

Wild seedlings of a tree endemic on granite outcrops show no evidence of inbreeding depression
Nicole BezemerCentre of Excellence in Natural Resource Management, The University of Western Australia, 35 Stirling Terrace, Albany, WA 6330, Australia and Botanic Gardens and Parks Authority, Fraser Avenue, Kings Park and Botanic Garden, West Perth, Western Australia 6005. Email: nicole.bezemer@research.uwa.edu.au
Australian Journal of Botany 66(1) 39-47 https://doi.org/10.1071/BT17175
Submitted: 21 September 2017 Accepted: 25 November 2017 Published: 4 January 2018
Abstract
Plant communities on granite outcrops are characterised by high biodiversity and endemism, and strong population genetic structuring. The floristic diversity and structure of inselberg flora receives much attention in international conservation research, yet our understanding of how recruitment influences population genetic structure of plants endemic to granite outcrops is limited. To determine the immediate genetic consequences of recruitment and to quantify mating and dispersal patterns by parentage analysis, I conducted an opportunistic study following a wildfire in a stand of the lignotuberous tree Eucalyptus caesia Benth. (Myrtaceae) at Boyagin Nature Reserve. The entire adult stand (n = 180) plus seedlings (n = 115) were genotyped with 14 microsatellite loci. Compared with adults, seedlings had low heterozygosity and a high inbreeding co-efficient, but similar levels of allelic diversity. There was no evidence of post-germination selection against homozygous seedlings over the 20 month survey period. Seedlings that resulted from cross-pollination were more heterozygous than seedlings resulting from selfing, but could not be distinguished based on height measurements or survivorship. Parentage analysis in CERVUS revealed mostly limited seed dispersal, assuming that candidate parents closest to seedlings were the maternal parent. By comparison, pollen movement was more extensive, yet still restricted to plants within the stand. Recruitment has increased the population size, but appears insufficient to enhance genetic diversity in the Boyagin stand of E. caesia. Nonetheless, E. caesia appears adept at persisting as extremely small populations.
Additional keywords: genetic diversity, granite-outcrop endemism, parentage analysis, recruitment, seed and pollen dispersal.
Introduction
Inselbergs are isolated rocky outcrops or mountains, often of granitic or gneissic origin, that support biota distinct from that of surrounding forest or grassland landscapes (Porembski and Barthlott 2012). The floristic diversity, high endemism and uniqueness of vegetation communities of inselberg habitats have been a focal point of international conservation research (Groger and Barthlott 1996; Porembski et al. 1996; 1997). Around the globe, inselberg habitats are severely threatened by quarrying, recreational destruction, urbanisation, livestock, and invasion of exotic plants (Porembski et al. 2016; Fitzsimons and Michael 2017).
Strong genetic differentiation amongst populations is widely reported for plants endemic to granite outcrops (Wyatt et al. 1992; Barbará et al. 2007; Byrne and Hopper 2008; Tapper et al. 2014; Hmeljevski et al. 2017), although some populations do maintain connectivity (Gevaert et al. 2013; Gonçalves-Oliveira et al. 2017). Much less is known about the immediate effects of recruitment on the population genetics of granite-endemic flora, perhaps due to the sporadic and sometimes inefficacious nature of recruitment (Vlasáková 2013; Hmeljevski et al. 2014). Harsh environmental conditions that characterise rocky habitats, such as high radiation, extreme temperatures, high runoff and low soil moisture may impede recruitment or cause high mortality of new recruits (Szarzynski 2000). Additional constraints such as lack of rock crevices suitable for seedling establishment may constrain population growth (Yates and Ladd 2004). Combining demographic studies with recruitment genetics is critical to develop appropriate genetic management strategies for small, fragmented populations.
In small plant populations, highly outcrossed mating systems and widespread pollen dispersal may ensure high levels of heterozygosity in canopy or soil-stored seed crops as they develop over time (Krauss et al. 2009; Ayre et al. 2010; Breed et al. 2015b). Pulse recruitment from genetically diverse seed crops thus may buffer genetic decline in isolated populations (Ayre et al. 2009). In the absence of cross-pollination, selection against homozygous progeny may enhance genetic diversity. Post-dispersal selection against homozygotes has been demonstrated in long-term field trials of selfed and outcrossed seedlings, and genetic surveys of recruits in wild populations (Hardner and Potts 1997; Roberts et al. 2014). It is unclear whether harsh environmental conditions that exert strong selection pressure will lead to rapid mortality of inbred progeny that is observable over short time periods.
Pollination systems involving highly mobile animals, in particular nectar-feeding birds, are theorised to maintain genetic connectivity between fragmented populations (Byrne et al. 2007; Breed et al. 2015a; Krauss et al. 2017). However, low population densities, insularity and human disturbance may lead to reduced levels of pollen migration and restricted mating patterns, even in plants with bird pollinators (Llorens et al. 2012; Coates et al. 2013). Most studies that document mating patterns do so by sampling seeds held in canopy- or soil-stored seedbanks – few have focused on seedling recruits that form the next generation (Barrett et al. 2005; Jones et al. 2010; Roberts et al. 2014). Our understanding of genetic structure in the typically small proportion of seeds that germinate and survive to contribute to the standing population is therefore limited.
In south-west Australia, granite outcrops support hyper-diverse plant communities and provide a refuge for fire-sensitive biota in an otherwise seasonally dry and fire-prone landscape (Hopper et al. 1990, 1997). A large proportion of plants on granite outcrops are obligate seeders that may be killed by fire (Yates et al. 2003a) whereas other plants require disturbance, such as fire, to trigger recruitment (Burrows 2013). The present study opportunistically investigates recruitment genetics in Eucalyptus caesia Benth. (Myrtaceae), an endemic of granite outcrops that is known from 27 small, disjunct populations in the Wheatbelt Region of Western Australia (Bezemer et al. 2016) including two populations recently documented by N. Bezemer and S. D. Hopper. Recruitment from seed in E. caesia is rare, associated with the occurrence of fire, and seldom recorded in the wild – as observed in other mallee eucalypts (Wellington and Noble 1985a; Davies and Myerscough 1991; Yates et al. 2003b). Seed dispersal in eucalypts occurs predominantly by gravity and is considered to be highly restricted (Potts and Wiltshire 1997), yet studies that provide direct genetic confirmation of this assumption are lacking (Booth 2017). Genetic connectivity is usually maintained via long distance pollen movement in gravity-dispersed species. In several eucalypts, bird pollination has been linked to increased gene flow (Southerton et al. 2004; Hopper 2009; Breed et al. 2015b). However, in a population of the predominantly bird-pollinated E. caesia, mating appears spatially restricted, despite high levels of multiple paternity (Bezemer et al. 2016).
I used a field survey of adult plants and seedling growth and survivorship over 20 months, in combination with microsatellite genotyping and parentage analysis, to test two hypotheses: (i) that seedlings will predominantly result from mating between plants within the study population; and (ii) that levels of genetic diversity in 20 month old seedlings will reflect that of adults despite high levels of inbreeding, because of selection against inbred seedlings.
Materials and methods
Study area and species
The study was conducted on a natural stand of Eucalyptus caesia Benth. at Boyagin Nature Reserve in the Central Wheatbelt region of Western Australia (32°28ʹ21.0ʹʹS 116°52ʹ23.5ʹʹE). E. caesia is monoecious, with a mean realised outcrossing rate of 0.72 reported for the Boyagin stand (Bezemer et al. 2016). Populations of E. caesia are characterised by low genetic diversity and high levels of genetic differentiation (Moran and Hopper 1983; Byrne and Hopper 2008). In the winter months, large pink flowers produce copious amounts of nectar that appear to favour pollination by nectar-feeding birds such as honeyeaters, although flowers are also visited by insects – including the European honeybee (Hopper 1981). Although recruitment has been rarely observed in E. caesia, asexual reproduction via lignotubers is common. Individual plants appear to have a capacity for extreme longevity based on the size of lignotubers, inferred population age structure, and presence of large clonal patches (Hopper et al. 1982; Bezemer et al. 2016).
Field data collection
Commencing 5 months after a wildfire, I conducted on-ground searches for E. caesia seedlings over seven intermittent surveys (1 October 2015, 28 October 2015, 6 December 2015, 14 January 2016, 17 March 2016, 25 August 2016 and 6 April 2017). The aim was to locate as many seedlings as possible, rather than conduct a transect-style survey. A total of 70 h was spent searching near all adult plants. When no seedlings were found in unburnt areas after the first two surveys, searches were concentrated in burnt areas. The global positioning system (GPS) co-ordinates (UTM easting and northing) of all seedlings were recorded with a Trimble GeoExplorer differential GPS (Digital Mapping Solutions, Perth, WA, Australia), and for seedlings with more than two expanded leaves, a sample collected for genetic analysis. This precautionary strategy was employed to reduce the risk of killing very small seedlings by sampling. Seedling height was measured as the length (cm) between the base of the stem and terminal node, and repeat measurements taken at subsequent surveys.
Microsatellite genotyping
DNA was extracted from seedling leaf samples following a CTAB method modified from Doyle and Doyle (1987). Fourteen microsatellite loci were amplified with a QIAGEN Multiplex PCR Kit (Qiagen, Venio, Limburg, Netherlands), following the manufacturer’s instructions. Loci included EGM30, EGM47, EGM14, EGM12, EGM34, EGM35, EMBRA10, EMBRA7, EMBRA20, EMBRA18, EMCRC11, EV22, EV23, EV28 and ES140 (Brondani et al. 1998; Glaubitz et al. 2001; Steane et al. 2001; Bradbury et al. 2013; Nevill et al. 2013). Multiplex mixes and PCR conditions follow Bezemer et al. (2016), except for modified annealing temperatures of 53°C for EMBRA18, EMCRC11 and EV22, and 65°C for EV23, EV28 and ES140. To optimise amplification peaks, the concentrations of several primer pairs were increased or reduced from a 2 µm concentration.
Amplified loci were visualised with an automated 3500 Genetic Analyser with 0.2 µl 500 LIZ size standard/10 µl Hi-Di Formamide (Applied Biosystems, Thermo Fischer Scientific, Melbourne, Vic., Australia). Alleles were scored manually with Geneious 7.1.4 (Drummond et al. 2011). Positive and negative controls were included in each 96 well plate and electropherogram profiles double-checked for all samples. For some primers, changing primer manufacturer from Applied Biosystems to Alpha DNA (Montreal, Canada) resulted in a consistent shift in allele size; for example alleles were always four base pairs larger for EMBRA10 primers manufactured by AB than Alpha DNA. To confirm Mendelian inheritance (i.e. all progeny should have at least one maternally-inherited allele), all primers were screened with different trees and a sample of their progeny.
Genetic diversity and structure
Genetic diversity parameters including Na (number of alleles), number of alleles with a frequency of ≥ 5% (Na freq. ≥ 5%), Ne (effective number of alleles), HE (expected heterozygosity), HO (observed heterozygosity), and F (fixation index, also called the inbreeding co-efficient) were calculated in GenAlEx ver. 6.5 (Peakall and Smouse 2012) for adults and seedlings, and for seedling subsets that were present at repeat surveys. To test for genetic differentiation of adults and seedlings, genetic distances between all multi-locus genotypes were calculated and a principle co-ordinates analysis (PCoA) performed, and genetic differentiation measures including GST, GʹST N, GʹST H, GʹST, DEST and P (GST) calculated. Fewer seedlings were included in the genetic analysis than reported in the on-ground survey results because some seedlings were too small to sample and died before the next survey.
Parentage assignment
Parentage assignment was conducted by categorical allocation in CERVUS ver.3.0.3 with strict confidence levels set at 95% and relaxed confidence levels of 80%, and critical Logarithm of Odds (LOD) scores were used to determine confidence of assignments (Kalinowski et al. 2007). Two analyses were performed – first, with all reproductive adults as candidates and second, with duplicate genotypes of all candidates (to allow detection of selfing). Candidate parents were grouped according to Euclidean distance from seedlings. For each parent-pair, the individual nearest/furthest to the seedling was assumed the maternal parent/pollen donor respectively (Guidugli et al. 2016). I used these results to infer realised pollen and seed dispersal distances.
To test for differences in growth rate or survivorship of seedlings resulting from self- versus cross-pollination, the minimum number of days alive, growth (taken as height at the last measurement) and growth (cm)/(minimum days alive) were calculated. Seedlings were assumed to be at least seven days old upon initial sampling (based on the presence of at least two expanded leaves), and the minimum days alive calculated as the number of days from the initial to the final observation (at which point seedlings were either still alive or recorded as dead or missing) plus seven. For example, a seedling first recorded on 28 October 2015 and noted as dead on 25 August 2016 may have died anywhere between 17 March 2016 to 25 August 2016 and was therefore alive for a minimum of 145 days.
Results
Seedling survey
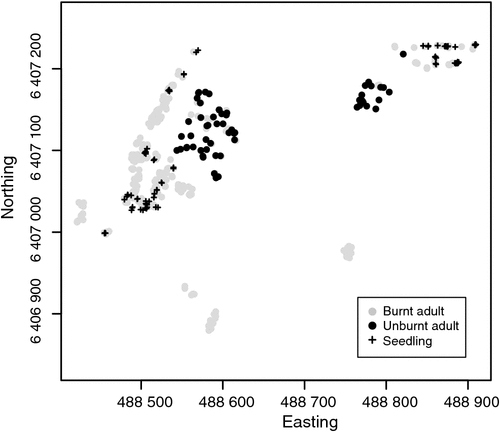
A total of 118 seedlings were found; these were easily distinguishable from coppice growth from adult plants by the presence of dissected cotyledons or glaucous leaves, and by differences in leaf shape and stem thickness (see Fig. S1, available as Supplementary Material to this paper). Seedlings were located only underneath the canopies of burnt adult plants, except for two seedlings which were ~2 m downslope from the canopy edge of the nearest adult (Fig. 1).
|
Genetic diversity
There were some differences in genetic variation between the adult and seedling cohorts, with a 20% reduction in observed heterozygosity (HO adults = 0.582 and HO seedlings = 0.464) and fixation indices of –0.005 in adults and 0.181 in seedlings (Table 1). Allelic diversity (Na, Na freq. <5%, and Ne) was similar for adults and seedlings, but nine rare alleles detected in adults were absent in the sampled seedlings. Despite an increase in seedling number from 31 individuals in the initial survey (1 October 2015) to 92 in the third survey (6 December 2015), and subsequent reduction to 32 seedlings in the final survey (6 April 2017), observed heterozygosity and fixation index of seedling samples did not change to match that of adults. There was little genetic differentiation of adult and seedling cohorts (GST = 0.006 ± 0.002 (s.e.), Fig. S2).

|
Parentage assignment
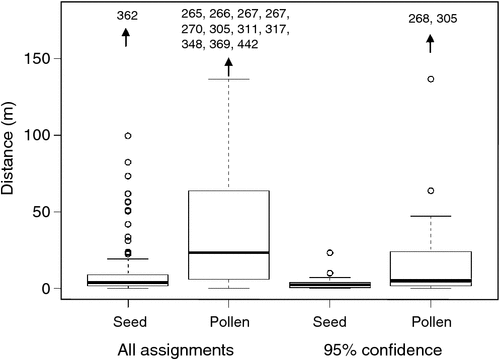
Out of 115 seedlings analysed, 34 candidate parent matches had critical logarithm of odd (LOD) scores meeting the strict confidence threshold and a further 49 matches met the relaxed confidence threshold. Most likely parents were assigned for an additional 22 seedlings. Parents were not assigned for 10 seedlings for which more than two candidate parents were identified (but a pool of potential parents from within the study site was identified). Overall, an absence of non-local alleles suggested no evidence of gene flow from outside the study population that was represented in new recruits. The mean LOD score of all assignments was 12.9 ± 0.5 (s.e.), whereas that for presumed selfs was higher at 18.1 ± 1.2 (s.e., Fig. S3).
A total of 19 seedlings (16% of the total analysed) were identified as products of self-pollination. Considering all assignments, mean distance to the nearest parent was significantly lower than mean distance to the furthest parent (14.6 ± 3.8 (s.e.) vs 67.8 ± 9.4 (s.e.) m, P < 0.001). The same was true when only assignments that met the 95% confidence threshold were considered (6.6 ± 1.8 (s.e.) vs 39.9 ± 12.9 (s.e.) m, P = 0.021; Fig. 2). Maximum recorded dispersal was 362 m (seed) and 442 m (pollen) for all assignments, 82 m (seed) and 353 m (pollen) at the relaxed confidence threshold, and 23 m (seed) and 305 m (pollen) for assignments meeting the 95% confidence threshold. The mean distance between all adults (and mean pollen dispersal expected under random mating) was 158 m, while the mean distance between unburnt adults was significantly lower at 105 m (P < 0.001).
|
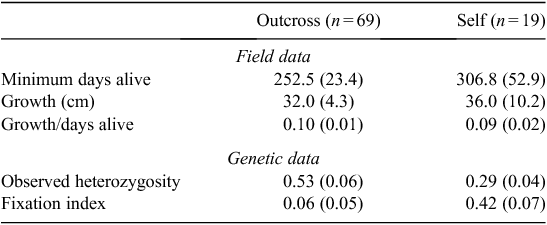
Outcrossed seedlings neither outlived nor outgrew seedlings resulting from selfing (Table 2; Fig. 3), despite being more heterozygous as a group (HO selfs = 0.29 ± 0.04 (s.e.) and HO outcrosses = 0.53 ± 0.06 (s.e.)). Mean growth rates and survivorship of selfed and outcrossed seedlings did not vary significantly (P = 0.723 for growth rate and P = 0.302 for survivorship), although selfs performed slightly better. Of 32 seedlings still alive at the last survey, nine seedlings were selfs, amongst which was the tallest seedling (185 cm).
|

|
Discussion
A recent wildfire in a stand of Eucalyptus caesia in Boyagin Nature Reserve triggered a large-scale recruitment episode that was the first for at least 40 years, but probably much longer based on the size distribution of adults (Hopper et al. 1982; Bezemer et al. 2016). Recruitment in the wild has only ever been recently observed in one other population, also following fire (Yates et al. 2003b). As predicted, parentage analysis revealed that mating is spatially restricted within the study population, with mostly limited seed dispersal but more extensive pollen movement. Contrary to expectations, the inbreeding co-efficient of seedlings remained high even with rapid seedling mortality, and relatively inbred offspring did not have reduced growth rates or survivorship. Population genetic diversity of E. caesia at Boyagin was not enhanced following the documented recruitment episode.
Post-fire recruitment genetics
Post-germination selection against homozygous progeny was not evident over the 20 month survey period. Although there was low heterozygosity and high fixation in seedlings compared with adults, no increasing/decreasing trends in these measures over the survey period were evident. Seedlings that resulted from cross-pollination had greater heterozygosity compared with the products of selfing, but these groups could not be distinguished based on height measurements or survivorship. In contrast, strong patterns of selection have been documented in other plants with mixed mating systems, including Eucalyptus grandis and Grevillea macleayana (Hardner and Potts 1997; Roberts et al. 2014). In the E. grandis common garden experiment, selection was evident after 4 years, corresponding with canopy closure and increased competition. In E. caesia, low seedling densities may render intra-specific competition leading to self-thinning negligible (Westoby 1984). However, inbreeding depression may be more pronounced and more readily detectable with age (Hardner and Potts 1995), with the close-to-zero inbreeding co-efficient of adults at Boyagin suggesting that inbred individuals are eventually selected against. Environmental variability may have confounded selection patterns in the study population (Yates and Ladd 2004; Bailey et al. 2012), although in wild subpopulations of G. macleayana, selection favouring outcrossed progeny was evident despite variability in seedling microsites (Roberts et al. 2014). E. caesia has a root morphology specialised to exploit water resources held in fissures in the underlying rock (Poot et al. 2012), which may explain the distribution of surviving seedlings at the study site.
Selection against homozygotes may have occurred before commencement of the survey. The proportion of outcrossed progeny in canopy-stored fruit of six families from the same population of E. caesia was less than that of recruits in the present study (0.72 vs 0.84; Bezemer et al. 2016), although this could be an artefact of fewer maternal plants included in the Bezemer et al. (2016) study. However, mean pollen dispersal distance in the present study was nearly twice that previously recorded in an open-pollinated seed crop (67.8 ± 9.4 (s.e.) vs 39.4 ± 3.9 (s.e.) m, see Bezemer et al. 2016). These results may indicate selection against progeny resulting from near-neighbour mating during early- or late-stage fruit development, preferential germination of heterozygous seeds (Kennington and James 1997), or, undocumented death of inbred offspring soon after germination. It is also possible that the study population may have undergone purging of deleterious recessive alleles (Byers and Waller 1999; Crnokrak and Barrett 2002). Extant populations of E. caesia appear to have a long history of isolation, over the course of which purging, in combination with genetic drift and limited gene flow, may have contributed to low levels of genetic variation (Byrne and Hopper 2008), but empirical testing of purging is difficult in E. caesia given the longevity of individuals.
Seed and pollen dispersal
Seedlings were generally only found beneath the canopies of adult plants, supporting the prediction of predominant seed dispersal by gravity in eucalypts (House 1997). There was a tendency for seedlings to occur downslope from maternal plants. Previous studies in mallee eucalypts such as Eucalyptus incrassata and Eucalyptus luehmanniana have assumed that seedlings found beneath the canopy of adult conspecifics originated from those plants (Wellington and Noble 1985b; Davies and Myerscough 1991). The present study is among the first to provide direct genetic confirmation of restricted seed dispersal patterns. The assumption that the closest candidate parent to the seedling is the maternal parent is valid because the alternative of seed dispersal towards the pollen parent is implausible and biologically inexplicable (Guidugli et al. 2016). Occasional seed dispersal over relatively large distances was recorded in the present study, perhaps as a consequence of transport by parrots as they leave feeding sites, by water transport along drainage lines, or by convectional updrafts during a fire (Kirkpatrick 1977; He et al. 2004; Southerton et al. 2004). Although rare, such movement may be critical for local population expansion (Vivian et al. 2008).
Mean pollen dispersal distance exceeded that of seed dispersal, and mating occurred frequently between non-neighbours in the study population. Nonetheless, mating was restricted to individuals within the study site, as previously reported for open-pollinated progeny in the same population (Bezemer et al. 2016). The limitation of both studies to a single flowering season may have resulted in failure to detect pollen migration, and so the possibility of mating between populations cannot be discounted. In contrast to the apparently restricted mating documented at Boyagin, frequent pollen exchange has been reported for similarly small or fragmented plant populations (Byrne et al. 2007, 2008; Lander et al. 2010; Breed et al. 2015a; Guidugli et al. 2016). Highly mobile pollinators such as nectar-feeding birds are thought to maintain genetic connectivity between populations (Krauss et al. 2017), yet gene flow appears rare or lacking in the apparently bird-adapted E. caesia (Byrne and Hopper 2008). E. caesia may have other mechanisms in place to cope with genetic and geographic insularity, such as extreme longevity of individuals, biological specialisation or enhanced resilience, or cytogenetic processes conserving genetic variation despite inbreeding (Hopper 2009). Conservation management strategies such as assisted migration or genetic rescue are gaining popularity (Frankham 2015; Whiteley et al. 2015), but may be inappropriate for species such as E. caesia that have intrinsically low genetic diversity and a naturally fragmented and strongly genetically differentiated population structure (Weeks et al. 2011).
Population response to fire
The Boyagin population has a large canopy-stored seed bank, and sixty-eight percent of the standing population was burnt, yet intensive on-ground searches yielded only 118 seedlings. The observed density of less than 10 seedlings per hectare at the study site is considerably lower than post-fire seedling densities reported for other eucalypts, for instance, 7000 seedlings ha–1 in E. incrassata (Wellington and Noble 1985a), 6000 seedlings ha–1 in Eucalyptus tetrodonta (Bowman and Panton 1993) and up to 60 seedlings m2 in Eucalyptus salmonophloia (Yates et al. 1994). Similarly divergent recruitment densities occur between the woody shrub Clusia blattopila, a granite inselberg specialist, and its less restricted congener Clusia nemorosa (Vlasáková 2013). Interestingly, seedling density at the study site was less than recorded for E. caesia at Chidaarcooping Nature Reserve, where nearly 5 seedlings m2 were recorded 10 months after a fire (Yates et al. 2003b).
At Boyagin, low levels of recruitment may be a consequence of predation of seeds by ants, preventing the accumulation of seeds in the soil over long inter-fire intervals (Yates et al. 1995; Bezemer et al. 2013), or a combination of low soil deposition/ high water runoff that washes seed away (Szarzynski 2000). Nonetheless, seedling survivorship was high (with 30% of seedlings still alive at the final survey) compared with other granite outcrop specialists (Yates et al. 2003b, 2007; Vlasáková 2013). At Boyagin, it is interesting to note that seedlings were absent from some burnt patches (see Fig. 1), perhaps as a result of the wildfire reaching a high intensity that killed canopy or soil-stored seed (Auld and Denham 2006; Bezemer et al. 2013).
Large-scale disturbance such as wildfire may be critical to stimulate recruitment in E. caesia (Yates et al. 2003b), but may also have adverse consequences in small plant populations. A reduction in the number of reproductive adult plants may result in loss of attractiveness to pollinators and lower reproductive potential (Menz et al. 2011; Phillips et al. 2014). High intensity fires may contribute to population contraction by killing plants or increasing mortality rates (Wellington and Noble 1985a; Moreno and Oechel 1991). At Boyagin, some ramets have not resprouted, warranting further investigation of the impact of fire.
Conclusions
Wildfire represents a rare but potentially important opportunity for large-scale recruitment in granite-outcrop endemics such as Eucalyptus caesia. The addition of 32 individuals at Boyagin (if all survive to adulthood) represents a substantial expansion of the population. Similar recruitment events are unlikely to occur frequently. It is unclear whether failure to detect selection against homozygous progeny over the survey period is a consequence of the survey duration, environmental variability of microsites, or purging of genetic loads. Further experimentation through common garden trials may provide clarification. Genetic mixing through frequent non-neighbour mating between plants within the study site may delay inbreeding, but appears insufficient to enhance genetic diversity in the Boyagin stand of E. caesia. Here, mating system and pollen dispersal analyses in additional populations would be useful to shed light on the generality of this conclusion. Nonetheless, E. caesia appears adept at persisting as extremely small populations.
Conflicts of interest
The author declares no conflicts of interest.
Acknowledgements
I thank SD Hopper, DG Roberts and SL Krauss for their outstanding supervision and stimulating conversations on E. caesia, JD Roberts and AM Weinstein for providing style edits on a draft of the manuscript, and M Byrne, FAO Silveira, and an anonymous reviewer for providing insightful comments that improved the manuscript. This research was supported by an Australian Postgraduate Award and Holsworth Wildlife Research Endowment awarded to NB, and an ARC Grant (DP140103357) awarded to SDH, SLK and RD Phillips. A licence to take flora for scientific purposes within Department of Parks and Wildlife lands was obtained (SW018638).
References
Auld TD, Denham AJ (2006) How much seed remains in the soil after a fire? Plant Ecology 187, 15–24.| How much seed remains in the soil after a fire?Crossref | GoogleScholarGoogle Scholar |
Ayre DJ, Ottewell KM, Krauss SL, Whelan RJ (2009) Genetic structure of seedling cohorts following repeated wildfires in the fire‐sensitive shrub Persoonia mollis ssp. nectens. Journal of Ecology 97, 752–760.
| Genetic structure of seedling cohorts following repeated wildfires in the fire‐sensitive shrub Persoonia mollis ssp. nectens.Crossref | GoogleScholarGoogle Scholar |
Ayre DJ, O’Brien E, Ottewell K, Whelan R (2010) The accumulation of genetic diversity within a canopy-stored seed bank. Molecular Ecology 19, 2640–2650.
| The accumulation of genetic diversity within a canopy-stored seed bank.Crossref | GoogleScholarGoogle Scholar |
Bailey TG, Davidson NJ, Close DC (2012) Understanding the regeneration niche: microsite attributes and recruitment of eucalypts in dry forests. Forest Ecology and Management 269, 229–238.
| Understanding the regeneration niche: microsite attributes and recruitment of eucalypts in dry forests.Crossref | GoogleScholarGoogle Scholar |
Barbará T, Martinelli G, Fay M, Mayo S, Lexer C (2007) Population differentiation and species cohesion in two closely related plants adapted to neotropical high‐altitude ‘inselbergs’, Alcantarea imperialis and Alcantarea geniculata (Bromeliaceae). Molecular Ecology 16, 1981–1992.
| Population differentiation and species cohesion in two closely related plants adapted to neotropical high‐altitude ‘inselbergs’, Alcantarea imperialis and Alcantarea geniculata (Bromeliaceae).Crossref | GoogleScholarGoogle Scholar |
Barrett LG, He T, Lamont BB, Krauss SL (2005) Temporal patterns of genetic variation across a 9‐year‐old aerial seed bank of the shrub Banksia hookeriana (Proteaceae). Molecular Ecology 14, 4169–4179.
| Temporal patterns of genetic variation across a 9‐year‐old aerial seed bank of the shrub Banksia hookeriana (Proteaceae).Crossref | GoogleScholarGoogle Scholar |
Bezemer N, Kirkpatrick JB, Wood JA (2013) The effect of recent fire history on the abundance and viability of large seeds in the soil of sclerophyll forest in Tasmania, Australia. Papers and Proceedings of the Royal Society of Tasmania 147, 41–50.
Bezemer N, Krauss SL, Phillips RD, Roberts DG, Hopper SD (2016) Paternity analysis reveals wide pollen dispersal and high multiple paternity in a small isolated population of the bird-pollinated Eucalyptus caesia (Myrtaceae). Heredity 117, 460–471.
| Paternity analysis reveals wide pollen dispersal and high multiple paternity in a small isolated population of the bird-pollinated Eucalyptus caesia (Myrtaceae).Crossref | GoogleScholarGoogle Scholar | 1:STN:280:DC%2BC2szhtFeksA%3D%3D&md5=973d5080a7c557a744319f88b17654f0CAS |
Booth TH (2017) Going nowhere fast: a review of seed dispersal in eucalypts. Australian Journal of Botany
| Going nowhere fast: a review of seed dispersal in eucalypts.Crossref | GoogleScholarGoogle Scholar |
Bowman DMJS, Panton WJ (1993) Differences in the stand structure of Eucalyptus tetrodonta forests between Elcho Island and Gunn-Point, northern Australia. Australian Journal of Botany 41, 211–215.
| Differences in the stand structure of Eucalyptus tetrodonta forests between Elcho Island and Gunn-Point, northern Australia.Crossref | GoogleScholarGoogle Scholar |
Bradbury D, Smithson A, Krauss SL (2013) Development and testing of new gene-homologous EST-SSRs for Eucalyptus gomphocephala (Myrtaceae). Applications in Plant Sciences 1, 1300004
| Development and testing of new gene-homologous EST-SSRs for Eucalyptus gomphocephala (Myrtaceae).Crossref | GoogleScholarGoogle Scholar |
Breed MF, Ottewell KM, Gardner MG, Marklund MHK, Dormontt EE, Lowe AJ (2015a) Mating patterns and pollinator mobility are critical traits in forest fragmentation genetics. Heredity 115, 108–114.
| Mating patterns and pollinator mobility are critical traits in forest fragmentation genetics.Crossref | GoogleScholarGoogle Scholar | 1:STN:280:DC%2BC3sbltVeiug%3D%3D&md5=55af50b8432ebdba301ef4af49d33ca4CAS |
Breed MF, Ottewell KM, Gardner MG, Marklund MHK, Stead MG, Harris JBC, Lowe AJ (2015b) Mating system and early viability resistance to habitat fragmentation in a bird-pollinated eucalypt. Heredity 115, 100–107.
| Mating system and early viability resistance to habitat fragmentation in a bird-pollinated eucalypt.Crossref | GoogleScholarGoogle Scholar | 1:STN:280:DC%2BC3s7mvFKqsg%3D%3D&md5=20cfaeecc1d91d6a5f3ada5a2ce883baCAS |
Brondani RPV, Brondani C, Tarchini R, Grattapaglia D (1998) Development, characterization and mapping of microsatellite markers in Eucalyptus grandis and E. urophylla. Theoretical and Applied Genetics 97, 816–827.
| Development, characterization and mapping of microsatellite markers in Eucalyptus grandis and E. urophylla.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaK1cXotVaht7o%3D&md5=23960f68fa1d19a22731815f1ddf465eCAS |
Burrows ND (2013) Fire dependency of a rock-outcrop plant Calothamnus rupestris (Myrtaceae) and implications for managing fire in south-western Australian forests. Australian Journal of Botany 61, 81–88.
| Fire dependency of a rock-outcrop plant Calothamnus rupestris (Myrtaceae) and implications for managing fire in south-western Australian forests.Crossref | GoogleScholarGoogle Scholar |
Byers DL, Waller DM (1999) Do plant populations purge their genetic load? Effects of population size and mating history on inbreeding depression. Annual Review of Ecology and Systematics 30, 479–513.
| Do plant populations purge their genetic load? Effects of population size and mating history on inbreeding depression.Crossref | GoogleScholarGoogle Scholar |
Byrne M, Hopper SD (2008) Granite outcrops as ancient islands in old landscapes: evidence from the phylogeography and population genetics of Eucalyptus caesia (Myrtaceae) in Western Australia. Biological Journal of the Linnean Society. Linnean Society of London 93, 177–188.
| Granite outcrops as ancient islands in old landscapes: evidence from the phylogeography and population genetics of Eucalyptus caesia (Myrtaceae) in Western Australia.Crossref | GoogleScholarGoogle Scholar |
Byrne M, Elliott CP, Yates CJ, Coates DJ (2007) Extensive pollen dispersal in a bird-pollinated shrub, Calothamnus quadrifidus, in a fragmented landscape. Molecular Ecology 16, 1303–1314.
| Extensive pollen dispersal in a bird-pollinated shrub, Calothamnus quadrifidus, in a fragmented landscape.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2sXkvVKruro%3D&md5=ca8e464b1102cddc57941183816dd4d6CAS |
Byrne M, Elliott CP, Yates CJ, Coates DJ (2008) Maintenance of high pollen dispersal in Eucalyptus wandoo, a dominant tree of the fragmented agricultural region in Western Australia. Conservation Genetics 9, 97–105.
| Maintenance of high pollen dispersal in Eucalyptus wandoo, a dominant tree of the fragmented agricultural region in Western Australia.Crossref | GoogleScholarGoogle Scholar |
Coates DJ, Williams MR, Madden S (2013) Temporal and spatial mating-system variation in fragmented populations of Banksia cuneata, a rare bird-pollinated long-lived plant. Australian Journal of Botany 61, 235–242.
| Temporal and spatial mating-system variation in fragmented populations of Banksia cuneata, a rare bird-pollinated long-lived plant.Crossref | GoogleScholarGoogle Scholar |
Crnokrak P, Barrett SC (2002) Perspective: purging the genetic load: a review of the experimental evidence. Evolution 56, 2347–2358.
| Perspective: purging the genetic load: a review of the experimental evidence.Crossref | GoogleScholarGoogle Scholar |
Davies SJ, Myerscough PJ (1991) Postfire demography of the wet-mallee Eucalyptus luehmanniana F.Muell (Myrtaceae). Australian Journal of Botany 39, 459–466.
| Postfire demography of the wet-mallee Eucalyptus luehmanniana F.Muell (Myrtaceae).Crossref | GoogleScholarGoogle Scholar |
Doyle JJ, Doyle JL (1987) A rapid DNA isolation procedure for small quantities of fresh leaf tissue. Phytochemical Bulletin 19, 11–15.
Drummond AJ, Ashton B, Buxton S, Cheung M, Cooper A, Duran C et al. (2011). Geneious V 7.1.4. Created by Biomatters. Available from http://www.geneious.com/ [verified 11 December 2017].
Fitzsimons JA, Michael DR (2017) Rocky outcrops: a hard road in the conservation of critical habitats. Biological Conservation 211, 36–44.
| Rocky outcrops: a hard road in the conservation of critical habitats.Crossref | GoogleScholarGoogle Scholar |
Frankham R (2015) Genetic rescue of small inbred populations: meta‐analysis reveals large and consistent benefits of gene flow. Molecular Ecology 24, 2610–2618.
| Genetic rescue of small inbred populations: meta‐analysis reveals large and consistent benefits of gene flow.Crossref | GoogleScholarGoogle Scholar |
Gevaert SD, Mandel JR, Burke JM, Donovan LA (2013) High genetic diversity and low population structure in Porter’s sunflower (Helianthus porteri). Journal of Heredity 104, 407–415.
| High genetic diversity and low population structure in Porter’s sunflower (Helianthus porteri).Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3sXmtF2jtbY%3D&md5=fce523ee10259e7cf693231dd0d2467dCAS |
Glaubitz J, Emebiri L, Moran G (2001) Dinucleotide microsatellites from Eucalyptus sieberi: inheritance, diversity, and improved scoring of single-base differences. Genome 44, 1041–1045.
| Dinucleotide microsatellites from Eucalyptus sieberi: inheritance, diversity, and improved scoring of single-base differences.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD38XjvVOgug%3D%3D&md5=8076ffc15752daf22e2d558bc3e58044CAS |
Gonçalves-Oliveira RC, Wöhrmann T, Benko-Iseppon AM, Krapp F, Alves M (2017) Maria das Graças LW, Weising K (2017) Population genetic structure of the rock outcrop species Encholirium spectabile (Bromeliaceae): the role of pollination vs. seed dispersal and evolutionary implications. American Journal of Botany 104, 868–878.
| Maria das Graças LW, Weising K (2017) Population genetic structure of the rock outcrop species Encholirium spectabile (Bromeliaceae): the role of pollination vs. seed dispersal and evolutionary implications.Crossref | GoogleScholarGoogle Scholar |
Groger A, Barthlott W (1996) Biogeography and diversity of the inselberg (Laja) vegetation of southern Venezuela. Biodiversity Letters 3, 165–179.
| Biogeography and diversity of the inselberg (Laja) vegetation of southern Venezuela.Crossref | GoogleScholarGoogle Scholar |
Guidugli M, Nazareno A, Feres J, Contel E, Mestriner M, Alzate-Marin A (2016) Small but not isolated: a population genetic survey of the tropical tree Cariniana estrellensis (Lecythidaceae) in a highly fragmented habitat. Heredity 116, 339–347.
| Small but not isolated: a population genetic survey of the tropical tree Cariniana estrellensis (Lecythidaceae) in a highly fragmented habitat.Crossref | GoogleScholarGoogle Scholar | 1:STN:280:DC%2BC28rnt1Cqtw%3D%3D&md5=51ac8fab49adf9837d1d4449c0b3fe7cCAS |
Hardner C, Potts B (1995) Inbreeding depression and changes in variation after selfing in Eucalyptus globulus ssp. globulus. Silvae Genetica 44, 46–54.
Hardner CM, Potts BM (1997) Postdispersal selection following mixed mating in Eucalyptus regnans. Evolution 51, 103–111.
| Postdispersal selection following mixed mating in Eucalyptus regnans.Crossref | GoogleScholarGoogle Scholar |
He T, Krauss SL, Lamont BB, Miller BP, Enright NJ (2004) Long‐distance seed dispersal in a metapopulation of Banksia hookeriana inferred from a population allocation analysis of amplified fragment length polymorphism data. Molecular Ecology 13, 1099–1109.
| Long‐distance seed dispersal in a metapopulation of Banksia hookeriana inferred from a population allocation analysis of amplified fragment length polymorphism data.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2cXktl2rs7s%3D&md5=3948d4c7777aa6ff320ea4d356d3fdb4CAS |
Hmeljevski KV, Freitas L, Domingues R, Pereira AR, Cancio AS, Andrade ACS, Machado MA, Viccini LF, Forzza RC (2014) Conservation assessment of an extremely restricted bromeliad highlights the need for population-based conservation on granitic inselbergs of the Brazilian Atlantic Forest. Flora - Morphology, Distribution, Functional Ecology of Plants 209, 250–259.
| Conservation assessment of an extremely restricted bromeliad highlights the need for population-based conservation on granitic inselbergs of the Brazilian Atlantic Forest.Crossref | GoogleScholarGoogle Scholar |
Hmeljevski KV, Nazareno AG, Leandro Bueno M, Reis MS, Forzza RC (2017) Do plant populations on distinct inselbergs talk to each other? A case study of genetic connectivity of a bromeliad species in an Ocbil landscape. Ecology and Evolution 7, 4704–4716.
| Do plant populations on distinct inselbergs talk to each other? A case study of genetic connectivity of a bromeliad species in an Ocbil landscape.Crossref | GoogleScholarGoogle Scholar |
Hopper SD (1981) Honeyeaters and their winter food plants on granite rocks in the central wheatbelt of Western Australia. Wildlife Research 8, 187–197.
| Honeyeaters and their winter food plants on granite rocks in the central wheatbelt of Western Australia.Crossref | GoogleScholarGoogle Scholar |
Hopper SD (2009) OCBIL theory: towards an integrated understanding of the evolution, ecology and conservation of biodiversity on old, climatically buffered, infertile landscapes. Plant and Soil 322, 49–86.
| OCBIL theory: towards an integrated understanding of the evolution, ecology and conservation of biodiversity on old, climatically buffered, infertile landscapes.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD1MXhtVWisrnN&md5=c6be5b90f584177b989c3c111fd3a652CAS |
Hopper SD, Campbell NA, Moran GF (1982) Eucalyptus caesia, a rare mallee of granite rocks from south-western Australia. In ‘Species at risk. Research in Australia’. (Ed. RH Groves) pp. 46–61. (Australian Academy of Science: Canberra)
Hopper SD, Van Leeuwen S, Brown A, Patrick S (1990) ‘Western Australia’s endangered flora.’ ‘Department of Conservation and Land Management: Perth)
Hopper SD, Brown AP, Marchant NG (1997) Plants of Western Australian granite outcrops. Journal of the Royal Society of Western Australia 80, 141–158.
House SM (1997) Reproductive biology of eucalypts. In ‘Eucalypt ecology: individuals to ecosystems’. (Eds J Williams, J Woinarski) pp. 30–55. (Cambridge University Press: Cambridge, UK)
Jones AG, Small CM, Paczolt KA, Ratterman NL (2010) A practical guide to methods of parentage analysis. Molecular Ecology Resources 10, 6–30.
| A practical guide to methods of parentage analysis.Crossref | GoogleScholarGoogle Scholar |
Kalinowski ST, Taper ML, Marshall TC (2007) Revising how the computer program CERVUS accommodates genotyping error increases success in paternity assignment. Molecular Ecology 16, 1099–1106.
| Revising how the computer program CERVUS accommodates genotyping error increases success in paternity assignment.Crossref | GoogleScholarGoogle Scholar |
Kennington JW, James SH (1997) The effect of small population size on the mating system of a rare clonal mallee, Eucalyptus argutifolia (Myrtaceae). Heredity 78, 252–260.
| The effect of small population size on the mating system of a rare clonal mallee, Eucalyptus argutifolia (Myrtaceae).Crossref | GoogleScholarGoogle Scholar |
Kirkpatrick JB (1977) Eucalypt invasion in southern California. The Australian Geographer 13, 387–393.
| Eucalypt invasion in southern California.Crossref | GoogleScholarGoogle Scholar |
Krauss SL, He T, Barrett LG, Lamont BB, Enright NJ, Miller BP, Hanley ME (2009) Contrasting impacts of pollen and seed dispersal on spatial genetic structure in the bird-pollinated Banksia hookeriana. Heredity 102, 274–285.
| Contrasting impacts of pollen and seed dispersal on spatial genetic structure in the bird-pollinated Banksia hookeriana.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD1MXit1Okurg%3D&md5=6c4388bb585fb62de189d9d77f44016fCAS |
Krauss SL, Phillips RD, Karron JD, Johnson SD, Roberts DG, Hopper SD (2017) Novel consequences of bird pollination for plant mating. Trends in Plant Science 22, 395–410.
| Novel consequences of bird pollination for plant mating.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC2sXks1Smtbs%3D&md5=0763864a8fc39780a0dafd937e04a327CAS |
Lander TA, Boshier DH, Harris SA (2010) Fragmented but not isolated: contribution of single trees, small patches and long-distance pollen flow to genetic connectivity for Gomortega keule, an endangered Chilean tree. Biological Conservation 143, 2583–2590.
| Fragmented but not isolated: contribution of single trees, small patches and long-distance pollen flow to genetic connectivity for Gomortega keule, an endangered Chilean tree.Crossref | GoogleScholarGoogle Scholar |
Llorens TM, Byrne M, Yates CJ, Nistelberger HM, Coates DJ (2012) Evaluating the influence of different aspects of habitat fragmentation on mating patterns and pollen dispersal in the bird-pollinated Banksia sphaerocarpa var. caesia. Molecular Ecology 21, 314–328.
| Evaluating the influence of different aspects of habitat fragmentation on mating patterns and pollen dispersal in the bird-pollinated Banksia sphaerocarpa var. caesia.Crossref | GoogleScholarGoogle Scholar | 1:STN:280:DC%2BC387itlSmtQ%3D%3D&md5=940dc6fe718ee0a72400f7f3a4b31d39CAS |
Menz MHM, Phillips RD, Winfree R, Kremen C, Aizen MA, Johnson SD, Dixon KW (2011) Reconnecting plants and pollinators: challenges in the restoration of pollination mutualisms. Trends in Plant Science 16, 4–12.
| Reconnecting plants and pollinators: challenges in the restoration of pollination mutualisms.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3MXjvFOgsw%3D%3D&md5=f3dd6fd345596fe9a28b9258b5a6be27CAS |
Moran GF, Hopper SD (1983) Genetic diversity and the insular population structure of the rare granite rock species, Eucalyptus caesia Benth. Australian Journal of Botany 31, 161–172.
| Genetic diversity and the insular population structure of the rare granite rock species, Eucalyptus caesia Benth.Crossref | GoogleScholarGoogle Scholar |
Moreno JM, Oechel WC (1991) Fire intensity and herbivory effects on postfire resprouting of Adenostoma fasciculatum in southern California chaparral. Oecologia 85, 429–433.
| Fire intensity and herbivory effects on postfire resprouting of Adenostoma fasciculatum in southern California chaparral.Crossref | GoogleScholarGoogle Scholar | 1:STN:280:DC%2BC1czotVGmuw%3D%3D&md5=92e2f6077c1f12613b3a4ca104881fabCAS |
Nevill PG, Bradbury D, Jørgensen T, Krauss S, Samaraweera S, Gardner MG (2013) Microsatellite primers identified by 454 sequencing in the floodplain tree species Eucalyptus victrix (Myrtaceae). Applications in Plant Sciences 1, 1200402
| Microsatellite primers identified by 454 sequencing in the floodplain tree species Eucalyptus victrix (Myrtaceae).Crossref | GoogleScholarGoogle Scholar |
Peakall R, Smouse PE (2012) GenAlEx 6.5: genetic analysis in Excel. Population genetic software for teaching and research – an update. Bioinformatics 28, 2537–2539.
| GenAlEx 6.5: genetic analysis in Excel. Population genetic software for teaching and research – an update.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC38XhsVehtbjI&md5=5f64d61431a2b6d2b1a54d37d1d4b8e1CAS |
Phillips RD, Steinmeyer F, Menz MHM, Erickson TE, Dixon KW (2014) Changes in the composition and behaviour of a pollinator guild with plant population size and the consequences for plant fecundity. Functional Ecology 28, 846–856.
| Changes in the composition and behaviour of a pollinator guild with plant population size and the consequences for plant fecundity.Crossref | GoogleScholarGoogle Scholar |
Poot P, Hopper SD, van Diggelen JM (2012) Exploring rock fissures: does a specialized root morphology explain endemism on granite outcrops? Annals of Botany 110, 291–300.
| Exploring rock fissures: does a specialized root morphology explain endemism on granite outcrops?Crossref | GoogleScholarGoogle Scholar |
Porembski S, Barthlott W (2012) ‘Inselbergs: biotic diversity of isolated rock outcrops in tropical and temperate regions.’ (Springer Science & Business Media: New York)
Porembski S, Szarzynski J, Mund JP, Barthlott W (1996) Biodiversity and vegetation of small‐sized inselbergs in a West African rain forest (Taı, Ivory Coast). Journal of Biogeography 23, 47–55.
| Biodiversity and vegetation of small‐sized inselbergs in a West African rain forest (Taı, Ivory Coast).Crossref | GoogleScholarGoogle Scholar |
Porembski S, Seine R, Barthlott W (1997) Inselberg vegetation and the biodiversity of granite outcrops. Journal of the Royal Society of Western Australia 80, 193–199.
Porembski S, Silveira FA, Fiedler PL, Watve A, Rabarimanarivo M, Kouame F, Hopper SD (2016) Worldwide destruction of inselbergs and related rock outcrops threatens a unique ecosystem. Biodiversity and Conservation 25, 2827–2830.
| Worldwide destruction of inselbergs and related rock outcrops threatens a unique ecosystem.Crossref | GoogleScholarGoogle Scholar |
Potts BM, Wiltshire RJ (1997) Eucalypt genetics and genecology. In ‘Eucalypt ecology: individuals to ecosystems’. (Eds J Williams and J Woinarski) pp. 56–91. (Cambridge University Press: Cambridge, UK)
Roberts DG, Ottewell KM, Whelan RJ, Ayre DJ (2014) Is the post-disturbance composition of a plant population determined by selection for outcrossed seedlings or by the composition of the seedbank? Heredity 112, 409–414.
| Is the post-disturbance composition of a plant population determined by selection for outcrossed seedlings or by the composition of the seedbank?Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3sXhvVOhsrbF&md5=951e574b12b5a88e9770546c3534d52eCAS |
Southerton SG, Birt P, Porter J, Ford HA (2004) Review of gene movement by bats and birds and its potential significance for eucalypt plantation forestry. Australian Forestry 67, 44–53.
| Review of gene movement by bats and birds and its potential significance for eucalypt plantation forestry.Crossref | GoogleScholarGoogle Scholar |
Steane D, Vaillancourt R, Russell J, Powell W, Marshall D, Potts B (2001) Development and characterisation of microsatellite loci in Eucalyptus globulus (Myrtaceae). Silvae Genetica 50, 89–91.
Szarzynski J (2000) Xeric islands: environmental conditions on inselbergs. In ‘Inselbergs’. pp. 37–48. (Springer: New York)
Tapper SL, Byrne M, Yates C, Keppel G, Hopper S, Van Niel K, Schut A, Mucina L, Wardell‐Johnson G (2014) Isolated with persistence or dynamically connected? Genetic patterns in a common granite outcrop endemic. Diversity & Distributions 20, 987–1001.
| Isolated with persistence or dynamically connected? Genetic patterns in a common granite outcrop endemic.Crossref | GoogleScholarGoogle Scholar |
Vivian LM, Cary GJ, Bradstock RA, Gill A (2008) Influence of fire severity on the regeneration, recruitment and distribution of eucalypts in the Cotter River Catchment, Australian Capital Territory. Austral Ecology 33, 55–67.
| Influence of fire severity on the regeneration, recruitment and distribution of eucalypts in the Cotter River Catchment, Australian Capital Territory.Crossref | GoogleScholarGoogle Scholar |
Vlasáková B (2013) Sexual regeneration on a tropical inselberg in French Guiana: an inselberg specialist versus a species with a wider ecological amplitude. Plant Ecology 214, 279–290.
| Sexual regeneration on a tropical inselberg in French Guiana: an inselberg specialist versus a species with a wider ecological amplitude.Crossref | GoogleScholarGoogle Scholar |
Weeks AR, Sgro CM, Young AG, Frankham R, Mitchell NJ, Miller KA, Byrne M, Coates DJ, Eldridge MDB, Sunnucks P, Breed MF, James EA, Hoffmann AA (2011) Assessing the benefits and risks of translocations in changing environments: a genetic perspective. Evolutionary Applications 4, 709–725.
| Assessing the benefits and risks of translocations in changing environments: a genetic perspective.Crossref | GoogleScholarGoogle Scholar |
Wellington AB, Noble IR (1985a) Post-fire recruitment and mortality in a population of the mallee Eucalyptus incrassata in semi-arid, south-eastern Australia. Journal of Ecology 73, 645–656.
| Post-fire recruitment and mortality in a population of the mallee Eucalyptus incrassata in semi-arid, south-eastern Australia.Crossref | GoogleScholarGoogle Scholar |
Wellington AB, Noble IR (1985b) Seed dynamics and factors limiting recruitment of the mallee Eucalyptus incrassata in semi-arid, south-eastern Australia. Journal of Ecology 73, 657–666.
| Seed dynamics and factors limiting recruitment of the mallee Eucalyptus incrassata in semi-arid, south-eastern Australia.Crossref | GoogleScholarGoogle Scholar |
Westoby M (1984) The self-thinning rule. Advances in Ecological Research 14, 167–225.
| The self-thinning rule.Crossref | GoogleScholarGoogle Scholar |
Whiteley AR, Fitzpatrick SW, Funk WC, Tallmon DA (2015) Genetic rescue to the rescue. Trends in Ecology & Evolution 30, 42–49.
| Genetic rescue to the rescue.Crossref | GoogleScholarGoogle Scholar |
Wyatt R, Evans EA, Sorenson JC (1992) The evolution of self-pollination in granite outcrop species of Arenaria (Caryophyllaceae). VI. Electrophoretically detectable genetic variation. Systematic Botany 17, 201–209.
| The evolution of self-pollination in granite outcrop species of Arenaria (Caryophyllaceae). VI. Electrophoretically detectable genetic variation.Crossref | GoogleScholarGoogle Scholar |
Yates CJ, Ladd PG (2004) Breeding system, pollination and demography in the rare granite endemic shrub Verticordia staminosa ssp. staminosa in south-west Western Australia. Austral Ecology 29, 189–200.
| Breeding system, pollination and demography in the rare granite endemic shrub Verticordia staminosa ssp. staminosa in south-west Western Australia.Crossref | GoogleScholarGoogle Scholar |
Yates CJ, Hobbs RJ, Bell RW (1994) Landscape-scale disturbances and regeneration in semi-arid woodlands of southwestern Australia. Pacific Conservation Biology 1, 214–221.
| Landscape-scale disturbances and regeneration in semi-arid woodlands of southwestern Australia.Crossref | GoogleScholarGoogle Scholar |
Yates CJ, Taplin R, Hobbs RJ, Bell RW (1995) Factors limiting the recruitment of Eucalyptus salmonophloia in remnant woodlands. II. Postdispersal seed predation and soil seed reserves. Australian Journal of Botany 43, 145–155.
| Factors limiting the recruitment of Eucalyptus salmonophloia in remnant woodlands. II. Postdispersal seed predation and soil seed reserves.Crossref | GoogleScholarGoogle Scholar |
Yates CJ, Abbott I, Hopper SD, Coates DJ, Burrows N (2003a) Fire as a determinant of rarity in the south-west Western Australian global biodiversity hotspot. In ‘Fire in ecosystems of south-west Western Australia: impacts and management’. (Eds I Abbott, ND Burrows) pp. 395–420. (Backhuys Publishers: Leiden, The Netherlands)
Yates CJ, Hopper SD, Brown A, Leeuwen S (2003b) Impact of two wildfires on endemic granite outcrop vegetation in Western Australia. Journal of Vegetation Science 14, 185–194.
| Impact of two wildfires on endemic granite outcrop vegetation in Western Australia.Crossref | GoogleScholarGoogle Scholar |
Yates CJ, Ladd PG, Coates DJ, McArthur S (2007) Hierarchies of cause: understanding rarity in an endemic shrub Verticordia staminosa (Myrtaceae) with a highly restricted distribution. Australian Journal of Botany 55, 194–205.
| Hierarchies of cause: understanding rarity in an endemic shrub Verticordia staminosa (Myrtaceae) with a highly restricted distribution.Crossref | GoogleScholarGoogle Scholar |