

Climate change or tree disease: challenges for diagnosing causes of forest die-off
George Matusick A * , Katinka X. Ruthrof A B , Peter Scott C and Giles E. St. J. Hardy A
A * , Katinka X. Ruthrof A B , Peter Scott C and Giles E. St. J. Hardy A
A
B
C
Australian Journal of Botany - https://doi.org/10.1071/BT23039
Submitted: 14 May 2023 Accepted: 10 October 2023 Published online: 6 November 2023
Abstract
Forest ecosystems experience compositional and structural changes as species’ environmental envelopes shift with climate change. Extreme climate events and pests/pathogens are driving these ecosystem changes. Determining which of the two potential drivers is causing a particular forest die-off can be challenging. In south-western Australia, widespread forest die-off in 2011 coincided with extremely hot and dry conditions. It occurred in a forest ecosystem that has historically experienced Phytophthora cinnamomi root disease (Phytophthora dieback).
To determine whether the causal agent of Phytophthora dieback, P. cinnamomi, was associated with forest die-off in the Northern Jarrah Forest.
A combination of direct (isolation of pathogen) and indirect (survey of susceptible indicator plant species) measurements were taken inside and outside patches of forest experiencing the die-off.
There was no consistent association between die-off patches and the presence of P. cinnamomi. P. cinnamomi was isolated from 3 of 33 control plots and 3 of 33 die-off plots. Although several plant species susceptible to P. cinnamomi were absent from die-off plots, the findings were inconsistent across species. This may be explained by plant tolerance to high temperatures and drought.
P. cinnamomi was not the proximate cause of the observed die-off in the Northern Jarrah Forest in 2011.
Novel disturbance caused by extreme climate events can mimic damage caused by certain pests/pathogens. More research is needed to determine the tolerances of plants to extreme temperature and drought conditions to disentangle abiotic and biotic drivers of tree die-off.
Keywords: climate change, drought, eucalypt dieback, Eucalyptus marginata, extreme climate event, jarrah, Phytophthora cinnamomi, Phytophthora dieback.
Introduction
Changes in species’ environmental envelopes with climate change are expected to result in compositional, structural and ecosystem-type shifts in the coming decades in many regions (Anadón et al. 2014; Taylor et al. 2017). The direct effects of climate change and the influence of forest pests and pathogens are expected to be the proximate mechanisms driving these shifts (Beckage et al. 2008; Dukes et al. 2009). Therefore, understanding the relative roles of these two mechanisms is vital for determining the rate and direction ecosystems will shift and developing potential mitigation strategies.
Management of Phytophthora diseases, which are among the most destructive tree diseases worldwide (Hansen 2015; Burgess et al. 2017), is likely to become increasingly complex because of climate change. Soil-borne Phytophthora species rely on soil moisture for movement, reproduction and infection of hosts, making the disease susceptible to climate change-driven moisture changes (La Porta et al. 2008; Sturrock et al. 2011). Whereas some future climate scenarios, including drying, may reduce the severity of Phytophthora diseases in certain regions, other regions may experience an increase in severity of the disease due to optimal conditions for the manifestation of the phenomenon (Aguayo et al. 2014; Homet et al. 2019). Phytophthora diseases may also interact with climate drivers of tree stress to cause greater tree die-off and mortality (Corcobado et al. 2014). Finally, climate change is likely to result in tree damage that is rarely documented, which has the potential to mimic disease symptoms and lead to misdiagnosis. This effect of climate change may be particularly important for root disease caused by Phytophthora species, because disease diagnosis and mapping are commonly symptom-driven (Hardy et al. 2007; Wilson et al. 2012), owing to the high costs and frequent false negatives of isolation and culture methods (Hüberli et al. 2000).
The Mediterranean climate region of south-western Australia provides a model region to highlight the management challenges and ecosystem effects of climate change and introduced Phytophthora pathogens on forest ecosystems. The region has experienced declining precipitation (Timbal et al. 2006; Delworth and Zeng 2014) and increasing temperatures in recent decades (Hope et al. 2015), resulting in significant declines in streamflow (Liu et al. 2019) and groundwater levels (Liu et al. 2019; McFarlane et al. 2020), among other changes. Most ecosystems have also been affected by the introduction of species of the soil-borne water mold, Phytophthora (Shearer et al. 2007; Ritchie et al. 2021). Corresponding with these climate and disease changes, a wide variety of forest and woodland tree species have experienced episodes of tree die-off, crown decline and tree mortality (Hooper and Sivasithamparam 2005; Dakin et al. 2010; Matusick et al. 2012, 2013; Paap et al. 2017; Wentzel et al. 2018; Ritchie et al. 2021). Phytophthora cinnamomi, one of the most destructive plant pathogens worldwide (Burgess et al. 2017; Sena et al. 2018), remains the predominant pathogen threat to ecosystems in south-western Australia (Shearer et al. 2004, 2007), causing significant dieback. It kills plants throughout the South-West Botanical Province (Shearer et al. 2004), including in the Banksia woodland ecosystem (Shearer and Dillon 1996), the hyper-diverse Stirling Range (Wills 1993) and the jarrah forest ecosystem (McDougall et al. 2002). Understorey plant species, especially those in the Proteaceae, Fabaceae and Ericaceae (among others), are especially susceptible to disease. Since its discovery in the jarrah forest ecosystem, P. cinnamomi has been considered the cause of progressive crown dieback, leading to mortality of stands of overstorey jarrah (Eucalyptus marginata) trees (‘Phytophthora dieback’). However, more recently, the direct role of P. cinnamomi in the death of jarrah is under debate (Davison 2015, 2018). Regardless of the role of P. cinnamomi in jarrah’s decline and death, it remains a clear threat to the vegetative biodiversity of the jarrah forest ecosystem. The future sustainable use of the jarrah forest for recreation, conservation and bauxite mining is contingent on the successful management of P. cinnamomi (Colquhoun and Hardy 2000).
Mapping the distribution of P. cinnamomi is the most critical component of Phytophthora dieback management. Various direct and indirect methods have been used to map P. cinnamomi in the jarrah forest (Dell et al. 2005). Research has increased the accuracy and decreased the cost of detecting the pathogen’s presence through direct isolation (Aghighi et al. 2016), and in situ and ex situ baiting using soil and susceptible plant material (Burgess et al. 2021). However, these methods continue to be impractical for mapping large geographic areas or complex patterns of disease distribution and are mostly used strategically to supplement indirect disease mapping methods. The approach most relied on includes using the presence/absence of species known to be susceptible to the disease (Koch 2007). Disease symptoms and pathogen isolation are most common on the ‘dieback front’ (McDougall et al. 2002), and by identifying the presence of symptoms in susceptible species, skilled ‘dieback interpreters’ develop maps of the leading edge of infested patches. Then, the disease can often be effectively contained by avoiding the movement of infested soil and plant material from infested to not infested areas (Dunstan et al. 2010). Therefore, a thorough understanding of the stressors and potential causes of plant symptom development is necessary for accurate P. cinnamomi mapping.
In 2011, extensive areas of the Northern Jarrah Forest (NJF) experienced rapid canopy die-off (Fig. 1) associated with extreme climatic conditions, including a heatwave-compounded drought (Matusick et al. 2013). The die-off was most common on sites containing rocky soils with low water-holding capacity, at high elevations, on steep slopes surrounding rock outcrops, in xeric areas (Brouwers et al. 2013; Andrew et al. 2016), with shallow bedrock and thin soils (McGrath et al. 2023), and most severe on sites experiencing a long-term precipitation decline (Matusick et al. 2018). Forest vegetation surrounding rock outcrops experiences waterlogging during precipitation events, and excessive soil drying during drought (Withers 2000; Schut et al. 2014). These soil conditions, including repeated cycles of waterlogging (root hypoxia), are known to stress jarrah directly (Davison and Tay 1985), as well as potentially increase the P. cinnamomi infection in jarrah roots (Davison and Tay 1987). Increasing the complexity, one of the largest and most common plant species used for mapping P. cinnamomi infestation (Banksia grandis) experienced widespread die-off during the heatwave-compounded drought event in 2011, exhibiting symptoms similar to those of P. cinnamomi infection (Steel et al. 2019). The unprecedented die-off of B. grandis, and other species susceptible to P. cinnamomi, presented the opportunity for confusion and uncertainty when mapping Phytophthora dieback. Whereas the die-off event was strongly associated with extreme climatic conditions, the role of P. cinnamomi remains unclear. Thus, this study aimed to determine whether there was a consistent association between P. cinnamomi presence and areas of the NJF experiencing die-off in 2011.

Methods
Study area
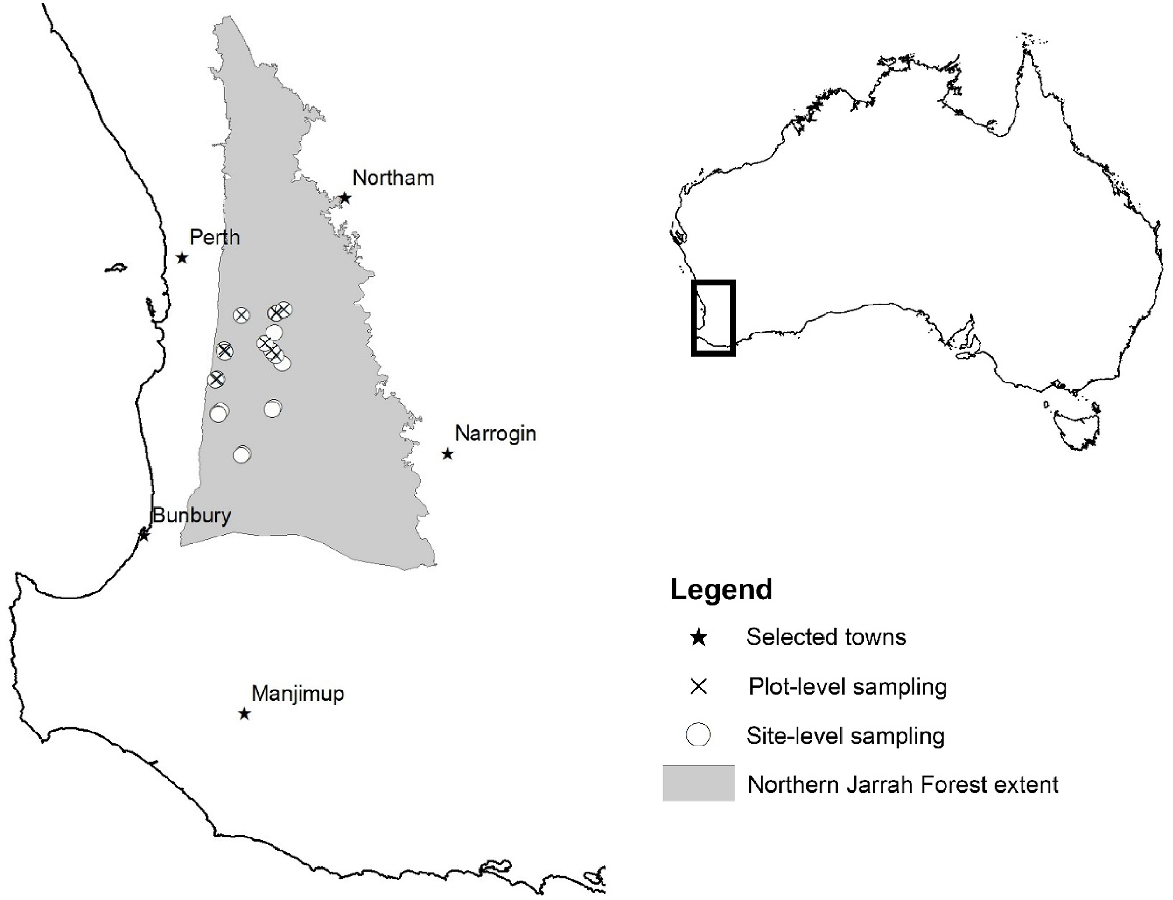
The Northern Jarrah Forest (NJF) occupies 1.1 million ha on the Darling Plateau, a topographic feature that parallels the western coast approximately 20–30 km inland, with undulating topography ranging from ~200 m to ~300 m above sea level (Fig. 2). Several prominent peaks rise above the plateau (~500–600 m above sea level). Most of the NJF has a lateritic duricrust resulting in surface soils dominated by coarse gravels. The bedrock is granitic, which has led to the formation of a clay subsoil occupying the space between the duricrust and granite basement (Churchward and Dimmock 1989). Clay depths vary greatly from non-existent surrounding rock outcrops up to 40 m (Dell et al. 1983). There is a slight north–south temperature gradient within the NJF, and a strong west–east precipitation gradient, historically ranging from ~1200 mm year−1 in the west to ~630 mm year−1 in the east (Gentilli 1989). Corresponding with the relatively good growing conditions in the west, and poorer conditions in the east, the forest structure is significantly denser and taller in the west than the relatively short, open forest in the east. The NJF has historically been used for timber production (Bradshaw 1999). However, since the 1970s, the forest has increasingly been managed for multiple uses, including bauxite mining, recreation and biodiversity conservation.
P. cinnamomi
The soil-borne water mold, P. cinnamomi, was thought to be introduced to south-western Australia prior to 1920. A combination of factors led to the movement and proliferation of the pathogen throughout large portions of the NJF between 1940 and 1970, including a lack of understanding and appreciation for the magnitude of the disease problem and extensive road construction with potentially infested gravel material (Dell and Malajczuk 1989). Today, P. cinnamomi is known to occur throughout the western forest margin and across all topographic positions. Moving inland from the infested escarpment, the disease-affected forest becomes increasingly concentrated in drainage and lower-slope topographic positions, because P. cinnamomi moves readily through the movement of soil water (McDougall et al. 2002).
Site selection
Shortly following the cessation of the forest die-off event in NJF in 2011, 20 symptomatic areas were selected randomly from a larger sample detected during an aerial survey in May 2011 (see Matusick et al. (2013) for details). The boundary of each area affected by die-off, defined as having >70% of trees exhibiting dying or recently killed crowns, was delineated accurately by using a differential GPS. Three points were selected randomly within each die-off patch, representing the centre of a 6-m radius (0.011 ha) circular plot. Because die-off patches were clearly defined, control areas that exhibited minimal crown symptoms could be close to symptomatic ones. Therefore, three control plots were established approximately 20 m from the boundary of the forest experiencing die-off.
Site sampling
Traditionally, the presence and distribution of P. cinnamomi have been determined at the site level in advance of timber harvest or other projects involving soil movement. Site-level sampling provides the greatest possible chance of detecting P. cinnamomi. For each study site (Fig. 2), including the area within and surrounding die-off patches, the presence of P. cinnamomi was evaluated on the basis of the interpreter guidelines (Department of Conservation and Land Management 2001). The guidelines involve assessing the interpretability of the site, detecting and demarcating obvious infested areas and establishing transects through less obvious portions of the site. Samples of symptomatic, susceptible plant species and soil surrounding the plants were collected and baited to detect P. cinnamomi (see below for plot sampling methods). The result of the investigation determined whether P. cinnamomi was present within or near delineated die-off patches.
Plot sampling
In addition to site-level sampling, study plots were also sampled in 11 of the 20 study sites. Plot-level sampling was more random than site-level sampling and was conducted to directly examine the association between vegetation health and P. cinnamomi presence. Each 6-m radius plot was divided into four equal quadrants (north-west, north-east, south-east, south-west). Within each quadrant, two 25 cm × 25 cm × 5 cm soil monoliths were collected. Samples were preferentially taken near symptomatic understorey plants susceptible to P. cinnamomi (examples in Table 1).
S, susceptible; MS, moderately susceptible; HS, highly susceptible; SP, susceptible but persistent.
Phytophthora baiting and isolation
Each soil sample was sifted using a sieve to isolate coarse roots (>2 mm diameter). The sieve was sterilised between the samples each time. A sample of coarse roots and sifted soil was taken from each sample location and bulked for each plot quadrant. This process resulted in four soil samples and four root samples from each plot. Soil and root samples were baited within 2 days of field collection in a baiting container (15 cm long × 5 cm wide × 5 cm deep). Phytophthora species baiting followed a protocol developed by the Centre for Phytophthora Science and Management (CPSM), Murdoch University (Burgess et al. 2021). Sample material was saturated with distilled water for 24 h, followed by complete flooding. Young Quercus spp. (Quercus suber and Quercus ilex) leaves were floated on each baiting tray following flooding. Baits were checked daily for discoloration and symptoms of infection for 20 days. When symptoms were detected, diseased leaf tissue was cut into sections and plated on Phytophthora-selective media (NARPH, Hüberli et al. 2000). Representatives of all Pythiaceae colonies were isolated. However, only P. cinnamomi was identified to species on the basis of morphological characteristics.
Understorey species survey
The presence of native plant species that are known to be highly susceptible to P. cinnamomi in the 6-m study plots suggests that the pathogen may not be present. To examine this indirect evidence for the absence of P. cinnamomi, all understorey and mid-storey species were recorded within each of the 120 plots, spanning all 20 die-off patches, in late winter/spring (June–October 2011). Each of the plants was categorised into P. cinnamomi susceptibility classes on the basis of McDougall (2005) (Table 2). Here, we report results on 10 common plant taxa that are known to be either highly or moderately susceptible to P. cinnamomi infection and mortality (McDougall 2005) (Table 1). A binary logistic regression using SAS software (SAS ver. 9.4, Cary, NC, USA) was implemented to explore the association between plot health (die-off vs control) and presence/health condition of each of the focal 10 species.
| Susceptibility rating | Description | References | |
|---|---|---|---|
| HS: Highly susceptible | Species highly susceptible, i.e. species that are frequently and consistently killed in the wild, following infection by P. cinnamomi, and/or appear to decline or be rare on infested sites | Includes scale categories 10, 11 and 12 of Wills (1993) and Barrett (1996), and groups 3 and 5 of Shearer and Dillon (1995) | |
| MS: Moderately susceptible or variable susceptibility | Species that are often killed following infection by P. cinnamomi in the wild, but many plants that commonly survive | Includes scale categories 7, 8 and 9 of Wills (1993) and Barrett (1996), and group 4 of Shearer and Dillon (1995) | |
| LS: Low susceptibility | Species that are rarely but occasionally found dead on infested sites | Includes scale categories 4, 5 and 6 of Wills (1993) and Barrett (1996), and group 2 of Shearer and Dillon (1995) | |
| S: Susceptible and thought to be affected | Degree of susceptibility not documented | ||
| SP: Susceptible but persistent | Species that are frequently killed following infection by P. cinnamomi in the wild, but which persist on affected sites through effective reproductive strategies | ||
| SV: Variable susceptibility | Plants may be commonly killed on some infested sites but appear unaffected on others – this may be attributable to genetic differences among populations or differences in site characteristics that influence plant responses | ||
| FR: Field resistant or tolerant | Species that appear to be unaffected by P. cinnamomi in the wild when it is present and for which deaths in the field can rarely be associated with infection by P. cinnamomi | Includes scale categories 1, 2 and 3 of Wills (1993) and Barrett (1996), and group 1 of Shearer and Dillon (1995) | |
| Q: Not known to be directly affected | Rarely found on affected sites (and may be affected either directly through infection or changes in habitat) |
Results
By using established site-level interpretation approaches, P. cinnamomi was detected at 3 of 20 sites (Table 3). This included one detection within a patch experiencing canopy die-off and three detections outside patches of canopy die-off.
| Site number | Eucalyptus marginata trees (%) | E. marginata trees dying or recently killed (%) | Phytophthora cinnamomi site-level interpretation (1 = positive detection) | Plot-level P. cinnamomi samplingA | P. cinnamomi detection (%) | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| Control plots | Die-off plots | Control plots | Die-off plots | Within die-off patch | Outside die-off patch | Control plots | Die-off plots | |||
| 1 | 20 | 85 | 6 | 33 | 0 | 0 | 1 | 0 | 0 | |
| 2 | 80 | 89 | 0 | 0 | 1 | 1 | 1 | 33 | 0 | |
| 5 | 77 | 99 | 0 | 89 | 0 | 0 | 1 | 33 | 0 | |
| 27 | 89 | 79 | 6 | 100 | 0 | 0 | 1 | 0 | 0 | |
| 48 | 71 | 91 | 15 | 90 | 0 | 0 | 1 | 0 | 0 | |
| 59 | 94 | 89 | 0 | 96 | 0 | 0 | 0 | |||
| 74 | 63 | 95 | 0 | 100 | 0 | 0 | 1 | 0 | 0 | |
| 80 | 78 | 66 | 29 | 33 | 0 | 0 | 1 | 33 | 66 | |
| 87 | 100 | 43 | 0 | 60 | 0 | 0 | 0 | |||
| 96 | 65 | 99 | 5 | 67 | 0 | 0 | 0 | |||
| 98 | 60 | 62 | 0 | 92 | 0 | 0 | 0 | |||
| 99 | 78 | 60 | 0 | 95 | 0 | 0 | 0 | |||
| 152 | 73 | 86 | 0 | 53 | 0 | 1 | 0 | |||
| 155 | 41 | 69 | 9 | 49 | 0 | 0 | 0 | |||
| 165 | 66 | 85 | 3 | 82 | 0 | 0 | 1 | 0 | 33 | |
| 166 | 73 | 92 | 12 | 66 | 0 | 0 | 1 | 0 | 0 | |
| 181 | 49 | 71 | 0 | 100 | 0 | 0 | 1 | 0 | 0 | |
| 183 | 92 | 61 | 11 | 33 | 0 | 0 | 1 | 0 | 0 | |
| 232 | 100 | 95 | 0 | 32 | 0 | 0 | 0 | |||
| 240 | 77 | 98 | 0 | 88 | 0 | 1 | 0 | |||
Of the 11 sites where P. cinnamomi was assessed at the plot level via isolation, P. cinnamomi was detected at four sites. At a plot level, P. cinnamomi was found at 3 of 33 plots in control forest, not experiencing die-off, and at 3 of 33 plots in forest experiencing die-off. P. cinnamomi was found using both site-level and plot-level methods on one site. In total, including both site- and plot-level methods of detection, P. cinnamomi was detected at 6 of the 20 total study sites. There were situations where plot-level sampling detected P. cinnamomi and site-level interpretation did not (Sites 80 and 165).
Plot health (die-off vs control) contributed significantly to explaining the presence of 2 of 10 plant species known to be susceptible to P. cinnamomi (Table 4). Adenanthos barbiger and B. grandis were more likely to occur in control plots than in plots experiencing die-off. Other species had similar trends, including Persoonia longifolia and Boronia fastigata, but plot health was not a significant predictor at P = 0.05. Among the sites where P. cinnamomi was confirmed present (either by site- or plot-level methods), the 10 susceptible plant species were found on ~50% of control plots and ~28% of die-off plots.
| Species | Control plots (%) | Die-off plots (%) | X2 | P-value | |
|---|---|---|---|---|---|
| Adenanthos barbiger | 45 | 40 | 8.3835 | 0.0038 | |
| Banksia grandis | 65 | 25 | 12.8024 | 0.0003 | |
| Boronia fastigiata | 30 | 10 | 2.7332 | 0.0983 | |
| Hibbertia amplexicaulis | 65 | 45 | 0.7078 | 0.4002 | |
| Leucopogon capitellatus | 35 | 30 | 0.0577 | 0.8102 | |
| Leucopogon verticillatus | 30 | 30 | 2.1021 | 0.1471 | |
| Persoonia elliptica | 10 | 10 | 0.6755 | 0.4111 | |
| Persoonia longifolia | 80 | 45 | 3.6078 | 0.0575 | |
| Xanthorrhoea gracilis | 70 | 55 | 1.2449 | 0.2645 | |
| Xanthorrhoea preissii | 75 | 90 | 1.681 | 0.1948 |
Discussion
There was no evidence of a consistent association between P. cinnamomi and forest experiencing die-off, which would be required to attribute the die-off to P. cinnamomi infestation. Instead, P. cinnamomi was confirmed on a small subset of sites. These findings are unsurprising because P. cinnamomi is known to infest sites throughout the NJF (McDougall et al. 2002). However, the presence of Phytophthora root pathogens does not automatically imply disease, because specific environmental factors must be aligned with pathogen biology and host susceptibility (Shearer and Crane 2014). From vegetation surveys, there was minimal indirect evidence of the association between P. cinnamomi and forest die-off in 2011. Certain plant species susceptible to P. cinnamomi were found less frequently on die-off plots. However, this pattern was not consistent across all indicator plant species. We consider the observed differences in plant species between sites to represent weak evidence since these differences may be explained by tolerance to drought and heat, which is largely unknown. For example, B. grandis, commonly used for mapping P. cinnamomi infestations because of its high susceptibility, appears less tolerant of drought stress than do other mid-storey species in the NJF from field observations (Matusick et al. 2013; Steel et al. 2019). The low frequency of P. cinnamomi isolation and inconsistent indirect evidence of P. cinnamomi infestation leads us to conclude that P. cinnamomi was not consistently associated with the forest die-off patches and was not the primary cause of the die-off event in the NJF.
Uncertainty in the weeks and months following the onset of die-off in the NJF regarding the cause of the damage highlights an understudied challenge climate change is likely to impose on forest managers. Most research regarding the interaction between climate change and biotic pests of forests focuses on shifts in geographic distributions (Bergot et al. 2004; Ikegami and Jenkins 2018), changes in pest biology (Mitton and Ferrenberg 2012) and host resistance (Cudmore et al. 2010). However, tree and forest die-off can be associated with extreme climatic events and biotic sources of tree stress (Shaw et al. 2005; Navarro-Cerrillo et al. 2022), creating a potential source of uncertainty regarding the proximate cause. Indeed, several widespread forest die-off events have led to conflicting reports of the proximate causes of die-off because of the association with both abiotic and biotic tree stressors (Jurskis and Turner 2002; Close et al. 2009; Negrón et al. 2009; de Sampaio e Paiva Camilo-Alves et al. 2013). When both extreme climate stress and Phytophthora root diseases are present, the cause of die-off may be particularly difficult to diagnose because Phytophthora species can be challenging to isolate consistently from tissues (Tsao 1990; Bowman et al. 2007), and disease may be diagnosed from host symptomology (Spencer 1984; Burgess et al. 2009). Such was the case for the NJF following the die-off in 2011, where climate stress and P. cinnamomi were suspected of causing the tree die-off. This research suggests that determining causation on the basis of symptomology alone may no longer be valid in many parts of the NJF under a warming and drying climate.
The unprecedented response of terrestrial and marine organisms to climatic extremes in south-western Australia (Ruthrof et al. 2018) highlights the need for basic research regarding the tolerance of species to extreme temperatures, especially when combined with drought. During and following the heatwave-compounded drought in the NJF, plants were observed responding in ways that had only ever been associated with P. cinnamomi infection, fueling confusion regarding the cause of the stress. Basic understanding of temperature and moisture tolerances is unknown for most species in south-western Australia and other regions experiencing similar climate extremes (Breshears et al. 2021). Filling the knowledge gap in species tolerances to extreme temperatures and moisture regimes will be important for interpreting the causes of novel disturbance in the future (Tesky et al. 2015). Once a library of stress tolerances among predominant plant species in different ecosystems is developed, this information could be compiled and disseminated in extension publications and field guides to assist professionals in discriminating among potential stress agents.
In summary, here we have highlighted one understudied phenomenon likely to occur with greater frequency under continued climate change, the challenge of diagnosing proximate causes of novel disturbances. This problem is expected to be particularly acute when biotic stress agents and abiotic sources of stress are both present prior to die-off, and when Phytophthora root pathogens are suspected. In the example reported here, a collection of direct and indirect evidence was analysed to conclude that P. cinnamomi was not the proximate cause of the observed die-off in the NJF. South-western Australia can be seen as a climate-change hotspot (Andrich and Imberger 2013; Foster et al. 2014), experiencing extreme effects of climate change, and can therefore highlight the novel challenges imposed by a shifting climate.
Declaration of funding
This research was conducted as part of the Western Australian State Centre of Excellence for Climate Change and Woodland and Forest Health, a partnership between private industry, community groups, Universities and the Government of Western Australia.
Acknowledgements
The authors thank the Western Australian Department of Biodiversity, Conservation and Attractions for accessing field sites, and Treena Burgess, Cielito Marbus and Bill Dunstan (Murdoch University) for assistance in collecting field samples and interpreting Phytophthora dieback on study sites.
References
Aghighi S, Burgess TI, Scott JK, Calver M, Hardy GESJ (2016) Isolation and pathogenicity of Phytophthora species from declining Rubus anglocandicans. Plant Pathology 65, 451-461.
| Crossref | Google Scholar |
Aguayo J, Elegbede F, Husson C, Saintonge F-X, Marçais B (2014) Modeling climate impact on an emerging disease, the Phytophthora alni-induced alder decline. Global Change Biology 20, 3209-3221.
| Crossref | Google Scholar | PubMed |
Anadón JD, Sala OE, Maestre FT (2014) Climate change will increase savannas at the expense of forests and treeless vegetation in tropical and subtropical Americas. Journal of Ecology 102, 1363-1373.
| Crossref | Google Scholar |
Andrew ME, Ruthrof KX, Matusick G, Hardy GESJ (2016) Spatial configuration of drought disturbance and forest gap creation across environmental gradients. PLoS ONE 11, e0157154.
| Crossref | Google Scholar | PubMed |
Andrich MA, Imberger J (2013) The effect of land-water contrast on the sensitivity of the regional climate of south-west Western Australia to changes in background climate conditions. Climate Dynamics 41, 875-895.
| Google Scholar |
Beckage B, Osborne B, Gavin DG, Pucko C, Siccama T, Perkins T (2008) A rapid upward shift of a forest ecotone during 40 years of warming in the Green Mountains of Vermont. Proceedings of the National Academy of Sciences 105, 4197-4202.
| Crossref | Google Scholar |
Bergot M, Cloppet E, Pérarnaud V, Déqué M, Marçais B, Desprez-Loustau M-L (2004) Simulation of potential range expansion of oak disease caused by Phytophthora cinnamomi under climate change. Global Change Biology 10, 1539-1552.
| Crossref | Google Scholar |
Bowman KD, Albrecht U, Graham JH, Bright DB (2007) Detection of Phytophthora nicotianae and P. palmivora in citrus roots using PCR–RFLP in comparison with other methods. European Journal of Plant Pathology 119, 143-158.
| Crossref | Google Scholar |
Bradshaw FJ (1999) Trends in silvicultural practices in the native forests of Western Australia. Australian Forestry 62, 255-264.
| Crossref | Google Scholar |
Breshears DD, Fontaine JB, Ruthrof KX, Field JP, Feng X, Burger JR, Law DJ, Kala J, Hardy GESJ (2021) Underappreciated plant vulnerabilities to heat waves. New Phytologist 231, 32-39.
| Crossref | Google Scholar | PubMed |
Brouwers N, Matusick G, Ruthrof K, Lyons T, Hardy G (2013) Landscape-scale assessment of tree crown dieback following extreme drought and heat in a Mediterranean eucalypt forest ecosystem. Landscape Ecology 28, 69-80.
| Crossref | Google Scholar |
Burgess TI, Webster JL, Ciampini JA, White D, Hardy GESJ, Stukely MJC (2009) Re-evaluation of Phytophthora species isolated during 30 years of vegetation health surveys in Western Australia using molecular techniques. Plant Disease 93, 215-223.
| Crossref | Google Scholar | PubMed |
Burgess TI, Scott JK, McDougall KL, Stukely MJC, Crane C, Dunstan WA, Brigg F, Andjic V, White D, Rudman T, Arentz F, Ota N, Hardy GESJ (2017) Current and projected global distribution of Phytophthora cinnamomi, one of the world’s worst plant pathogens. Global Change Biology 23, 1661-1674.
| Crossref | Google Scholar | PubMed |
Burgess TI, López-Villamor A, Paap T, Williams B, Belhaj R, Crone M, Dunstan W, Howard K, Hardy GESJ (2021) Towards a best practice methodology for the detection of Phytophthora species in soils. Plant Pathology 70, 604-614.
| Crossref | Google Scholar |
Cho JJ (1983) Variability in susceptibility of some Banksia species to Phytophthora cinnamomi and their distribution in Australia. Plant Disease 67, 869-871.
| Crossref | Google Scholar |
Close DC, Davidson NJ, Johnson DW, Abrams MD, Hart SC, Lunt ID, Archibald RD, Horton B, Adams MA (2009) Premature decline of Eucalyptus and altered ecosystem processes in the absence of fire in some Australian forests. The Botanical Review 75, 191-202.
| Crossref | Google Scholar |
Colquhoun IJ, Hardy GESJ (2000) Managing the risks of Phytophthora root and collar rot during bauxite mining in the Eucalyptus marginata (Jarrah) forest of Western Australia. Plant Disease 84, 116-127.
| Crossref | Google Scholar | PubMed |
Corcobado T, Cubera E, Juárez E, Moreno G, Solla A (2014) Drought events determine performance of Quercus ilex seedlings and increase their susceptibility to Phytophthora cinnamomi. Agricultural and Forest Meteorology 192-193, 1-8.
| Crossref | Google Scholar |
Cudmore TJ, Björklund N, Carroll AL, Staffan Lindgren B (2010) Climate change and range expansion of an aggressive bark beetle: evidence of higher beetle reproduction in naïve host tree populations. Journal of Applied Ecology 47, 1036-1043.
| Crossref | Google Scholar |
Dakin N, White D, Hardy GESJ, Burgess TI (2010) The opportunistic pathogen, Neofusicoccum australe, is response for crown dieback of peppermint (Agonis flexuosa) in Western Australia. Australasian Plant Pathology 39, 202-206.
| Crossref | Google Scholar |
Davison EM (2015) How Phytophthora cinnamomi became associated with the death of Eucalyptus marginata – the early investigations into jarrah dieback. Australasian Plant Pathology 44, 263-271.
| Crossref | Google Scholar |
Davison EM (2018) Relative importance of site, weather and Phytophthora cinnamomi in the decline and death of Eucalyptus marginata – jarrah dieback investigations in the 1970s to 1990s. Australasian Plant Pathology 47, 245-257.
| Crossref | Google Scholar |
Davison EM, Tay FCS (1985) The effect of waterlogging on seedlings of Eucalyptus marginata. New Phytologist 101, 743-753.
| Crossref | Google Scholar |
Davison EM, Tay FCS (1987) The effect of waterlogging on infection of Eucalyptus marginata seedlings by Phytophthora cinnamomi. New Phytologist 105, 585-594.
| Crossref | Google Scholar |
de Sampaio e Paiva Camilo-Alves C, da Clara MIE, de Almeida Ribeiro NMC (2013) Decline of Mediterranean oak trees and its association with Phytophthora cinnamomi: a review. European Journal of Forest Research 132, 411-432.
| Crossref | Google Scholar |
Dell B, Bartle JR, Tacey WH (1983) Root occupation and root channels of jarrah forest subsoils. Australian Journal of Botany 31(6), 615-627.
| Crossref | Google Scholar |
Dell B, Hardy GESJ, Vear K (2005) History of Phytophthora cinnamomi management in Western Australia. In ‘A Forest Conscienceness: Proceedings 6th National Conference of the Australian Forest History Society’. (Eds MC Calver, H Bigler-Cole, G Bolton, J Dargavel, A Gaynor, P Horwitz, J Mills, G Wardell-Johnston) pp. 391–406. (Millpress Science Publishers: Rotterdam, Netherlands)
Delworth TL, Zeng F (2014) Regional rainfall decline in Australia attributed to anthropogenic greenhouse gases and ozone levels. Nature Geoscience 7, 583-587.
| Crossref | Google Scholar |
Dukes JS, Pontius J, Orwig D, Garnas JR, Rodgers VL, Brazee N, Cooke B, Theoharides KA, Stange EE, Harrington R, Ehrenfeld J, Gurevitch J, Lerdau M, Stinson K, Wick R, Ayres M (2009) Responses of insect pests, pathogens, and invasive plant species to climate change in the forests of northeastern North America: what can we predict? Canadian Journal of Forest Research 39, 231-248.
| Crossref | Google Scholar |
Dunstan WA, Rudman T, Shearer BL, Moore NA, Paap T, Calver MC, Dell B, Hardy GESJ (2010) Containment and spot eradication of a highly destructive, invasive plant pathogen (Phytophthora cinnamomi) in natural ecosystems. Biological Invasions 12, 913-925.
| Crossref | Google Scholar |
Foster SD, Griffin DA, Dunstan PK (2014) Twenty years of high-resolution sea surface temperature imagery around Australia: inter-annual and annual variability. PLoS ONE 9, e100762.
| Crossref | Google Scholar |
Groves E, Hollick P, Hardy G, McComb J (2009) Appendix 2 Western Australian natives susceptible to Phytophthora cinnamomi. Murdoch University, WA, Australia. Available at https://www.cpsm-phytophthora.org/downloads/natives_susceptible.pdf [Accessed 28 October 2023]
Hansen EM (2015) Phytophthora species emerging as pathogens of forest trees. Current Forestry Reports 1, 16-24.
| Crossref | Google Scholar |
Hardy GEStJ, Vear K, O’Gara E, Williams NM (2007) Detection, diagnosis and mapping of native areas infested by Phytophthora species in Western Australia. Brazilian Phytopathology 32, S45-S46.
| Google Scholar |
Hill TCJ (1990) Dieback diseases and other Phytophthora spp. in the Northern Kwongon. In ‘Nature, conservation, landscape and recreational values of the Lesueur area. A report to the Environmental Protection Authority from the Department of Conservation and Land Management’. (Eds A Burbidge, SD Hopper, S van Leeuwen) pp. 89–97. (EPA: WA, Australia)
Homet P, González M, Matías L, Godoy O, Pérez-Ramos IM, García LV, Gómez-Aparicio L (2019) Exploring interactive effects of climate change and exotic pathogens on Quercus suber performance: damage caused by Phytophthora cinnamomi varies across contrasting scenarios of soil moisture. Agricultural and Forest Meteorology 276-277, 107605.
| Crossref | Google Scholar |
Hooper RJ, Sivasithamparam K (2005) Characterization of damage and biotic factors associated with the decline of Eucalyptus wandoo in southwest Western Australia. Canadian Journal of Forest Research 35, 2589-2602.
| Crossref | Google Scholar |
Hope P, Abbs D, Bhend J, Chiew F, Church J, Ekström M, Kirono D, Lenton A, Lucas C, McInnes K, Moise A, Monselesan D, Mpelasoka F, Timbal B, Webb L, Whetton P (2015) Southern and south-western flatlands cluster report, climate change in Australia projections for Australia’s natural resource management regions: cluster reports. CSIRO and Bureau of Meteorology, Australia.
Hüberli D, Tommerup IC, Hardy GESJ (2000) False-negative isolations or absence of lesions may cause mis-diagnosis of diseased plants infected with Phytophthora cinnamomi. Australasian Plant Pathology 29, 164-169.
| Crossref | Google Scholar |
Ikegami M, Jenkins TAR (2018) Estimate global risks of a forest disease under current and future climates using species distribution model and simple thermal model – Pine Wilt disease as a model case. Forest Ecology and Management 409, 343-352.
| Crossref | Google Scholar |
Jurskis V, Turner J (2002) Eucalypt dieback in eastern Australia: a simple model. Australian Forestry 65, 87-98.
| Crossref | Google Scholar |
Koch JM (2007) Alcoa’s mining and restoration process in south western Australia. Restoration Ecology 15, S11-S16.
| Crossref | Google Scholar |
La Porta N, Capretti P, Thomsen IM, Kasanen R, Hietala AM, Von Weissenberg K (2008) Forest pathogens with higher damage potential due to climate change in Europe. Canadian Journal of Plant Pathology 30, 177-195.
| Crossref | Google Scholar |
Liu N, Harper RJ, Smettem KRJ, Dell B, Liu S (2019) Responses of streamflow to vegetation and climate change in southwestern Australia. Journal of Hydrology 572, 761-770.
| Crossref | Google Scholar |
Matusick G, Ruthrof KX, Hardy GSJ (2012) Drought and heat triggers sudden and severe dieback in a dominant mediterranean-type woodland species. Open Journal of Forestry 2, 183-186.
| Crossref | Google Scholar |
Matusick G, Ruthrof KX, Brouwers NC, Dell B, Hardy GSJ (2013) Sudden forest canopy collapse corresponding with extreme drought and heat in a mediterranean-type eucalypt forest in southwestern Australia. European Journal of Forest Research 132, 497-510.
| Crossref | Google Scholar |
Matusick G, Ruthrof KX, Kala J, Brouwers NC, Breshears DD, Hardy GESJ (2018) Chronic historical drought legacy exacerbates tree mortality and crown dieback during acute heatwave-compounded drought. Environmental Research Letters 13, 095002.
| Crossref | Google Scholar |
McCredie TA, Dixon KW, Sivasithamparam K (1985) Variability in the resistance of Banksia L. f. species to Phytophthora cinnamomi Rands. Australian Journal of Botany 33, 629-637.
| Crossref | Google Scholar |
McDougall KL, Hardy GESJ, Hobbs RJ (2002) Distribution of Phytophthora cinnamomi in the northern jarrah (Eucalyptus marginata) forest of Western Australia in relation to dieback age and topography. Australian Journal of Botany 50, 107-114.
| Crossref | Google Scholar |
McDougall KL (2005) The responses of native Australian plant species to Phytophthora cinnamomi. Appendix 4. In ‘Management of Phytophthora cinnamomi for biodiversity conservation in Australia: Part 2. National best practice’. (Eds E O’Gara, K Howard, B Wilson, GESJ Hardy) pp. 1–52. (Department of the Environment and Heritage: Canberra, ACT, Australia)
McFarlane D, George R, Ruprecht J, Charles S, Hodgson G (2020) Runoff and groundwater responses to climate change in South West Australia. Journal of the Royal Society of Western Australia 103, 9-27.
| Google Scholar |
McGrath G, Fontaine JB, Van Dongen R, Hyde J, Leopold M, Matusick G, Ruthrof KX (2023) Geophysics reveals forest vulnerability to drought. Ecohydrology
| Crossref | Google Scholar |
Mitton JB, Ferrenberg SM (2012) Mountain pine beetle develops an unprecedented summer generation in response to climate warming. The American Naturalist 179, E163-E171.
| Crossref | Google Scholar | PubMed |
Navarro-Cerrillo RM, González-Moreno P, Ruiz-Gómez FJ, Sánchez-Cuesta R, Gazol A, Camarero JJ (2022) Drought stress and pests increase defoliation and mortality rates in vulnerable Abies pinsapo forests. Forest Ecology and Management 504, 119824.
| Crossref | Google Scholar |
Negrón JF, McMillin JD, Anhold JA, Coulson D (2009) Bark beetle-caused mortality in a drought-affected ponderosa pine landscape in Arizona, USA. Forest Ecology and Management 257, 1353-1362.
| Crossref | Google Scholar |
Paap T, Brouwers NC, Burgess TI, Hardy GESJ (2017) Importance of climate, anthropogenic disturbance and pathogens (Quambalaria coyrecup and Phytophthora spp.) on marri (Corymbia calophylla) tree health in southwest Western Australia. Annals of Forest Science 74, 62.
| Crossref | Google Scholar |
Podger FD (1972) Phytophthora cinnamomi, a cause of lethal disease in indigenous plant communities in Western Australia. Phytopathology 62, 972-981.
| Crossref | Google Scholar |
Ritchie AL, Svejcar LN, Ayre BM, Bolleter J, Brace A, Craig MD, Davis B, Davis RA, van Etten EJB, Fontaine JB, Fowler WM, Froend RH, Groom C, Hardy GESJ, Hooper P, Hopkins AJM, Hughes M, Krauss SL, Leopold M, Miller BP, Miller RG, Ramalho CE, Ruthrof KX, Shaw C, Stevens JC, Tangney R, Valentine LE, Veneklaas EJ, Hobbs RJ (2021) A threatened ecological community: research advances and priorities for Banksia woodlands. Australian Journal of Botany 69, 53-84.
| Crossref | Google Scholar |
Ruthrof KX, Breshears DD, Fontaine JB, Froend RH, Matusick G, Kala J, Miller BP, Mitchell PJ, Wilson SK, van Keulen M, Enright NJ, Law DJ, Wernberg T, Hardy GESJ (2018) Subcontinental heat wave triggers terrestrial and marine, multi-taxa responses. Scientific Reports 8, 13094.
| Crossref | Google Scholar | PubMed |
Schut AGT, Wardell-Johnson GW, Yates CJ, Keppel G, Baran I, Franklin SE, Hopper SD, Van Niel KP, Mucina L, Byrne M (2014) Rapid characterization of vegetation structure to predict refugia and climate change impacts across a global biodiversity hotspot. PLoS ONE 9, e82778.
| Crossref | Google Scholar | PubMed |
Sena K, Crocker E, Vincelli P, Barton C (2018) Phytophthora cinnamomi as a driver of forest change: implications for conservation and management. Forest Ecology and Management 409, 799-807.
| Crossref | Google Scholar |
Shaw JD, Steed BE, DeBlander LT (2005) Forest inventory and analysis (FIA) annual inventory answers the question: what is happening to Pinyon–Juniper woodlands? Journal of Forestry 103, 280-285.
| Google Scholar |
Shearer BL, Crane CE (2014) Phytophthora cinnamomi disease expression and habitat suitability of soils on a topographic gradient across a coastal plain from dunes to forested peneplain. Australasian Plant Pathology 43, 131-142.
| Crossref | Google Scholar |
Shearer BL, Dillon M (1995) Susceptibility of plant species in Eucalyptus marginata forest to infection by Phytophthora cinnamomi. Australian Journal of Botany 43, 113-134.
| Crossref | Google Scholar |
Shearer BL, Dillon M (1996) Impact and disease centre characteristics of Phytophthora cinnamomi infestations of Banksia woodlands on the Swan Coastal Plain, Western Australia. Australian Journal of Botany 44, 79-90.
| Crossref | Google Scholar |
Shearer BL, Crane CE, Cochrane A (2004) Quantification of the susceptibility of the native flora of the South-West Botanical Province, Western Australia, to Phytophthora cinnamomi. Australian Journal of Botany 52, 435-443.
| Crossref | Google Scholar |
Shearer BL, Crane CE, Barrett S, Cochrane A (2007) Phytophthora cinnamomi invasion, a major threatening process to conservation of flora diversity in the South-west Botanical Province of Western Australia. Australian Journal of Botany 55, 225-238.
| Crossref | Google Scholar |
Spencer RD (1984) Detecting dieback in native eucalypt forests using aerial photographs. Australian Forestry 47, 243-249.
| Crossref | Google Scholar |
Steel EJ, Fontaine JB, Ruthrof KX, Burgess TI, Hardy GESJ (2019) Changes in structure of over- and midstory tree species in a Mediterranean-type forest after an extreme drought-associated heatwave. Austral Ecology 44, 1438-1450.
| Crossref | Google Scholar |
Sturrock RN, Frankel SJ, Brown AV, Hennon PE, Kliejunas JT, Lewis KJ, Worrall JJ, Woods AJ (2011) Climate change and forest diseases. Plant Pathology 60, 133-149.
| Crossref | Google Scholar |
Taylor AR, Boulanger Y, Price DT, Cyr D, McGarrigle E, Rammer W, Kershaw JA, Jr. (2017) Rapid 21st century climate change projected to shift composition and growth of Canada’s Acadian Forest Region. Forest Ecology and Management 405, 284-294.
| Crossref | Google Scholar |
Tesky R, Wertin T, Bauweraerts I, Ameye M, McGuire MA, Steppe K (2015) Responses of tree species to heat waves and extreme heat events. Plant, Cell & Environment 38, 1699-1712.
| Crossref | Google Scholar |
Timbal B, Arblaster JM, Power S (2006) Attribution of the late-twentieth-century rainfall decline in southwest Australia. Journal of Climate 19, 2046-2062.
| Crossref | Google Scholar |
Titze JF, Palzer CR (1969) Host list of Phytophthora cinnamomi Rands with special reference to Western Australia. Forestry and Timber Bureau, Department of National Development. Technical. Note No. 1.
| Google Scholar |
Tsao PH (1990) Why many phytophthora root rots and crown rots of tree and horticultural crops remain undetected. EPPO Bulletin 20, 11-17.
| Crossref | Google Scholar |
Wentzel JJ, Craig MD, Barber PA, Hardy GESJ, Fleming PA (2018) Tuart (Eucalyptus gomphocephala) decline is not associated with other vegetation structure and composition changes. Australasian Plant Pathology 47, 521-530.
| Crossref | Google Scholar |
Wills RT (1993) The ecological impact of Phytophthora cinnamomi in the Stirling Range National Park, Western Australia. Australian Journal of Ecology 18, 145-159.
| Crossref | Google Scholar |
Wilson BA, Zdunic K, Kinloch J, Behn G (2012) Use of remote sensing to map occurrence and spread of Phytophthora cinnamomi in Banksia woodlands on the Gnangara Groundwater System, Western Australia. Australian Journal of Botany 60, 495-505.
| Crossref | Google Scholar |
Withers PC (2000) Overview of granite outcrops in Western Australia. Journal of the Royal Society of Western Australia 83, 103-108.
| Google Scholar |