

Livestock guardian dog protection of free-range poultry from the red fox
S. Roddick A , T. L. Kreplins A B , H. T. Kobryn A and P. A. Fleming A *
A B , H. T. Kobryn A and P. A. Fleming A *
A Centre for Terrestrial Ecosystem Science and Sustainability, Harry Butler Institute, Murdoch University, 90 South Street, Murdoch, WA 6150, Australia.
B Department of Primary Industries and Regional Development, 75 York Road, Northam, WA 6401, Australia.
Animal Production Science 62(13) 1290-1302 https://doi.org/10.1071/AN21229
Submitted: 28 April 2021 Accepted: 10 March 2022 Published: 26 April 2022
© 2022 The Author(s) (or their employer(s)). Published by CSIRO Publishing. This is an open access article distributed under the Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License (CC BY-NC-ND)
Abstract
Context: Predation of layer chickens is a major issue for free-range egg producers. Using livestock guardian dogs (LGD) to protect free-ranging poultry is a possible option for producers, although there is little published literature regarding how the dogs protect chickens.
Aims: This case study was conducted at a free-range egg production farm in Western Australia, where red foxes (Vulpes vulpes) were a common predator of chickens prior to introducing Maremma LGD. We investigated LGD responses to experimental cues that might indicate fox incursion (fox urine and calls).
Methods: Four dogs were GPS tracked and monitored using camera traps. Over the first week, experimental fox cues were set out around the paddock boundaries, alternating with ‘non-cue’ experimental control nights. We recorded whether the LGD altered (1) their space use, (2) activity patterns (movement speed), or (3) behaviour in response to these cues. We also recorded (4) distances between LGD from known sightings of foxes.
Key results: The Maremmas appeared to work independently of each other, covering separate areas. There was no significant difference in overnight home range area by experimental fox cue treatment, but there was a significant (P < 0.001) treatment × dog interaction term for distance moved. Three dogs spent most of their time at night around the chicken shelters and generally increased distances moved on experimental fox cue nights. The fourth dog was more bonded to people and did not alter its movements. Paradoxically, dogs rested more and barked less on experimental fox cue nights; however, we recorded foxes on camera traps placed around the chicken shelters on 17 of the 23 nights of monitoring, and the high background activity level of foxes on this property compromised our experimental control (nights without experimental fox cues). The dogs did not move towards known fox sightings.
Conclusions: The Maremmas in this trial closely guarded the chicken shelters rather than maintaining the entire paddock as a predator-exclusion zone.
Implications: Understanding how guardian dogs behave when challenged by potential predators will help increase producers’ confidence in the efficacy of these dogs as a viable method to protect livestock from predation threat.
Keywords: animal welfare, behaviour, GPS, guardian dog, livestock predation, Maremma, poultry, predator.
Introduction
Consumer demand for welfare-friendly food choices has driven change in many food production industries. Some of the greatest changes have been witnessed by the egg production industry, with consumer demands globally driving movement from intensive conventional cage eggs to a range of more welfare-friendly alternatives (Australian Eggs 2021). For example, the use of conventional cages has been replaced by the use of enriched/furnished cages in some European countries (e.g. Sweden, Switzerland and Austria; Appleby 2003), and legislation was passed in 2014 to ban conventional cages in Australian Capital Territory Australia (Brennan 2014).
In place of conventional cages, there has been increasing demand for free-range egg production over the past decade. While the behaviour of free-range chickens has greater consumer appeal, free-range chickens are much more vulnerable to predators than are those under intensive production systems, and predation is one of the main challenges for this industry (Knierim 2006; Van de Weerd et al. 2009). A major predator of outdoor livestock is the red fox (Vulpes vulpes; Saunders et al. 1995; Fleming et al. 2016), which has the widest geographic distribution of any non-domesticated carnivore (Statham et al. 2014). These opportunistic predators were introduced into Australia and have quickly spread to become established across most of the continent (Fairfax 2019). The impacts of foxes on livestock, particularly poultry, were recognised within only a few years of their introduction (Saunders et al. 2010). Today, it is predicted that there is a population of approximately 1.7 million foxes in Australia (Stobo-Wilson et al. 2022), imposing substantial costs on producers and the environment (Bradshaw et al. 2021). It is estimated that foxes cause A$28 million loss due to lamb predation, and they represent a significant proportion of the estimated A$46 million spent by broad acre farmers and livestock producers on vertebrate pest control (McLeod 2016). These cost estimates largely ignore their significant impact on poultry production.
Losses to predators can be difficult to quantify in many livestock industries (Bestman and Bikker-Ouwejan 2020), especially where the predator removes the killed animal (Fleming et al. 2016). Foxes are commonly reported on free-range layer farms (Scott et al. 2018). Domestic poultry is also recorded in the diets of foxes; some studies have recorded a low occurrence of poultry in fox diets (0.4–6% frequency of occurrence; Latham 1950; Coman 1973; Croft and Hone 1978; Dawson et al. 2016), while other studies have reported much greater values (14–36% frequency of occurrence; Latham 1950; Korschgen 1959). Differences among studies in the reported occurrence in fox diet are likely to reflect differences in poultry numbers present with location. Recognition of poultry losses to predators can also vary due to different flock sizes and methods of survey (Knierim 2006), making direct comparison among studies and production systems difficult. Losses to predators (predator not specified) exceeding 20 birds per production period have been reported for 9% of 96 surveyed Swiss free-range farms (each with more than 500 hens; mean 3042 ± 3600 birds per farm; Häne et al. 2000). In Britain, losses to foxes amount to 2 ± 17% (mean ± s.d.) across 50 surveyed farms (35–130 000 hens, median 11 000), with three-quarters of respondents identifying losses to red foxes (Moberly et al. 2004). However, losses could reach up to 100% of small flocks of less than 200 birds (Heydon and Reynolds 2000).
With an increased customer demand for free-range eggs, there is an increasing proportion of the layer hen flocks that are exposed to potential predation risk. The need for predator control for free-range poultry is therefore of increasing importance. However, this shift to socially acceptable free-range production has also seen a growing demand for non-lethal forms of predator control (Gehring et al. 2010), further increasing the challenge for poultry producers.
Livestock guardian dogs (LGD) originated across central Europe and Asia centuries ago to help protect livestock against predators. LGD are generally large-bodied breeds (35–45 kg, 65 cm or higher at the shoulders; Smith et al. 2010). They are bonded to livestock from an early age, have been bred to be calm and inoffensive around livestock in their behaviour and appearance, and do not engage in stalking or intimidation towards livestock, as would be expected from herding dogs (van Bommel 2010). LGD have been effective protection against predators across many parts of the world (e.g. Yilmaz et al. 2015). In Australia, a common breed is the Maremmano-Abruzzese Sheepdog (‘Maremma’), which originates from Italy. These LGD are primarily used to guard small stock such as sheep, goats and poultry (van Bommel and Johnson 2012; King et al. 2015). A survey of 150 Australian livestock producers with LGD indicated that 21% of respondents used them to guard free-range poultry (van Bommel and Johnson 2012). Understanding the behaviour of LGD around layer farms could benefit poultry producers and potentially increase use of these dogs for livestock protection.
A number of studies have quantified the behaviour of LGD around livestock (principally sheep; Van Eeden et al. 2018). LGD directly respond to threat, using pursuit and vocalisation (barking, growling) to deter predators (e.g. Shivik 2006; van Bommel and Johnson 2015), which can also be effective through alerting people (Eklund et al. 2017). LGD may also confront a potential predator, chasing and intimidating them into retreat, or sometimes the dog will even attack and kill the predator (Matt Longworth, pers. comm.; van Bommel 2010). LGD may also indirectly protect livestock by maintaining territories, using scent marking to advertise their presence and deterring predators from the area around livestock (van Bommel and Johnson 2015).
Tracking studies suggest that LGD use both direct and indirect defence of livestock. Zingaro et al. (2018) used GPS collars to monitor movements and proximity to sheep flocks for 29 LGD (mostly Maremmas) across 11 farms in Italy. They found that the dog’s proximity to their stock varied with environment, but that the dogs never left their flock unattended. Similarly, van Bommel and Johnson (2014) GPS-tracked 14 Maremmas across three Australian sheep properties to record movement patterns of the dogs, and also identified responses to playback of dingo calls and presentation of dingo urine (van Bommel and Johnson 2015). These Maremmas spent an average of 90% of their time with the livestock and showed territorial behaviour, vocalising and scent marking on the range margins (van Bommel and Johnson 2014). Maremmas always responded to playback of a dingo call. Dingo urine incited a response, while a control cue (distilled water) never received a response (although in a separate trial, captive dingoes showed minimal differences in their responses to urine marks of LGD compared with water controls; van Bommel and Johnson 2017). In contrast, Allen et al. (2017) GPS tracked eight Maremma LGD as well as six free-ranging wild dogs (dingoes, feral domestic dogs and their hybrids; Canis familiaris) for 7 months on an Australian sheep property and found that the wild dogs regularly spent time inside the LGD dog’s territories, although direct physical altercations seldom occurred. Allen et al. (2017) suggested that rather than maintaining an exclusive territory, the Maremmas defended their flocks by boisterous vocalisation. These studies suggest possible differences in how groups of LGD respond to incursion by a predator, dependent on their environment and the nature of the predator or cues about predator presence.
Effectiveness of active forms of livestock defence is likely to depend on the numbers of stock that LGD are protecting, as well as the distribution of livestock and predators across the landscape. Consequently, their defence methods for extensive sheep or goat systems are likely to be different from how the dogs guard poultry. There has been no published study quantifying the behaviour patterns of LGD around free-range poultry. We therefore used camera traps and GPS tracking collars to investigate movement patterns of LGD used to guard free-range chicken flocks and tested a manipulation where we provided olfactory (fox urine) and auditory (playback of fox calls) experimental cues that would suggest fox presence. We quantified LGD (1) space use, (2) activity patterns (speed of movement), and (3) behaviour (resting, walking, running, barking and smelling/urination) by experimental fox cue treatment. We also recorded (4) the minimum distances between the LGD and known fox incursions into the paddock (foxes captured on camera traps).
Materials and methods
This work was completed under Murdoch University’s Animal Ethics Committee permit IRMA2844/16.
Study site
The study site was a free-range egg farm situated in Kauring, Western Australia, approximately 124 km east of Perth. The study was conducted between 2 and 24 August 2016. Average daily temperature during August was 12.3°C, with a maximum of 20.9°C and a minimum of 5.9°C. Rainfall during August 2016 was 57.7 mm, making it the second-highest monthly average in the year of study after July (Bureau of Meteorology 2020). On this farm, there were two paddocks with laying chickens guarded by Maremma LGD (Fig. 1). To train these Maremmas, they were introduced to chickens at an early age and the puppies were penned in a fenced-off kennel area next to a chicken shelter (and therefore could see but not interact with the chickens). Time spent with the chickens by the Maremmas was slowly increased over successive weeks; with the puppies full time in with the chickens from 12 weeks old (Matt Longworth, pers. comm.). The hens and Maremmas were all free-ranging and not penned at night.

|
Paddock 1 was a 27-ha olive grove with 3000 chickens (Hyline brown) spread over five open-plan chicken shelters (various dome ecoshelters or metal sheds; Fig. 1). Each shelter was the focal point for introduction of a cohort of hens (although they were free to disperse from these areas), and therefore the paddock contained chickens of a range of laying ages. There were six working Maremmas (three males and three females, aged 14 months to 4 years; all animals were entire, Table 1) and a litter of four 6-week old puppies from one of the bitches. Although they were free-roaming at the time of the study, neither the pups or the females were tracked in this study as it was thought that pregnancy or lactating could alter their responses to predators. Egg production during the study period was low in comparison to previous years, with approximately 700–800 eggs/day, which were collected during daylight hours by a team of farm workers. The perimeter wire fence was 1.5 m high with every second strand of the six strands electrified to keep the Maremmas and chickens inside the paddocks; a fox could easily pass through this fencing and were regularly observed to do so (for the paddocks as well as for fencing around the farm more generally; Matt Longworth, pers. comm.). A residential dwelling, where 4–6 farm workers resided, was located about 50 m south of the paddock. The Maremmas were fed outside the chicken shelter furthermost from the worker’s residence as a conscious effort to move the Maremmas away from proximity to the farm residence. The Maremmas ranged in sociability, with one extremely friendly and social Maremma and others that were less sociable or even anti-social towards people. Actual chicken numbers lost to foxes were unknown; overall losses were estimated by the landholder at about 800 individuals over a 12-month period (the sum for both paddocks) but included deaths due to disease/illness. Foxes had been sighted within Paddock 1 on multiple occasions; the Maremmas had caught and killed a juvenile fox in the 6 months prior to the study.

|
Paddock 2 was effectively a 0.411-ha enclosure with approximately 100 chickens (mixed breeds) and two chicken shelters (Table 1). There were three Maremmas (two entire males: 2 and 4 years old, and one entire 4-year-old female). Paddock 2 was about 250 m from the farm residence. The rate of egg production in Paddock 2 (not recorded; farm manager, pers. comm.) was lower than that in Paddock 1, although no foxes had been recorded in the paddock by the manager. Paddock 2 was surrounded by a 1-m-high electrified perimeter fence of a construction similar to that of other fences on the farm. Maremmas in Paddock 2 were fed near one of the chicken shelters.
All Maremmas were fed daily in the morning by the farm workers; often the dogs were also reported to eat chicken eggs.
Experimental design
We used camera traps and GPS tracking to compare Maremma behaviour (camera traps: presence along the fence-line boundaries for each paddock; and GPS tracking: home-range area by using GPS tracking) for nights with and without experimental cues to simulate the presence of a red fox. Each experimental fox cue was set out in the late afternoon (16:00 hours) and retrieved the next morning (08:00 hours). The paddock was then rested for 1 day/night. The following three combinations of experimental fox cue were used:
-
On Day 3 of the trial, female fox urine (collected from a carcass derived from a landscape-scale control action; Red Card for the Red Fox) was pipetted into a new plastic urine specimen jar and placed immediately outside the paddock fences overnight. The fox urine was stored in a plastic jar at room temperature between sessions.
-
On Day 5 of the trial, fox calls (downloaded from http://www.angelfire.com/ar2/thefoxden/sounds.html; foxcry.wav) were played through a portable mp3 player placed immediately outside each of the paddock fences at 16:00 hours and played for 5 min at 2-h intervals over the next 16 h.
-
On Day 7 of the trial (9 August), both fox urine and fox call were used together.
No experimental fox cues were used for the last 2 weeks of monitoring (10–24 August).
Monitoring methods
Space use
GPS trackers (i-gotU GT-120 travel logger, Australia) were fitted daily to four Maremma collars to record their movements (Table 1), including three males from Paddock 1 and one male from Paddock 2; the second male in Paddock 2 was not approachable to collar. The tracking devices were set to record GPS location (latitude and longitude), speed of activity, and duration of activity every minute. Generally, the units lasted 24 h before requiring 2 h to recharge; rarely, they lost power after 6 h.
In ArcGIS (ESRI, USA), kernel density estimates (KDE; 50% and 90% percentage volume contours; PVC) were formulated for each Maremma (quartic kernel function, cell size = 2 m, search radius (smoothing parameter) = 20 m, using the approach by Silverman (1986)) over the entire 22-day study period (all data points) for visualisation. We also calculated the dogs’ nocturnal space use (there were no farm workers in the paddock at night and this is also when foxes were likely to be active) for each night to allow statistical comparison of nightly space use by fox treatment (two separate analyses: experimental control nights vs (1) experimental fox cue nights each considered separately, or (2) experimental fox cue nights all together) using a Kruskal–Wallis ANOVA. To aid visualisation (as per Fig. 2), we created a hexagonal grid (each hexagonal cell with an area of 120 m2) used to display counts of night-time GPS locations by experimental fox cue treatment within each hexagon cell.

|

|
Activity patterns (speed of movement)
To identify how activity patterns were influenced by the experimental fox cue treatments, we compared the dog’s speed (speed of movement estimated for each minute by the i-gotU GPS tracking software as dependent variable) by using a generalised linear mixed model in R (R Core Team 2018). For this analysis, fox treatment (two separate analyses: experimental control nights vs (1) experimental fox cue nights each considered separately, or (2) experimental fox cue nights all together), dog ID (to account for individual differences) and the treatment × dog ID interaction term were included as predictor variables. Because there was a circadian pattern in speed, we also included sine-hour (with maximum values at midday and minimum values at mid-night) as a covariate. We tested for model fit using the quartile–quartile plot fit function in ‘DHARMa’ (Hartig and Lohse 2020), which indicated overdispersion of residuals due to a high proportion of zeros (i.e. the dog was inactive). A negative binomial fit was inappropriate because the data were non-integers, and therefore a tweedie GLM was fitted to these data with the alpha value set to maximise normality of residuals as indicated using the ‘tweedie’ package (Dunn 2017). We used the ggpredict function in ‘ggeffects’ (Lüdecke 2018) to plot the predicted speeds with circadian variation in activity speed held constant, and used ‘emmeans’ (Lenth et al. 2018) to conduct pairwise Tukey analysis by experimental fox cue treatment.
LGD behaviour classification
To capture the LGD and fox activity, Reconyx™ XR6 Ultrafire Covert video-camera traps (Reconyx, Holmen, USA) were attached to steel posts around the perimeter of each paddock, with cameras positioned 450 mm above the ground (this height was aimed to maximise capture rates of the dogs, as we had not initially thought that foxes would enter the paddock) and set to 45° angle down the boundary fence facing into the paddock. The camera traps were set to high sensitivity and collected 10-s duration videos, with no lag phase. We initially positioned camera traps around the border of the paddocks, as we presumed that this would be where the Maremmas would be likely to defend the paddock from intrusion by foxes. However, our results indicated that the Maremmas did not respond by moving out to the paddock boundary in response to experimental fox cues; subsequently, on completion of the experimental manipulations, we re-positioned cameras around the chicken huts to increase capture of activity. During the first phase of the experiment, when experimental fox cues were presented, camera traps were placed in 10 positions around the edges of Paddock 1 (seven cameras were positioned on the boundary fence, the other three positions were located in from the boundary fence), excluding the northern corner as it had dense bush which the landholder had observed that the Maremmas did not use, and in four locations around the boundary of Paddock 2. Two camera traps were set in front of the experimental fox cues. During the second phase of the experiment, when no experimental fox cues were presented, three camera traps from around the boundary of Paddock 1 and the four camera traps from Paddock 2 were all moved to in front of the chicken huts in Paddock 1 (Fig. 1).
Video footage was observed using image viewer (Microsoft image viewer) and scored for behaviour of the LGD (resting, walking, running, barking, smelling/urination, eating), time of day (day/night), and experimental fox cue treatment (cue present yes/no). We compared the proportion of camera events where we identified Maremmas barking, running, smelling/urinating, walking, or resting for experimental fox cue nights (total 43 camera events over three nights of monitoring) by Pearson’s chi test, with expected values based on proportions calculated for the same behavioural categories for experimental control nights (total 674 camera events over 20 control nights), acknowledging that the camera positions had changed for phase 2 of monitoring.
Distances between LGD from known sightings of foxes
For fox sightings from camera traps, we categorised their behaviour (walking, running, smelling/urination, eating), and calculated the approximate distance (to the nearest 25 m) to each LGD wearing a GPS tracker, by using ArcGIS Euclidean distance function (ESRI, USA).
Results
In Paddock 1, where three male Maremmas were tracked simultaneously, these dogs worked independently of each other and covered separate areas of the paddock (Fig. 2). Dog A spent time at each of the chicken shelters, but the majority of his time was spent around Shelters 4 and 5. Dog B was observed across a large proportion of Paddock 1 and in all chicken shelters except Shelter 3, and was observed on camera interacting with hens in the shelter in the evening, herding and even appearing to pick up animals in his jaws. The space occupied by Dogs A and B for the 22-day experiment were concentrated around the chicken shelters (Fig. 2a, e). Dog C moved along the front fence, his patrol route typically followed farm workers in the paddock (who performed maintenance, as well as daily egg collection and animal husbandry inspection) and he covered a smaller area than did the other two dogs (Fig. 2i). Dog D’s KDE for the 22-day experiment showed that he spent most of his time in the larger chicken shelter in Paddock 2 (Fig. 2m). We predominantly recorded hens on cameras located around the shelters; hens were rarely observed on the cameras located around the perimeter of Paddock 1 (hens were more likely seen at the border of Paddock 2, due to its smaller size).
Space use
The four collared LGD did not significantly increase their home range area on experimental fox cue nights compared with experimental control nights (Table 2), as there was no statistically significant difference in area used for the three experimental fox cue nights considered separately or all together, from experimental control nights (the area covered on experimental fox cue nights was within the 95% confidence intervals of the means for experimental control nights, Table 1). This lack of statistical significance of the treatment effect for area covered was largely because the pattern of responses was not consistent among dogs (Fig. 2).
Activity patterns (speed of movement)
The dogs showed individual differences in their responses to the presentation of experimental fox cues. Dogs A and D both moved significantly faster on experimental fox cue nights than on experimental control nights (Fig. 3). Predicted speeds (after time of day was accounted for in the analyses) indicated that Dog A increased his activity speed by 154% and Dog D by 120% for experimental fox cue nights. Dog B increased his average speed by 114% for experimental fox cue nights, although this difference was not significant. Dog C spent the majority of his time closest to people and significantly decreased his average speed on experimental fox cue nights (85% of the average speed for experimental control nights).

|
LGD behaviour classification
We recorded a different profile of LGD behaviour classification (resting, walking, running, barking and smelling/urination behaviour recorded on video cameras) for experimental fox cue nights compared with experimental control nights (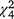 = 16.28, P = 0.003; Fig. 4). Paradoxically, the LGD were more often observed resting (
= 16.28, P = 0.003; Fig. 4). Paradoxically, the LGD were more often observed resting (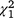 = 6.02, P = 0.014) and less often recorded barking (
= 6.02, P = 0.014) and less often recorded barking (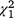 = 4.99, P = 0.025) on experimental fox cue nights (noting that seven cameras had been moved for Phase 2 of the experiment, as cameras positioned around the paddock boundary during Phase 1 had shown little activity). Additionally, when LGD were observed on camera traps on experimental fox cue nights, they were not observed on those cameras that were positioned near the cues.
= 4.99, P = 0.025) on experimental fox cue nights (noting that seven cameras had been moved for Phase 2 of the experiment, as cameras positioned around the paddock boundary during Phase 1 had shown little activity). Additionally, when LGD were observed on camera traps on experimental fox cue nights, they were not observed on those cameras that were positioned near the cues.

|
Distances between LGD and known sightings of foxes
No foxes were seen on camera within the smaller Paddock 2. However, foxes were recorded on cameras on 17 of the 23 nights of monitoring in Paddock 1, with 25 independent camera-trap events. On these occasions, foxes were captured on cameras that averaged 64 ± 43 m from the chicken shelters. While most observations of foxes showed them running or walking through the paddock, foxes were twice observed on camera eating something (which appeared to be eggs), and in another instance a fox was seen to carry what looked to be an egg. A fox was also observed urinating, another appeared to be smelling the ground. Foxes were observed on cameras positioned within 50 m of the chicken shelters on 15 occasions, including being observed on cameras in front of chicken shelters in the middle of the paddock on five occasions. No LGD was seen on camera near or after the fox on any occasion. At the time that a fox was seen on camera, GPS records indicated that Dog A was an average of 243 ± 159 m away; he was at another chicken shelter on 11 of 15 potential occasions. Dog B was 471 ± 140 m away (n = 7 occasions). Dog C was 362 ± 167 m away (n = 19 occasions) when foxes were seen on camera and was at the fence closest to the farmworker’s residence on all occasions. Note that the number of responses vary because of collar failure or flat batteries at the time of the respective fox incursions.
Discussion
Three of the four dogs (Dog A, B and D) that we monitored appeared to be strongly bonded to the chickens they were guarding, spending the majority of their time overnight around the chicken shelters and increasing their speed of activity on experimental fox cue nights. However, despite the dogs’ activity, foxes were confirmed present within Paddock 1 on three quarters of the nights that we monitored and may well have been present, just not captured on camera, on the other nights. Foxes were observed on camera consuming or carrying eggs, but we had no record of predation on chickens during the monitoring period and chicken losses on this property had anecdotally reduced markedly since the introduction of the LGD. Therefore, although the dogs were not excluding foxes from around the chicken shelters, they were likely to have still been reducing predation on the hens.
Our data does not suggest that the Maremmas excluded foxes from entering Paddock 1. The dogs were not commonly seen on the camera traps positioned around the paddock perimeter and their GPS tracking collars showed that Dogs A and B spent the majority of their time immediately around the chicken shelters (Dog C showed preference for the edge of the paddock closest to the worker’s residence). Furthermore, fox presence in Paddock 1, including the activity of foxes on cameras positioned in front of the chicken shelters, indicates that the Maremmas did not maintain the paddock as a predator-exclusion zone. Rather than responding to all predator incursions, they appeared to maintain a small predator-free zone immediately around the chicken huts. The observation that foxes intruded into the free-range chicken paddock suggests a similar scenario as reported by Allen et al. (2017), who found that wild dogs would regularly spend time inside the LGD territories, or McGrew and Blakesley (1982), who noted that Komondor LGD stayed very close to the sheep at all times to prevent predation by coyotes (Canis latrans), actively defending only when they had to. When presented with an experimental predator cue (dingo call or urine), van Bommel and Johnson 2015 reported that LGD were more likely to actively respond when the cue was nearby rather than at a distance (although the dogs did move up to 570 m away from the stock to challenge a perceived threat). These results together support the presumption that LGD actively defend a small area around their livestock.
We set out to determine whether LGD would actively respond to cues suggesting fox presence (fox urine and calls), but we found no statistically significant difference in area covered (home range area) for nights with or without experimental fox cues. This result could have been due to the small number of experimental fox cue nights (n = 3 nights; which reduces the power for statistical analysis), individual differences in responses among dogs, or masking by the substantial amount of background fox activity on the farm, meaning that our experimental control treatment was not sufficient contrast to the experimental fox cues. We did find that three of the four dogs tracked generally increased their activity on experimental fox cue nights (two dogs significantly increased their average speeds), which may have increased their ability to monitor the paddock on these nights.
It is possible that a group of five adult dogs in the 27-ha Paddock 1 would find it difficult to simultaneously monitor ∼3000 chickens distributed across five shelters, and therefore it is probably not surprising that there were fox incursions into this paddock. By contrast, the LGD in Paddock 2 had minimal area to maintain and their presence in any part of the paddock may have been sufficient deterrent to foxes (no fox incursions were recorded for this paddock during our period of monitoring). The numbers and distribution of the birds are therefore likely to influence the effectiveness of LGD protecting free-range chickens.
Individual differences in dog responses
While the other three tracked dogs spent the majority of their time around the chicken shelters and generally increased the distances they moved on experimental fox cue nights, Dog C spent the majority of his time at night at the front fence close to the worker’s residence. This dog did not change his behaviour on any occasion of a confirmed fox sighting in the paddock and even significantly decreased his activity speeds on experimental fox cue nights, suggesting that he was not responding to potential threat to the chickens. A similar observation for LGD has been noted previously (Rust et al. 2013), with inattentiveness being the primary cause of LGD removals on South African farms, accounting for 33% of removals. However, the same study also reported that retraining to more deeply entrench the bond with livestock is possible, with one in three dogs with behaviour problems responding to corrective training (Rust et al. 2013).
The individual differences in space use and activity that we found are likely to translate to differences in guarding behaviour and responses to disturbance by a potential predator. van Bommel and Johnson 2015 found that individual dogs responded differently to a stimulus; one dog raised the alarm, one or two investigated, and one or two remained with their livestock. Similarly, McGrew and Blakesley (1982) noted individual differences in responses, with aggressive dogs being more effective at protecting sheep; the authors note that breeding for more ‘guardian like’ characteristics can improve guarding ability of Komodor LGD.
The Maremmas in the present study were all relatively young working animals. The oldest males in Paddock 1 (Dog A) and Paddock 2 (Dog D) were both 4 years of age. By contrast, Dogs B and C were both 2 years old and likely to not yet have been at their full potential, as guardian Maremmas are technically not mature until about 2 years of age (van Bommel 2010). An animal’s age and experience can influence their behavioural responses to predator cues, with older dogs being less active but more likely to associate with their livestock (Rust et al. 2013; van Bommel and Johnson 2014; Zingaro et al. 2018). Mixing groups of dogs according to their personality and age can therefore potentially increase their ability to guard livestock as a group (van Bommel 2010).
Limitations of the study
We had only two separate paddocks on this farm, which differed in size, numbers of chickens and numbers of dogs. There was therefore likely to be differences in vulnerability of the chickens to foxes. Ideally, this study could be repeated on additional properties to identify whether the LGD behavioural responses we recorded are common to incursion by foxes.
The size of the local fox population in the study area was not known, although foxes were regularly seen on parts of the farm away from the paddocks and a juvenile fox had been killed by the LGD in Paddock 1 within 6 months of this study. We also have no indication of whether or not fox abundance or activity had been reduced by the presence of the LGD. With very little background information on fox activity on the farm, in retrospect, our experimental design was naïve. We recorded foxes on camera traps within the paddock on 17 of 23 (74%) nights of monitoring, suggesting that fox scents and sounds were present even for our experimental control nights, undermining the effect of our experimental manipulation.
It was expected that the guarding activities of LGD (i.e. running, barking and scent marking; McGrew and Blakesley 1982) would increase on nights with greater perceived threat (i.e. experimental fox cues). However, in contrast to our prediction, we recorded significantly less barking and more resting on cameras placed around the paddock boundary for experimental fox cue nights. While three dogs were generally more active on experimental fox cue nights, their activity was concentrated around the chicken shelters. In contrast, Dog C did not alter his behaviour and spent most of his time at the paddock boundary, near the farmworkers’ residence; he was therefore likely to have contributed disproportionately to behaviour analysed from sightings of LGD on the boundary cameras on these nights (we could not distinguish among individuals from the nocturnal camera footage).
Another factor that could have compromised the experimental treatment effect in our study is that the fox urine used may not have stimulated the LGD’s guarding instinct due to the lack of glandular secretions generally associated with a fox urine marking (Henry 1977), or the way it was presented and stored (in a plastic container). Similarly, the fox calls may not have been believeble to the LGD, being distorted through recording and playback. It may have been more useful to use a distressed chicken call to illicit an active response. We also established our experimental design assuming that the dogs might maintain the whole paddock as an exclusive territory, and therefore placed our experimental fox cues around the paddock perimeter. Given that foxes entered the paddock and were commonly seen around the chicken huts, placing the cues closer to the chicken huts would be interesting to test, as it could represent a more realistic scenario for the dogs to respond to.
While it would be unethical to remove the guardian dogs for an experimental manipulation, and there were no control paddocks to observe fox behaviour in the absence of LGD, other experimental designs are suggested for future research. It could be possible to track foxes and LGD simultaneously to determine whether foxes avoided the dogs, as has been undertaken to simultaneously observe the behaviour of wild dogs and LGD (Allen et al. 2017). Repetition of this study at other farms with working LGD is also recommended, being mindful that each farm will have different environmental conditions, housing setup for the chickens, and numbers of dogs and livestock.
Conclusions
Our study was a novel experiment looking at the guardianship of free-ranging layer chickens by Maremma LGD. The four GPS-tracked LGD were active in their movements throughout the paddocks they were assigned to, but those in Paddock 1 did not maintain an exclusion zone or prevent foxes from entering the paddock. We also found no significant increase in home range area that might suggest that these LGD changed their space use to actively defend the paddock boundaries against an intrusion or a perceived intrusion of a fox. Instead, the LGD guarded the chicken shelters closely and three of the dogs increased their activity (average speed of movement) in response to experimental fox cues. Outside this experiment, the LGD had killed foxes that were found dead inside Paddock 1, and loss of chickens on the farm was anecdotally noted as reduced after introduction of the dogs.
Maremmas have a knack for changing their behaviour to suit their environment and the animals they are protecting, as somewhat evident from the substantial individual differences in responses we recorded. The few previous studies investigating Maremmas have examined other types of livestock or predators, and measured different aspects of the dogs’ behaviour. Generalising across studies is therefore not possible, and further data are still required. Livestock producers are increasingly selecting Maremmas for predator control in free-ranging poultry and will therefore benefit from greater understanding of how these dogs are doing their jobs. The fox is a persistent and opportunist predator but may have met its match in LGD.
Data availability
Raw data will be shared upon reasonable request to the corresponding author.
Conflicts of interest
The authors declare no conflicts of interest.
Declaration of funding
This work was funded by the Royalties for Regions Boosting Biosecurity fund BBD RDF14-00034.
Acknowledgements
The authors thank Matt Longworth for allowing access to his stock and LGD.
References
Allen LR, Stewart-Moore N, Byrne D, Allen BL (2017) Guardian dogs protect sheep by guarding sheep, not by establishing territories and excluding predators. Animal Production Science 57, 1118–1127.| Guardian dogs protect sheep by guarding sheep, not by establishing territories and excluding predators.Crossref | GoogleScholarGoogle Scholar |
Appleby MC (2003) The European Union ban on conventional cages for laying hens: history and prospects. Journal of Applied Animal Welfare Science 6, 103–121.
| The European Union ban on conventional cages for laying hens: history and prospects.Crossref | GoogleScholarGoogle Scholar | 12909526PubMed |
Australian Eggs (2021) Annual Report 2021. (Australian Eggs: Sydney, NSW, Australia) Available at https://www.australianeggs.org.au/who-we-are/annual-reports [Accessed 25 March 2022]
Bestman M, Bikker-Ouwejan J (2020) Predation in organic and free-range egg production. Animals 10, 177
| Predation in organic and free-range egg production.Crossref | GoogleScholarGoogle Scholar |
Bradshaw CJA, Hoskins AJ, Haubrock PJ, Cuthbert RN, Diagne C, Leroy B, Andrews L, Page B, Cassey P, Sheppard AW, Courchamp F (2021) Detailed assessment of the reported economic costs of invasive species in Australia. NeoBiota 67, 511–550.
| Detailed assessment of the reported economic costs of invasive species in Australia.Crossref | GoogleScholarGoogle Scholar |
Brennan M (2014) ‘ACT bans battery cages and sow stalls.’ Available at https://www.aph.gov.au/About_Parliament/Parliamentary_Departments/Parliamentary_Library/FlagPost/2014/February/ACT_bans_battery_cages_and_sow_stalls#:∼:text=On%2025%20February%202014%20the,by%20the%20ACT%20Legislative%20Assembly [Accessed 25 March 2022]
Bureau of Meteorology (2020) Climate data online. Rainfall data for Station number 010165. Bureau of Meteorology. Available at http://www.bom.gov.au/climate/data/ [Accessed 20 April 2020]
Coman BJ (1973) The diet of red foxes, Vulpes vulpes L., in Victoria. Australian Journal of Zoology 21, 391–401.
| The diet of red foxes, Vulpes vulpes L., in Victoria.Crossref | GoogleScholarGoogle Scholar |
Croft JD, Hone LJ (1978) The stomach contents of foxes, Vulpes vulpes, collected in New South Wales. Australian Wildlife Research 5, 85–92.
| The stomach contents of foxes, Vulpes vulpes, collected in New South Wales.Crossref | GoogleScholarGoogle Scholar |
Dawson SJ, Crawford HM, Huston RM, Adams PJ, Fleming PA (2016) How to catch red foxes red handed: identifying predation of freshwater turtles and nests. Wildlife Research 43, 615–622.
| How to catch red foxes red handed: identifying predation of freshwater turtles and nests.Crossref | GoogleScholarGoogle Scholar |
Dunn PK (2017) Tweedie: evaluation of tweedie exponential family models. R package version 2.3.
Eklund A, López-Bao JV, Tourani M, Chapron G, Frank J (2017) Limited evidence on the effectiveness of interventions to reduce livestock predation by large carnivores. Scientific Reports 7, 2097
| Limited evidence on the effectiveness of interventions to reduce livestock predation by large carnivores.Crossref | GoogleScholarGoogle Scholar | 28522834PubMed |
Fairfax RJ (2019) Dispersal of the introduced red fox (Vulpes vulpes) across Australia. Biological Invasions 21, 1259–1268.
| Dispersal of the introduced red fox (Vulpes vulpes) across Australia.Crossref | GoogleScholarGoogle Scholar |
Fleming PA, Dundas SJ, Lau YYW, Pluske JR (2016) Predation by red foxes (Vulpes vulpes) at an outdoor piggery. Animals 6, 60
| Predation by red foxes (Vulpes vulpes) at an outdoor piggery.Crossref | GoogleScholarGoogle Scholar |
Gehring TM, VerCauteren KC, Landry J-M (2010) Livestock Protection Dogs in the 21st century: is an ancient tool relevant to modern conservation challenges? BioScience 60, 299–308.
| Livestock Protection Dogs in the 21st century: is an ancient tool relevant to modern conservation challenges?Crossref | GoogleScholarGoogle Scholar |
Häne M, Huber-Eicher B, Fröhlich E (2000) Survey of laying hen husbandry in Switzerland. World’s Poultry Science Journal 56, 21–31.
| Survey of laying hen husbandry in Switzerland.Crossref | GoogleScholarGoogle Scholar |
Hartig F, Lohse L (2020) DHARMa: residual diagnostics for hierarchical (multi-level/mixed) regression models. R package version 0.3.
Henry JD (1977) The use of urine marking in the scavenging behaviour of the red fox (Vulpes vulpes). Behaviour 61, 82–106.
| The use of urine marking in the scavenging behaviour of the red fox (Vulpes vulpes).Crossref | GoogleScholarGoogle Scholar | 869875PubMed |
Heydon MJ, Reynolds JC (2000) Fox (Vulpes vulpes) management in three contrasting regions of Britain, in relation to agricultural and sporting interests. Journal of Zoology 251, 237–252.
| Fox (Vulpes vulpes) management in three contrasting regions of Britain, in relation to agricultural and sporting interests.Crossref | GoogleScholarGoogle Scholar |
King K, Wallis R, Wallis A, Peucker A, Williams D (2015) Successful protection against canid predation on little penguins (Eudyptula minor) in Australia using maremma guardian dogs: the ‘Warrnambool method’. International Journal of Arts & Sciences 8, 139
Knierim U (2006) Animal welfare aspects of outdoor runs for laying hens: a review. NJAS-Wageningen Journal of Life Sciences 54, 133–145.
| Animal welfare aspects of outdoor runs for laying hens: a review.Crossref | GoogleScholarGoogle Scholar |
Korschgen LJ (1959) Food habits of the red fox in Missouri. The Journal of Wildlife Management 23, 168–176.
| Food habits of the red fox in Missouri.Crossref | GoogleScholarGoogle Scholar |
Latham RM (1950) ‘The food of predaceous animals in northeastern United States.’ (Pennsylvania Game Commission: Harrisburg, PA, USA)
Lenth R, Singmann H, Love J, Buerkner P, Herve M (2018) Emmeans: estimated marginal means, aka least-squares means. R package version 1(1), 3.
Lüdecke D (2018) ggeffects: tidy data frames of marginal effects from regression models. Journal of Open Source Software 3, 772
| ggeffects: tidy data frames of marginal effects from regression models.Crossref | GoogleScholarGoogle Scholar |
McGrew JC, Blakesley CS (1982) How komondor dogs reduce sheep losses to coyotes. Journal of Range Management 35, 693–696.
| How komondor dogs reduce sheep losses to coyotes.Crossref | GoogleScholarGoogle Scholar |
McLeod R (2016) Cost of pest animals in NSW and Australia, 2013–14. eSYS Development Pty Ltd. Report prepared for the NSW Natural Resources Commission.
Moberly RL, White PCL, Harris S (2004) Mortality due to fox predation in free-range poultry flocks in Britain. The Veterinary Record 155, 48–52.
| Mortality due to fox predation in free-range poultry flocks in Britain.Crossref | GoogleScholarGoogle Scholar | 15285283PubMed |
R Core Team (2018) ‘R: a language and environment for statistical computing.’ (R Core Team: Vienna, Austria)
Rust NA, Whitehouse-Tedd KM, MacMillan DC (2013) Perceived efficacy of livestock-guarding dogs in South Africa: implications for cheetah conservation. Wildlife Society Bulletin 37, 690–697.
| Perceived efficacy of livestock-guarding dogs in South Africa: implications for cheetah conservation.Crossref | GoogleScholarGoogle Scholar |
Saunders G, Coman B, Kinnear J, Braysher M (1995) ‘Managing vertebrate pests: foxes.’ (Australian Government Publishing: Canberra, Australia)
Saunders GR, Gentle MN, Dickman CR (2010) The impacts and management of foxes Vulpes vulpes in Australia. Mammal Review 40, 181–211.
| The impacts and management of foxes Vulpes vulpes in Australia.Crossref | GoogleScholarGoogle Scholar |
Scott AB, Phalen D, Hernandez-Jover M, Singh M, Groves P, Toribio J-ALML (2018) Wildlife presence and interactions with chickens on Australian commercial chicken farms assessed by camera traps. Avian Diseases 62, 65–72.
| Wildlife presence and interactions with chickens on Australian commercial chicken farms assessed by camera traps.Crossref | GoogleScholarGoogle Scholar | 29620454PubMed |
Shivik JA (2006) Tools for the edge: what’s new for conserving carnivores. BioScience 56, 253–259.
| Tools for the edge: what’s new for conserving carnivores.Crossref | GoogleScholarGoogle Scholar |
Silverman BW (1986) ‘Density estimation for statistics and data analysis.’ (Chapman and Hall: New York, NY, USA)
Smith ME, Linnell JDC, Odden J, Swenson JE (2010) Review of methods to reduce livestock depradation: I. guardian animals. Acta Agriculturae Scandinavica, Section A – Animal Science 50, 279–290.
| Review of methods to reduce livestock depradation: I. guardian animals.Crossref | GoogleScholarGoogle Scholar |
Statham MJ, Murdoch J, Janecka J, Aubry KB, Edwards CJ, Soulsbury CD, Berry O, Wang Z, Harrison D, Pearch M, Tomsett L, Chupasko J, Sacks BN (2014) Range-wide multilocus phylogeography of the red fox reveals ancient continental divergence, minimal genomic exchange and distinct demographic histories. Molecular Ecology 23, 4813–4830.
| Range-wide multilocus phylogeography of the red fox reveals ancient continental divergence, minimal genomic exchange and distinct demographic histories.Crossref | GoogleScholarGoogle Scholar | 25212210PubMed |
Statsoft Inc (2007) ‘Statistica (data analysis software system) version 8.0.’ (Statsoft Inc: Tulsa, OK, USA)
Stobo-Wilson AM, Murphy BP, Legge SM, Caceres-Escobar H, Chapple DG, Crawford HM, Dawson SJ, Dickman CR, Doherty TS, Fleming PA, Garnett ST, Gentle M, Newsome TM, Palmer R, Rees MW, Ritchie EG, Speed J, Stuart J-M, Suarez-Castro AF, Thompson E, Tulloch A, Turpin JM, Woinarski JCZ (2022) Counting the bodies: estimating the numbers and spatial variation of Australian reptiles, birds and mammals killed by two invasive mesopredators. Diversity and Distributions
| Counting the bodies: estimating the numbers and spatial variation of Australian reptiles, birds and mammals killed by two invasive mesopredators.Crossref | GoogleScholarGoogle Scholar |
van Bommel L (2010) ‘Guardian dogs: best practice manual for the use of livestock guardian dogs.’ (Invasive Animals Cooperative Research Centre: Canberra, ACT, Australia)
van Bommel L, Johnson CN (2012) Good dog! Using livestock guardian dogs to protect livestock from predators in Australia’s extensive grazing systems. Wildllife Research 39, 220–229.
| Good dog! Using livestock guardian dogs to protect livestock from predators in Australia’s extensive grazing systems.Crossref | GoogleScholarGoogle Scholar |
van Bommel L, Johnson CN (2014) Where do livestock guardian dogs go? Movement patterns of free-ranging Maremma sheepdogs. PLoS ONE 9, e111444
| Where do livestock guardian dogs go? Movement patterns of free-ranging Maremma sheepdogs.Crossref | GoogleScholarGoogle Scholar | 25353319PubMed |
van Bommel L, Johnson CN (2015) How guardian dogs protect livestock from predators: territorial enforcement by Maremma sheepdogs. Wildlife Research 41, 662–672.
| How guardian dogs protect livestock from predators: territorial enforcement by Maremma sheepdogs.Crossref | GoogleScholarGoogle Scholar |
van Bommel L, Johnson CN (2017) Olfactory communication to protect livestock: dingo response to urine marks of livestock guardian dogs. Australian Mammalogy 39, 219–226.
| Olfactory communication to protect livestock: dingo response to urine marks of livestock guardian dogs.Crossref | GoogleScholarGoogle Scholar |
Van De Weerd HA, Keatinge R, Roderick S (2009) A review of key health-related welfare issues in organic poultry production. World’s Poultry Science Journal 65, 649–684.
| A review of key health-related welfare issues in organic poultry production.Crossref | GoogleScholarGoogle Scholar |
van Eeden LM, Eklund A, Miller JRB, et al. (2018) Carnivore conservation needs evidence-based livestock protection. PLoS Biology 16, e2005577
| Carnivore conservation needs evidence-based livestock protection.Crossref | GoogleScholarGoogle Scholar | 30226872PubMed |
Yilmaz O, Coskun F, Ertugrul M (2015) Livestock damage by carnivores and use of livestock guardian dogs for its prevention in Europe: a review. Journal of Livestock Science 6, 23–35.
Zingaro M, Salvatori V, Vielmi L, Boitani L (2018) Are the livestock guarding dogs where they are supposed to be? Applied Animal Behaviour Science 198, 89–94.
| Are the livestock guarding dogs where they are supposed to be?Crossref | GoogleScholarGoogle Scholar |