

Feasibility of nocturnal thermal drone surveys for detecting endangered mahogany gliders (Petaurus gracilis) in tropical lowland woodlands
Emmeline Bernadette Barrett Norris A * , Yiyin Chang B , Stephen Jackson C D E and Conrad J. Hoskin B
A * , Yiyin Chang B , Stephen Jackson C D E and Conrad J. Hoskin B
A
B
C
D
E
Abstract
Reliable detection methods are essential for monitoring threatened species. Yet detection remains challenging for low-density populations of nocturnal, canopy-dwelling mammals. The endangered mahogany glider (Petaurus gracilis), endemic to lowland woodlands in north Queensland (Qld), Australia, is difficult to survey using conventional survey methods, which can be labour-intensive and yield low detection rates. We tested the feasibility of using thermal drones for detecting mahogany gliders by conducting four consecutive nocturnal flights over a 64-ha woodland fragment that supports a high-density population. Six individuals were detected within 2 h while flying the drone 10–30 m above the canopy and using oblique camera angles. We identified gliders in thermal imagery by their size, long tail and gliding behaviour, with no visible disturbance observed. These preliminary results indicate that thermal drones can detect mahogany gliders under certain conditions. With further validation, this approach could complement existing techniques for presence–absence surveys, population assessments and behavioural observations.
Keywords: arboreal mammal, fauna survey, Petauridae, population monitoring, presence–absence, remote sensing, RPAS, UAV.
Introduction
Understanding habitat occupancy and use is critical for informing conservation management of threatened species (Lindenmayer et al. 2012; Lindenmayer 2023; Numminen et al. 2023). Arboreal gliding mammals pose a significant challenge in this context due to their nocturnal behaviour, small size and rapid movements, frequently avoiding detection by observers. Spotlighting, camera trapping and cage trapping are widely used to study glider species, providing information on occupancy, abundance and genetic connectivity, but they are often labour-intensive and can alter natural behaviour (Jackson and Johnson 2002; Berris et al. 2022; Lumsden et al. 2022; Smith et al. 2022). Thermal drone surveys offer a complementary approach that can provide valuable preliminary data on the presence and habitat use of arboreal mammals, helping to guide and refine more intensive monitoring programs (Zhang et al. 2023; Wearn et al. 2024).
The mahogany glider (Petaurus gracilis) is endemic to a narrow band of coastal lowland woodland in tropical north Queensland (Qld), Australia (IUCN 2008; Jackson 2023). Within its range, ~50% of suitable habitat has been cleared, leading to the species’ listing as Endangered (Jackson et al. 2011; Jackson and Diggins 2021). Recent distribution modelling indicates patchy occurrence and uncertainty regarding its distribution and population densities (Chang et al. 2023). The species’ population trajectory remains difficult to monitor due to its low population density and cryptic nature (Jackson 2000a). Spotlighting and cage trapping can greatly underestimate gliding marsupial presence. For example, Goldingay and Taylor (2022) estimated that spotlighting and trapping yielded equally low detection probabilities of ~22% for single-night surveys of squirrel gliders (Petaurus norfolcensis). Similarly, Lindenmayer et al. (2001) found a spotlighting detection rate of ~26% for collared greater gliders (Petauroides volans) occurring within a survey area in southern New South Wales (NSW). Arboreal camera traps require extensive deployment effort and typically return low detection rates (Chang et al. 2025b). Cage trapping can provide valuable demographic and genetic data but it is invasive and unsuitable for routine population monitoring (Jackson 2000a; Chang et al. 2025a).
Limitations of conventional survey methods have prompted the need to trial alternative techniques that may improve detection rates while reducing effort and disturbance to animals. Handheld thermal cameras and scopes can improve detection of some arboreal mammals compared to spotlighting, but dense forests limit their effectiveness by occluding heat signatures in the canopy (Vinson et al. 2020; Pocknee et al. 2021; Underwood et al. 2022). Conversely, thermal drones are advantageous for detecting canopy-dwelling species as their heat signatures are less likely to be obscured by vegetation when viewed from above. Thermal drones have already demonstrated strong potential for detecting arboreal mammals in Australia, including koalas (Phascolarctos cinereus) in temperate forests (Beranek et al. 2021), Bennett’s tree kangaroos (Dendrolagus bennettianus) in tropical rainforests (Norris and Larson 2025) and spectacled flying-foxes (Pteropus conspicillatus) in various tropical forest types (Norris 2025a). Although concerns persist that warm tropical environments may preclude adequate thermal contrast, recent studies indicate that high detection rates are possible under favourable conditions, such as in overcast conditions or at night (Norris and Larson 2025; Rahman et al. 2025).
The ecology of the mahogany glider makes it well-suited for thermal drone detection. As a strictly nocturnal species, its peak activity occurs at night when cooler air temperatures and the absence of solar radiation enhance thermal contrast. Mahogany gliders typically forage in moderately open mid- to upper-canopy strata (Jackson 2000b), facilitating a clearer aerial line of sight. Despite its small body size (<500 g), its distinctive gliding behaviour, with glides of up to 60 m between trees, further aids detection in thermal imagery. Misidentification with the sympatric inland sugar glider (Petaurus notatus) is possible but unlikely because it is much smaller (60–115 g; 290–345 mm long) with a shorter, less bushy tail and typically occupies the mid- to lower canopy (Jackson 2000a; Cremona et al. 2023; Goldingay et al. 2024). Given these distinctions, high-resolution thermal imagery may support morphological assessments comparable to spotlighting or camera trapping. Although mahogany gliders occur near squirrel glider populations in the southern Wet Tropics, these two species are not known to be sympatric (Chang et al. 2025a).
Thermal drones have not previously been used to survey mahogany gliders. This study aimed to: (1) assess the feasibility of detecting mahogany gliders by using nocturnal thermal drone surveys, and (2) document flight conditions associated with detections to guide future research.
Methods
Thermal drone surveys were conducted on 30 May 2025 in an isolated woodland fragment at Bambaroo (–18.86704, 146.184063), approximately 84 km north-west of Townsville in tropical north Qld (Fig. 1a). The region has a tropical savanna climate with a distinct wet season (November–April) and drier winter months. The 64-ha study site comprises predominantly open eucalypt woodland on gently sloping alluvial plains, with a ~20–28 m high canopy and an open understory (Fig. 1b; Neldner et al. 2023). The fragment is bounded to the south by a highway and sugarcane railway, and elsewhere by cleared sugarcane farmland (Fig. 2). The site was chosen because recent cage- and camera-trapping studies reported a relatively high density of mahogany gliders in the region (Chang et al. 2023) and no other glider species are known to occur within the fragment (Chang et al. 2023), minimising the risk of false-positive detections.
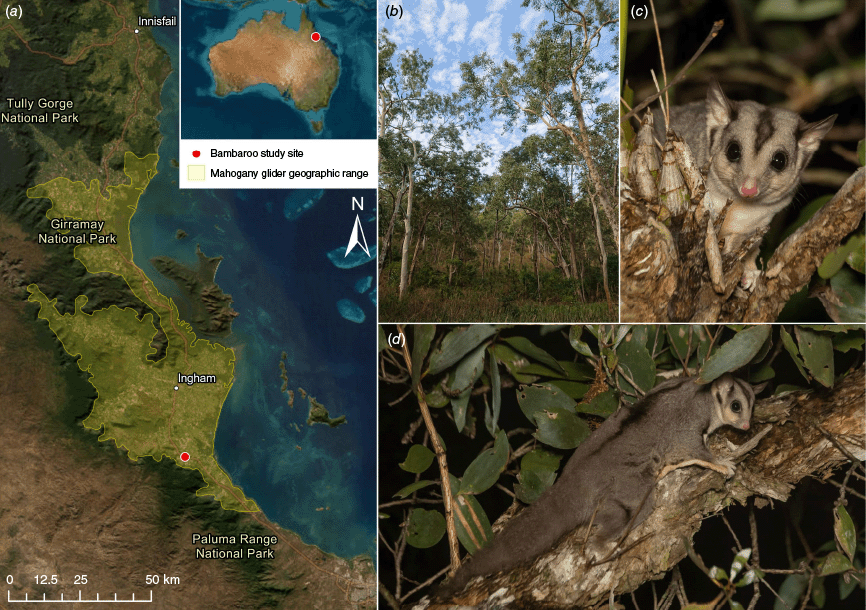
Location and context of the mahogany glider (Petaurus gracilis) thermal drone surveys: (a) map of the southern Wet Tropics region in north Queensland, Australia, showing the geographic range of the mahogany glider (yellow boundary) (IUCN 2008) and the Bambaroo study site (red dot), with the inset showing the site location relative to Australia; (b) open eucalypt woodland habitat surveyed at Bambaroo; (c) close-up of a mahogany glider highlighting the dark facial stripe (not unique to mahogany gliders); and (d) full-body view of a mahogany glider, showing the long, furred tail and dorsal stripe, which is more prominent in mahogany gliders than other glider species. Images by (b) Y. Chang and (c, d) K. Palmer.
Thermal drone flight paths and mahogany glider (Petaurus gracilis) detections at Bambaroo, north Queensland, on 30 May 2025. The four flight routes followed approximately parallel transects over the 64-ha woodland fragment and are shown in red (Flight 1, starting 8:40 pm), orange (Flight 2, starting 9:22 pm), yellow (Flight 3, starting 9:57 pm) and grey (Flight 4, starting 10:30 pm). Points indicate locations where mahogany gliders were detected, with point size corresponding to the number of individuals observed and colour matching the flight in which the detection occurred. The drone launch and landing site is marked with an ‘H’ at the southern boundary of the fragment.
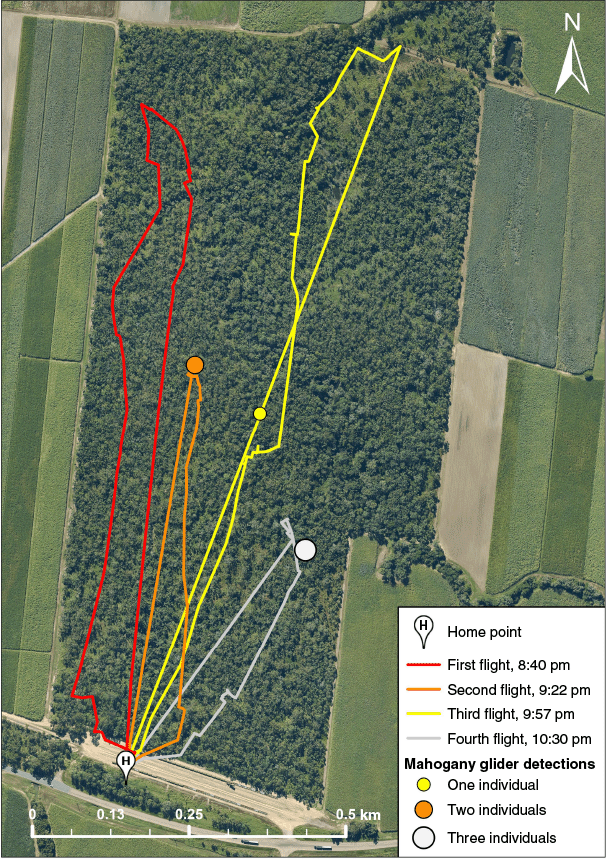
Four consecutive flights were conducted between 8:40 pm and 11:10 pm (Table 1) using a DJI Matrice 350 RTK drone equipped with a DJI Zenmuse H30T payload, which includes a high-resolution (1280 × 1024 pixel) thermal infrared sensor and Night Scene and near-infrared (NIR) Illumination modes for visual identification under low-light conditions (DJI Technology Co., Shenzhen, China; Supplementary Table S1). Weather conditions were recorded at 5-min intervals using a Kestrel 5200 weather meter (Nielsen-Kellerman Co., Boothwyn, PA, USA). Manual flights followed approximately parallel transects (Fig. 2). Flight parameters, including flight height above ground level (AGL), speed and gimbal pitch (i.e. vertical camera angle), were varied heuristically by the pilot (E. B. B. Norris) to gauge which settings might improve mahogany glider detection (Table 1). All flights complied with Civil Aviation Safety Authority (CASA) regulations.
| Flight no. | Start time | Flight duration (min) | Distance flown (km) | Mean air temperature (°C) | Mean relative humidity (%) | P. gracilis detections | |
|---|---|---|---|---|---|---|---|
| 1 | 8:40 pm | 30:44 | 2.4 | 21.0 | 84.3 | 0 | |
| 2 | 9:22 pm | 28:32 | 1.5 | 20.5 | 81.3 | 2 | |
| 3 | 9:57 pm | 30:17 | 2.7 | 20.4 | 84.0 | 1 | |
| 4 | 10:30 pm | 30:50 | 1.2 | 20.4 | 78.5 | 3 |
Thermal video was recorded continuously from take-off to landing. When a hotspot resembling a mammal was detected, the drone was brought to a hover, and the thermal zoom function was used to investigate its shape, size and behaviour. If the characteristics were consistent with a mammal, the drone was flown closer to improve image clarity. Mahogany gliders were identified based on their size, long tail, rapid slinking movements through the canopy and distinctive rectangular shape with patagium outstretched while gliding between trees (Fig. 3). In some cases, Night Scene and NIR Illumination modes were activated to aid visual species identification by providing detailed views of key morphological traits such as pelage patterning and facial markings (Fig. 1c–d; Fig. 4). Coordinates of confirmed sightings were recorded in the DJI Pilot 2 app using the drone’s onboard laser rangefinder.
Thermal video stills showing all six mahogany gliders (Petaurus gracilis) detected during nocturnal drone surveys conducted at Bambaroo, north Queensland, on 30 May 2025: (a, b) first and second individuals detected during the second flight; (c) third individual observed during the third flight; (d) three gliders detected concurrently during the fourth flight; and (e, f) the fifth and sixth gliders captured while gliding during the same flight. Yellow circles in panels (c) and (d) indicate glider locations.

Identification of a mahogany glider (Petaurus gracilis) using dual imaging modes during nocturnal thermal drone surveys: (a) thermal video frame showing the glider as a bright heat signature in the canopy, and (b) concurrent video frame captured using the zoom camera with Night Scene mode and near-infrared (NIR) Illumination activated, revealing the species’ distinctive dorsal stripe while foraging on a branch.

Thermal and Night Scene footage were reviewed post-flight to confirm detections and identify any additional animals that may have been overlooked during surveys. Weather data were downloaded from the Kestrel weather meter, and flight logs with global positioning system (GPS) metadata were retrieved via the AirData platform (https://airdata.com/). Drone flight paths and confirmed sightings were mapped in ArcGIS Pro (Esri, Redlands, CA, USA) (Fig. 2).
Results and discussion
Nocturnal thermal drone surveys yielded six mahogany glider detections over a 2-h flight period (Table 1; Fig. 2). All confirmed detections were made when flying 10–30 m above the canopy, at slower flight speeds (~1–3 m s−1), and using a gimbal pitch angle between approximately −20° and −30° (Table 2). Given this was a single-night feasibility study, these settings are not necessarily optimal for maximising detection. Detection probability in thermal drone surveys is shaped by interactions between flight altitude, speed, gimbal pitch and sensor performance, as well as vegetation structure, weather and animal behaviour (Pinel-Ramos et al. 2024; Reinegger et al. 2025). While similar flight parameters have proven effective for detecting some arboreal mammal species (Rahman et al. 2022; Gazagne et al. 2023; Norris and Larson 2025), other studies found greater success using different settings, such as higher altitudes or nadir gimbal pitch (Burke et al. 2019; Santos et al. 2023; Pinel-Ramos et al. 2024; Seegobin et al. 2024). Formal, replicated surveys comparing detection rates across different times, weather conditions and flight parameters are required to quantify detection errors and evaluate trade-offs across parameter space (Baxter and Hamilton 2018; Brack et al. 2018).
| Flight no. | Detection no. | Detection time | Flight altitude (m) | Flight speed (m s−1) | Gimbal pitch angle (°) | |
|---|---|---|---|---|---|---|
| 2 | 1 | 9:45 pm | 57.2 | 2.2 | –27.4 | |
| 2 | 52.8 | 2.7 | –24.8 | |||
| 3 | 3 | 10:10 pm | 32.2 | 1.2 | –21.3 | |
| 4 | 4 | 10:39 pm | 38.7 | 2.4 | –21.2 | |
| 5 | ||||||
| 6 |
Single parameter values attributed to multiple detections indicate that those individuals were detected concurrently. Altitude is above ground level (AGL). Negative gimbal pitch angle values represent the vertical camera angle directed below the horizontal plane (i.e. 0°), with nadir being at –90°.
All mahogany glider detections were identified in thermal imagery by body size, a long tapering tail and distinctive gliding behaviour (Fig. 3; Supplementary Video S1; Norris 2025b). In one case, Night Scene and NIR Illumination also revealed the dark dorsal stripe (Fig. 4; Supplementary Video S1, Norris 2025b); however, these imaging modes were seldom useful for species confirmation because the gliders’ rapid movements across swaying branches generally prevented the camera from focussing. Where morphologically similar species co-occur, definitive identification may require supplementary verification, such as by using drone-mounted searchlights (Beranek et al. 2021; Ryan et al. 2025). Compared to visual verification by ground-based observers with spotlights, which may cause delays and missed opportunities for species confirmation, dual-gimbal systems that carry a thermal camera payload and a searchlight allow near-simultaneous detection and visual confirmation from the drone (Ryan et al. 2025). Future research would benefit from comparing these visual identification methods for mahogany gliders and other petaurid species to assess how they impact survey efficiency, identification accuracy and the disturbance responses of target species.
No mahogany gliders were detected during the first flight. Several factors could explain this lack of detections. The earlier time of night may have prevented adequate thermal contrast because ground surfaces, tree trunks and branches retained more heat compared to later surveys (Supplementary Video S1; Norris 2025b). Additionally, the higher altitudes (60–80 m AGL) at which this flight was conducted may have decreased glider detectability by increasing the ground sampling distance (GSD) and reducing the number of pixels representing animals’ thermal signatures (Burke et al. 2019). A large macropod, likely an agile wallaby (Notamacropus agilis), was detected near the forest edge during this survey, suggesting that the smaller size and potentially less exposed position of mahogany gliders’ thermal signatures may have limited detectability. Alternatively, gliders may have been absent from the eastern portion of the site during the first flight, reinforcing the need to resolve detection probabilities for mahogany gliders before thermal drones can be adopted as a systematic survey tool.
False positive errors caused by double counting remain possible for detections separated in time and space across flights due to the highly mobile nature of mahogany gliders. Whereas individuals observed concurrently within the second and fourth flights can be treated as unique, the glider detected during the third flight may have been one of the two seen earlier, given the short distance (125 m) and time interval (25 min) between detections (Table 1; Fig. 2). Mahogany gliders are capable of gliding up to 60 m and making several glides in quick succession (Jackson 2000c), rendering movement between detection points within this timeframe plausible. Although less likely, double counting cannot be excluded for the fourth flight, where the nearest prior detection was 230 m away (Fig. 2). Individuals may also have moved out of surveyed areas between flights, increasing false-negative errors. Such false-positive and -negative errors are a known limitation of thermal drone surveys using parallel transects, particularly for highly mobile species that can easily move between flight lines within the survey period (Brack et al. 2018; Schultz et al. 2024). Addressing these biases will require further research. One potential approach is to incorporate animal movement data, such as GPS-tracking, in statistical models to account for the likelihood of individuals traversing flight lines during survey windows (Schultz et al. 2024).
These preliminary results suggest that thermal drones are a feasible tool for detecting endangered mahogany gliders in tropical lowland woodlands, offering a minimally invasive way to collect preliminary data on species presence and habitat use. In this study, six individuals were detected in just 2 h of survey effort, highlighting the potential of this approach to provide rapid insights into glider activity in the surveyed area. While established methods such as cage trapping, camera trapping and spotlighting remain essential for estimating occupancy, abundance and genetic connectivity, they are often labour-intensive, limited in the area they can cover, and yield low detection rates (Jackson 2000a; Chang et al. 2025a). Thermal drones therefore provide a useful complement to these approaches by rapidly identifying areas of activity and guiding the design of more targeted, intensive monitoring programs. Further studies are needed to evaluate the broader applicability of thermal drones across habitats and seasons to determine how best they can be integrated within existing survey frameworks.
Importantly, thermal drones did not elicit visible disturbance responses from mahogany gliders in this study. All individuals exhibited natural behaviours such as foraging, gliding and grooming while the drone hovered less than 20 m above (Supplementary Video S1; Norris 2025b). This suggests that thermal drones may be suitable for behavioural studies, offering a distinct advantage over camera traps or spotlighting, which often fail to capture fine-scale behaviours in fast-moving or canopy-dwelling species. Additionally, the ability to observe gliders behaving naturally in situ presents opportunities to identify key denning and foraging sites, which can be geotagged using the drone’s onboard rangefinder for subsequent ground-truthing.
Findings from this single-night feasibility study suggest that thermal drones are most immediately valuable for rapid presence-absence surveys of mahogany gliders at sites flagged as potential habitat (Chang et al. 2023). Efficient confirmation of occupancy could facilitate area prioritisation for detailed assessments of population density (Chang et al. 2025b) and cage-trapping to obtain genetic samples needed to resolve taxonomy and guide conservation management (Chang et al. 2025a). With further methodological refinement and explicit estimation of detection probability, it may also be possible to use spatially and temporally replicated drone-derived count data in conjunction with N-mixture models for population estimation across the species’ range (see Trinh-Dinh et al. 2024; Ryan et al. 2025). In this complementary role, thermal drones have the potential to expand survey coverage across areas that are difficult to access, and open new avenues for behavioural observations to strengthen the evidence base for conservation of this endangered species.
Data availability
Thermal video footage from all four flights is archived and available via figshare (Norris 2025b). Associated detection records from the drone surveys are available from the corresponding author upon reasonable request.
Conflicts of interest
The authors declare that there are no conflicts of interest regarding the publication of this manuscript.
Declaration of funding
This research was funded by the National Environmental Science Program (NESP) Resilient Landscapes Hub. The funding body had no role in the study design, data collection, analysis, or interpretation, nor in the decision to publish this manuscript.
Acknowledgements
The authors are grateful to the NESP Resilient Landscapes Hub for providing funding that supported the purchase of the drone and thermal camera used in this study. We thank the reviewers for their constructive comments, which improved the readability of this manuscript.
References
Baxter, P. W. J., and Hamilton, G. (2018). Learning to fly: integrating spatial ecology with unmanned aerial vehicle surveys. Ecosphere 9(4), e02194.
| Crossref | Google Scholar |
Beranek, C. T., Roff, A., Denholm, B., Howell, L. G., and Witt, R. R. (2021). Trialling a real-time drone detection and validation protocol for the koala (Phascolarctos cinereus). Australian Mammalogy 43(2), 260-264.
| Crossref | Google Scholar |
Berris, K. K., Ruykys, L., Goldingay, R. L., Morrant, D. S., Packer, J. G., Descovich, K., Matthews, A., Coetsee, A., Nicolson, V., Horsup, A., Moseby, K., Jose, H. L., Gill, S. J., and Taggart, D. A. (2022). Wombats, bandicoots, bilbies, possums and gliders. In ‘Wildlife Research in Australia: Practical and Applied Methods’. (Eds B. P. Smith, H. P. Waudby, C. Alberthsen, J. O. Hampton.) pp. 401–411. (CSIRO Publishing: Melbourne.)
Brack, I. V., Kindel, A., and Oliveira, L. F. B. (2018). Detection errors in wildlife abundance estimates from Unmanned Aerial Systems (UAS) surveys: Synthesis, solutions, and challenges. Methods in Ecology and Evolution 9, 1864-1873.
| Crossref | Google Scholar |
Burke, C., Rashman, M. F., Longmore, S. N., McAree, O., Glover-Kapfer, P., Ancrenaz, M., and Wich, S. A. (2019). Successful observation of orangutans in the wild with thermal-equipped drones. Journal of Unmanned Vehicle Systems 7(3), 235-257.
| Crossref | Google Scholar |
Chang, Y., Bertola, L. V., and Hoskin, C. J. (2023). Species distribution modelling of the endangered Mahogany Glider (Petaurus gracilis) reveals key areas for targeted survey and conservation. Austral Ecology 48, 289-312.
| Crossref | Google Scholar |
Chang, Y., Bertola, L. V., Zenger, K. R., and Hoskin, C. J. (2025a). Conservation genetics of mahogany gliders and their complex evolutionary relationship with squirrel gliders. Conservation Genetics 26, 731-750.
| Crossref | Google Scholar |
Chang, Y., Lin, C., and Hoskin, C. J. (2025b). Estimating the abundance of an endangered arboreal marsupial using camera traps and an integrated species distribution model. Ecology and Evolution 15, e72037.
| Crossref | Google Scholar |
Gazagne, E., Gray, R. J., Ratajszczak, R., Brotcorne, F., and Hambuckers, A. (2023). Unmanned aerial vehicles (UAVs) with thermal infrared (TIR) sensors are effective for monitoring and counting threatened Vietnamese primates. Primates 64(4), 407-413.
| Crossref | Google Scholar | PubMed |
Goldingay, R. L., and Taylor, B. D. (2022). Comparison of spotlighting and trapping in population surveys of the squirrel glider. Australian Mammalogy 44(2), 208-212.
| Crossref | Google Scholar |
Goldingay, R. L., Jackson, S. M., Winter, J. W., Harley, D. K. P., Bilney, R. J., Quin, D. G., Smith, G. C., Taylor, B. D., and Kavanagh, R. P. (2024). What’s in a name? Selection of common names among new and revised species of Australian mammals, and the case of the sugar glider. Australian Mammalogy 46, AM23017.
| Crossref | Google Scholar |
Jackson, S. M. (2000a). Population dynamics and life history of the mahogany glider, Petaurus gracilis, and the sugar glider, Petaurus breviceps, in North Queensland. Wildlife Research 27, 21-37.
| Crossref | Google Scholar |
Jackson, S. M. (2000b). Habitat relationships of the mahogany glider, Petaurus gracilis, and the sugar glider, Petaurus breviceps. Wildlife Research 27, 39-48.
| Crossref | Google Scholar |
Jackson, S. M. (2000c). Glide angle in the genus Petaurus and a review of gliding in mammals. Mammal Review 30, 9-30.
| Crossref | Google Scholar |
Jackson, S. M., and Johnson, C. N. (2002). Time allocation to foraging in the mahogany glider Petaurus gracilis (Marsupialia, Petauridae) and a comparison of activity times in exudivorous and folivorous possums and gliders. Journal of Zoology (London) 256, 271-277.
| Crossref | Google Scholar |
Jackson, S. M., Morgan, G., Kemp, J. E., Maughan, M., and Stafford, C. M. (2011). An accurate assessment of habitat loss and current threats of the mahogany glider (Petaurus gracilis). Australian Mammalogy 33, 82-92.
| Crossref | Google Scholar |
Lindenmayer, D. B. (2023). Forest biodiversity declines and extinctions linked with forest degradation: A case study from Australian tall, wet forests. Land 12(3), 528.
| Crossref | Google Scholar |
Lindenmayer, D. B., Cunningham, R. B., Donnelly, C. F., Incoll, R. D., Pope, M. L., Tribolet, C. R., Viggers, K. L., and Welsh, A. H. (2001). How effective is spotlighting for detecting the great glider (Petauroides volans)?. Wildlife Research 28, 105-109.
| Crossref | Google Scholar |
Lindenmayer, D. B., Likens, G. E., Anderson, A., Bowman, D., Bull, C. M., Burns, E., Dickman, C. R., Hoffmann, A. A., Keith, D. A., Liddell, M. J., Lowe, A. J., Metcalfe, D. J., Phinn, S. R., Russell-Smith, J., Thurgate, N., and Wardle, G. M. (2012). Value of long-term ecological studies. Austral Ecology 37(7), 745-757.
| Crossref | Google Scholar |
Lumsden, L. F., Waudby, H. P., Meek, P. D., Crisp, H. A., Cripps, J. K., Cristescu, R., Jamieson, L. T., Nelson, J. L., Teixeira, D., Hodgens, N., Rutter, N., Morrant, D. S., Bower, D. S., Dickman, C. R., and Soennichsen, K. (2022). Detecting and monitoring wildlife remotely. In ‘Wildlife Research in Australia: Practical and Applied Methods’. (Eds B. P. Smith, H. P. Waudby, C. Alberthsen, J. O. Hampton.) pp. 75–95. (CSIRO Publishing: Melbourne.)
Norris, E. B. B. (2025a). Drone surveys cause less disturbance than ground-based surveys in endangered spectacled flying-foxes (Pteropus conspicillatus). Australian Mammalogy 47, AM25010.
| Crossref | Google Scholar |
Norris, E. (2025b). Thermal drone video footage of mahogany gliders (Petaurus gracilis) recorded at Bambaroo, north Queensland, Australia on 30 May 2025. figshare. Media. doi:10.6084/m9.figshare.29484896
Norris, E. B. B., and Larson, J. (2025). Thermal drones are highly effective for detecting elusive Bennett’s tree kangaroos (Dendrolagus bennettianus) in Australia’s tropical rainforests. Australian Mammalogy 47, AM24053.
| Crossref | Google Scholar |
Numminen, E., Jokinen, M., Linden, A., and Vanhatalo, J. (2023). Species ecology can bias population estimates. Biological Conservation 283, 110115.
| Crossref | Google Scholar |
Pinel-Ramos, E. J., Aureli, F., Wich, S., Longmore, S., and Spaan, D. (2024). Evaluating thermal infrared drone flight parameters on spider monkey detection in tropical forests. Sensors 24(17), 5659.
| Crossref | Google Scholar | PubMed |
Pocknee, C. A., Lahoz-Monfort, J. J., Martin, R. W., and Wintle, B. A. (2021). Cost-effectiveness of thermal imaging for monitoring a cryptic arboreal mammal. Wildlife Research 48, 625-634.
| Crossref | Google Scholar |
Rahman, D. A., Sitorus, A. B. Y., and Condro, A. A. (2022). From coastal to montane forest ecosystems, using drones for multi-species research in the tropics. Drones 6(1), 6.
| Crossref | Google Scholar |
Rahman, D. A., Putro, H. R., Mufawwaz, T. A., Rinaldi, D., Yudiarti, Y., Prabowo, E. D., Arief, H., Sihite, J., and Priantara, F. R. N. (2025). Developing a new method using thermal drones for population surveys of the world’s rarest great ape species, Pongo tapanuliensis. Global Ecology and Conservation 58, e03463.
| Crossref | Google Scholar |
Reinegger, R. D., Bissessur, P., Meereechpersad, I., Bhanda, G., Gazagne, E., and Jones, G. (2025). Improving primate detection using thermal infrared imaging: availability and observer errors in drone surveys of mixed tropical forests. International Journal of Remote Sensing 46(11), 4345-4373.
| Crossref | Google Scholar |
Ryan, S. A., Southwell, D. M., Beranek, C. T., Clulow, J., Jordan, N. R., and Witt, R. R. (2025). Estimating the landscape-scale abundance of an arboreal folivore using thermal imaging drones and binomial N-mixture modelling. Biological Conservation 309, 111207.
| Crossref | Google Scholar |
Santos, P. M., Kaizer, M. D., Moreira, D. O., Melo, F. R. D., and Mendes, S. L. (2023). Every flight is a surprise: First records of the southern maned three-toed sloth (Bradypus crinitus: Bradypodidae) through drones. Mammalia 87(3), 223-227.
| Crossref | Google Scholar |
Schultz, E. A., Ellison-Neary, N., Boudreau, M. R., Street, G. M., Jones, L. R., Evans, K. O., and Iglay, R. B. (2024). On the move: Influence of animal movements on count error during drone surveys. Ecology and Evolution 14(10), e70287.
| Crossref | Google Scholar | PubMed |
Seegobin, V. O., Oleksy, R. Z., and Florens, F. B. V. (2024). Foraging habitat quality of an Endangered mass-culled flying fox is reduced by alien plant invasion and improved by alien plant control. Journal for Nature Conservation 78, 126569.
| Crossref | Google Scholar |
Smith, B. P., Waudby, H. P., Dickman, C. R., Soennichsen, K., Mills, C., Howe, A., Woolley, L. A., Griffiths, S. R., Cox, T. E., Clayton, J., Penton, C., Morrant, D. S., Letnic, M., Cullen, D., Stobo-Wilson, A., Taggart, D. A., Gleeson, D., Cripps, J. K., Gill, M. J., and MacDonald, A. J. (2022). Observing wildlife and its signs. In ‘Wildlife Research in Australia: Practical and Applied Methods’. (Eds B. P. Smith, H. P. Waudby, C. Alberthsen, J. O. Hampton.) pp. 42–74. (CSIRO Publishing: Melbourne.)
Trinh-Dinh, H., Wearn, O. R., Ngoprasert, D., Wich, S., and Savini, T. (2024). A drone-based population survey of Delacour’s langur (Trachypithecus delacouri) in the karst forests of northern Vietnam. Biological Conservation 300, 110841.
| Crossref | Google Scholar |
Underwood, A. H., Derhe, M. A., and Jacups, S. (2022). Thermal imaging outshines spotlighting for detecting cryptic, nocturnal mammals in tropical rainforests. Wildlife Research 49, 491-499.
| Crossref | Google Scholar |
Vinson, S. G., Johnson, A. P., and Mikac, K. M. (2020). Thermal cameras as a survey method for Australian arboreal mammals: a focus on greater gliders. Australian Mammalogy 42, 367-374.
| Crossref | Google Scholar |
Wearn, O. R., Trinh-Dinh, H., Ma, C. Y., Khac Le, Q., Nguyen, P., Van Hoang, T., Van Luong, C., Van Hua, T., Van Hoang, Q., Fan, P. F., and Duc Nguyen, T. (2024). Vocal fingerprinting reveals a substantially smaller global population of the Critically Endangered cao vit gibbon (Nomascus nasutus) than previously thought. Scientific Reports 14(1), 416.
| Crossref | Google Scholar | PubMed |
Zhang, H., Turvey, S. T., Pandey, S. P., Song, X., Sun, Z., and Wang, N. (2023). Commercial drones can provide accurate and effective monitoring of the world’s rarest primate. Remote Sensing in Ecology and Conservation 9(6), 775-786.
| Crossref | Google Scholar |