

Influence of moult and location on patterns of daily movement by Egyptian Geese in South Africa
Mduduzi Ndlovu A B , Graeme S. Cumming A and Philip A. R. Hockey A CA Percy FitzPatrick Institute of African Ornithology, University of Cape Town, Rondebosch 7701, South Africa.
B Corresponding author. Email: mduduzindlovu@gmail.com
C Deceased.
Emu 114(1) 23-29 https://doi.org/10.1071/MU12108
Submitted: 22 November 2012 Accepted: 20 March 2013 Published: 2 August 2013
Abstract
The daily movements of 18 Egyptian Geese (Alopochen aegyptiaca) in South Africa were recorded using satellite telemetry. General additive mixed models were used to explain the distances moved by the birds at different times of the day, correcting for sex, site, season and individual. Distances moved by birds in the mesic, winter-rainfall, south-western region (Strandfontein) of South Africa were compared with movements in the semi-arid, summer-rainfall region (Barberspan) of northern South Africa. Moulting birds all remained at single wetlands during moult, displayed crepuscular patterns of activity and nocturnal retreats to safety. At the end of moult, birds dispersed away from moulting sites. The longest daily distances travelled by individuals took place mainly just before moult was due to start or within the first 10 days after completing moult, providing strong evidence that Egyptian Geese undertake moult migrations. The daily pattern of movement was strongly skewed, with >70% of flights being short (<10 km) and longer flights mainly occurring at night (88%). There were considerable differences in patterns of dispersal between birds at the summer-rainfall site and the winter-rainfall one. Summer-rainfall birds undertook long-distance movements between wetlands whereas Winter-rainfall birds appeared to move randomly and over short distances.
Additional keywords: Afrotropical region, telemetry, waterfowl.
Introduction
Movements of waterfowl at different scales are fundamental to individual fitness and population dynamics (Levin 1974; Fauchald and Tveraa 2006). Local movements of waterfowl enable individuals to search for food, mates and escape predators, whereas longer movements are important in avoiding interspecific competition and locating breeding and moulting sites. These movements arise from behavioural decisions by individuals in response to environmental heterogeneity at multiple spatial and temporal scales (Pyke 1984; Zollner and Lima 1999; Morales and Ellner 2002), internal states, life-history demands and the history of the individual bird (Nathan et al. 2008). Despite the relevance of movements to fitness strategies, however, there are few fine-scale studies that follow movements of individual waterfowl over long periods of time, although Roshier et al. (2006) conducted a study of daily movements of Grey Teal (Anas gracilis) over 15 months in Australia. Most previous studies of waterfowl movement have focussed on a particular biological context, such as dispersal (e.g. Oatley and Prŷs-Jones 1986; Underhill et al. 1999), foraging (e.g. Petrie and Rogers 1997) or migration (e.g. Gudmundsson et al. 1995; Dugger 1997; Miller et al. 2005).
Until recently, our understanding of patterns of waterfowl movements and use of habitat was limited largely by our inability to track individual birds for long periods of time over large geographical areas (Lindberg and Walker 2007). Since the inception of small, lightweight satellite transmitters, the movement patterns of waterfowl in the northern hemisphere are increasingly being explored (e.g. Gudmundsson et al. 1995; Dugger 1997; Miller et al. 2005).
Most movement studies of waterfowl have been of long-distance migrants and few studies have focussed on Afrotropical waterfowl. Although there are ringing returns that describe movements of African waterfowl between wetlands (Oatley and Prŷs-Jones 1986; Underhill et al. 1999), the fine-scale daily movements of waterfowl at different times of the year are undescribed. Petrie and Rogers (1997), in the first telemetry study on waterfowl in Africa, described the movement patterns of White-faced Whistling-Ducks (Dendrocygna viduata) from Nelspruit in South Africa. The study had a sample size of only two, the transmitters functioned for <8 months, and the authors did not evaluate daily movements. More recently, Cappelle et al. (2011) used satellite telemetry to study the movement patterns of Garganey (Anas querquedula), Comb Duck (Sarkidiornis melanotos), White-faced Whistling-Duck and Fulvous Whistling-Duck (Dendrocygna bicolor) in parts of central and West Africa in order to understand pathways of influenza transmission, and Cumming et al. (2012) have analysed the seasonal movements of Egyptian Geese and Red-billed Teal (Anas erythrorhyncha). Neither of these two studies, however, evaluated daily movements.
Most studies of movements to date have focussed on movement of waterfowl between breeding sites (e.g. Seymour 1991; Milstein 1993) and non-breeding sites (Robertson and Cooke 1999). Less is known about moult migrations (Salomonsen 1968; Anderson et al. 1992; Bollinger and Derksen 1996) and movements, if any, during moult (Fleskes et al. 2010). Further, most studies of moult migrations among waterfowl come from migratory northern hemisphere species, such as female Mallards (Anas platyrhynchos) (Fleskes et al. 2010), Gadwall (A. strepera) (Szymczak and Rexstad 1991) and American Black Duck (A. rubripes) (Bowman and Brown 1992). In South Africa, it is well known that more than 70% of the South African Shelduck (Tadorna cana) population moults at only 23 locations (Geldenhuys 1981) but movement to and from moulting sites, and that of flightless moulting birds, have not been quantified.
As a first contribution to a more detailed understanding of the patterns of daily movements of an Afrotropical duck, data for Egyptian Geese were analysed during the (1) flightless moulting period and (2) flying periods in two different rainfall regions (summer and winter) of South Africa.
Materials and methods
Study sites
The study was conducted at Barberspan Nature Reserve (26°33′S, 25°37′E) (hereafter referred to as Barberspan) in the North West Province, and at Strandfontein Sewage Works (34°05′S, 18°32′E) (hereafter referred to as Strandfontein) in the Western Cape Province, South Africa.
Barberspan is in north-western South Africa, in a transitional area between the grassland and savanna biomes. The north-western region of South Africa is semi-arid, receives summer rainfall and consists of undulating plains rising to 900 m above sea level. The area is characterised by large depressions, or pans, where rainfall collects during the wet summer season. Barberspan Nature Reserve is surrounded by crop and sunflower farms in a vast grasslands biome (Mucina et al. 2006). Large numbers of waterfowl and waders use one large permanent pan inside the reserve for foraging, roosting and moulting (Milstein 1975; Taylor et al. 1999). However, few Egyptian Geese were observed to breed (fewer than 10 pairs during peak breeding season) and forage at the pan in the reserve. Egyptian Geese flew out at sunrise and at sunset to forage in the surrounding cropland but spent the rest of the day and night roosting along the shore of and on islands in the pan (Milstein 1975; M. Ndlovu, pers. obs.). Barberspan nature reserve is a Ramsar site, based in part on the waterbird populations of the wetland in the nature reserve (Cowan and Marneweck 1996).
Strandfontein Sewage Works is on the Atlantic Ocean coast in the south-western corner of the country. The area has a Mediterranean climate and vegetation, with wet winters and warm, dry summers and plant communities dominated by shrubs. The surrounding area has strong gradients of human population density, ranging from a large city (Cape Town, with a metropolitan population of 3.7 million people) to sparsely settled agricultural and rural areas. The dominant agriculture of the area consists of viticulture and crop cultivation. The sewage farm is hydrologically managed and some ponds have open mudflats that provide roosting sites for waterfowl and feeding areas for waders. Three pairs of Egyptian Geese bred at this site during the study period, and the other birds captured here bred in nearby suburban parks (Mangnall and Crowe 2002).
Telemetry
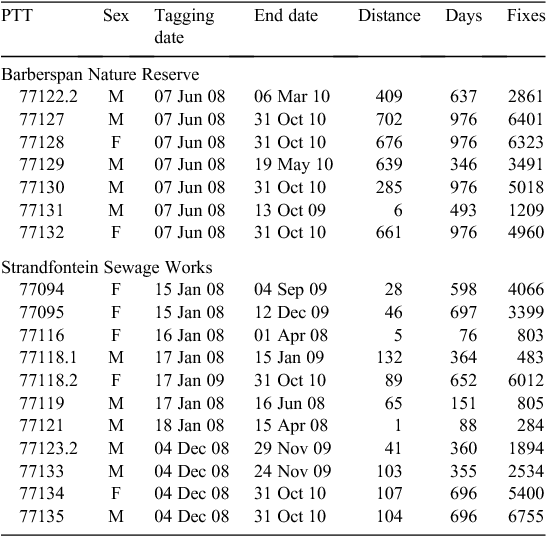
Between 15 January and 4 December 2008, 21 Egyptian Geese (8 at Barberspan, 13 at Strandfontein) that had just completed moult of flight-feathers were caught using walk-in traps placed near the edge of the water and baited with maize. Blood samples were taken from each tagged bird, stored in a lysis buffer solution and later sent to the University of Cape Town genetics lab for sex determination. A DNA sex test was carried out following the Griffiths et al. (1998) method. Mean weight of the 21 Geese was 2299 g ± 49 (mean ± 1 s.e.). The Geese were fitted with solar-powered, satellite GPS platform transmitter terminals (PTTs) (Microwave Telemetry Inc, Columbia, MD, USA; http://www.microwavetelemetry.com, accessed 20 November 2012) (Table 1). Transmitters each weighing 30 g (~1.3% of mean weight of study animals at capture) and were attached using a backpack harness (total Teflon ribbon weighing ~5 g for each harness), as described in detail by Cumming and Ndlovu (2011). Birds were all moved to a central processing point ~500 m away from the traps. It took an average of 15–20 min to fit a single transmitter on a bird, and the bird was released thereafter (Cumming and Ndlovu 2011).
|
The transmitters were set to capture data every 2 h (i.e. 12 points per day). The transmitters provided high-quality GPS fixes accurate to within 20 m. Distances covered between successive fixes were computed as straight-line distances. Satellite reception in southern Africa is excellent and detailed visual inspection of the data indicated no obviously incorrect points within the dataset. In addition, the analysis used thousands of data points and the most important movements were fairly long, meaning that small errors in location would be both difficult to screen out and irrelevant to the final conclusions. We manually removed testing locations and stationary points from the end of data series (i.e. where transmitters had fallen off or the animal was dead).
Determination of moult period
The mean moult-cycle period of Egyptian Geese in South Africa is 366 ± 73 days (n = 30; Milstein 1993) and Egyptian Geese are flightless and restricted to a moult-site for at least 28.5 days (Ndlovu et al. 2010). Egyptian Geese in South Africa are generally terrestrial grazers and moult in permanent pans during the dry season (Halse 1985; Milstein 1993; Ndlovu et al. 2010). Only birds that had just completed moult (before the moult flocks depart from the study site) were used for this study. Given the above information, timing of moult within the tracking data was determined by overlaying fixes on a Google map (www.google.earth.com, accessed 15 January 2012) identifying a continuous period of restricted movement (not more than 1 km from the water body) of a tagged bird for at least 30 days within a permanent wetland and occurring during the dry season. To further validate moulting seasons, bimonthly point counts and routine captures were carried out at each of the study sites.
Data analysis
General additive mixed models (GAMM) were used to investigate distances moved by Egyptian Geese in the summer- and winter-rainfall regions of South Africa at different times of the day, considering individual, sex, site of capture and season as potential influences on patterns of movements. General linear models (GLM) were inappropriate for these analyses because distances moved were non-linearly related to time of day and two of the covariates, sex and site, were binary. GAMM overcome the limitations of GLM by identifying non-linearities using flexible non-linear spline smoothing but preserve the ability of GLM to construct complex models with easy interpretability of predictor-response relationships (Hastie and Tibshirani 1990; Wood 2006).
One-way analysis of variance (ANOVA) with a 95% confidence interval level was also used to test for differences in daily distances moved by Egyptian Geese from Barberspan and Strandfontein. All analyses were carried out-using the R programming language for statistical computing (R Development Core Team 2010) using the vegan package mgcv (Wood 2006).
Results
In total, 18 of the PTTs that were deployed functioned for >50 days or gave more than 100 fixes, or both, and were used in the analysis. Data from three inconsistently transmitting PTTs (77120, 77117 and 77122.1) were omitted. One of the eight Barberspan birds (PTT 77177.1) was killed by a Black-backed Jackal (Canis mesomelas) at the site 6 days after tagging. Seven PTTs (77127, 77128, 77130, 77118.2, 77134, 77135) were still functional and transmitting on 31 October 2010, the day when data collection stopped (Table 1).
Movement during moult
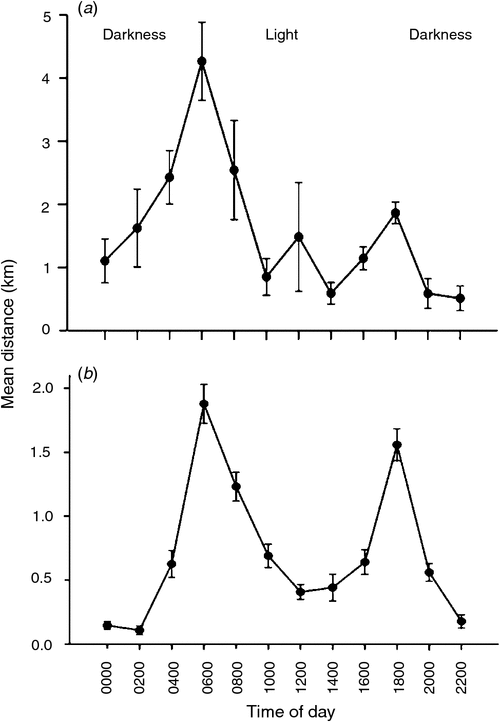
All moulting Egyptian Geese remained at the wetland where they were captured until after moult was completed. GAMM analysis showed that (correcting for the various random factors), the distance moved by moulting birds was significantly associated with time of day (F = 44.45, P < 0.001, adjusted R2 = 0.2898, n = 5798), that site had a significant effect on distance moved (F = 0.04, P = 0.034), and that sex had no effect on distance moved (F = 0.036, P = 0.849). Daily distances moved by individual birds in moult within a site did not differ significantly (Barberspan: F = 26.35, P = 0.052, n = 5; Strandfontein: F = 44.21, P = 0.06, n = 11) and Barberspan birds moved further on a daily basis than did Strandfontein birds (4.32 km ± 1.01 v. 1.95 km ± 0.76; F = 119.05, P < 0.001, n = 2). Moulting birds travelled furthest between 0400 and 0600 hours (GMT+2). During the moulting period, Barberspan sunrise was between 0600 and 0700 hours and sunset was between 1800 and 1900 hours whereas Strandfontein sunrise was between 0500 and 0600 hours and sunset was between 1900 and 2000 hours (Kruger and Esterhuyse 2005).
Movement when not moulting
Of the six birds captured at Barberspan with transmitters that functioned for at least 339 days, four birds moved >600 km from the point of capture and the other two moved 409 and 285 km from Barberspan (Table 1). In contrast, none of the 11 birds captured at Strandfontein travelled >132 km from the site. Four birds moved >100 km from Strandfontein, and the rest moved <90 km. PTT 77120 malfunctioned 27 days after deployment, at which time the bird was still at Strandfontein, but the transmitter was recovered 382 days later when the bird was shot by a farmer at Stellenbosch, 65 km from Strandfontein.
The GAMM analysis showed that time of day (F = 18.85, P < 0.001), bird location (entered in GAMM as a random variable; latitude × longitude; F = 14.47, P < 0.001) and month after moult (F = 7.81, P = 0.002) were the best predictors of distance moved (adjusted R2 = 0.129, n = 24271); sex of the bird had no effect on distance moved (F = 36.99, P = 0.52). Bird location was determined as the GPS position (±7 m) of the bird at a given fix. Daily distances moved by individual birds tagged from the same moulting site differed significantly (Barberspan: F = 4.25, P < 0.001, n = 8; Strandfontein: F = 54.16, P < 0.001, n = 9). Egyptian Geese that moulted at Barberspan moved, on average, significantly further per day (14 km ± 0.71 s.e.) than those from Strandfontein (6.6 km ± 0.19) (F = 83.85, P < 0.001).
On a daily basis, most movements by non-moulting Egyptian Geese (n = 4321) were between 1 and 10 km daily (Table 2). There were 85 instances of birds tagged at Barberspan moving >100 km in a day and 21 of these were in the first month after moult was completed (Month 1) and 16 in Month 11 after moult. The other 43 were in Months 2–10. In contrast, there were only two movements of birds captured at Strandfontein >100 km in a day, and both were by the same bird.

|
Egyptian Geese tagged at Barberspan moved furthest in the morning, between 0400 and 0800 hours (Fig. 1a). Egyptian Geese tagged at Strandfontein also moved furthest in the morning (0400–0800 hours), but undertook some longer movement distances in the late afternoon (1600–1800 hours; Fig. 1b). At both sites, there was a lull in activity during the middle of the day (1000–1600 hours), and least movement occurred at night between 2000 and 0200 hours.
|
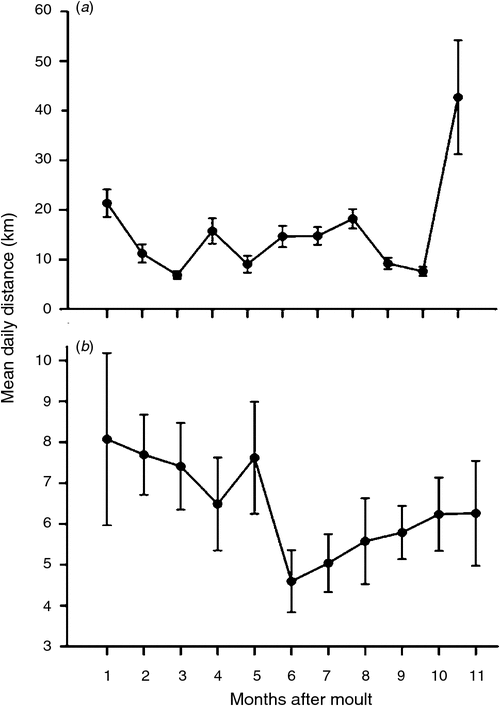
In addition to being linked to time of day, movement patterns were also linked to the timing of moult. Birds from Barberspan travelled the furthest immediately before moult (Month 11), and again travelled longer distances immediately after moult (Month 1) , though not as far as before moult (Fig. 2a). For the rest of the year, distances moved daily varied little (Fig. 2a). Like their counterparts in the summer-rainfall region, birds tagged at Strandfontein moved furthest in the month immediately after moult (Month 1), but the distances moved were not significantly different from those travelled in the subsequent 4 months. In Month 6 after moult, distances moved daily decreased sharply then increased steadily until Month 11, immediately before moult began (Fig. 2b).
|
Outside the moulting period, birds from Barberspan never returned to their moulting site, whereas non-moulting birds from Strandfontein returned to the moulting site an average of 2.31 times ± 0.02 per year, although each return visit lasted only 2–4 h. Four of the five birds satellite-tagged at Barberspan returned to Barberspan to moult the following year. Two of those same four birds moulted at Barberspan in the third year, whereas the other two birds moulted at two other sites <300 km away (Table 1). Between moults, three Barberspan birds dispersed maximum distances of over 600 km and the other two birds moved maximum distances of 409 and 285 km from Barberspan. Of the seven birds satellite-tagged at Strandfontein, only one returned to moult there the following year. One bird moulted at a nearby pan (5 km away), two moulted at estuaries less than 30 km away and one moulted at a farm dam 56 km away. Two birds satellite-tagged at Strandfontein did not moult the following year and returned to moult at Strandfontein in the third year. None of the seven Strandfontein-tagged birds travelled more than 132 km from the moult site. Four birds dispersed maximum distances of 103–132 km, whereas the remainder travelled less than 90 km from Strandfontein.
Discussion
Our data show that moulting birds at both Barberspan and Strandfontein moved furthest (displacement distances on a daily basis) at sunrise, with a second but smaller peak in distance moved before or after sunset. During the period of inactivity (as indicated by telemetry and validated by field observations) between dawn and sunset, birds spent most of their time either sitting on the shore or roosting in large flocks on sandy islands inside the water while loafing and preening, and not moving far (as determined by two-hourly fixes). Similar observations of Egyptian Geese loafing during the day have been reported by Halse (1985) and Gyemesi and Lensink (2010) and they both concluded that Egyptian Geese need ~4.5 h of foraging to satisfy their nutritional needs and dedicate more daytime for self-maintenance especially when birds are growing new feathers. The reduction in foraging time during the day in moulting birds is also a phenotypic flexibility strategy to reduce wing loading by reducing body mass, which ultimately shortens the flightless period (Ndlovu et al. 2010). Given that a moulting Egyptian Geese is flightless for at least 29 days (Halse 1985; Ndlovu et al. 2010) and the risk of predation is high for a flightless bird (Milstein 1993), another reason why moulting Egyptian Geese do not travel long distances away from the water during the day time is possibly to avoid detection and predation by terrestrial predators (Halse 1985; Milstein 1993). At night, birds swim towards open water inside the pan for safety and some roost on islands. These crepuscular patterns of activity and nocturnal retreats to safety are compatible with behaviour in response to the predation risks posed by nocturnal predators, such as Black-backed Jackals. As the predators return to their dens at dawn (Estes 1992), the birds come ashore to graze.
Moulting Egyptian Geese at Barberspan travelled farther on a daily basis than moulting birds at Strandfontein. This probably relates to the differing sizes of the two wetlands. Barberspan has a surface area of water of ~1700 ha compared with only 319 ha at Strandfontein, meaning that flightless moulting Egyptian Geese at Barberspan swim longer distances to cross the wetland or to travel from roosting islands to graze onshore.
At the end of moult, Egyptian Geese dispersed from their moulting sites. Most long daily movements by individuals were within the first 10 days after moult finished and just before the next moult began. The long-distance travel paths summarise these movements and provide compelling evidence that these birds undertaking moult migrations (Table 1, see fig. 2 in Cumming et al. 2012). The existence of moult migrations by waterfowl in southern Africa has long being suspected (e.g. Geldenhuys 1981; Milstein 1993) but has never been proven, not least because of the disparate directions in which migrating individuals move (see Cumming et al. 2012). This study, together with our previous larger-scale analysis of the same dataset (Cumming et al. 2012), thus provides empirical evidence of such migration and some indication of the distances that birds travel during these migrations. It is not clear why Egyptian Geese tagged at Barberspan travel such long distances (>600 km in the case of four individuals) to moult again at Barberspan, given that these birds bypass several seemingly suitable moulting sites en route. It is very likely that similar migrations take place among South African Shelducks, a species whose global population moults at only a few wetlands (Geldenhuys 1981). Although general seasonal patterns of movement in relation to the timing of moult were concordant in both groups of birds, there were considerable differences in the details of movement paths between Barberspan and Strandfontein birds (Table 1; Cumming et al. 2012).
The daily movements of Egyptian Geese can be divided into short- and long-distance travels. Short distances reflect foraging within a discrete area, typically with movements between a roosting wetland and a grazing pasture, or between wetlands. Long flights were predominantly undertaken at night (88%) and were associated with semi-permanent changes in location. The pattern of daily movement is strongly skewed with >70% of flights being over distances of <10 km (Table 2).
Birds tagged at Strandfontein typically flew back to Strandfontein at least twice between moults, but would spend at most 4 h there before returning to their previous location. Similar prospecting flights have been observed in Grey Teal (Roshier et al. 2006). In contrast, birds at Barberspan did not return to the moulting site between moults.
Simmons et al. (1999) suggest that waterfowl can find temporary wetlands using visual cues, such as formation of thunderclouds that precede major rain fronts. Although this may be true over spatial scales of <100 km, it seems an unlikely explanation for night-time flights by Egyptian Geese that flew to isolated waterbodies in dry regions of Botswana, Namibia and the Northern Cape Province. Rather, these observations suggest that Egyptian Geese at Barberspan interact with their environment at very broad scales and that their long-distance flights are heavily influenced by experience and spatial memory (Nathan et al. 2008; Roshier et al. 2008; Cumming et al. 2012). Birds from Strandfontein, by contrast, travelled shorter distances and more frequently, thus sampling their surroundings more constantly in a manner that would be more characteristic of nomadism than experience (Nathan et al. 2008; Cumming et al. 2012).
There were no clear, sex-related differences in any of the movement parameters examined in this study. Given that Egyptian Geese form long-term pair-bonds (Milstein 1993), this is not unexpected. Birds from the resource-dense, winter-rainfall area remained near their moulting site, whereas birds from the semi-arid, resource-sparse, summer-rainfall area travelled much further afield and did not return to their moulting site between moults. Once Egyptian Geese from Barberspan had arrived at their post-moult destinations, most of their daily movements were over short distances, similar to the movement patterns of White-faced Whistling-Duck in the same region (Petrie and Rogers 1997). White-faced Whistling-Ducks stay several months in one area and frequently undertake short daily flights between wetlands or from wetlands to terrestrial foraging sites (Petrie and Rogers 1997). A similar pattern of short-distance daily movement is also shown by Grey Teal in Australia (Roshier et al. 2006). These birds travel mean distances of 2.9–25.2 km day–1, similar to the distances travelled daily by Egyptian Geese in the present study. The longest daily distance recorded for a Grey Teal is 332 km (Roshier et al. 2006), exactly half the longest daily distance travelled by an Egyptian Goose (Table 2). A Comb Duck tagged in Malawi had a displacement distance of 655 km in 285 days, similar to displacement distances of Egyptian Geese from Barberspan (Cappelle et al. 2011).
Barberspan birds undertake long-distance directed movements that cannot be interpreted as random wanderings between adjacent wetlands. Movements of Strandfontein birds, in contrast, do appear to be unpredictable (see fig. 2 in Cumming et al. 2012) and over short distances, fit the expectations of nomadism (Bennetts and Kitchens 2000; Mueller and Fagan 2008). It appears that the proximate cues for movement by Strandfontein birds act at the local landscape level, whereas birds from Barberspan rely more on memory (possibly genetically ‘hard-wired’) when travelling long distances. This suggestion is reinforced by the consistency with which individual birds used the same general flight-path to and from their moulting sites in successive years (Cumming et al. 2012). A similar dichotomy in movement patterns has been documented among dispersing, juvenile African Black Oystercatchers (Haematopus moquini). Some move by a process of diffusion dispersal, covering fairly short distances, whereas others both bypass apparently suitable resources and traverse extensive stretches of inhospitable coastline to congregate at sites >1500 km from their natal sites and outside the breeding range of the species (Hockey et al. 2003). In the Oystercatchers, it has been mooted that short-distance movements may be driven by proximate environmental cues whereas longer-distance movements are under genetic control (Hockey et al. 2003). Our results suggest that Egyptian Geese have equally divergent movement strategies that may be determined more by where they moult than where they breed. The fact that they are over-dispersed when breeding and under-dispersed when moulting suggests that suitable moulting sites, certainly for those birds that live in the semi-arid regions, may be the limiting resource that ultimately determines their life history.
The differences in daily movement distances and distances moved to preferred moulting sites between populations within different rainfall regions probably reflect differences in predation pressure and the dispersion of permanent wetlands between south-western and north-eastern South Africa, suggesting that broad-scale features have some influence in shaping the daily movement patterns and moult-migration patterns of Egyptian Geese. Although the findings confirm some known aspects of semi-arid waterfowl ecology (for prior documentation of the influence of wetland distribution on movement patterns, see Petrie and Rogers 1997; Roshier and Reid 2003; Roshier et al. 2006), it is important to note that Egyptian Geese from the two sites appear to show different patterns of movements, despite evidence from ringing that there is a limited exchange of birds between these two sites (Underhill et al. 1999).
Acknowledgements
We thank Gregory Mutumi, David Nkosi and many other volunteers for their help with field work. Timothy Reed helped with GAMM analysis. We are grateful to the Percy FitzPatrick Institute staff, particularly Hilary Buchanan, Tania Jansen, Chris Tobler, Rob Little, Anthea Links, Margaret Koopman and Lionel Mansfield, for all the logistical support rendered during this study. We also thank Sampie van der Merwe and Barberspan Bird Sanctuary staff (particularly Amos Koloti, Sydwell Setuki, Mmapula Kakgodi and Andrew Mvundle) for supporting this project at Barberspan. For assistance in working at Strandfontein we are grateful to Morné Carstens, Eben Olderwagen, Dalton Gibbs and the Strandfontein Wastewater Treatment works management staff. Permits for this research were issued by Cape Nature and the North West Parks Board. The project was funded through a USAID grant awarded to G.S. Cumming under the Wildlife Conservation Society’s Global Avian Influenza Network for Surveillance (GAINS) program. Further funding came from the Department of Science and Technology through the National Research Foundation, Centre of Excellence at the Percy FitzPatrick Institute and the KW Johnstone Doctoral Package Scholarship. The first and second authors acknowledge the memory of the third author, the late Phil Hockey.
References
Anderson, M. G., Rhymer, J. M., and Rohwer, F. C. (1992). Philopatry, dispersal and the genetic structure of waterfowl populations. In ‘Ecology and Management of Breeding Waterfowl’. (Eds B. D. J. Batt, A. D. Afton, M. G. Anderson, C. D. Ankney, D. H. Johnson, J. A. Kadlec and G. L. Krapu.) pp. 365–395. (University of Minnesota Press: Minneapolis, MN.)Bennetts, R. E., and Kitchens, W. M. (2000). Factors influencing movement probabilities of a nomadic food specialist: proximate foraging benefits or ultimate gains from exploration? Oikos 91, 459–467.
| Factors influencing movement probabilities of a nomadic food specialist: proximate foraging benefits or ultimate gains from exploration?Crossref | GoogleScholarGoogle Scholar |
Bollinger, K. S., and Derksen, D. V. (1996). Demographic characteristics of molting Black Brant near Teshekpuk Lake, Alaska. Journal of Field Ornithology 67, 141–158.
Bowman, T. D., and Brown, P. W. (1992). Site fidelity of male Black Ducks to a molting area in Labrador. Journal of Field Ornithology 63, 32–34.
Cappelle, J., Iverson, S. A., Takekawa, J. Y., Newman, S. H., Dobman, T., and Gaidet, N. (2011). Implementing telemetry on new species in remote areas: recommendations from a large-scale satellite tracking study of African waterfowl. Ostrich 82, 17–26.
| Implementing telemetry on new species in remote areas: recommendations from a large-scale satellite tracking study of African waterfowl.Crossref | GoogleScholarGoogle Scholar |
Cowan, G. I., and Marneweck, G. C. (1996). South African National Report to the Ramsar Convention 1996. Department of Environmental Affairs and Tourism, Pretoria.
Cumming, G. S., and Ndlovu, M. (2011). Satellite telemetry of Afrotropical ducks: methodological details and assessment of success rates. Journal of African Zoology 46, 425–434.
| Satellite telemetry of Afrotropical ducks: methodological details and assessment of success rates.Crossref | GoogleScholarGoogle Scholar |
Cumming, G. S., Gaidet, N., and Ndlovu, M. (2012). Linking dispersal and biogeography: comparative analysis of movements of southern African ducks at three different latitudes using satellite telemetry. Journal of Biogeography 39, 1401–1411.
| Linking dispersal and biogeography: comparative analysis of movements of southern African ducks at three different latitudes using satellite telemetry.Crossref | GoogleScholarGoogle Scholar |
Dugger, B. D. (1997). Factors influencing the onset of spring migration in Mallards. Journal of Field Ornithology 68, 331–337.
Estes, R. (1992). Jackals. In ‘The Behavior Guide to African Mammals: Including Hoofed Mammals, Carnivores and Primates’. pp. 90–97. (University of California Press: Berkeley, CA.)
Fauchald, P., and Tveraa, T. (2006). Hierarchical patch dynamics and animal movement. Oecologia 149, 383–395.
| Hierarchical patch dynamics and animal movement.Crossref | GoogleScholarGoogle Scholar | 16794832PubMed |
Fleskes, J. P., Mauser, D. M., Yee, J. L., Blehert, D. S., and Yarris, G. S. (2010). Flightless and post-molt survival and movements of female Mallards molting in Klamath Basin. Waterbirds 33, 208–220.
| Flightless and post-molt survival and movements of female Mallards molting in Klamath Basin.Crossref | GoogleScholarGoogle Scholar |
Geldenhuys, J. N. (1981). Moult and moult localities of the South African Shelduck. Ostrich 52, 129–134.
| Moult and moult localities of the South African Shelduck.Crossref | GoogleScholarGoogle Scholar |
Griffiths, R., Double, M. C., Orr, K., and Dawson, R. J. G. (1998). A DNA test to sex most birds. Molecular Ecology 7, 1071–1075.
| A DNA test to sex most birds.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaK1cXlslGmt7c%3D&md5=7279a17064467487f4f088122e32cff5CAS | 9711866PubMed |
Gudmundsson, G. A., Benvenuti, S., Alerstam, T., Papi, F., Lilliendahl, K., and Akesson, S. (1995). Examining the limits of flight and orientation performance: satellite tracking of Brent Geese migrating across Greenland ice-cap. Proceedings of the Royal Society of London – B. Biological Science 261, 73–79.
| Examining the limits of flight and orientation performance: satellite tracking of Brent Geese migrating across Greenland ice-cap.Crossref | GoogleScholarGoogle Scholar |
Gyemesi, A., and Lensink, R. (2010). Risk analysis of the Egyptian Goose in the Netherlands. (Bureau Waardenburg bv; Ministry of Agriculture, Nature and Food Quality, Invasive Allen Species Team: Wageningen, the Netherlands.) Available at http://www.buwa.nl/fileadmin/buwa_upload/Bureau_Waardenburg_rapporten/09-617_Egyptian_Goose_final_draft_30-09-2010_web.pdf [Verified 15 May 2013].
Halse, S. A. (1985). Activity budgets of Spurwinged and Egyptian Geese at Barberspan during winter. Ostrich 56, 104–110.
| Activity budgets of Spurwinged and Egyptian Geese at Barberspan during winter.Crossref | GoogleScholarGoogle Scholar |
Hastie, T., and Tibshirani, R. (1990). ‘Generalized Additive Models.’ (Chapman and Hall: New York.)
Hockey, P. A. R., Leseberg, A., and Loewenthal, D. (2003). Dispersal and migration of juvenile African Black Oystercatchers (Haematopus moquini). Ibis 145, 114–123.
| Dispersal and migration of juvenile African Black Oystercatchers (Haematopus moquini).Crossref | GoogleScholarGoogle Scholar |
Kruger, A. C., and Esterhuyse, D. J. (2005). Climate of South Africa: Sunshine and Cloudiness, WS46. South African Weather Service, Pretoria.
Levin, S. A. (1974). Dispersion and population interactions. American Naturalist 108, 207–228.
| Dispersion and population interactions.Crossref | GoogleScholarGoogle Scholar |
Lindberg, M. S., and Walker, J. (2007). Satellite telemetry in avian research and management: sample size considerations. Journal of Wildlife Management 71, 1002–1009.
| Satellite telemetry in avian research and management: sample size considerations.Crossref | GoogleScholarGoogle Scholar |
Mangnall, M. J., and Crowe, T. M. (2002). Population dynamics and physical and financial impacts to cereal crops of the Egyptian Goose Alopochen aegyptiacus on the Agulhas Plain, Western Cape, South Africa. Agriculture, Ecosystems & Environment 90, 231–246.
| Population dynamics and physical and financial impacts to cereal crops of the Egyptian Goose Alopochen aegyptiacus on the Agulhas Plain, Western Cape, South Africa.Crossref | GoogleScholarGoogle Scholar |
Miller, M. R., Takekawa, J. Y., Fleskes, J. P., Orthmeyer, D. L., Casazza, M. L., and Perry, W. M. (2005). Spring migration of Northern Pintails from California’s Central Valley wintering area tracked with satellite telemetry: routes, timing, and destinations. Canadian Journal of Zoology 83, 1314–1332.
| Spring migration of Northern Pintails from California’s Central Valley wintering area tracked with satellite telemetry: routes, timing, and destinations.Crossref | GoogleScholarGoogle Scholar |
Milstein, P. le S. (1975). The biology of Barberspan, with special reference to the avifauna. Ostrich 83, 1–74.
Milstein, P. le S. (1993). A study of the Egyptian Goose (Alopochen aegyptiacus). Ph.D. Thesis, Department of Zoology, University of Pretoria, Pretoria.
Morales, J. M., and Ellner, S. P. (2002). Scaling up animal movements in heterogeneous landscapes: the importance of behavior. Ecology 83, 2240–2247.
| Scaling up animal movements in heterogeneous landscapes: the importance of behavior.Crossref | GoogleScholarGoogle Scholar |
Mucina, L., Rutherford, M. C., and Powrie, L. W. (2006). Vegetation Atlas of South Africa, Lesotho and Swaziland. In ‘The Vegetation of South Africa, Lesotho and Swaziland’. (Eds L. Mucina and M. C. Rutherford.) pp. 748–789. (South African National Biodiversity Institute: Pretoria.)
Mueller, T., and Fagan, W. F. (2008). Search and navigation in dynamic environments – from individual behaviors to population distributions. Oikos 117, 654–664.
| Search and navigation in dynamic environments – from individual behaviors to population distributions.Crossref | GoogleScholarGoogle Scholar |
Ndlovu, M., Cumming, G. S., Hockey, P. A. R., and Bruinzeel, L. W. (2010). Phenotypic flexibility of a southern African duck Alopochen aegyptiaca: do northern hemisphere paradigms apply? Journal of Avian Biology 41, 558–564.
| Phenotypic flexibility of a southern African duck Alopochen aegyptiaca: do northern hemisphere paradigms apply?Crossref | GoogleScholarGoogle Scholar |
Nathan, R., Getz, W. M., Revilla, E., Holyoak, M., Kadmona, R., Saltz, D., and Smouse, P. E. (2008). A movement ecology paradigm for unifying organismal movement research. Proceedings of the National Academy of Sciences of the United States of America 105, 19 052–19 059.
| A movement ecology paradigm for unifying organismal movement research.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD1cXhsFamurfP&md5=a0e7d7f6b372117f62850865cc889ca1CAS |
Oatley, T. B., and Prŷs-Jones, R. P. (1986). A comparative analysis of movements of southern African waterfowl (Anatidae), based on ringing recoveries. South African Journal of Wildlife Research 16, 1–6.
Petrie, S. A., and Rogers, K. H. (1997). Satellite tracking of White-faced Whistling Ducks in a semiarid region of South Africa. Journal of Wildlife Management 61, 1208–1213.
| Satellite tracking of White-faced Whistling Ducks in a semiarid region of South Africa.Crossref | GoogleScholarGoogle Scholar |
Pyke, G. H. (1984). Optimal foraging theory: a critical review. Annual Review of Ecology and Systematics 15, 523–575.
| Optimal foraging theory: a critical review.Crossref | GoogleScholarGoogle Scholar |
R Development Core Team (2010). ‘R: A Language and Environment for Statistical Computing.’ (R Foundation for Statistical Computing: Vienna.) Available at http://www.R-project.org [Verified 15 January 2012].
Robertson, R. J., and Cooke, F. (1999). Winter philopatry in migratory waterfowl. Auk 116, 20–34.
| Winter philopatry in migratory waterfowl.Crossref | GoogleScholarGoogle Scholar |
Roshier, D. A., and Reid, J. R. W. (2003). On animal distributions in dynamic landscapes. Ecography 26, 539–544.
| On animal distributions in dynamic landscapes.Crossref | GoogleScholarGoogle Scholar |
Roshier, D. A., Klomp, N. I., and Asmus, M. (2006). Movement of a nomadic waterfowl, Grey Teal (Anas gracilis), across inland Australia – results from satellite telemetry spanning fifteen months. Ardea 94, 461–475.
Roshier, D. A., Asmus, M. W., and Klaassen, M. (2008). What drives long-distance movements in Grey Teal Anas gracilis in Australia? Ibis 150, 474–484.
| What drives long-distance movements in Grey Teal Anas gracilis in Australia?Crossref | GoogleScholarGoogle Scholar |
Salomonsen, F. (1968). The moult migration. Wildfowl 19, 5–24.
Seymour, N. R. (1991). Philopatry in male and female American Black Ducks. Condor 93, 189–191.
| Philopatry in male and female American Black Ducks.Crossref | GoogleScholarGoogle Scholar |
Simmons, R. E., Barnard, P., and Jamieson, I. G. (1999). What precipitates influxes of wetland birds to ephemeral pans in arid landscapes? Observations from Namibia. Ostrich 70, 145–148.
| What precipitates influxes of wetland birds to ephemeral pans in arid landscapes? Observations from Namibia.Crossref | GoogleScholarGoogle Scholar |
Szymczak, M. R., and Rexstad, E. A. (1991). Harvest distribution and survival of a Gadwall population. Journal of Wildlife Management 55, 592–600.
| Harvest distribution and survival of a Gadwall population.Crossref | GoogleScholarGoogle Scholar |
Taylor, P. B., Navarro, R. A., Wren-Sargent, M., Harrison, J. A., and Kieswetter, S. L. 1999. TOTAL CWAC Report: Coordinated Waterbird Counts in South Africa, 1992–97. Avian Demography Unit, University of Cape Town, Cape Town.
Underhill, L. G., Tree, A. J., Oschadleus, H. D., and Parker, V. (1999). ‘Review of Ring Recoveries of Waterbirds in Southern Africa.’ (Avian Demography Unit, University of Cape Town: Cape Town.)
Wood, S. N. (2006). ‘Generalized Additive Models: An Introduction with R.’ (Chapman and Hall and CRC Press: Boca Raton, FL.)
Zollner, P. A., and Lima, S. L. (1999). Search strategies for landscape-level interpatch movements. Ecology 80, 1019–1030.
| Search strategies for landscape-level interpatch movements.Crossref | GoogleScholarGoogle Scholar |