

Optimising camera trap height and model increases detection and individual identification rates for a small mammal, the numbat (Myrmecobius fasciatus)
Anke Seidlitz A D , Kate A. Bryant A , Nicola J. Armstrong B , Michael Calver A and Adrian F. Wayne C
A D , Kate A. Bryant A , Nicola J. Armstrong B , Michael Calver A and Adrian F. Wayne C
A Environmental and Conservation Sciences, Murdoch University, Murdoch, WA 6150, Australia.
B Mathematics and Statistics, Murdoch University, Murdoch, WA 6150, Australia.
C Department of Biodiversity, Conservation and Attractions, Locked Bag 2, Manjimup, WA 6258, Australia.
D Corresponding author. Email: anke.seidlitz@gmx.net
Australian Mammalogy 43(2) 226-234 https://doi.org/10.1071/AM20020
Submitted: 14 February 2020 Accepted: 17 June 2020 Published: 13 July 2020
Journal Compilation © Australian Mammal Society 2021 Open Access CC BY-NC-ND
Abstract
Camera traps are widely used to collect data for wildlife management, but species-specific testing is crucial. We conducted three trials to optimise camera traps for detecting numbats (Myrmecobius fasciatus), a 500–700-g mammal. We compared detection rates from (1) Reconyx PC900 camera traps installed at heights ranging from 10–45 cm, and (2) Reconyx PC900, Swift 3C standard and wide-angle camera traps with differing detection zone widths. Finally, we compared elevated, downward-angled time-lapse cameras installed at heights ranging from 1–2 m to obtain dorsal images for individual numbat identification. Camera traps set at 25 cm had the highest detection rates but missed 40% of known events. During model comparison, Swift 3C wide-angle camera traps recorded 89%, Swift 3C standard 51%, and Reconyx PC900 37% of known events. The number of suitable images from elevated, downward-angled cameras, depicting dorsal fur patterns, increased with increasing camera height. The use of well regarded camera trap brands and generic recommendations for set-up techniques cannot replace rigorous, species-specific testing. For numbat detection, we recommend the Swift 3C wide-angle model installed at 25-cm height. For individual numbat identification, elevated, downward-angled time-lapse cameras were useful; however, more research is needed to optimise this technique.
Additional keywords: comparative camera trap study, Reconyx PC900, Swift 3C, wide angle, wildlife detection.
Introduction
Camera traps are increasingly used to detect and monitor wildlife for conservation management (Rowcliffe and Carbone 2008; Burton et al. 2015; Meek et al. 2015b) so it is important to position cameras carefully to obtain robust data and to choose the most effective camera model (Meek et al. 2016; Jacobs and Ausband 2018; Apps and McNutt 2018a). Motion-activated camera traps with passive infrared (PIR) sensors trigger when objects (e.g. animals) with a different surface temperature from the background move within the detection zone (Welbourne et al. 2016). Missed detections (where animals are present but not detected) and missed opportunities to identify individual animals should be minimised. This is best achieved by species-specific optimisation of camera trap models and set-up techniques (e.g. Glen et al. 2013; Taylor et al. 2014; Meek and Vernes 2016).
The height of camera traps above the ground can influence animal detection rates. The effects of large differences in camera trap height have been investigated in several studies. Camera traps in lower positions were found to be more effective by Meek et al. (2016), who investigated instalment heights of 90 cm and 350 cm (targeting wild dogs, foxes and feral cats), and by Swann et al. (2004), testing camera traps at 20 cm and 120 cm (targeting warm water bottles and a human mimicking animal movement). While Newey et al. (2015) had better detection rates from camera traps in higher positions (120 cm versus 60 cm) when targeting sheep, Jacobs and Ausband (2018) had inconclusive results from camera traps at 60 cm and 300 cm targeting a variety of species native to Idaho, USA. In these studies, at least one of the compared camera heights substantially exceeded 50 cm, making results inconclusive for small species. The only study that tested camera-trap heights in small increments between 20 cm and 122 cm was conducted by Apps and McNutt (2018a). However, a large dog (68-cm shoulder height) was used to trigger camera-traps, making results again uninformative for small species.
As well as height, the choice of camera model is important. Detection rates may be affected by technological differences, such as trigger speed (time between animal detection and image recording) (Swann et al. 2004; Wellington et al. 2014; Meek et al. 2015a) and width of the PIR sensor detection zone (Meek et al. 2014; Wellington et al. 2014; Fancourt et al. 2018). When comparing Reconyx PC900 camera traps (40° PIR detection zone, 0.2-s trigger speed) with Ltl Acorn Ltl-5310A wide-angle camera traps (120° PIR detection zone, 0.8 s trigger speed), Fancourt et al. (2018) found that the wide-angle camera trap models had higher detection rates for rabbits. This demonstrates that it is important to consider the combination of model specifications (e.g. width of the sensor detection zone, and trigger speed) together, rather than separately. Other model differences that may affect animal detection include the camera’s software, pyroelectric sensor, and type and arrangement of Fresnel lenses (Welbourne et al. 2016; Apps and McNutt 2018b).
Camera traps may also be useful in identifying individual animals for species with unique markings, such as tigers, leopards, hyenas and Australian dasyurids (Karanth 1995; Jackson et al. 2006; Hohnen et al. 2013; Tichon et al. 2017; Rowland et al. 2020). Identifying individuals allows the use of capture–recapture modelling to obtain robust population size estimates (Royle et al. 2014). To obtain images for individual animal identification, camera traps are commonly set up horizontally with the camera facing parallel to the ground (Karanth 2002). Images from camera traps set up horizontally typically show characteristic markings from animal flanks or heads. But some species have characteristic markings only on their backs. A vertical camera trap set-up with the camera pointing vertically downward from 1.5–2 m above ground was found useful for individual recognition of skunks using dorsal fur patterns (Theimer et al. 2017). However, a vertical set-up reduces the field of view and detection zone of PIR-triggered camera traps (Smith and Coulson 2012) and therefore decreases detection rates (Taylor et al. 2014; Nichols et al. 2017). To overcome these limitations, typically baits are used to attract target species to a vertical camera station (De Bondi et al. 2010; Smith and Coulson 2012; Taylor et al. 2014; Nichols et al. 2017). However, baiting may be undesirable (e.g. if it biases study results: McCoy et al. 2011; Stewart et al. 2019) or impossible (e.g. when species are not attracted by bait: Burrows and Christensen 2002). In such cases, a different approach could be useful to overcome the field of view and detection zone restrictions experienced in a vertical camera set-up. One solution worthy of testing is to use elevated cameras (e.g. at 2 m height) with a moderate downward angle (e.g. 45°) to obtain images for individual animal identification of a species with characteristic markings on their backs.
Questions of the optimum height, appropriate camera trap model, and opportunities for individual identification were all prominent in developing monitoring strategies for the endangered Australian endemic marsupial, the numbat (Myrmecobius fasciatus), in one of its two remaining natural populations – the Upper Warren region in south-west Western Australia. Camera traps may detect elusive animals occurring in low densities (Kucera and Barrett 2011) but have not been tested for their efficiency for numbats. Since camera trap optimisation before field studies can reduce missed detections (Taylor et al. 2014), we designed this study to optimise camera traps with the use of captive numbats in zoo enclosures.
We had three aims. First, we aimed to determine the optimal PIR sensor height for camera traps to maximise detection rates for numbats, which have a body-core height of ~10–15 cm. We used the widely available Reconyx PC900 camera trap model (Glover-Kapfer et al. 2019) commonly used by researchers and government agencies in Australia (Meek et al. 2015b). Second, we aimed to determine whether detection rates from Reconyx PC900 camera traps (42° PIR detection zone) can be increased by using two models with wider detection zones (the Swift 3C standard and wide-angle; 52° and 110° PIR detection zone respectively). Third, we aimed to establish whether elevated, downward-angled cameras can be used to obtain suitable images for individual numbat identification from dorsal patterns, and to test which camera height (1 m, 1.25 m, 1.5 m, 1.75 m, or 2 m above ground) would be most appropriate. Because the main interest regarded image suitability and camera height for individual numbat identification, we used cameras set to time-lapse mode for this part of the study to avoid problems faced with restricted detection zones from downward-angled PIR motion-sensors (see Apps and McNutt 2018a). We are unaware of any other publication that evaluated optimal camera trap height for a small mammal such as the numbat, that compared detection rates from Reconyx PC900 and Swift 3C models, or that tested elevated, angled time-lapse cameras for identification of individual animals.
Materials and methods
Study species
The numbat is a small (500–700 g) diurnal marsupial (Cooper 2011) that can be individually recognised by characteristic dorsal pelage patterns. It is difficult to detect numbats because of their solitary lifestyle, large home range (which can exceed 100 ha) (Christensen et al. 1984), and elusive nature. Numbats feed exclusively on termites (Calaby 1960); there are no known lures or baits to attract numbats to traps (Burrows and Christensen 2002) so this remains a topic for investigation. The numbat occurs today in only 1% of its former range, which covered much of the southern half of the Australian continent before European settlement (Friend 1990). It is listed as endangered nationally under Australia’s Environment Protection and Biodiversity Conservation Act 1999, and internationally under the IUCN Red List of Threatened Species (Friend and Burbidge 2008). Numbats occur now in several reintroduction sites (see details in Department of Parks and Wildlife 2017), and in two remaining natural populations at Dryandra Woodland and the Upper Warren region, both located in south-west Western Australia. There is a paucity of adequate monitoring information for the Upper Warren region (see Wayne 2018 for area map) due to the lack of effective survey methods to assess population trends or size.
Perth Zoo and enclosures
This study was conducted in a zoo environment to ensure numbat detection rates were sufficiently high for a comparative camera study. Perth Zoo in Western Australia keeps numbats in captivity as part of a breeding program for reintroduction purposes (Mawson and Lambert 2017). Two types of fenced numbat enclosures were used: one L-shaped enclosure (8 m × 11 m plus 4 m × 6 m) and three rectangular enclosures (5 m × 3 m). Each enclosure contained areas of bare ground, native vegetation, nesting and climbing structures, feeding areas, and a single adult numbat. The three trials (PIR motion-sensor height; comparison of camera trap models; time-lapse camera height for identification of individual animals) took place between June and September 2017.
Cameras tested
We tested the following camera models: Reconyx PC900 and HC600 (RECONYX, LLP, Holmen, WI, USA), and the Swift 3C standard and wide-angle model (Outdoor Cameras Australia, Toowoomba, Qld, Australia). We had intended to use only the Reconyx PC900 model, but one HC600 model was provided by error in the delivery of cameras. It was included in the trial because differences between PC900 and HC600 models are predominantly related to software functions, with the camera trap settings required for our trials available in both models. PC900 and HC600 models could therefore be treated as equivalent. The Swift 3C is a distributer-branded, low-cost camera trap. Details on camera specifications and accessories are given in Table 1. We chose Reconyx PC900 and HC600 camera traps because they are commonly used by the Department of Biodiversity, Conservation and Attractions for camera trap studies in the Upper Warren region. Swift 3C camera traps were chosen because they are available as standard and wide-angle models. The Reconyx cameras were used in assessments of the ideal height of camera traps for numbat detection and for identifying individual numbats, while both Reconyx and Swift cameras were used to determine differences in detection rates between camera trap models.

|
PIR motion-sensor height
To determine which camera trap height is most suitable for numbat detection, 12 Reconyx camera traps (11 × PC900 and 1 × HC600 models) were deployed in three rectangular numbat enclosures. In each of the three enclosures, four camera traps were stacked vertically, and firmly secured to a wooden plank with their PIR motion-sensors positioned at 10 cm, 25 cm, 40 cm, and 55 cm height above ground (Fig. 1a). Camera trap lenses were aimed horizontally, facing the ~2.5-m-distant central area of the enclosures. Camera traps were set to take three pictures at 1-s intervals per trigger, and with a quiet period of 15 s between triggers. The 15-s quiet period was used to reduce multiple detections of numbats remaining in their favourite areas for an extended time within the detection zone. PIR motion-sensor sensitivity was set to high. Camera traps were set to operate for four days; however, one Reconyx PC900 camera, positioned at 10 cm, failed to operate from the morning of Day 4 for unknown reasons. The Day-4 data of all camera traps in that enclosure were therefore excluded from the analysis.

|
In addition to the above, a time-lapse camera taking a picture every 2 s, was installed in each enclosure (Reconyx PC900 model set to time-lapse mode with motion detection disabled). The time-lapse cameras were aimed in the same general direction as the camera traps and were installed at 20 cm height. Time-lapse cameras were used to approximate numbat movements in front of cameras that were missed by the camera traps because they take pictures at set intervals independent of animal movements.
Comparison of camera trap models
To determine differences in detection rates between camera trap models with different detection zone width, three different camera traps (see Table 1) were attached side-by-side to a wooden plank ~2 cm apart (Fig. 1b). Initial camera positions were determined randomly, and, within enclosures, camera traps were rotated daily so that each camera occupied each position twice. The camera traps operated over six days in three rectangular numbat enclosures. Camera traps were aimed horizontally towards the centre of the enclosure with their PIR motion-sensors positioned at ~25 cm above ground. Cameras were set to take three images per trigger with no quiet period between triggers to maximise possible detections. We did not use a 15-s quiet period between triggers because camera trap positions were rotated daily. Therefore, cameras were not advantaged/disadvantaged by constant positioning (e.g. facing to/away from frequently visited enclosure areas). Reconyx PC900 models were set to have a 1-s interval between pictures. This was to approximate the picture recording interval time of Swift 3C models, which do not allow adjustment of this function. PIR motion-sensor sensitivity of all camera traps was set to high.
As with the height trial, a Reconyx PC900 camera in time-lapse mode was installed in each enclosure. The time-lapse cameras, taking an image every 2 s, were positioned centrally, below the camera traps at ~10 cm above ground, and aimed in the same general direction. This time-lapse camera placement was chosen to maximise the field-of-view overlap with the camera traps. A positioning above the camera traps would have caused the time-lapse camera to miss numbats moving close to the camera station.
Time-lapse camera height for identification of individual animals
To determine the most suitable camera height above ground for obtaining dorsal images for identification of individual numbats, five Reconyx PC900 cameras were installed in each of three numbat enclosures (two rectangular and one L-shaped enclosure). Cameras were stacked vertically and attached to wooden poles, at 1 m, 1.25 m, 1.5 m, 1.75 m, and 2 m height (Fig. 1c). Maximum camera height was limited by enclosure height. Cameras were angled to centre images on a point on the ground ~2 m from the attachment pole. Cameras were set to time-lapse mode, taking one image every 2 s from 0800 to 1600 hours, which is when numbat activity was most likely. Batteries and SD cards were changed daily before 0800 hours. The time-lapse cameras operated over eight days to allow enough time to collect sufficient identifications of individual numbats for comparison.
Data collection and analysis
For the ‘PIR motion-sensor height’ and the ‘Comparison of camera trap models’ trials, camera time and date settings were synchronised during camera set-up and were visible on each image. This allowed direct comparison of images from camera traps and time-lapse cameras. Images were viewed using FastStone Image Viewer 6.2 (FastStone Soft 2019). We compared the number of numbat detections (also referred to as detection rates) from camera traps and time-lapse cameras. We defined a numbat movement in front of cameras as an event, and a detection as an event being recorded, resulting in an image depicting a numbat partially or wholly, by at least one camera trap or time-lapse camera. Using the synchronised time and date stamps on images for reference, each event was recorded as either detected or missed for each camera. Since all detections in each enclosure were of the same animal, we did not use a quiet period between successive detections which is often used in camera trap studies to ensure independence of animal detections (see Meek et al. 2014 for detailed definition).
For the third trial, time-lapse images were scanned for images depicting numbats. A numbat movement within the field-of-view of time-lapse cameras resulted in sequences of images depicting that numbat. If a sequence of images contained one or more clear images of the numbat’s dorsal pelage pattern, and thereby allowing individual numbat identification, then the identification was recorded and counted. Identifications were totalled for each time-lapse camera per day.
Numbat detections or individual identifications from the different trials were analysed using linear regression models in R 3.5.0 (R Core Team 2018). Diagnostic plots for linear regression analysis were inspected to ensure model fit. Models were fitted with the main covariates of interest ‘camera trap height’, ‘camera trap model’, and ‘time-lapse camera height’ for the three trials respectively. Since different numbats in each enclosure may have had different activity levels, and because those activity levels may have changed each day, the covariates ‘enclosure’ and ‘day’ were used to improve model fit. Both covariates were treated as fixed to investigate the effects of the different days while the number of enclosures was too small to allow it to be random. Predictive models were compared using the AIC (Akaike Information Criterion: Akaike 1974) function in R, and models with the lowest values were selected for further analysis (Burnham and Anderson 2002). Further analysis comprised a Tukey’s HSD test to compare all possible pairs of means (Tukey 1949).
Results
PIR motion-sensor height trial
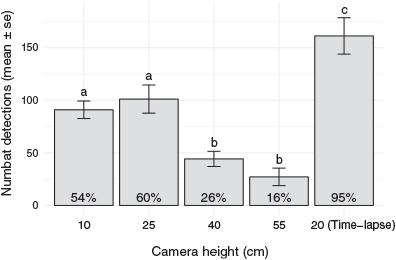
There were 1869 events where at least one camera trap or time-lapse camera recorded a numbat moving in front of a camera. Most observed events were recorded by cameras at 25 cm above ground (60%), while cameras installed at 55 cm recorded the lowest percentage (16%) of events (Fig. 2). The time-lapse cameras in each enclosure recorded 95% of observed events. Camera trap height was found to be significantly associated with the number of detections (F14,40 = 18.91, P < 0.0001). The initial model, which included the explanatory variable ‘camera height’ only, had an AIC of 564.08. The value of the AIC decreased with the addition of ‘enclosure’ (546.49), ‘day’ (551.96), ‘enclosure+day’ (534.22), and ‘enclosure*day’ (526.20). Therefore, the model with the interaction term was used as the final model. Camera traps installed at 10 cm and 25 cm had significantly higher mean detection rates than those at 40 cm and 55 cm (Fig. 2).
|
Comparison of camera trap models
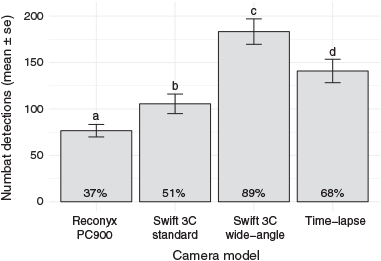
Cameras recorded a total of 3703 events where at least one camera trap or time-lapse camera recorded a numbat movement. The Swift 3C wide-angle model recorded the highest percentage (89%), and the Reconyx PC900 model the lowest percentage (37%) of observed events (Fig. 3). Detection rates of time-lapse cameras were lower than those of the Swift 3C wide-angle camera trap model. Camera trap models were significantly associated with the number of detections (F20,51 = 17.53, P < 0.0001). The initial model, which included the explanatory variable ‘camera model’ only, had an AIC of 766.45. The value of the AIC decreased with the addition of ‘enclosure’ (750.49), ‘day’ (753.90), ‘enclosure+day’ (728.93), and ‘enclosure*day’ (691.93). Therefore, the model with the interaction term was used as the final model. Swift 3C wide-angle camera traps had significantly more mean detections than time-lapse cameras, Swift 3C standard camera traps and Reconyx PC900 camera traps (Fig. 3).
|
Time-lapse camera height for identification of individual animals
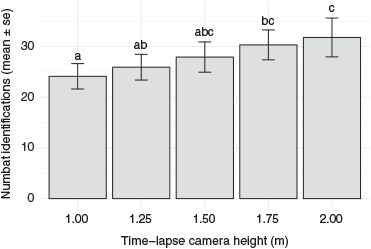
Images depicting the numbats’ characteristic dorsal pelage patterns (Fig. 4) allowed a total of 2102 individual identifications. Camera height was found to be significantly associated with the number of individual identifications (F18,56 = 26.14, P < 0.0001). The initial model, which included the explanatory variable ‘camera height’ only, had an AIC of 587.67. The value of the AIC decreased with the addition of ‘enclosure’ (582.41), ‘day’ (541.83), ‘enclosure+day’ (525.45), and ‘enclosure*day’ (452.11). As with the other trials, the model with the interaction term was used as the final model. The number of individual numbat identifications differed between camera trap heights, with cameras at 2 m height having significantly more identifications than those at 1 m and 1.25 m height (Fig. 5).

|
|
Discussion
Camera trap optimisation trials are a critical preparational step for camera trap studies. Such trials can help optimise camera trap set-up techniques and identify limitations. If convenient situations (such as captive colonies) exist and the trial is well designed, informative results can be obtained within a few days. If camera trap limitations are overlooked, animal population data from camera trap studies may be inadequate or flawed and misinform management. We conducted three trials that (1) identified a suitable camera trap height for numbat detection, (2) allowed us to significantly improve detection rates by choosing a better camera trap model, and (3) informed us that elevated, downward-angled time-lapse cameras can be used to identify numbat individuals. The results are applicable to monitoring of the numbat and potentially to other small mammals.
PIR motion-sensor height trial
It is recommended that PIR sensors of camera traps be aimed at the body core of target species to best detect the animal’s surface heat and movement (Meek et al. 2012; Wearn and Glover-Kapfer 2017). The detection rates observed in this study (where the target had a core body height of ~10–15 cm) confirmed this and show that a slightly higher sensor placement (e.g. 25 cm) is acceptable and possibly advantageous, especially when near-ground obstructions such as vegetation and undulating ground may restrict the camera’s detection zone. Vegetation (grass) was found to obscure camera traps set close to the ground in a trial by Glen et al. (2013) of stoats (Mustela erminea), cats (Felis catus) and hedgehogs (Erinaceus europaeus). Even though we used Reconyx PC900 camera traps in this study, we are confident our results are applicable to other camera trap models with the same PIR sensor types, such as Swift 3C camera traps. While near-ground obstructions are likely to be encountered in the field, possibly obstructing camera traps installed in low positions (e.g. 10 cm), an installation above 25 cm could allow small mammals to stay undetected when passing close to the camera traps, below the detection zone. We therefore recommend an installation height of ~25 cm above ground for small mammals similar to numbats, when using camera traps with PIR sensors similar to those of Reconyx PC900 camera traps.
While exploring differences in detection rates of camera traps at different heights, it was established that the Reconyx PC900 camera traps at the preferred height (25 cm) missed 40% of all known events. This highlights the importance of testing camera traps on target species to learn about potential limitations such as missed events. The number of missed events in camera trap studies is seldom known, and can only be approximated when animal movements are controlled (e.g. Apps and McNutt (2018a) used monitored dog movements to trigger camera traps) or when time-lapse or continuously recording video cameras are used concurrently with camera traps (Glen et al. 2013; Newey et al. 2015; Jumeau et al. 2017; Urbanek et al. 2019). Similar to this study, Jumeau et al. (2017) found in a study of small animals at wildlife underpasses that Reconyx HC600 camera traps missed 43% of mammal movements recorded from continuous video cameras. In contrast, Reconyx RC55 camera trap models missed only 5% of animal visits recorded by a video camera (Dixon et al. 2009). The Reconyx RC55 has the same fast trigger speed (0.2 s) and PIR motion-sensor zone (40°) as the PC900 and HC600, so these models should perform similarly. Perhaps the higher detection rates in the study by Dixon et al. (2009) are due to a difference in environment or study animal. One clear difference in that study is that cameras were aimed at bait stations that attract animals to stay for extended periods within the camera trap’s detection zone, increasing the probability of detection (Gil-Sánchez et al. 2011; Monterroso et al. 2011; Rovero et al. 2013). As there are no known baits for numbats (Burrows and Christensen 2002), it was important to seek a more suitable camera trap model to reduce the missed detections experienced in this study with Reconyx PC900 models.
Comparison of camera trap models
By using camera traps with wider detection zones, we significantly increased the detection rates achieved by the Reconyx PC900 camera trap model. The Swift 3C wide-angle camera traps (110° detection zone) detected 89%, while the Reconyx PC900 models (40° detection zone) detected 37% of all known events. With a wider detection zone, the Swift 3C wide-angle camera traps also detected more events than the time-lapse camera (Reconyx PC900 model) set to 2-s intervals. These results are important to future camera trap studies of numbats and similar species. Without this preparatory camera trap trial, unawareness of camera trap limitations could have resulted in missing >60% of possible numbat detections. These limitations would likely substantially compromise the power and confidence in the inferences, and therefore the value of subsequent studies. While performance differences between camera traps of different brands may derive from multiple factors other than detection zone width (e.g. type and arrangement of Fresnel lenses), the detection rates of the Swift 3C standard model (52° detection zone), which detected 51% of known events show that the width of PIR sensor detection zones alone can significantly affect detection rates as the Swift 3C standard and wide-angle models are otherwise identical. While increased detection rates of Swift 3C wide-angle cameras were expected, given that this camera model effectively samples a larger area, we acknowledge that differences may have been inflated by possible non-random movements of numbats in enclosures. For example, this might occur if numbats preferred running along fence lines, which were within the wide-angle cameras’ but outside the standard cameras’ field of view. The results of this trial and the study of Fancourt et al. (2018), who also found improved detection rates from camera traps with wider detection zones, may increase awareness of the importance of PIR detection-zone width in wildlife studies. Camera traps with wider detection zones than standard models may increase detection rates in a wide range of animals.
Time-lapse camera height for identification of individual animals
We have shown that individual numbat identification is possible from images derived from elevated, downward-angled cameras, and that the number of images depicting dorsal fur patterns increased with increasing camera height. Optimal camera height depends on several factors. Increasing camera height increases the field of view, and therefore the sample area. There will also be an optimal distance range for cameras to detect animals within the field of view. An increased height also may allow a better perspective of the animal’s dorsal area, improving identification opportunities. However, an increase in camera height results in a decrease in image quality as the distance to the animal increases. These factors must be understood and balanced for optimal results, with due consideration for operator safety if climbing is required to position cameras. Here we have used time-lapse cameras but there are challenges with the use of time-lapse cameras in the field. When set to take images at very short intervals (e.g. 5 s or less), time-lapse cameras have high energy and image storage requirements, necessitating either frequent servicing and/or large battery and data storage capacities. Both may not be practical and/or feasible for field studies. Using motion-activated camera traps with inbuilt PIR sensors would solve these problems; however, aiming PIR sensors downward reduces the camera’s detection zone (see fig. 3 in Apps and McNutt 2018b). A reduced detection zone may not hinder animal detection when cameras can be aimed at areas they visit (e.g. recurrently used pathways) or when animals can be attracted to the camera station by bait. Neither option applies to numbats. A possible solution for numbat studies could be the use of external PIR motion sensors that trigger cameras wirelessly or by cable attachment. This would allow setting camera traps at elevated, angled positions while placing the motion sensor at 25-cm height facing parallel to the ground. Further trials are required to explore this option.
Limitations
This study did not evaluate the long-term reliability, usability and cost-effectiveness of camera trap models. Also, image quality was not formally assessed; however, we deem image quality of models tested here generally as suitable for identifying animals to species level in field situations, as well as identifying numbat individuals from pelage patterns when the animal is captured within 2 m from the camera. It is likely that numbat individuals could be identified at further distances; however, we were not able to reliably determine this due to limitations in the zoo enclosures (e.g. vegetation restricting the view of camera traps). Additionally, the number of false triggers (images that did not result from numbat presence, e.g. detection of moving vegetation) per camera trap was not evaluated. False triggers were low in this study in zoo enclosures. Further trials are needed to validate and extend results from this study under field conditions.
Conclusions
Animal population data from camera trap studies are widely used to inform wildlife management. Camera trap studies can be substantially improved by first conducting well designed, short camera trap trials. Such trials help to optimise camera trap set-up, identify limitations, improve animal detection rates, and therefore lead to better outcomes from camera studies for species management. Positioning camera traps with their PIR sensor 25 cm above ground and using Swift 3C wide-angle camera traps has been shown here to significantly increase the detection of numbats. Further research is recommended to enhance techniques for individual numbat identification using their characteristic dorsal pelage patterns, which was shown to be possible by using elevated, downward-angled time-lapse cameras.
Conflicts of interest
The authors declare no conflicts of interest.
Acknowledgements
This study was conducted under permit NS2951/17 of the Murdoch University Animal Ethics Committee, and project number 2017-4 of the Perth Zoological Parks Authority Research and Animal Ethics Committees. We thank Perth Zoo and staff members for their invaluable assistance and support. We are very grateful for the loans of cameras from WWF-Australia, Perth, Western Australia and from Outdoor Cameras Australia, Toowoomba, Queensland. Furthermore, we thank WWF-Australia for obtaining and managing project funds. This project was supported by funding from the Western Australian Government’s State NRM Program (A16002) and Murdoch University.
References
Akaike, H. (1974). A new look at the statistical model identification. IEEE Transactions on Automatic Control 19, 716–723.| A new look at the statistical model identification.Crossref | GoogleScholarGoogle Scholar |
Apps, P., and McNutt, J. W. (2018a). Are camera traps fit for purpose? A rigorous, reproducible and realistic test of camera trap performance. African Journal of Ecology 56, 710–720.
| Are camera traps fit for purpose? A rigorous, reproducible and realistic test of camera trap performance.Crossref | GoogleScholarGoogle Scholar |
Apps, P. J., and McNutt, J. W. (2018b). How camera traps work and how to work them. African Journal of Ecology 56, 702–709.
| How camera traps work and how to work them.Crossref | GoogleScholarGoogle Scholar |
Burnham, K. P., and Anderson, D. R. (2002). ‘Model Selection and Multimodel Inference: A Practical Information-Theoretic Approach.’ 2nd edn. (Springer: New York.)
Burrows, N. D., and Christensen, P. E. S. (2002). Long-term trends in native mammal capture rates in a jarrah forest in south-western Australia. Australian Forestry 65, 211–219.
| Long-term trends in native mammal capture rates in a jarrah forest in south-western Australia.Crossref | GoogleScholarGoogle Scholar |
Burton, A. C., Neilson, E., Moreira, D., Ladle, A., Steenweg, R., Fisher, J. T., Bayne, E., and Boutin, S. (2015). Wildlife camera trapping: a review and recommendations for linking surveys to ecological processes. Journal of Applied Ecology 52, 675–685.
| Wildlife camera trapping: a review and recommendations for linking surveys to ecological processes.Crossref | GoogleScholarGoogle Scholar |
Calaby, J. H. (1960). Observations on the banded ant-eater Myrmecobius f. fasciatus Waterhouse (Marsupialia), with particular reference to its food habits. Proceedings of the Zoological Society of London 135, 183–207.
| Observations on the banded ant-eater Myrmecobius f. fasciatus Waterhouse (Marsupialia), with particular reference to its food habits.Crossref | GoogleScholarGoogle Scholar |
Christensen, P., Maisey, K., and Perry, D. (1984). Radiotracking the numbat, Myrmecobius fasciatus, in the Perup Forest of Western Australia. Wildlife Research 11, 275–288.
| Radiotracking the numbat, Myrmecobius fasciatus, in the Perup Forest of Western Australia.Crossref | GoogleScholarGoogle Scholar |
Cooper, C. E. (2011). Myrmecobius fasciatus (Dasyuromorphia: Myrmecobiidae). Mammalian Species 43, 129–140.
| Myrmecobius fasciatus (Dasyuromorphia: Myrmecobiidae).Crossref | GoogleScholarGoogle Scholar |
De Bondi, N., White, J. G., Stevens, M., and Cooke, R. (2010). A comparison of the effectiveness of camera trapping and live trapping for sampling terrestrial small-mammal communities. Wildlife Research 37, 456–465.
| A comparison of the effectiveness of camera trapping and live trapping for sampling terrestrial small-mammal communities.Crossref | GoogleScholarGoogle Scholar |
Department of Parks and Wildlife (2017). Numbat (Myrmecobius fasciatus) Recovery Plan. Prepared by J. A. Friend and M. J. Page. Wildlife Management Program No. 60, Department of Parks and Wildlife, Perth.
Dixon, V., Glover, H. K., Winnell, J., Treloar, S. M., Whisson, D. A., and Weston, M. A. (2009). Evaluation of three remote camera systems for detecting mammals and birds. Ecological Management & Restoration 10, 156–158.
| Evaluation of three remote camera systems for detecting mammals and birds.Crossref | GoogleScholarGoogle Scholar |
Fancourt, B. A., Sweaney, M., and Fletcher, D. B. (2018). More haste, less speed: pilot study suggests camera trap detection zone could be more important than trigger speed to maximise species detections. Australian Mammalogy 40, 118–121.
| More haste, less speed: pilot study suggests camera trap detection zone could be more important than trigger speed to maximise species detections.Crossref | GoogleScholarGoogle Scholar |
FastStone Soft (2019). FastStone Image Viewer for Windows. Available at: www.faststone.org [accessed 20 February 2019].
Friend, J. A. (1990). The numbat Myrmecobius fasciatus (Myrmecobiidae): history of decline and potential for recovery. Proceedings of the Ecological Society of Australia 16, 369–377.
Friend, T., and Burbidge, A. (2008). Myrmecobius fasciatus. The IUCN Red List of Threatened Species 2016: e.T14222A21949380. Available at: http://dx.doi.org/10.2305/IUCN.UK.2016-2.RLTS.T14222A21949380.en [accessed 15 October 2019].
Gil-Sánchez, J. M., Moral, M., Bueno, J., Rodríguez-Siles, J., Lillo, S., Pérez, J., Martín, J. M., Valenzuela, G., Garrote, G., Torralba, B., and Simón-Mata, M. Á. (2011). The use of camera trapping for estimating Iberian lynx (Lynx pardinus) home ranges. European Journal of Wildlife Research 57, 1203–1211.
| The use of camera trapping for estimating Iberian lynx (Lynx pardinus) home ranges.Crossref | GoogleScholarGoogle Scholar |
Glen, A. S., Cockburn, S., Nichols, M., Ekanayake, J., and Warburton, B. (2013). Optimising camera traps for monitoring small mammals. PLoS One 8, .
| Optimising camera traps for monitoring small mammals.Crossref | GoogleScholarGoogle Scholar | 24039978PubMed |
Glover-Kapfer, P., Soto-Navarro, C. A., and Wearn, O. R. (2019). Camera-trapping version 3.0: current constraints and future priorities for development. Remote Sensing in Ecology and Conservation 5, 209–223.
| Camera-trapping version 3.0: current constraints and future priorities for development.Crossref | GoogleScholarGoogle Scholar |
Hohnen, R., Ashby, J., Tuft, K., and McGregor, H. (2013). Individual identification of northern quolls (Dasyurus hallucatus) using remote cameras. Australian Mammalogy 35, 131–135.
| Individual identification of northern quolls (Dasyurus hallucatus) using remote cameras.Crossref | GoogleScholarGoogle Scholar |
Jackson, R. M., Roe, J. D., Wangchuk, R., and Hunter, D. O. (2006). Estimating snow leopard population abundance using photography and capture–recapture techniques. Wildlife Society Bulletin 34, 772–781.
| Estimating snow leopard population abundance using photography and capture–recapture techniques.Crossref | GoogleScholarGoogle Scholar |
Jacobs, C. E., and Ausband, D. E. (2018). An evaluation of camera trap performance – what are we missing and does deployment height matter? Remote Sensing in Ecology and Conservation 4, 352–360.
| An evaluation of camera trap performance – what are we missing and does deployment height matter?Crossref | GoogleScholarGoogle Scholar |
Jumeau, J., Petrod, L., and Handrich, Y. (2017). A comparison of camera trap and permanent recording video camera efficiency in wildlife underpasses. Ecology and Evolution 7, 7399–7407.
| A comparison of camera trap and permanent recording video camera efficiency in wildlife underpasses.Crossref | GoogleScholarGoogle Scholar | 28944025PubMed |
Karanth, K. U. (1995). Estimating tiger Panthera tigris populations from camera-trap data using capture–recapture models. Biological Conservation 71, 333–338.
| Estimating tiger Panthera tigris populations from camera-trap data using capture–recapture models.Crossref | GoogleScholarGoogle Scholar |
Karanth, K. U. (2002). ‘Monitoring Tigers and Their Prey: A Manual for Wildlife Researchers, Managers and Conservationists in Tropical Asia.’ (Centre for Wildlife Studies: Bangalore, India.)
Kucera, T. E., and Barrett, R. H. (2011). A history of camera trapping. In ‘Camera Traps in Animal Ecology: Methods and Analyses’. (Eds A. F. O’Connell, J. D. Nichols, and K. U. Karanth.) pp. 9–26. (Springer: New York.)
Mawson, P. R., and Lambert, C. (2017). Challenges of operating a multi-species breeding-for-release facility at Perth Zoo, Australia. International Zoo Yearbook 51, 165–174.
| Challenges of operating a multi-species breeding-for-release facility at Perth Zoo, Australia.Crossref | GoogleScholarGoogle Scholar |
McCoy, J. C., Ditchkoff, S. S., and Steury, T. D. (2011). Bias associated with baited camera sites for assessing population characteristics of deer. Journal of Wildlife Management 75, 472–477.
| Bias associated with baited camera sites for assessing population characteristics of deer.Crossref | GoogleScholarGoogle Scholar |
Meek, P. D., and Vernes, K. (2016). Can camera trapping be used to accurately survey and monitor the Hastings River mouse (Pseudomys oralis)? Australian Mammalogy 38, 44–51.
| Can camera trapping be used to accurately survey and monitor the Hastings River mouse (Pseudomys oralis)?Crossref | GoogleScholarGoogle Scholar |
Meek, P. D., Ballard, G., and Fleming, P. (2012). An introduction to camera trapping for wildlife surveys in Australia. PestSmart Toolkit publication, Invasive Animals Cooperative Research Centre, Canberra.
Meek, P. D., Ballard, G., Claridge, A., Kays, R., Moseby, K., O’Brien, T., O’Connell, A., Sanderson, J., Swann, D. E., Tobler, M., and Townsend, S. (2014). Recommended guiding principles for reporting on camera trapping research. Biodiversity and Conservation 23, 2321–2343.
| Recommended guiding principles for reporting on camera trapping research.Crossref | GoogleScholarGoogle Scholar |
Meek, P. D., Ballard, G.-A., and Fleming, P. J. S. (2015a). The pitfalls of wildlife camera trapping as a survey tool in Australia. Australian Mammalogy 37, 13–22.
| The pitfalls of wildlife camera trapping as a survey tool in Australia.Crossref | GoogleScholarGoogle Scholar |
Meek, P. D., Ballard, G.-A., Vernes, K., and Fleming, P. J. S. (2015b). The history of wildlife camera trapping as a survey tool in Australia. Australian Mammalogy 37, 1–12.
| The history of wildlife camera trapping as a survey tool in Australia.Crossref | GoogleScholarGoogle Scholar |
Meek, P. D., Ballard, G. A., and Falzon, G. (2016). The higher you go the less you will know: placing camera traps high to avoid theft will affect detection. Remote Sensing in Ecology and Conservation 2, 204–211.
| The higher you go the less you will know: placing camera traps high to avoid theft will affect detection.Crossref | GoogleScholarGoogle Scholar |
Monterroso, P., Alves, P. C., and Ferreras, P. (2011). Evaluation of attractants for non-invasive studies of Iberian carnivore communities. Wildlife Research 38, 446–454.
| Evaluation of attractants for non-invasive studies of Iberian carnivore communities.Crossref | GoogleScholarGoogle Scholar |
Newey, S., Davidson, P., Nazir, S., Fairhurst, G., Verdicchio, F., Irvine, R. J., and van der Wal, R. (2015). Limitations of recreational camera traps for wildlife management and conservation research: a practitioner’s perspective. Ambio 44, 624–635.
| Limitations of recreational camera traps for wildlife management and conservation research: a practitioner’s perspective.Crossref | GoogleScholarGoogle Scholar | 26508349PubMed |
Nichols, M., Glen, A. S., Garvey, P., and Ross, J. (2017). A comparison of horizontal versus vertical camera placement to detect feral cats and mustelids. New Zealand Journal of Ecology 41, 145–150.
| A comparison of horizontal versus vertical camera placement to detect feral cats and mustelids.Crossref | GoogleScholarGoogle Scholar |
R Core Team (2018). R: a language and environment for statistical computing. Available at: http://www.r-project.org [accessed 20 December 2019].
Rovero, F., Zimmermann, F., Berzi, D., and Meek, P. (2013). “Which camera trap type and how many do I need?” A review of camera features and study designs for a range of wildlife research applications. Hystrix, the Italian Journal of Mammalogy 24, 148–156.
| “Which camera trap type and how many do I need?” A review of camera features and study designs for a range of wildlife research applications.Crossref | GoogleScholarGoogle Scholar |
Rowcliffe, J. M., and Carbone, C. (2008). Surveys using camera traps: are we looking to a brighter future? Animal Conservation 11, 185–186.
| Surveys using camera traps: are we looking to a brighter future?Crossref | GoogleScholarGoogle Scholar |
Rowland, J., Hoskin, C. J., and Burnett, S. (2020). Camera traps are an effective method for identifying individuals and determining the sex of spotted-tailed quolls (Dasyurus maculatus gracilis). Australian Mammalogy , .
| Camera traps are an effective method for identifying individuals and determining the sex of spotted-tailed quolls (Dasyurus maculatus gracilis).Crossref | GoogleScholarGoogle Scholar |
Royle, J. A., Chandler, R. B., Sollmann, R., and Gardner, B. (2014). ‘Spatial Capture–Recapture.’ (Elsevier Academic Press: Oxford.)
Smith, J. K., and Coulson, G. (2012). A comparison of vertical and horizontal camera trap orientations for detection of potoroos and bandicoots. Australian Mammalogy 34, 196–201.
| A comparison of vertical and horizontal camera trap orientations for detection of potoroos and bandicoots.Crossref | GoogleScholarGoogle Scholar |
Stewart, F. E. C., Volpe, J. P., and Fisher, J. T. (2019). The debate about bait: a red herring in wildlife research. Journal of Wildlife Management 83, 985–992.
| The debate about bait: a red herring in wildlife research.Crossref | GoogleScholarGoogle Scholar |
Swann, D. E., Hass, C. C., Dalton, D. C., and Wolf, S. A. (2004). Infrared-triggered cameras for detecting wildlife: an evaluation and review. Wildlife Society Bulletin 32, 357–365.
| Infrared-triggered cameras for detecting wildlife: an evaluation and review.Crossref | GoogleScholarGoogle Scholar |
Taylor, B. D., Goldingay, R. L., and Lindsay, J. M. (2014). Horizontal or vertical? Camera trap orientations and recording modes for detecting potoroos, bandicoots and pademelons. Australian Mammalogy 36, 60–66.
| Horizontal or vertical? Camera trap orientations and recording modes for detecting potoroos, bandicoots and pademelons.Crossref | GoogleScholarGoogle Scholar |
Theimer, T. C., Ray, D. T., and Bergman, D. L. (2017). Camera angle and photographic identification of individual striped skunks. Wildlife Society Bulletin 41, 146–150.
| Camera angle and photographic identification of individual striped skunks.Crossref | GoogleScholarGoogle Scholar |
Tichon, J., Rotem, G., and Ward, P. (2017). Estimating abundance of striped hyenas (Hyaena hyaena) in the Negev Desert of Israel using camera traps and closed capture–recapture models. European Journal of Wildlife Research 63, 5.
| Estimating abundance of striped hyenas (Hyaena hyaena) in the Negev Desert of Israel using camera traps and closed capture–recapture models.Crossref | GoogleScholarGoogle Scholar |
Tukey, J. W. (1949). Comparing individual means in the analysis of variance. Biometrics 5, 99–114.
| Comparing individual means in the analysis of variance.Crossref | GoogleScholarGoogle Scholar | 18151955PubMed |
Urbanek, R. E., Ferreira, H. J., Olfenbuttel, C., Dukes, C. G., and Albers, G. (2019). See what you’ve been missing: an assessment of Reconyx® PC900 Hyperfire cameras. Wildlife Society Bulletin 43, 630–638.
| See what you’ve been missing: an assessment of Reconyx® PC900 Hyperfire cameras.Crossref | GoogleScholarGoogle Scholar |
Wayne, A. (2018). Insights from multi-species mammal monitoring programs in the Upper Warren, Western Australia. In ‘Monitoring Threatened Species and Ecological Communities’. (Eds S. Legge, D. B. Lindenmayer, N. M. Robinson, B. C. Scheele, D. M. Southwell, and B. A. Wintle.) pp. 179–192. (CSIRO Publishing: Melbourne.)
Wearn, O. R., and Glover-Kapfer, P. (2017). Camera-trapping for conservation: a guide to best-practices. WWF Conservation Technology, Series 1, Woking, UK.
Welbourne, D. J., Claridge, A. W., Paull, D. J., and Lambert, A. (2016). How do passive infrared triggered camera traps operate and why does it matter? Breaking down common misconceptions. Remote Sensing in Ecology and Conservation 2, 77–83.
| How do passive infrared triggered camera traps operate and why does it matter? Breaking down common misconceptions.Crossref | GoogleScholarGoogle Scholar |
Wellington, K., Bottom, C., Merrill, C., and Litvaitis, J. A. (2014). Identifying performance differences among trail cameras used to monitor forest mammals. Wildlife Society Bulletin 38, 634–638.
| Identifying performance differences among trail cameras used to monitor forest mammals.Crossref | GoogleScholarGoogle Scholar |