

Carbon sequestration in woody biomass of mulga (Acacia aneura) woodlands: confidence in prediction using the carbon accounting model FullCAM
Keryn I. Paul A and Stephen H. Roxburgh A *A
Abstract
Decreased grazing and/or cessation of land clearing across Australia’s rangelands are being used to promote carbon sequestration through regeneration of woody biomass, predominately in Acacia aneura (mulga) woodlands. Changes in carbon stock are predicted using the carbon accounting model FullCAM. We collated datasets to assess the level of confidence in applying FullCAM to mulga regeneration across south-western Queensland and north-western New South Wales, with respect to model accuracy, specificity, and comprehensiveness. We found that FullCAM predictions were moderately accurate, with independent verification sites (N = 102) indicating model efficiencies of 48–70% and bias of −3.50 to −0.99 Mg DM ha−1, depending on calculation method. To ensure accuracy and to reduce risks of over-prediction, it is recommended FullCAM should be limited to sites with regeneration ages of ≤25 years and with levels of pre-existing above-ground biomass less than approximately 5 Mg DM ha−1. The paucity of data from mulga ecosystems in central and western Australia was identified as an important research gap. Regarding specificity, FullCAM has been calibrated to average rates of regeneration, generalised across a range of vegetation types, disturbance histories, and grazing management practices. This generalisation ensures accuracy when applied over broad spatial domains, but may limit the model’s accuracy at specific locations. For example, at the site scale, long-term grazing exclosure experiments (N = 34) have shown a wide range of regeneration outcomes (−0.52 to 1.85 Mg DM ha−1 year−1, with an average of 0.29 Mg DM ha−1 year−1), with site-scale contributors to this variability including the proportion of mulga in the total biomass, and the degree of change in grazing intensity (e.g. exclusion of livestock only, cf. exclusion of livestock plus native and feral animals). Regarding model comprehensiveness, new field data suggest that FullCAM could be extended to include standing dead pools of woody biomass, which contribute, on average, 17% of total woody biomass in mulga woodlands.
Keywords: abatement, Acacia aneura, baseline, FullCAM, grazing, regeneration, regrowth, standing dead.
Introduction
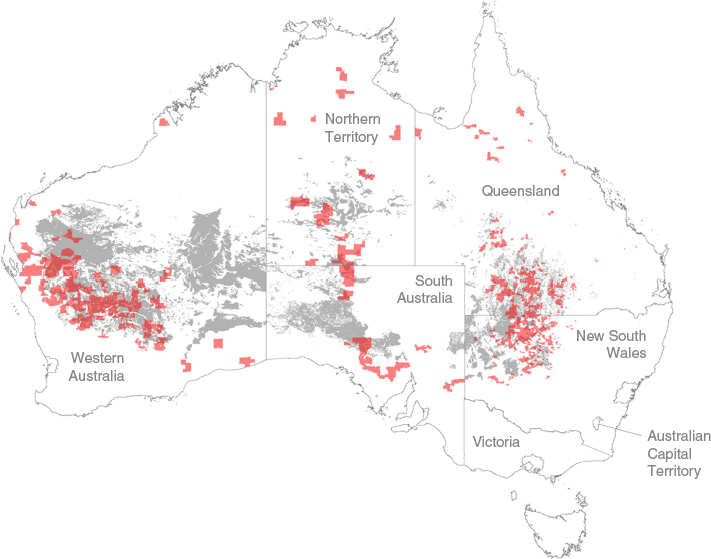
Rangelands occupy about 54% of the Earth’s land surface, and much of this area is grazed woodlands with woody biomass that may be managed in a way that contributes to climate change mitigation (e.g. Yirdaw et al. 2017; Feng et al. 2020). In Australia, this is of particular interest for the 9.2 million km2 (12% of the continent) of acacia woodlands, which are largely dominated by the native tree mulga (Acacia aneura F.Muell. Ex Benth.; Everist 1969; Howden et al. 2001; Baumber et al. 2020). Mulga is commonly used as fodder via direct browsing or following cutting/pushing (hereafter referred to as clearing). About 5% (0.42 million km2) of this acacia woodland area currently encompasses ‘Human-Induced Regeneration of a Permanent Even-aged Native Forests’ projects (HIR projects; AG 2020) (Fig. 1).
Map of Australia, indicating the location of acacia woodland (grey shading, as described by Pasut et al. 2025) and overlayed with current HIR project areas (red shading, source: https://data.gov.au/data/dataset/erf_project_mapping). Internal boundaries delineate Australian states and territories.
These HIR projects receive Australian carbon credit units (ACCUs) for sequestering carbon in live woody biomass, litter and coarse woody debris as the site regenerates to achieve the ‘forest’ cover threshold of 20% canopy cover and 2 m tall (AG 2018), typically following changed grazing management (Macintosh et al. 2024a). Indeed, where mulga tree foliage is within reach, even light browsing can suppress growth (Brown 1985), and so a change of management to release this suppression has potential to accumulate biomass in resulting regeneration (Harrington 1979; Moore et al. 2001; Witt et al. 2011; Daryanto et al. 2013). Throughout this paper, we use the term ‘grazing’ to refer generally to grazing land management and the overall impacts that grazing has on vegetation. The term ‘browsing’ is used when specifically referring to the direct impacts of animals consuming woody vegetation.
The assumption on which HIR activities are based is that historical land management has reduced or ‘suppressed’ the biomass below its potential, and following release from suppression, the vegetation is assumed to fully recover. Note that if significant loss of ecosystem integrity has occurred, for example, through the loss of the topsoil, then regeneration may be compromised, and complete recovery may not occur. One of the conditions for project approval under the HIR rules is the demonstration of regeneration potential within the proposed project area.
In the more productive parts of mulga woodlands in south-western Queensland and north-western New South Wales (Fig. 1), where there is a history of clearing and re-clearing, a mosaic landscape of mulga trees of varying ages and heights has been created to ensure availability of mulga fodder, especially during periods of drought (e.g. Page et al. 2008; Bowen et al. 2022). Mulga clearing involves branches being cut from trees, or by pushing or pulling trees so that the relatively nutritious canopy leaves become available for domestic stock, following which mulga plants are allowed to regrow or regenerate before the process is repeated (Fensham et al. 2012). The spatial extent of clearing in mulga is variable, and can range from localised and spatially patchy impacts, such as clearing in ‘windrows’ or strips, through to the broadscale clearing of more extensive areas. In more productive regions, some mulga-dominated woodlands have been cleared and oversown with the exotic perennial grass, buffel grass (Cenchrus ciliaris L.), with a treeless state being maintained through repeated clearing to remove seedlings (Fensham et al. 2012). For HIR projects, cessation of clearing is an additional HIR activity (Macintosh et al. 2024a).
Regarding other disturbances, mulga is generally resistant to fire, owing to the low connectivity of ground cover, and although mulga is capable of re-sprouting vegetatively following low-intensity fire, it is readily killed by high-intensity fire. Widespread fire therefore occurs only after significant rainfall and the subsequent curing of the resulting rain-promoted herbaceous understorey; however, because mass seeding is triggered by rainfall, this also provides the capacity and the mechanism for subsequent community recovery (Wright and Zuur 2014; Wright and Fensham 2017). Silcock et al. (2017) also noted the importance of drought-induced mortality in eastern Australia mulga communities, with mortality being up to 20% following the millennium drought at the start of this century.
Australian laws regulating ACCUs (AG 2020) require that the methodology for estimation of sequestration of biomass carbon in HIR projects be based on the Full Carbon Accounting Model (FullCAM, Richards and Brack 2004). This model is applied at the continental-scale to estimate net greenhouse-gas emissions from the land sector as part of Australia’s National Inventory Report (AG 2024). FullCAM simulates carbon in live biomass, standing dead, fallen debris, soil, and any carbon dioxide (CO2)-C emitted from decomposition and fires (Forrester et al. 2024). The amount of carbon available to cycle through these pools is determined by the quantity of CO2-C sequestered from the atmosphere due to growth of trees/shrubs; primarily driven by an empirical tree yield formula (TYF) (Eqn 1). The TYF predicts accumulation of live above-ground woody biomass (AGB) in a stand of trees/shrubs of uniform age, from seed/seedling to maturity (Waterworth et al. 2007). Three versions of FullCAM are legislated for use in HIR projects, depending on the version of the method under which projects are reporting, including the following: the Reforestation Modelling Tool (RMT, ver. 1.0.1.16.815); the 2016 public release version of FullCAM (ver. 4.1.6.19417); and the current 2020 public release version of FullCAM (ver. 6.20.03.0827). Changes in woody biomass for all three versions use the TYF, but differ in underlying parameters, and hence model predictions and confidence in these (e.g. Roxburgh and Paul 2019). Although only approximately 20% of HIR projects use the 2020 FullCAM version, because the HIR method was closed to new projects in September 2023, and because the 2020 version was the first to have a specific growth calibration for regenerating native vegetation, the FullCAM 2020 version is the focus of investigation here. At the time of writing a 2025 beta-release version of FullCAM has been released for consultation, which embeds the same growth calibration parameters for regenerating vegetation as the 2020 version.
The tree yield formula (TYF):
where
ΔAGBLiveRegen = current annual increment in live above-ground biomass attributable to regeneration (Mg DM ha−1 year−1, where DM is the oven-dry mass);
A1, A2 = age (years) in Years 1 and 2 respectively;
M = spatial input layer of maximum AGB in minimally disturbed native vegetation (Mg DM ha−1) (Roxburgh et al. 2019);
FPI/FPIave = spatial input layers (FPI and FPIave) used to calculate the fraction of the forest productivity index (FPI) over the period A1–A2 when expressed relative to the average over the years 1970–2017. The FPI is the sum of key site factors (soil type, fertility and climate) driving growth, regardless of the type of planting or its age (Kesteven et al. 2004). This index acts to modify the growth increment up or down, depending on whether the FPI/FPIave is >1.0 or <1.0 respectively.
k = 2 × G − 1.25, where G = 12.53 years for natural regeneration in land managed for grazing (Paul and Roxburgh 2020).
By using a wide range of datasets collected from across Australian woodlands managed for grazing (N = 573), the TYF was calibrated for woody biomass of native vegetation regenerating post-removal of grazing suppression and/or cessation of clearing, and hence, is applicable to HIR projects (Paul and Roxburgh 2020). This entailed calibration of the G parameter, which is largely attributable to the predicted rate of AGB accumulation towards a maximum of M (Eqn 1). Across the 573 calibration sites, a G of 12.53 years provided negligible bias and maximised model precision (Paul and Roxburgh 2020).
Given the current (and potential future) application of FullCAM for quantifying sequestration in regenerating native vegetation, including mulga, the aim of this study was to explore the level of confidence in application of the current version of the model for regenerating native vegetation. This was achieved through collating datasets to explore the level of confidence in FullCAM in terms of its: (i) accuracy and domain of application over which there is likely to be negligible model bias, particularly with respect to stand age (the age of the trees that regenerated following suppression removal) and presence of pre-existing biomass; (ii) specificity in simulation of management changes; and (iii) comprehensiveness in simulating all pools of woody biomass. Although a number of issues have been raised regarding the implementation and performance of HIR projects (e.g. Macintosh et al. 2022a, 2022b, 2024a, 2024b), we limit discussion here to only those that directly involve FullCAM and its broader use in simulating native forest regeneration.
Accuracy and domain of application
As FullCAM was designed to be applied in Australia at the continental and regional scales (Richards and Brack 2004), calibration of the FullCAM empirical TYF entailed trade-offs in model precision to ensure model utility with respect to availability of input datasets at these scales, yet ensuring that the model was overall unbiased (Paul and Roxburgh 2020). Only moderate efficiencies of TYF prediction are therefore expected. Indeed, when fitting the G parameter during TYF calibration to 573 regenerating woodland sites managed for grazing, bias was effectively zero and efficiency of prediction was 60% (Paul and Roxburgh 2020). When this calibration was verified by using a relatively small dataset of 49 independent sites (many from HIR project areas), bias was found to be negligible (averaging −0.73 Mg DM ha−1) and efficiency of prediction was 42% (Paul and Roxburgh 2022).
Because of imperfect TYF precision, when applying FullCAM to specific locations in mulga woodlands, there are likely to be under- or over-estimates of AGB. To inform overall model performance, it is useful to quantify this uncertainty, and its decrease over time with on-going model improvements. For example, the recent TYF calibrations of Paul and Roxburgh (2020) resulted in an accuracy gain of site-level prediction of 29% in young stands of natural regeneration, and 88% in mature stands (Roxburgh and Paul 2019).
As new datasets have become available since initial TYF verification (Paul and Roxburgh 2022), the first objective of this study was therefore to use an expanded verification dataset to refine estimates of model accuracy when simulating natural regeneration in Australian woodlands managed for grazing.
In Australia’s woodlands, including mulga woodlands, regeneration of live above-ground woody biomass (AGBLiveRegen) often occurs among pre-existing trees and shrubs, providing a pre-existing baseline of live above-ground woody biomass (AGBLiveBase) (Fig. 2; Supplementary Fig. S1), yet FullCAM TYF accounts only for AGBLiveRegen (Eqn 1, Paul and Roxburgh 2020, 2022). Where HIR activities are implemented, land surveys are required to stratify land into assessment cells of a prescribed size, and to exclude areas with forest cover and, therefore, relatively high AGBLiveBase (AG 2019). However, because vegetation cover is spatially variable, remaining eligible assessment cells of land often still have some AGBLiveBase. For example, in a remote-sensing study, Beare and Chambers (2021) showed that prior to commencement of regeneration activity in HIR projects that were predominantly mulga woodlands, 28–35% of the area was sparsely covered with woody vegetation (where stand-level canopy cover was 5–20%) and another 11–13% was densely covered (where stand-level canopy cover was ≥20%).
Composition of total above-ground woody biomass (AGBTotal) into regenerating and baseline components of live and dead pools. Although AGBLiveRegen, AGBLiveBase, AGBDeadRegen and AGBDeadBase components are often common in mulga woodlands and were measured (Supplementary material Part D), FullCAM TYF predicts only accumulation of AGBLiveRegen, with carbon stocks in the other pools of AGB being predicted in response to FullCAM simulation of regular mortality, as well as the history of disturbance events, e.g. fire and clearing (Forrester et al. 2025b).
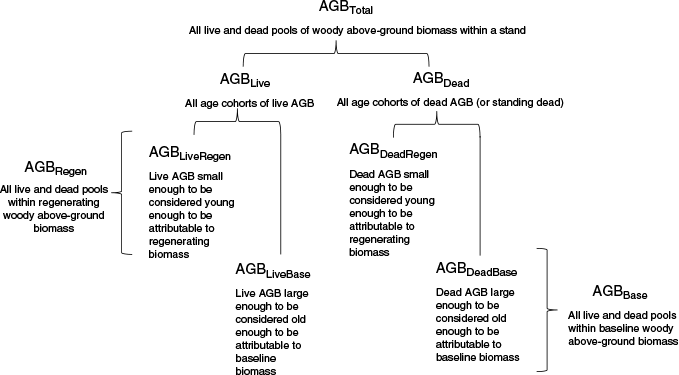
Given that the presence of AGBLiveBase is common in HIR project areas (Beare and Chambers 2021), concerns have been raised that application of FullCAM TYF could lead to over-prediction of AGBLiveRegen (Macintosh et al. 2022a, 2022b). This is because in theory, presence of AGBLiveBase has the potential to limit the growth of AGBLiveRegen relative to that predicted by FullCAM, because the TYF was formulated as an empirical yield curve predicting growth of AGBLiveRegen from negligible levels to a maximum (M) that the site can support on the basis of available resources (Eqn 1). Therefore, the TYF is effectively ‘blind’ to the presence of AGBLiveBase, and, consequently, ‘blind’ to the fact that AGBLiveRegen may be limited by (a) only a proportion of the maximum biomass potential being available, because AGBLiveBase already accounts for a proportion of the site carrying capacity, M, and (b) possible competition with AGBLiveBase for water, nutrients and light (Supplementary material Part A).
This raises two questions regarding the appropriate domain of application to minimise risks of TYF over-prediction in HIR projects. First, should application be restricted to sites of relatively low AGBLiveBase, given as AGBLiveBase increases, then the likelihood of AGBLiveRegen being suppressed by pre-existing AGBLiveBase also increases, leading to actual AGBLiveRegen not attaining TYF-predicted AGBLiveRegen? Second, should application be restricted to sites of relatively young age, given that as stand age increases, the likelihood that actual AGBLiveRegen is less than TYF-predicted AGBLiveRegen increases, with early AGBLiveRegen assumed to suffer less competition with AGBLiveBase, but with suppression of growth increasing over time? Another complicating factor is that the extent of over-prediction may also depend on the interaction of age and amount of AGBLiveBase, with an increasing risk of over-prediction with an increasing stand age, and with an increasing AGBLiveBase as a proportion of M (Supplementary material Part A).
To address these questions, we assessed the importance of these factors by using observations of AGBLiveRegen from across both calibration and expanded verification datasets to (a) assess whether model bias increases with the amount of AGBLiveBase or stand age, and (b) provide a recommended domain of TYF application, with respect to amounts of pre-existing biomass (AGBLiveBase) and stand age.
Specificity: approach used to simulate impacts of management change
The premise of FullCAM application in HIR projects is that AGBLiveRegen increases in accordance with the TYF, following removal of suppression owing to grazing and/or a cessation of clearing (Paul and Roxburgh 2020). It is therefore of paramount importance to verify that FullCAM can accurately simulate changes in woody biomass in regenerating mulga woodlands.
The TYF calibration dataset (N = 573) used by Paul and Roxburgh (2020) for the FullCAM 2020 release did not include explicit data on how grazing management was modified to promote regeneration of woody biomass at each of the calibration sites. Moreover, such data are not available at the continental or regional scales over which FullCAM was designed to simulate (Richards and Brack 2004). This was also the case with the earlier FullCAM releases, although for those versions only generic growth parameters (not specifically calibrated to regenerating vegetation) were applied (e.g. Roxburgh and Paul 2019). Therefore, to facilitate model utility, simulations in FullCAM do not explicitly represent changes in grazing regime, and the TYF calibration for natural regeneration was generalised with respect to changed grazing management (Paul and Roxburgh 2020). The only option currently available in FullCAM to allow temporal variability in grazing pressure on woody biomass is for the user to apply a ‘growth pause’; a nominated period over which TYF-predicted growth increments are assumed to be zero (AG 2020). However, application of such ‘growth pauses’ remains unvalidated against field data (Supplementary material Part B).
There may be variation in rates of regeneration post-removal of grazing pressure on the basis of two factors. The first is sensitivity of AGB to grazing. In an assessment of AGB regeneration in response to removal of grazing in different types of woodlands across Australia, Forrester et al. (2025a) found very mixed responses, but with the highest positive responses to grazing reduction found in acacia woodlands, which were predominantly mulga woodlands and, hence, woodlands containing relatively large amounts of the fodder tree mulga (e.g. Everist 1969). The second factor is the extent of change in grazing management and, thus, the magnitude to which suppression of regeneration is removed. This may vary with the type and quantity of livestock and feral animals present, as well as quantity of feed available, which in turn will be affected by a combination of past climate and fire events that directly or indirectly affect woody vegetation (Paul et al. 2016). For example, there can be increased tree/shrub mortality because of reduced capture of overland flow of water when excessive grazing has removed perennial grass cover (e.g. Anderson and Hodgkinson 1997; Munro et al. 2009).
Building on the work of Forrester et al. (2025a), but focusing on mulga woodlands where HIR projects predominate, another objective of this study was to explore implications of the generalisation of the TYF with respect to changed grazing management. This included quantifying how regeneration post-removal of grazing suppression is influenced by factors not yet specifically accounted for in the TYF, namely (a) sensitivity of the woody vegetation to browsing pressure, and (b) magnitude to which this suppression is removed.
Comprehensiveness: inclusion of standing dead biomass pools
Previous work has indicated that there are large amounts of AGBDead and associated coarse woody debris (CWD) in grazed woodland regions of Australia (e.g. Tongway et al. 1989; Witt et al. 2011; Macdonald et al. 2015), and that these biomass pools can persist in the landscape for decades (Sinclair 2004). Therefore, recent advances in the capability of FullCAM have included separating the dead pools of woody biomass into CWD, and dead material residing in standing senesced individuals (AGBDead, Fig. 2, Supplementary Fig. S11), leading to improved accuracy of prediction of post-disturbance carbon cycling in response to fires and clearing (Paul and Roxburgh 2024; Forrester et al. 2025b).
FullCAM-predicted carbon cycling in AGBDead pools has only recently been calibrated (Paul and Roxburgh 2024; Forrester et al. 2025b). A final objective was therefore to explore merit in expanding the comprehensiveness of FullCAM by accounting for AGBDead through implementation of the latest version of FullCAM and its calibrations. This requires quantification of typical amounts of AGBDead in mulga woodlands, and assessing whether this is significantly influenced by factors such as management (e.g. differences between cleared and uncleared sites), or stand age.
Methods
Accuracy and domain of application
Building on previous independent verification of FullCAM-predicted woody biomass in regenerating mulga woodlands (Paul and Roxburgh 2022; N = 41 sites), additional verification sites (defined as an area of common management history and vegetation and soil type) were sourced to more comprehensively represent mulga woodlands (N = 102 sites, Fig. 3, Table S1). All components of AGB were measured at each of these sites (Fig. 2, Supplementary material Part D). For 33 of these sites (with a subset of these (N = 8) also being paired with mature vegetation sites), trees/shrubs were sampled from an area of 100 ha by using an average (±s.d.) of 16 ± 7 (range 5–35) transects of 100 m by 5 m, placed randomly (location and direction) (Fig. S6). In the remaining 69 sites, all trees/shrubs were measured in (a) 61 exclosures listed in Table 1 (together with additional mature vegetation sites paired with a subset of these (N = 3)), with plot areas ranging from 0.06 to 1.00 ha depending on the experiment, and (b) eight sites with 90 m by 90 m plots, or areas of 0.81 ha, described below in the section 'Specificity: approach used to simulate impacts of management change'.
Location of the sites (Tables 1–3, S1) measured for live above-ground biomass (AGBLiveRegen and AGBLiveBase) and standing dead biomass (AGBDead/AGBTotal) of woody vegetation within the mulga regions of south-western Queensland and western New South Wales, and identification of where these sites are in relation to acacia woodland (grey shading, described by Pasut et al. 2025) and HIR project areas (red shading).

| Property | Location coordinates | Age (years) | Plot size (ha) | N | N | N | N | Source | |
|---|---|---|---|---|---|---|---|---|---|
| SFull-RFull | SFull-RPartial | SFull-RPartial | Open-Grazed | ||||||
| 16 (Qld)A | −25.94, 145.72 | 58 | 0.22 | 1 | 0 | 0 | 0 | B. Witt, pers. comm. | |
| 17–1983 (Qld) | −27.60, 145.80 | 39 | 0.22 | 1 | 0 | 0 | 1 | J. Mills, pers. comm.. | |
| 13 (Qld) | −26.74, 145.09 | 38 | 0.25 | 1 | 0 | 0 | 2 | J. Mills, pers. comm. | |
| Croxdale (Qld)B | −26.46, 146.13 | 41 | 0.25 | 1 | 0 | 1 | 1 | J. Mills, pers. comm. | |
| 18–1996 (Qld) | −27.66, 145.77 | 26 | 0.30 | 1 | 0 | 0 | 1 | K. Hodgkinson, pers. comm. | |
| 7 (Qld) | −27.85, 143.99 | 28 | 0.30 | 1 | 0 | 1 | 2 | K. Hodgkinson, pers. comm. | |
| 6 (Qld) | −26.43, 146.82 | 27 | 0.33 | 1 | 0 | 1 | 1 | K. Hodgkinson, pers. comm. | |
| 14 (Qld) | −26.85, 143.94 | 28 | 0.31 | 1 | 0 | 1 | 1 | K. Hodgkinson, pers. comm. | |
| 5 (Qld) | −27.91, 146.04 | 28 | 0.33 | 1 | 0 | 1 | 1 | K. Hodgkinson, pers. comm. | |
| Glenvue (NSW) | −30.98, 145.97 | 27 | 0.30 | 1 | 0 | 1 | 1 | K. Hodgkinson, pers. comm. | |
| 19-K (NSW) | −29.26, 145.32 | 27 | 0.30 | 1 | 0 | 1 | 1 | K. Hodgkinson, pers. comm. | |
| 24-1 (NSW)C | −30.95, 146.46 | 48 | 0.90 | 1 | 0 | 0 | 1 | K. Hodgkinson, pers. comm. | |
| 24-2 (NSW)C | −30.95, 146.46 | 48 | 0.77 | 1 | 0 | 0 | 1 | K. Hodgkinson, pers. comm. | |
| 15–East (NSW) | −30.96, 145.10 | 45 | 0.25 | 1 | 0 | 0 | 1 | K. Hodgkinson, pers. comm. | |
| 15–West (NSW) | −30.96, 145.10 | 45 | 0.25 | 1 | 0 | 0 | 1 | K. Hodgkinson, pers. comm. | |
| 19-1 (NSW) | −29.27, 145.44 | 32 | 0.15 | 1 | 0 | 0 | 1 | D. Eldridge, pers. comm. | |
| 19-1P (NSW)D | −29.27, 145.44 | 32 | 0.47 | 1 | 0 | 0 | 1 | D. Eldridge, pers. comm. | |
| 19-2 (NSW) | −29.27, 145.45 | 32 | 1.00 | 1 | 0 | 0 | 1 | D. Eldridge, pers. comm. | |
| 19-2P (NSW)D | −29.27, 145.45 | 32 | 0.25 | 1 | 0 | 0 | 1 | D. Eldridge, pers. comm. | |
| 19-3 (NSW) | −29.25, 145.48 | 32 | 0.53 | 1 | 0 | 0 | 1 | D. Eldridge, pers. comm. | |
| 19-3P (NSW)D | −29.25, 145.48 | 32 | 0.48 | 1 | 0 | 0 | 1 | D. Eldridge, pers. comm. | |
| Goorninya East (Qld) | −28.66, 144.37 | 8 | 0.06 | 0 | 1 | 1 | 2 | G. Carroll, J. Silcock, pers. comm. | |
| Goorninya West (Qld) | −28.61, 144.30 | 8 | 0.06 | 0 | 1 | 1 | 2 | G. Carroll, J. Silcock, pers. comm. | |
| Kyabra (Qld) | −28.62, 144.40 | 8 | 0.06 | 0 | 1 | 1 | 2 | G. Carroll, J. Silcock, pers. comm. | |
| Tolee (Qld) | −28.53, 144.24 | 8 | 0.06 | 0 | 1 | 1 | 2 | G. Carroll, J. Silcock, pers. comm. | |
| Paradise (Qld) | −28.66, 144.65 | 8 | 0.06 | 0 | 1 | 1 | 2 | G. Carroll, J. Silcock, pers. comm. | |
| Tareen (Qld) | −28.56, 144.32 | 8 | 0.06 | 0 | 1 | 1 | 2 | G. Carroll, J. Silcock, pers. comm. | |
| Werai (Qld) | −28.66, 144.90 | 29 | 0.16 | 0 | 1 | 0 | 1 | M. Page, pers. comm. | |
| Boorara-1 (Qld) | −28.67, 144.28 | 29 | 0.08 | 0 | 1 | 0 | 1 | M. Page, pers. comm. | |
| Boorara-2 (Qld) | −28.69, 144.28 | 29 | 0.04 | 0 | 1 | 0 | 1 | M. Page, pers. comm. | |
| Boorara-3 (Qld) | −28.69, 144.23 | 29 | 0.04 | 0 | 1 | 0 | 1 | M. Page, pers. comm. | |
| 27 (NSW)E | −31.77, 145.56 | 72 | 0.30 | 1 | 0 | 0 | 1 | S. Ogill, pers. comm. | |
| Monamby (QLD) | −26.64, 145.38 | 58 | 0.16 | 21G | 0 | 0 | 2 | I. Beale, S. Bray, pers. comm. | |
| Boatmans (QLD) | −27.24, 146.98 | 59 | 0.45F | 21G | 0 | 0 | 1 | I. Beale, S. Bray, pers. comm. |
The number of plots (N) in each experiment varied, but each experiment generally had a ‘control’ or 'open-grazed' plot and one or more exclosure plots, including: SFull-RFull, exclosure with meshed wire at the base of a fence at least 1.5–2.0 m high, thereby excluding macropods as well as livestock; SPartial-RFull, exclosure with meshed wire at the base of a fence at least 1.5–2.0 m high, but with livestock being removed from the property only 2 years after the experiment was established, and; SFull-RPartial, exclosure with a wire fence of about 1.1 m high, thereby excluding only livestock.
| Management | Code | N | |
|---|---|---|---|
| Supressed (S) regeneration from high-intensity grazing, followed by regeneration (R) promoted through management of total grazing pressure: S-R | S-R | 84 (2.3) | |
| Mechanical and/or chemical clearing (C) followed by 3–20 years of supressed (S) regeneration owing to grazing, and then regeneration (R) promoted through management of total grazing pressure: C-S-R | C-S-RA | 24 (3.3) | |
| Mechanical and/or chemical clearing (C) followed by supressed (S) regeneration from high-intensity grazing: C-S | C-SB | 4 (2.0) | |
| Mechanical and/or chemical clearing (C) followed by regeneration (R) promoted through management of total grazing pressure grazing: C-R | C-R | 14 (14.0) |
Values in parentheses were the numbers of sites where default M was replaced with a higher M′ taken from an observed AGBLive measured in adjacent undisturbed vegetation (first number in parentheses), or within the site itself (second number in parentheses).
| Property | Coordinates of approximate location | N | FCGB (m2 m−2) | AGBTotal (Mg DM ha−1) | |
|---|---|---|---|---|---|
| 0.81 ha plots | |||||
| QLD001 | −23.11, 143.89 | 3 | 0.11–0.36 | 4.35–13.7 | |
| QLD004A | −25.77, 145.31 | 3 | 0.14–0.18 | 3.33–10.8 | |
| NSW003 | −31.88, 100.86 | 4 | 0.22–0.80 | 21.6–55.9 | |
| NSW004 | −29.22, 145.13 | 1 | 0.60 | 60.2 | |
| QLD012 | −27.73, 143.78 | 6 | 0.02–0.24 | 2.98–46.0 | |
| QLD014A | −27.43, 146.37 | 3 | 0.29–0.65 | 19.5–34.8 | |
| Total | 20 | 0.02–0.80 | 0.83–60.2 |
Each of the 102 sites were categorised into four different groups based on management regimes considered (S-R, C-S-R, C-S and C-R), with this categorisation and abbreviations being described in Table 2. The 24 sites categorised as C-S-R comprised two separate cohorts of regeneration, thereby requiring two separate calculations of AGBLiveRegen based on the assumed stand age, including only the (i) older initial cohort of AGBLiveRegen post-clearing (C → S-R), when categorised as C-S-R, and (ii) younger cohort of AGBLiveRegen that commenced later post-removal of suppression (C-S → R), when categorised S-R.
It was important to separate the AGBLiveRegen component of AGBLive, given this is what FullCAM TYF predicts when simulating the yield of regenerating woody biomass (Eqn 1). Methods used to separate ‘observed’ AGBLiveRegen for locations with existing AGBLiveBase have been outlined previously (Paul and Roxburgh 2020), and are briefly summarised in Supplementary material Part D. In short, pre-existing biomass that had established prior to the regenerating cohort was identified statistically through analysis of the diameter distribution of individuals within the site, and those individuals greater than a specified limit (which varied by plant functional type and stand age), were classified as belonging to AGBLiveBase.
Of the sites used in this analysis (Table 2), 19% were paired with adjacent ‘M’ sites (additional sites measured but not listed in Table 2) that contained mature and relatively undisturbed vegetation (Fig. S7). A further 8% of sites had an observed AGBLive of >M, indicating that M was an underestimate for those sites. Hence, when applying the TYF to sites listed in Table 2, two alternative sets of predictions were obtained, including (i) uninformed prediction, where M applied was the default value for location of the site based on the spatial input layer (Roxburgh et al. 2019), and (ii) where possible, a more accurate data-constrained prediction. This data-constrained estimate of M (notated as M′) was generally based on AGBLive observed from adjacent plots. However, at sites where observed AGBLive was greater than the default M, the M′was taken to be equal to observed AGBLive to curtail the under-prediction that would have resulted had default M been applied.
Predicted AGBLiveRegen was obtained for each site by applying FullCAM (2020 public release, Australian Government 2020) with the year of regeneration being the year of removal of suppression from grazing (S, in S-R sites) or the year post-cessation of clearing (C, in C-S-R, C-S or C-R sites), and with the initial AGBLiveRegen assumed to be 2.0 Mg DM ha−1, consistent with Paul and Roxburgh (2020). Across all the sites (Table 2), FullCAM-predicted that AGBLiveRegen at the age at which the site was measured was compared with that observed, and average bias and the efficiency of prediction (Soares et al. 1995) were calculated, where bias of an individual site was calculated as [predicted AGBLiveRegen] − [observed AGBLiveRegen], with negative values indicating that the FullCAM-predicted AGB was less than observed. This was undertaken using both the uninformed AGBLiveRegen prediction, and the data-constrained AGBLiveRegen prediction (to focus on verification of G, with M′ applied).
To test the risk of AGBLiveRegen over-prediction when applying the TYF to sites with AGBLiveBase (Macintosh et al. 2022a, 2022b), bias was plotted against AGBLiveBase and AGBLiveBase/M, as well as the age of the cohort of regeneration being modelled. This was undertaken for the new datasets collected for model verification (Table 2, N = 102), together with datasets originally collated for model calibration (Paul and Roxburgh 2020, N = 573). To aid presentation of results across sites of differing AGBLiveRegen, bias was expressed as log predicted AGBLiveRegen minus log observed AGBLiveRegen. Results were complemented with the additional analyses outlined in Supplementary material Part A, investigating in greater detail the range of values of AGBLiveBase and stand age across the model calibration dataset of Paul and Roxburgh (2020), and the sensitivity of the FullCAM-calibrated G parameter to AGBLiveBase. When taken together, these analyses were used to provide a recommended temporal and spatial domain of TYF application.
Specificity: approach used to simulate impacts of management change
We analysed datasets collected from 34 long-term (average age 32 years, ±standard deviation of 16 years) grazing exclosure experiments from mulga woodlands (Table 1). These were drawn from a subset of the wider dataset collected by Forrester et al. (2025a) from across all areas of Australian woodlands. Because of a paucity of exclosure experiments in westerns parts of Australia’s woodlands (Forrester et al. 2025a), our analysis was limited to mulga woodlands from the eastern parts of Australia (Fig. 3). Although such experiments can provide valuable insights into the impacts of grazing, it is important to note at any given site the difference in response between exclosure plots and their surrounding land area reflects changes that are a consequence of both grazing pressure per se, in addition to clearing and other activities undertaken as part of broader grazing management.
At all mulga woodlands within which these 34 long-term experiments were located (Table 1), there were macropods, and possibly also feral animals (predominantly goats and rabbits), present over some periods of time. In the older experiments, the predominant livestock was sheep (sites 13, 17, 18, 19, 24, Croxdale), or a mix of sheep and cattle (Werai, Boorara). Cattle have been replacing sheep in this region over the past six decades (e.g. Bowen et al. 2022). Therefore, predominant livestock were cattle in the younger experiments, and in the latter years of the older experiments, with the exception being Sites 17–19, where sheep grazing continued.
Exclosure plots were generally paired with adjacent grazed areas to establish open-grazed or ‘control’ plots of similar characteristics, including size, soil type, landscape position, and previous management. The only exceptions were (a) Site 16, where, unlike the exclosure plot, open-grazed areas were recently cleared of woody vegetation by the land manager, and (b) the Monamby and Boatmans sites, where, unlike the exclosure plots, open-grazed plots were not thinned of woody vegetation at the commencement of the experiment (Table 1).
Exclosure experiments varied in terms of both number of replicate plots and type of exclosures established (Table 1). At all experiments, differences in AGBLiveRegen observed in the exclosure and adjacent open-grazed plots were used to estimate Regeneration post-removal of grazing Suppression (S-R), but with the extent of change in grazing management varying. Exclosures were caterogised into those with:
meshed wire (at least at the base) fence at least 1.5–2.0 m high (Fig. S9), assumed to remove suppression owing to grazing by goats, macropods and rabbits as well as livestock, thereby facilitating the full potential for AGB regeneration, SFull-RFull (N = 22); open-grazed plots were continuously grazed throughout the experiment, by a combination of domestic, native and non-native (feral) animals;
fencing similar to (i), therefore having full potential for AGB regeneration, but with adjacent open-grazed plots having only partial suppression because livestock (but neither macropods nor feral animals) were removed from the property 2 years after the experiment commenced, SPartial-RFull (N = 10), and;
wire fence of about 1.1 m high (Fig. S9), assumed to exclude suppression from livestock grazing only, thereby also facilitating only partial AGB regeneration with respect to the potential, SFull-RPartial (N = 14). Open-grazed plots were continuously grazed throughout the experiment, by a combination of domestic, native and non-native (feral) animals.
Exclosure and open-grazed plots were each measured for components of AGB (Fig. 2), as described in detail by Forrester et al. (2025a), with a brief description of methods being provided in Supplementary material Part D. It was assumed that at the time the experiment was established, AGB and its components were the same across areas of land containing the exclosure and its associated open-grazed plot. For each exclosure plot within each experiment, an estimate of the regeneration attributable to the removal of grazing suppression (ΔAGBRegen) was calculated as follows:
where
ΔAGBRegen = regeneration of live and dead AGB (Mg DM ha−1 year−1),
(AGBRegen)Exclosure = AGBLiveRegen and AGBDeadRegen pools in the exclosure plot,
(AGBRegen)Open-grazed = AGBLiveRegen and AGBDeadRegen pools in the open-grazed plot, and
Age = age of the experiment, or years since the exclosure fence was erected (years).
Because of the decadal timescales being considered, and uncertainty over past management and disturbance histories at each site, no attempt was made to model sequestration directly. However, to explore implications of applying the TYF, generalised for predicting natural regeneration following changes in grazing management, two types of analyses were undertaken. First, to assess the impact of sensitivity of vegetation to grazing, we tested whether there was a statistically significant relationship between ΔAGBRegen and average proportion of total AGB attributable to mulga in exclosure experiments. Anecdotal observations made during field assessments provided further commentary on how the impact of grazing management on ΔAGBRegen may also depend on height of mulga relative to the height of browsing.
Second, to assess the impact of magnitude of grazing suppression removal on ΔAGBRegen, average ΔAGBRegen was compared across datasets (Table 1) from (a) the three contrasting categories of exclosures where magnitude of this removed suppression differed, and (b) SFull-RFull or SPartial-RFull exclosures and the adjacent SFull-RPartial exclosure observed at the 14 experiments that had more than one type of exclosure plot.
Comprehensiveness: inclusion of standing dead biomass pools
To assess the potential to improve the comprehensiveness of FullCAM by expanding the pools of biomass to include AGBDead (e.g. ‘standing dead’, Fig. S11), an indication of the relative importance of this pool was assessed by calculation of AGBDead/AGBTotal for 151 study sites (Tables 1–3). To make up these 151 study sites, an additional 20 sites (or plots) were added (Table 3) to those sites already described above (Tables 1, 2). These additional sites (Table 3) represented 19% of the total acacia-dominant vegetation dataset described by Pasut et al. (2025) and were selected for further study here as they were defined as mulga woodlands; being located from within the mulga region studied (Fig. 1), and for which the predominant acacia species was Acacia aneura. Statistical analysis (ANOVA or linear regression) of these data explored whether AGBDead/AGBTotal is influenced by management (e.g. differed between cleared and uncleared sites), or stand age.
Results and discussion
Accuracy and domain of application
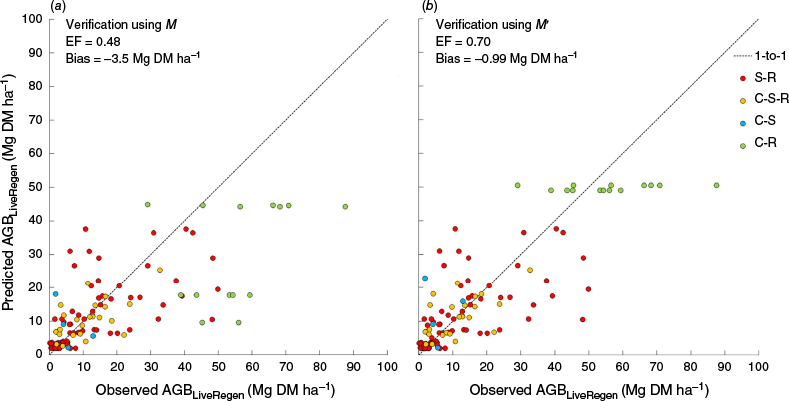
When testing FullCAM TYF-predicted AGBLiveRegen by using the 102 independent verification sites, bias was found to average −3.5 Mg DM ha−1 when applying M (Fig. 4a), and −0.99 Mg DM ha−1 when applying a data-informed estimate of M, M′ (Fig. 4b). Because minimising overall bias is paramount for a model applied at continental (or regional) scales, this provides confidence in application of FullCAM across these scales (Paul and Roxburgh 2020). Model efficiencies of TYF-predicted AGBLiveRegen across the 102 independent verification sites was 48% when applying M (Fig. 4a), and 70% when applying a data-informed estimate of M, M′ (Fig. 4b). Finding model efficiencies in the order of 48–70% was consistent with the model efficiency of 60% attained across the 573 calibration datasets (Paul and Roxburgh 2020).
The relationship between observed and predicted AGBLiveRegen in S-R, C-S-R, C-S and C-R stands listed in Tables 1, S1, when applying (a) M and (b) M′. The number of regenerating sites shown was 126, with 24 of these sites being represented twice because they were included in both S-R and C-S-R.
These results provided confidence that when aggregated over multiple stands of regeneration across the study area (Fig. 3), there is minimal risk of bias, i.e. a divergence between modelled and actual rates of regeneration of woody biomass, providing the model is applied consistently to the way in which it was calibrated, with an assumed growth trajectory towards M that remains unaffected by competitive suppression from any pre-existing AGBLiveBase. A caveat is that further work is required to verify TYF predictions of AGBLiveRegen for regions where HIR projects are common, but were outside our study region, e.g. mulga woodlands of western and central Australia (Fig. 1).
The poorer model performance when applying M (cf. M′) indicates that, in the sample studied (Fig. 3), default values of M may be under-estimated. This may be possible because, like the TYF itself, M is a generalised product that aims to provide high accuracy at the continental (or regional) scale but is highly uncertain at any given point location (Roxburgh et al. 2019). Uncertainties were relatively high for mulga woodlands because, of the 5739 sites of undisturbed and mature native vegetation across Australia used to calibrate M (Roxburgh et al. 2019), only 5.7% were from these woodlands. Therefore, further work is also required to decrease the uncertainty in M.
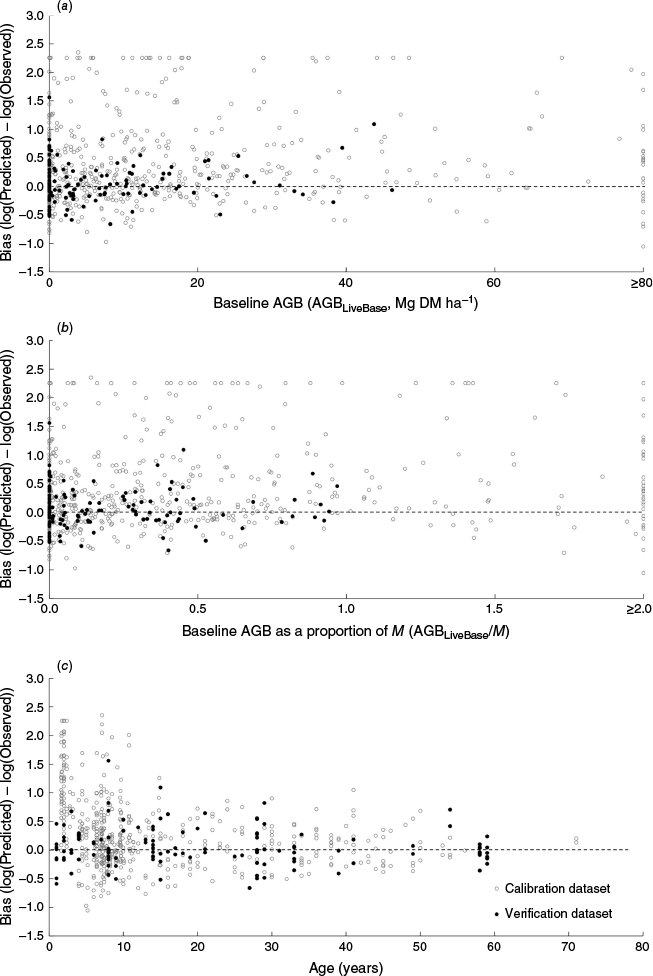
Consistent with earlier findings of Paul and Roxburgh (2022), there was no indication within either the calibration or verification datasets that bias (towards over-prediction) increased with the amount of AGBLiveBase (Fig. 5a, b) or stand age (Fig. 5c). There was also no indication that over-prediction bias was higher in calibration or validation sites where both age and AGBLiveBase were high (Fig. S4), although definitive conclusions cannot be drawn because of the limited domain of the current data (dominated by younger stands; Fig. 6), and the large variability and statistical non-significance of the trends.
Relationships between bias in predicted live regenerating above-ground biomass (AGBLiveRegen) and (a) live baseline above-ground biomass (AGBLiveBase), (b) baseline above-ground biomass as a proportion of M (AGBLiveBase/M), and (c) age of the AGBLiveRegen, for both the calibration (Paul and Roxburgh 2020) and verification (Tables 2, S1) datasets.
Distribution of FullCAM TYF calibration sites (N = 573) with respect to stand age and amount of pre-existing baseline biomass (AGBLiveBase). Red vertical line corresponds with the calibrated G parameter of 12.53 years, and the red horizontal line and grey area show the mean predicted AGB at a canopy cover of 20% and the associated 90% prediction interval, based on data from Pasut et al. (2025). In Australia, vegetation canopy cover in excess of 20% corresponds to the formal definition of forest cover, above which establishment of new HIR projects is ineligible. The prediction interval indicates that 90% of sites at 20% canopy cover are expected to have an AGBLiveBase between 6.8 and 36.2 Mg DM ha−1, with a mean of 17.8 Mg DM ha−1 Note that the prediction interval is asymmetric around the mean on the natural scale because the underlying regression required logarithmic transformation to remove heteroscedasticity.
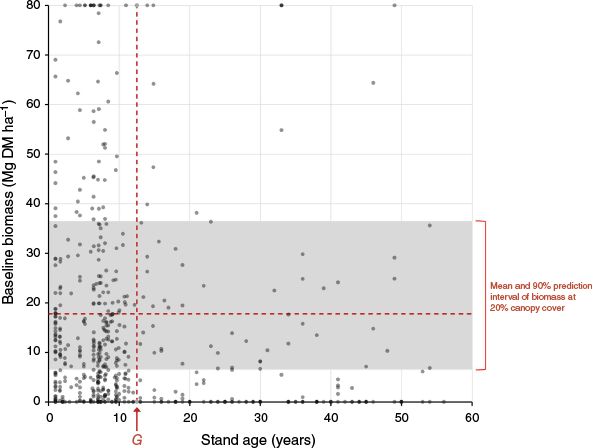
Calibration of the G parameter in the TYF (Eqn 1) was found to be insensitive to the presence of AGBLiveBase within the calibration dataset (Fig. S5), thereby providing confidence in assumptions made by Paul and Roxburgh (2020) when undertaking this calibration that (a) although the AGBLiveBase forms part of M, excluding this baseline for the purposes of calibration did not appear to influence the robustness of the calibration of G, and (b) for the calibration sites used, AGBLiveBase was young enough, and the AGBLiveBase was low enough for additional inter-tree competition for site resources resulting from the presence of AGBLiveBase to have a negligible impact on AGBLiveRegen.
Whilst we consistently found no evidence for over-prediction bias with the calibration and validation datasets available (Figs 5, S4), examination of the data domain of stand AGBLiveBase and stand age over which the model was calibrated (Fig. 6) showed that any extrapolation must be treated with caution, as discussed in the following two sections.
Across the dataset of 573 sites originally used to calibrate the 2020 version of FullCAM (Paul and Roxburgh 2020), 40% of the data were sourced from sites with low levels of AGBLiveBase (i.e. less than 5 Mg DM ha−1, corresponding approximately to the lower limit of data defining the AGBTotal associated with 20% canopy closure based on data from Pasut et al. (2025) (6.8 Mg DM ha−1; Fig. 6); and with 67% of the data sourced from sites with AGBLiveBase less than 15 Mg DM ha−1, corresponding approximately to the mean AGBTotal associated with 20% canopy closure (17.8 Mg DM ha−1; Fig. 6).
Regarding the recommended spatial domain of model application, risk of over-prediction will be lowest when FullCAM's TYF is restricted to sites that have AGBLiveBase that is appreciably less than the biomass expected at 20% canopy cover (Fig. 6). Therefore, to ensure that sites have sufficient potential for regrowth, and to minimise the risk of pre-existing vegetation preventing AGBRegen reaching M, we suggest an upper limit of 5 Mg DM ha−1 for AGBLiveBase. This corresponds to approximately 13% of the average maximum above-ground biomass potential (M) across the 573 calibration sites (Fig. 6). These are conservative recommendations, with analysis suggesting that there is likely to be minimal (although not zero) risk of over-prediction when the model is applied to sites that may have higher AGBLiveBase, but are still below the 20% ‘forest’ cover threshold required to meet HIR new project eligibility (AG 2019) (Figs 6, S4, S5). Moreover, some existing mature trees are required to provide seed stock (Burrows et al. 1988, 1990), and therefore regeneration may require at least a small amount of AGBLiveBase. This is consistent with the procedure used to calibrate the model, which assumed an initial AGB of 2 Mg DM ha−1 (Paul and Roxburgh 2020).
Across the FullCAM calibration dataset, 71% of the data were from sites with AGBLiveRegen of a relatively young age (i.e. less than the calibrated value of G = 12.53 years) (Fig. 6). Regarding the recommended temporal domain of model application, the most conservative approach to limiting the risk of over-prediction would be to restrict FullCAM TYF to sites with regeneration ages less than approximately G, noting that additional analysis suggests application of the model up to 25 years of age is likely to have minimal risk of over-prediction when AGBLiveBase is present. Although TYF application appears sound for prediction of AGBLiveRegen up to 25 years old, these results nevertheless do not preclude the eventual outcome over the longer term, on the basis ecological first principles, that as regenerating stands mature, FullCAM TYF will eventually result in an over-prediction of AGBLiveRegen in the presence of AGBLiveBase (Figs S2, S3). Moreover, for a given amount of AGBLiveBase, the degree of over-prediction will also be influenced by the temporal pattern of competition from AGBLiveBase, which is currently poorly understood (Supplementary material Part A).
Specificity: approach used to simulate impacts of management change
Across the 34 exclosure experiments (comprising 46 plot-level comparisons; Table 1), average (±standard deviation) ΔAGBRegen (Eqn 2) was 0.29 (±0.51) Mg DM ha−1 year−1 (Fig. 7a). These results are consistent with those of other studies (e.g. Moore et al. 2001; Witt et al. 2011; Daryanto et al. 2013) that have shown that removal of grazing suppression in mulga woodlands often (but not always) results in an accumulation of AGB, but with marked among-site variability in the magnitude of the response. High variability in outcomes for soil organic carbon sequestration following change in grazing management have also been found (McDonald et al. 2023; Henry et al. 2024). Although the overall trends are not statistically significant for AGB, and thus no firm conclusions can be validly drawn from the analysis (Muff et al. 2022), further exploration of the same database by Forrester et al. (2025a) did detect statistically significant increases in fine above-ground litter for Acacia aneura following grazing exclusion (Fig. 4 in Forrester et al. 2025a).
Impact of removal of grazing suppression on (a) ΔAGBRegen across all grazing exclosure experiments listed in Table 1, and (b) for the subset of grazing exclosure experiments that had two different types of exclosures (Fig. S10, Table 1), increased ΔAGBRegen with macropod/feral animal exclusion in addition to livestock, calculated as the additional ΔAGBRegen in SFull-RFull or SPartial-RFull exclosures when compared to adjacent SFull-RPartial exclosures within the same experiment. The box and whisker plots indicate the range, together with the 75th, 50th and 25th percentile.
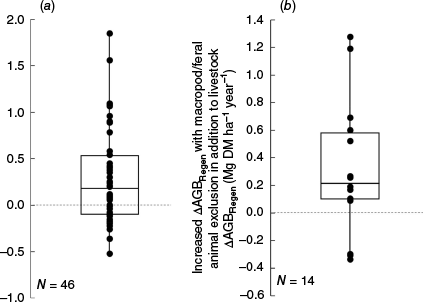
The high variability evident in Fig. 7 is likely to be due to the complexity of the processes that affect woody biomass and its regeneration in rangelands, which include spatial variability in topography and soil type, climate, past disturbance history such as clearing, fire and drought, differences among sites in historic grazing intensity and species composition, and interactions among these factors. In particular, climatic conditions experienced during the exclosure experiment may directly affect the efficacy of changed grazing management in promoting regeneration (e.g. Anderson and Hodgkinson 1997). Therefore, it is possible that the high variability in ΔAGBRegen may be partly explained by whether drought conditions were experienced during the exclosure experiments. It follows that to observe a positive ΔAGBRegen response to grazing removal, exclosure experiments may typically need to be decades old to encompass at least one drought period during which livestock may browse mulga almost exclusively because of poor grass quality (Casburn and Atkinson 2016). It is also possible that some of the exclosure experiments (Table 1) were too young to accurately reflect long-term impacts of grazing on ΔAGBRegen.
Despite the overall non-significant response to grazing (Fig. 7), the potential for mulga to respond to grazing removal was evident in the results obtained from the SFull-RFull subset of exclosures, where all types of grazing (livestock as well as macropods and feral animals) were excluded, and open-grazed plots were continually supressed by grazing livestock. In these exclosures, ΔAGBRegen was found to significantly (P < 0.05) increase with increasing mulga as a proportion of total AGB (R2 = 0.42, N = 22, Fig. 8a). Similar significant (P < 0.05) results were found using ΔAGBTotal (R2 = 0.31, N = 22, data not shown). However, such relationships were not evident when differences in grazing pressure between the exclosure and open-grazed plots were less pronounced (N = 24, Fig. 8b). Overall, in SFull-RFull exclosures, ΔAGBRegen averaged 0.57 ± 0.54 Mg ha−1 year−1 (N = 22, Fig. 8a), whereas in other exclosures (SPartial-RFull or SFull-RPartial), ΔAGBRegen averaged only 0.04 ± 0.31 Mg ha−1 year−1 (N = 24, Fig. 8b). The importance of difference in total grazing pressure influencing ΔAGBRegen was further supported by the finding that ΔAGBRegen averaged 0.32 ± 0.48 Mg ha−1 year−1 higher in exclosures where all grazing was excluded when compared with adjacent exclosures where only livestock grazing was excluded (Fig. 7b, N = 14). This was consistent with earlier studies demonstrating that, like cattle and sheep, overgrazing by macropods, goats and rabbits can also affect AGB of mulga woodlands (Norbury and Norbury 1993; McAlpine et al. 1999; Munro et al. 2009; Mutze 2016).
Observed ΔAGBRegen across experiments with differing average proportions of AGB attributable to mulga for (a) SFull-RFull exclosures, and (b) SPartial-RFull and SFull-RPartial exclosures. Black dashed line indicates the linear regression for SFull-RFull datasets.
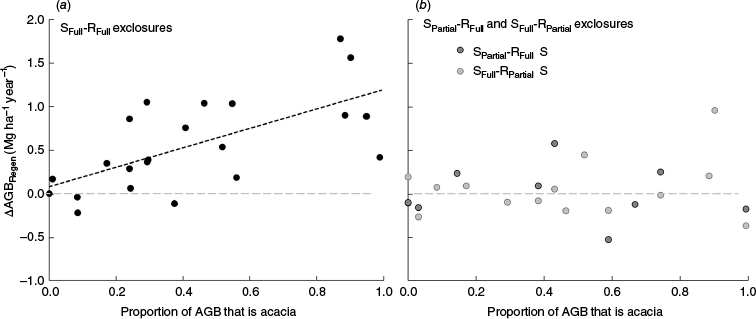
In addition to overall grazing pressure and the amount of mulga within a woodland site, there were some indications that ΔAGBRegen may also be influenced by the predominant height of mulga trees relative to the height of browsing (∼1.2 m; Brown 1985). For example, in open-grazed plots where mulga trees had crowns >1.2 m high owing to being long-term uncleared (Monamby and Boatmans sites), even after almost six decades, AGBTotal remained higher than in adjacent S-R exclosures where mulga had initially been partially cleared (Fig. S10a). Further, at Site 15 where the land owner reported that goats had recently (past ~10 years) breached the exclosure fence, during field assessments of individual tree/shrub biomass there was anecdotal evidence that goats were affecting only small regenerating mulga within this exclosure, with little impact on taller trees from earlier cohorts of regeneration. Further work is required to assess the impacts of mulga tree heights relative to browsing heights more generally.
In summary, our findings (Figs 7, 8) suggest that the extent of regeneration of woody biomass post-change in grazing management depends on the extent of both mulga (and probably also its height relative to height of browsing) and grazing pressure removed (as influenced by type of exclosure fences used, and/or the degree of initial suppression), and the interaction between these, but with additional nuances in response likely to be attributable to factors such as whether there is coincidence with a period of drought. With respect to FullCAM, these factors were not able to be explicitly accounted for in the TYF calibration, with the TYF being generalised with respect to changed grazing management to facilitate model utility (Paul and Roxburgh 2020). Moreover, data on extent of grazing pressure removed at each of the 573 calibration sites were not available, and so increasing TYF site specificity through application of ‘growth pauses’ remained untested (Fig. S8).
Generalising the TYF with respect to changed grazing management is a likely key contributing factor to the large site-to-site variability in TYF bias (Figs 4, 5), with over- or under-prediction of regeneration of woody biomass being anticipated at the site scale. Because changes to grazing is the main activity being implemented in many HIR projects (Macintosh et al. 2024a), increased confidence in prediction of climate-change mitigation attributable to a HIR project may be attained by increasing the specificity with which FullCAM simulates this management change.
Comprehensiveness: inclusion of standing dead biomass pools
Across the 151 sites for which live and dead pools of AGB were measured (Tables 1–3), average (±standard deviation) AGBDead/AGBTotal was 0.17 ± 0.20 (Fig. 9). There was evidence that clearing history significantly (P < 0.05) influenced AGBDead/AGBTotal, averaging 0.16 ± 0.18 in uncleared sites (N = 109), and only 0.11 ± 0.09 in generally younger sites with recent clearing histories (N = 42), and therefore presumably less time to accumulate AGBDead from natural mortality. Across the entire dataset (N = 151) where regeneration was predominantly attributable to removal of grazing suppression rather than just cessation of clearing, there was no statistical evidence that AGBDead/AGBTotal increases with either stand age or site AGBTotal. There are clearly many interacting factors influencing AGBDead/AGBTotal in mulga woodlands, with these likely to be not just clearing and regular mortality, but also drought and fire events (Forrester et al. 2025b).
Histogram of the proportion of sites within the combined datasets (Tables 1–3) that had differing amounts of AGBDead/AGBTotal.
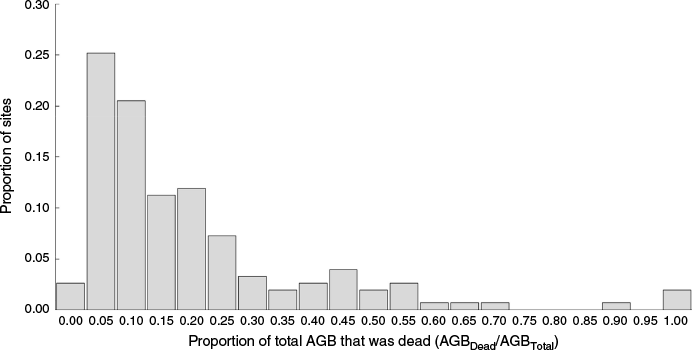
Because AGBDead typically accounts for an average of about 17% of the total on-site woody biomass in mulga woodlands (Fig. 9), there is merit in expanding the pools of biomass considered when applying FullCAM to regenerating mulga, and how AGBDead responds to both regular tree mortality and simulated disturbance events such as clearing, fire, and drought (Paul and Roxburgh 2024; Forrester et al. 2025b).
Conclusions
Confidence in the generalised FullCAM model to predict sequestration of carbon in regeneration of woody biomass is high at continental (or regional) scales, which is of paramount importance for Australia’s National Inventory Reporting, and for ensuring overall integrity of regeneration across a portfolio of projects seeking to promote mulga regeneration. A caveat being further testing is still required for mulga woodlands in central and western Australia where there is currently a paucity of data.
FullCAM is calibrated to ensure broad utility and unbiased predictions at regional-to-continental scales, with the consequence that prediction accuracy can be low at any given location. The generality of the model calibrations has implications in the context of HIR-type project activities. To minimise risks of over-prediction bias at local (or project) scales, restrictions are required to the spatial and temporal domain of application of FullCAM's generalised TYF calibration for natural regeneration. Recommended restrictions include limiting application of the model to areas with pre-existing biomass less than approximately 5 Mg DM ha−1, and limiting the temporal domain to regeneration ages of ≤25 years. Specificity in simulation of changed grazing management is also limited because the generalised TYF does not explicitly account for differing sensitivity of the woody vegetation to browsing (e.g. proportion of woody biomass that was mulga), nor the magnitude of total grazing pressure removal (e.g. exclusion of livestock only, or also macropods and ferals). These limitations on domain of application and specificity of the generalised TYF could be overcome through HIR-type projects undertaking monitoring of biomass post implementation of specific HIR activities, with the biomass measurements obtained then being used to inform project-specific TYF calibrations, including initialisng the model for baseline biomass. These project-specific TYF calibrations could be implemented in a revised version of FullCAM. We also conclude the comprehensiveness of FullCAMcould be improved through the inclusion of standing dead pools of woody biomass, because they are not currently accounted for.
Data availability
Site-level data for model verification are provided in the Supplementary material. Some property-level information including property names and locations has been anonymised due to confidentiality agreements.
Declaration of funding
The work was funded by the Department of Climate Change, Energy, the Environment and Water, Canberra.
Acknowledgements
Mel Piper is thanked for managing the field work planning and logistics. Andrew Ford, Alex Drew, Joel Armstrong, Micah Davis, Tim Hogarty, Matt Bradford, Wendy Cook, Brenton Brooks, Matt Charmley and Branka Culvenor are thanked for their assistance with field work. Konrad Muller, Max Collett, Lyon Young, Jacqui England, Senani Karunaratne and Dan Rosauer are thanked for reviewing an earlier draft of this paper. Research scientists listed in Table 1 as well as Green Collar Group, Climate Friendly, TEM, Bush Heritage Australia, Corporate Carbon, and CO2 Australia are thanked for providing access to properties and/or reserach trial sites, and for providing auxiliary information on management changes implemented to induced regeneration of woody biomass.
References
Australian Government (2018) Carbon Credits (Carbon Farming Initiative) (Human-Induced Regeneration of a Permanent even-aged Native Forests–1.1) Methodology Determination 2013 made under section 106 of the Carbon Credits (Carbon Farming Initiative) Act 2011. Available at https://www.legislation.gov.au/Details/F2018C00125
Australian Government (2019) ‘Guidelines on stratification, evidence and records or projects under the Human-Induced Regeneration of a Permanent Even-Aged Native Forest and Native Forest from Managed Regrowth methods.’ (Clean Energy Regulator, Australian Government (AG): Canberra, ACT, Australia) Available at https://cer.gov.au/node/3783
Australian Government (2020) Requirements for use of the Full Carbon Accounting Model (FullCAM) with the Emissions Reduction Fund (ERF) Methodology Determination: Carbon Credits (Carbon Farming Initiative) (Human Induced Regeneration of a Permanent Even Aged Native Forest—1.1). Methodology Determination 2013. Version 3.0 (published and in force from 1 September 2020) Available at https://www.dcceew.gov.au/sites/default/files/documents/final_fullcam_guideline_human-induced_regeneration_of_a_permanent_even-a.pdf
Australian Government (2024) National Greenhouse Gas Inventory Report: 2014. Australian Government submission to the United Nations Framework Convention on Climate Change and its Kyoto Protocol. Australia’s National Greenhouse Accounts. (Department of Climate Change, Energy, the Environment and Water: Canberra, ACT, Australia) Available at https://www.dcceew.gov.au/climate-change/publications/national-inventory-report-2022
Anderson VJ, Hodgkinson KC (1997) Grass-mediated capture of resource flows and the maintenance of banded mulga in a semi-arid woodland. Australian Journal of Botany 45, 331-342.
| Crossref | Google Scholar |
Baumber A, Waters C, Cross R, Metternicht G, Simpson M (2020) Carbon farming for resilient rangelands: people, paddocks and policy. The Rangeland Journal 42, 293.
| Crossref | Google Scholar |
Beare S, Chambers R (2021) ‘Human induced regeneration: a spatiotemporal study.’ (AnalytEcon: Berry, NSW, Australia) Available at https://www.dcceew.gov.au/sites/default/files/documents/human-induced-regeneration-spatiotemporal-study.pdf
Bowen MK, Chudleigh F, Sallur NM, Sommerfield J (2022) Opportunities to build resilience of beef cattle properties in the mulga lands of south-western Queensland, Australia. The Rangeland Journal 44, 115-128.
| Crossref | Google Scholar |
Brown RF (1985) The growth and survival of young mulga (Acacia aneura F.Muller) trees under different levels of grazing. Australian Rangeland Journal 7, 143-148.
| Crossref | Google Scholar |
Burrows WH, Scanlan JC, Anderson ER (1988) Plant ecological relations in open forests, woodlands and shrublands. In ‘Native pastures in Queensland: the Resources and their Management’. Queensland Department of Primary Industries Information Series Q187023. (Eds WH Burrows, JC Scanlan, MT Rutherford) pp. 72–90. (Queensland Government: Brisbane, Qld, Australia)
Burrows WH, Carter JO, Scanland JC, Andrson ER (1990) Management of savannas for livestock production in north-east Australia: contrast across the tree–grass continuum. Journal of Biogeography 17, 503-512.
| Crossref | Google Scholar |
Casburn G, Atkinson T (2016) ‘Using mulga as a forage supplement for livestock in droughts. PrimeFact 1487, August 2016.’ (NSW Department of Primary Industries, NSW Government) Available at https://www.dpi.nsw.gov.au/__data/assets/pdf_file/0003/671421/using-mulga-as-a-forage-supplement-for-livestock-in-droughts.pdf
Daryanto S, Eldridge DJ, Throop HL (2013) Managing semi-arid woodlands for C storage: grazing and shrub effects on above- and belowground C. Agriculture Ecosystems & Environment 169, 1-11.
| Crossref | Google Scholar |
Feng X, Stafford-Smith M, Zhang L, Lyu N, Lyu Y, Fu B, Xu Z (2020) Global dryland ecosystem programme (Global-DEP): Australasian consultation result. Journal of Soils and Sediments 20, 1807-1810.
| Crossref | Google Scholar |
Fensham RJ, Dwyer JM, Eyre TJ, et al. (2012) The effect of clearing on plant composition in mulga (Acacia aneura) dry forest, Australia. Austral Ecology 37(2), 183-192.
| Crossref | Google Scholar |
Forrester DI, England JR, Paul KI, Rosauer DF, Roxburgh SH (2024) Modelling carbon flows from live biomass to soils using the full Carbon Accounting Model (FullCAM). Environmental Modelling and Software 177, 106064.
| Crossref | Google Scholar |
Forrester DI, England JR, Pasut C, Paul KI, Roxburgh SH, Wang YP (2025b) Calibration of the FullCAM model for Australian native vegetation. Ecological Modelling In press.
| Google Scholar |
Forrester DI, England JR, Ng EL, Piper M, Hodgkinson K, Bray SG, Roxburgh SH, Paul KI (2025a) How does grazing exclusion in Australia’s rangelands impact biomass and debris carbon stocks? The Rangeland Journal 47, RJ24028.
| Crossref | Google Scholar |
Harrington G (1979) The effects of feral goats and sheep on the shrub populations in a semi-arid woodland. The Rangeland Journal 1, 334.
| Crossref | Google Scholar |
Henry B, Allen D, Badgery W, et al. (2024) Soil carbon sequestration in rangelands: a critical review of the impacts of major management strategies. The Rangeland Journal 46(3), RJ24005.
| Crossref | Google Scholar |
Howden SM, Moore JL, McKeon GM, Carter JO (2001) Global change and the mulga woodlands of southwest Queensland: greenhouse gas emissions, impacts, and adaptation. Environment International 27, 161-166.
| Crossref | Google Scholar | PubMed |
Macdonald BCT, Gillen J, Tuomi S, Newport J, Barton PS, Manning AD (2015) Can coarse woody debris be used for carbon storage in open grazed woodlands? Journal of Environmental Quality 44, 1037-1332.
| Crossref | Google Scholar |
Macintosh A, Butler D, Evans MC, Ansell D, Waschka M (2022a) Fixing the Integrity Problems with Australia’s Carbon Market. (Australian National University: Canberra, ACT) Available at https://law.anu.edu.au/files/2023-10/short_-_erf_reform_june_2022_final.pdf
Macintosh A, Butler D, Evans MC, Larraondo PR, Ansell D, Gibbons P (2022b) The ERF’s Human-Induced Regeneration (HIR): what the Beare and Chambers Report Really Found and a Critique of its Method. (Australian National University: Canberra, ACT, Australia) Available at https://law.anu.edu.au/files/2024-01/What%20the%20Beare%20and%20Chambers%20Report%20Really%20Found%20and%20a%20Critique%20of%20its%20Method%20%2816%20March%202022%29.pdf
Macintosh A, Butler D, Larraonda P, Evans MC, Ansell D, Waschka M, Fensham R, Eldridge D, Lindenmayer D, Gibbons P, Summerfield P (2024a) Australian human-induced native forest regeneration carbon offset projects have limited impact on changes in woody vegetation cover and carbon removals. Communications Earth and Environment 5, 149.
| Crossref | Google Scholar |
Macintosh A, Evans MC, Butler D, et al. (2024b) Non-compliance and under-performance in Australian human-induced regeneration projects. The Rangeland Journal 46(5), RJ24024.
| Crossref | Google Scholar |
McAlpine CA, Grigg GC, Mot JJ, Sharma P (1999) Influence of landscape structure on kangaroo abundance in a disturred semi-arid woodland of Queensland. Rangeland Journal 21, 104-134.
| Crossref | Google Scholar |
McDonald SE, Badgery W, Clarendon S, et al. (2023) Grazing management for soil carbon in Australia: a review. Journal of Environmental Management 347, 119146.
| Crossref | Google Scholar | PubMed |
Moore JL, Howden SM, McKeon GM, Carter JO, Scanlan JC (2001) The dynamics of grazed woodlands in southwest Queensland Australia and their effect on greenhouse gas emissions. Environment International 27, 147-153.
| Crossref | Google Scholar | PubMed |
Muff S, Nilson EB, O’Hara RB, Nater CR (2022) Rewriting results in the language of evidence. Trends in Ecology and Evolution 37, 203-210.
| Crossref | Google Scholar | PubMed |
Munro NT, Moseby KE, Read JL (2009) The effects of browsing by feral and re-introduced native herbivores on seedling survivorship in the Australian rangelands. The Rangeland Journal 31, 417-426.
| Crossref | Google Scholar |
Mutze G (2016) Barking up the wrong tree? Are livestock or rabbits the greater threat to rangeland biodiversity in southern Australia? The Rangeland Journal 38, 523-531.
| Crossref | Google Scholar |
Norbury GL, Norbury DC (1993) Distribution of red kangaroos in relation to range regeneration. Rangeland Journal 15, 3-11.
| Crossref | Google Scholar |
Page MJ, Witt GB, Noël MV, Slaughter G, Beeton RJS (2008) Economic and environmental analysis of fodder harvesting practices associated with mulga (Acacia aneura) and fire management practices in the mulga lands of south western Queensland. Final Report to Australian Government, Department of the Environment, Water, Heritage and the Arts. (School of Natural and Rural Systems Management, University of Queensland: Australia)
Pasut C, England JR, Piper M, Roxburgh SH, Paul KI (2025) Aboveground biomass relationship with canopy cover and vegetation structure to improve carbon change monitoring in rangelands. Ecosphere 16(4), e70231.
| Crossref | Google Scholar |
Paul KI, Roxburgh SH (2020) Predicting carbon sequestration of woody biomass following land restoration. Forest Ecology and Management 460, 117838.
| Crossref | Google Scholar |
Paul KI, Roxburgh SH (2022) ‘Verification of FullCAM’s Tree Yield Formula for Regenerating Systems.’ (CSIRO: Australia) 10.25919/wtap-j381
Paul KI, Roxburgh SH (2024) A national accounting framework for fire and carbon dynamics in Australian savannas. International Journal of Wildland Fire 33, WF23104.
| Crossref | Google Scholar |
Richards GP, Brack CL (2004) A continental biomass stock and stock change estimation approach for Australia. Australian Forestry 67, 284-288.
| Crossref | Google Scholar |
Roxburgh SH, Karunaratne SB, Paul KI, Lucas RM, Armston JD, Sun J (2019) A revised above-ground maximum biomass layer for the Australian continent. Forest Ecology and Management 432, 264-275.
| Crossref | Google Scholar |
Silcock JL, Drimer J, Fraser J, Fensham RJ (2017) Inability of fire to control vegetation dynamics in low-productivity mulga (Acacia aneura)-dominated communities of eastern Australia. International Journal of Wildland Fire 26, 896-905.
| Crossref | Google Scholar |
Sinclair R (2004) Persistence of dead trees and fallen timber in the arid zone: 76 years of data from the T.G.B. Osborn Vegetation Reserve, Koonamore, South Australia. The Rangeland Journal 26, 111-122.
| Crossref | Google Scholar |
Soares P, Tome M, Skovsgaard JP, Vanclay JK (1995) Evaluating a growth model for forest management using continuous forest inventory data. Forest Ecology and Management 71, 251-265.
| Crossref | Google Scholar |
Tongway DJ, Ludwig JA, Whitford WG (1989) Mulga log mounds: fertile patches in the semi-arid woodlands of eastern Australia. Australian Journal of Ecology 14, 263-268.
| Crossref | Google Scholar |
Waterworth RM, Richards GP, Brack CL, Evans DMW (2007) A generalised hybrid process-empirical model for predicting plantation forest growth. Forest Ecology and Management 238, 231-243.
| Crossref | Google Scholar |
Witt GB, Noel MV, Bird MI, Beeton RJS, Menzies NW (2011) C sequestration and biodiversity restoration potential of semi-arid mulga lands of Australia interpreted from long-term grazing exclosures. Agriculture Ecosystems and Environment 141, 108-118.
| Crossref | Google Scholar |
Wright BR, Fensham RJ (2017) Fire after a mast year triggers mass recruitment of slender mulga (Acacia aptaneura), a desert shrub with heat-stimulated germination. American Journal of Botany 104, 1474-1483.
| Crossref | Google Scholar | PubMed |
Wright BR, Zuur AF (2014) Seedbank dynamics after masting in mulga (Acacia aptaneura): implications for post-fire regeneration. Journal of Arid Environments 107, 10-17.
| Crossref | Google Scholar |
Yirdaw E, Tigabu M, Monge M (2017) Rehabilitation of degraded dryland ecosystems – review. Silva Fennica 51, sf.1673.
| Crossref | Google Scholar |