

Faecal particle size and tooth wear of the koala (Phascolarctos cinereus)
William Ellis A B E , Rachael Attard C , Stephen Johnston B , Peter Theileman D , Allan McKinnon D and David Booth CA Centre for Mined Land Rehabilitation, Sustainable Minerals Institute, The University of Queensland, Brisbane, Qld 4072, Australia.
B Wildlife Biology Unit, School of Agriculture and Food Sciences, The University of Queensland, Gatton, Qld 4343, Australia.
C School of Biological Science, The University of Queensland, Brisbane, Qld 4072, Australia.
D Queensland Department of Environment and Heritage Protection, Moggill Koala Hospital, Moggill, Qld 4070, Australia.
E Corresponding author. Email: w.ellis@uq.edu.au
Australian Mammalogy 36(1) 90-94 https://doi.org/10.1071/AM13033
Submitted: 10 October 2013 Accepted: 3 December 2013 Published: 4 March 2014
Abstract
We used computer-aided image analysis of leaf fragment particles found in faecal pellets of 45 koalas, representing the range of tooth wear in this species, to investigate how tooth wear in the koala influences faecal particle sizes. Although the range of sizes of particles produced did not vary between koalas across different tooth wear classes, with all koalas producing small, medium and large particles, koalas with advanced tooth wear produced a greater proportion of larger particles. This observation may prove useful for demographic population analyses based on scat surveys since the broad age class of individual koalas can be estimated from faecal pellet analysis. Older koalas produced faecal pellets containing a higher proportion of the largest-sized particle sizes (those greater than 0.59 mm2) than either young or mature koalas but there was no difference detected between mature and young koalas.
Additional keywords: faecal pellets, population age structure, tooth wear.
Introduction
Faecal pellet searches are often used in surveys to detect koala (Phascolactos cinereus) presence and examination of the leaf fragments that constitute the pellets can reveal the diet of the koalas that produced them (Ellis et al. 1999). The leaf particle sizes in the digestive tract of koalas have been shown to reflect tooth wear, which, in turn, is an index of age and consequently an important parameter in understanding the dynamics and life history of species (Logan and Sanson 2002b). Non-invasive methods of evaluating koala distribution, such as mapping faecal pellet distributions, are important tools for conserving such cryptic species because they produce an unequivocal record of presence with minimal effort (Rhodes et al. 2011; Ellis et al. 2013). The analysis of faecal pellet distributions has been particularly useful for studying koala population trends (Seabrook et al. 2011) because they can have large home ranges of up to 100 ha (Ellis et al. 2002) and are often difficult to observe during surveys of their habitat (Dique et al. 2003; Phillips and Callaghan 2011; Woosnam-Merchez et al. 2012). In addition to information about which trees koalas use and eat, recent work suggests that fresh pellets may be analysed to produce information on endocrine profiles (Davies et al. 2013; Narayan et al. 2013) and genetic information for free-ranging koalas (Wedrowicz et al. 2013). It is necessary to identify the age distribution of a population to examine and understand population trends (Caughley and Gunn 1993): for koalas, the collection and analysis of skeletal material has revealed patterns of mortality across a wide geographical range (Melzer et al. 2011), but perhaps faecal pellet collections could be used to non-invasively collect data on the age structure of a population in situ.
Digestive efficiency in the koala, as for other herbivores, is related to food particle size in the gut, which, in turn, is primary determined by mastication (Logan and Sanson 2002a). The efficiency by which koalas can produce small particles is dictated by tooth wear: animals with worn teeth produce fewer small particles, and such animals may have to depend on strategies such as merycism (rumination-like behaviour) (Logan 2001) or selection of younger leaves to produce small particles. Indeed, the relationship between tooth wear and digestive efficiency is considered so close in koalas that tooth wear is described as the most significant consequence of ageing because it impacts on many life-history attributes such as resource protection and socialisation (Logan and Sanson 2002b). Ultimately, like other herbivores, the decrease in chewing efficiency is accompanied by a decrease in nutritional uptake and loss of body condition until the koala becomes too weak to survive (Lanyon and Sanson 1986).
Previous studies have revealed a relationship between tooth wear and particle size in the digestive tract of the koala (Logan and Sanson 2000): here we test whether such a relationship exists in the faeces of koalas, which could provide a tool for demographic estimation based on faecal sampling. Our testable hypothesis was that faecal particle size could be used to calculate chronological age in koalas.
Materials and methods
Study animals and faecal samples
Faecal samples used in this study were sourced from koalas at the Queensland Department of Environment and Heritage Protection Koala Hospital, at Moggill (Queensland), where sick, injured and orphaned koalas from south-east Queensland are bought for care. The koalas that provided faecal pellets for this study were trauma victims that were placed alone in small aluminium holding cages (1 m × 0.5 m × 0.5 m) in the hospital ward at the Moggill facility upon arrival. Faecal pellets were collected within 24 h of arrival to ensure that faeces contained plant material from food consumed before admission; a minimum of three individual koalas per tooth-wear category were sampled in this study (Table 1).

|
Each koala was also assigned to a tooth-wear category from 1 to 13+ (determined by P. Thieleman for all subjects), as described in Gordon (1991) (Table 1). To ensure that a comparative set of otherwise healthy koalas was sampled, each koala was initially assessed and assigned a body condition score between 1 and 10 (Ellis and Carrick 1992), as determined by a single judge (P. Thieleman) for consistency. Younger koalas with poor body condition scores (less than 6) were excluded, since poor body condition in young koalas may indicate a nutritional deficiency due to undiagnosed health conditions that could compromise the project. However, we had to include older koalas with a body condition score of less than 6 because (a) a decrease in body condition is expected to occur with advanced stages of tooth wear (Skogland 1988) and (b) particularly for the very advanced tooth-wear classes, no individuals with body score greater than 6 were encountered.
Particle size analysis
Ten faecal pellets were collected from each koala; five of these were used to produce microscopic slide preparations for faecal particle size analysis. These pellets were first crushed between the fingers and the resulting crushed material placed into a 4 cm × 4 cm plastic weighboat (Cole Parmer, Australia) containing 50 mL of water and a single drop of dishwashing liquid (Colgate-Palmolive Co, USA) to help separate the particles. After mixing with a plastic stirring rod to further break up any aggregations of material, the weigh boat was agitated by hand and 1 mL of mixture drawn into a plastic pipette. This mixture was expelled onto a 76 mm × 25 mm microscope slide, left exposed to dry for ~2 h at room temperature and stored uncovered. Three such slides were created for each faecal pellet, resulting in 15 slides per koala.
Computer-aided image analysis was used to analyse faecal particle size because it is an accurate and rapid method of comparing the measurements of numerous non-geometric particles (Logan and Sanson 2000). Each slide was viewed under a compound microscope under ×40 magnification, using a Q Imaging Micropublisher 3.3 RTV camera mounted on the microscope. ‘Q Capture’ (Q Capture Suite Plus, Q Imaging, BC, Canada) software was used to capture five images per slide, which allowed measurement of at least 100 particles per slide. Hence, for each pellet, at least 1500 particles were measured, resulting in at least 7500 particles being measured for each koala. Each resulting image was then converted to greyscale so objects could be distinguished clearly from the background using the software package ‘IrfanView’ (Irfan Skiljan, http://www.irfanview.com). The visible area of each particle was measured using the image analysis software ‘Image J’ (National Institute of Health, Research Services Branch, http://rsbweb.nih.gov), using a 1-mm scale object for reference, to obtain an indication of particle size. Very small particles and bacterial cells were excluded from image analysis because the magnification of the microscope was not sufficient to detect particles of this size (Logan and Sanson 2000). The ‘Image J’ program measures and records particle size in terms of pixels, and the 1-mm scale object measured 1300 pixels; hence, this scaling factor was used to convert pixels to square millimetres, which are the units we report here. To ascertain the sampling protocol required to ensure that all size classes of particles present on a slide were detected, we captured multiple images from nine sample slides and compared particle size across the cumulative data from each successive slide [similar in fashion to the species area curve concept (Preston 1962)]; this approach confirmed that five slides were sufficient to accurately estimate the frequency of all size classes.
Statistical analysis
Particles in the faeces were sorted into seven size classes based on an arbitrary measure for analysis. These size classes were: less than 5.9 × 10−6 mm2, 6.0 × 10−6–5.9 × 10−5 mm2, 6.0 × 10−5–5.9 × 10−4 mm2, 6.0 × 10−4–5.9 × 10−3 mm2, 6.0 × 10−3–5.9 × 10−2 mm2, 6.0 × 10−2–0.59 mm2 and particles greater than 0.59 mm2. Thereafter we conducted simple statistical tests to ascertain whether the material presented on the slide could be interpreted to reveal the tooth wear of the koala. First, we compared particle size variation between tooth wear classes, testing whether older koalas (with greater tooth wear) produced a broader spread of particle sizes. Second, we compared the average size of particles across the range of tooth wear classes, to test whether the average particle size on the slides we made was correlated with tooth wear. Finally, we tested the relationship between the proportional representation of the largest particles in slides and their corresponding tooth wear classes, again attempting to provide a simple test of age using slides made from faecal pellets. To determine whether the variation between each tooth wear category was significantly different, a weighted regression model was fitted to the log10-transformed data and compared with an unweighted regression model of the same data using a ratio test to test for heteroscedasticity. We then conducted a non-parametric correlation analysis of average particle size for each tooth wear category to assess the relationship between these measures and finally we conducted the same test using the proportion of largest-sized fragments for each tooth wear class.
Statistical analyses were performed using the software package R 2.9.2. (Venables and Smith, R Development Core Team) and StatPlus (AnalystSoft, www.analystsoft.com).
Results
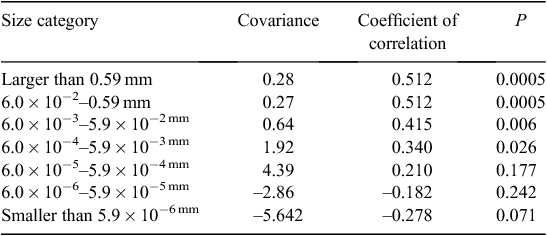
The average size of particles in faeces was significantly correlated with tooth wear classes (weighted regression analysis P = 1 × 10−4, n = 45). Faecal pellets from koalas with tooth wear between Class 1 and Class 7 contained significantly fewer of the largest-sized particles (>0.59 mm2) than those from koalas with tooth wear of Class 7 or above (t14 = 2.81, P = 0.00694). We calculated the covariance between the proportions of each size class of particles in the faeces and the tooth wear class of the koala. As expected, the proportions of the largest particles increased with advancing tooth wear while the proportions of the smallest particles decreased as tooth wear increased (Table 2). Coefficients of correlation between these variables (proportion of particles that were in each size class) were non-significant except for the three largest particle classes, the proportions of all of which were correlated with tooth wear (Table 2).
|
Most of particles were smaller than 2.95 × 10−5 mm2 in size, and within this size range most particles were less than 6.0 × 10−6 mm2. These extremely small particles, however, comprised less than 1% of biomass (assuming two-dimensional particles, less if calculated in three dimensions), and we are not confident that these represent leaf fragments rather than endogenous cells, sloughed gut lining or even microbes. Together, size Classes 1, 2 and 3 (5.9 × 10−6 mm2, 6.0 × 10−6–5.9 × 10−5 mm2 and 6.0 × 10−5–5.9 × 10−4 mm2) accounted for less than 7% of biomass identified in the slides, yet they constituted the most abundant fragments identified (95% of fragments from koalas of tooth wear Classes 1–7, 93% of fragments from koalas of tooth wear Classes >7). From a biological perspective, the amount of material that falls into each size class (and hence its gut passage rate and digestibility) should determine its impact on the life history of the koala. Hence, it follows that biologically meaningful analyses should be conducted on these size classes and this was evidenced in our further analyses.
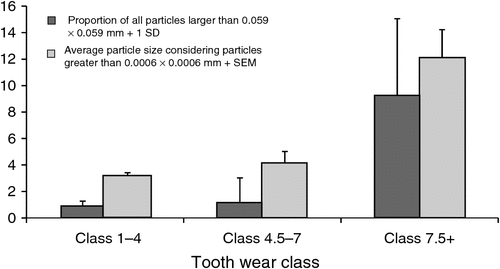
Koalas with advanced tooth wear (greater than Class 7) produced faeces that contained a greater standard deviation in average particle size when compared with koalas with less tooth wear (Fig. 1). This was statistically assessed using a ratio test comparing a weighted and unweighted regression model (P = 0.0227, n = 45).
|
The analysis of variation in particle size between individual tooth wear classes was affected by small sample sizes for each tooth wear class (Table 1), so koalas were further assigned to age groups ‘young’ (tooth wear Classes 1–4), ‘mature’ (tooth wear Classes 4–7) and ‘old’ (tooth wear Classes >7). Tests of heteroscedacity revealed unequal variances between each of these groups when compared for most particle size classes. Only for the second largest particle size group (0.06–0.59 mm2), for comparison between young and mature koalas, were the variances equal (F = 1.14, Fcrit = 3.049, P(2 tailed) = 0.79032).
Older koalas produced a higher proportion of the largest-sized particle sizes (those greater than 0.59 mm2) than either young (t11 = 3.10059, P = 0.01) or mature koalas (t11 = 3.00565, P = 0.006) but there was no difference detected between mature and young koalas (t15 = 0.86311, P = 0.20083) (Fig. 1).
Discussion
Koalas have recently been added to the list of species protected by the Environment Protection and Biodiversity Conservation Act (Cth) 1999 in Australia, which attempts to protect the habitat of protected species as well as the individuals of that species. Faecal pellet surveys are a key element in identifying koala habitat, and these searches have also been used to investigate elements of the ecology of this species, including tree use (Callaghan et al. 2011) and diet (Ellis et al. 2002). Tooth wear in the koala is, with some exceptions (Melzer et al. 2011), closely linked to age of the koala (Gordon 1991). By investigating the particle size distribution of these pellets, we aimed to test the potential for pellets to reveal information about the koala population age structure at locations without the need for intensive methods such as capture.
We proposed to develop a tool, based simply on fragment size in faecal pellets, that could be used to estimate the tooth wear (and hence age) of a koala. Our results were equivocal with regard to an association between particle size variation and age in the koala, and we did not find evidence among our samples to support the hypothesis that the average size of faecal particles indicated age accurately for koalas. However, we did find a significant increase in the proportion of large particles in faecal pellets from koalas with advanced tooth wear. For example, using our size classes, a koala is likely to be in age Class 7 or above if the proportion of faecal particles greater than 0.06 mm2 in area exceeds 2%. Our data provide evidence that faecal pellets produced by older koalas can be distinguished from those produced by younger koalas, on the basis of the proportion of faecal particles that are greater than 0.06 mm2.
Although the proportion of larger particles in faeces increases as tooth wear increases (Fig. 1), the distribution of particle sizes relative to the average particle size is not sufficient to distinguish tooth wear classes. Hence, the spread of particle sizes in faecal pellets does not appear to be a useful index of the age of a koala, a finding that concurs with that of Lanyon and Sanson (1986): the youngest molar wear class produced a similar ratio of small to large particles in digesta from the stomach as seen in the oldest molar wear class in their study.
Our results suggest that by focusing attention on the largest particles that we identified in the faeces of koalas, a guide to the age structure of a koala population can be produced, but the information gathered in this manner may be imprecise. While our approach to sorting particles into size classes made the basic analyses straightforward, the results indicate that paying greater attention to variation within the larger size classes should reveal more about the tooth wear of the koala that produced the pellet.
Lanyon and Sanson (1986) investigated particle size in the stomach and caecum of koalas, determining particle size distributions using a wet sieving technique. They concluded that koalas with very advanced stages of tooth wear had an increased proportion of larger particles in the stomach compared with younger koalas with minimal tooth wear. Despite this, they found that within the caecum there was only a small increase in the proportion of the largest particle size class. Therefore koalas with advanced tooth wear still exhibited some selective retention of proportionally smaller particles in the caecum, even though they were producing fewer small particles. Our study also found that koalas with advanced tooth wear were able to produce the small particles – as indicated by the similar faecal particle size distribution of old and young koalas. Hence the present study confirms the findings of Lanyon and Sanson (1986), that older koalas may have the ability to compensate for reduced masticatory effectiveness as indicated by the high proportion of small particles in the faeces of animals of all tooth wear categories.
Tooth wear not only has adverse effects on the feeding ecology of koalas, but has the potential to interfere with their physiology, behaviour and fecundity (Logan and Sanson 2002c). Logan and Sanson (2002b) found that male koalas with advanced tooth wear spent more time and effort feeding, with a daily increase in the amount of leaves consumed, and an increase in chew rate, chews per leaf, and total number of chews. This means that male koalas invested more time and energy in processing each leaf and spent less time moving between trees, resulting in a decrease in home-range size and sociality. This could reduce their reproductive effort and the number of offspring they sire, and it also explains why we may not find changes in the faecal pellet particle sizes we expected to see accompany advancing tooth wear. However, like most species, advancing age is not likely to be accompanied by a linear incremental decrease in masticatory efficiency across the life span of the koala. Tooth wear in koalas will reflect their chronological age, depending on factors such as diet over time. From the perspective of demographic forecasting, it may be more important to identify the ratio of immature, mature and old individuals in a population and this is what our attention to the larger particles and comparison of grouped toothwear classes (Fig. 1) attempted. While our method does not currently result in an accurate and repeatable index of koala age, the proportion of larger-sized particles in the faeces of koalas can be used to distinguish pellets produced by old koalas from those of younger individuals.
Acknowledgements
Queensland Department of Environment and Heritage Protection (DEHP) provided the Scientific Purposes Permits for this research (WISP00491302) and all animal procedures were carried out in accordance with University of Queensland Animal Ethics approval (ZOO/ENT/115/04/RT). Rebecca Larkin of DEHP provided valuable veterinary assistance, as did the volunteers at the Moggill Koala Hospital. Simon Blomberg assisted with statistical analyses and an anonymous reviewer provided valuable direction.
References
Callaghan, J., McAlpine, C., Thompson, J., Mitchell, D., Bowen, M., Rhodes, J., de Jong, C., Sternberg, R., and Scott, A. (2011). Ranking and mapping koala habitat quality for conservation planning on the basis of indirect evidence of tree-species use: a case study of Noosa Shire, south-eastern Queensland. Wildlife Research 38, 89–102.| Ranking and mapping koala habitat quality for conservation planning on the basis of indirect evidence of tree-species use: a case study of Noosa Shire, south-eastern Queensland.Crossref | GoogleScholarGoogle Scholar |
Caughley, G., and Gunn, A. (1993). Dynamics of large herbivores in deserts: kangaroos and caribou. Oikos 67, 47–55.
| Dynamics of large herbivores in deserts: kangaroos and caribou.Crossref | GoogleScholarGoogle Scholar |
Davies, N. A., Gillett, A., McAlpine, C., Seabrook, L., Baxter, G., Lunney, D., and Bradley, A. (2013). The effect of ACTH upon faecal glucocorticoid excretion in the koala. The Journal of Endocrinology 219, 1–12.
| The effect of ACTH upon faecal glucocorticoid excretion in the koala.Crossref | GoogleScholarGoogle Scholar |
Dique, D. S., de Villiers, D. L., and Preece, H. J. (2003). Evaluation of line-transect sampling for estimating koala abundance in the Pine Rivers Shire, south-east Queensland. Wildlife Research 30, 127–133.
| Evaluation of line-transect sampling for estimating koala abundance in the Pine Rivers Shire, south-east Queensland.Crossref | GoogleScholarGoogle Scholar |
Ellis, W., and Carrick, F. N. (1992). Total body water and the estimation of fat in the koala (Phascolarctos cinereus). Australian Veterinary Journal 69, 229–231.
| Total body water and the estimation of fat in the koala (Phascolarctos cinereus).Crossref | GoogleScholarGoogle Scholar | 1449462PubMed |
Ellis, W., Carrick, F., Lundgren, P., Veary, A., and Cohen, B. (1999). The use of faecal cuticle examination to determine the dietary composition of koalas. Australian Zoologist 31, 127–133.
Ellis, W., Melzer, A., Carrick, F. N., and Hasegawa, M. (2002). Tree use, diet and home range of the koala (Phascolarctos cinereus) at Blair Athol, central Queensland. Wildlife Research 29, 303–311.
| Tree use, diet and home range of the koala (Phascolarctos cinereus) at Blair Athol, central Queensland.Crossref | GoogleScholarGoogle Scholar |
Ellis, W., FitzGibbon, S., Melzer, A., Wilson, R., Johnston, S., Bercovitch, F., Dique, D., and Carrick, F. (2013). Koala habitat use and population density: using field data to test the assumptions of ecological models. Australian Mammalogy 35, 160–165.
| Koala habitat use and population density: using field data to test the assumptions of ecological models.Crossref | GoogleScholarGoogle Scholar |
Gordon, G. (1991). Estimation of the age of the koala, Phascolarctos cinereus (Marsupialia: Phascolarctidae) from tooth wear and growth. Australian Mammalogy 14, 5–12.
Gordon, G., McGreevy, D., and Lawrie, B. (1988). Koala populations in Queensland. In ‘Biology of the Koala’. (Eds K. Handasyde and G. Sanson.) pp. 95–95. (Surrey Beatty: Sydney.)
Lanyon, J. M., and Sanson, G. D. (1986). Koala (Phascolarctos cinereus) dentition and nutrition. 2. Implications of tooth wear in nutrition. Journal of Zoology. Series A 209, 169–181.
Logan, M. (2001). Evidence for the occurrence of rumination-like behaviour, or merycism, in the koala (Phascolarctos cinereus, Goldfuss). Journal of Zoology 255, 83–87.
| Evidence for the occurrence of rumination-like behaviour, or merycism, in the koala (Phascolarctos cinereus, Goldfuss).Crossref | GoogleScholarGoogle Scholar |
Logan, M., and Sanson, G. (2000). Measuring fine ingesta/digesta particle size distributions. Wildlife Research 27, 191–194.
| Measuring fine ingesta/digesta particle size distributions.Crossref | GoogleScholarGoogle Scholar |
Logan, M., and Sanson, G. (2002a). The effect of tooth wear on the feeding behaviour of free-ranging koalas (Phascolarctos cinereus, Goldfuss). Journal of Zoology 256, 63–69.
| The effect of tooth wear on the feeding behaviour of free-ranging koalas (Phascolarctos cinereus, Goldfuss).Crossref | GoogleScholarGoogle Scholar |
Logan, M., and Sanson, G. (2002b). The effects of tooth wear on activity patterns of free-ranging koalas (Phascolarctos cinereus Goldfuss). Australian Journal of Zoology 50, 281–292.
| The effects of tooth wear on activity patterns of free-ranging koalas (Phascolarctos cinereus Goldfuss).Crossref | GoogleScholarGoogle Scholar |
Logan, M., and Sanson, G. (2002c). The association of tooth wear with sociality of free-ranging male koalas (Phascolarctos cinereus Goldfuss). Australian Journal of Zoology 50, 621–626.
| The association of tooth wear with sociality of free-ranging male koalas (Phascolarctos cinereus Goldfuss).Crossref | GoogleScholarGoogle Scholar |
Melzer, A., Ellis, W., Gordon, G., Tucker, G., Kindness, R., and Carrick, F. (2011). Unusual patterns of tooth wear among koalas Phascolarctos cinereus from St Bees Island, Queensland, require re-evaluation of criteria for aging koalas by tooth-wear class. Australian Zoologist 35, 550–554.
| Unusual patterns of tooth wear among koalas Phascolarctos cinereus from St Bees Island, Queensland, require re-evaluation of criteria for aging koalas by tooth-wear class.Crossref | GoogleScholarGoogle Scholar |
Narayan, E., Webster, K., Nicolson, V., Mucci, A., and Hero, J. (2013). Non-invasive evaluation of physiological stress in an iconic Australian marsupial: the koala (Phascolarctos cinereus). General and Comparative Endocrinology. , .
| Non-invasive evaluation of physiological stress in an iconic Australian marsupial: the koala (Phascolarctos cinereus).Crossref | GoogleScholarGoogle Scholar | 24140710PubMed |
Phillips, S., and Callaghan, J. (2011). The Spot Assessment Technique: a tool for determining localised levels of habitat use by koalas Phascolarctos cinereus. Australian Zoologist 35, 774–780.
| The Spot Assessment Technique: a tool for determining localised levels of habitat use by koalas Phascolarctos cinereus.Crossref | GoogleScholarGoogle Scholar |
Preston, F. W. (1962). The canonical distribution of commonness and rarity: Part I. Ecology 43, 185–215.
| The canonical distribution of commonness and rarity: Part I.Crossref | GoogleScholarGoogle Scholar |
Rhodes, J., Lunney, D., Moon, C., Matthews, A., and McAlpine, C. (2011). The consequences of using indirect signs that decay to determine species’ occupancy. Ecography 34, 141–150.
| The consequences of using indirect signs that decay to determine species’ occupancy.Crossref | GoogleScholarGoogle Scholar |
Seabrook, L., McAlpine, C., Baxter, G., Rhodes, J., Bradley, A., and Lunney, D. (2011). Drought-driven change in wildlife distribution and numbers: a case study of koalas in south west Queensland. Wildlife Research 38, 509–524.
| Drought-driven change in wildlife distribution and numbers: a case study of koalas in south west Queensland.Crossref | GoogleScholarGoogle Scholar |
Skogland, T. (1988). Tooth wear by food limitation and its life history consequences in wild reindeer. Oikos 51, 238–242.
| Tooth wear by food limitation and its life history consequences in wild reindeer.Crossref | GoogleScholarGoogle Scholar |
Wedrowicz, F., Karsa, M., Mosse, J., and Hogan, F. (2013). Reliable genotyping of the koala (Phascolarctos cinereus) using DNA isolated from a single faecal pellet. Molecular Ecology Resources 13, 634–641.
| Reliable genotyping of the koala (Phascolarctos cinereus) using DNA isolated from a single faecal pellet.Crossref | GoogleScholarGoogle Scholar | 23582171PubMed |
Woosnam-Merchez, O., Cristescu, R., Dique, D., Ellis, B., Beeton, R., Simmonds, J., and Carrick, F. (2012). What faecal pellet surveys can and can’t reveal about the ecology of koalas Phascolarctos cinereus. Australian Zoologist , .
| What faecal pellet surveys can and can’t reveal about the ecology of koalas Phascolarctos cinereus.Crossref | GoogleScholarGoogle Scholar |