

Awn length variation in Australia’s most widespread grass, Themeda triandra, across its distribution
Annette M. Cavanagh A * , Robert C. Godfree B and John W. Morgan A
A * , Robert C. Godfree B and John W. Morgan A
A
B
Abstract
Themeda triandra (kangaroo grass) is the most widespread plant species in Australia. Using herbarium specimens, we quantified awn-length variation across its distribution in relation to climate. We found that awn lengths were shortest around the eastern and southern coast of Australia (with short awns also concentrated in the Pilbara), increasing in length progressively inland and towards the north. There was a significant positive relationship between awn length and mean annual temperature. Further research is needed to understand why awn lengths vary, and how awn-length variation affects seed and seedling fitness in natural habitats.
Keywords: biogeography, grasses, grasslands, plant adaptation, plant ecology, Poaceae, polyploidy, seed ecology.
Introduction
Seed dispersal is an important step in the life cycle of plants. The persistence of a non-clonal plant relies on the ability of its seeds to move away from the parent plant and successfully bury at sites that have optimal conditions for germination, establishment, growth and reproduction. Awned diaspores have been shown to influence autochorous dispersal and burial (Murbach 1900; Peart 1979; Peart and Clifford 1987; Adams and Tainton 1990; Sindel et al. 1993; Garnier and Dajoz 2001; Cavanagh et al. 2020). Awns are frequent in Australian native grasses and vary in their morphology across species (Cavanagh et al. 2019). Given that awns are a large resource investment for many grass species, and they are likely to have important plant fitness benefits (Adams and Tainton 1990), it is surprising that more consideration has not been given to this variation.
The length of awns can vary both among and within plant populations (Garnier and Dajoz 2001). The broad distribution of some grass species in Australia may, in part, be due to their ability to move seeds across the soil surface and facilitate their burial into soil away from potential threats where germination can take place (Peart 1979; Stamp 1989; Cavanagh et al. 2020). This may be particularly pertinent to Australia’s most widespread plant species, Themeda triandra Forssk. (Poaceae; kangaroo grass) (Gallagher 2016), a C4 grass species thought to have arrived in Australia only 1.3 million years ago (Dunning et al. 2017) but now growing in all parts of the continent except permanently wet, very dry, very cold, saline or heavily shaded sites. Across its range, T. triandra exhibits substantial ecotypic variation (Dunning et al. 2022).
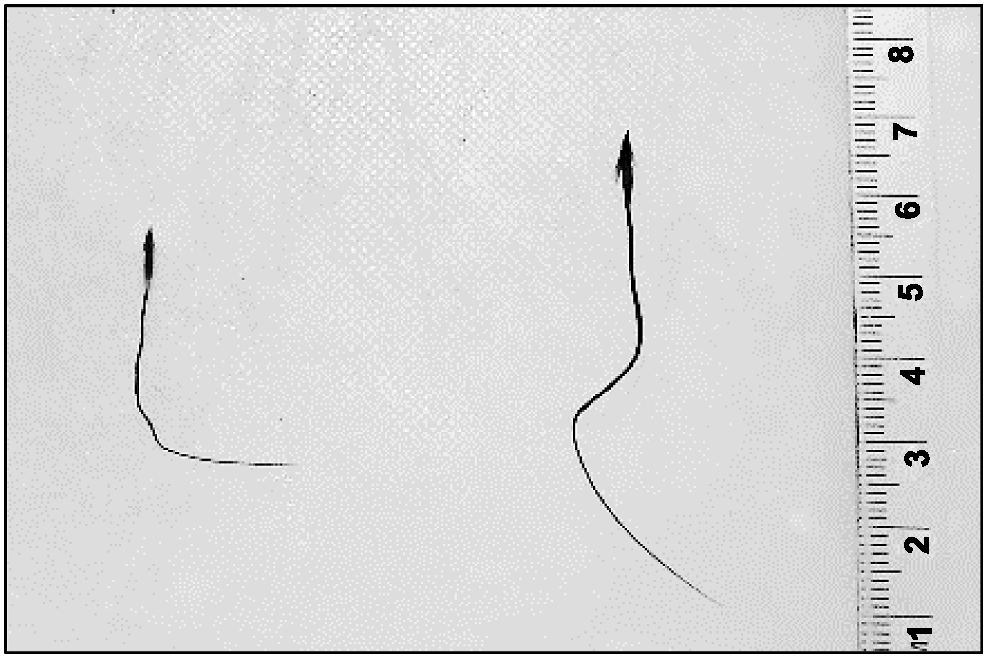
T. triandra has a bigeniculate, hygroscopic awn (Fig. 1), meaning that the awn twists and untwists in response to changes in humidity (Sindel et al. 1993). This mechanical movement allows the awn to act as a lever, facilitating forward movement on a surface and then bracing of the diaspore as it twists itself into the soil (Murbach 1900; Stamp 1989; Johnson and Baruch 2014). Surface movements of T. triandra awned diaspores are among the highest of Australian native grasses (Cavanagh et al. 2020), but it is unclear whether, and how, awn length varies in this species. The objective of this study was to quantify how awn length of T. triandra varies across the broad range of climates that the species occupies.
Materials and methods
Herbarium specimens from the Australian National Herbarium (Canberra, ACT) were used to determine awn lengths for 206 Themeda triandra specimens that had been collected from across the distribution of the species in Australia. Four diaspores were removed from each specimen; the length of the awn was measured (millimetres) with digital callipers, with the awn stretched as straight as possible to facilitate measurement. The mean awn lengths were then calculated for each specimen. Seed length was measured in the same way. Climate data for each site were obtained from Worldclim ver. 2 (Fick and Hijmans 2017) and linear regressions were performed on awn length and both mean annual precipitation (millimetres) and mean annual temperature (°C).
Results
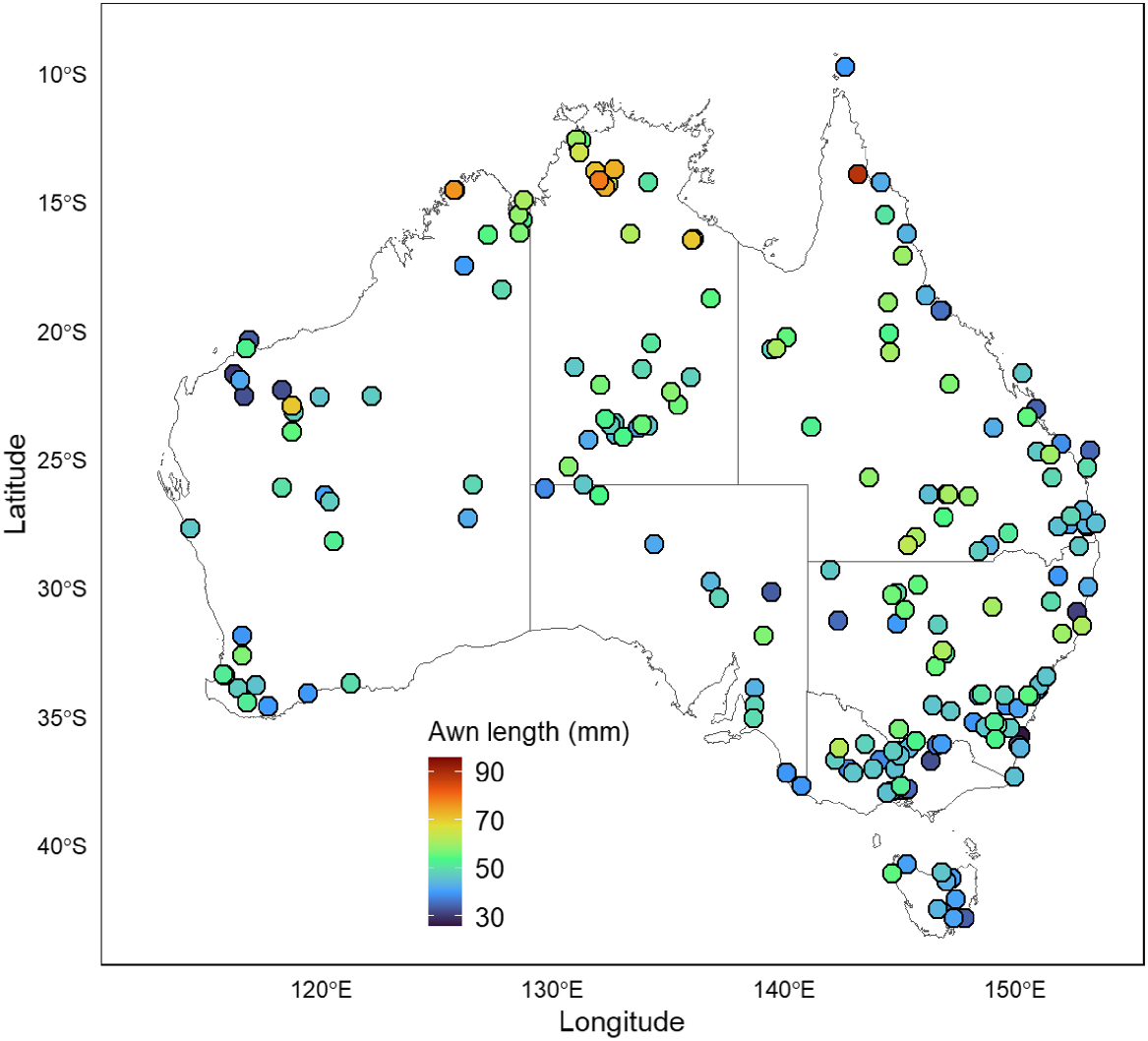
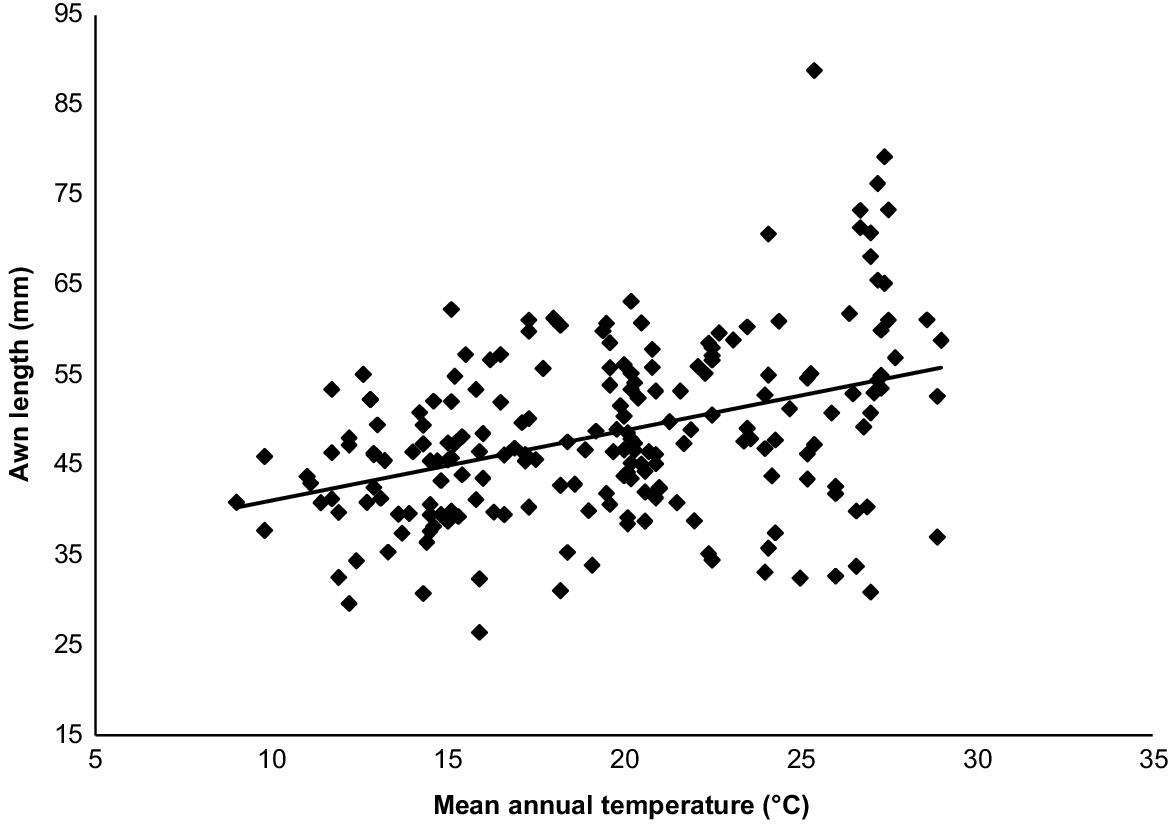
Awn lengths ranged from 26.5 mm to 88.8 mm. Awn lengths were generally shortest around the eastern and southern coast of Australia, with awn length increasing progressively inland (particularly west of the Great Dividing Range) and towards the north (Fig. 2). Exceptions to this general observation can be seen in the Pilbara region of northern Western Australia where awns are short. The longest awns were found in specimens collected in the tropical savanna biome of northern Australia. There was no relationship between awn length and mean annual precipitation (r = 0.05, P = 0.50), but there was a significant positive relationship between awn length and mean annual temperature (awn length (mm) = 0.7753 (mean annual temperature) + 33.345, r = 0.39, P < 0.0001; Fig. 3). Seed length was positively associated with awn length (seed length (mm) = 0.0516 (awn length (mm)) + 6.0885, r = 0.57, P < 0.0001).
Discussion
Themeda triandra had longer awn lengths with an increasing mean annual temperature across its distribution. This implies that the way that diaspores move and bury will vary, given that species with longer bigeniculate awns move further and bury deeper than do species with other awn types (Cavanagh et al. 2020). There may be three, non-mutually exclusive explanations at play for the observation of longer awns in hotter environments.
First, ploidy of T. triandra varies across Australia. At least five ploidy variants of T. triandra have been documented throughout Australia, with populations of tetraploids found mainly in xeric inland areas and diploids around the cooler and wetter eastern and southern coasts (Hayman 1960; Godfree et al. 2017; Dunning et al. 2022). It is thought that Australia was colonised twice by T. triandra, with the independent colonisations representing two different ecotypes adapted to different environments (Dunning et al. 2022). Tetraploid grass species typically have longer awns than do diploids (Godfree et al. 2017) and, so, this is likely to be the most parsimonious explanation for the observed awn-length variation in this study. Indeed, Ahrens et al. (2020) found that genomic variation (including ploidy) in T. triandra was strongly correlated with climate. Further detailed investigation of ploidy type, distribution and awn length is therefore warranted. In other species, polyploids have bigger seeds, a faster rate of germination and larger and taller seedlings than do diploids (Chan et al. 2022), suggesting that morphological variation in diaspore traits is likely to have functional implications for T. triandra.
Second, fire intensity has been found to influence the awn length of diaspores within a species (Garnier and Dajoz 2001). In northern Australia, tropical savannas burn very frequently (Russell-Smith et al. 2020) and, hence, there may be an adaptive advantage to burying deeply into soils where vegetation is burnt frequently, thereby escaping direct fire effects.
Third, awn length was longer at higher mean annual temperature. Soil baking over several days can kill seeds (Egley 1990) and, in northern Australian climates, consistently high temperatures, combined with a dry season and high temperatures post-fire in burnt soils, may place seeds at risk of soil-heating mortality. Diaspores with longer awns have significantly greater burial depths than medium- or short-awned diaspores (Cavanagh et al. 2020) and, as such, deeper burial may reduce risks of high-temperature exposure in surface soils. The shorter awns observed in T. triandra from the Pilbara (of what are thought to be polyploid populations; Dunning et al. 2022) may be because these habitats are quite different from those elsewhere in north-western Australia. Here, populations are found on cracking claypans that are frequently inundated by water (Dunning et al. 2022), perhaps reducing the magnitude or effect of soil baking. Further to this, Cavanagh et al. (2020) found that some grass species with particularly long awns did not bury as deeply as expected and were species that are typically found on floodplains or in soils with high organic matter, meaning that the necessity of assisted burial in these soil types may be lower. Further study on the performance of awns in different soil types should be considered.
Conclusions
To conclude, we have shown that there is substantial variation in awn length across the distribution of Themeda triandra. Such variation is likely to play a role in ‘safe-site’ selection across the species’ range and may be an outcome of differences in ploidy across the range. Quantifying the relationship between ploidy and awn length is an obvious starting point to better understand the reasons for this variation. Regardless, further research is needed to understand how observable variation in awn length of T. triandra with climate, and more broadly among native grasses, affects fitness in natural habitats. Avoidance of hazards (such as fire, drought) is one potential explanation worthy of further investigation. This may contribute to a better understanding of how (or why) T. triandra has come to occupy much of the continent in such a short time.
Data availability
The data that support this study will be shared upon reasonable request to a.m.cavanagh@outlook.com.au.
Conflicts of interest
J. W. M. is Co-Editor in Chief of Australian Journal of Botany. To mitigate this potential conflict of interest they were blinded from the review process. The authors declare no other conflicts of interest.
Declaration of funding
Friends of Grasslands, ACT, provided financial assistance (Grant reference: FSP2017-03).
Author contributions
A. M. C. and R. C. G. conceptualised the study; A. M. C. collected the data; all authors participated in analysis and interpretation of the results; J. W. M. wrote the first draft that all authors edited.
Acknowledgements
The Australian National Herbarium and National Research Collections Australia, CSIRO, Canberra, provided access to plant specimens for study. Alex Godfree assisted with data collection. Luke Florence assisted with data presentation. Sue Bryceson provided valuable feedback on the manuscript.
References
Adams KM, Tainton NM (1990) The function of the hygroscopic awn of Themeda triandra. Journal of the Grassland Society of Southern Africa 7, 271-273.
| Crossref | Google Scholar |
Ahrens CW, James EA, Miller AD, Scott F, Aitken NC, Jones AW, Lu-Irving P, Borevitz JO, Cantrill DJ, Rymer PD (2020) Spatial, climate and ploidy factors drive genomic diversity and resilience in the widespread grass Themeda triandra. Molecular Ecology 29, 3872-3888.
| Crossref | Google Scholar | PubMed |
Cavanagh AM, Godfree RC, Morgan JW (2019) An awn typology for Australian native grasses (Poaceae). Australian Journal of Botany 67, 309-334.
| Crossref | Google Scholar |
Cavanagh AM, Morgan JW, Godfree RC (2020) Awn morphology influences dispersal, microsite selection and burial of Australian native grass diaspores. Frontiers in Ecology and Evolution 8, 581967.
| Crossref | Google Scholar |
Chan JCS, Ooi MKJ, Guja LK (2022) Polyploidy but not range size is associated with seed and seedling traits that affect performance of Pomaderris species. Frontiers in Plant Science 12, 779651.
| Crossref | Google Scholar |
Dunning LT, Liabot A-L, Olofsson JK, Smith EK, Vorontsova MS, Besnard G, Simpson KJ, Lundgren MR, Addicott E, Gallagher RV, Chu Y, Pennington RT, Christin P-A, Lehmann CER (2017) The recent and rapid spread of Themeda triandra. Botany Letters 164, 327-337.
| Crossref | Google Scholar |
Dunning LT, Olofsson JK, Papadopulos AST, Hibdige SGS, Hidalgo O, Leitch IJ, Baleeiro PC, Ntshangase S, Barker N, Jobson RW (2022) Hybridisation and chloroplast capture between distinct Themeda triandra lineages in Australia. Molecular Ecology 31, 5846-5860.
| Crossref | Google Scholar | PubMed |
Egley GH (1990) High-temperature effects on germination and survival of weed seeds in soil. Weed Science 38, 429-435.
| Crossref | Google Scholar |
Fick SE, Hijmans RJ (2017) Worldclim 2: new 1-km spatial resolution climate surfaces for global land areas. International Journal of Climatology 37, 4302-4315.
| Crossref | Google Scholar |
Gallagher RV (2016) Correlates of range size variation in the Australian seed-plant flora. Journal of Biogeography 43, 1287-1298.
| Crossref | Google Scholar |
Garnier LKM, Dajoz I (2001) Evolutionary significance of awn length variation in a clonal grass of fire-prone savannas. Ecology 82, 1720-1733.
| Crossref | Google Scholar |
Godfree RC, Marshall DJ, Young AG, Miller CH, Mathews S (2017) Empirical evidence of fixed and homeostatic patterns of polyploid advantage in a keystone grass exposed to drought and heat stress. Royal Society Open Science 4, 170934.
| Crossref | Google Scholar | PubMed |
Hayman DL (1960) The distribution and cytology of the chromosome races of Themeda australis in southern Australia. Australian Journal of Botany 8, 58-68.
| Crossref | Google Scholar |
Johnson EE, Baruch Z (2014) Awn length variation and its effect on dispersal unit burial of Trachypogon spicatus (Poaceae). Revista de Biología Tropical 62, 321-326.
| Crossref | Google Scholar | PubMed |
Murbach L (1900) Note on the mechanics of the seed-burying awns of Stipa avenacea. Botanical Gazette 30, 113-117.
| Crossref | Google Scholar |
Peart MH (1979) Experiments on the biological significance of the morphology of seed-dispersal units in grasses. Journal of Ecology 67, 843-863.
| Crossref | Google Scholar |
Peart MH, Clifford HT (1987) The influence of diaspore morphology and soil-surface properties on the distribution of grasses. Journal of Ecology 75, 569-576.
| Crossref | Google Scholar |
Russell-Smith J, Edwards AC, Sangha KK, Yates CP, Gardener MR (2020) Challenges for prescribed fire management in Australia’s fire-prone rangelands – the example of the Northern Territory. International Journal of Wildland Fire 29, 339-353.
| Crossref | Google Scholar |
Sindel BM, Davidson SJ, Kilby MJ, Groves RH (1993) Germination and establishment of Themeda triandra (kangaroo grass) as affected by soil and seed characteristics. Australian Journal of Botany 41, 105-117.
| Crossref | Google Scholar |
Stamp NE (1989) Efficacy of explosive vs. hygroscopic seed dispersal by an annual grassland species. American Journal of Botany 76, 555-561.
| Crossref | Google Scholar |