

Recruitment of Melaleuca quinquenervia (Myrtaceae) in the fringing forests of the Myall Lakes, NSW, Australia
Anne Baumann A and Alison Hewitt B *
B *
A Anne Clements and Associates Pty Limited, Environmental and Botanical Consultants, PO Box 1623, North Sydney 2059, NSW, Australia.
B Hawkesbury Institute for the Environment, Western Sydney University, Locked Bag 1797, Penrith, NSW 2751, Australia.
Australian Journal of Botany 71(3) 127-145 https://doi.org/10.1071/BT22070
Submitted: 4 July 2022 Accepted: 10 March 2023 Published: 4 April 2023
© 2023 The Author(s) (or their employer(s)). Published by CSIRO Publishing. This is an open access article distributed under the Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License (CC BY-NC-ND)
Abstract
Context: Melaleuca quinquenervia is a widespread, keystone species of eastern Australian coastal forested wetlands. Populations of M. quinquenervia in the fringes of Myall Lakes, New South Wales, Australia exhibit low numbers of juveniles, infrequent recruitment and a dominance of mature individuals, raising concerns about stand replacement from conservation managers. In contrast, M. quinquenervia often recruits in large numbers and is highly invasive in wetlands around the world.
Aims: To explore factors that limit recruitment of M. quinquenervia within its native range.
Methods: Field deployed seed traps; soil-core sampling; a burial longevity trial and depth of emergence experiment; laboratory temperature, salinity and light germination experiments; a transplantation experiment; monitoring of field germination; and a litter/leachate experiment.
Key results: Low seed viability (5–26%) is offset by high canopy seed loads (250 000 to 120 million seeds per tree) released continuously throughout the year with a peak fall in summer (2000 to 5000 viable seeds per m2 per month), coinciding with optimal germination temperatures. Seedling emergence is reduced below 5 mm soil depth, there is a very low soil seedbank reserve, and germination is light-dependant. M. quinquenervia exhibited reduced germination percentage and rate as well as reduced survival of seedlings at salinities higher than 15 dS m−1 EC (electrical conductivity). The most favourable sites for seedling establishment are permanently moist with low salinity, protected from wave action via sedge vegetation, and sheltered from prevailing winds. There is an inhibitory effect of Casuarina leachate on germination.
Conclusions: Germination in the field is episodic, and tied to specific environmental conditions. Highest losses occur at the seedling establishment phase.
Implications: Future research avenues and management recommendations are provided.
Keywords: allelopathy, Casuarina glauca, coastal communities, Melaleuca, salinity, seed germination, seedling growth, transplantation experiment, wetlands, viability.
Introduction
The native Australian plant species Melaleuca quinquenervia (Cav.) S.T.Blake (broad-leaved paperbark tree) is a weed of global significance (Global Invasive Species Database 2021). Listed as noxious in Asia, Central and South America, South Africa and the Pacific (Dray et al. 2006; van Kleunen et al. 2015; van Wyk and Jacobs 2015), the species has proven to be an invasive coloniser and difficult to eradicate (Bodle et al. 1994; Tipping et al. 2016; Martin et al. 2019; Tipping et al. 2021). In the Florida Everglades, for example, where M. quinquenervia was first introduced from Sydney in 1886 to drain swamps and increase agricultural land (Turner et al. 1997; Dray et al. 2006), it now infests and outcompetes native plants due to its ability to release seeds in high numbers (Myers 1983), to resprout following fire (Cronk and Siobhan-Fenessey 2001) and to tolerate submergence following flooding (Serbesoff-King 2003).
By contrast, M. quinquenervia in Australia exhibits infrequent recruitment in the field, and environmental parameters to germination and seedling survival are unknown. As far back as 1939 at the Myall Lakes in New South Wales (NSW), low numbers of seedlings, infrequent recruitment, ‘marked stepping’ of size classes and population profiles dominated by mature to old individuals, have been noted (Osborn and Robertson 1939; Myerscough and Carolin 1986; Martin 2002; Baumann 2008). However, recruitment at the Myall Lakes does not appear tied to fire events (Baumann 2008), with fire frequencies in the Myall Lakes National Park of between 5 and 15 years (NSW National Parks and Wildlife Service 2002; Mooney and Maltby 2006; NSW Department of Planning Infrastructure and Environment 2021). The conditions around its occasional, episodic regeneration at the Myall Lakes, and its feared loss as older trees senesce, were the focus of this study.
Melaleuca quinquenervia is a keystone species of several Australian coastal wetland plant communities, including threatened ‘Swamp Sclerophyll Forest on Coastal Floodplains of the NSW North Coast, Sydney Basin and South East Corner bioregions’ (Keith 2004; NSW Scientific Committee 2012). It provides food and shelter for native aquatic and terrestrial fauna, with particular importance as a flowering food source for flying foxes (Pteropus sp.) that are wide-ranging pollinators and seed dispersers of many native plant species (Eby and Lunney 2002; Welbergen et al. 2020). Coastal wetland communities in Australia have been extensively affected by land-clearing, levee construction, drainage work and filling for agriculture and urbanisation (Zoete and Davie 2000; NSW Office of Environment and Heritage 2012). At the Myall Lakes in NSW, where acid-sulfate soils are largely unsuitable for agriculture and protection has been provided in the National Parks Estate (NSW National Parks and Wildlife Service 2002; Great Lakes Council 2009), swamp sclerophyll forest persists and M. quinquenervia fringes the lakeshore of some 8000 hectares of fresh to brackish water.
Given the distribution of M. quinquenervia around and at the lakes edge in both fresh and brackish areas, it was deemed ecologically important to determine the effect of fluctuating salinity on seed germination and subsequent establishment. Van der Moezel and Bell (1987) reported decreased percentage and rate of germination of Australian Melaleuca species with an increasing salinity, and little germination at 200 mM NaCl (~20 dS m−1). However, the study did not include M. quinquenervia.
The effects of temperature, natural light versus dark, and burial depth on germination and seedling emergence of M. quinquenervia were also investigated and reported as factors long considered among the most important influences on these processes (Harper 1977; Bewley et al. 2013). An examination of the effects of seed desiccation on germination success is also reported, because seeds on the foreshores and fringing forests of the Myall Lakes are subjected to repeated wetting or drying owing to the ever-changing lake and water table levels.
The influence of litter and leachates of M. quinquenervia, and that of co-occurring plants such as Casuarina glauca Sieber and Zostera capricorni Asch. on germination success, were investigated in pot experiments. Three hypotheses tested were as follows: (a) germination percentage is reduced by litter cover; (b) seedling emergence percentage is reduced by litter cover; and (c) germination of M. quinquenervia is influenced by leachate from its own species or from Casuarina glauca, either positively or negatively.
Two field-based studies were conducted to address knowledge gaps concerning seedling establishment and recruitment in the field. First, the fate of seedlings from an unanticipated opportune natural germination event was observed and documented for 1 year to inform knowledge of conditions for successful seedling establishment. Second, a translocation experiment was conducted to study field survival of transplanted glasshouse-grown seedlings to investigate the many physical and biological factors that determine the most favourable environments for juvenile establishment.
This study aims to explore key processes involved in regeneration of M. quinquenervia within the fringing forests of the Myall Lakes and to determine the factors most likely to limit these processes in its native range. The study is, thus, useful to conservation efforts in Australia and potentially informative to ongoing work internationally, where the focus is firmly on suppression.
The study aimed to quantify the number and viability of seeds within the canopy available for germination on the foreshores of the Myall Lakes. The quantity and timing of seed fall and the quantity, depth and longevity of a soil seedbank were also investigated for the first time in Australia.
Materials and methods
Study species
Melaleuca quinquenervia (Fig. 1) is a small–medium-sized tree that is naturally distributed in a thin belt along the eastern coast of Australia from Botany Bay in Sydney to near Rockhampton in Queensland, beyond which it extends more sparsely to the tip of Cape York, southern New Guinea, adjacent parts of Indonesia and New Caledonia (Jacobs and Pickard 1981; Boland et al. 2006; Brophy et al. 2013). It occurs in coastal wetlands within about 40 km of the coast, generally between 0 and 100 m above sea level, in warm subhumid to hot-humid climates with an annual rainfall of 900–1400 mm over neutral to acid, deep sandy soils stained black with humus and a pH range of 2.5–6.5 (Marcar et al. 1995; Benson and McDougall 1998; Boland et al. 2006; NSW Scientific Committee 2012).

|
Study area
Myall Lakes National Park (32°26′S, 152°24′E) (Fig. 2) is a Ramsar-listed coastal lake system in NSW, Australia, made up of three inter-connecting, shallow (2.4–3.7 m depth) fresh, brackish and saline water bodies over ~10 000 ha (NSW National Parks and Wildlife Service 2002; Ramsar 2021). The lakes drain a catchment of ~78 000 ha of mainly agricultural land, flowing from the Upper Myall River, and Nerong and Boolambayte creeks (Mid Coast Council 2018). Freshwater also enters the lakes via groundwater drainage from the sand mass of the eastern shoreline (Atkinson et al. 1981) and from rainfall. Water exits the lakes via the Lower Myall River flowing 20 km south into Port Stephens (Palmer et al. 2000).

|
The lake fringing forest of M. quinquenervia trees at Myall Lakes extends back from the lakeshore up to a distance of 20–150 m. This area has a high water table and soils and lake silts of high nutrient and organic content as well as areas of low nutrient, wave washed sands (Myerscough and Carolin 1986). Thirteen study sites were sampled around the lake shore; these sites were largely chosen because of their accessibility by vehicle (Table 1, Fig. 2).

|
Estimation of Melaleuca quinquenervia canopy seed loads
Canopy seed load was estimated from three sites (two mature trees per site were selected as being typical of each location in terms of size and vigour) (Table 1). Fruit load per tree was first estimated by multiplying the average number of capsules per infructescence (the mean of five replicate infructescences) by the average number of infructescences per branch (the mean of five typical branches) by the number of branches on the tree. Binoculars were used, and infructescences and branches were selected randomly. Canopy seed load was derived by multiplying the fruit load per tree by the average number of seeds per fruit (capsule; 250–300). Seeds per capsule were estimated from a random sample of 18 capsules (three per mature tree per site). Individual capsules were left for 4 weeks on a bench and seeds passively expelled or were dislodged with a small brush. Tree heights (estimated by eye) and diameter at breast height (dbh) were recorded.
Melaleuca quinquenervia seed viability
Seed capsules were collected from 12 trees (four per age class of young, mature and over-mature) on the northern side of Casuarina Picnic Area (Table 1). Trees were allocated to age class by measuring their trunk diameter at breast height (dbh) and by observing developmental patterns characteristic of the species. Capsules were stored in paper bags, allowed to dry for 3 weeks on a laboratory bench and then gently tapped or brushed to expel contents. Five replicates of 100 seeds per tree were sown over moistened filter paper in Petri dishes. The 60 petri dishes were arranged randomly into a germination room at 26°C, with constant irradiance by fluorescent light. The number of seeds germinated was recorded. Remaining seeds were tested for viability by soaking in 0.5% 2,3,5 tetrazolium chloride for 6 days at 25°C, then cutting with a scalpel to observe internal stain uptake of tetrazolium chloride (TTC; International Seed Testing Association 2000).
Seed rain
The mass and timing of seed fall over 12 months was estimated from one trap per site at six sites throughout the fringing forest (Table 1). Traps were constructed from green Ripstop® nylon, with a polar fleece section in the middle (to allow drainage) and suspended with rope from four steel posts driven into the ground to a height of approximately 1 m. Each trap formed an upside-down pyramid with base measurements 400 mm × 400 mm, and a central peak weighed down by a large fishing sinker. The design proved durable and effective in ground that was intermittently wet or dry, depending on lake water levels and an often windy environment. Traps were installed under the outer third of the tree canopy and contents were collected and replaced at near monthly intervals, and, hence, a total of 11 times over 12 months.
Trap contents were sieved and the larger litter items such as leaves, capsules and flowering parts were removed. The remaining fine material was hand-aspirated using a threshing tray down to seed only. Seed fall was calculated as g m−2. Four samples of 0.05 g from each trap each month were spread on agar (7 g L−1) in 9-mm Petri dishes and placed into a germination room at 25°C with constant irradiance by fluorescent light. The numbers of seeds that germinated (as indicated by radicle emergence) were counted, the total number of viable seeds per collection was calculated and converted to the number of viable seeds per m2 per location.
Quantification of the soil seedbank
Twenty soil cores (50 mm in diameter and 80 mm in depth) were taken under and around the canopy of one M. quinquenervia at each of three sites (four samples in each of five directions extending outward from the trunk in 1 m steps) (Table 1). Mature trees were selected that were more than 10 m from any other M. quinquenervia trees. Cores were kept moist while being transported back to the laboratory, then spread evenly onto trays containing wet vermiculite with the surface soil at one end of the tray and the 80 mm depth at the other. Trays were kept in a microclimate with 25–18°C alternating day–night temperatures (conditions ideal for germination – see later). Germinants of M. quinquenervia were identified and recorded at the three- to four-leaf stage when they display a distinctive leaf shape, parallel venation, and a strong smell of ‘tea tree’ oil on pinching the leaves, enabling their discrimination from other species.
Seed-burial longevity trial
Seed longevity in the soil was tested over 12 months by burying 12 parcels each of 50 potentially viable or ‘full’ seeds at a depth of 20 mm (the maximum depth at which seeds were found in the soil-core study). Burial was under a mature M. quinquenervia canopy in the sandy foreshore soil at Mungo Brush in April 2006 (the experimental site chosen because of its location in a fenced exclusion zone, unlikely to be disturbed).
Parcels were made from nylon-mesh stocking material (Van et al. 2005), their positions being marked with a peg. Each month, one parcel was removed from the soil, returned to the lab and opened. The seeds were examined under a microscope, germination or deterioration was recorded, then seeds were placed onto moist filter paper in a Petri dish and put in the seed-germination room at 25°C with constant irradiance by fluorescent light for 21 days. The remaining, ungerminated seeds were tested for viability with TTC per methods outlined above.
Effect of temperature on germination
Seeds from five foreshore sites (Table 1) were collected and mixed together in equal proportions (provenance was not being tested). Twenty-five seeds were sown per Petri dish half-filled with agar at 7 g L−1, sealed with parafilm and placed into growth chambers at six constant temperatures of 5°C, 10°C, 15°C, 20°C, 25°C and 30°C and three alternating (night/day) temperatures: 5°C/20°C, 12°C/27°C and 18°C/33°C each with a 12:12 h fluorescent light:dark photoperiod cycle. Each treatment (Petri dish of 25 seeds) was replicated eight times. The three alternating temperature regimes were selected to simulate temperatures occurring at the Myall Lakes in summer (18°C/33°C), winter (5°C/20°C), and spring and autumn (12°C/27°C) (nearest weather station data from Bulahdelah).
Germination was recorded every day for the first 2 weeks and then three times a week until there was no further germination for 2 weeks. The 5°C and 10°C treatments were transferred into the 25°C cabinet after showing no germination at Day 54. Mean germination percentage was recorded along with the speed of germination as time to 85% of the final germination achieved (T85).
Effect of light versus dark on germination
Seeds were germinated either in the light or in the dark in two replicate experiments. In each, there were six Petri dishes with 200 M. quinquenervia seeds per dish. Seeds were placed on filter paper and moistened to glistening with distilled water and the Petri dishes were wrapped in clear Parafilm. Experiment 1 was set up and kept under natural light conditions filtered through white plastic in a microclimate room at 25°C and a 12:12 h light:dark photoperiod cycle. Experiment 2 was set up and inspected in a dark room under green ‘safelight’, the Petri dishes were additionally wrapped in black plastic followed by aluminium foil then placed into a lightproof box lined with black plastic before being moved to the microclimate room.
Seeds exposed to the light were checked for germination every few days. Seeds in the dark were checked under a green ‘safe’ light in the dark room at Day 20 and Day 63. On Day 63, the seeds that were in the dark were transferred into the light. At the end of the trial (Day 80), remaining ungerminated seeds were tested for viability with TTC.
Effect of depth of planting on germination
Five replicates, each of 25 seeds, were sown at five different depths (0, 5, 10, 20 and 30 mm), into pots of 95 mm width, 100 mm depth containing medium–fine nutrient-poor sand from the Myall Lakes foreshore zone. Sand was first inundated with water, allowed to settle and the surface was levelled before a piece of filter paper was placed on the surface and seeds sown into small pinholes in the paper. Seeds were covered to the required depth, pots were labelled and placed randomly within trays in a glasshouse at 18°C/25°C alternating night/day temperature and filtered daylight, and were watered from the bottom up with tap water. Pots were checked daily for seedling emergence.
Effect of salinity on germination
Artificial seawater was made to a modified formula from McLean and Ivimey Cook (1941), then diluted to achieve the desired electrical conductivities (EC). Two experiments were conducted in a germination room at 25°C. (1) Five replicates of 25 seeds were sown over filter paper in Petri dishes at seven levels of salinity (2, 6, 7, 10, 15.5, 20 and 28 dS m−1 to cover the naturally occurring range at the Myall Lakes) plus a control of deionised water. Germinants were recorded daily for 14 days. (2) Five replicates of 25 seeds were sown over an 18 mm depth of sand sourced from the Myall Lakes foreshore zone in Petri dishes watered with artificial seawater diluted to six levels of salinity (2, 5, 10, 15, 20 and 25 dS m−1) plus a control of deionised water. Germinants were recorded daily for 22 days. After 22 days, seedlings were transplanted into pots by carefully transferring the entire contents of the Petri dish onto a pot of foreshore zone sand. These pots were put on trays in a microclimate of 18°C/25°C alternating night/day temperature under natural lighting and watered from the bottom up with the corresponding strengths of artificial seawater. Final survival percentage was calculated at 8 weeks.
Seed-desiccation experiment
Four replicates of 25 potentially viable seeds from Angophora Picnic Area (Table 1) were sown onto filter paper in Petri dishes that had been watered with 12 mL of deionised water. The dishes were sealed with parafilm and placed in an incubation room at 25°C (±1°C), and the seeds were allowed to imbibe for nine increasing lengths of time: 1, 1.5, 2, 2.5, 3, 3.5, 4, 4.5 and 5 days. After their specified imbibition period, the seeds were dried out for 5 days by removing the lid of the Petri dish (Ren and Tao 2003) and then left in the incubation room where humidity ranged from 39% to 40%. Dishes were re-watered with deionised water and resealed after 5 days allowing the seeds to re-imbibe. Germination numbers and observations were recorded for 20 days after the drying and re-wetting periods. Ungerminated seeds were then tested for viability by staining with TTC.
Litter/leachate experiment
Casuarina glauca cladodes, M. quinquenervia leaves and Zostera capricorni stems and leaves (herein referred to as litter) were collected from the foreshore areas at the Myall Lakes. Casuarina and Melaleuca leachate was made by soaking litter in water for 10 days at a ratio of 1:5 (v/v) dry litter to water.
Pots (95 mm in diameter, 100 mm deep) were filled with washed medium-grade sand and sown with 75 seeds of M. quinquenervia from Myall Shores and The Wells. There were the following eight treatments, each replicated four times: (1) Melaleuca leachate; (2) Melaleuca litter plus leachate; (3) Casuarina leachate; (4) Casuarina litter plus leachate; (5) Melaleuca litter; (6) Casuarina litter; (7) Zostera capricorni litter; and (8) control (no leachate or litter). Litter cover was observed to be a range of thicknesses in the field. A minimum thickness of 15 mm of M. quinquenervia leaves (about four layers) was considered the minimum amount to completely cover the surface of the pot with no gaps between the leaves. Casuarina litter and seagrass treatment pots were covered to the same thickness. Pots were watered from below with the treatment leachate or with tap water if the treatment prescribed no leachate. Numbers germinated (radicle emergence) were recorded and final germination percentage was calculated on Day 39 along with days taken to reach 50% of the final germination percentage (T50).
Survival and mortality from a natural germination event
In January/February 2006, a natural germination event occurred on the Myall Lakes foreshore. Bands and patches of seedlings 10–50 mm tall with seedling numbers estimated to be in the thousands were noted on wet sand exposed by a receding lake level. Five sites with different aspects around the lake were selected for monitoring the growth and survival of seedlings – three on the Myall Lake (Neranie Sands, Neranie (Little Beach) and One Horse Sands), and two on the Bombah Broadwater (Angophora Picnic Area and Casuarina Picnic Area) (Table 1).
A 1-m2 quadrat subdivided into four 0.25 m2 subquadrats was pegged out with wooden stakes within the seedling zone at each site in April 2006. Seedlings growing within each quadrat were plotted using a thin nylon grid and their position and height were recorded. Foreshore sites were visited on or near monthly for 15 months. At each visit, seedling numbers and heights were recorded, photographs taken, and lake levels (depth to the water table using a hand-auger and tape measure) and lake salinity (using an electrical conductivity (EC) meter (Salttestr EC11)) were recorded. General observations were documented, such as the presence of lake foam, salt spray, tannins in the water or seagrass washing over quadrats. Germinant survival, growth, mortality and new germination events were derived from these data and survival curves were constructed.
Seedling transplantation experiment
Melaleuca seed was collected from four sites (Paperbark Picnic Area, West Legges, Mayers Point and One Horse Sands; Table 1) and kept separately to prevent mixing of provenances. Seeds were mixed with sand, sieved and sprinkled into labelled propagating cells containing pasteurised seed-raising mix (Stewart and Stewart 1995), then kept in trays in a glasshouse with alternating night/day temperatures of 18°C/25°C under natural light. A version of the wetland/bog method (Ralph 2003) was employed, with cells watered from the bottom up. Seedlings were thinned to one per cell at the two- to three-leaf stage. At the four- to five-leaf stage, the cells were transplanted into 125 mm forestry tubes containing pasteurised potting mix to prevent any introduction of pathogens to the National Park.
Seedlings were grown in the glasshouse for 3 months and then hardened-off (acclimated to outside conditions) on a raised bench. When seedlings were 6 months old and approximately 200–250 mm tall, the 24 most uniform in height from each provenance were selected for transplanting. Transplantation took place in late April 2006 when air temperatures were cooling down, but soil temperatures were expected to remain warm with autumn rains anticipated.
Seedlings were transported in trays to their respective sites of origin and planted directly into the lake fringe vegetation along a transect perpendicular to the shoreline. Distances between the 24 seedlings differed by up to a factor of two (0.75–1.5 m apart) owing to the varying width of the paperbark fringe, e.g. steeper rocky headlands tended to have a narrower fringe of M. quinquenervia than lower, flatter, lake shore areas). Seedlings were watered once at planting with freshwater carried to the site.
Seedlings were protected initially with green tree bags that were removed 7 months after planting. The heights of each seedling were recorded at the time of planting (27 April 2006) then every 2 or 3 months for 10 months and again at 15 months. Seedling growth was measured as the difference in height at 6 months and 15 months from the height at planting. Depth to water table was measured several times during the experiment and salinity of the water table and lake water was measured as per the seedling survival investigation above.
Data analysis
Data with respect to seed viability were analysed by a two-factor ANOVA (trees nested within maturity class). Data for mean percentage germination and T85 in the temperature experiment, mean percentage germination in the light experiments, emergence percentage in the depth of planting experiment, mean percentage germination and T50 in the salinity experiment and the litter/leachate experiment were analysed via series of one-way ANOVA, by using the statistical package Genstat (7th edn, https://vsni.co.uk/software/genstat). Heteroscedasticity was assessed using Levene’s test, and percentage data were arcsine transformed as required. Post hoc comparisons of means were made using Fishers protected least significant differences (lsds) at P = 0.05 (Clewer and Scarisbrick 2013). Graphs were constructed in SigmaPlot (V7).
For the transplantation experiment, diary notes and photographs were reviewed alongside Bureau of Meteorology (BOM) weather data, lake level records by Manly Hydraulics (Government Water Engineers) and our own lake salinity data. Survival curves were constructed and transplant height was graphed against distance from shore at each site after 6 and 15 months (see Baumann 2008).
Results
Canopy seed load
There was a 500-fold variation in the number of seeds held within the canopies of trees, each tree holding from several hundred thousand to over one hundred million seeds (Table 2). The number of seeds in the trees at Mayers Point were fewer, corresponding to the generally smaller size of the trees than those at the other sites (Table 2).

|
Seed viability
Seed viability from individual trees at Casuarina Picnic Area ranged from 5% to 26%. Germination percentages were equal to viability, with no remaining viable seeds that did not germinate during the experiment. The effect of plant maturity (size) was found to be highly significant (F2,59 = 59.43, P < 0.001), with the young plant class having a significantly higher mean seed viability of 18.25% than for seeds produced by the mature and over mature plant classes (8.9–10.65%), which themselves did not significantly (P > 0.05) differ from one another.
Seed rain
The mean number of viable seeds falling per square metre across sites was higher in the warmer months, with a clear decrease during autumn/winter (Fig. 3). Seed fall followed a similar trend at all locations, except Mayers Point, which decreased after an October peak and never recovered.

|
Quantification of soil seedbanks
Nil to eight seeds germinated per soil core, with an average of 1.1–3 viable seeds per soil core per site, resulting in estimates of 560–1528 viable seeds per square metre in the soil across the sites. None of the core samples showed any viable seeds below a depth of 20 mm.
Seed-burial longevity trial
There was no decline in seed survival after 2 months of burial, with 100% germination in retrieved seed (Fig. 4). Viability remained high after 6 months of burial (91.5%), but was noticeably reduced after 7 months (52%) and further reduced to 36% at 12 months (Fig. 4). Observation of the seeds each month did not show any signs of insect damage or disintegration, seeds appeared black and still intact. A polynomial curve fitted to the data gave the best fit (R2 = 0.84) leading to a prediction of complete loss of viable seed by about 15 months (Fig. 4).

|
Effect of temperature on germination
The fastest germination was at 18°C/33°C (P = 0.05), with 85% germination by Day 4, followed by constant 30°C and then 25°C (Fig. 5). The remaining temperatures had little or no germination effect at this time. By Day 20 the 18°C/33°C, 30°C, 25°C, 20°C and 12°C/27°C treatments had reached near to 100% germination, with no significant differences among them (Fig. 5). By Day 54, the 5°C and 10°C treatments still had little or no effect on germination (Table 3).

|

|
The three fastest rates of germination were at 25°C, 30°C and 18°C/33°C, with no significant difference in time taken to 85% germination (Fig. 5). However, they were significantly more favourable than the remaining temperature treatments. The 5°C/20°C treatment along with 15°C had the slowest germination rate, with an average T85 of 20 and 22 days respectively, and neither 5°C nor 10°C reached 85% within the duration of the experiment (54 days; Fig. 5). After 54 days, when the remaining ungerminated seeds in the coldest 5°C and 10°C cabinets were transferred to the 25°C cabinet, they commenced rapid germination, with an average germination by Day 3 of 17% (the 5°C treatment) and 72.3% (the 10°C treatment).
Effect of light versus dark on germination
Melaleuca quinquenervia had a significantly greater germination percentage in the light treatment than in dark (F1,11 = 335.12, P < 0.001), with no seeds having germinated by Day 20 in the dark treatments, and only 11.1% by Day 63. In contrast, seeds kept in light conditions germinated from Day 6 (average = 72%) onward and four of five dishes had reached 100% germination by Day 12. Once exposed to light, seeds that had been kept in the dark germinated rapidly, reaching 100% within 7 days.
Effect of depth of planting on germination
Emergence from 5 mm was significantly reduced compared with that of surface-sown seed (F1,9 = 47.04, P < 0.001). There was no emergence of seedlings from the depths of 10, 20, 25 or 30 mm by 80 days.
Emergence of the surface-sown seeds (0 mm) had a time to 50% emergence averaging 20.6 days. On further investigation at the end of the trial, it was found that seeds as deep as 30 mm had germinated, but after 16 weeks none was able to reach or penetrate the surface of the soil and the experiment was terminated. The ungerminated seeds were retrieved from several pots and put on moist filter paper in a Petri dish and incubated at 27°C. Within 2 days, the majority had germinated.
Effect of salinity on germination
Germination on filter paper
Increasing salinity up to 10 dS m−1 was found to have little effect on final germination percentages, which was 97.6% (Table 4). There was a significant decrease in germination with an increasing salinity from 10 to 28 dS m−1 (F7,39 = 313.08, P < 0.001); at the highest salinity strength tested (i.e. 28 dS m−1), no seeds germinated. Thus, it appears the limit for germination occurs somewhere between 20 and 28 dS m−1 (Table 4).
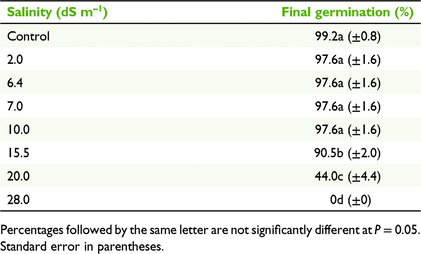
|
Germination on sand substrate
Final percentage germination was not significantly affected by salinity up to 10 dS m−1 (Table 5). Final percentage germination was significantly lower at 15 dS m−1 (84%) and was substantially and significantly lower again at 25 dS m−1 (down to 6.4%; F6,34 = 72.64, P < 0.05). The median germination rate (1/T50) was unaffected up to 5 dS m−1, after which it decreased at a constant rate with an increasing salinity (Table 5).
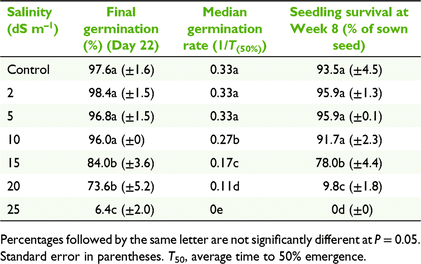
|
Survival percentage for the transplanted seedlings decreased with an increasing salinity. The level of tolerance for survival of 8-week-old seedlings showed a significant reduction from 15 dS m−1 (78% survival) and another significant reduction at 20 dS m−1 (10% survival), with no survival at 25 dS m−1. Morphological differences were also observed from 10 dS m−1 onward, these being a reduction in the number of leaves developed and increasingly broader and shorter leaves. Some roots were exposed at the sand surface and grey in colour, progressing to black at 20 dS m−1.
Seed-desiccation experiment
Increasing periods of imbibition prior to imposing desiccation saw increasing numbers of seeds germinate and concomitant decreasing numbers of germinants surviving desiccation (Table 6). Imbibition for 1 and 1.5 days resulted in no germination prior to desiccation. Once being re-wet, the majority of these seeds germinated. Imbibition for 2.5 days or longer resulted in the majority of seeds germinating, with few of them surviving subsequent desiccation (Table 6). ANOVA found no significant difference in final mean germination percentage among imbibition periods (F8,35 = 0.9, P = 0.53).
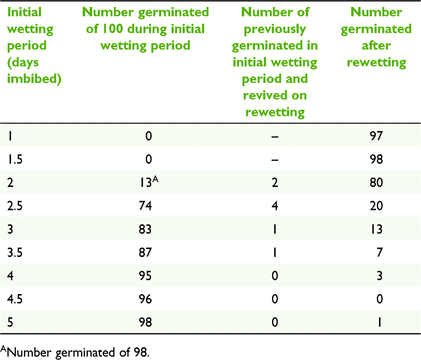
|
Litter/leachate experiment
Leachate-only treatments
The first signs of germination were seen on Day 8. On Day 14, when seedlings were checked under the microscope, the roots in both leachate treatments but not the control showed some signs of reaction to toxicity, i.e. browning of roots, necrosis of root tips and loss of geotropism (roots turning away from sand surface and pointing upward). There was no significant difference in final germination at Day 39 between the control and the M. quinquenervia leachate treatments (Fig. 6). However, germination percentage in the C. glauca leachate was significantly lower than in the M. quinquenervia leachate and the control (F2,11 = 8.27, P = 0.009). treatments. T50 was not significantly different between either of the leachate treatments or the control. By Day 25, all three treatments averaged over 80% germination (Fig. 6) and the seedlings were at cotyledon stage,

|
Litter treatments
By Day 32, seedlings of M. quinquenervia were visibly observed emerging through the C. glauca cladodes. No seedlings emerged through the M. quinquenervia leaves, or the seagrass, which had formed a dense mat. On Day 32, the litter was carefully lifted to determine whether seeds had germinated underneath. Germination had occurred under all litter-cover treatments, including the seagrass; however, because of an uncommon occurrence of cutworm infestation (moth larvae) under all the litter types, exact numbers could not be counted. Seedlings that had not emerged and were still present and intact among the moth larvae infestations were observed to have elongated hypocotyls, little or no root development and a lack of colour. Some seedlings under the Casuarina litter treatments were healthier in appearance, with green cotyledons.
Survival and mortality from a natural germination event
One hundred per cent mortality was recorded in all quadrats at One Horse Sands by 2 June 2006, at Little Neranie by 3 July 2006, at Angophora Picnic Area on 4 July 2006, and at Casuarina Picnic Area on 31 July 2006. All seedlings in the locations more broadly (outside of the quadrats established) were also noted to have died. Only seedlings at the fifth site, Neranie Sands, survived through until 11 February 2007, with two prolific germination events recorded there on 9 December 2006 and 11 February 2007 (Table 7). With rising lake levels in winter, seedling mortality was much higher for small seedlings (1–50 mm tall) than for larger size classes (see site-level survival curves in Baumann 2008).
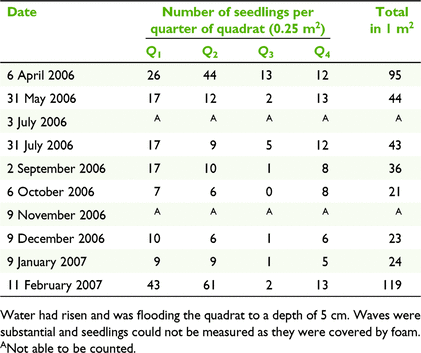
|
Seedling transplantation experiment
Survival of the 6-month-old transplanted seedlings was consistently high at all four sites for the first 6 months (Fig. 7). This was despite the fact that rainfall through autumn and into late winter was unusually low compared with normal monthly averages. After 9 months (6 January), between two and three seedlings had died at each site and after 15 months (7 July), there was a significant reduction in numbers at Paperbark Picnic Area and Mayers Point, and a slight drop at One Horse Sands and West Legges (Fig. 7).

|
Once the tree bags were removed (after 7 months), seedling growth was noted to be affected by herbivory and insect attack at all four sites. Leaves were chewed and growth tips removed. Some seedlings showed the ability to withstand a certain amount of herbivory by developing new shoots at the point where the tip was removed; others developed epicormic shoots towards the base.
There were no obvious similarities or trends in transplant growth among the four sites in relation to distance from the shoreline. Growth differed at each site and was more subtly affected by lake levels, groundwater and lake salinity (as well as damage from herbivory).
Discussion
This series of controlled laboratory experiments, glasshouse and field investigations and the opportune monitoring of seedling survival from a natural germination event, has enabled us to confirm the key environmental parameters required for the episodic recruitment of M. quinquenervia in the fringing forest at the Myall Lakes in NSW, Australia.
Canopy seed load
Canopy seed loads of several hundred thousand to over one hundred million seeds per tree were substantially higher than seed-load estimates reported from congeners, such as, for example, M. styphelioides at 25, 638 per tree, M. nodosa 106, 099 per shrub and M. deanei 28, 783 per shrub (Hewitt et al. 2014). The wide range in numbers of seeds held within the canopies of individual M. quinquenervia trees reflects the combined factors of capsule numbers and the size and density of canopies, and would also vary with a plant’s vigour, pollinator activity and insect infestation (Jenkins et al. 2010). It is also noted that this study was conducted in winter when flowering had finished and immature fruits were forming on top of a previous year’s fruit load. Melaleuca quinquenervia appear to be highly fecund and the size of its canopy seedbank in the fringing forests of the Myall Lakes is large and unlikely to be a limiting factor to recruitment of the plant.
Seed viability
The low seed viability found in this study (5–26%) is comparable with that in earlier work by Meskimen (1962) who found viability rates of between 3% and 28% for M. quinquenervia, and Rayachhetry et al. (1998) who reported that an average 15% of canopy-stored seeds of M. quinquenervia contained embryos and were, therefore, potentially viable; both studies were conducted in the species’ invaded range of eastern USA.
Low seed viability appears typical of the genus more widely, with consistent reports of low viability (filled seed as a proportion of chaff), but high germinability from viable seed (see Hewitt et al. 2014, and references therein). Low seed viability can reflect a number of factors such as failed seed set or abortion by self-pollen, pollinator limitation, resource limitation or pre-dispersal seed predation (Sutherland 1986; Fenner et al. 2005). Low seed viability in M. quinquenervia would appear to be more than offset by the high seed numbers produced each year by mature trees.
Seed rain
Seed rain was high and continuous all year, albeit with a peak during summer approximately 10 times that of winter. A likely explanation for the lower seed fall at one site than at others in this study is a previous mass release of seed (although with no resultant seedling survival) triggered by a fire, which occurred at that site in the October, preceding trap installation (Table 1). The seasonal pattern to seed fall indicates that conditions are most suitable for seed release over summer when generous numbers of seeds fall and are potentially available for germination.
Presence of a soil seedbank
This study has shown the minor role played by the soil seedbank for M. quinquenervia in its regeneration, and that most of the seeds in soil are located at or within the first few millimetres of the soil surface. Despite surface seeds being more vulnerable to predation and environmental extremes than are more deeply buried seeds (Van et al. 2005), not being buried at great depth is likely to be advantageous for M. quinquenervia because the seeds lack a large food reserve for germinating seedlings to draw on during emergence to the soil surface and prior to the first leaves being photosynthetically active (Baskin and Baskin 1998). Shallow soil-burial depth also ensures seeds’ access to light, which was found to be a germination requirement for M. quinquenervia.
Seed-burial longevity trial
Melaleuca quinquenervia appears to have a shorter longevity in the soil than do other Australian species that have a persistent soil seedbanks, such as the fire-prone myrtaceaous species Kunzea ambigua and K. capitata (Auld et al. 2000). These species were found to have seed half lives in the soil greater than 2 years. Although the canopy is a consistent source of new seeds, for M. quinquenervia, the soil seedbank study at the Myall Lakes will suggest a complete loss of viability by 15 months. The implication of the finding for conservation indicates the importance of tree retention, particularly of older trees with their much larger seed output. In contrast, in invaded regions, control may be best focused on tree removal prior to the next seasons fruiting to reduce propagule pressures, with only a 1-year follow-up needed to target removal of new seedling recruits. This assertion is consistent with recent work by Matthys et al. (2022) who reported similar seedbanking strategies in another invasive Australian melaleuca, M. rugolusa, in South Africa.
Seed survival is no doubt influenced by the environment in which it is buried. Van et al. (2005) found the rate of loss of viability appeared to depend on the moisture content of the soil, with M. quinquenervia seeds buried at wet sites losing viability much faster than those buried in dry sites. Expected seed survival would depend, therefore, on the frequency of inundation in the area. Soil type is also a factor, with Van et al. (2005) finding that M. quinquenervia seed survived longer in sandier soils (up to 2.3 years). Studies on seed survival and germinability in different soil types would be especially interesting, given the high recruitment reported from overseas studies. Soil properties such as oxygen content, density and chemical composition, would also be worth investigating in relation to longevity of seed.
Consistent with previous germination studies in M. quinquenervia (Rayachhetry et al. 1998; Van et al. 2005), and the genus more broadly (Robinson et al. 2006; Hewitt et al. 2015), this study showed little difference between germination figures and viability figures.
Effect of temperature on germination
The optimum constant temperature for the germination of M. quinquenervia seeds in this study was 25°C, which is in agreement with Ralph (2003) and Myers (1975, cited in Myers 1983). The alternating temperature of 18°C/33°C had a faster germination rate and was, therefore, the optimum in this experiment, indicating summer as the likely best time for germination. It is well known that fluctuating temperatures can have a stimulatory effect on germination (Thompson and Grime 1983), the physiological mechanisms apparently tied to increased levels of specific germination promoting hormones and enzymes (Ozden et al. 2021). The 18°C/33°C regime had a significantly faster rate of germination than did the 12°C/27°C regime. This may be due to the warmer night temperature (18°C) rather than the warmer day temperature (33°C), as suggested by the more rapid rate of germination at 25°C than at 30°C. It is unlikely to be due to any difference in the size of temperature fluctuation, as they both differed by 15°C. The increased germination at warmer temperatures suggests that maximum germination in the field would occur over summer, with 4–9 days required for natural seed fall to obtain ~85% germination.
The slow germination in the cooler temperature treatments suggests that germination is unlikely to occur during winter when optimum conditions would need to be maintained for 20–60 days for natural seed fall to obtain ~85% germination, on the basis of a continuous number of days experiencing the same temperatures. However, the cold temperatures did not appear to have any detrimental effect on the seeds’ ability to achieve 100% germination once they were returned to optimum temperatures.
Effect of light versus dark on germination
Melaleuca quinquenervia has a clear dependence on light for germination, which contrasts with the report of White (1988) that germination was not stimulated by light. The stringent methods used in this experiment to exclude light from treatments via the use of a green ‘safe’ light (Withrow and Price 1957) in a dark room may account for this discrepancy. This is not to say that germination cannot occur in shaded conditions, as was observed by Meskimen (1962). The wavelength required for germination in these situations requires further study. However, it is clear that the complete absence of light prevents germination, with seed subsequently moved to the light germinating within days.
Effect of depth of planting on germination
Seed germination and emergence were significantly higher from the soil surface, suggesting that in the field even if seeds do lodge deeper down the soil profile, the few that might germinate would have little or no chance of emerging. The fact that seeds germinated to a depth of 30 mm in the experiment indicated that enough light was available at this depth in lakeshore sand for germination; however, the seeds did not have enough resources to reach the surface. The insulating effects of burial below a few centimetres might also mute the stimulatory effects of temperature fluctuations on germination, as reported by Probert (2000).
Effect of salinity on germination
This study confirmed the importance of freshwater for germination of M. quinquenervia seeds, with no reduction in final percentage in salinities up to 10 dS m−1 and a significant reduction at salinities from 15 to 20 dS m−1. These higher salt concentrations are typical during low water levels at the Myall lakes, with the Bombah Broadwater area record being as high as 21.8 dS m−1 during this period of study. The lack of germination at 28 dS m−1 on the filter paper, and the extremely low germination at 25 dS m−1 on sand, suggest that the limit for germination lies somewhere between these two salinities. This is comparable to the tolerance level reported for the germination of M. ericifolia that is completely inhibited at 25 dS m−1 (Ladiges et al. 1981; Robinson et al. 2006).
Seed-desiccation experiment
It is clear from this study that M. quinquenervia seeds have the ability to withstand desiccation of at least once during the critical development stage of the disapore, in this case prior to radicle protrusion. This is not an unusual response for pioneer tree species with no ability of the young seedling to withstand desiccation, and germination after the critical stage being always fatal (Daws et al. 2007). The results of this experiment are similar to those of Battaglia’s (1993) study of Eucalyptus delagatensis in that once seeds begin to rupture their seedcoats, the effects of dehydration were usually fatal.
The most important factor in ecological terms, and as determined from this experiment, is that for germination and initial seedling establishment to be successful, it is crucial that the supply of moisture is continual throughout this period. An interruption can be tolerated within the first few days of water uptake, prior to radicle protrusion, but no later. Optimum conditions for establishment, therefore, would be those that are continually moist. This could generally be satisfied in summer in the fringing forests of the Myall Lakes when the peak summer seed fall corresponds with the time of highest rainfall; the wettest months being late summer and early autumn, thereby ensuring ideal warm, moist conditions for recruitment.
Litter/leachate experiment
This study has suggested that the presence and amount of litter in the field is a likely determinant of both emergence and survival of M. quinquenervia seedlings. Germination is possible under litter from both M. quinquenervia and C. glauca and also under seagrass (Zostera sp.), provided litter density is low enough to allow light to penetrate. However, emergence was greatly suppressed under M. quinquenervia leaves and appeared impossible through the thick, matted layer of seagrass. The implication of these results is that recruitment is more likely to be successful at a site where the M. quinquenervia leaf litter has been cleared, for example, by fire or flood, or perhaps disturbed by the activities of foraging birds and animals, and where stranded seagrass does not accumulate.
However, the layer of Casuarina cladodes did not prevent M. quinquenervia seedling emergence, because the cladodes both let in more light and provided gaps for the seedling to push through. Hence, there was general association of the two species.
The small but significant reduction in germination under C. glauca leachate versus M. quinquenervia leachate and control pots suggests that there is an inhibitory effect produced by the Casuarina leachate. Allelopathy, or the release of compounds that affect the growth or germination of neighbouring species, involves chemicals such as phenolic acids, flavonoids, terpenoids, alkaloids and tannins (Kruse et al. 2000; Singh et al. 2003). Hachani et al. (2019) reported significantly decreased rates of germination, delayed root and shoot elongation and a significant negative correlation between dry mass accumulation in Triticum durum seedlings and increased leachate concentrations from C. glauca. Jacob and Nair (1999) also noticed an inhibitory effect of leaf extract from Casuarina on germination and growth behaviour of rice and cowpea. Such an inhibitory effect may be the explanation for the noticeably reduced understorey vegetation found under Casuarina trees in general. An analysis of leachate content to identify the allelopathic compounds therein and the concentrations at which those compounds become inhibitory, could be promising for areas where M. quinquenervia is a noxious weed.
Survival and mortality from a natural germination event
Many factors were seen to prevent seedling survival up to 12 months following a large natural germination event along the eastern Broadwater in 2006. The main factors causing death were derived from the detailed diary and photographs. These included (1) wave action and rising lake levels washing away seedlings, (2) the effects of salinity, including both inundation and salt spray, and (3) complete burial of seedlings under dense strandlines of seagrass that was dumped onto the lake shore as lake levels rose, mostly in the more saline Broadwater. These three factors combined tended to kill seedlings closest to the water’s edge.
The only area found to be suitable for both germination and seedling establishment was within the sedge vegetation on the sand flats at Neranie Sands. Areas of reed vegetation with their deep, tough rhizomes binding the sands dissipated the strongest forces of incoming waves, thereby protecting the seedlings. The characteristics of the site appear to have been as follows: (1) salinities, although quite high at times, were lower than in other areas of the lakes; (2) a sheltered position, away from the prevailing north-easterly winds during summer when seedlings were young and sensitive to desiccation; (3) a flat terrain that was permanently moist; and (4) sedge vegetation offering some protection from destructive wave action and debris.
Results are consistent with work by Zacks et al. (2018) who reported greater seedling survival and growth of wetland species M. squarrosa and Leptospermum lanigerum under moist but not permanently flooded conditions in both glasshouse experiments and field studies at the Yellingbo wetlands of south-eastern Australia.
Seedling transplantation experiment
In contrast to previous plant translocation studies by de Jong (2000), Howard (1973), Jusaitis (2005) and Osunkoya and Creese (1997), this onsite seedling establishment experiment was not limited to a small number of selected microsites. The establishment of a transect across the entire width of the foreshore zone of four widely separated sites across the National Park, subjected the seedling transplants to a greater range of topographic and environmental conditions, enabling an investigation of a range of factors influencing seedling establishment, growth and survival. Several factors stood out as being important in seedling establishment of M. quinquenervia. These were (1) the availability of moisture, (2) the ability to withstand the physical forces of a lake side, such as wave wash and burial by seagrass, (3) the level of salinity and effects of salt spray, (4) the frequency and duration of inundation, and (5) herbivory and insect attack.
The transect experiment showed clearly that 6 m from the lakes edge at each site is a zone of high mortality, with 6-month-old seedlings planted here exhibiting a low chance of survival. The submergence of seedlings in this zone tended to cause leaf abscission and discolouration, which, when combined with insect attack and herbivory above the water line, retarded growth and, in many cases, was the apparent cause of death. At all sites, the ‘frontline’ seedlings suffered damage and, mostly, eventual death. Salt spray, physical damage to stems and dislodgement by wave action or complete plant burial under a heavy, 200 mm or thicker, strandline layer of seagrass also caused mortality.
Although seedling survival further than 6 m from the lakes edge was generally higher, mortality occurred up to a further 7.2 m at Paperbark Picnic Area and 4.5 m at West Legges where plants were exposed to both higher salinity (up to 17.4 dS m−1 and 21.8 dS m−1 respectively) and choppy wave action owing to south westerly winds. At one site only (Mayers Point), the seedling closest to shore survived, possibly owing to a lower salinity at that site (up to 8.3 dS m−1) and/or the tempering of destructive wave action by the presence of reed vegetation (Phragmites australis and Typha orientalis).
The fastest seedling growth occurred in the areas that experienced intermittent soaking of the soil, but not permanent inundation with salty water, i.e. Mayers Point and the low-lying swampland at West Legges. This is in agreement with Myers (1983) and (White 1988) who found significantly greater plant heights in saturated soil than in well drained soil, but that submergence of seedlings retards growth and eventually causes death. Studies on the success of revegetation with M. ericifolia at wetlands in Victoria, Australia, found significantly lower rates of seedling survival in waterlogged sites, with the best results being obtained from plantings into raised hummocks combined with more mature plantings (Raulings et al. 2007).
The seedlings that survived the inundation in our study were generally taller, with more leaves extending above the water line. van der Moezel et al. (1991) also found the combined effects of salinity and submergence to be detrimental to M. quinquenervia seedling survival; however, they used very high salinity (up to 63 dS m−1) compared with those recorded in our study. A study by White (1988) investigated the influence of salinity on the distribution of M. quinquenervia seedlings and found that increasing salinity inhibited growth, with a marked decrease in seedling height at salinity above 16 dS m−1.
Of note, at Paperbark Picnic Area and One Horse Sands, the foreshore zone is backed by remnant dunes that have a deep sandy profile. Rainfall accumulating in the dune sand would create an underlying, freshwater table in the foreshore zone with a considerable amount of downward pressure towards the lake, as described by Stratford et al. (2013) and Abesser et al. (2017). Hence, it was often the case that when the lake water was quite saline here, the underlying groundwater, often right up to the water’s edge, remained quite fresh. This was no doubt an important factor for initial survival of the foreshore seedlings.
These observations, combined with experimental results from this study, suggest that favourable conditions for seedling establishment could be met, by a receding lake level, for an extended period, such as during a drought, long enough to allow growth of seedlings to a stage where they are able to withstand a subsequent rise in lake level. Alternatively, a severely wet period causing a flood, followed by receding lake levels, may provide suitable conditions for germination on the wet soil, cleared of litter, during drawdown. This would have to coincide with high temperatures suitable for germination. Indeed, this restriction in time and space for seed release and germination during warm periods of wetland drawdown is consistent with studies of other wetland species such as Taxodium distichum (Middleton 2000) and M. ericifolia (Hamilton-Brown et al. 2009).
Conclusions and management recommendations
We have shown that survival during seedling establishment appears to be the limiting process in the recruitment of M. quinquenervia in the fringing forest at the Myall Lakes in NSW, Australia. Whereas germination depends on the synchronisation of warm summer temperatures and peak summer seed supply falling from parent trees, subsequent seedling establishment is likely to occur only where seed lodges in wet surface locations that do not dry out, at least until roots can access groundwater. In addition, seedlings require protection from wave wash, complete inundation and seagrass coverage in the foreshore zone, and lower salinity (<15 dS m−1) are crucial for seedling survival.
Key management recommendations for natural-area managers on the basis of this study are as follows:
-
Maintaining a reduction of wave wash from recreational boating activities near sensitive shoreline areas and establishment of No Wash Zones that exclude powerboats would offer some protection, with buffering vegetation such as sedges and reeds providing suitable microsites for germination.
-
Increasing public awareness of the presence of fragile seedlings may prevent the unnecessary trampling or disturbance of young seedlings on foreshore areas, such as Neranie Sands, where there is evidence of watercraft entering the sedge flats. More visible policing by boating authorities to restrict boating access to some foreshore areas would also assist regeneration.
-
Some public amenities may need to be relocated away from older trees to ensure public safety and conservation of more fecund mature trees.
-
In the cases where existing older trees have been removed for public safety, park management may decide to employ more speedy methods of establishment, rather than relying on natural recruitment. In this case, moderate-scale plantings of local provenance seedlings may be appropriate in the zones where survival was found to be highest. Where sandy, reedy flats do not exist, the ideal planting zone would be in the area back from the edge of the lake, away from the salt spray zone. Consideration should be given to the presence of underlying freshwater tables as a means of moisture supply when selecting locations for newly planted seedlings. If the seedlings were likely to be subject to inundation, chances of survival would increase if they were planted at a fairly established size such as 1 m whips (young trees up to 2 years old).
-
Protection of young seedlings with tree guards, bags or fences is recommended as herbivory from mammals and insects was found to be a major cause of mortality in these studies.
-
In the event of receding floodwaters during summer, it may be possible to find more areas where germination has occurred. These may benefit from protective fencing if they are located within high public-use camping or recreational zones.
-
Continued monitoring of sapling recruitment, lake levels and salinity would add to our understanding of M. quinquenervia requirements and provide useful information for future ecological studies.
Data availability
All data are available from the authors on request.
Conflicts of interest
The authors declare no conflicts of interest.
Declaration of funding
Funding was provided to the University from the NSW National Parks and Wildlife Service, Hunter Region, to investigate the likelihood of natural recruitment from the (mostly ageing) iconic paperbarks of the Myall Lakes National Park, camping grounds.
Acknowledgements
This work was completed while Anne Baumann was a doctoral student at the University of Sydney under primary supervision of Dr Peter Martin. We are grateful to Professor Paul Holford for reproducing all graphs in SigmaPlot and for reviewing early drafts.
References
Abesser C, Clarke D, Hughes AG, Robins NS (2017) Modelling small groundwater systems: experiences from the Braunton Burrows and Ainsdale coastal dune systems, UK. Journal of Coastal Conservation 21, 595–614.| Modelling small groundwater systems: experiences from the Braunton Burrows and Ainsdale coastal dune systems, UK.Crossref | GoogleScholarGoogle Scholar |
Atkinson G, Hutchings P, Johnson M, Johnson WD, Melville MD (1981) An ecological investigation of the Myall Lakes region. Australian Journal of Ecology 6, 299–327.
| An ecological investigation of the Myall Lakes region.Crossref | GoogleScholarGoogle Scholar |
Auld TD, Keith DA, Bradstock RA (2000) Patterns in longevity of soil seedbanks in fire-prone communities of south-eastern Australia. Australian Journal of Botany 48, 539–548.
| Patterns in longevity of soil seedbanks in fire-prone communities of south-eastern Australia.Crossref | GoogleScholarGoogle Scholar |
Baskin CC, Baskin JM (1998) ‘Seeds: ecology, biogeography and evolution of dormancy and germination.’ (Academic Press: San Diego, CA, USA)
Battaglia M (1993) Seed germination physiology of Eucalyptus delegatensis R.T.Baker in Tasmania. Australian Journal of Botany 41, 119–136.
| Seed germination physiology of Eucalyptus delegatensis R.T.Baker in Tasmania.Crossref | GoogleScholarGoogle Scholar |
Baumann A (2008) Regeneration of broad-leaved paperbark trees in the fringing forest of the Myall Lakes. Doctoral thesis, Sydney University, NSW, Australia.
Benson D, McDougall L (1998) Ecology of Sydney plant species Part 6: Dicotyledon family Myrtaceae. Cunninghamia 5, 808–987.
Bewley J, Bradford K, Hilhorst H, Nonogaki H (2013) ‘Seeds: physiology of development, germination and dormancy.’ 3rd edn. (Springer: New York, NY, USA)
Bodle MJ, Ferriter AP, Thayer DD (1994) The biology, distribution, and ecological consequences of Melaleuca quinquenervia in the Everglades. In ‘Everglades: the ecosystem and its restoration’. (Eds SM Davies, JC Ogden) pp. 341–355. (St Lucie Press: Delray Beach, FL, USA)
Boland DJ, Brooker MIH, Chippendale G, Hall N, Hyland B, Johnston R, Kleinig D, McDonald M, Turner J (2006) ‘Forest Trees of Australia.’ (CSIRO Publishing: Melbourne, Vic., Australia)
Brophy JJ, Craven LA, Doran JC (2013) ‘Melaleucas: their botany, essential oils and uses.’ (Australian Centre for International Agricultural Research (ACIAR))
Clewer AG, Scarisbrick DH (2013) ‘Practical statistics and experimental design for plant and crop science.’ (John Wiley & Sons)
Cronk K, Siobhan-Fenessey M (2001) ‘Wetland Plants: biology and ecology.’ (Lewis Publishers: Boca Raton, FL, USA)
Daws MI, Bolton S, Burslem DFRP, Garwood NC, Mullins CE (2007) Loss of desiccation tolerance during germination in neo-tropical pioneer seeds: implications for seed mortality and germination characteristics. Seed Science Research 17, 273–281.
| Loss of desiccation tolerance during germination in neo-tropical pioneer seeds: implications for seed mortality and germination characteristics.Crossref | GoogleScholarGoogle Scholar |
de Jong N (2000) Woody plant restoration and natural regeneration in wet meadow at Coomonderry Swamp on the south coast of New South Wales. Marine and Freshwater Research 51, 81–89.
| Woody plant restoration and natural regeneration in wet meadow at Coomonderry Swamp on the south coast of New South Wales.Crossref | GoogleScholarGoogle Scholar |
Dray FA, Bennett BC, Center TD (2006) Invasion history of Melaleuca quinquenervia (Cav.) ST Blake in Florida. Castanea 71, 210–225.
| Invasion history of Melaleuca quinquenervia (Cav.) ST Blake in Florida.Crossref | GoogleScholarGoogle Scholar |
Eby P, Lunney D (2002) ‘Managing the grey-headed flying-fox Pteropus poliocephalus as a threatened species: a context for the debate.’ (Royal Zoological Society of NSW)
Fenner MK, Fenner M, Thompson K (2005) ‘The ecology of seeds.’ (Cambridge University Press)
Global Invasive Species Database (2021) Melaleuca quinquenervia. Available at www.invasivespecies.net/database [Accessed February 2022]
Great Lakes Council (2009) ‘Great Lakes Water Quality Improvement Plan: Wallis, Smith and Myall Lakes.’ (Great Lakes Council: Forster, NSW, Australia)
Hachani C, Abassi M, Lazhar C, Lamhamedi MS, Béjaoui Z (2019) Allelopathic effects of leachates of Casuarina glauca Sieb. Ex Spreng. and Populus nigra L. on germination and seedling growth of Triticum durum Desf. under laboratory conditions. Agroforestry Systems 93, 1973–1983.
| Allelopathic effects of leachates of Casuarina glauca Sieb. Ex Spreng. and Populus nigra L. on germination and seedling growth of Triticum durum Desf. under laboratory conditions.Crossref | GoogleScholarGoogle Scholar |
Hamilton-Brown S, Boon PI, Raulings E, Morris K, Robinson R (2009) Aerial seed storage in Melaleuca ericifolia Sm. (Swamp Paperbark): environmental triggers for seed release. Hydrobiologia 620, 121–133.
| Aerial seed storage in Melaleuca ericifolia Sm. (Swamp Paperbark): environmental triggers for seed release.Crossref | GoogleScholarGoogle Scholar |
Harper JL (1977) ‘Population biology of plants.’ (Academic Press: London, UK)
Hewitt A, Holford P, Renshaw A, Haigh AM, Morris E (2014) Population structure, seed loads and flowering phenology in three common (Melaleuca styphelioides, M. thymifolia, M. nodosa) and one rare (M. deanei) Melaleuca (Myrtaceae) species of the Sydney region. Australian Journal of Botany 62, 286–304.
| Population structure, seed loads and flowering phenology in three common (Melaleuca styphelioides, M. thymifolia, M. nodosa) and one rare (M. deanei) Melaleuca (Myrtaceae) species of the Sydney region.Crossref | GoogleScholarGoogle Scholar |
Hewitt A, Holford P, Renshaw A, Stone G, Morris EC (2015) Seed size and the regeneration niches of one rare (Melaleuca deanei) and three common (Melaleuca styphelioides, Melaleuca thymifolia and Melaleuca nodosa) Melaleuca (Myrtaceae) species of the Sydney region. Austral Ecology 40, 661–671.
| Seed size and the regeneration niches of one rare (Melaleuca deanei) and three common (Melaleuca styphelioides, Melaleuca thymifolia and Melaleuca nodosa) Melaleuca (Myrtaceae) species of the Sydney region.Crossref | GoogleScholarGoogle Scholar |
Howard TM (1973) Studies in the Ecology of Nothofagus Cunninghamii Oerst. i Natural regeneration on the Mt. Donna Buang Massif, Victoria. Australian Journal of Botany 21, 67–78.
| Studies in the Ecology of Nothofagus Cunninghamii Oerst. i Natural regeneration on the Mt. Donna Buang Massif, Victoria.Crossref | GoogleScholarGoogle Scholar |
International Seed Testing Association (2000) ‘Tetrazolium Testing Handbook.’ (Association of Official Seed Analysts: CA, USA)
Jacob J, Nair AM (1999) Allelopathic effect of leaf leachates of multipurpose trees. Allelopathy Journal 6, 81–86.
Jacobs SWL, Pickard J (1981) ‘Plants of New South Wales: a census of the cycads, conifers and angiosperms.’ (National Herbarium of New South Wales: Royal Botanic Gardens, Sydney, NSW, Australia)
Jenkins ME, Morrison DA, Auld T (2010) Estimating seed bank accumulation and dynamics in three obligate-seeder Proteaceae species. Quantitative Biology 1, 1–15.
Jusaitis M (2005) Translocation trials confirm specific factors affecting the establishment of three endangered plant species. Ecological Management & Restoration 6, 61–67.
| Translocation trials confirm specific factors affecting the establishment of three endangered plant species.Crossref | GoogleScholarGoogle Scholar |
Keith DA (2004) ‘Ocean Shores to Desert Dunes: The Native Vegetation of New South Wales and the ACT.’ (The Department of Environment and Conservation: Hurstville, NSW, Australia)
Kruse M, Strandberg M, Strandberg B (2000) Ecological effects of allelopathic plants – a review. NERI Technical Report No. 315. National Environmental Research Institute, Silkeborg, Denmark.
Ladiges PY, Foord PC, Willis R (1981) Salinity and waterlogging tolerance of some populations of Melaleuca ericifolia Smith. Australian Journal of Ecology 6, 203–215.
| Salinity and waterlogging tolerance of some populations of Melaleuca ericifolia Smith.Crossref | GoogleScholarGoogle Scholar |
Marcar N, Crawford D, Leppert P, Jovanovic T, Floyd R, Farrow R (1995) ‘Trees for saltland: a guide to selecting native species for Australia.’ (CSIRO Publishing: Melbourne, Vic., Australia)
Martin P (2002) Conservation status of Broadleaved Paperbark trees in the lake-fringe forests of the Myall Lakes National Park. Report for NSW National Parks and Wildlife Service.
Martin MR, Tipping PW, Mack MC, Reddy KR (2019) Direct herbivory by biological control agents, and the consequent disruption of plant-nutrient-feedback cycles, combine to reduce the invasiveness of Melaleuca quinquenervia (Myrtaceae). Biological Control 135, 89–94.
| Direct herbivory by biological control agents, and the consequent disruption of plant-nutrient-feedback cycles, combine to reduce the invasiveness of Melaleuca quinquenervia (Myrtaceae).Crossref | GoogleScholarGoogle Scholar |
Matthys C, Jubase N, Visser V, Geerts S (2022) Distribution of Melaleuca rugulosa (Schlechtendal ex Link) Craven (Myrtaceae) in South Africa: assessment of invasiveness and feasibility of eradication. South African Journal of Botany 148, 228–237.
| Distribution of Melaleuca rugulosa (Schlechtendal ex Link) Craven (Myrtaceae) in South Africa: assessment of invasiveness and feasibility of eradication.Crossref | GoogleScholarGoogle Scholar |
McLean RC, Ivimey Cook WR (1941) ‘Plant Science Formulae.’ (Macmillan & Co.: London, UK)
Meskimen G (1962) A silvicultural study of the Melaleuca tree in South Florida. Master of Science. University of Florida.
Mid Coast Council (2018) Waterways and Catchment Report. Mid Coast Council, NSW, Australia.
Middleton B (2000) Hydrochory, seed banks, and regeneration dynamics along the landscape boundaries of a forested wetland. Plant Ecology 146, 167–181.
| Hydrochory, seed banks, and regeneration dynamics along the landscape boundaries of a forested wetland.Crossref | GoogleScholarGoogle Scholar |
Mooney SD, Maltby EL (2006) Two proxy records revealing the late Holocene fire history at a site on the central coast of New South Wales, Australia. Austral Ecology 31, 682–695.
| Two proxy records revealing the late Holocene fire history at a site on the central coast of New South Wales, Australia.Crossref | GoogleScholarGoogle Scholar |
Myers RL (1983) Site susceptibility to invasion by the exotic tree Melaleuca quinquenervia in southern Florida. Journal of Applied Ecology 20, 645–658.
| Site susceptibility to invasion by the exotic tree Melaleuca quinquenervia in southern Florida.Crossref | GoogleScholarGoogle Scholar |
Myerscough PJ, Carolin RC (1986) The vegetation of the Eurunderee sand mass, headlands and previous islands in the Myall Lakes area New South Wales. Cunninghamia 1, 399–466.
NSW Department of Planning Infrastructure and Environment (2021) NPWS Fire History – Wildfires and prescribed burns dataset. Available at https://datasets.seed.nsw.gov.au/dataset/fire-history-wildfires-and-prescribed-burns-1e8b6 [Accessed June 2022]
NSW National Parks and Wildlife Service (2002) Myall Lakes National Park, Little Broughton Island and Stormpetrel Nature Reserves Plan of Management. NSW National Parks and Wildlife Service.
NSW Office of Environment and Heritage (2012) Myall Lakes Ramsar site ecological character description. Office of Environment and Heritage, NSW, Australia.
NSW Scientific Committee (2012) NSW Scientific Committee – final determination ‘Swamp Sclerophyll Forest on Coastal Floodplains of the NSW North Coast, Sydney Basin and South East Corner bioregions’ – Determination to make a minor amendment to Part 3 of Schedule 1 of the Threatened Species Conservation Act. (NSW Scientific Committee)
Osborn T, Robertson RN (1939) A reconnaissance survey of the vegetation of the Myall Lakes. Proceedings of the Linnean Society of New South Wales 64, 279–296.
Osunkoya O, Creese RG (1997) Population structure, spatial pattern and seedling establishment of the grey mangrove, Avicennia marina var. australasica, in New Zealand. Australian Journal of Botany 45, 707–725.
| Population structure, spatial pattern and seedling establishment of the grey mangrove, Avicennia marina var. australasica, in New Zealand.Crossref | GoogleScholarGoogle Scholar |
Ozden E, Light ME, Demir I (2021) Alternating temperatures increase germination and emergence in relation to endogenous hormones and enzyme activities in aubergine seeds. South African Journal of Botany 139, 130–139.
| Alternating temperatures increase germination and emergence in relation to endogenous hormones and enzyme activities in aubergine seeds.Crossref | GoogleScholarGoogle Scholar |
Palmer D, Fredericks D, Smith C, Heggie D (2000) Nutrients from sediments. AGSO Research. Australian Geological Survey Organisation Research Newsletter 33, 1–4.
Probert RJ (2000) The role of temperature in seed dormancy and germination. In ‘The ecology of regeneration in plant communities’, Vol. 2. (Ed. M Fenner) pp. 261–292. (CAB International)
Ralph M (2003) ‘Growing Australian native plants from seed for revegetation, tree planting, and direct seeding.’ (Murray Ralph/Bushland Horticulture)
Ramsar (2021) ‘The List of Wetlands of International Importance.’ (The Secretariat of the Convention on Wetlands: Ramsar, Iran Rue Mauverney Gland, Switzerland)
Raulings EJ, Boon PI, Bailey PC, Roache MC, Morris K, Robinson R (2007) Rehabilitation of Swamp Paperbark (Melaleuca ericifolia) wetlands in south-eastern Australia: effects of hydrology, microtopography, plant age and planting technique on the success of community-based revegetation trials. Wetlands Ecology and Management 15, 175–188.
| Rehabilitation of Swamp Paperbark (Melaleuca ericifolia) wetlands in south-eastern Australia: effects of hydrology, microtopography, plant age and planting technique on the success of community-based revegetation trials.Crossref | GoogleScholarGoogle Scholar |
Rayachhetry MB, Van TK, Center TD (1998) Regeneration potential of the canopy-held seeds of Melaleuca quinquenervia in south Florida. International Journal of Plant Sciences 159, 648–654.
| Regeneration potential of the canopy-held seeds of Melaleuca quinquenervia in south Florida.Crossref | GoogleScholarGoogle Scholar |
Ren J, Tao L (2003) Effect of hydration–dehydration cycles on germination of seven Calligonum species. Journal of Arid Environments 55, 111–122.
| Effect of hydration–dehydration cycles on germination of seven Calligonum species.Crossref | GoogleScholarGoogle Scholar |
Robinson RW, Boon PI, Bailey P (2006) Germination characteristics of Melaleuca ericifolia Sm.(swamp paperbark) and their implications for the rehabilitation of coastal wetlands. Marine and Freshwater Research 57, 703–711.
| Germination characteristics of Melaleuca ericifolia Sm.(swamp paperbark) and their implications for the rehabilitation of coastal wetlands.Crossref | GoogleScholarGoogle Scholar |
Serbesoff-King K (2003) Melaleuca in Florida: a literature review on the taxonomy, distribution, biology, ecology, economic importance and control measures. Journal of Aquatic Plant Management 41, 98–112.
Singh H, Batish D, Kaur S, Kohli R (2003) Phytotoxic interference of Ageratum conyzoides with wheat (Triticum aestivum). Journal of Agronomy and Crop Science 189, 341–346.
| Phytotoxic interference of Ageratum conyzoides with wheat (Triticum aestivum).Crossref | GoogleScholarGoogle Scholar |
Stewart D, Stewart R (1995) ‘From seeds to leaves: a complete guide to growing Australian trees and shrubs from seed.’ (Agmedia: Melbourne, Vic., Australia)
Stratford CJ, Robins NS, Clarke D, Jones L, Weaver G (2013) An ecohydrological review of dune slacks on the west coast of England and Wales. Ecohydrology 6, 162–171.
| An ecohydrological review of dune slacks on the west coast of England and Wales.Crossref | GoogleScholarGoogle Scholar |
Sutherland S (1986) Patterns of fruit-set: what controls fruit–flower ratios in plants? Evolution 40, 117–128.
| Patterns of fruit-set: what controls fruit–flower ratios in plants?Crossref | GoogleScholarGoogle Scholar |
Thompson K, Grime JP (1983) A comparative study of germination responses to diurnally-fluctuating temperatures. Journal of Applied Ecology 20, 141–156.
| A comparative study of germination responses to diurnally-fluctuating temperatures.Crossref | GoogleScholarGoogle Scholar |
Tipping PW, Martin MR, Nimmo KR, Smart MD, Wear EW (2016) Food web associations among generalist predators and biological control agents of Melaleuca quinquenervia. Biological Control 101, 52–58.
| Food web associations among generalist predators and biological control agents of Melaleuca quinquenervia.Crossref | GoogleScholarGoogle Scholar |
Tipping PW, Martin MR, Foley JR, Pierce RM, Gettys LA (2021) The influence of two wildfires and biological control agents on the population dynamics of Melaleuca quinquenervia in a seasonally inundated wetland. Invasive Plant Science and Management 14, 3–8.
| The influence of two wildfires and biological control agents on the population dynamics of Melaleuca quinquenervia in a seasonally inundated wetland.Crossref | GoogleScholarGoogle Scholar |
Turner CE, Center TD, Burrows DW, Buckingham GR (1997) Ecology and management of Melaleuca quinquenervia, an invader of wetlands in Florida, USA. Wetlands Ecology and Management 5, 165–178.
| Ecology and management of Melaleuca quinquenervia, an invader of wetlands in Florida, USA.Crossref | GoogleScholarGoogle Scholar |
Van TK, Rayamajhi MB, Center TD (2005) Seed longevity of Melaleuca quinquenervia: a burial experiment in south Florida. Journal of Aquatic Plant Management 43, 39–42.
Van der Moezel PG, Bell DT (1987) The effect of salinity on the germination of some Western Australian Eucalyptus and Melaleuca species. Seed Science and Technology 15, 239–246.
van der Moezel PG, Pearce-Pinto GVN, Bell DT (1991) Screening for salt and waterlogging tolerance in Eucalyptus and Melaleuca species. Forest Ecology and Management 40, 27–37.
| Screening for salt and waterlogging tolerance in Eucalyptus and Melaleuca species.Crossref | GoogleScholarGoogle Scholar |
van Kleunen M, Dawson W, Essl F, et al. (2015) Global exchange and accumulation of non-native plants. Nature 525, 100–103.
| Global exchange and accumulation of non-native plants.Crossref | GoogleScholarGoogle Scholar |
van Wyk E, Jacobs L (2015) Prospects for extirpating small populations of the wetland invader Melaleuca quinquenervia from South Africa: a case study from the Western Cape region. African Journal of Aquatic Science 40, 299–306.
| Prospects for extirpating small populations of the wetland invader Melaleuca quinquenervia from South Africa: a case study from the Western Cape region.Crossref | GoogleScholarGoogle Scholar |
Welbergen JA, Meade J, Field HE, Edson D, McMichael L, Shoo LP, Praszczalek J, Smith C, Martin JM (2020) Extreme mobility of the world’s largest flying mammals creates key challenges for management and conservation. BMC Biology 18, 101
| Extreme mobility of the world’s largest flying mammals creates key challenges for management and conservation.Crossref | GoogleScholarGoogle Scholar |
White E (1988) The Distribution and population dynamics of Melaleuca quinquenervia (Cav.) ST.Blake., Masters thesis, University of New England, Armidale, NSW, Australia.
Withrow RB, Price L (1957) A darkroom safelight for research in plant physiology. Plant Physiology 32, 244–248.
| A darkroom safelight for research in plant physiology.Crossref | GoogleScholarGoogle Scholar |
Zacks G, Greet J, Walsh CJ, Raulings E (2018) The flooding tolerance of two critical habitat-forming wetland shrubs, Leptospermum lanigerum and Melaleuca squarrosa, at different life history stages. Australian Journal of Botany 66, 500–510.
| The flooding tolerance of two critical habitat-forming wetland shrubs, Leptospermum lanigerum and Melaleuca squarrosa, at different life history stages.Crossref | GoogleScholarGoogle Scholar |
Zoete T, Davie J (2000) Arguing for conservation of wetlands in landscapes with intensifying land use pressures: Melaleuca quinquenervia South-East Queensland. In ‘Conservation in production environments: managing the matrix’. (Eds J Craig, N Mitchell, DA Saunders) pp. 633–643. (Surrey Beatty: Sydney, NSW, Australia)