits: what proportion of the species need to be measured? Applied Vegetation Science 10, 91–96.
| Sampling plant functional traits: what proportion of the species need to be measured?Crossref | GoogleScholarGoogle Scholar |
Pammenter NW, Van der Willigen C (1998) A mathematical and statistical analysis of the curves illustrating vulnerability of xylem to cavitation. Tree Physiology 18, 589–593.
| A mathematical and statistical analysis of the curves illustrating vulnerability of xylem to cavitation.Crossref | GoogleScholarGoogle Scholar |
Papió C, Trabaud L (1990) Structural characteristics of fuel components of five Mediterraean shrubs. Forest Ecology and Management 35, 249–259.
| Structural characteristics of fuel components of five Mediterraean shrubs.Crossref | GoogleScholarGoogle Scholar |
Parkhurst DF (1994) Diffusion of CO2 and other gases inside leaves. New Phytologist 126, 449–479.
| Diffusion of CO2 and other gases inside leaves.Crossref | GoogleScholarGoogle Scholar |
Parkhurst DF, Loucks OL (1972) Optimal leaf size in relation to environment. Journal of Ecology 60, 505–537.
| Optimal leaf size in relation to environment.Crossref | GoogleScholarGoogle Scholar |
Parton W, Silver WL, Burke IC, Grassens L, Harmon ME, Currie WS, King JY, Adair EC, Brandt LA, Hart SC, Fasth B (2007) Global-scale similarities in nitrogen release patterns during long-term decomposition. Science 315, 361–364.
| Global-scale similarities in nitrogen release patterns during long-term decomposition.Crossref | GoogleScholarGoogle Scholar |
Patiño S, Lloyd J, Paiva R, Baker TR, Quesada CA, Mercado LM, Schmerler J, Schwarz M, Santos AJB, Aguilar A, Czimczik CI, Gallo J, Horna V, Hoyos EJ, Jimenez EM, Palomino W, Peacock J, Peña-Cruz A, Sarmiento C, Sota A, Turriago JD, Villanueva B, Vitzthum P, Alvarez E, Arroyo L, Baraloto C, Bonal D, Chave J, Costa ACL, Herrera R, Higuchi N, Killeen T, Leal E, Luizão F, Meir P, Monteagudo A, Neill D, Núñez-Vargas P, Peñuela MC, Pitman N, Priante Filho N, Prieto A, Panfil SN, Rudas A, Salomão R, Silva N, Silveira M, Soares de Almeida S, Torres-Lezama A, Vásquez-Martínez R, Vieira I, Malhi Y, Phillips OL (2009) Branch xylem density variations across the Amazon Basin. Biogeosciences 6, 545–568.
| Branch xylem density variations across the Amazon Basin.Crossref | GoogleScholarGoogle Scholar |
Patiño S, Fyllas NM, Baker TR, Paiva R, Quesada CA, Santos AJB, Schwarz M, ter Steege H, Phillips OL, Lloyd J (2012) Coordination of physiological and structural traits in Amazon forest trees. Biogeosciences 9, 775–801.
| Coordination of physiological and structural traits in Amazon forest trees.Crossref | GoogleScholarGoogle Scholar |
Paula S, Pausas JG (2006) Leaf traits and resprouting ability in the Mediterranean basin. Functional Ecology 20, 941–947.
| Leaf traits and resprouting ability in the Mediterranean basin.Crossref | GoogleScholarGoogle Scholar |
Paula S, Pausas JG (2011) Root traits explain different foraging strategies between resprouting life histories. Oecologia 165, 321–331.
| Root traits explain different foraging strategies between resprouting life histories.Crossref | GoogleScholarGoogle Scholar |
Paula S, Arianoutsou M, Kazanis Paula S, Arianoutsou M, Kazanis Paula S, Arianoutsou M, Kazanis (2009) Fire-related traits for plant species of the Mediterranean Basin. Ecology 90, 1420
| Fire-related traits for plant species of the Mediterranean Basin.Crossref | GoogleScholarGoogle Scholar |
Pausas JG (1997) Resprouting of Quercus suber in NE Spain after fire. Journal of Vegetation Science 8, 703–706.
| Resprouting of Quercus suber in NE Spain after fire.Crossref | GoogleScholarGoogle Scholar |
Pausas JG, Bradstock RA (2007) Fire persistence traits of plants along a productivity and disturbance gradient in mediterranean shrublands of south-east Australia. Global Ecology and Biogeography 16, 330–340.
| Fire persistence traits of plants along a productivity and disturbance gradient in mediterranean shrublands of south-east Australia.Crossref | GoogleScholarGoogle Scholar |
Pausas JG, Alessio GA, Moreira B, Corcobado G (2012) Fires enhance flammability in Ulex parviflorus. New Phytologist 193, 18–23.
| Fires enhance flammability in Ulex parviflorus.Crossref | GoogleScholarGoogle Scholar |
Peco B, Traba J, Levassor C, Sánchez AM, Azcárate FM (2003) Seed size, shape and persistence in dry Mediterranean grass and scrublands. Seed Science Research 13, 87–95.
| Seed size, shape and persistence in dry Mediterranean grass and scrublands.Crossref | GoogleScholarGoogle Scholar |
Pérez-Harguindeguy N, Díaz S, Vendramini F, Cornelissen JHC, Gurvich DE, Cabido M (2003) Leaf traits and herbivore selection in the field and in cafeteria experiments. Austral Ecology 28, 642–650.
| Leaf traits and herbivore selection in the field and in cafeteria experiments.Crossref | GoogleScholarGoogle Scholar |
Pickett STA, Kempf JS (1980) Branching patterns in forest shrubs and understory trees in relation to habitat. New Phytologist 86, 219–228.
| Branching patterns in forest shrubs and understory trees in relation to habitat.Crossref | GoogleScholarGoogle Scholar |
Pierce S, Winter K, Griffiths H (2002) Carbon isotope ratio and the extent of daily CAM use by Bromeliaceae. New Phytologist 156, 75–83.
| Carbon isotope ratio and the extent of daily CAM use by Bromeliaceae.Crossref | GoogleScholarGoogle Scholar |
Pisani JM, Distel RA (1998) Inter- and intraspecific variations in production of spines and phenols in Prosopis caldenia and Prosopis flexuosa. Journal of Chemical Ecology 24, 23–36.
| Inter- and intraspecific variations in production of spines and phenols in Prosopis caldenia and Prosopis flexuosa.Crossref | GoogleScholarGoogle Scholar |
Pons J, Pausas JG (2007) Acorn dispersal estimated by radio-tracking Oecologia 153, 903–911.
| Acorn dispersal estimated by radio-trackingCrossref | GoogleScholarGoogle Scholar |
Poorter L, Bongers F (2006) Leaf traits are good predictors of plant performance across 53 rain forest species. Ecology 87, 1733–1743.
| Leaf traits are good predictors of plant performance across 53 rain forest species.Crossref | GoogleScholarGoogle Scholar |
Poorter H, Garnier E (1999) Ecological significance of relative growth rate and its components. In ‘Handbook of functional plant ecology’. (Eds FI Pugnaire, F Valladares) pp. 81–120 (Marcel Dekker: New York)
Poorter H, Garnier E (2007) The ecological significance of variation in relative growth rate and its components. In ‘Functional plant ecology’. (Eds FI Pugnaire, F Valladares) pp. 67–100 (CRC press: Boca Raton, FL)
Poorter H, Lewis C (1986) Testing differences in relative growth rate: a method avoiding curve fitting and pairing. Physiologia Plantarum 67, 223–226.
| Testing differences in relative growth rate: a method avoiding curve fitting and pairing.Crossref | GoogleScholarGoogle Scholar |
Poorter H, Nagel O (2000) The role of biomass allocation in the growth response of plants to different levels of light, CO2, nutrients and water: a quantitative review [published erratum appears in Australian Journal of Plant Physiology 27, 1191]. Australian Journal of Plant Physiology 27, 595–607.
| The role of biomass allocation in the growth response of plants to different levels of light, CO2, nutrients and water: a quantitative review [published erratum appears in Australian Journal of Plant Physiology 27, 1191].Crossref | GoogleScholarGoogle Scholar |
Poorter H, Rozendaal DMA (2008) Leaf size and leaf display of thirty-eight tropical tree species. Oecologia 158, 35–46.
| Leaf size and leaf display of thirty-eight tropical tree species.Crossref | GoogleScholarGoogle Scholar |
Poorter H, Welschen RAM (1993). Analyzing variation in RGR in terms of the underlying carbon economy. In ‘Methods in comparative plant ecology – a laboratory manual’. (Eds GAF Hendry, JP Grime) pp. 107–110 (Chapman and Hall: London)
Poorter L, Bongers L, Bongers F (2006) Architecture of 54 moist-forest tree species: traits, trade-oofs, and functional groups. Ecology 87, 1289–1301.
| Architecture of 54 moist-forest tree species: traits, trade-oofs, and functional groups.Crossref | GoogleScholarGoogle Scholar |
Poorter L, Wright SJ, Paz H, Ackerly DD, Condit R, Ibarra-Manríquez G, Harms KE, Licona JC, Martínez-Ramos M, Mazer SJ, Muller-Landau HC, Peña-Claros M, Webb CO, Wright IJ (2008) Are functional traits good predictors of demographics rates? Evidence from five neotropical forests. Ecology 89, 1908–1920.
| Are functional traits good predictors of demographics rates? Evidence from five neotropical forests.Crossref | GoogleScholarGoogle Scholar |
Poorter H, Niinemets Ü, Poorter L, Wright IJ, Villar R (2009) Causes and consequences of variation in leaf mass per area (LMA): a meta-analysis. New Phytologist 182, 565–588.
| Causes and consequences of variation in leaf mass per area (LMA): a meta-analysis.Crossref | GoogleScholarGoogle Scholar |
Poorter L, Kitajima K, Mercado P, Chubiña J, Melgar I, Prins HHT (2010) Resprouting as a persistence strategy of tropical forest trees: its relation with carbohydrate storage and shade tolerance. Ecology 91, 2613–2627.
| Resprouting as a persistence strategy of tropical forest trees: its relation with carbohydrate storage and shade tolerance.Crossref | GoogleScholarGoogle Scholar |
Poorter H, Niklas KJ, Reich PB, Oleksyn J, Poot P, Mommer L (2012) Tansley review. Biomass allocation to leaves, stems and roots: meta-analyses of interspecific variation and environmental control. New Phytologist 193, 30–50.
| Tansley review. Biomass allocation to leaves, stems and roots: meta-analyses of interspecific variation and environmental control.Crossref | GoogleScholarGoogle Scholar |
Poschlod P, Kiefer S, Tränkle U, Bonn S (1998) Plant species richness in calcareous grasslands as affected by dispersability in space and time. Applied Vegetation Science 1, 75–91.
| Plant species richness in calcareous grasslands as affected by dispersability in space and time.Crossref | GoogleScholarGoogle Scholar |
Poschlod P, Kleyer M, Tackenberg O (2000) Databases on life history traits as a tool for risk assessment in plant species. Zeitschrift für Ökologie und Naturschutz 9, 3–18.
Poschlod P, Tackenberg O, Bonn S (2005) Plant dispersal potential and its relation to species frequency and coexistence. In ‘Vegetation ecology’. (Ed. E Van der Maarel). pp. 147–171 (Blackwell: Oxford, UK)
Pregitzer KS, DeForest JL, Burton AJ, Allen MF, Ruess RW, Hendrick RL (2002) Fine root architecture of nine North American trees. Ecology 72, 293–309.
Preston KA, Ackerly DD (2003) Hydraulic architecture and the evolution of shoot allometry in contrasting climates. American Journal of Botany 90, 1502–1512.
| Hydraulic architecture and the evolution of shoot allometry in contrasting climates.Crossref | GoogleScholarGoogle Scholar |
Price CA, Symonova O, Mileyko Y, Hilley T, Weitz J (2011) Leaf extraction and analysis framework graphical user interface: segmenting and analyzing the structure of leaf veins and areoles. Plant Physiology 155, 236–245.
| Leaf extraction and analysis framework graphical user interface: segmenting and analyzing the structure of leaf veins and areoles.Crossref | GoogleScholarGoogle Scholar |
Puigdefábregas J, Pugnaire FI (1999) Plant survival in arid environments. In ‘Handbook of functional plant ecology’. (Eds FI Pugnaire, F Valladares) pp. 381–405 (Marcel Dekker: New York)
Putz FE, Phyllis DC, Lu K, Montalvo A, Aiello A (1983) Uprooting and snapping of trees: structural determinants and ecological consequences. Canadian Journal of Forest Research 13, 1011–1020.
| Uprooting and snapping of trees: structural determinants and ecological consequences.Crossref | GoogleScholarGoogle Scholar |
Pyankov VI, Gunin PD, Tsoog S, Black CC (2000) C-4 plants in the vegetation of Mongolia: their natural occurrence and geographical distribution in relation to climate. Oecologia 123, 15–31.
| C-4 plants in the vegetation of Mongolia: their natural occurrence and geographical distribution in relation to climate.Crossref | GoogleScholarGoogle Scholar |
Rabotnov TA (1950) The life cycle of perennial herbaceous plants in meadow coenoses. Trudy Botanicheskogo Instituta Akademii Nauk SSSR Seriia III 6, 7–204.
Raunkiaer C (1934) ‘The life forms of plants and statistical plant geography.’ (Clarendon Press: Oxford, UK)
Read J, Stokes A (2006) Plant biomechanics in an ecological context. American Journal of Botany 93, 1546–1565.
| Plant biomechanics in an ecological context.Crossref | GoogleScholarGoogle Scholar |
Rebollo S, Milchunas DG, Noy-Meir I, Chapman PL (2002) The role of a spiny plant refuge in structuring grazed shortgrass steppe plant communities. Oikos 98, 53–64.
| The role of a spiny plant refuge in structuring grazed shortgrass steppe plant communities.Crossref | GoogleScholarGoogle Scholar |
Rees M, Osborne CP, Woodward FI, Hulme SP, Turnbull LA, Taylor SH (2010) Partitioning the components of relative growth rate: how important is plant size variation? American Naturalist 176, E152–E161.
| Partitioning the components of relative growth rate: how important is plant size variation?Crossref | GoogleScholarGoogle Scholar |
Reich PB (2002) Root-shoot relations: optimality in acclimation and adaptation or the ‘Emperor’s new clothes? In ‘Plant roots: the hidden half’. (Eds Y Waisel, A Eshel, U Kafkafi) pp. 314–338 (Marcel Dekker: New York)
Reich PB, Uhl C, Walters MB, Ellsworth DS (1991a) Leaf lifespan as a determinant of leaf structure and function among 23 Amazonian tree species. Oecologia 86, 16–24.
| Leaf lifespan as a determinant of leaf structure and function among 23 Amazonian tree species.Crossref | GoogleScholarGoogle Scholar |
Reich PB, Walters MB, Ellsworth DS (1991b) Leaf age and season influence the relationships between leaf nitrogen, leaf mass per area and photosynthesis in maple and oak trees. Plant, Cell & Environment 14, 251–259.
| Leaf age and season influence the relationships between leaf nitrogen, leaf mass per area and photosynthesis in maple and oak trees.Crossref | GoogleScholarGoogle Scholar |
Reich PB, Walters MB, Ellsworth DS (1992) Leaf life-span in relation to leaf, plant and stand characteristics among diverse ecosystems. Ecological Monographs 62, 365–392.
| Leaf life-span in relation to leaf, plant and stand characteristics among diverse ecosystems.Crossref | GoogleScholarGoogle Scholar |
Reich PB, Walters MB, Ellsworth DS (1997) From tropics to tundra: global convergence in plant functioning. Proceedings of the National Academy of Sciences, USA 94, 13730–13734.
| From tropics to tundra: global convergence in plant functioning.Crossref | GoogleScholarGoogle Scholar |
Reich PB, Tjoelker MG, Walters MB, Vanderklein DW, Bushena C (1998) Close association of RGR, leaf and root morphology, seed mass and shade tolerance in seedlings of nine boreal tree species grown in high and low light. Functional Ecology 12, 327–338.
| Close association of RGR, leaf and root morphology, seed mass and shade tolerance in seedlings of nine boreal tree species grown in high and low light.Crossref | GoogleScholarGoogle Scholar |
Reich PB, Ellsworth DS, Walters MB, Vose JM, Gresham C, Volin JC, Bowman WD (1999) Generality of leaf trait relationships: a test across six biomes. Ecology 80, 1955–1969.
| Generality of leaf trait relationships: a test across six biomes.Crossref | GoogleScholarGoogle Scholar |
Reich PB, Tilman D, Naeem S, Ellsworth DS, Knops J, Craine J, Wedin D, Trost J (2004) Species and functional group diversity independently influence biomass accumulation and its response to CO2 and N. Proceedings of the National Academy of Sciences, USA 101, 10 101–10 106.
| Species and functional group diversity independently influence biomass accumulation and its response to CO2 and N.Crossref | GoogleScholarGoogle Scholar |
Reich PB, Tjoelker MG, Pregitzer KS, Wright IJ, Oleksyn J, Machado JL (2008) Scaling of respiration to nitrogen in leaves, stems and roots of higher land plants. Ecology Letters 11, 793–801.
| Scaling of respiration to nitrogen in leaves, stems and roots of higher land plants.Crossref | GoogleScholarGoogle Scholar |
Reich PB, Oleksyn J, Wright IJ, Niklas KJ, Hedin L, Elser J (2010) Evidence of a general 2/3-power law of scaling leaf nitrogen to phosphorus among major plant groups and biomes. Proceedings. Biological Sciences 277, 877–883.
| Evidence of a general 2/3-power law of scaling leaf nitrogen to phosphorus among major plant groups and biomes.Crossref | GoogleScholarGoogle Scholar |
Reyes G, Brown S, Chapman J, Lugo AE (1992) Wood densities of tropical tree species. General technical report S0-88 US Department of Agriculture, Forest Service, Southern Forest Experiment Station, New Orleans, LA.
Richter M (1992) Methods of interpreting climatological conditions based on phytomorphological characteristics in the cordilleras of the Neotropics. Plant Research and Development 36, 89–114.
Robertson GP, Coleman DC, Bledsoe CS, Sollins P (1999) ‘Standard soil methods for long-term ecological research.’ (Oxford University Press: Oxford, UK)
Rodriguez-Calcerrada J, Atkin OK, Robson TM, Zaragoza-Castells J, Gil L, Aranda I (2010) Thermal acclimation of leaf dark respiration of beech seedlings experiencing summer drought in high and low light environments. Tree Physiology 30, 214–224.
| Thermal acclimation of leaf dark respiration of beech seedlings experiencing summer drought in high and low light environments.Crossref | GoogleScholarGoogle Scholar |
Römermann C, Tackenberg O, Poschlod P (2005a) How to predict attachment potential of seeds to sheep and cattle coat from simple morphological seed traits. Oikos 110, 219–230.
| How to predict attachment potential of seeds to sheep and cattle coat from simple morphological seed traits.Crossref | GoogleScholarGoogle Scholar |
Römermann C, Tackenberg O, Poschlod P (2005b) Buoyancy. In ‘The LEDA traitbase collecting and measuring standards’. (Eds IC Knevel, RM Bekker, D Kunzmann, M Stadler, K Thompson) pp. 124–127 (Groningen University: Groningen, The Netherlands)
Römermann C, Tackenberg O, Poschlod P (2005c) External animal dispersal (epizoochory). In ‘The LEDA traitbase collecting and measuring standards’. (Eds IC Knevel, RM Bekker, D Kunzmann, M Stadler, K Thompson) pp. 127–129 (Groningen University: Groningen, The Netherlands)
Roth-Nebelsick A, Uhl D, Mosbrugger V, Kerp H (2001) Evolution and function of leaf venation architecture: a review. Annals of Botany 87, 553–566.
| Evolution and function of leaf venation architecture: a review.Crossref | GoogleScholarGoogle Scholar |
Rothermel RC (1972) A mathematical model for predicting fire spread in wildland fuels. USDA Forest Service Research Paper INT. (USDA Forest Service: Washington, DC)
Roumet C, Urcelay C, Díaz S (2006) Suites of root traits differ between annual and perennial species growing in the field. New Phytologist 170, 357–368.
| Suites of root traits differ between annual and perennial species growing in the field.Crossref | GoogleScholarGoogle Scholar |
Royer DL, McElwain JC, Adams JM, Wilf P (2008) Sensitivity of leaf size and shape to climate within Acer rubrum and Quercus kelloggii. New Phytologist 179, 808–817.
| Sensitivity of leaf size and shape to climate within Acer rubrum and Quercus kelloggii.Crossref | GoogleScholarGoogle Scholar |
Rozema J, Bijwaard P, Prast G, Broekman R (1985) Ecophysiological adaptations of coastal halophytes from foredunes and salt marshes. Vegetatio 62, 499–521.
| Ecophysiological adaptations of coastal halophytes from foredunes and salt marshes.Crossref | GoogleScholarGoogle Scholar |
Rozema J, Weijers S, Broekman R, Blokker P, Buizer B, Werleman C, Yaqine HE, Hoogedoorn H, Mayoral Fuertes M, Cooper E (2009) Annual growth of Cassiope tetragona as a proxy for Arctic climate: developing correlative and experimental transfer functions to reconstruct past summer temperature on a millennial time scale. Global Change Biology 15, 1703–1715.
| Annual growth of Cassiope tetragona as a proxy for Arctic climate: developing correlative and experimental transfer functions to reconstruct past summer temperature on a millennial time scale.Crossref | GoogleScholarGoogle Scholar |
Rundel PW (1991) Shrub life-forms. In ‘Response of plants to multiple stresses’. (Eds HA Mooney, WE Winner, EJ Pell) pp. 345–370 (Academic Press: San Diego, CA)
Ryser P, Bernardi J, Merla A (2008) Determination of leaf fresh mass after storage between moist paper towels: constraints and reliability of the method. Journal of Experimental Botany 59, 2461–2467.
| Determination of leaf fresh mass after storage between moist paper towels: constraints and reliability of the method.Crossref | GoogleScholarGoogle Scholar |
Sack L, Frole K (2006) Leaf structural diversity is related to hydraulic capacity in tropical rain forest trees. Ecology 87, 483–491.
| Leaf structural diversity is related to hydraulic capacity in tropical rain forest trees.Crossref | GoogleScholarGoogle Scholar |
Sack L, Holbrook NM (2006) Leaf hydraulics. Annual Review of Plant Biology 57, 361–381.
| Leaf hydraulics.Crossref | GoogleScholarGoogle Scholar |
Sack L, Grubb PJ, Marañón T (2003) The functional morphology of juvenile plants tolerant of strong summer drought in shaded forest understories in southern Spain. Plant Ecology 168, 139–163.
| The functional morphology of juvenile plants tolerant of strong summer drought in shaded forest understories in southern Spain.Crossref | GoogleScholarGoogle Scholar |
Sack L, Cornwell WK, Santiago LS, Barbour MM, Choat B, Evans JR, Munns R, Nicotra A (2010) A unique web resource for physiology, ecology and the environmental sciences: PrometheusWiki. Functional Plant Biology 37, 687–693.
| A unique web resource for physiology, ecology and the environmental sciences: PrometheusWiki.Crossref | GoogleScholarGoogle Scholar |
Sage RF (2001) Environmental and evolutionary preconditions for the origin and diversification of the C4 photosynthetic syndrome. Plant Biology 3, 202–213.
| Environmental and evolutionary preconditions for the origin and diversification of the C4 photosynthetic syndrome.Crossref | GoogleScholarGoogle Scholar |
Santiago LS, Goldstein G, Meinzer FC, Fisher JB, Machado K, Woodruff D, Jones T (2004) Leaf photosynthetic traits scale with hydraulic conductivity and wood density in Panamanian forest canopy trees. Oecologia 140, 543–550.
| Leaf photosynthetic traits scale with hydraulic conductivity and wood density in Panamanian forest canopy trees.Crossref | GoogleScholarGoogle Scholar |
Scarff FR, Westoby M (2006) Leaf litter flammability in some semi-arid Australian woodlands. Functional Ecology 20, 745–752.
| Leaf litter flammability in some semi-arid Australian woodlands.Crossref | GoogleScholarGoogle Scholar |
Schenk HJ, Jackson RB (2002) The global biogeography of roots. Ecological Monographs 72, 311–328.
| The global biogeography of roots.Crossref | GoogleScholarGoogle Scholar |
Scholander PF (1966) The role of solvent pressure in osmotic systems. Proceedings of the National Academy of Sciences, USA 55, 1407–1414.
| The role of solvent pressure in osmotic systems.Crossref | GoogleScholarGoogle Scholar |
Schurr FM, Bond WJ, Midgley GF, Higgins SI (2005) A mechanistic model for secondary seed dispersal by wind and its experimental validation. Journal of Ecology 93, 1017–1028.
| A mechanistic model for secondary seed dispersal by wind and its experimental validation.Crossref | GoogleScholarGoogle Scholar |
Schweingruber FH (1996) ‘Tree rings and environment. Dendroecology.’ (Haupt-Verlag: Bern, Switzerland)
Schweingruber FH, Poschlod P (2005) Growth rings in herbs and shrubs: life span, age determination and stem anatomy. Forest Snow and Landscape Research 79, 195–415.
Schwilk DW, Ackerly DD (2001) Flammability and serotiny as strategies: correlated evolution in pines. Oikos 94, 326–336.
| Flammability and serotiny as strategies: correlated evolution in pines.Crossref | GoogleScholarGoogle Scholar |
Seibt U, Rajabi A, Griffiths H, Berry JA (2008) Carbon isotopes and water use efficiency: sense and sensitivity. Oecologia 155, 441–454.
| Carbon isotopes and water use efficiency: sense and sensitivity.Crossref | GoogleScholarGoogle Scholar |
Seiwa K, Kikuzawa K (1996) Importance of seed size for the establishment of seedlings of five deciduous broad-leaved tree species. Vegetatio 123, 51–64.
| Importance of seed size for the establishment of seedlings of five deciduous broad-leaved tree species.Crossref | GoogleScholarGoogle Scholar |
Shipley B (1995) Structured interspecific determinants of specific leaf area in 34 species of herbaceous angiosperms. Functional Ecology 9, 312–319.
| Structured interspecific determinants of specific leaf area in 34 species of herbaceous angiosperms.Crossref | GoogleScholarGoogle Scholar |
Shipley B, Vu TT (2002) Dry matter content as a measure of dry matter concentration in plants and their parts. New Phytologist 153, 359–364.
| Dry matter content as a measure of dry matter concentration in plants and their parts.Crossref | GoogleScholarGoogle Scholar |
Singer MC (2000) Reducing ambiguity in describing plant-insect interactions: ‘preference’, ‘acceptability’ and ‘electivity’. Ecology Letters 3, 159–162.
| Reducing ambiguity in describing plant-insect interactions: ‘preference’, ‘acceptability’ and ‘electivity’.Crossref | GoogleScholarGoogle Scholar |
Skinner FK, Rotenberg Y, Neumann AW (1989) Contact angle measurements from the contact diameter of sessile drops by means of a modified axisymmetric drop shape analysis. Journal of Colloid and Interface Science 130, 25–34.
| Contact angle measurements from the contact diameter of sessile drops by means of a modified axisymmetric drop shape analysis.Crossref | GoogleScholarGoogle Scholar |
Smith WK, Bell DT, Shepherd KA (1998) Associations between leaf structure, orientation, and sunlight exposure in five Western Australian communities. American Journal of Botany 85, 56–63.
| Associations between leaf structure, orientation, and sunlight exposure in five Western Australian communities.Crossref | GoogleScholarGoogle Scholar |
Southwood TRE (1977) Habitat, the templet for ecological strategies? Journal of Animal Ecology 43, 337–365.
Southwood TRE, Brown VK, Reader PM (1986) Leaf palatability, life expectancy and herbivore damage. Oecologia 70, 544–548.
| Leaf palatability, life expectancy and herbivore damage.Crossref | GoogleScholarGoogle Scholar |
Sperry JS (2003) Evolution of water transport and xylem structure. International Journal of Plant Sciences 164, S115–S127.
| Evolution of water transport and xylem structure.Crossref | GoogleScholarGoogle Scholar |
Sperry JS, Donnelly JR, Tyree MT (1988) A method for measuring hydraulic conductivity and embolism in xylem. Plant, Cell & Environment 11, 35–40.
| A method for measuring hydraulic conductivity and embolism in xylem.Crossref | GoogleScholarGoogle Scholar |
Sperry JS, Meinzer FC, McCulloh KA (2008a) Safety and efficiency conflicts in hydraulic architecture: scaling from tissues to trees. Plant, Cell & Environment 31, 632–645.
| Safety and efficiency conflicts in hydraulic architecture: scaling from tissues to trees.Crossref | GoogleScholarGoogle Scholar |
Sperry JS, Taneda H, Bush SE, Hacke UG (2008b) Evaluation of centrifugal methods for measuring xylem cavitation in conifers, diffuse and ring-porous angiosperms. New Phytologist 177, 558–568.
Srivastava DS, Cadotte MW, MacDonald AAM, Marushia RG, Mirotchnick N (2012) Phylogenetic diversity and the functioning of ecosystems. Ecology Letters 15, 637–648.
| Phylogenetic diversity and the functioning of ecosystems.Crossref | GoogleScholarGoogle Scholar |
Staver AC, Bond WJ, February EC (2011) History matters: tree establishment variability and species turnover in an African savanna. Ecosphere 2, art49
| History matters: tree establishment variability and species turnover in an African savanna.Crossref | GoogleScholarGoogle Scholar |
Stephens SL, Gordon DA, Martin RE (1994) Combustibility of selected domestic vegetation subjected to desiccation. In ‘12th conference on fire and forest meteorology’, Jekyll Island, GA. Available from: http://www.cnr.berkeley.edu/stephens-lab/Publications/Stephensetal_93_%20Comb_Veg.pdf [Verified 22 February 2013].
Steudle E (2001) Water uptake by plant roots: an integration of views. In ‘Recent advances of plant root structure and function’. (Eds O Gašparíková, M Ciamporová, I Mistrík, F Baluška) pp. 71–82 (Kluwer Academic Publishers: Dordrecht, The Netherlands)
Stich B, Piepho H-P, Schulz B, Melchinger AE (2008) Multi-trait association mapping in sugar beet (Beta vulgaris L.). Theoretical and Applied Genetics 117, 947–954.
| Multi-trait association mapping in sugar beet (Beta vulgaris L.).Crossref | GoogleScholarGoogle Scholar |
Strauss SY, Agrawal AA (1999) The ecology and evolution of plant tolerance to herbivory. Trends in Ecology & Evolution 14, 179–185.
| The ecology and evolution of plant tolerance to herbivory.Crossref | GoogleScholarGoogle Scholar |
Swenson NG, Enquist BJ (2008) The relationship between stem and branch specific gravity and the ability of each measure to predict leaf area. American Journal of Botany 95, 516–519.
| The relationship between stem and branch specific gravity and the ability of each measure to predict leaf area.Crossref | GoogleScholarGoogle Scholar |
Tackenberg O (2003) Modeling long distance dispersal of plant diaspores by wind. Ecological Monographs 73, 173–189.
| Modeling long distance dispersal of plant diaspores by wind.Crossref | GoogleScholarGoogle Scholar |
Tackenberg O, Poschlod P, Bonn S (2003) Assessment of wind dispersal potential in plant species. Ecological Monographs 73, 191–205.
| Assessment of wind dispersal potential in plant species.Crossref | GoogleScholarGoogle Scholar |
Taiz L, Zeiger E (2010) ‘Plant physiology.’ (The Benjamin/Cummings Publishing: Redwood City, CA)
Tamm CO (1972) Survival and flowering of some perennial herbs. II. The behaviour of some orchids on permanent plots. Oikos 23, 23–28.
| Survival and flowering of some perennial herbs. II. The behaviour of some orchids on permanent plots.Crossref | GoogleScholarGoogle Scholar |
Taylor B, Parkinson D (1988) A new microcosm approach to litter decomposition studies. Canadian Journal of Botany 66, 1933–1939.
Temminghoff EEJM, Houba VJG (2004) ‘Plant analyses procedures.’ (Kluwer Academic Publishers: Dodrecht, The Netherlands).
Tennant D (1975) A test of a modified line intersect method of estimating root length. Journal of Ecology 63, 995–1001.
| A test of a modified line intersect method of estimating root length.Crossref | GoogleScholarGoogle Scholar |
Thomas SC (1996) Asymptotic height as a predictor of growth and allometric characteristics in Malaysian rain forest trees. American Journal of Botany 83, 556–566.
| Asymptotic height as a predictor of growth and allometric characteristics in Malaysian rain forest trees.Crossref | GoogleScholarGoogle Scholar |
Thompson K, Band SR, Hodgson JG (1993) Seed size and shape predict seed persistence in the soil. Functional Ecology 7, 236–241.
| Seed size and shape predict seed persistence in the soil.Crossref | GoogleScholarGoogle Scholar |
Thompson K, Bakker JP, Bekker RM (1997) ‘The soil seed bank of North West Europe: methodology, density and longevity.’ (Cambridge University Press: Cambridge, UK)
Tjoelker MG, Oleksyn J, Reich PB (2001) Modelling respiration of vegetation: evidence for a general temperature-dependent Q10 Global Change Biology 7, 223–230.
| Modelling respiration of vegetation: evidence for a general temperature-dependent Q10Crossref | GoogleScholarGoogle Scholar |
Turner NC (1988) Measurement of plant water status by the pressure chamber technique. Irrigation Science 9, 289–308.
| Measurement of plant water status by the pressure chamber technique.Crossref | GoogleScholarGoogle Scholar |
Turner IM (1994) Sclerophylly: primarily protective? Functional Ecology 8, 669–675.
| Sclerophylly: primarily protective?Crossref | GoogleScholarGoogle Scholar |
Tyree MT, Sperry JS (1989) Vulnerability of xylem to cavitation and embolism. Annual Review of Plant Physiology and Plant Molecular Biology 40, 19–36.
| Vulnerability of xylem to cavitation and embolism.Crossref | GoogleScholarGoogle Scholar |
Tyree MT, Zimmermann MH (2002) ‘Xylem structure and the ascent of sap.’ (Springer-Verlag: Berlin)
Uhl D, Mosbrugger V (1999) Leaf venation density as a climate and environmental proxy: a critical review and new data. Palaeogeography, Palaeoclimatology, Palaeoecology 149, 15–26.
| Leaf venation density as a climate and environmental proxy: a critical review and new data.Crossref | GoogleScholarGoogle Scholar |
Vaieretti MV, Díaz S, Vile D, Garnier E (2007) Two measurement methods of leaf dry matter content produce similar results in a broad range of species. Annals of Botany 99, 955–958.
| Two measurement methods of leaf dry matter content produce similar results in a broad range of species.Crossref | GoogleScholarGoogle Scholar |
Valette J-C (1997) Inflammabilities of Mediterranean species. In ‘Forest fire risk and management. EUR 16719 EN’. (Ed. P Balabanis, F Eftichidis, R Fantechi) pp. 51–64 (European Commission, Environment and Quality of Life: Brussels)
Van Altena C, Van Logtestijn RSP, Cornwell WK, Cornelissen JHC (2012) Species composition and fire: non-additive mixture effects on ground fuel flammability. Frontiers in Plant Science 3, 63
| Species composition and fire: non-additive mixture effects on ground fuel flammability.Crossref | GoogleScholarGoogle Scholar |
Van der Pijl L (1982) ‘Principles of dispersal in higher plants.’ (Springer-Verlag: Berlin)
Van Gelder HA, Poorter L, Sterck FJ (2006) Wood mechanics, allometry, and life-history variation in a tropical rain forest tree community. New Phytologist 171, 367–378.
| Wood mechanics, allometry, and life-history variation in a tropical rain forest tree community.Crossref | GoogleScholarGoogle Scholar |
Van Groenendael JM, Klimeš L, Klimešova J, Hendriks RJJ (1997) Comparative ecology of clonal plants. In ‘Plant life histories’. (Eds JL Harper, J Silvertown, M Franco) pp. 191–209 (Cambridge University Press: Cambridge, UK)
Vendramini F, Díaz S, Gurvich DE, Wilson PJ, Thompson K, Hodgson JG (2002) Leaf traits as indicators of resource-use strategy in floras with succulent species. New Phytologist 154, 147–157.
| Leaf traits as indicators of resource-use strategy in floras with succulent species.Crossref | GoogleScholarGoogle Scholar |
Veneklaas EJ, Poorter L (1998) Growth and carbon partitioning of tropical tree seedlings in contrasting light environments. In ‘Inherent variation in plant growth’. (Eds H Lambers, H Poorter, MMI Van Vuuren) pp. 337–361 (Backhuys Publishers: Leiden, The Netherlands)
Veneklaas EJ, Zagt RJ, Van Leerdam A, Van Ek R, Broekhoven AJ, Van Genderen M (1990) Hydrological properties of the epiphyte mass of a montane tropical rain forest, Colombia. Vegetatio 89, 183–192.
| Hydrological properties of the epiphyte mass of a montane tropical rain forest, Colombia.Crossref | GoogleScholarGoogle Scholar |
Vesk PA, Westoby M (2004) Sprouting ability across diverse disturbances and vegetation types worldwide. Journal of Ecology 92, 310–320.
| Sprouting ability across diverse disturbances and vegetation types worldwide.Crossref | GoogleScholarGoogle Scholar |
Vile D, Garnier E, Shipley B, Laurent G, Navas M-L, Roumet C, Lavorel S, Díaz S, Hodgson JG, Lloret F, Midgley GF, Poorter H, Rutherford MC, Wilson PJ, Wright IJ (2005) Specific leaf area and dry matter content estimate thickness in laminar leaves. Annals of Botany 96, 1129–1136.
| Specific leaf area and dry matter content estimate thickness in laminar leaves.Crossref | GoogleScholarGoogle Scholar |
Violle C, Navas M-L, Vile D, Kazakou E, Fortunel C, Hummel I, Garnier E (2007) Let the concept of trait be functional! Oikos 116, 882–892.
| Let the concept of trait be functional!Crossref | GoogleScholarGoogle Scholar |
Wahl S, Ryser P (2000) Root tissue structure is linked to ecological strategies of grasses. New Phytologist 148, 459–471.
| Root tissue structure is linked to ecological strategies of grasses.Crossref | GoogleScholarGoogle Scholar |
Walters MB, Reich PB (1999) Low-light carbon balance and shade tolerance in the seedlings of woody plants: do winter deciduous and broad-leaved evergreen species differ? New Phytologist 143, 143–154.
| Low-light carbon balance and shade tolerance in the seedlings of woody plants: do winter deciduous and broad-leaved evergreen species differ?Crossref | GoogleScholarGoogle Scholar |
Wand SJE, Midgley GG, Jones MH, Curtis PS (1999) Responses of wild C4 and C3 grasses (Poaceae) species to elevated atmospheric CO2 concentration: a meta-analytical test of current theories and perceptions. Global Change Biology 5, 723–741.
| Responses of wild C4 and C3 grasses (Poaceae) species to elevated atmospheric CO2 concentration: a meta-analytical test of current theories and perceptions.Crossref | GoogleScholarGoogle Scholar |
Weiher E, Clarke GDP, Keddy PA (1998) Community assembly rules, morphological dispersion, and the coexistence of plant species. Oikos 81, 309–322.
| Community assembly rules, morphological dispersion, and the coexistence of plant species.Crossref | GoogleScholarGoogle Scholar |
Weiher E, Van der Werf A, Thompson K, Roderick M, Garnier E, Eriksson O (1999) Challenging Theophrastus: a common core list of plant traits for functional ecology. Journal of Vegetation Science 10, 609–620.
| Challenging Theophrastus: a common core list of plant traits for functional ecology.Crossref | GoogleScholarGoogle Scholar |
Westoby M (1998) A leaf-height-seed (LHS) plant ecology strategy scheme. Plant and Soil 199, 213–227.
| A leaf-height-seed (LHS) plant ecology strategy scheme.Crossref | GoogleScholarGoogle Scholar |
Westoby M, Cunningham SA, Fonseca C, Overton J, Wright IJ (1998) Phylogeny and variation in light capture area deployed per unit investment in leaves: designs for selecting study species with a view to generalizing. In ‘Variation in growth rate and productivity of higher plants’. (Eds H Lambers, H Poorter H, MMI Van Vuuren) pp. 539–566 (Backhuys Publishers: Leiden, The Netherlands)
Westoby M, Warton D, Reich PB (2000) The time value of leaf area. American Naturalist 155, 649–656.
| The time value of leaf area.Crossref | GoogleScholarGoogle Scholar |
Westoby M, Falster D, Moles A, Vesk P, Wright I (2002) Plant ecological strategies: some leading dimensions of variation between species. Annual Review of Ecology and Systematics 33, 125–159.
| Plant ecological strategies: some leading dimensions of variation between species.Crossref | GoogleScholarGoogle Scholar |
Whittaker RH (1975) ‘Communities and ecosystems.’ 2nd edn. (Macmillan Publishing: New York)
Will H, Tackenberg O (2008) A mechanistic simulation model of seed dispersal by animals. Journal of Ecology 96, 1011–1022.
| A mechanistic simulation model of seed dispersal by animals.Crossref | GoogleScholarGoogle Scholar |
Williamson GB, Wiemann MC (2010) Measuring wood specific gravity…Correctly. American Journal of Botany 97, 519–524.
| Measuring wood specific gravity…Correctly.Crossref | GoogleScholarGoogle Scholar |
Wilson PJ, Thompson K, Hodgson JG (1999) Specific leaf area and leaf dry matter content as alternative predictors of plant strategies. New Phytologist 143, 155–162.
| Specific leaf area and leaf dry matter content as alternative predictors of plant strategies.Crossref | GoogleScholarGoogle Scholar |
Withington JM, Reich B, Oleksyn J, Eissenstat DM (2006) Comparisons of structure and life span in roots and leaves among temperate trees. Ecological Monographs 76, 381–397.
| Comparisons of structure and life span in roots and leaves among temperate trees.Crossref | GoogleScholarGoogle Scholar |
Witkowski ETF, Lamont BB (1991) Leaf specific mass confounds leaf density and thickness. Oecologia 88, 486–493.
Wong S-C, Cowan IR, Farquhar GD (1979) Stomatal conductance correlates with photosynthetic capacity. Nature 282, 424–426.
| Stomatal conductance correlates with photosynthetic capacity.Crossref | GoogleScholarGoogle Scholar |
Wong S-C, Cowan IR, Farquhar GD (1985a) Leaf conductance in relation to rate of CO2 assimilation. I. Influence of nitrogen nutrition, phosphorus nutrition, photon flux density, and ambient partial pressure of CO2 during ontogeny. Plant Physiology 78, 821–825.
| Leaf conductance in relation to rate of CO2 assimilation. I. Influence of nitrogen nutrition, phosphorus nutrition, photon flux density, and ambient partial pressure of CO2 during ontogeny.Crossref | GoogleScholarGoogle Scholar |
Wong S-C, Cowan IR, Farquhar GD (1985b) Leaf conductance in relation to rate of CO2 assimilation. II. Effects of short-term exposures to different photon flux densities. Plant Physiology 78, 826–829.
| Leaf conductance in relation to rate of CO2 assimilation. II. Effects of short-term exposures to different photon flux densities.Crossref | GoogleScholarGoogle Scholar |
Wong S-C, Cowan IR, Farquhar GD (1985c) Leaf conductance in relation to rate of CO2 assimilation. III. Influences of water stress and photoinhibition. Plant Physiology 78, 830–834.
| Leaf conductance in relation to rate of CO2 assimilation. III. Influences of water stress and photoinhibition.Crossref | GoogleScholarGoogle Scholar |
Wright IJ, Cannon K (2001) Relationships between leaf lifespan and structural defences in a low-nutrient, sclerophyll flora. Functional Ecology 15, 351–359.
| Relationships between leaf lifespan and structural defences in a low-nutrient, sclerophyll flora.Crossref | GoogleScholarGoogle Scholar |
Wright W, Illius AW (1995) A comparative study of the fracture properties of 5 grasses. Functional Ecology 9, 269–278.
| A comparative study of the fracture properties of 5 grasses.Crossref | GoogleScholarGoogle Scholar |
Wright W, Vincent JFK (1996) Herbivory and the mechanics of fracture in plants. Biological Reviews of the Cambridge Philosophical Society 71, 401–413.
| Herbivory and the mechanics of fracture in plants.Crossref | GoogleScholarGoogle Scholar |
Wright IJ, Westoby M (2002) Leaves at low versus high rainfall: coordination of structure, lifespan and physiology. New Phytologist 155, 403–416.
| Leaves at low versus high rainfall: coordination of structure, lifespan and physiology.Crossref | GoogleScholarGoogle Scholar |
Wright IJ, Clifford HT, Kidson R, Reed ML, Rice BL, Westoby M (2000) A survey of seed and seedling characters in 1744 Australian dicotyledon species: cross-species trait correlations and correlated trait-shifts within evolutionary lineages. Biological Journal of the Linnean Society. Linnean Society of London 69, 521–547.
| A survey of seed and seedling characters in 1744 Australian dicotyledon species: cross-species trait correlations and correlated trait-shifts within evolutionary lineages.Crossref | GoogleScholarGoogle Scholar |
Wright IJ, Westoby M, Reich PB (2002) Convergence towards higher leaf mass per area in dry and nutrient-poor habitats has different consequences for leaf life span. Journal of Ecology 90, 534–543.
| Convergence towards higher leaf mass per area in dry and nutrient-poor habitats has different consequences for leaf life span.Crossref | GoogleScholarGoogle Scholar |
Wright IJ, Reich PB, Westoby M, Ackerly DD, Baruch Z, Bongers F, Cavender-Bares J, Chapin T, Cornelissen JHC, Diemer M, Flexas J, Garnier E, Groom PK, Gulias J, Hikosaka K, Lamont BB, Lee T, Lee W, Lusk C, Midgley JJ, Navas ML, Niinemets U, Oleksyn J, Osada N, Poorter H, Poot P, Prior L, Pyankov VI, Roumet C, Thomas SC, Tjoelker MG, Veneklaas EJ, Villar R (2004) The worldwide leaf economics spectrum. Nature 428, 821–827.
| The worldwide leaf economics spectrum.Crossref | GoogleScholarGoogle Scholar |
Wright IJ, Falster DS, Pickup M, Westoby M (2006) Cross-species patterns in the coordination between leaf and stem traits, and their implications for plant hydraulics. Physiologia Plantarum 127, 445–456.
| Cross-species patterns in the coordination between leaf and stem traits, and their implications for plant hydraulics.Crossref | GoogleScholarGoogle Scholar |
Wright IJ, Ackerly D, Bongers F, Harms KE, Ibarra-Manríquez G, Martínez-Ramos M, Mazer SJ, Muller-Landau HC, Paz H, Pitman NCA, Poorter L, Silman MR, Vriesendorp CF, Webb CO, Westoby M, Wright SJ (2007) Relationships among ecologically important dimensions of plant trait variation in seven neotropical forests. Annals of Botany 99, 1003–1015.
| Relationships among ecologically important dimensions of plant trait variation in seven neotropical forests.Crossref | GoogleScholarGoogle Scholar |
Yeo AR (1983) Salinity resistance: physiologies and prices. Physiologia Plantarum 58, 214–222.
| Salinity resistance: physiologies and prices.Crossref | GoogleScholarGoogle Scholar |
Zanne AE, Chapman CA, Kitajima K (2005) Evolutionary and ecological correlates of early seedling morphology in east African trees and shrubs. American Journal of Botany 92, 972–978.
| Evolutionary and ecological correlates of early seedling morphology in east African trees and shrubs.Crossref | GoogleScholarGoogle Scholar |
Zanne AE, Westoby M, Falster DS, Ackerly DD, Loarie SR, Arnold SEJ, Coomes DA (2010) Angiosperm wood structure: global patterns in vessel anatomy and their relation to wood density and potential conductivity. American Journal of Botany 97, 207–215.
| Angiosperm wood structure: global patterns in vessel anatomy and their relation to wood density and potential conductivity.Crossref | GoogleScholarGoogle Scholar |
Zhu JK (2001) Plant salt tolerance. Trends in Plant Science 6, 66–71.
| Plant salt tolerance.Crossref | GoogleScholarGoogle Scholar |
Zimmermann MH, Jeje AA (1981) Vessel-length distribution in stems of some American woody plants. Canadian Journal of Botany 59, 1882–1892.
| Vessel-length distribution in stems of some American woody plants.Crossref | GoogleScholarGoogle Scholar |
Zinke PJ (1962) The pattern of influence of individual forest trees on soil properties. Ecology 43, 130–133.
| The pattern of influence of individual forest trees on soil properties.Crossref | GoogleScholarGoogle Scholar |
Zotz G, Ziegler H (1997) The occurrence of crassulacean metabolism among vascular epiphytes from central Panama. New Phytologist 137, 223–229.
| The occurrence of crassulacean metabolism among vascular epiphytes from central Panama.Crossref | GoogleScholarGoogle Scholar |
Zwieniecki MA, Melcher PJ, Holbrook NM (2001) Hydrogel control of xylem hydraulic resistance in plants. Science 291, 1059–1062.
| Hydrogel control of xylem hydraulic resistance in plants.Crossref | GoogleScholarGoogle Scholar |
Zwieniecki MA, Melcher PJ, Boyce CK, Sack L, Holbrook NM (2002) Hydraulic architecture of leaf venation in Laurus nobilis L. Plant, Cell & Environment 25, 1445–1450.
| Hydraulic architecture of leaf venation in Laurus nobilis L.Crossref | GoogleScholarGoogle Scholar |
Appendix 1. Summary of plant traits

|