

A vertical bait station for black rats (Rattus rattus) that reduces bait take by a sympatric native rodent
Frances Zewe A B E , Paul Meek A B C , Hugh Ford D and Karl Vernes AA Ecosystem Management, School of Environmental and Rural Sciences, University of New England, Armidale, NSW 2351, Australia.
B Invasive Animals Cooperative Research Centre, Orange Agricultural Institute, Locked Bag 6001, Orange, NSW 2800, Australia.
C Vertebrate Pest Research Unit, NSW Department of Primary Industries, Orange Agricultural Institute, Locked Bag 6001, Orange, NSW 2800, Australia.
D Zoology, School of Environmental and Rural Sciences, University of New England, Armidale, NSW 2351, Australia.
E Corresponding author. Email: fzewe@myune.edu.au
Australian Mammalogy 36(1) 67-73 https://doi.org/10.1071/AM13010
Submitted: 25 April 2013 Accepted: 12 October 2013 Published: 13 December 2013
Abstract
Novel bait stations can be used as a targeted method of delivering bait by exploiting behavioural traits of the target species. On Muttonbird Island, New South Wales, the black rat (Rattus rattus) has been baited to aid the conservation of the island’s wedge-tailed shearwater (Ardenna pacifica) colony, which may result in poisoning of the sympatric swamp rat (Rattus lutreolus). We aimed to design a bait station that R. rattus could reach, but that R. lutreolus could not. We found that 11 (92%) of 12 captive R. rattus reached the bait chambers by climbing a 50-cm vertical pipe, whereas only four (18%) of 22 R. lutreolus reached these bait stations. In a field trial on Muttonbird Island R. rattus entered the bait chamber on an average of 5.3 events per night of vertical bait station deployment, but R. lutreolus did not enter the stations. In a field trial on the mainland at a site with a high density of R. lutreolus, this species was detected in one vertical bait station five times, equating to an average of 0.017 events per night of vertical bait station deployment. We conclude that R. rattus readily climbs a 50-cm pipe to enter the bait station, whereas R. lutreolus rarely or never does on Muttonbird Island or at the mainland site.
Additional keywords: camera trap, island conservation, seabirds, swamp rat (Rattus lutreolus).
Introduction
Many island ecosystems have been threatened by invasive rodents, particularly black rats (Rattus rattus), brown rats (R. norvegicus) and house mice (Mus musculus) (Atkinson 1985). Invasive rodents have been implicated in the decline and extinction of insular seabirds (Atkinson 1985; Towns et al. 2006; Jones et al. 2008; Banks and Hughes 2012) and mammals (Burbidge et al. 1997; Burbidge 1999; Burbidge and Manly 2002; Harris 2009; Banks and Hughes 2012). Predation by exotic rats on Australian Islands was recognised as a key threatening process under the EPBC Act 1999 in 2006, and soon afterwards a Threat Abatement Plan was prepared by the Australian Government (DEWHA 2009). The eradication of exotic rodents from islands has led to the recovery of endemic fauna (e.g. Jouventin et al. 2003; Lorvelec and Pascal 2005). However, some islands are highly prone to reinvasion because they are near the mainland, or connected to it by human-made structures. In these cases, permanent eradication of invasive rodents is impossible and therefore sustained control may be needed.
Muttonbird Island, located 500 m off the north coast of New South Wales and connected to the mainland by a breakwall, is home to a large breeding colony of wedge-tailed shearwaters (Ardenna pacifica). Although the species is not threatened, nearly 30 species of seabirds that breed in Australia are threatened or near-threatened, often due to rodent predation (Garnett et al. 2011). Most breed on remote islands, but Muttonbird Island attracts over 100 000 visitors annually, providing a unique opportunity for educating the public about the threats experienced by seabirds. Volunteer bird-banders have been monitoring a subset of the A. pacifica population on the island for ~40 years, with their data suggesting a declining population on the island and poor breeding success in recent years (Narelle Swanson, pers. comm.).
The exotic R. rattus and the native swamp rat (R. lutreolus) coexist on the island (Egan 2008). Given the impact of R. rattus on seabirds elsewhere, predation of eggs and chicks by introduced rodents is listed as a potential threat to A. pacifica in the Muttonbird Island Plan of Management (NSW National Parks and Wildlife Service 2009). Furthermore, the island population of R. lutreolus differs genetically from that on the neighbouring mainland, suggesting that it is a long-term isolate on the island (CPCG 2010). Because R. lutreolus and R. rattus forage at ground level, and are of similar size, the resident R. lutreolus on Muttonbird Island are at risk of being poisoned during any attempt to control R. rattus.
Two published studies have demonstrated the ability of vertical bait stations to deliver bait to R. rattus but not to coexisting similar-sized native rodents: the Anacapa Island deer mice (Peromyscus maniculatus anacapae) (~25 g) on Anacapa Island, USA (Erickson et al. 1990), and the Santa Fe rice rat (Oryzomys bauri) (~65 g), the large Fernandina rice rat (Nesoryzomys narboroughi) (~78 g) and the Santiago Galapagos mouse (N. swarthi) (~98 g) in the Galapagos Islands (Phillips et al. 2007). Differences in the climbing ability between R. rattus, which is an agile and competent climber (Watts and Aslin 1981; Erickson et al. 1990; Morris 2002; Phillips et al. 2007), and the non-target species, which all climb less, have been used to design bait stations in these studies. R. lutreolus relies heavily on runways along the ground in dense ground cover (Fox and Monamy 2007), and so may not be well adapted for climbing. On the basis of this apparent lack of climbing ability, we postulated that vertical bait stations could exclude R. lutreolus. Therefore, the aim of this study was to test the hypothesis that R. rattus would climb to bait stations raised vertically to 50 cm in the field and laboratory, whereas R. lutreolus would be unable or unwilling to climb such bait stations.
Materials and methods
Site descriptions
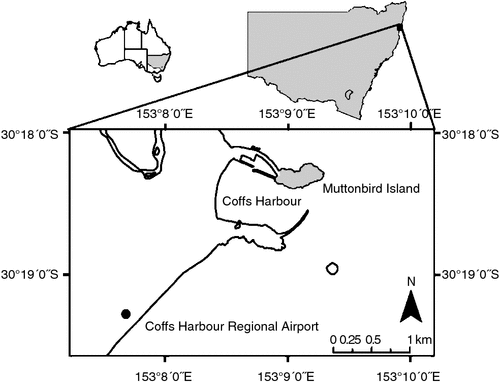
Two study sites were used: the 8-ha Muttonbird Island (30°18′S, 153°09′E), located off Coffs Harbour, on the New South Wales mid-north coast, Australia; and a heathland site on the mainland near the Coffs Harbour Regional Airport (30°18′S, 153°07′E) (Fig. 1). Muttonbird Island is a nature reserve to protect the A. pacifica colony, which breeds from August to May (Swanson and Merritt 1974). The island is connected to the mainland by a 500-m-long breakwall that forms the northern wall of the Coffs Harbour International Marina. A paved walking track running east–west along the island provides pedestrian access to view the A. pacifica colony.
|
The vegetation of Muttonbird Island primarily consists of low-lying grasses, sedges and herbs, including Commelina cyanea, Dianella caerulea, Kennedia rubicunda and Senecio lautus. Flagellaria indica and Cupaniopsis anacardioides occur in patches on the island and the exotic Lantana camara is common on the northern edge. The Coffs Harbour Regional Airport (CHRA) site is heathland with Xanthorrhoea and Banksia species interspersed with stands of Leptospermum polygalifolium, bordered to the east by a narrow stand of Melaleuca quinquinervia swamp forest.
Study species
Egan (2008) estimated the density of R. rattus and R. lutreolus on the island to be 6.5 and 9 animals ha–1 respectively in 2008. There were also 53 M. musculus per ha, based on mark–recapture, although recent estimates suggest much higher populations of M. musculus and R. lutreolus (P. Meek, unpublished data). The western (landward) section of the island was baited to control R. rattus before each A. pacifica breeding season from 2007 to 2010 using a second-generation anticoagulant rodenticide presented in 20 conventional ground-based bait stations. Sporadic baiting also occurred along the breakwall.
Vertical bait stations
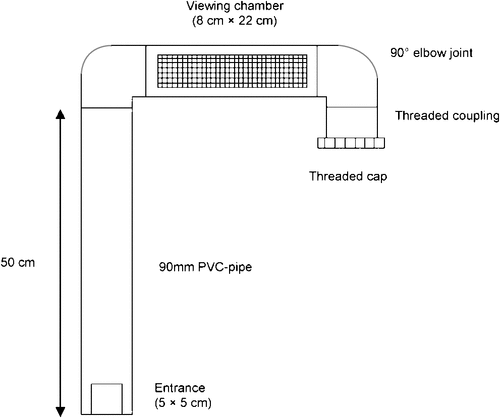
The vertical bait station we tested was modified from a design used in the Galapagos Islands by Phillips et al. (2007). Polyvinyl chloride (PVC) pipe (90 mm diameter) cut into 50-cm, 30-cm and 10-cm lengths attached to 90° bends and screw cap ends were used to construct each bait station (Fig. 2). A 5 × 5 cm square hole cut at the base of the upright pipe allowed rodents to enter the bait station, and reach the bait by climbing the vertical section. We securely mounted each bait station on two star pickets using plastic cable ties in the field. The internal surface of the PVC pipe was not modified. The external surfaces of the PVC-pipe and fittings were spray-painted with green and brown to camouflage them in the field so as to reduce detection and tampering by the general public. The material cost about AU$9 per bait station in 2010. The bait chamber was a 30-cm horizontal section with a viewing chamber to photograph and identify animals entering it, using camera traps. A section (8 × 22 cm) was removed from the horizontal pipe and covered by a piece of 12.5-mm aviary mesh slightly larger than the viewing chamber (Fig. 2).
|
Captive trials
The climbing ability of the two rat species was tested by a series of captive trials conducted in an indoor laboratory at the University of New England Field Station, Arrawarra Headland, ~30 km north of Muttonbird Island. We trapped R. lutreolus for the trials on Muttonbird Island (n = 10) and at the CHRA site (n = 12). We trapped R. rattus on Muttonbird Island (n = 9) and at a residence near Coffs Harbour (n = 3). Rats were housed individually in cylindrical mesh enclosures (70 × 80 cm), each with one vertical bait station. Rats were provided with water ad lib, a cardboard box with shredded paper in it for a nest box and 6 g of commercial rat meal every day of the trial.
The trials allowed for staggered entry of rats into captivity on the basis of our success in trapping them, and started the day after each animal was captured and transported to the field station. Rats had access to each elevation of the bait station for 24 h unless otherwise stated. On the first day of the trial the bait station, with no food, was laid on its side and not elevated. On the second day the bait station was also laid on its side, but was baited with 5 g of peanut butter and rolled oat mixture. Supplementary food (6 g commercial rat meal) was also provided each day outside each bait station so the rats could access it without climbing. Then the bait stations were progressively raised vertically to 10 cm, 25 cm and finally to 50 cm by increasing the length of the upright section of PVC-pipe, to determine how high the rats would climb. Each rat was given a maximum of four nights to enter the elevated bait station, starting with a height of 10 cm. If the rat successfully entered, then the following day the station was elevated to the next height until the maximum height of 50 cm was reached. Failure to enter the station after four nights at any height resulted in the conclusion of the trial for that animal.
The bait stations were monitored with a Reconyx PC90 RapidFire camera trap (Reconyx Inc., Holmen, WI, USA) by cutting holes in the perspex lid of the holding cages corresponding to the infrared sensor and lens and placing the camera horizontally face-down above each cage. The cameras were set to capture five photographs at 1-s intervals after being triggered, followed by a 30-s stand-by period. Therefore two methods were used to confirm successful entry into the bait station in the captive trials: (1) the consumption of bait from the station (by comparing initial and final bait weights), and (2) the examination of camera trap photographs for images of the rodent in the bait chamber.
Rodent enclosures and bait stations were spot-cleaned daily by removing soiled bedding and faecal material from the cages, and thoroughly cleaned with a dilute solution of dishwashing detergent and hot water, rinsed in fresh water and air-dried before housing a new rat. After the trial the sex, reproductive condition, mass and length of the pes, manus, head, body (snout to vent) and tail of each rat were recorded before release (R. lutreolus) or euthanasia (R. rattus).
Muttonbird Island field trial
Two vertical bait stations (VBS1 and VBS2) were set up on the eastern side of Muttonbird Island 10 m apart. This part of the island was chosen because of its low density of A. pacifica burrows. Two different brands of camera trap were used during the Muttonbird Island field trial: the Reconyx PC90 RapidFire and the PixController DigitalEye (PixController Inc., Export, PA, USA). Both cameras used passive infrared sensors to detect animal movement, but the Reconyx has an infrared flash whereas the PixController has a xenon flash. One of each camera type was deployed on a tripod at each bait station, which was baited with peanut butter and rolled oat mixture for three nights (i.e. 6 camera-nights). A small portion of peanut butter and rolled oat mixture was scattered around the base of the bait station in every field trial as a form of prefeeding, as employed by Phillips et al. (2007) in the Galapagos Islands. However, we found this technique to be largely ineffective because we deployed the bait stations during the day and ants consumed most of the scattered bait before dark. Therefore photos of rats actually feeding on this scattered bait were rare. In order to reduce the risk of theft, cameras were deployed at 1800 hours and collected at 0700 hours daily. At VBS1 the PixController and Reconyx cameras were set to take a single photograph with a 10- and 15-s stand-by period respectively; at VBS2 both cameras were set to take a single photograph with a 30-s stand-by period.
The Muttonbird Island field trial was conducted in November when the A. pacifica were digging burrows and mating, which limited our ability to move off the pedestrian path to deploy cameras and check bait stations, due to the risk of causing burrow collapse. In addition, the island was visited frequently by the public, so there was a risk to camera security. This meant that cameras were deployed only at night, which may have underestimated visitation by R. lutreolus individuals, which are partially diurnal (Braithwaite 1977; Watts and Aslin 1981; Meek et al. 2012). We therefore conducted another trial at the CHRA, which has a large population of R. lutreolus and two previous instances of R. rattus occurring on the northern edge of the site. Few people visit this site, so we were able to deploy the cameras continuously throughout the trial.
Coffs Harbour Regional Airport field trial
Ten bait stations were mounted on star pickets at intervals of 20 m along two transects. The vegetation surrounding each bait station and its camera was cleared with hand tools. A 30 × 40 cm section of black non-slip mat was placed at the entrance to the bait station to provide a contrasting background to assist animal identification in the photographs taken by the cameras. The bait stations were baited with peanut butter and rolled oat mixture encapsulated within a tea strainer and securely fastened inside the bait station to ensure the bait was present throughout the trial.
The first and second trials were conducted in March and November/December of the same year, respectively, and the bait stations were removed from the field between trials. Due to camera availability, two different models of Reconyx camera with the same functionality were used for the two field trials: a PC90 RapidFire for the first trial (38 camera-nights) and a HC600 HyperFire (Reconyx Inc., Holmen, WI, USA) for the second trial (250 camera-nights). In the first trial cameras were mounted on tripods 145 cm from the bait stations, which we hoped would detect rodent movement at both the entrance and viewing chamber of the stations. However, on one occasion in the second trial, we found R. lutreolus scats in the bait chamber, with no corresponding photograph taken by the camera. We presumed that this was due to the placement of the camera, so after setting the camera at various distances from the station we found that the optimal distance for exclusively detecting animals in the viewing chamber was 50 cm, and so we moved the cameras to this distance for the second half of the second trial. After this time any R. lutreolus near the entrance of the station would have been missed, which was acceptable because we knew that R. lutreolus were present at the site, and our aim was to determine whether they were able to climb into the bait chamber.
Data analysis
Photographs of animals visiting the bait stations were catalogued according to date, time and activity. An event was defined as the detection of an animal until the activity of the animal changed. Key morphological features between the two rat species, such as tail length relative to body length, and ear size and morphology (Watts and Aslin 1981), allowed us to identify rats to species level in most photographs from the field trials. Each event was classified as a ‘probable’ or ‘definite’ detection of a species, depending on our confidence in identifying each animal to species level.
We used Fisher’s Exact Test to test for differences in the proportion of rats of each species that entered the bait station at each height. Analysis of variance (ANOVA) and unpaired t-tests were used to test for a difference in body mass between the R. lutreolus and R. rattus tested in the captive trials, and to determine any relationship between body mass and maximum height climbed. Minitab for Windows 15.1 was used for statistical analysis.
Results
Captive trials
All R. rattus and R. lutreolus entered the unelevated bait stations and those elevated to 10 cm (Fig. 3). R. rattus and R. lutreolus varied in their ability to climb to 25 cm (Table 1), but the difference was not significant (Fisher’s Exact Test, P = 0.069). A greater proportion of R. rattus (92%) than R. lutreolus (18%) climbed to 50 cm (Fisher’s Exact Test, P < 0.001). Although more R. lutreolus from CHRA than from Muttonbird Island climbed to 25 cm, this was marginally non-significant (Fisher’s Exact Test, P = 0.0557).

|

|
The mean mass (±s.e.) of the rats used in the captive trials was 127.8 ± 18.7 g for R. rattus (n = 12), 121.0 ± 8.7 g for Muttonbird Island R. lutreolus (n = 10) and 102.7 ± 7.7 g for CHRA R. lutreolus (n = 12), which were not significantly different (P = 0.373). The mean mass of the R. rattus did not differ from that of the R. lutreolus from the two populations combined (P = 0.409). There was no relationship between body mass of R. lutreolus and maximum height climbed (ANOVA, F = 0.030, d.f. = 21, P = 0.864). The one R. lutreolus from Muttonbird Island and three R. lutreolus from CHRA that climbed to 50 cm were all males. One R. lutreolus was found dead the morning after the third night of the trial at 50 cm, but as it had failed to climb to 50 cm the two previous nights the data were included in the analysis.
Muttonbird Island field trial
The animals photographed in and around the bait station were identified to species in 223 cases. Of these, 210 (94%) were R. rattus, with four R. lutreolus (2%), seven Pacific black ducks (Anas superciliosus) (3%) and two A. pacifica (1%). No M. musculus were photographed at the stations.
R. rattus were photographed in the viewing chamber in 32 (15%) events, indicating that they had climbed the 50-cm vertical section of the bait station (Fig. 4). This equates to an average of 5.3 successful entries per bait station per night of deployment. Because rats were not marked we do not know how many individual R. rattus actually climbed the vertical bait station. Two R. rattus were present in 50 (24%) of the camera trap images with one or both in the bait station, or around the base of the station. R. rattus were climbing externally on the bait station or on the star pickets in 28 (13%) events, on the black mat near at the entrance to the bait station in 94 (45%) events and in the area surrounding the black mat in 56 (27%) events. All four R. lutreolus events were of individual animals near the entrance to the bait station, with no successful entries detected.

|
Coffs Harbour Regional Airport field trial
R. lutreolus accounted for a total of 56 definite and three probable detections in the trials at the CHRA. They were most commonly photographed on the mat (36 events) or in the vegetation surrounding the vertical bait station (13 events). R. lutreolus were also observed placing their forepaws on the external surface of the upright PVC-pipe (three events) and feeding on bait scattered at the entrance to the bait station (one event). One R. lutreolus was photographed in the viewing chamber in five events at the same bait station in the second field trial, which equates to 0.017 successful entries per bait station per night of deployment. The size of the rat was consistent across photographs, but rats were not marked so we cannot confirm whether it was the same animal or not. A single R. rattus was photographed climbing externally on the bait station but no other rodents were detected. Other species present at the vertical bait stations included two detections of swamp wallabies (Wallabia bicolor), one detection of a pheasant coucal (Centropus phasianinus) and 13 detections of Australian ravens (Corvus coronoides).
Discussion
This experiment evaluated whether the introduced R. rattus and the native R. lutreolus were physically able to climb and enter a vertical bait station. In the captive trial, most of the R. rattus (11 of 12; 92%) climbed the vertical bait station to a height of 50 cm, which supports our hypothesis that R. rattus are good climbers. The apparently good climbing ability of black rats was mirrored in the field trial on Muttonbird Island where R. rattus were observed in the bait chamber in 32 occasions over 6 camera-trap-nights. Our result is similar to the 28 (93%) of 30 R. rattus that reached 45 cm in the study by Erickson et al. (1990). Unlike in our trial, Erickson et al. (1990) used toxic bait in the bait chamber, and so visitation was inferred from death of the rat. Erickson et al. (1990) suggested that R. rattus climb a vertical pipe by adopting a ‘spread-eagled’ stance and exerting pressure onto the insides of the pipe. If this is so, then larger rats may be better equipped for climbing. Our results may support this suggestion as the only R. rattus that failed to climb to 50 cm was a small animal. It is, however, unlikely that absolute size is the only determinant of climbing ability because the R. lutreolus used in the trial were no lighter, on average, than the R. rattus.
In contrast to the results from the R. rattus trials, few R. lutreolus from Muttonbird Island (1 of 10; 10%) and the CHRA site (3 of 12; 25%) climbed to 50 cm in the captive trial. This does not support our hypothesis that R. lutreolus are unable to climb. It is likely that R. lutreolus were more motivated to climb in the captive trials than the field trials because they had little other food and were restricted in where they could move. This is similar to the findings of Weerakoon and Banks (2011) who showed that black rats were more willing to eat peanut butter and oats mixture with higher doses of Rhodamine B when they had to work to access other foods. A R. lutreolus was photographed in one of the 10 vertical bait stations at CHRA five times. It is possible that all photographs were of the same R. lutreolus repeatedly entering the station, though the animals were not marked so we cannot confirm this. So a small percentage of R. lutreolus are physically able, and willing, to climb to 50 cm; however, we suggest that this would be a much lower value than would enter conventional ground-based bait stations. Although the proportion that climbed to 50 cm did not differ significantly between mainland and Muttonbird Island swamp rats, mainland rats may be better climbers. Hence, whereas bait stations at 50 cm above the ground should lead to minimal mortality of swamp rats on Muttonbird Island, further trials should be conducted on mainland sites before using 50-cm bait stations there, as more swamp rats may be killed.
Camera traps were effective in identifying mammalian visitors to the bait stations. By placing a wire mesh window near the bait chamber we could photograph and identify most rats that reached the bait chamber and hence infer the likelihood of each species being poisoned if we had used toxic bait. We initially had a problem with the horizontal banding pattern of the Reconyx camera’s detection zone, which we addressed by moving the camera closer to the bait station. It is important that the users of camera traps determine precisely the nature of the array of the passive infrared sensor (detection zone) of their cameras. For example, in our study it was crucial that animals in the relatively narrow viewing chamber were detected, or visits by rats may be missed. We used the removal of bait as a second measure for detecting visitors to the bait station in the captive trials and are confident that we did not miss any visits by the rats.
A baiting program using just vertical bait stations may be limited if not all R. rattus enter them. Erickson et al. (1990) suggested that this problem was not unique to vertical bait stations, as some rats may also fail to visit conventional bait stations. A study of bait station preferences of R. rattus in New Zealand found that 75%, 87% and 100% of wild-caught captive R. rattus entered a wooden box, yellow plastic pipe and wooden tunnel bait stations respectively (Spurr et al. 2007). The similarity in the proportions of R. rattus that entered conventional bait stations in Spurr et al. (2007) and vertical bait stations in this study suggests that few R. rattus would avoid using vertical bait stations. Other factors that may affect the likelihood of rodents entering bait stations include their density and social behaviour, attractiveness of the bait, availability of natural foods, weather conditions and the number of bait stations (Erickson et al. 1990). While we are confident that the use of vertical bait stations would reduce the incidence of swamp rats consuming toxic bait, future research should focus on any population-level reduction in R. rattus numbers that could be achieved by deploying vertical bait stations during rodent control campaigns.
Acknowledgements
This research was conducted under the University of New England Animal Ethics Committee Authority No. AEC09/163, NSW Scientific Licence No. S13004 and Forests NSW Special Purposes Permit for Research No. 48911. We thank Stuart Green for assisting in the Muttonbird Island R. lutreolus field trial and Ray Ward and Jeff Taylor for design and construction of the rat enclosures. We are grateful to Narelle Swanson for providing extensive data on numbers and breeding success of A. pacifica. FZ was supported by the Office of Environment and Heritage (formerly the NSW Department of Environment, Climate Change and Water) and the Northern Rivers Catchment Management Authority (NRCMA) ‘Caring for our Country’ funding program, and would like to thank the staff at the Coffs Harbour OEH office, particularly Ann Walton, Glenn Storrie and Dean Egan, for their support and encouragement.
References
Atkinson, I. A. E. (1985). The spread of commensal species of Rattus to oceanic islands and their effect on island avifaunas. In ‘Conservation of Island Birds: Case Studies for the Management of Threatened Island Birds’. ICBP Technical Publication No. 3. (Ed. P. J. Moors.) pp. 35–81. (International Council for Bird Preservation: Cambridge.)Banks, P. B., and Hughes, N. K. (2012). A review of the evidence for potential impacts of black rats (Rattus rattus) on wildlife and humans in Australia. Wildlife Research 39, 78–88.
| A review of the evidence for potential impacts of black rats (Rattus rattus) on wildlife and humans in Australia.Crossref | GoogleScholarGoogle Scholar |
Braithwaite, R. W. (1977). Preliminary observations on the activity patterns of Rattus lutreolus and other Victorian small mammals. The Victorian Naturalist 94, 216–219.
Burbidge, A. (1999). Conservation values and management of Australian islands for non-volant mammal conservation. Australian Mammalogy 21, 67–74.
Burbidge, A., and Manly, B. F. J. (2002). Mammal extinctions on Australian islands: causes and conservation implications. Journal of Biogeography 29, 465–473.
| Mammal extinctions on Australian islands: causes and conservation implications.Crossref | GoogleScholarGoogle Scholar |
Burbidge, A., Williams, M. R., and Abbott, I. (1997). Mammals on Australian islands: factors influencing species richness. Journal of Biogeography 24, 703–715.
| Mammals on Australian islands: factors influencing species richness.Crossref | GoogleScholarGoogle Scholar |
CPCG (2010). Assessment of genetic structure of Rattus lutreolus from Muttonbird Island, Coffs Harbour, NSW. Report to the Department of Environment, Climate Change and Water. Centre for Plant Conservation Genetics, Southern Cross University.
DEWHA (2009). Threat abatement plan to reduce the impacts of exotic rodents on biodiversity on Australian offshore islands of less than 100 000 ha. Department of the Environment, Water, Heritage and the Arts, Canberra.
Egan, D. (2008). Rodent population, diet, habitat usage and their management implications for Muttonbird Island, Coffs Harbour, NSW. B.Nat.Res.(Honours) Thesis, University of New England, Armidale.
Erickson, W. A., Marsh, R. E., and Halvorson, W. L. (1990). A roof rat bait station that excludes deer mice. Wildlife Society Bulletin 18, 319–325.
Fox, B. J., and Monamy, V. (2007). A review of habitat selection by the swamp rat, Rattus lutreolus (Rodentia: Muridae). Austral Ecology 32, 837–849.
| A review of habitat selection by the swamp rat, Rattus lutreolus (Rodentia: Muridae).Crossref | GoogleScholarGoogle Scholar |
Garnett, S. T., Szabo, J. K., and Dutson, G. (2011). ‘The Action Plan for Australian Birds 2010.’ (CSIRO Publishing: Melbourne.)
Harris, D. B. (2009). Review of negative effects of introduced rodents on small mammals on islands. Biological Invasions 11, 1611–1630.
| Review of negative effects of introduced rodents on small mammals on islands.Crossref | GoogleScholarGoogle Scholar |
Jones, H. P., Tershy, B. R., Zavaleta, E. S., Croll, D. A., Keitt, B. S., Finkelstein, M. E., and Howald, G. R. (2008). Severity of the effects of invasive rats on seabirds: a global review. Conservation Biology 22, 16–26.
| Severity of the effects of invasive rats on seabirds: a global review.Crossref | GoogleScholarGoogle Scholar | 18254849PubMed |
Jouventin, P., Bried, J., and Micol, T. (2003). Insular bird populations can be saved from rats: a long-term experimental study of white-chinned petrels Procellaria aequinoctialis on Ile de la Possession (Crozet Archipelago). Polar Biology 26, 371–378.
Lorvelec, O., and Pascal, M. (2005). French attempts to eradicate non-indigenous mammals and their consequences for native biota. Biological Invasions 7, 135–140.
| French attempts to eradicate non-indigenous mammals and their consequences for native biota.Crossref | GoogleScholarGoogle Scholar |
Meek, P. D., Zewe, F., and Falzon, G. (2012). Temporal activity patterns of the swamp rat (Rattus lutreolus) and other rodents in north-eastern New South Wales, Australia. Australian Mammalogy 34, 223–233.
| Temporal activity patterns of the swamp rat (Rattus lutreolus) and other rodents in north-eastern New South Wales, Australia.Crossref | GoogleScholarGoogle Scholar |
Morris, K. D. (2002). The eradication of the black rat (Rattus rattus) on Barrow and adjacent islands off the north-west coast of Western Australia. In ‘Turning the Tide: The Eradication of Invasive Species’. (Eds C. R. Veitch and M. N. Clout.) pp. 219–225. (IUCN SSC Invasive Species Specialist Group: Gland, Switzerland & Cambridge, UK.)
NSW National Parks and Wildlife Service (NPWS) (2009). Muttonbird Island Nature Reserve plan of management. New South Wales NPWS North Coast Region, NSW Department of Environment and Climate Change, Sydney.
Phillips, R. B., Harris, D. B., and Snell, H. W. (2007). Bait stations for detection and control of alien rats in Galapagos. The Journal of Wildlife Management 71, 2736–2742.
| Bait stations for detection and control of alien rats in Galapagos.Crossref | GoogleScholarGoogle Scholar |
Spurr, E. B., Morriss, G. A., Turner, J., O’Connor, C. E., and Fisher, P. (2007). Bait station preferences of ship rats. DOC Research & Development Series No. 271. New Zealand Department of Conservation, Wellington.
Swanson, N. M., and Merritt, F. D. (1974). The breeding cycle of the wedge-tailed shearwater on Mutton Bird Island, N.S.W. The Australian Bird Bander 12, 3–9.
Towns, D. R., Atkinson, I. E. A., and Daugherty, C. H. (2006). Have the harmful effects of introduced rats on islands been exaggerated? Biological Invasions 8, 863–891.
| Have the harmful effects of introduced rats on islands been exaggerated?Crossref | GoogleScholarGoogle Scholar |
Watts, C. H. S., and Aslin, H. J. (1981). ‘The Rodents of Australia.’ (Angus and Robertson: Sydney.)
Weerakoon, M. K., and Banks, P. B. (2011). Not just a matter of taste: palatability of bait markers is influenced by the need to search for alternative food. Wildlife Research 38, 596–602.
| Not just a matter of taste: palatability of bait markers is influenced by the need to search for alternative food.Crossref | GoogleScholarGoogle Scholar |