

Influence of selected environmental variables on the germination of Eucalyptus costata
Joseph Stapleton A , Callum Measham-Pywell A and Singarayer Florentine A B *
A B *
A
B
Abstract
Eucalyptus costata is a widespread eucalypt species used in restoration works. It is common to the Mallee, a vegetation type in the semiarid temperate region of southern Australia, characterised by hot summers and mild winters, generally reliable but highly variable rainfall, and nutrient poor soils composed primarily of sands. An understanding of the germination biology of Eucalyptus costata may assist in its successful establishment. We studied the impacts of light, temperature, salinity, moisture and sowing depth on the germination of E. costata in the laboratory and considered the results in a restoration context. Germination was lowest under the highest temperature regime (35°C/25°C), high salt concentrations (>100 mM) and low water potentials (taking 14 MPa of humidity per day to germinate). Sowing depths greater than 2 cm reduced seedling emergence. These results suggest that E. costata’s germination requirements are shallow sowing in low salinity soils during periods of cool or mild wet weather, consistent with winter and spring planting.
Keywords: arid conditions, Australian Mallee, burial depth, drought salinity, Eucalyptus, germination, germination temperature, restoration works.
Introduction
Seeds generally have an optimal range of conditions under which germination is maximised (Dalziell et al. 2022). However, germination biology is understudied compared with other seed traits, despite the potentially valuable contributions it can make towards restoration activities aimed at returning native vegetation to degraded landscapes (Jiménez-Alfaro et al. 2016).
Arid ecosystems may particularly benefit from an understanding of germination biology, particularly for their restoration following degrading anthropogenic activities (Williams 1999; Munro et al. 2009; Verhoeven et al. 2020), because the extreme environmental conditions that may be evident including low soil water (Callister and Westbrooke 2017; Huang et al. 2019), extreme temperatures (Houston 2006) and saline soils (Stavi et al. 2021) make restoration particularly challenging.
Plants in arid rangelands have developed a range of distinct germination strategies (Gutterman 1994). For example, many seeds exhibit dormancy where a seed will forgo germination until an environmental trigger, such as exposure to water, is experienced initiating germination (Poschlod et al. 2013). Other species may produce large quantities of seeds that rapidly germinate over a variety of temperatures, increasing the possibility of germination in an environment prone to significant environmental change (Duncan et al. 2019).
The Mallee area of south-eastern Australia is an arid environment that may benefit from such studies. Mallee is characterised by a canopy mostly composed of low-growing multi-stemmed eucalypt (mallee) species growing on sandy soil. The Mallee has been extensively cleared for agriculture since the late 19th century (White 2006), leaving much of the overgrazed Mallee vegetation fragmented and facing issues such as weed competition and erosion, the latter affecting site vegetation even after grazing activities have ceased (Calder 1990; White 2006). In ex-agricultural and grazing areas, efforts are being made to restore these lands and promote biodiversity recovery (Calder 1990).
One key Mallee species is E. costata Behr & F.Muell. (previously E. incrassata), a eucalypt species widespread across the Mallee region of southern Australia (Commonwealth of Australia 1988). Although being a good choice for restoration programmes, there is limited germination biology information on this species. We undertook studies on E. costata with the aim to (i) determine its optimal germination conditions considering light, temperature, soil salinity, drought tolerance and sowing/burial depth; and (ii) outline the management approaches for improving restoration work in the field based on these findings.
Materials and methods
Seed collection and preparation
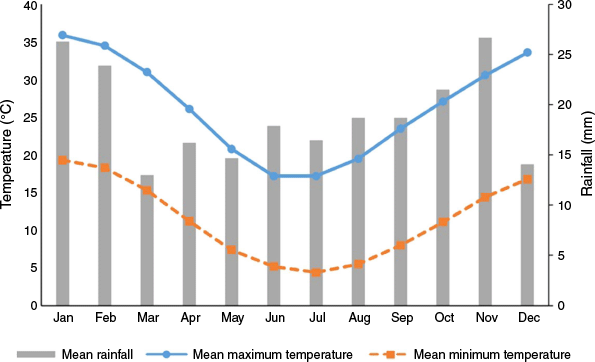
Seed capsules were collected during February 2024 from 16 mature E. costata trees at Federation University Australia’s Nanya Research Station (33.12°09′8.370″S, 141.3°17′30.20″E), in far south-western New South Wales, 200 km south of Broken Hill (Fig. 1). The vegetation primarily consisted of old-growth mallee on sandy soil (Westbrooke 2012). The climate is semiarid, with mean maximum summer temperatures above 30°C and mean winter minima below 10°C (Bureau of Meteorology 2024, accessed from http://www.bom.gov.au/climate/data/). Annual rainfall is typically less than 250 mm, with the highest rainfall usually occurring in late spring to summer, and the lowest in autumn (Fig. 2).
Mean monthly temperature and rainfall data for the region around Nanya Research Station (Bureau of Meteorology 2024, accessed from http://www.bom.gov.au/climate/data/). Rainfall data are from Wentworth (Scotia Sanctuary) station number: 47105, (33.21°S, 141.17°E); and temperature data from Pooncarie Mail Agency, station number: 47029, (33.39°S, 142.57°E).
After collection, seed capsules were placed in labelled bags and transported to the Federation University Seed Ecology laboratory, Mount Helen, within 2 days. The bags were left in the laboratory at room temperature to dry for 18 days, during which the seeds fell out of the capsules into the bag. Seeds remaining within the capsules were shaken free before the mixture of chaff and seeds was passed through a set of sieves with 2 mm mesh to avoid the loss of smaller seeds, and seeds were then separated from the remaining chaff by hand. Mean seed mass (0.72 ± 0.062 g) was determined by weighing a total of 105 seeds from 25 capsules. Prior to germination trials, seeds were surface sterilised by placing them in a sieve and rinsing five times with 1% (v/v) hypochlorite solution and then rinsing with reverse-osmosis (RO) water.
Seed germination trials
For all germination trials, 90 mm × 15.8 mm Petri dishes were used. Whatman No. 11 filter paper was moistened with sterilised RO water or the appropriate treatment solution to ensure adequate moisture for germination. Twenty seeds were placed on the moistened filter paper in a 4 × 5 grid. All dishes were sealed with two layers of parafilm, then placed into an incubator (Thermoline Scientific, temperature and humidity cabinet, Model TRISLH-495-1-s.d., Vol. 240, Sydney, Australia). The incubator was fitted with cool-white, fluorescent lamps that provided a photosynthetic photon flux of 40 mmol m−2 s−1. Seeds were checked three times a week for 41 days, and were considered germinated when the radicle length was at least 2 mm, and the cotyledons had emerged from the seed coat (Ferrari and Parera 2015). There were three replicates with 20 seeds each used in each treatment as described below.
Light and temperature trials
The effects of light and temperature on E. costata germination were determined by exposing seeds to day/night temperatures of 35°C/25°C, 30°C/20°C, 25°C/15°C, and 13°C/5°C under 12/12 h light/dark conditions, and 24 h dark conditions. A double layer of aluminium foil was placed around each Petri dish for the 24 h dark conditions. The 25°C/15°C trial under 12/12 h light/dark conditions, provided the best germination results. Consequently, these light conditions were used for the remainder of the trials described below.
Osmotic stress trials
The effects of water availability on E. costata germination were investigated by wetting the filter paper with pre-mixed aqueous solutions containing polyethylene glycol 8000, giving osmotic potentials of −0.1, −0.2, −0.4, −0.6 and −0.8 MPa.
Salinity trials
Premixed saline solutions containing 25, 50, 100, 150, and 200 mM of NaCl were applied to the filter paper to investigate the effect of substrate salinity on germination
Sowing/burial depth trials
River sand was autoclaved to denature seeds or propagules. Five centimetres of sand was added to 15 black plastic pots (10 cm × 12 cm × 11 cm) lined with paper towels to prevent sand loss. The sand was moistened, and 20 sterilised seeds placed in a 4 × 5 grid on the exposed surface of each pot. Seed planting depth was achieved by covering seeds with sand to depths of 0, 0.5, 1, 2, and 3 cm. The pots were placed on two large plastic trays (36 cm × 43 cm × 8 cm) containing RO water allowing water to permeate by capillary action. Trays were placed in an incubator under 25°C/15°C with 12/12 h light/dark conditions.
Data analysis
Light, temperature, salinity and burial depth analysis
Data analysis was conducted using R (ver. 4.2.3; R Core Team 2024). The Germina R package (ver. 2.1.4, https://cran.r-project.org/web/packages/GerminaR/index.html) constructed by Lozano-Isla et al. (2019) was used to calculate the germination proportion (GRP) referring to the percentage of seeds that complete germination, and mean germination time (MGT) for the seeds under different light, temperature and salinity treatments. For burial depth analysis, the GRP and MGT were also used as a surrogate for emergence. The GRP was calculated using the formula:
MGT refers to the mean time, in days, taken for seeds to germinate, calculated using the following formula:
where ni is the number of seeds germinated at the ith time, k is the last day of evaluation, N is the total number of seeds in each replicate and ti is the time from the beginning of the experiment till the ith observation (Lozano-Isla et al. 2019).
The results of the GRP and MGT calculations were non-normally distributed. Therefore, a Kruskal–Wallis test (Kruskal and Wallis 1952) was used to determine significance. Pairwise comparison were made using a Dunn’s test (Dunn 1964).
Osmotic stress data analysis
Hydro-time-to-event models were created to estimate the germination percentage of seeds at different water potentials using the drcte (ver. 1.0.30, https://cran.r-project.org/web/packages/drcte/index.htm; Onofri 2022) and drcSeedGerm R packages (ver. 1.0.1, https://cran.r-project.org/web/packages/drcSeedGerm/index.html; Onofri 2018). Time-to-event models are a type of mathematical model used to predict the time till an event, such as germination, occurs. For this trial, a type of model, referred to as hydro-time-to-event, were used, which predict the time course of germination under different water potentials. This provides information on the minimum water potential threshold that must be crossed before germination can begin and how long this takes to achieve (Bradford 2002). The analysis was performed by creating multiple hydro-time-to-event models by using different cumulative distribution functions. An Akaike’s information criterion was then used to determine which of these models provided the best fit for the data. It was found that the best-fitting model used a Weibull (Type I) distribution as developed by Mesgaran et al. (2013), which used the following equation:
where P is the proportion of germinated seeds, t is the time in days, is a water potential function, b is the slope, θH is a hydrotime constant (referring to the humidity content in mega Pascals (MPa) per unit time taken for a seed to germinate), and σ is the scale parameter.
This model was then applied to the data and the results were analysed via a graphical display. Information included the base water potential (referring to the predicted lowest water potential at which a seed can germinate), and the hydro-time constant (Bradford 2002).
Results
Response to light and temperature regime
Light regime had no significant effect on either GRP or MGT. Conversely, temperature regime was significant, with significantly fewer seeds germinating at 35°C/25°C than at the 25°C/15°C or 13°C/5°C regimes (Fig. 3a).
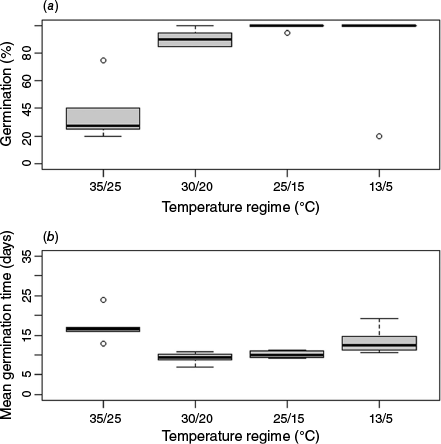
Germination was significantly slower at 35°C/25°C than at 30°C/20°C or 25°C/15°C (Fig. 3b). Germination at 35°C/25°C was not significantly different from that at 13°C/5°C, with germination in the latter being significantly slower than at 30°C/20°C.
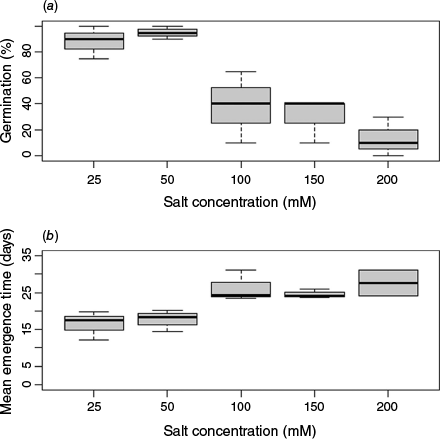
Response to salt concentrations
Salt concentration significantly affected seed GRP. Germination under 25 and 50 mM concentrations was generally higher than at the 100–200 mM range (Fig. 4a). There was a significant effect for salt concentration on MGT, with seeds in the 25–50 mM range germinating faster than those in the 100–200 mM range (Fig. 4b).
Response to sowing/burial depth
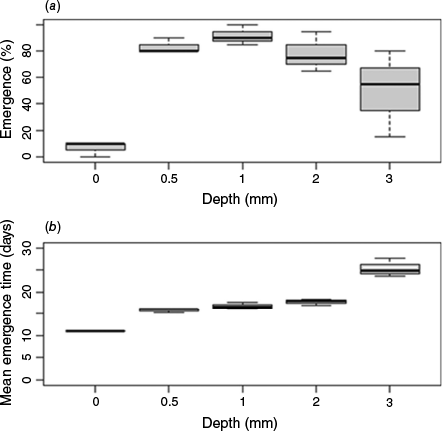
Burial depth significantly affected seed emergence, with emergence of surface sown seeds significantly lower than that of seeds buried at 1 cm depth where overall emergence was highest (Fig. 5a). Burial depth significantly affected MGT, with surface sown seeds emerging faster than seeds buried at 3 cm (Fig. 5b). Seeds at the other burial depths (0.5, 1, 2 cm) had emergence rates between these two extremes, although these rates did not differ significantly.
Response to changes in water potential
At water potentials greater than −0.4 MPa, the number of germinating seedlings declined sharply (Supplementary Fig. S1). The mean base water potential (the threshold above which water potential is sufficient for germination to occur) was about −0.4 MPa, although this result was not significant (P = 0.405). The hydro-time constant was found to be significant (P =6.812 × 10−10) and was calculated to be about 14 MPa day–1 (Table 1).
Discussion
Eucalyptus costata germination was lowest at 35°C/25°C, a response similar to that reported by Wellington (1981). It is possible that this species remains dormant during unfavourable environmental conditions, such as periods of high temperature, potentially as a strategy to avoid germination when hot and dry conditions would increase seedling mortality (Duncan et al. 2019). This dormancy could serve as an adaptive mechanism to enhance survival. However, some ungerminated seeds appeared to have softened, and also in the absence of a seed viability assessment on the ungerminated seeds, it is difficult to confirm whether dormancy or a loss of viability is responsible. As such, further investigation is needed to clarify the underlying strategy. The germination rate was least under both the hottest and coolest temperature regimes, and the slowing of germination under cool temperatures was consistent, with results observed in other eucalypt species (Ruiz-Talonia et al. 2018; Stapleton et al. 2024).
Reported response of eucalypts to changing water potential is inconsistent. The recorded response in this study suggests that E. costata seeds are sensitive to water-limited conditions, which contradicts both Facelli and Ladd (1996) with E. costata, and other studies on semiarid eucalypt species where germination at low water potentials of about −1.0 MPa and hydro-time constants of 5.1 MPa day–1 were recorded (Arnold et al. 2014; Stapleton et al. 2024). Conversely, Emery and Collette (2021), by using several eucalypt species (E. melanophloia, E. pilligaensis, E. populnea subsp. bimbil and E. viridis) from the semiarid brigalow (Acacia harpophylla) region of north-western New South Wales, Australia, found these species to be drought intolerant, with germination being inhibited at water potentials below −0.25 MPa. Given the recorded preference for milder temperatures, it is suggested that E. costata similarly prefers periods of mild wet weather before germination. The provenance region of the seeds in this study is known to experience periods of flooding (Florentine et al. 2015), which may provide opportunities of high soil moisture suitable for germination.
The E. costata response to salinity indicates a preference for germinating in low-salinity soils, a result similar to that of Mechergui et al. (2019) on the eucalypt species they tested. However, given that the E. costata still showed some limited germination under high salinity levels, the species may have some tolerance to saline soils. Such a tolerance would be useful in the Mallee region, given the common occurrence there of highly saline soil (Westbrooke 2012).
Eucalyptus costata germinated well at depths of about 0.5–1 cm, and this preference matches observations by Facelli and Ladd (1996), who showed that E. costata seeds are able to tolerate being covered by leaf litter. However, Ruiz Talonia et al. (2017) found that although the floodplain eucalypt species they tested emerged best under a similar depth of 0.6 cm when the soil was dry, under wet conditions, the seeds preferred to germinate on the soil surface. The preference for shallow burial depths found in eucalypt species is likely to be due to the small size of eucalypt seeds, because smaller seeds tend to have smaller nutritional stores that limit potential emergence depths (Grant et al. 1996; Bond et al. 1999).
In its natural setting, E. costata appears most likely to germinate after or during high-rainfall periods at milder times of the year such as spring and winter. This result is consistent with field studies, which found that E. costata seeds can germinate in winter (Wellington and Noble 1985). It follows that when direct seeding E.costata, matching sowing to spring or winter, particularly in years of predicted high rainfall, may also improve emergence results. Although summer has a higher average rainfall, the high temperatures could limit germination. Autumn temperatures are within the favourable germination range, although the lower average rainfall at that time could reduce germination percentage.
This study used laboratory-based experiments on the germination behaviour of E. costata, but there may be different germination and emergence results from field trials because conditions in the field will differ from the sterile conditions found in the laboratory (Ronnenberg et al. 2008; El-Keblawy et al. 2018). However, we believe the knowledge generated from these trials can offer valuable guidance for restoration practitioners working with species such as this, helping improve restoration outcomes. Given the low tolerance of saline conditions, the majority of E. costata seeds would likely prefer to be sown at a distance from the highly saline salt lakes found around the seed collection area (Westbrooke 2012). A light burial, either in sandy soil or leaf litter, would also improve germination conditions.
Data availability
The dataset used during the study is available from the corresponding author upon request.
Acknowledgements
The authors thank Michael McBain for creating Fig. 1. The authors also acknowledge Dr Paul Novelly, Editor-in-Chief of The Rangeland Journal, and the anonymous reviewers for their constructive feedback on earlier versions of this paper, which significantly improved its quality.
Author contributions
Callum Measham-Pywell and Singarayer Florentine contributed to the study concept and design. Trial setup and data collection were performed by Callum Measham-Pywell as part of his final year Undergraduate research project unit. Data analysis was performed by Joseph Stapleton. Report write-up was primarily performed by Joseph Stapleton, with Callum Measham-Pywell contributing to the Methods section. All authors have reviewed and approved the final paper.
References
Arnold S, Kailichova Y, Knauer J, Ruthsatz AD, Baumgartl T (2014) Effects of soil water potential on germination of co-dominant Brigalow species: implications for rehabilitation of water-limited ecosystems in the Brigalow Belt bioregion. Ecological Engineering 70, 35-42.
| Crossref | Google Scholar |
Bond WJ, Honig M, Maze KE (1999) Seed size and seedling emergence: an allometric relationship and some ecological implications. Oecologia 120(1), 132-136.
| Crossref | Google Scholar | PubMed |
Bradford KJ (2002) Applications of hydrothermal time to quantifying and modeling seed germination and dormancy. Weed Science 50(2), 248-260.
| Crossref | Google Scholar |
Bureau of Meteorology (2024) Climate Data Online Vol. 2024. (Commonwealth of Australia) Available at http://www.bom.gov.au/climate/data/
Callister K, Westbrooke M (2017) A field trial to test effects of watering, seed addition and disturbance on perennial species recruitment in Belah woodland. Ecological Management and Restoration 18(3), 246-249.
| Crossref | Google Scholar |
Dalziell EL, Lewandrowski W, Commander LE, Elliott CP, Erickson TE, Tudor EP, Turner SR, Merritt DJ (2022) Seed traits inform the germination niche for biodiverse ecological restoration. Seed Science and Technology 50(2), 103-124.
| Crossref | Google Scholar |
Duncan C, Schultz NL, Good MK, Lewandrowski W, Cook S (2019) The risk-takers and -avoiders: germination sensitivity to water stress in an arid zone with unpredictable rainfall. AoB Plants 11(6), plz066.
| Crossref | Google Scholar | PubMed |
Dunn OJ (1964) Multiple comparisons using rank sums. Technometrics 6(3), 241-252.
| Crossref | Google Scholar |
El-Keblawy A, Gairola S, Elsheikh EA, Hussain MI, Abhilash P (2018) Variability in seed germination behaviour of six grasses under laboratory and natural field conditions: implications for restoration of degraded lands in subtropical arid deserts. Tropical Ecology 59(4), 715-726.
| Google Scholar |
Emery NJ, Collette JC (2021) Drought stress affects the germination of four co-occurring Eucalyptus species from north-west New South Wales. Australian Journal of Botany 69(3), 143-151.
| Crossref | Google Scholar |
Facelli JM, Ladd B (1996) Germination requirements and responses to leaf litter of four species of eucalypt. Oecologia 107(4), 441-445.
| Crossref | Google Scholar | PubMed |
Ferrari FN, Parera CA (2015) Germination of six native perennial grasses that can be used as potential soil cover crops in drip-irrigated vineyards in semiarid environs of Argentina. Journal of Arid Environments 113, 1-5.
| Crossref | Google Scholar |
Florentine S, Milberg P, Di Stefano J, Westbrooke M, Graz P (2015) Decade-long response of arid-land mallee vegetation to fire, flooding and grazing in south-eastern Australia. Journal of Arid Environments 121, 7-14.
| Crossref | Google Scholar |
Grant CD, Bell DT, Koch JM, Loneragan WA (1996) Implications of seedling emergence to site restoration following bauxite mining in Western Australia. Restoration Ecology 4(2), 146-154.
| Crossref | Google Scholar |
Gutterman Y (1994) Strategies of seed dispersal and germination in plants inhabiting deserts. The Botanical Review 60(4), 373-425.
| Crossref | Google Scholar |
Houston J (2006) Evaporation in the Atacama Desert: an empirical study of spatio-temporal variations and their causes. Journal of Hydrology 330(3–4), 402-412.
| Crossref | Google Scholar |
Huang L, Wang D, Yao L, Li X, Wang D, Du Q, Zhang Y, Hou X, Guo Y (2019) Primary limitation on vegetation productivity shifts from precipitation in dry years to nitrogen in wet years in a degraded arid steppe of Inner Mongolia, northern China. Journal of Soils and Sediments 19(2), 544-556.
| Crossref | Google Scholar |
Jiménez-Alfaro B, Silveira FAO, Fidelis A, Poschlod P, Commander LE (2016) Seed germination traits can contribute better to plant community ecology. Journal of Vegetation Science 27(3), 637-645.
| Crossref | Google Scholar |
Kruskal WH, Wallis WA (1952) Use of ranks in one-criterion variance analysis. Journal of the American Statistical Association 47(260), 583-621.
| Crossref | Google Scholar |
Lozano-Isla F, Benites-Alfaro OE, Pompelli MF (2019) GerminaR: an R package for germination analysis with the interactive web application ‘Germinaquant for R’. Ecological Research 34(2), 339-346.
| Crossref | Google Scholar |
Mechergui K, Jaouadi W, Naghmouchi S, Alsubeie M, Khouja ML (2019) Micromorphological observation of eucalyptus seeds, multivariate statistical analyses and modeling of their germination under salt stress and osmotic constraint. Cerne 25(2), 156-171.
| Crossref | Google Scholar |
Mesgaran MB, Mashhadi HR, Alizadeh H, Hunt J, Young KR, Cousens RD (2013) Importance of distribution function selection for hydrothermal time models of seed germination. Weed Research 53(2), 89-101.
| Crossref | Google Scholar |
Munro NT, Moseby KE, Read JL (2009) The effects of browsing by feral and re-introduced native herbivores on seedling survivorship in the Australian rangelands. Rangeland Journal 31(4), 417-426.
| Crossref | Google Scholar |
Onofri A, Benincasa P, Mesgaran MB, Ritz C (2018) Hydrothermal-time-to-event models for seed germination. European Journal of Agronomy 101, 129-139.
| Crossref | Google Scholar |
Onofri A, Mesgaran MB, Ritz C (2022) A unified framework for the analysis of germination, emergence, and other time-to-event data in weed science. Weed Science 70(3), 259-271.
| Crossref | Google Scholar |
Poschlod P, Abedi M, Bartelheimer M, Drobnik J, Rosbakh S, Saatkamp A (2013) Seed Ecology and Assembly Rules in Plant Communities. In ‘Vegetation Ecology’. (Eds E van der Maarel, J Franklin) pp. 164–202. (John Wiley & Sons) 10.1002/9781118452592.ch6
R Core Team (2024) R: a language and environment for statistical computing, 2022.7.2.576. (R Foundation for Statistical Computing: Vienna, Austria). Available at https://www.R-project.org/
Ronnenberg K, Wesche K, Hensen I (2008) Germination ecology of Central Asian Stipa spp: differences among species, seed provenances, and the importance of field studies. Plant Ecology 196(2), 269-280.
| Crossref | Google Scholar |
Ruiz Talonia L, Reid N, Gross CL, Whalley RDB (2017) Germination ecology of six species of Eucalyptus in shrink-swell vertosols: moisture, seed depth and seed size limit seedling emergence. Australian Journal of Botany 65(1), 22-30.
| Crossref | Google Scholar |
Ruiz-Talonia L, Carr D, Smith R, Whalley RDB, Reid N (2018) Effect of temperature and light on germination of 10 species of Eucalyptus from north-western NSW. Australian Journal of Botany 66(8), 657-666.
| Crossref | Google Scholar |
Stapleton J, Turner SR, Warne D, Florentine S (2024) Linking ex situ germination to in situ direct seeding for landscape scale restoration efforts in the semiarid Mallee region of Victoria, Australia. Restoration Ecology 33, e14220.
| Crossref | Google Scholar |
Stavi I, Thevs N, Priori S (2021) Soil salinity and sodicity in drylands: a review of causes, effects, monitoring, and restoration measures. Frontiers in Environmental Science 9, 712831.
| Crossref | Google Scholar |
Verhoeven EM, Murray BR, Dickman CR, Wardle GM, Greenville AC (2020) Fire and rain are one: extreme rainfall events predict wildfire extent in an arid grassland. International Journal of Wildland Fire 29(8), 702-711.
| Crossref | Google Scholar |
Wellington AB, Noble IR (1985) Post-fire recruitment and mortality in a population of the mallee Eucalyptus incrassata in semi-arid south-eastern Australia. Journal of Ecology 73(2), 645-656.
| Crossref | Google Scholar |
Westbrooke M (2012) The pastoral history, biological and cultural significance of the Scotia Country, far western New South Wales. Proceedings of the Linnean Society of New South Wales 134, A55-A68.
| Google Scholar |
White MD (2006) The mallee vegetation of north western Victoria. Proceedings of the Royal Society of Victoria 118(2), 229-243.
| Google Scholar |
Williams WD (1999) Salinisation: a major threat to water resources in the arid and semi-arid regions of the world. Lakes and Reservoirs: Research and Management 4(3–4), 85-91.
| Crossref | Google Scholar |