

Productivity of diverse forage brassica genotypes exceeds that of oats across multiple environments within Australia’s mixed farming zone
Lucinda J. Watt A , Lindsay W. Bell A , Brett D. Cocks A , Anthony D. Swan B , Rebecca S. Stutz B , Andrew Toovey C and Joanne De Faveri D
A , Lindsay W. Bell A , Brett D. Cocks A , Anthony D. Swan B , Rebecca S. Stutz B , Andrew Toovey C and Joanne De Faveri D
A CSIRO Agriculture and Food, PO Box 102, Toowoomba, Qld 4350, Australia.
B CSIRO Agriculture and Food, PO Box 1700, Canberra, ACT 2601, Australia.
C CSIRO Agriculture and Food, Private Bag 5, Wembley, WA 6913, Australia.
D CSIRO Data 61, PO Box 780, Atherton, Qld 4883, Australia.
E Corresponding author. Email: lucy.watt@csiro.au
Crop and Pasture Science 72(5) 393-406 https://doi.org/10.1071/CP21034
Submitted: 22 January 2021 Accepted: 6 April 2021 Published: 1 June 2021
Journal Compilation © CSIRO 2021 Open Access CC BY
Abstract
In many areas of Australia’s mixed farming zone, cropping rotations are dominated by cereals and some areas have few suitable broadleaf alternatives. Forage brassicas are widely used in high rainfall livestock systems, but this study shows that several genotypes offer an alternative to forage oats in drier environments within Australia’s mixed farming zone. We compared a diverse set of forage brassica genotypes sown in autumn and winter with benchmark species, principally oats, across 10 experimental site-years. In both early (800–1300 growing degree days after sowing) and late (1600–2100 growing degree days after sowing) grazing windows, several forage brassica genotypes had forage production similar or superior to oats and dual-purpose canola. Many forage brassica genotypes produced higher yields of metabolisable energy (ME) and crude protein (CP), particularly in the late grazing window. In the early grazing window, Rival and Green globe turnips and HT-R24 forage rape consistently produced ~15% above the site mean for all productivity measures, whereas kale produced ~40% less than the site mean. In the late grazing window, oats produced the greatest amount of edible biomass (~44% higher than the site mean); however, Goliath and HT-R24 forage rapes, Pallaton raphanobrassica and dual-purpose canola had the highest yields of ME and CP (~16% higher than the site mean). Green globe turnip, Hunter leafy turnip and Regal kale produced ~22% less than the site mean in this late grazing window. Multi-environment analyses revealed no genotype × environment interactions within the early grazing window. In the late grazing window, there were several genotypic adaptations, particularly for Pallaton, which performed better in low–medium production environments than the other genotypes. We show that forage brassicas offer superior yields of ME and CP, indicating that they may be better able to meet the energy and protein demands of grazing livestock than forage oats.
Keywords: arid environments, crude protein, feedbase, metabolisable energy, multi-environment analysis.
Introduction
Forage brassicas (members of the Brassicaceae family) are beneficial to livestock systems because they have higher digestibility and higher metabolisable energy (ME) than grass-based forages grown in the same season, and have comparable crude protein (CP) contents (Lindsay et al. 2007; Sun et al. 2012; Barry 2013). Generally, they have high vernalisation requirements to maintain vegetative growth (Paridaen and Kirkegaard 2015), and thus nutritive value, for long periods. Because of these nutritive characteristics, forage brassicas are widely used as summer–autumn feed in Australian and New Zealand dairy farming (Moate et al. 1996) and sheep and beef systems (Paridaen and Kirkegaard 2015) in higher rainfall regions. They often replace or complement dry pasture and/or crop residues or lucerne pastures (Nie et al. 2020; Raeside et al. 2020) that are otherwise utilised during this period. In New Zealand, forage brassicas represent 70% of all cultivated crops in dairy and meat-producing livestock grazing systems (Lindsay et al. 2007; Dumbleton et al. 2012).
A broad range of forage brassicas has been developed for livestock systems, including several new commercially available genotypes with improved attributes compared with older varieties. Forage brassicas used commercially include a range of different species and interspecific hybrids serving diverse roles in livestock systems. Bulb turnips (Brassica campestris var. rapa L.) and swedes (B. napus var. napobrassica L.) have above- and belowground (bulb) biomass that can be grazed by livestock and are generally held as a standover feed until required and grazed in a single cycle or event. Kale (B. oleracea var. acephala L.) provides only aboveground biomass but grazing is typically managed in the same way as bulb turnips, although it can regrow after light grazing. Forage rape (B. napus var. biennis L.) and hybrid brassicas such as leafy turnip (B. rapa var. rapifera L. and B. campestris × napus) and raphanobrassica (B. oleracea var. acephala L. × Raphanus sativus L.) provide only aboveground biomass but are suitable for multiple grazings; however, different variety groups are recommended for different grazing windows. Even within these different forage ‘types’, forage brassicas are genetically diverse, with most evolving from three diploid species (B. oleracea, B. rapa, B, nigra) and three crosses of these diploid species (allotetraploid: B. napus, B. juncea, B. carinata) (Waters and Schaal 1996) as well as some interspecific hybrid genotypes. As a result, there are often considerable functional differences (phenology, plant allocation, nutritive value) between genotypes of the same type and/or species (de Ruiter et al. 2009).
It is clear that forage brassicas provide significant benefits to livestock systems in higher rainfall regions; however, in Australia’s mixed farming zone, they have the potential to diversify the forage grown on-farm and may help to fill feed gaps and reduce risks in forage-dependant livestock systems (Bell et al. 2018). The mixed farming zone is a major feature of Australia’s agricultural industry, occupying ~70 Mha of land (Healy et al. 2013; Bell et al. 2014) with a wide variety of soil types (e.g. black Vertosols to sands) and a comparatively hot and dry climate (Bell et al. 2020). Traditionally, crops were grown in rotation with self-regenerating ley pastures, but a recent shift towards more intensive cropping and use of annual forages and short-term pastures (1–3 years) in this zone (Howieson et al. 2000) has resulted in fewer forage options to fill winter feed gaps, and these options are largely limited to cereals such as forage oats (Avena sativa L.), dual-purpose wheat (Triticum aestivum L.), barley (Hordeum vulgare L.) and triticale (T. aestivum L. × Secale cereale L.). As a result, cropping rotations remain cereal dominated, and a greater diversity of broadleaf options is desirable to manage weeds and soilborne pests and diseases (Smith et al. 2004; Kirkegaard et al. 2014; Angus et al. 2015). However, in semi-arid subtropic regions, broadleaf options such as canola are rarely used in crop rotations owing to unreliable winter rainfall and a shorter growing season, which limits the productivity and overall profitability of the crop (Robertson and Holland 2004). Preliminary studies undertaken in southern Queensland (Qld) and northern New South Wales (NSW) under dry winter growing conditions identified some forage brassica genotypes that have production potential of >5 t/ha of dry matter (DM), which is comparable or superior to other commonly used forage cereals and alternative legumes (Bell et al. 2020). However, that study included relatively few genotypes from a narrow set of forage brassica species/types. In areas with reliable rainfall >450 mm, the increased adoption of dual-purpose canola (Brassica napus var. annua L.), which can be grazed by livestock in winter and harvested for grain in summer (Kirkegaard et al. 2016), indicates the desire of producers to diversify their forage options. The expansion of dual-purpose canola into the medium rainfall environments has the potential to contribute to on-farm profitability (McCormick et al. 2012) and has also heightened interest in the use of forage brassicas within this zone.
Although significant opportunities are apparent, there is limited knowledge and information about the suitability of the range of forage brassicas for use in drier environments within Australia’s mixed farming zone. In the present study, we aimed to address these limitations by evaluating the productivity of a broad range of forage brassica genotypes across multiple environments spanning different agro-climatic regions of the mixed farming zone, and comparing them to relevant benchmark species, principally oats and canola. We show that forage brassicas provide a useful forage alternative across many different environments, and thus offer more diversified forage options for livestock producers.
Materials and methods
Site locations and design
Edible biomass and yields of ME and CP of a broad range of forage brassica genotypes were evaluated across multiple environments and compared with benchmark forage species. The rationale for the selection of the forage brassica genotypes included in this study was to test a diversity of plant types (i.e. leaf type, bulb producing and hybrids) and cultivars within type that varied in functional traits, and thus, were expected to vary in productivity potential and adaptation. A multi-environment trial analysis was used to assess whether genotypes were niche specific or had broader potential use across the mixed farming zone.
The core experimental sites varied considerably in geographical location and soil type (Fig. 1). Eight experimental site-years were derived from two growing seasons (2018 and 2019) at four core experimental sites: Tummaville and Condamine in southern Qld, Iandra in central west NSW, and York in the central wheatbelt of Western Australia (WA) (Fig. 1). At each core site, eight or nine forage brassica genotypes were compared with the benchmark species of dual-purpose canola and forage oats. The cultivars of dual-purpose canola and forage oats used at each site were selected based on relevance to the region and sowing window (Table 1). Forage oats failed to germinate at the York 2019 site. Another two satellite sites in northern NSW, at Armatree and Pine Ridge (Fig. 1), established in 2019 compared eight forage brassica genotypes (including six genotypes used at the core sites) with forage barley, which is commonly used in this region (Table 1). Data from the satellite sites were included only in the multi-environment trial analyses. The details for each of the experimental sites, including location, soil type, sowing date, rainfall and irrigation received over growing season, and nitrogen (N) applied, are presented in Table 1.

|

|
All sites except the Pine Ridge 2019 satellite site included four replicates of genotypes arranged in a randomised complete block design. The Pine Ridge site had only three replicates. Plots at all sites varied in size from 40 to 70 m2 with 5–7 planting rows and row spacings 250–320 mm, depending on area and equipment available. Seed was sown at depths of 20–50 mm by knife point with a press wheel (at Tummaville 2018, Condamine 2018 and 2019, York 2018 and 2019) or tines (at Tummaville 2019, Iandra 2018 and 2019). The forage brassicas were sown at a set sowing rate (Table 2), and starter fertiliser (Granulock Z: 11% N; 21.8% P; 4% S; 1% Zn; Incitec Pivot, Melbourne) was applied at sowing.

|
The sites were monitored regularly for weed and insect infestations, fungal infections and diseases, and appropriate management was implemented when needed.
Forage sampling, processing and nutritive value analysis
At all sites, plant biomass samples were taken at several, but varying, times during the growing season based on there being adequate bulb and/or biomass for collection. To allow for cross-site comparisons with different sowing dates and environmental conditions, forage production among genotypes was compared within two phenological windows that represented an early (800–1300 growing degree days (GDD) after sowing) and late (1600–2100 GDD after sowing) grazing window. GDD were calculated based on the sum of average daily temperature after sowing, with a base temperature of 0°C. The base temperature for canola is also 0°C (Vigil et al. 1997), and so this was considered the most appropriate approach to deal with the diversity of genotypes. However, because of drought conditions and biomass availability, some sites were sampled within only one of these grazing windows.
Forage biomass was collected from quadrats (0.5–1.0 m2) in the central rows of each replicated plot. Both aboveground (leaf/petiole and stem) and belowground (bulb, for bulb species) portions were collected when sufficient bulb biomass had accumulated. Bulb/belowground biomass was not collected within the early grazing window at the Tummaville 2018 and Condamine 2019 core sites, or at the Armatree 2019 and Pine Ridge 2019 satellite sites.
Prior to drying, bulb and stem portions were chopped to a size of ~20 mm2. All forage samples were weighed and then dried in an oven at 60–80°C until constant dry weight and re-weighed for calculation of DM content and total edible biomass (t DM/ha).
At core sites, forage nutritive value was determined from biomass samples taken at each sampling time (above- and belowground portions were analysed separately). In the late grazing window at the Tummaville 2018 site, canola cv. Wahoo was in the late reproductive phase (seedset) and hence was not analysed for nutritive value. Samples were ground through a 1-mm sieve and stored in an airtight container. Forage samples were analysed for DM digestibility (DMD) (DMD corrected, DMDC) and total N at the CSIRO Rural Research Laboratory, Floreat, WA. Near-infrared reflectance spectrophotometry (NIRS; SpectraStar 2500X rotating top window system (680–2500 nm); Unity Scientific) was used to predict DMDC. Spectral data were collected by InfoStar3-11-1.exe software and converted using spc.in.exe software (Unity Scientific) for forage nutritive value predictions via a CSIRO feed calibration developed using a trimmed wave specification employing the wavelengths 1350–2500 nm, at intervals of 1 nm. The performance of the calibration equations was assessed by using R2 value and standard error of validation as criteria; for DMDC, R2 was 0.89 and standard error of validation was 1.61. A 10% subsample from the test set was also analysed by using wet chemistry for further validation of the predictions. Metabolisable energy was calculated from predicted DMDC for in vivo conditions using AFIA standards (AFIA 2014), as: 0.172 × DMDC – 1.707. Total N was measured by combustion (Dumas method; AFIA 2014) in a CN628 N Analyser (LECO). Crude protein (as % of DM) was calculated as: total N (% of DM) × 6.25.
Yields of ME (Eqn 1) and CP (Eqn 2) were calculated within each grazing window to account for both the biomass and nutritive value characteristics of the forages:
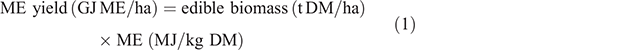
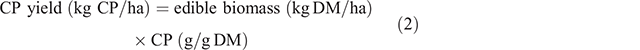
Site characterisations
Owing to the diversity of the environments where these forage brassicas were grown, soil cores were taken before sowing at all core sites to characterise for soil available water and nutrients. Four to six soil cores were taken to 1.5 or 1.8 m depth across each replicate block (n = 4 blocks). Soil cores were partitioned into soil layers 0–150, 150–300, 300–600, 600–900, 900–1200, 1200–1500 and 1500–1800 mm and split in half lengthwise. Each half was placed into separate soil bags for soil water and nutrient determinations, and layers within each replicate block were bulked into one representative sample. Soil water samples were weighed (taring bag weight) immediately following collection and subsequently dried at 100–105°C for 3–5 days (until constant dry weight) to determine gravimetric soil water content. Volumetric soil water was calculated using assumed bulk density values for previously characterised soils that were nearby the sites. Soil nutrient samples were dried at 40°C for 3–5 days, finely ground and stored in an airtight container before analysis at the CSBP Soil and Plant Analysis Laboratory, Bibra Lake, WA. Soil samples from the top four layers underwent a basic set of analyses for soil characterisation: the standard CSBP test including electrical conductivity, pH (in water and CaCl2), NO3, NH4, Colwell P, Colwell K, sulfur (KCl 40), organic carbon (Walkley Black). All other samples underwent analyses for NO3 and NH4 only.
Statistical analyses
Edible biomass and yields of ME and CP for each core experimental site within the early and late grazing windows were analysed with linear mixed models using REML with the lme4 package (Bates et al. 2015) in the statistical software program R (R Foundation, Vienna). All data were initially tested for normality and heteroscedasticity of the residuals and all data met these criteria. In all analyses, genotype was fitted as a fixed effect and replicate block was fitted as a random effect. Results were considered significant with a P-value ≤0.05.
A multi-environment trial analysis (Smith et al. 2005) for edible biomass and yields of ME and CP at both core and satellite sites was done by using linear mixed models with ASReml-R (Butler et al. 2017) to correlate the genetic (forage genotype) effects across environments, and employing factor analytic models (Smith et al. 2001). This method also accounted for other non-genetic sources of variation including experimental design and spatial location of experimental plots using the approach of Gilmour et al. (1997) and allowed each experiment to have a separate residual model. The factor analytic models provide an approximation to the fully unstructured covariance matrix (with a reduction in the number of parameters required) to model the genotype × environment interaction by estimating a separate genetic variance for each experiment and all pairs of genetic covariances between experiments (Kelly et al. 2007).
A square root transformation was applied to the edible biomass data for better approximation of the assumptions for the analysis. Genotype predictions (best linear unbiased predictors, BLUPs) were calculated for each experiment for all analysed traits, and genotype was ranked within experiment (on a square root scale for edible biomass). For the two grazing windows, a regression on the environment mean was used to explore the genotype × environment interactions (Finlay and Wilkinson 1963). The environmental mean was based on the mean of the BLUPs for all genotypes to represent best the sites with low and high productivity potential (Lawes et al. 2016).
Results
The growing conditions at all sites were very challenging with well below average annual rainfall conditions; in-crop rainfall ranged from 26 to 233 mm at all sites, except Tummaville 2019 that received 328 mm due to irrigation (Table 1). At some sites, decile 1 drought conditions were reported. Some irrigation was applied at the Tummaville 2018 and 2019 sites; however, total water received by these sites was similar to, or just below, the average growing season rainfall. This set of different production environments resulted in a wide range in forage productivity potential, with the mean edible biomass production across the sites in the range 2.0–6.4 t DM/ha in the early grazing window, and 0.6–8.5 t DM/ha in the late grazing window (Tables 3 and 4).

|

|
Productivity of forage brassica genotypes compared with benchmark species
Early grazing window
At all sites sampled within the early grazing window (800–1300 GDD after sowing), at least six of the forage brassicas produced edible biomass and ME yield similar to or higher than oats. More forage brassicas (eight or all nine genotypes) produced CP yield similar to or higher than oats across these sites (Table 3). A clear example of the forage brassicas outperforming oats in this early window was at the Tummaville 2019 site, where the top three forage brassicas outperformed oats by around 1.8 times for yields of ME (94 vs 51 GJ/ha) and CP (1670 vs 963 kg/ha) (Table 3). This was also evident at the Iandra 2019 site (lower mean site production), where forage brassicas outperformed oats by ~1.9 times for yields of ME (41 vs 22 GJ/ha) and CP (770 vs 408 kg/ha) (Table 3). Compared with dual-purpose canola, forage brassicas also ranked well for all productivity measures, with at least seven forage brassicas producing yields similar or higher at four of the five sites, and at least four of the forage brassicas at the other site (Table 3). Dual-purpose canola produced below site average edible biomass and ME yield at both the Tummaville 2018 and York 2018 sites, likely due to the use of triazine-tolerant cvv. Wahoo and Bonito, respectively, in these experiments. The other sites, which used hybrid cvv. 45Y91CL and Hyola970CL, had edible biomass and ME yields similar to or higher than the site mean.
Of the forage brassicas, the best performing genotypes in this early grazing window were Green globe and Rival turnip, and HT-R24 forage rape, which consistently ranked ~15% above the site mean across all sites for all productivity measures, except for Rival performing poorly at the Tummaville 2018 site. Across at least three of the five sites sampled, Goliath forage rape and Hunter leafy turnip also ranked above the site mean for all productivity measures. Regal kale performed poorly across all sites in this early grazing window and was on average 40% lower than the site mean for all productivity measures (Table 3).
Late grazing window
In the late grazing window (1600–2100 GDD after sowing), oats often produced more edible biomass (ranking 44% higher than the site mean) than the forage brassicas (Table 4). However, the forage brassicas had higher nutritive value than oats and during this late grazing window; yields of ME and CP for oats were only 22% and 1% higher than the site mean, respectively. Within this late grazing window, at least seven of the forage brassicas produced edible biomass similar to or higher than oats at three of the six sites, but very few (if any) of the forage brassicas outperformed oats at the other three sites. Pallaton was the only forage brassica to produce edible biomass similar to oats at the York 2018 site. On the other hand, the forage brassicas performed better for CP yield, with eight or all nine of the forage brassica genotypes producing CP yields similar to or higher than oats at four of six sites, and four or six of the forage brassicas at the other two sites (Table 4). This was most evident at the Tummaville 2019 site and both years at Condamine, where the CP yield of oats was ~20% lower than the site mean, and the top three performing forage brassica genotypes at these three sites produced 1.5–2 times more CP yield than oats (Table 4). In the late grazing window, there were some instances where dual-purpose canola was reproductive, particularly in the 2018 season; in these cases, the ME content of canola was lower than of the forage brassica genotypes (Supplementary Materials Table S1). Compared with dual-purpose canola, several forage brassicas (ranging from three to all nine genotypes) produced edible biomass and ME yields similar to or higher than canola. Many more of the forage brassicas produced CP yields similar to or higher than canola (ranging from five to all nine genotypes) across all sites in this late grazing window (Table 4).
The best performing forage genotypes in the early grazing window were not the best performing genotypes in the late grazing window. Oats performed the best for edible biomass, but in terms of yields of ME and CP, the best performing genotypes (i.e. those ranked most consistently higher than the site mean) were Goliath and HT-R24 forage rapes, Pallaton raphanobrassica and canola, all of which produced on average 16% higher yields than site means. The lowest performing genotypes were Hunter leafy turnip, Green globe turnip and Regal kale, which produced ~22% less than the site mean for all productivity measures (Table 4).
Genotype × environment interactions
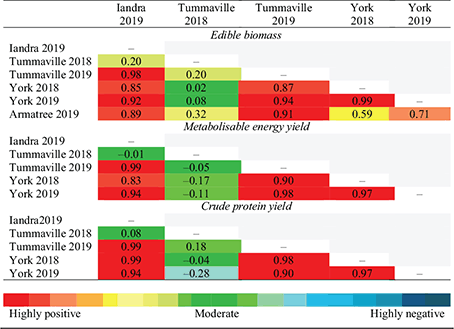
Across the range of production environments measured here, the multi-environment trial analysis revealed limited genotype × environment interactions in the early grazing window; however, in the late grazing window, several genotype × environment interactions were found for the various productivity measures (Tables 5 and 6). In the early grazing window, there were no genotype × environment interactions for edible biomass, but there were some slightly negative correlations for ME yield and particularly CP yield. These slightly negative correlations between sites always featured the Tummaville 2018 site (Table 5) and were strongly related to the lower performance of Rival at this site, which was inconsistent with the other sites (Fig. 2a–c). Within this early grazing window, all genotypes other than Rival had a stable performance ranking across the range of production environments. Relative to the other genotypes, Rival was highly responsive in higher production environments, as indicated by a steeper regression (Fig. 2a–c). To illustrate, Goliath had similar relative ME yield to Rival in low production environments, but a much lower relative performance in higher production environments (Fig. 2b).
|

|

|
In the late grazing window, some moderate negative correlations between experimental sites were identified, particularly for yields of ME and CP. In the late grazing window, there was no single forage brassica genotype that ranked consistently across all production environments for any of the productivity measures. Moderate negative correlations for ME yield were mostly related to the Tummaville 2019 site, and moderate negative correlations for CP yield were related to both the Tummaville 2019 and Condamine 2018 sites (Table 6). This indicated that several of the forage genotypes ranked very differently within the Tummaville 2019 experiment compared with their ranking at the sites that had lower production potential (Fig. 2e, f). This was most apparent for Hunter leafy turnip, which performed poorly for all productivity measures in environments with low–moderate production potential but quite favourably at the Tummaville 2019 site (Fig. 2d–f). This revealed that some genotypes, such as Hunter, are better suited to higher production environments than to low production environments (i.e. drier environments), where they were poorly adapted. The moderate negative correlations for CP yield related to the Condamine 2018 site were due to the relative ranking of several genotypes including oats, Graza, Winfred and Goliath, which varied considerably at this site compared with the York 2018 and Iandra 2018 sites (Figs 2d–f). It is not known why these genotypes responded so differently because they were all grown in low–moderate production environments. Of the forage brassicas, Pallaton had high relative productivity at sites with low–moderate mean production but was far less responsive at sites with high production potential in the late grazing window (Figs 2d–f).
Discussion
The most important finding of this study was that several forage brassica genotypes produced yields of ME and CP similar to or greater than forage oats across a range of environments spanning Australia’s mixed farming zone. Although the forage cereals produced more edible biomass in some instances in the late grazing window, they declined in nutritive value and yields of ME and CP relative to the forage brassicas. Compared with canola, a broad range of forage brassicas had edible biomass and yields of ME and CP that were similar or higher in the early grazing window, when canola would be typically grazed. In the late grazing window, comparisons were made between canola and the forage brassicas, although in some instances, the canola was already reproductive. This was most evident in many of the 2018 site-years where sowing was delayed because of insufficient early-season rainfall, and hence, faster developing spring canola cultivars (i.e. triazine-tolerant Wahoo and Bonito) were used. In a commercial system, grazing of canola would cease just before bud elongation (Sprague et al. 2015). The potential value of forage brassicas in drier environments was further highlighted by the ability of several genotypes, namely forage rape cultivars and Pallaton, to remain productive under very dry and challenging growing conditions across most sites within the late grazing window.
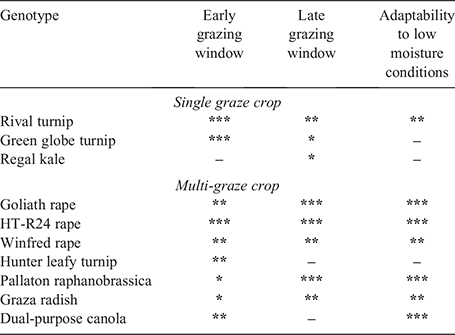
Measurement of yields of ME and CP was important in this study for quantifying forage value, because ME and CP are essential nutrient components for ruminant production (ACC 1990; Van Soest 1994). The results from this study confirm that forage brassicas grown in drier environments are able to maintain nutritive value for longer than cereal forages because they have an extended vegetative state. In this study, some genotypes such as Pallaton showed no signs of reproductive development throughout the experimental period, even into early summer (e.g. November or December; data not shown), and this is due to the high vernalisation requirements of many forage brassica genotypes. Forage brassica genotypes with particularly high vernalisation requirements, such as Pallaton and forage rapes (Paridaen and Kirkegaard 2015), are of considerable advantage to livestock production systems within the mixed farming zone, especially in those areas where crops are less exposed to vernalising temperatures (i.e. higher minimum temperatures), such as semi-arid subtropics. Thus, the integration of forage brassicas could extend the window during which nutritious feed is available for grazing, and hence provide greater flexibility around how they could be used to fill feed gaps when forage nutritive value or quantity is limiting livestock production. These attributes of forage brassicas are likely to be of major benefit in meat-producing livestock systems within drier environments, where higher ME and CP yields are necessary to maximise animal liveweight gain to meet target market specifications. Using the nutritive value attributes measured for the forages in this study (see Tables S1 and S2), the GrazFeed model (Freer et al. 1997) predicts average daily gain of lambs grazing forage brassicas in the early grazing window to be 280 g/head.day, compared with only 160 g/head.day for those grazing oats. Furthermore, the relative difference is larger in the late grazing window, where predicted daily growth rate of lambs was 94 g/head.day for oats and 250 g/head.day for forage brassicas. The high livestock productivity potential and fast-growing nature of several forage brassicas grown in diverse environments, such as Hunter, Green globe and Rival, will appeal to producers looking for an early-season forage of high nutritive value. However, forage brassicas such as Pallaton and forage rapes HT-R24 and Goliath are likely to be of greatest value later in the season when other forage options such as dual-purpose canola are reproductive and no longer suitable for grazing, and the feeding value of forage cereals is rapidly declining. This will make forage brassicas, particularly multi-graze species such as Pallaton and forage rapes, appealing to late-season livestock finishing systems within the mixed farming zone, especially in low production environments (i.e. low available moisture), where they were shown to reliably produce forage of high nutritive value (Table 7). However, the grazing management guidelines of forage brassica grown in drier environments will need to be better defined to ensure that productivity and utilisation by livestock is maximised.
|
The higher nutritive value of forage brassicas, as further highlighted in this study, may have other livestock systems benefits. Sheep fed forage brassicas have been shown to produce less methane per unit DM intake than those fed grasses (Sun et al. 2012, 2016), and this is correlated with ME and digestibility (Sun et al. 2012); the forage brassicas in this study were high in ME content (~11.8–12.0 MJ ME/kg DM) and DMD (~77–80%). Rumen conditions produced when lambs graze brassicas appear to decrease methanogenic activity during fermentation (Sun et al. 2015). Brassicas may also lower methane intensity because livestock can reach their target weights in less time, as demonstrated in the above GrazFeed model predictions. Given the high digestibility and ME content of the forage brassicas relative to the forage oats in this study, there is considerable potential for forage brassicas to reduce ruminant-derived carbon emissions. Despite the range of livestock production benefits, there are challenges to maximising production in brassica-fed livestock, including early suppression of intake, possibly associated with anti-nutritional factors (Nichol 2003), the risk of ruminal acidosis induced by insufficient neutral detergent fibre content (Westwood and Mulcock 2012), and the risk of photosensitisation where livestock intakes are not appropriately managed, such as break-feeding, as done in dairy systems (Collett and Matthews 2014). These issues can reduce the feeding value of forage brassicas and complicate their grazing management (Barry 2013), especially in extensive systems where greater livestock numbers and paddock size limit the rigorous monitoring of animals. Furthermore, low-yielding crops grown under hot, dry ‘drought stressed’ conditions such as those that occur in Australia’s mixed farming zone may also increase the risk of higher concentrations of anti-nutritional compounds and associated health disorders such as photosensitisation (Morton and Campbell 1997; Collett and Matthews 2014).
Among the diverse set of forage brassica genotypes tested in this study, we identified several genotypic adaptations that have implications for their use in the drier environments in Australia’s mixed farming zone. First, the forage rapes HT-R24 and Goliath produced edible biomass and yields of ME and CP that ranked most consistently above the site mean within both the early and late grazing windows (Table 7) across multiple environments. The consistently good performance of the forage rapes was not observed in the other forage brassicas, suggesting that these modern forage rape cultivars, and potentially other forage rape cultivars not included in this study, are viable in drier environments. This was particularly evident given the challenging growing conditions in this study. The ability of the forage rapes to remain productive under these dry conditions, relative to the other genotypes, indicates that they were able to access moisture to sustain late-season growth. This demonstrates that forage rapes are adaptable to environments with low available moisture (Table 7). Brassica species including forage rapes are known to have a deep root system (Paridaen and Kirkegaard 2015; Kirkegaard et al. 2021) that enables them to extract more soil water from deeper stores (Fletcher et al. 2010), which may explain their good performance under the dry conditions. However, root depth was not measured at any of the sites in this study and so this suggestion cannot be confirmed.
The productivity of Pallaton was lower than that of many other forage brassicas within the early grazing window; however, in the late grazing window, it performed well compared with other genotypes, particularly at sites with low moisture availability (Table 7). Why Pallaton is so persistent later in the season is unclear but may be attributable to its deep root system (A. Dumbleton, per. comm.) enabling it to persist under the dry conditions, in contrast to some of the other forage brassica genotypes. In both years at York in the WA wheatbelt, Pallaton yielded more than all other forage brassicas in the late grazing window and maintained green leaf far later into the season under the hot, dry Mediterranean summer climatic conditions. At all other sites, Pallaton yielded similarly to the other forage brassicas. An explanation for the lower early-season production from Pallaton than from the other forage brassicas may be the low plant populations that were achieved compared with the forage rapes (53%). Pallaton seed is 2.5 times larger than forage rapes cvv. Goliath and Winfred but was sown at the same rate in this study. Early biomass growth may have also been limited by a slower rate of leaf appearance (Adams et al. 2005; Fletcher et al. 2012). However, this cannot be confirmed because leaf appearance rate and canopy development of this new species has not yet been defined. Although raphanobrassica is a kale-cross hybrid, low plant populations did not appear to impact nutritive value in Pallaton, particularly late in the season, unlike kale, for which higher sowing rates are favourable to improve nutritive value by reducing plant stem size (de Ruiter et al. 2009). However, the functional traits of Pallaton are very different from those of older raphanobrassica cultivars (Harper and Compton 1980).
Our data suggest that Green globe and Hunter were much better suited to the early grazing window (Table 7) because they began to senesce in the late grazing window, when their relative production and yields of ME and CP were reduced. Green globe is known to have a higher rate of leaf senescence than Goliath and kale cvv. Gruner and Kestral, but it also has a higher rate of leaf appearance (Adams et al. 2005). In a rainfed system in north-west Tasmania, summer-grown turnips have shown more favourable early growth (90 days after sowing), yielding 1.9 times more than kale and 1.6 times more than forage rape (Neilsen et al. 2000). In our study, within the early grazing window, which was similar to 90 days after sowing, bulb turnips yielded on average 2.0–2.1 times more than kale cv. Regal, and up to 1.2 times more than the average of the forage rape genotypes. The very dry conditions in this study meant a higher rate of senescence of Green globe and Hunter, indicating that they are poorly adapted to dry conditions and unable to persist late in the season (Table 7). This poor adaptability to dry conditions may be caused by their shallower root systems (de Ruiter et al. 2009), which prevent them from extracting deeper soil moisture compared with the deeper rooted forage rape (Fletcher et al. 2010).
Finally, kale cv. Regal yielded poorly relative to the other forage brassicas within both grazing windows and across most environments in this study, which suggests that it has less application in these drier, hotter environments (Table 7). Kale biomass potential, in particular, is known to be significantly limited by insufficient soil water (Chakwizira et al. 2013; Chakwizira and De Ruiter 2014), and water use efficiency of kale is up to 2.5 times lower than other forage brassicas (Neilsen et al. 2000). In addition, leaf appearance rate of kale is lower than of other forage brassicas (Adams et al. 2005), and since thermal time affects leaf appearance, the cooler season sowing, particularly in the 2018 experiments, may have further impaired kale productivity, particularly in the early grazing window.
Guidelines for the agronomic management of forage brassicas that best suit drier environments are limited and there is likely potential to enhance performance further than is reported here. The sowing dates and sowing rates of the forage brassicas used in this study were largely a ‘best guess’ based on some adjustments to the growing guidelines for forage brassicas in higher rainfall systems. The growing conditions in several of those studies resulted in sowing dates that may have been less than optimal for forage brassicas (i.e. too early or too late). Sowing date is likely to be a significant factor because soil temperature at the time of sowing is known to be a critical driver of seedling emergence and early plant vigour in brassicas (Jacobs et al. 2001). Among our experimental sites, this is best demonstrated by considering the productivity of forage brassicas sown in the subtropical location of Tummaville in April 2019, which produced substantially more edible biomass than those sown in June of the previous year. The average daily temperatures at sowing at these two site-years were 20°C and 12°C, respectively. By comparison, the productivity of forage brassicas was similar in York for both site-years when they were sown within a week of each other. Further to this point, we investigated autumn–winter sown forage brassicas in this study; however, in higher rainfall systems, forage brassicas are typically sown in spring for a summer–autumn feed. The potential for spring-sown brassicas in areas of the mixed farming zone, particularly those with higher summer rainfall, should be explored, but likely targeting those genotypes with greater drought tolerance because drier and hotter environments, as occur in many areas of the mixed farming zone, are likely to be a considerable challenge to short- and long-term forage productivity (Bell et al. 2020). It is also possible that sowing rates of some genotypes within this study were suboptimal, notably Pallaton, and higher plant populations may have improved productivity within the early grazing window. However, in drier environments, reducing sowing rates may optimise productivity relative to input costs (Bell et al. 2020) and may be an acceptable trade-off for producers looking for forage options with relatively low upfront costs, similar to forage oats. Furthermore, in order to compete with forage oats as a reliable forage source in drier environments, better guidelines for optimal sowing depths of forage brassicas is warranted to ensure more reliable germination and establishment. In our study, forage oats exceeded plant establishment targets at three of the core sites, where forage brassicas were well below establishment targets and rarely reached or exceeded them (Table 2). Finally, the forage brassicas were not grazed in these experiments, and it is possible that the productivity of multi-graze species such as forage rapes, Hunter and Pallaton would have been enhanced compared with the management imposed, which required compromise among multiple species/types.
Conclusions
This multi-environment study has shown that, across a diverse range of environments spanning Australia’s mixed farming zone, several forage brassica genotypes offer forage production similar to or better than forage cereals and canola. Although forage brassicas did not always produce the most edible biomass, their high forage nutritive value meant that they often produced higher yields of ME and CP, particularly in the late grazing windows. This underlines the importance of considering the nutritive value of forages of different types when making comparisons and the need to calculate productivity measures such as yields of ME and CP rather than comparing only edible biomass production. Forage brassicas have the potential to diversify the on-farm feedbase and serve as a break-crop with livestock systems benefits within Australia’s mixed farming zone. The integration of forage brassicas within this zone will be of particular importance in regions where canola is not commonly grown and/or where late season feed gaps occur.
Conflicts of interest
The authors declare no conflicts of interest.
Acknowledgements
The data presented in this paper were collected and collated during a Meat & Livestock Australia Donor Co. project (P.PSH.1044: Improving the use of forage brassicas in mixed farming systems). The authors thank collaborative bodies including Matt Gardner and staff at Agricultural Marketing and Production Systems (AMPS) research, Tamworth, NSW, for taking biomass measurements at the satellite sites, and Rod and Nick Kershaw, Iandra, NSW, and Duane and Buddy Statham, Condamine, Qld, for the use of their land and assistance in managing the experiments at these sites. The authors also thank Rebecca Smith from Living Farm, York, WA, and Matt Burley and the staff from Kaylx, Toowoomba (Tummaville, Qld) for assisting with the sowing and agronomic management of these sites.
References
ACC (1990) ‘Feeding standards for Australian livestock: ruminants.’ (CSIRO Publishing: Melbourne)Adams C, Scott WR, Wilson DR, Purves L (2005) Dry matter accumulation and phenological development of four brassica cultivars sown in Canterbury. Agronomy New Zealand 35, 1–18.
AFIA (2014) ‘AFIA laboratory methods manual.’ (Australian Fodder Industry Association: Melbourne)
Angus JF, Kirkegaard JA, Hunt JR, Ryan MH, Ohlander L, Peoples MB (2015) Break crops and rotations for wheat. Crop & Pasture Science 66, 523–552.
| Break crops and rotations for wheat.Crossref | GoogleScholarGoogle Scholar |
Barry TN (2013) The feeding value of forage brassica plants for grazing ruminant livestock. Animal Feed Science and Technology 181, 15–25.
| The feeding value of forage brassica plants for grazing ruminant livestock.Crossref | GoogleScholarGoogle Scholar |
Bates D, Mächler M, Bolker BM, Walker SC (2015) Fitting linear mixed-effects models using lme4. Journal of Statistical Software 67, 1–48.
| Fitting linear mixed-effects models using lme4.Crossref | GoogleScholarGoogle Scholar |
Bell LW, Moore AD, Kirkegaard JA (2014) Evolution in crop-livestock integration systems that improve farm productivity and environmental performance in Australia. European Journal of Agronomy 57, 10–20.
Bell LW, Moore AD, Thomas DT (2018) Integrating diverse forage sources reduces feed gaps on mixed crop–livestock farms. Animal 12, 1967–1980.
| Integrating diverse forage sources reduces feed gaps on mixed crop–livestock farms.Crossref | GoogleScholarGoogle Scholar | 29198288PubMed |
Bell LW, Watt LJ, Stutz RS (2020) Forage brassicas have potential for wider use in drier, mixed crop–livestock farming systems across Australia. Crop & Pasture Science 71, 924–943.
| Forage brassicas have potential for wider use in drier, mixed crop–livestock farming systems across Australia.Crossref | GoogleScholarGoogle Scholar |
Butler DG, Cullis BR, Gilmour AR, Gogel BJ, Thompson R (2017) ‘ASReml-R reference manual. Version 4.’ Manual 176. (VSN International: Hemel Hempstead, UK) http://www.homepages.ed.ac.uk/iwhite/asreml/uop
Chakwizira E, De Ruiter JM (2014) Water use efficiency of forage kale crops. In ‘Proceedings 6th Australasian Dairy Science Symposium’. Hamilton, New Zealand. (Ed. J Roche) pp. 100–104. (Australasian Dairy Science Symposium)
Chakwizira E, Gillespie R, Maley S, George M, Michel A (2013) Water and nitrogen use efficiency of forage kale crops. Agronomy New Zealand 43, 1–16.
Collett M, Matthews Z (2014) Photosensitivity in cattle grazing Brassica crops. International Journal of Poisonous Plant Research 3, 6–21.
de Ruiter JM, Wilson D, Maley S, Fletcher A, Fraser T, Scott WR, Dumbleton A, Nichol WW (2009) ‘Management practices for forage brassicas.’ (Forage Brassica Development Group, New Zealand)
Dumbleton A, Gowers S, Conner A, Christie M, Kenny P, Mulcock H, Charteris B (2012) Cleancrop (TM) brassica system: the development of herbicide resistant brassica crops for New Zealand farming systems. In ‘Proceedings New Zealand Grassland Association’. pp. 25–30. (New Zealand Grassland Association)
Finlay KW, Wilkinson GN (1963) The analysis of adaptation in a plant-breeding programme. Australian Journal of Agricultural Research 14, 742–754.
| The analysis of adaptation in a plant-breeding programme.Crossref | GoogleScholarGoogle Scholar |
Fletcher AL, Sinton SM, Gillespie RN, Maley S, Sim RE, De Ruiter JM (2010) Drought response and water use efficiency of forage brassica crops. In ‘Proceedings Agronomy Society of New Zealand’. pp. 105–117. (Agronomy Society of New Zealand)
Fletcher AL, Chakwizira E, Maley S, George M (2012) Canopy development and radiation use efficiency of four forage brassica crops. In ‘Proceedings 16th Australian Agronomy Conference’. Armidale, NSW. (Ed. I Yunusa) (Australian Society of Agronomy)
Freer M, Moore A, Donnelly J (1997) GRAZPLAN: decision support systems for Australian grazing enterprises—II. The animal biology model for feed intake, production and reproduction and the GrazFeed DSS. Agricultural Systems 54, 77–126.
| GRAZPLAN: decision support systems for Australian grazing enterprises—II. The animal biology model for feed intake, production and reproduction and the GrazFeed DSS.Crossref | GoogleScholarGoogle Scholar |
Gilmour AR, Cullis BR, Verbyla AP (1997) Accounting for natural and extraneous variation in the analysis of field experiments. Journal of Agricultural, Biological & Environmental Statistics 2, 269–293.
| Accounting for natural and extraneous variation in the analysis of field experiments.Crossref | GoogleScholarGoogle Scholar |
Harper F, Compton IJ (1980) Sowing date, harvest date and the yield of forage brassica crops. Grass and Forage Science 35, 147–157.
| Sowing date, harvest date and the yield of forage brassica crops.Crossref | GoogleScholarGoogle Scholar |
Healy D, Gilmour P, Campbell J (2013) Grain and Graze 2 Impact Report. Report for Grains Research and Development Corporation. GRDC, Canberra, ACT.
Howieson JG, O’Hara GW, Carr SJ (2000) Changing roles for legumes in Mediterranean agriculture: developments from an Australian perspective. Field Crops Research 65, 107–122.
| Changing roles for legumes in Mediterranean agriculture: developments from an Australian perspective.Crossref | GoogleScholarGoogle Scholar |
Jacobs JL, Ward GN, McDowell AM, Kearney GA (2001) A survey on the effect of establishment techniques, crop management, moisture availability and soil type on turnip dry matter yields and nutritive characteristics in western Victoria. Australian Journal of Experimental Agriculture 41, 743–751.
| A survey on the effect of establishment techniques, crop management, moisture availability and soil type on turnip dry matter yields and nutritive characteristics in western Victoria.Crossref | GoogleScholarGoogle Scholar |
Kelly AM, Smith AB, Eccleston JA, Cullis BR (2007) The accuracy of varietal selection using factor analytic models for multi-environment plant breeding trials. Crop Science 47, 1063–1070.
| The accuracy of varietal selection using factor analytic models for multi-environment plant breeding trials.Crossref | GoogleScholarGoogle Scholar |
Kirkegaard JA, Conyers MK, Hunt JR, Kirkby CA, Watt M, Rebetzke GJ (2014) Sense and nonsense in conservation agriculture: principles, pragmatism and productivity in Australian mixed farming systems. Agriculture, Ecosystems & Environment 187, 133–145.
| Sense and nonsense in conservation agriculture: principles, pragmatism and productivity in Australian mixed farming systems.Crossref | GoogleScholarGoogle Scholar |
Kirkegaard JA, Lilley JM, Morrison MJ (2016) Drivers of trends in Australian canola productivity and future prospects. Crop & Pasture Science 67, i–ix.
| Drivers of trends in Australian canola productivity and future prospects.Crossref | GoogleScholarGoogle Scholar |
Kirkegaard JA, Lilley JM, Berry PM, Rondanini DP (2021) Canola. ‘Crop physiology case histories for major crops’. (Eds V Sadras, D Calderini) pp. 518–549. (Elsevier: Amsterdam)
Lawes RA, Huth ND, Hochman Z (2016) Commercially available wheat cultivars are broadly adapted to location and time of sowing in Australia’s grain zone. European Journal of Agronomy 77, 38–46.
| Commercially available wheat cultivars are broadly adapted to location and time of sowing in Australia’s grain zone.Crossref | GoogleScholarGoogle Scholar |
Lindsay CL, Kemp PD, Kenyon PR, Morris ST (2007) Summer lamb finishing on forage crops. In ‘Proceedings New Zealand Society of Animal Production’. pp. 121–125. (New Zealand Society of Animal Production)
McCormick JI, Virgona JM, Kirkegaard JA (2012) Growth, recovery, and yield of dual-purpose canola (Brassica napus) in the medium-rainfall zone of south-eastern Australia. Crop & Pasture Science 63, 635–646.
| Growth, recovery, and yield of dual-purpose canola (Brassica napus) in the medium-rainfall zone of south-eastern Australia.Crossref | GoogleScholarGoogle Scholar |
Moate PJ, Roche JR, Durling P (1996) Fodder turnips on Gippsland dairy farms. In ‘Proceedings Grassland Society of Victoria 37th Annual Conference’. Mornington, Vic. pp. 141–142. (Grassland Society of Victoria)
Morton JM, Campbell PH (1997) Disease signs reported in south-eastern Australian dairy cattle while grazing Brassica species. Australian Veterinary Journal 75, 109–113.
| Disease signs reported in south-eastern Australian dairy cattle while grazing Brassica species.Crossref | GoogleScholarGoogle Scholar | 9066967PubMed |
Neilsen JE, Rowe BA, Lane P (2000) Water use efficiency of four irrigated brassica forage crops. In ‘Proceedings Irrigation Association of Australia Conference’. Melbourne, Vic. pp. (Ed. G Connellan) pp. 136–139. (Irrigation Association of Australia: Melbourne)
Nichol WW (2003) Variation between forage rape (Brassica napus) cultivars for live-weight gain is due to anti-nutritional compounds. MSc. thesis, Lincoln University, New Zealand.
Nie ZN, Slocombe L, Behrendt R, Raeside M, Clark S, Jacobs JL (2020) Feeding lambs proportional mixtures of lucerne (Medicago sativa) and forage brassica (Brassica napus) grown under warm and dry conditions. Animal Production Science
| Feeding lambs proportional mixtures of lucerne (Medicago sativa) and forage brassica (Brassica napus) grown under warm and dry conditions.Crossref | GoogleScholarGoogle Scholar |
Paridaen A, Kirkegaard JA (2015) Forage canola (Brassica napus): spring-sown winter canola for biennial dual-purpose use in the high-rainfall zone of southern Australia. Crop & Pasture Science 66, 275–286.
| Forage canola (Brassica napus): spring-sown winter canola for biennial dual-purpose use in the high-rainfall zone of southern Australia.Crossref | GoogleScholarGoogle Scholar |
Raeside M., Byron J, Cameron F, MacDonald C, Partington D, Kearney GAB, Behrendt RA (2020) Spring-sown winter-type canola (Brassica napus) as summer–autumn forage for sheep production in southern Australia. Animal Production Science
| Spring-sown winter-type canola (Brassica napus) as summer–autumn forage for sheep production in southern Australia.Crossref | GoogleScholarGoogle Scholar |
Robertson MJ, Holland JF (2004) Production risk of canola in the semi-arid subtropics of Australia. Australian Journal of Agricultural Research 55, 525–538.
| Production risk of canola in the semi-arid subtropics of Australia.Crossref | GoogleScholarGoogle Scholar |
Smith AB, Cullis BR, Thompson R (2001) Analyzing variety by environment data using multiplicative mixed models and adjustments for spatial field trend. Biometrics 57, 1138–1147.
| Analyzing variety by environment data using multiplicative mixed models and adjustments for spatial field trend.Crossref | GoogleScholarGoogle Scholar |
Smith BJ, Kirkegaard JA, Howe GN (2004) Impacts of Brassica break-crops on soil biology and yield of following wheat crops. Australian Journal of Agricultural Research 55, 1–11.
| Impacts of Brassica break-crops on soil biology and yield of following wheat crops.Crossref | GoogleScholarGoogle Scholar |
Smith AB, Cullis BR, Thompson R (2005) The analysis of crop cultivar breeding and evaluation trials: an overview of current mixed model approaches. The Journal of Agricultural Science 143, 449–462.
| The analysis of crop cultivar breeding and evaluation trials: an overview of current mixed model approaches.Crossref | GoogleScholarGoogle Scholar |
Sprague SJ, Kirkegaard JA, Graham JM, Bell LW, Seymour M, Ryan M (2015) Forage and grain yield of diverse canola (Brassica napus) maturity types in the high-rainfall zone of Australia. Crop & Pasture Science 66, 260–274.
| Forage and grain yield of diverse canola (Brassica napus) maturity types in the high-rainfall zone of Australia.Crossref | GoogleScholarGoogle Scholar |
Sun XZ, Waghorn GC, Hoskin SO, Harrison SJ, Muetzel S, Pacheco D (2012) Methane emissions from sheep fed fresh brassicas (Brassica spp.) compared to perennial ryegrass (Lolium perenne). Animal Feed Science and Technology 176, 107–116.
| Methane emissions from sheep fed fresh brassicas (Brassica spp.) compared to perennial ryegrass (Lolium perenne).Crossref | GoogleScholarGoogle Scholar |
Sun X, Henderson G, Cox F, Molano G, Harrison SJ, Luo D, Janssen PH, Pacheco D (2015) Lambs fed fresh winter forage rape (Brassica napus L.) emit less methane than those fed perennial ryegrass (Lolium perenne L.), and possible mechanisms behind the difference. PLoS One 10, e0119697
| Lambs fed fresh winter forage rape (Brassica napus L.) emit less methane than those fed perennial ryegrass (Lolium perenne L.), and possible mechanisms behind the difference.Crossref | GoogleScholarGoogle Scholar | 26717241PubMed |
Sun X, Pacheco D, Luo D (2016) Forage brassica: a feed to mitigate enteric methane emissions? Animal Production Science 56, 451–456.
| Forage brassica: a feed to mitigate enteric methane emissions?Crossref | GoogleScholarGoogle Scholar |
Van Soest PJ (1994) ‘Nutritional ecology of the ruminant.’ (Cornell University Press: Ithaca, NY, USA)
Vigil MF, Anderson RL, Beard WE (1997) Base temperature and growing-degree-hour requirements for the emergence of canola. Crop Science 37, 844–849.
| Base temperature and growing-degree-hour requirements for the emergence of canola.Crossref | GoogleScholarGoogle Scholar |
Waters ER, Schaal BA (1996) Biased gene conversion is not occurring among rDNA repeats in the Brassica triangle. Genome 39, 150–154.
| Biased gene conversion is not occurring among rDNA repeats in the Brassica triangle.Crossref | GoogleScholarGoogle Scholar | 18469883PubMed |
Westwood CT, Mulcock H (2012) Nutritional evaluation of five species of forage brassica. In ‘Proceedings New Zealand Grassland Association’. pp. 31–38. (New Zealand Grassland Association)