

Investigating the effect of larval instar, post-harvest treatments, and substrate on the nutritional profile of black soldier fly larvae (Hermetia illucens)
Shanmugam Alagappan A B , Hung Hong C , Deirdre Mikkelsen A C , Sandra Olarte Mantilla A B , Peter James D , Olympia Yarger E , Louwrens Hoffman A B * and Daniel Cozzolino A B *
D , Olympia Yarger E , Louwrens Hoffman A B * and Daniel Cozzolino A B *
A
B
C
D
E
Handling Editor: Konstantinos Mountzouris
Abstract
Black soldier fly larvae (BSFL) is considered to be a sustainable animal feed ingredient, that feeds in different organic waste streams. It is well established that the nutritional profile of BSFL is influenced by the substrate used. However, the impact on the overall nutritional quality of the larvae when reared on food waste streams in an industrial setting is unknown.
The objective of this study is to determine the nutritional quality and chemical composition of BSFL from two instars (fifth and sixth) reared on five different food waste streams from two different waste facilities and post-harvest treatments.
BSFL were reared with five different waste streams, viz. soy waste (S), customised bread–vegetable waste (B), supermarket waste (WL), childcare centre waste (CC) and a food waste mixture (WB); two larvae instars (fifth and sixth) and post-harvest treatments (blanching and drying) were analysed for proximate composition, and amino, fatty acid and mineral contents.
The blanching and drying treatments did not affect the chemical composition of the larvae; however, they had a negative effect on fatty acid content and reduced the concentration of certain amino acids. The mineral content of BSFL was increased by drying, whereas blanching did not result in leaching except for sodium. The larval instar played a role in influencing the crude fat and crude protein content, with a higher fatty acid content in sixth instar BSFL. Mineral and amino acid profiles were found to be influenced by both the larval instar and the food waste stream used for rearing.
This study showed the interdependent relationship between substrate, instars, and the post-harvest treatments in influencing the nutritional properties of the larvae.
The outcomes of this study will assist manufacturers in substrate selection and enable them to custom tailor their operations to arrive at a product with desired nutritional properties.
Keywords: alternative proteins, black soldier fly larvae, circular economy, composition, food waste streams, nutritional quality, post-harvest treatments.
Introduction
Global food security is threatened as the world population is projected to increase up to 11 billion people by the end of this century (Erhard et al. 2023). A recent report of the Food and Agriculture Organization (FAO) suggests that the demand for animal proteins will increase by up to 14% by the year 2030, with a high demand for poultry meat (40%) followed by pork (34%), beef (20%), and goat (5%) meat (Organisation for Economic Co-operation and Development (OECD)/FAO 2021). More than 1.1 billion megagrams of animal feed are used in the farming of livestock annually, thereby utilising a significant amount of natural resources. The changing climate characterised by increased temperatures and varying rainfall patterns negatively influences the livestock production systems in several ways across the supply chain (Cheng et al. 2022). Therefore, it is of utmost importance to develop sustainable feed ingredients to meet increased meat demand for the growing population.
The use of insects, especially black soldier fly (Hermetia illucens L., Diptera: Stratiomyidae) larvae (BSFL) as an alternative protein source for livestock rearing has increased in several regions across the globe (Bessa et al. 2020; Siva Raman et al. 2022; Beyers et al. 2023). The BSFL is widely distributed in tropical and subtropical regions. After hatching from the egg, BSFL go through six morphological stages (instars) before becoming pupae (Kaya et al. 2021; Liew et al. 2022). The larvae of BSF can consume a wide variety of organic side streams over a period of 10–52 days where the fifth instar BSFL is increasingly being used as feed ingredient in livestock production (e.g. source of protein) (Alagappan et al. 2022a, 2022b). The BSFL reared on organic side streams are rich in protein (30–50%), fat (15–49%), fibre (20.4–30.5%) and various micronutrients (Nyakeri et al. 2017; Gold et al. 2018; Gao et al. 2019; Shumo et al. 2019a, 2019b). The amino acid and fatty acid profile of BSFL is well balanced and considered to be better than that of fish and soybean meal that are prevalently used as feedstuff in livestock farming (Heuel et al. 2021). Several feeding trials have also highlighted the suitability of BSFL as an alternative protein ingredient in diets for broilers, pigs, and fish (Maranga et al. 2022; Nampijja et al. 2023; Phaengphairee et al. 2023). Overall, BSFL are considered as more viable sustainable feed ingredient than are other conventional resources such as fish meal and soybean (Bessa et al. 2020).
Food waste contributes about 6% of the total anthropogenic greenhouse-gas emissions (Amicarelli et al. 2021). The food waste arising from households, restaurants, business places and other tertiary processing wastes comprises a diverse set of organic fractions rich in various macro- and micronutrients (Isibika et al. 2023). The use of such multi-stream food waste as substrate for rearing BSFL is becoming increasingly popular in commercial production facilities owing to the saprophagous nature of BSFL. However, it is noteworthy that the nutritional attributes of such multi-stream food waste vary with demographics, seasonal changes, and other underlying factors as reported by others (Barbi et al. 2020; Montevecchi et al. 2023).
The nutritional quality of the BSFL reared on such waste streams varies with several factors including rearing conditions, physiological age of the harvested larvae, and post-harvest treatments employed (Shumo et al. 2019a, 2019b; Ravi et al. 2020a, 2020b; Ribeiro et al. 2022; Yakti et al. 2022). However, it has been well established that the substrate used for rearing BSFL plays the most critical role in influencing the nutritional quality of BSFL (Gold et al. 2018, 2020). Previous studies have evaluated and highlighted the impact of specific macronutrients (e.g. carbohydrates, lipids and protein) and waste streams on BSFL nutrition in lab-scale systems (Cammack and Tomberlin 2017; Liu et al. 2018; Barragan-Fonseca et al. 2019; Danieli et al. 2019; Ewald et al. 2020; Gold et al. 2020).
This study was designed to address the following aims (1) to investigate the different nutritional attributes (chemical composition, amino acid, fatty acid, and mineral content) of BSFL reared on five different waste streams at two industrial production facilities and (2) to evaluate the effect of larval instar and post-harvest treatments on the nutritional quality of the larvae.
Materials and methods
Preparation of waste streams
In this study, a total of five different waste streams, viz. soy waste (S), customised bread–vegetable waste (B), supermarket waste (WL), childcare centre waste (CC) and a food waste mixture (WB) were used for rearing BSFL. The experiments with homogenous soy waste (S) and customised bread–vegetable waste (B) were conducted in commercial facility ‘A’, following their production standards. The soy waste obtained from a custard manufacturer was used as feed. The B was prepared by chopping and mixing bread (60% w/w) with the following fruits and vegetables: banana, apples, peaches, carrots, and potatoes (8% each w/w) sourced from a local supermarket (Alagappan et al. 2022a). The experiments with heterogenous food waste streams were conducted in a production facility ‘B’, following their production standards. The WL waste primarily comprised bread, meat, salads, pastries, potato chips and dairy products. The CC waste was constituted mainly of salads, bread and other cooked foods from a childcare facility. The WB was essentially a mixture of supermarket waste, fast-food waste, restaurant waste, childcare waste, and food waste from local retail outlets. The received waste was removed from its packaging material by using an industrial de-packager and ground using an industrial grinder prior to feeding to the larvae (Alagappan et al. 2023). It should be noted that the rearing conditions such as temperature, humidity, and other related processing conditions are not specified owing to confidentiality agreements with the commercial production facilities. However, these parameters (humidity and temperature) were controlled to ensure the maximum growth of the larvae under commercial conditions.
Rearing of BSFL
Two kilograms of ‘S’ and ‘B’ waste were thoroughly mixed and placed into each tray (n = 6 trays/waste; dimensions 60 cm × 40 cm × 12 cm) to which approximately 10,000 5-day old (chronological age) larvae were added. The trays were subjected to rearing conditions followed at facility ‘A’ and were fed ad libitum, with fresh feed being added as required. The ‘S’ required throughout the experiments was stored at 4°C and brought to room temperature before feeding to the BSFL. A subsample of this feed (~20 g) was stored at −20°C for future analysis. B diets were prepared fresh on the day of feeding and subsamples (~10 g) were collected on each day of feeding, stored at −20°C and were later pooled together for analysis. The fifth instar BSFL were harvested with sterile tweezers from random spots in the tray, following the first sighting of sixth instar (pre-pupae) larvae. The sixth instar BSFL was harvested similarly on sighting of the first pupae (Alagappan et al. 2022a). Approximately 20 g (100 larvae) were collected for both instars.
Ground WL, CC and WB waste was placed in trays (n = 3 trays/waste, dimensions 60 cm × 40 cm × 12 cm) where 15,000 5-day-old (chronological age) BSFL were added. The trays were then subjected to rearing conditions used in Facility ‘B’ and fed ad libitum. The feed for these experiments was prepared fresh on the day of feeding, a subsample was collected, stored at −20°C, and later, subsamples were pooled together for analysis. The fifth instar BSFL were collected as mentioned in section Rearing of BSFL with homogenous waste. It is to be noted that sixth instar BSFL were not collected from these experiments.
Post-harvest treatment of BSFL
Harvested BSFL samples from the above two experiments were subjected to the three processing treatments, viz. (a) homogenisation (unprocessed), where the whole live larvae samples collected from the trays were homogenised with a food processor (Cuisinart Food Processor, Australia); (b) blanching, where the whole live larvae collected were subjected to blanching at 100°C for 5 min and homogenised using a food processor (Cuisinart Food Processor, Australia); and (c) drying, wheren the whole live larvae were blanched according to the conditions mentioned above and subjected to drying in a commercial dehydrator at 70°C for 12 h, followed by homogenisation (Alagappan et al. 2023). It should be noted that blanching and drying treatments were conducted only on fifth instar BSFL obtained from heterogenous waste streams.
Chemical composition
The different food waste streams and the resulting BSFL were subjected to chemical analysis. Proximate analyses were conducted by the analytical services unit of the School of Agriculture and Food Sustainability (AGFS), The University of Queensland (St Lucia, Brisbane, Qld, Australia). Association of Official Analytical Chemists (AOAC) Methods 960.39 and 992.15 were used to determine crude fat (CF) and crude protein (CP) (total nitrogen × 5.62) respectively. AOAC Method 2002.04 was followed for the determination of neutral detergent fibre (NDF) (Association of Official Analytical Collaboration (AOAC) 2019). The method described by Karkalas (1985) was used to determine total starch content in the samples.
Mineral analysis
The different food waste streams and the resulting BSFL were subjected to mineral analysis. These analyses were conducted by the AGFS analytical services unit (Brisbane, Qld, Australia). All samples were subjected to drying and nitric acid pre-digestion prior to heavy metal analysis. Two millilitres of concentrated nitric acid was added to 0.1 g of samples. Pre-digestion was conducted for 2 h, followed by digestion at 100°C for 2 h. The samples were then made up with 15 mL of milliQ water. The solutions were then analysed with inductively coupled plasma–optical emission spectroscopy (ICP–OES) for different heavy metals at recommended wavelengths (Dolan and Capar 2002).
Amino acid analysis
Amino acid analysis was based on a previous report by Liyanaarachchi et al. (2020), with modifications and was conducted only on BSFL samples obtained from the different waste streams. Briefly, about 0.2 g of freeze dried and homogenised sample was placed in a 15 mL screw-capped glass tube under argon atmosphere, where 5 mL of 6 M HCl containing 0.1% (v/w) of phenol was added and vortexed for 5 s. The tubes were heated at 110°C for 22 h. After hydrolysis, the sample was transferred to a new 15 mL Falcon tube filled with 10 mL with MilliQ-water. The hydrolysed sample was filtered, and an aliquote of 500 μL of the filtered solution was placed into a 1.5 mL centrifuge tube with 100 μL of Norvaline (internal standard; 100 mg/L). Contents of the tube were dried under reduced pressure at 35°C, by using a vacuum concentrator (SpeedVac SPD140DDA, Thermo Fisher Scientific, USA). The pellet was resuspended with 1 mL of MilliQ-water for derivatisation. The derivatisation of amino acids was performed using 9-fluorenylmethoxycarbonyl chloride (Fmoc-Cl) according to Kahsay et al. (2022), with some modifications. Briefly, a prepared solution of 30 μL was mixed with 0.5 M borate buffer at pH 8.6 and 15 mM Fmoc-Cl in acetonitrile (ACN) (120 μL each). The sample was vortexed at room temperature for 5 min, then 120 μL of 0.3 M diphenylamine solution was added, following by 510 μL of MQ-water. The resultant solution was washed twice with 300 μL of n-hexane. The lower layer (200 μL) was mixed with 800 μL of 3:1 borate buffer–ACN and filtered through a 0.22 μm syringe filter for high-performance liquid chromatography (HPLC) analysis. The Nexera X2 UHPLC system included a system controller (CBM-30A), two pumps (LC-30AD), an autosampler (SIL-30AC), an oven (CTO-20AC) and a degasser (DGU-20A5R). The system was coupled to a RF-20AXS detector (Shimadzu, Kyoto, Japan). Chromatographic separation was performed on a reverse-phase Acquity UPLC BEH C18 column (150 mm × 2.1 mm i.d.,1.7 particle size; Waters, Dublin, Ireland) at a flow rate of 0.2 mL/min, with a column oven temperature of 30°C. The 29 min gradient elution was adjusted as follows: time(min)/%B of 37/0.0, 38/4.0, 39/4.1, 39/10, 47/14, 55/14.5, 56/22.0, 90/23, 95/24, 100/24.5, 100/26.5, 37/26.6 and 37/29; mobile phase B (45% acetonitrile, 45% methanol 10% MQ-water) and mobile phase A (sodium phosphate 10 mM, sodium borate 10 mM, pH 8.2). A Supelco 18-component amino acid mix standard for protein hydrolysates and a fluorescence detector (FLD) were used to detect and quantify amino acids with an excitation wavelength of 254 nm and emission wavelength of 630 nm.
Fatty acid analysis
Fatty acids analysis was conducted only on BSFL samples obtained from different waste streams. The lipid extraction method followed Ryckebosch et al. (2012), with some modifications. Briefly, approximately 0.2 g of dried and homogenised sample material was mixed with 2 mL methanol for 5 s by using a vortex mixer (Fisher Biotech, Perth, WA, Australia). The samples were sonicated for 15 min at room temperature with 100 W power in an ultrasonic bath (500 TD, SONICLEAN™, Australia). Chloroform (CHCl3, 2 mL) and MQ-water (2 mL) was added to the sample and vortexed for 5 s. The sample was then centrifuged at 3100g and 15°C (Centrifuge 5810 R, Eppendorf, Germany) for 5 min. The lower layer was collected into a glass 15 mL tube with a Pasteur pipette. The remaining pellet was re-extracted twice with a chloroform and methanol mixture (1:1, v/v). The supernatant was collected, and the combined solution was dried down under nitrogen flow at room temperature. The dried lipids and free fatty acids extracted from samples were converted into their corresponding fatty acid methyl esters (FAMEs) using acetyl chloride and were analysed according to Chua et al. (2019), with some alterations. In brief, the dried extract was dissolved in 20 mL of methanol and 200 μL of the resuspended lipid solution was mixed with 1 mL of 5% acetyl chloride in methanol in a 10 mL glass tube a with screw cap lid. The samples were then incubated at 100°C for 1 h and subsequently cooled to room temperature. Then, 1 mL of 0.9% NaCl solution and 2 mL of isooctane were added and vortexed for 5 s. The mixture was then centrifuged at 3000g and 15°C (Centrifuge 5810 R, Eppendorf) for 5 min, and 950 μL of the upper layer (isooctane) was collected and transferred to a GC vial together with methyl nonadecanoate (internal standard, 50 μL, 1 mg/mL) for fatty acid analysis. A Shimadzu GCMS-2010 Plus system (Shimadzu Scientific Instruments, Sydney, NSW, Australia) was used along with an Agilent DB-23 fused silica capillary column (60 m × 0.25 mm diameter i.d, 0.15 μm film thickness; Agilent Technologies, Santa Clara, CA, USA). The carrier gas used was ultra-pure helium with a constant linear velocity of 42.7 cm/s. The injection port temperature was set at 230°C, and 1 μL of the sample was injected in split mode with a split ratio of 5. The gradient temperature program started at an isocratic of 40°C for 1 min, then increased to 170°C at 30°C/min, and finally to 230°C at 3°C/min. The ion source and interface temperatures of the mass spectrometer were set at 200°C and 230°C respectively. The analysis was performed in full-scan mode with a mass range of 45–400 m/z. The fatty acids were identified using a Supelco 37-component FAME mix standard (Sigma-Aldrich) and verified using the National Institute of Standards and Technology (NIST14) library.
Data analysis
GraphPad Prism (ver. 10.0.2; see https://www.graphpad.com/features/prism-anova) was used for the univariate data analysis (mean, standard deviation, range and analysis of variance). One-way ANOVA was conducted at P = 0.05 to test for significant difference among means for the different samples (substrate and BSFL) for the various nutritional parameters analysed. Barlett’s test was performed to determine whether the samples have equal variance, which determined the suitable post hoc test selected; Bon-Tukey’s and Dunnett’s T3 test was performed at P < 0.05 to compare the sample groups that had equal and different variances respectively. Pearson correlation analysis was conducted with GraphPad Prism 10.0.2 for all measured nutritional attributes.
Results and discussion
Chemical and mineral composition of substrate
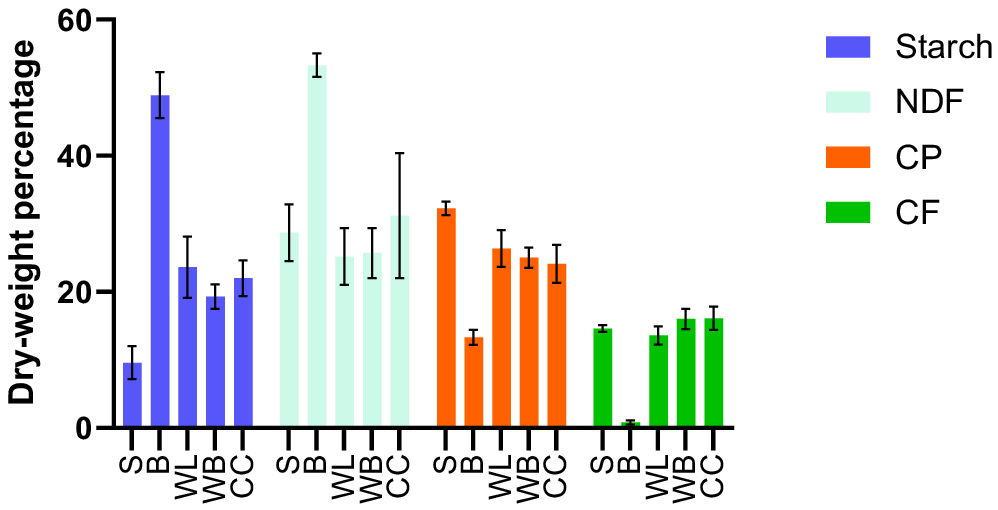
The chemical composition of the waste streams is shown in Fig. 1. The chemical compositions of the heterogenous substrates (WL, CC, and WB) were similar, with no significant differences (P > 0.05) in CP, CF, and NDF. Overall, both CP and CF contents of the heterogenous waste streams were comparable to those reported by other authors (Spranghers et al. 2017; Shumo et al. 2019a, 2019b; Barbi et al. 2020). Both CP and CF were lower, whereas starch and NDF were higher in B streams than with S diet.
Chemical composition of food waste streams used for rearing black soldier fly larvae. All values are reported as mean and standard deviation from three replicates. S, soy waste; B, bread vegetable diet; WB, waste mixture; WL, supermarket waste; CC, childcare centre waste; NDF, neutral detergent fibre; CP, crude protein; CF, crude fat.
The mineral composition of the different waste stream substrates (diets) is shown in Table 1. Calcium (Ca) was the dominant element, followed by potassium (K), sodium (Na), phosphorus (P), sulfur (S), and magnesium (Mg). The concentration of the above elements along with boron (B) and barium (Ba) (except Na) was higher (P < 0.05) in the S diet than in the B diet. No statistical differences were observed for selenium (Se) in both B and S diets. The mineral content of the B diet was similar to those reported by other researchers using vegetables and fruits as source of waste (Fischer and Romano 2021; Hashmi et al. 2021). However, the content of the S diet was relatively low compared with that in other reported studies using soybean as source of waste (Li et al. 2013; Rahman et al. 2021). No significant (P > 0.05) differences were found for Ca, K, B, and Se in samples from the heterogenous waste streams (WL, WB, and CC). The concentration of S was lower (P < 0.05), and Ba was higher (P < 0.05) in CC diet than in WB and WL diets. Differences in these elements can be explained by the ingredients in the waste streams analysed. The CC included salads, bread, and other cooked foods, whereas both WB and WL mainly consisted of meat, leftover foods, pastries, and other dairy products. Mg content also differed (P < 0.05) between CC and WB diets. Similar trends were observed between CC and WB diets for Na and P. Overall, the mineral content in all the diets agreed with those reported by other authors using similar food-waste streams (Shumo et al. 2019a, 2019b; Fitriana et al. 2022).
| Item | B (mg/kg) | Ca (mg/kg) | K (mg/kg) | Mg (mg/kg) | Na (mg/kg) | P (mg/kg) | S (mg/kg) | Ba (mg/kg) | Se (mg/kg) | |
|---|---|---|---|---|---|---|---|---|---|---|
| S | 17.7 ± 2.9a | 725.5 ± 44b | 3354.6 ± 189a | 461.7 ± 26.8a | 62 ± 3.1c | 1098.4 ± 67.2ab | 705.5 ± 30.2b | 1.6 ± 0.2b | 0.2 ± 0.3b | |
| B | 2.4 ± 1.2c | 541.6 ± 38.7c | 1851.8 ± 339.4b | 261.9 ± 22.6b | 2469.6 ± 271.9a | 675.8 ± 49.4c | 829.5 ± 49.9ab | 0.5 ± 0.1c | 0.3 ± 0.1b | |
| WL | 6.3 ± 1.1b | 3312.3 ± 186.3a | 2813.2 ± 75.7a | 308.4 ± 8.2bc | 2106.4 ± 124.5ab | 1136.7 ± 67.5ab | 934.8 ± 9.7a | 2.8 ± 0.3a | 1 ± 0.5ab | |
| WB | 4.4 ± 0.2bc | 3530.1 ± 109.6a | 2858.5 ± 55.7a | 301.4 ± 2.3cd | 2377.2 ± 97.7a | 1215.7 ± 34a | 959 ± 109.1a | 2.7 ± 0.5a | 1.3 ± 0.4a | |
| CC | 3.2 ± 0.4bc | 2346.7 ± 527.1abc | 2011 ± 97c | 239.4 ± 16.4d | 1712.5 ± 71b | 1062.2 ± 18.5b | 765.2 ± 87.1b | 1.4 ± 0.3b | 1.6 ± 1a |
All values are reported as mean and standard deviation from three replicates. Values in a column followed by the same letter are not significantly different (at P = 0.05).
S, soy waste; B, bread vegetable diet; WB, waste mixture; WL, supermarket waste; CC, childcare centre waste.
Effect of larval instar on nutritional quality
The chemical composition of the BSFL samples obtained from the different waste streams and diets at two different instars is reported in Fig. 2. NDF content was higher in sixth instar BSFL and was found to be significant (P < 0.05) for the fifth and sixth instar BSFL from B diets. No statistical differences were observed for NDF content in fifth and sixth larvae from S diets. The age of the BSFL did not influence the CP in the samples reared on the S diet. However, significant (P < 0.05) differences were observed for CF content in fifth and sixth instar BSFL fed with S. Previous research has shown that different agro-industry by-products with composition similar to that of the S diet have resulted in BSFL with reduced CP and higher CF contents (Gold et al. 2018, 2020; Siddiqui et al. 2022). It has also been reported that the CP of BSFL on reaching a certain threshold does not differ, even with changes in the CP content of the substrate (Barragán-Fonseca et al. 2018, 2019; Eggink et al. 2023). However, in our studies, CP values for sixth instar BSFL from S were found to be lower than those for the fifth instar. CF content in the sixth instar larvae has been reported to decrease during pupation. However, in our case, the CF values of sixth instar BSFL reared on S were found to be higher than those of the fifth instar larvae. The plausible reason for this discrepancy observed with CF and CP is that the larvae in this study from SW could have been harvested slightly before their actual intended life stage. Liu et al. (2017) reported that the CP and CF content of BSFL increased during the early stages of larval development.
Effect of larval instar on chemical composition of black soldier fly larvae from homogenous waste streams. All values are reported as mean and standard deviation from three replicates. 5IS and 6IS, fifth and sixth instar larvae from soy waste; 5IB and 6IB, fifth and sixth instar frass bread–vegetable diets; NDF, neutral detergent fibre; CP, crude protein; CF, crude fat.
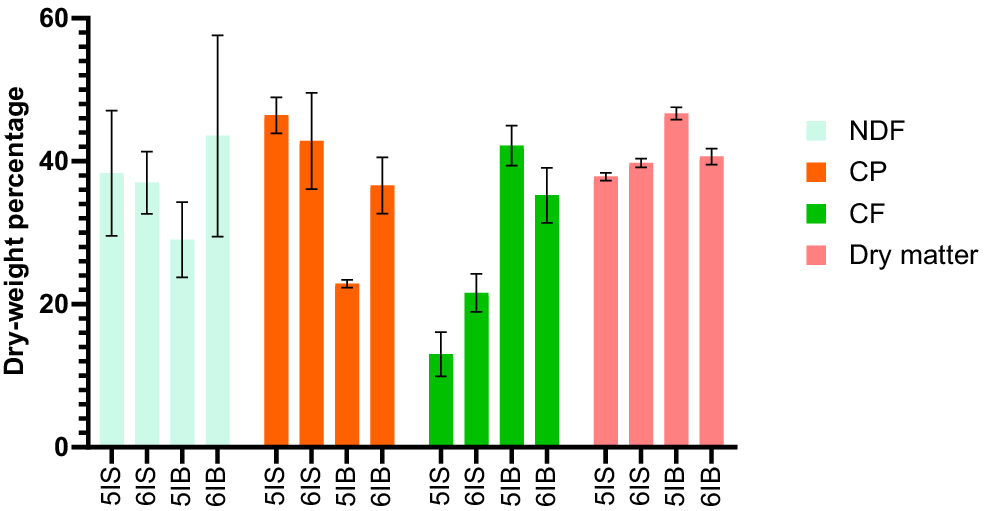
In case of the BSFL samples from the B diet, the CP of the sixth instar BSFL was higher (P < 0.05) than that of the fifth instar larvae. The increase in CP content observed for the sixth instar BSFL from B diet could be attributed to the increased distribution of chitin in the exoskeleton of BSFL pre-pupae (Rampure et al. 2023). The CF content, as stated earlier, was observed to decrease owing to the utilisation of fat stores by the pre-pupae.
The mineral compositions of BSFL reared from homogenous and heterogeneous wastes were similar, as reported by other authors (Shumo et al. 2019a, 2019b; Romano et al. 2023; Scieuzo et al. 2023) (Tables 2, 4). The fifth and sixth instar BSFL reared on the B diet did not differ in Ca and K (P > 0.05). No significant (P > 0.05) differences in the P and S content of the fifth and sixth instar BSFL reared on S diet were observed. Smets et al. (2020) reported that the concentration of Ca, K, Mg, P and Na can differ between the fifth and sixth instars. Similar trends were observed in this study; however, only for B and Mg. Minerals such as Na, P, S, Ba, and Se were not different (P > 0.05) despite the variations observed in the fifth and sixth instar BSFL grown on S diet. Similarly, minerals including K and Ca were not different (P < 0.05) for fifth and sixth instar BSFL from the B diets. It is noteworthy that Ca was found to be distributed at higher concentrations in the sixth instar BSFL from both S and B diets. This can be explained by the deposition of Ca in the cuticle at the later stage in the life cycle of the larvae (Do et al. 2021). Spranghers et al. (2017) also reported that the concentration of minerals in sixth instar BSFL reared with chicken feed, vegetable and restaurant waste remained unaltered irrespective of the substrate used. However, in this study, the concentration of most of the minerals evaluated varied in the sixth instar BSFL reared from the two waste streams. These results indicated that the waste substrate plays a greater role in determining the variation in mineral concentration than does the age of the larvae.
| Larval stage | B (mg/kg) | Ca (mg/kg) | K (mg/kg) | Mg (mg/kg) | Na (mg/kg) | P (mg/kg) | S (mg/kg) | Ba (mg/kg) | Se (mg/kg) | |
|---|---|---|---|---|---|---|---|---|---|---|
| 5IS | 11.5 ± 0.5a | 2946.4 ± 254.7c | 4666.6 ± 240.9a | 911.7 ± 67b | 188.1 ± 6.1c | 2902.6 ± 126.6ab | 1590.8 ± 48.8c | 1.1 ± 0.2c | 0.2 ± 0.1b | |
| 6IS | 7.5 ± 0.6b | 3678.7 ± 235a | 3523.4 ± 173.3b | 1254.7 ± 83.2a | 191.8 ± 9.9c | 3086.7 ± 202.5a | 1589.3 ± 73.8c | 1.1 ± 0.1c | 0.2 ± 0.2b | |
| 5IB | 1.3 ± 0.2c | 1842 ± 163.1b | 3635.6 ± 147.5b | 732 ± 74.3c | 1154.5 ± 80.5a | 1908.3 ± 147.6c | 1094 ± 44g | 1.5 ± 0.2a | 0.3 ± 0.2b | |
| 6IB | 0.7 ± 0.1d | 3233.6 ± 895.1abc | 3578.1 ± 218.9b | 1267.9 ± 92.7a | 653.8 ± 98.9b | 2694.9 ± 163b | 1285.9 ± 64.1de | 2.3 ± 0.3b | 0.4 ± 0.1a |
All values are reported as mean and standard deviation from three replicates. Values within a column followed by the same letter are not signficantly different (at P = 0.05).
5IS and 6IS, fifth and sixth instar larvae from soy waste; 5IB and 6IB, fifth and sixth instar frass bread–vegetable diets.
Amino acid composition
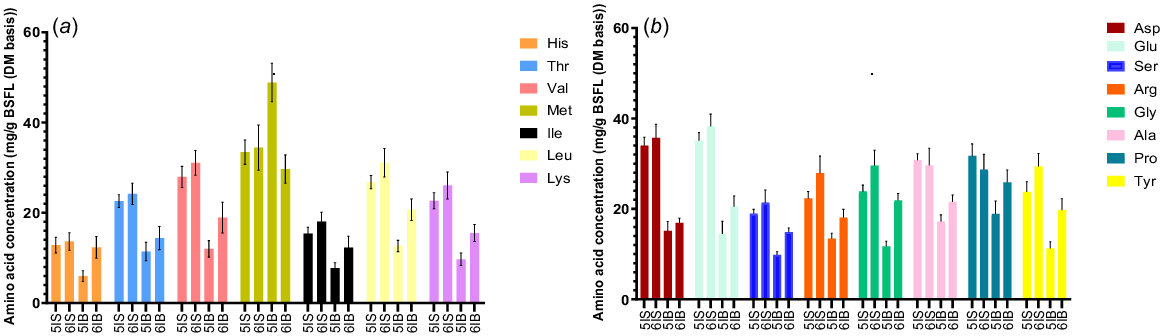
In the case of the amino acids (AA), the concentration of Lys was low whereas that of Met was high in all the samples (Fig. 3). This discrepancy in the concentration of these AA with those reported by other authors could be attributed to the differences in the geographical strain of BSFL employed in this study (Makkar et al. 2014). Similar trends were reported by other authors for Cys, Met, and Thr in the different BSFL samples reared in conventional soybean and fish meal as ingredients in animal feeds (Siddiqui et al. 2022). The AA composition of BSFL samples from the B diet were found to be relatively lower than that of samples from the S diet. It has been reported that the AA profile of BSFL varies on the basis of the substrate composition; hence, this could be attributed to differences in CP in the substrate (Ottoboni et al. 2018; Lalander et al. 2019). The amount of limiting AA (LAA) was higher (P < 0.05) for most of the AA in the sixth instar BSFL than in fifth instar larvae from the B diet except for Thr (no difference) and Met (lower in sixth instar). In samples from the S diet, Leu and Lys were the only LAA found to be at higher (P < 0.05) concentrations in the sixth instar BSFL. The range of non-limiting AA (NLAA) including Asp, Glu, Ser, Pro, and Ala did not differ (P > 0.05) among the two larval instars from the S diet. In samples from the B diet, the concentration of all NLAA except Asp was significantly higher (P < 0.05) in the sixth instar BSFL. Overall, it was observed that the AA content (except Met) in pre-pupae was higher than of the larvae. Smets et al. (2020) reported that the concentration of certain AAs increased in pre-pupae, which is in accordance with our findings. Overall, these results suggested that the amino acid content of the BSFL might be influenced by the larval instar and the substrate (waste).
Effect of larval instars on amino acid composition (mg/g DM of BSFL) of black soldier fly larvae from homogenous waste streams. (a) Concentration of limiting amino acids in fifth and sixth instar BSFL. (b) Concentration of non-limiting amino acids in fifth and sixth instar BSFL. All values are reported as mean and standard deviation from three replicates. 5IS and 6IS, fifth and sixth instar larvae from soy waste; 5IB and 6IB, fifth and sixth instar from bread–vegetable diets.
Fatty acid composition
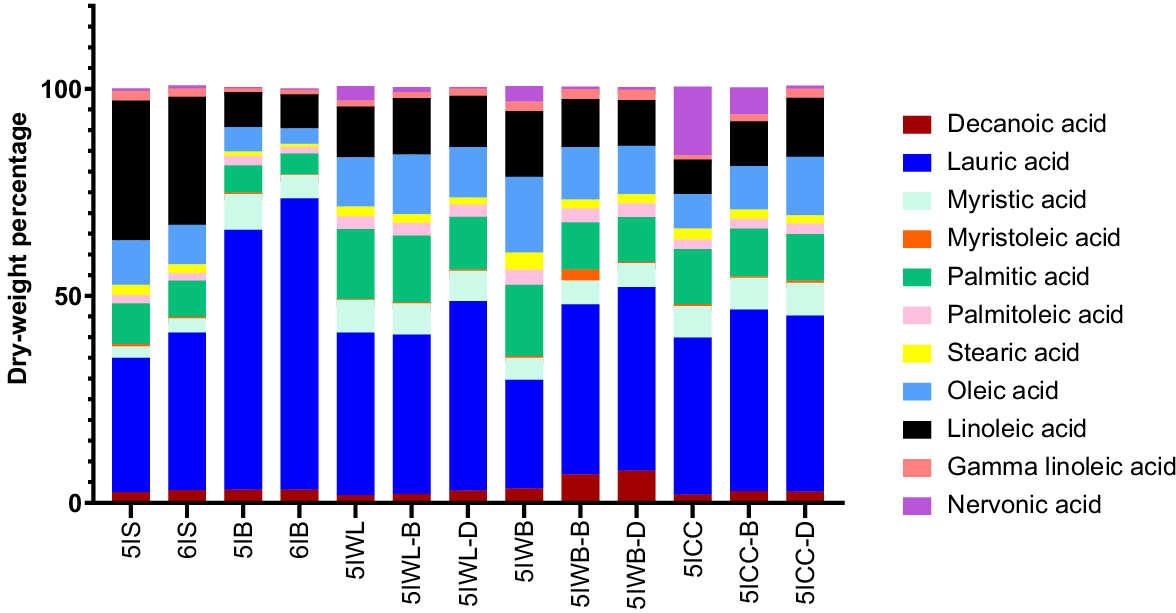
The relative distribution of fatty acids (FAs) in the BSFL samples is reported in Fig. 4. Saturated FAs (SFAs) were found to be predominant in BSFL samples reared on different waste streams. This was followed by poly-unsaturated (PUFA) and mono-unsaturated FAs (MUFA). Among the SFAs, lauric acid was found to be predominant (26.2–71.3%), followed by palmitic acid (4.9–16.9%) and myristic acid (0.8–4.1%). This trend was similar to that in previous findings reported by other authors (Liland et al. 2017; Meneguz et al. 2018; Ewald et al. 2020). BSFL is reported to synthesise lauric acid and other saturated fatty acids from carbohydrates present in the substrate (Liu et al. 2017; Spranghers et al. 2017). The same trend was observed in this study, where small differences were found for the concentration of other SFAs for the BSFL sourced from the different substrates. The distribution of PUFAs vary among the homogenised fifth instar BSFL from the different waste sources. This is because BSFL like many other species of insects, do not have the enzyme precursor needed for de novo synthesis of the unsaturated fatty acids (UFAs), and therefore their composition is highly influenced by the compositional properties of the substrate (Ewald et al. 2020).
Relative distribution of fatty acids in black soldier fly larvae reared with different food waste streams. All values are reported as mean and standard deviation from three replicates. 5IS and 6IS, fifth and sixth instar larvae from soy waste; 5IB and 6IB, fifth and sixth instar frass bread–vegetable diets; 5IWB, 5IWB-B, 5IWB-D, fifth instar larvae from waste mixture subjected to homogenisation, blanching, and drying respectively; 5IWL, 5IWL-B, 5IWL-D, fifth instar larvae from supermarket waste subjected to homogenisation, blanching and drying respectively; 5ICC, fifth instar larvae from childcare centre waste subjected to homogenisation, blanching and drying respectively.
The FA composition in the BSFL from the S and B diets is reported in Table 3. It was observed that the distribution of MUFAs such as lauric acid, myristic acid, linoleic acid and palmitoleic did not differ (P > 0.05) among the instars, but varied (P < 0.05) between S and B diets, and was relatively higher in BSFL than in B diet. Ewald et al. (2020) observed that BSFL fed with bread waste had higher lauric acid than did the other BSFL samples. This is attributed to the larvae’s ability to synthesise lauric acid from the carbohydrate content present in the substrate (bread), as stated earlier. Fatty acids including myristic acid, stearic acid, oleic acid, gamma linoleic acid and nervonic acid did not differ (P > 0.05) between the fifth and sixth instar BSFL grown on both S and B diets. The distribution of palmitic acid was higher (P < 0.05) in the sixth instar BSFL, but only for larvae reared on the S diet. Overall, the fatty acid content of BSFL was distributed at relatively higher concentrations in the sixth instar BSFL than in the fifth instar larvae. This is because BSF in their adult stages rely on their larval fat reserves for survival and other metabolic functions (Liu et al. 2017; Siddiqui et al. 2022).
| Larval stage | Decanoic acid (mg/kg) | Lauric acid (mg/kg) | Myristic acid (mg/kg) | Myristoleic acid (mg/kg) | Palmitic acid (mg/kg) | Palmitoleic acid (mg/kg) | Stearic acid (mg/kg) | Oleic acid (mg/kg) | Linoleic acid (mg/kg) | Gamma linoleic acid (mg/kg) | Nervonic acid (mg/kg) | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 5IS | 1.3 ± 0.3b | 17.1 ± 4.4b | 1.4 ± 0.9b | 0.3 ± 0.1a | 5 ± 2.5b | 1 ± 0.3b | 1.2 ± 0.4a | 5.5 ± 2.8a | 18.8 ± 9.9ab | 1.2 ± 0.6a | 0.3 ± 0.2a | |
| 6IS | 1.9 ± 0.5b | 24.4 ± 5.4b | 2.1 ± 1b | 0.2 ± 0.1a | 5.5 ± 1.9a | 1.1 ± 0.3b | 1.3 ± 0.4a | 5.9 ± 1.8ab | 20 ± 5.2a | 1.3 ± 0.6a | 0.4 ± 0.3a | |
| 5IB | 4.3 ± 1.2a | 86.1 ± 24.5a | 11.5 ± 3.3a | 0.5 ± 0.3a | 8.9 ± 2a | 3.1 ± 0.9a | 1.3 ± 0.2a | 7.9 ± 1.8ab | 11.4 ± 3.3b | 1.1 ± 0.4a | 0.3 ± 0.2a | |
| 6IB | 4.3 ± 1.2a | 98.2 ± 22.1a | 7.8 ± 2.5a | 0.2 ± 0.2a | 6.7 ± 2.2a | 2.1 ± 0.6a | 1.1 ± 0.4a | 5.1 ± 1.5b | 11.3 ± 3.1b | 1.4 ± 0.8a | 0.5 ± 0.3a |
All values are reported as mean and standard deviation from three replicates. Values in a column followed by the same letter are not significantly different (at P = 0.05).
5IS and 6IS, fifth and sixth instar larvae from soy waste; 5IB and 6IB, fifth and sixth instar frass bread–vegetable diets.
Effect of post-harvest treatments on nutritional quality
Ravi et al. (2020a, 2020b) observed that blanching and drying modifies the CP content in BSFL. However, in this study, the CP content of the BSFL samples reared on three different heterogenous waste streams did not differ (P > 0.05) from each other after the post-harvest treatments. This could be attributed to the drying conditions used. Zulkifli et al. (2022) dried BSFL samples with a conventional drying oven at different temperatures and time, which influenced the CP, CF, and crude fibre content of the BSFL samples. The CP content reported for the BSFL in these heterogenous trials (Fig. 5) is in accordance with other previously reported values. The CF content was found to be significantly (P < 0.05) lower for blanched samples in comparison with dried and unprocessed BSFL from WM. The CF content of these samples ranged between 14.7 and 39.4 dry weight percentage, with the lowest value being observed for blanched BSFL sample from WM waste stream. Unlike CP, CF content in BSFL samples varied greatly with changes in the carbohydrate content. Studies have shown the linear relationship between the CF content of BSFL and the carbohydrate content of the rearing substrate (Barragán-Fonseca et al. 2018, 2019; Eggink et al. 2023). Consequently, this observed discrepancy could be attributed to the variation observed in the CP to carbohydrate ratio of substrates used in our studies. The trends in results for NDF content were similar to that of CP content. The NDF values reported for BSFL in these trials are comparable to those of BSFL samples reared on similar diets (Shumo et al. 2019b). Overall, the carbohydrate content of BSFL from these trials are comparable to values reported in previous studies (Lu et al. 2022). Post-harvest treatments such as blanching and drying conducted in these studies did not significantly affect the CP, and fibre contents of BSFL.
Effect of post-harvest treatments on chemical composition of black soldier fly larvae from heterogenous waste streams. All values are reported as mean and standard deviation from three replicates. 5IWB, 5IWB-B, 5IWB-D, fifth instar larvae from waste mixture subjected to homogenisation, blanching, and drying respectively; 5IWL, 5IWL-B, 5IWL-D, fifth instar larvae from supermarket waste subjected to homogenisation, blanching and drying respectively; 5ICC, fifth instar larvae from childcare centre waste subjected to homogenisation, blanching and drying respectively; NDF, neutral detergent fibre; CP, crude protein; CF, crude fat.
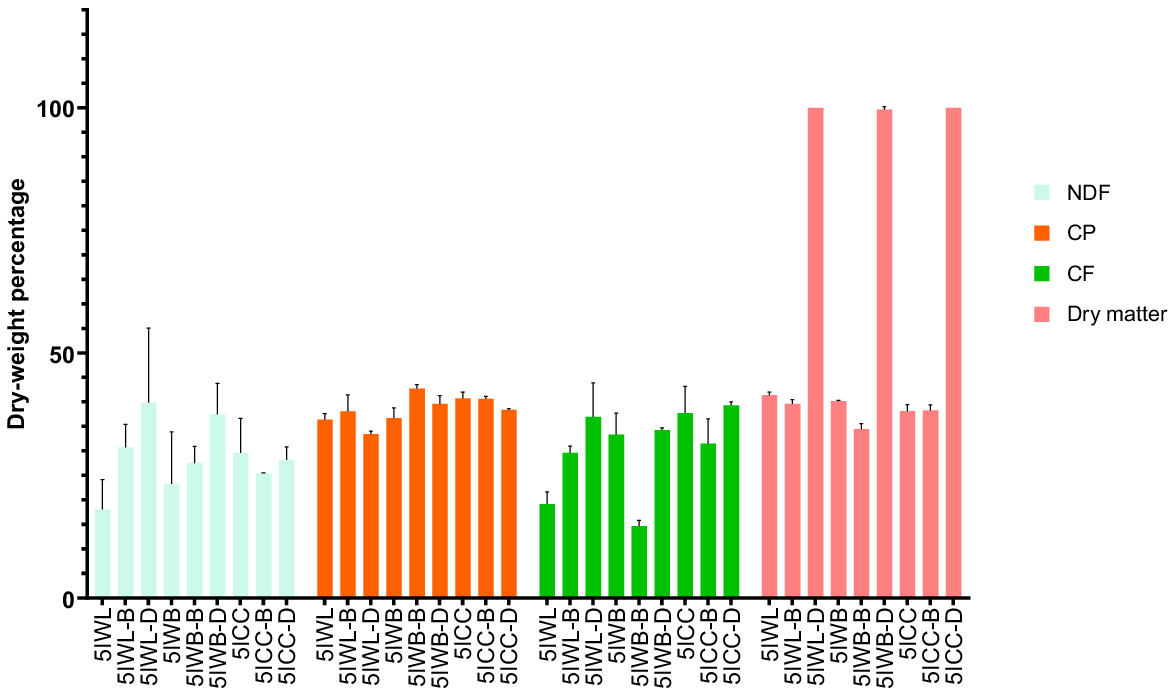
Calcium was found to be present at the highest concentration in the BSFL samples from heterogenous trials, followed by K, P, Mg, S, Na, Ba, B, and Se (Table 4). The order of distribution of minerals observed is comparable to that in previously published studies using a diverse sets of heterogeneous waste streams (Lu et al. 2022; Shah et al. 2022; Zulkifli et al. 2022). The concentration of B in unprocessed, blanched, and dried BSFL samples within the same waste streams did not differ (P > 0.05). Both Ca and P distribution in unprocessed and blanched BSFL from the same waste streams was found to be unaffected (P > 0.05). In the case of drying, Ca and P composition was found to be significantly higher (P < 0.05) than in unprocessed and blanched BSFL samples within the same waste streams. It is also noteworthy that Ca and P concentrations for dried BSFL from the different waste streams did not differ (P > 0.05). The concentrations reported in these experiments for Ca and P are similar to those of the values reported for BSFL samples reared on food waste streams subjected to similar processing conditions (Fitriana et al. 2022; Opoku et al. 2023).
| Item | B (mg/kg) | Ca (mg/kg) | K (mg/kg) | Mg (mg/kg) | Na (mg/kg) | P (mg/kg) | S (mg/kg) | Ba (mg/kg) | Se (mg/kg) | |
|---|---|---|---|---|---|---|---|---|---|---|
| 5IWL | 8.6 ± 1.4abd | 11765.8 ± 695c | 3968.4 ± 172.9bdg | 917.8 ± 44b | 596.1 ± 58.6abc | 2218.9 ± 48.7d | 1354.9 ± 32.2b | 4.4 ± 0.9bcd | 2.1 ± 0.7ab | |
| 5IWL-B | 8.6 ± 0.3ad | 11481.3 ± 610.2c | 3210 ± 94.5bdg | 860.6 ± 35.6b | 196.2 ± 29.1e | 1977.4 ± 65.4d | 1174.7 ± 31.4c | 3.4 ± 0.5bcd | 3 ± 1.1ab | |
| 5IWL-D | 7.6 ± 0.5ab | 30959.7 ± 5101.8abcd | 9946 ± 164.8ac | 2203.1 ± 102.7a | 564.2 ± 139.7abcde | 5326.3 ± 81.4c | 3067.8 ± 53.5a | 11.8 ± 1.5a | 2.3 ± 0.8ab | |
| 5IWB | 2.8 ± 0.6cd | 11926.4 ± 71.5ac | 3168 ± 100.3df | 807.1 ± 28.9b | 515.3 ± 63.8abcd | 1877.8 ± 53.6d | 1176.3 ± 22c | 1.5 ± 0.4d | 1.3 ± 0.4b | |
| 5IWB-B | 2.4 ± 0.7cd | 11297.9 ± 292.8cd | 2849.3 ± 97.9df | 816 ± 11.2b | 212.1 ± 5.9de | 1778.8 ± 43.2d | 1124.5 ± 40.3c | 1.4 ± 0.4d | 2.1 ± 1.1ab | |
| 5IWB-D | 3.1 ± 0.7bcd | 33513.2 ± 415.2a | 10186.8 ± 320.9ce | 2551.7 ± 99.2a | 750.4 ± 36.6a | 5649 ± 208.8b | 3419.4 ± 93.5a | 5.1 ± 0.3abc | 2.6 ± 0.2a | |
| 5ICC | 3.1 ± 1d | 9378.5 ± 892.9cd | 4031.9 ± 10.9g | 938.5 ± 40.2b | 490.1 ± 37.5b | 2633.8 ± 47.8d | 1262.2 ± 58.8bc | 3 ± 0.8cd | 1.5 ± 0.6ab | |
| 5ICC-B | 2 ± 0.3cd | 9383.6 ± 207.9c | 3421.3 ± 238.1fg | 878.6 ± 18b | 234.6 ± 18cde | 2407.9 ± 73.5d | 1199.3 ± 16.3c | 2.4 ± 0.7cd | 1.8 ± 1.2ab | |
| 5ICC-D | 2.4 ± 0.1cd | 24773.1 ± 627.3b | 11110.8 ± 230.9e | 2376.2 ± 13.9a | 763.3 ± 25.3a | 6758.5 ± 106.1a | 3299.1 ± 45.2a | 7.9 ± 1.1ab | 1.9 ± 1.3ab |
All values are reported as mean and standard deviation from three replicates. Values in a column followed by the same letter are not significantly different (at P = 0.05).
5IWB, 5IWB-B, 5IWB-D, fifth instar larvae from waste mixture subjected to homogenisation, blanching, and drying respectively; 5IWL, 5IWL-B, 5IWL-D, fifth instar larvae from supermarket waste subjected to homogenisation, blanching, and drying respectively; 5ICC, fifth instar larvae from childcare centre waste subjected to homogenisation, blanching and drying respectively.
The concentration of S and K did not differ significantly (P > 0.05) in unprocessed and blanched BSFL from the same waste streams. The K and S concentrations in blanched BSFL from all three waste streams were also similar (P > 0.05). Dried BSFL had significantly (P < 0.05) higher concentrations of K and S than did the blanched and unprocessed samples. However, it is noteworthy that the concentrations of K and S in dried BSFL from the different waste streams were not statistically different (P > 0.05). The levels of Se did not differ (P > 0.05) in any of the BSFL samples from the heterogenous trials. The distribution of Mg and Ba also did not differ (P > 0.05) between unprocessed and blanched BSFL samples from the three waste streams. Similar to K and S, the concentration of Mg and Ba in dried and blanched BSFL from the three different waste streams were similar (P > 0.05).
Blanching is commonly observed to leach out mineral content in food samples. However, all the minerals analysed in this study, except for Na, did not leach at statistically significant levels (P > 0.05) on blanching. The concentrations of minerals reported in these studies are in accordance with the values reported for BSFL samples reared in similar waste streams (Shumo et al. 2019a, 2019b; Ebeneezar et al. 2021; Zulkifli et al. 2022). Overall, blanching of BSFL samples according to the guidelines of International Platform of Insects for Food and Feed (IPIFF) (2022) did not significantly alter the retention capacity of various minerals considered essential in feedstock diets. Also, the drying resulted in an increase in concentration for certain minerals, including Ca, P, S, Mg, K, and Na. This can be attributed to the increase in ash content observed as the result of drying, which in turn is associated with the mineral content of the samples. Son et al. (2023) reported a similar trend in BSFL samples on blanching and drying treatments.
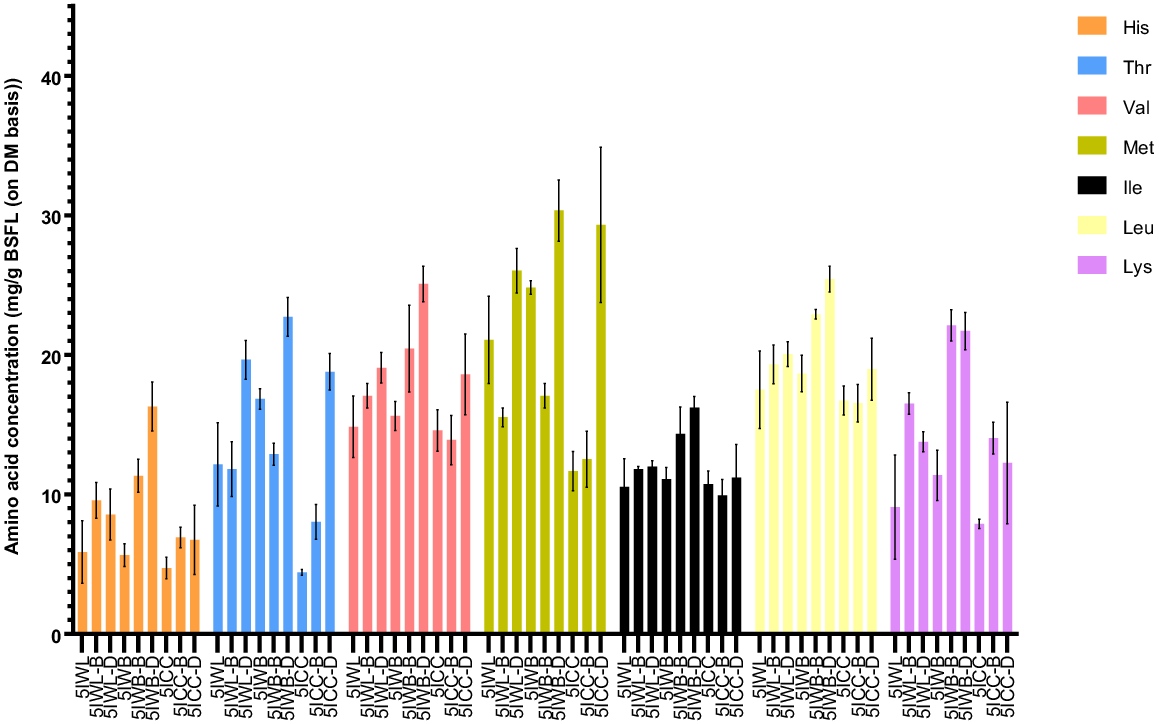
The distribution of different amino acids, including His, Met, Val, Ile, Leu, Ala, Pro, Arg, Gly and Tyr, in unprocessed BSFL from different waste streams did not differ signficantly (P > 0.05). This can be attributed to the similar AA composition for these amino acids in the diets used for rearing. Shumo et al. (2019a, 2019b) observed similar trends when rearing BSFL on different food waste streams with similar composition.
The concentrations of the LAA including Val, Thr, Ile, and Leu did not differ significantly (P > 0.05) (Fig. 6) for the unprocessed, blanched, and dried BSFL samples from the same waste streams, except for the dried samples from WB. His and Lys exhibited similar trends, but both dried and blanched samples from WB streams had higher concentrations of these AA than did unprocessed BSFL from the same waste stream. Methionine concentrationss of the unprocessed, blanched, and dried BSFL from the WB waste groups differed significantly (P < 0.05) from each other. Also, blanching was observed to reduce the methionine content of BSFL samples from the WL stream. In CC, the methionine concentration was not found to differ significantly (P > 0.05) across the samples. Son et al. (2023) subjected BSFL to two different processing treatments and observed similar trends in the results for the limiting amino acids.
Effect of post-harvest treatments on composition of limiting amino acids (mg/g DM of BSFL) in black soldier fly larvae from heterogenous waste streams. All values are reported as mean and standard deviation from three replicates. 5IWB, 5IWB-B, 5IWB-D, fifth instar larvae from waste mixture subjected to homogenisation, blanching, and drying respectively; 5IWL, 5IWL-B, 5IWL-D, fifth instar larvae from supermarket waste subjected to homogenisation, blanching and drying respectively; 5ICC, fifth instar larvae from childcare centre waste subjected to homogenisation, blanching and drying respectively.
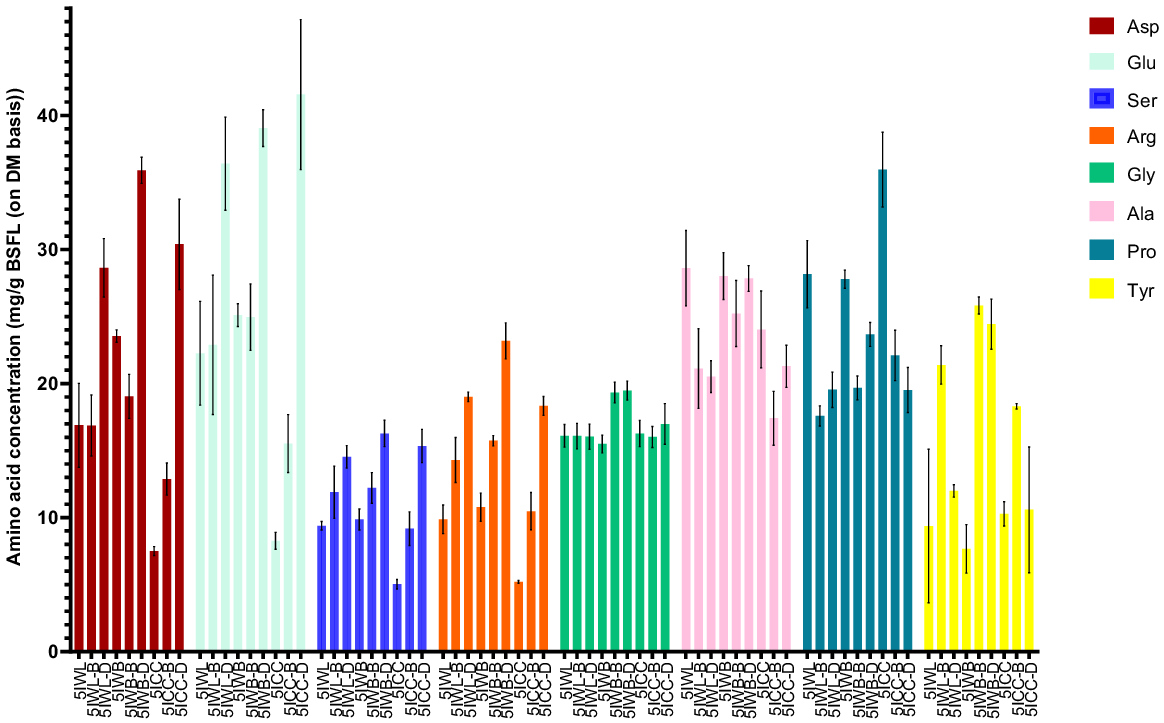
Drying resulted in an increase (P < 0.05) in the concentration of non-limiting amino acids (Fig. 7) including Asp, Glu, Ser, Arg, Gly, and Tyr compared with that of the unprocessed, blanched BSFL samples within the same waste stream. However, it is noteworthy that no difference (P > 0.05) was observed for the blanched and dried samples from specific waste streams for Asp from CC, Ser and Arg from WL, Gly from CC waste streams and, Tyr from WB and CC streams. Proline concentrations in unprocessed BSFL samples derived from the WL and CC streams were higher (P < 0.05) than in blanched samples but were unaffected by drying (except for CC). Blanching and drying treatment did not affect (P > 0.05) the concertation of proline in BSFL samples from the same group. Alanine distribution in the unprocessed, blanched, and dried BSFL samples was found to be unaffected within same streams (P > 0.05).
Impact of post-harvest treatments on composition of non-limiting amino acids (mg/g DM of BSFL) in black soldier fly larvae from heterogenous waste streams. All values are reported as mean and standard deviation from three replicates. 5IWB, 5IWB-B, 5IWB-D, fifth instar larvae from waste mixture subjected to homogenisation, blanching, and drying respectively; 5IWL, 5IWL-B, 5IWL-D, fifth instar larvae from supermarket waste subjected to homogenisation, blanching and drying respectively; 5ICC, fifth instar larvae from childcare centre waste subjected to homogenisation, blanching and drying respectively.
Son et al. (2023) observed that blanching accompanied by microwave drying and hot air drying altered only certain AA constituents in BSFL. Similarly, Zulkifli et al. (2022) subjected BSFL to different oven-drying and spray-drying conditions and observed a similar pattern in the results for amino acid composition. This trend, observed in previous studies and our studies, suggests that the AA profile of BSFL is influenced jointly by the type of AA, substrate and the processing treatment followed.
Decanoic and myristic acid contents of BSFL subjected to blanching and drying did not differ (P > 0.05) (Table 5) from that of unprocessed BSFL samples obtained from the same waste stream. Similar results were obtained for myristic, palmitoleic, linoleic and gamma linoleic acid, with one exception. The dried BSFL samples for fatty acids were found to be lower (P < 0.05) than the unprocessed BSFL from the WB diet. Lee et al. (2023) observed that drying treatments improved the composition of palmitic and stearic acid. Zulkifli et al. (2022) observed that the fatty acid composition in the BSFL samples was found to be affected by the different drying techniques employed. Therefore, it can be inferred that the deviations observed in our results can likely be attributed to the differences in the drying techniques and conditions followed.
| Item | Decanoic acid (mg/kg) | Lauric acid (mg/kg) | Myristic acid (mg/kg) | Myristoleic acid (mg/kg) | Palmitic acid (mg/kg) | Palmitoleic acid (mg/kg) | Stearic acid (mg/kg) | Oleic acid (mg/kg) | Linoleic acid (mg/kg) | Gamma linoleic acid (mg/kg) | Nervonic acid (mg/kg) | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 5IWL | 5.8 ± 0.3bc | 120.8 ± 3.2a | 24 ± 1.2bc | 0.6 ± 0.2ab | 51.6 ± 6a | 9.3 ± 0.9a | 6.8 ± 0.8b | 36.5 ± 3.4ab | 37.7 ± 3.5a | 4 ± 0.2a | 10.6 ± 0.6 a | |
| 5IWL-B | 6.7 ± 0.6ab | 117.1 ± 8.8ab | 23 ± 2.8ab | 0.7 ± 0.2ab | 48.6 ± 4.6a | 9.4 ± 1.3a | 6.3 ± 0.6b | 43.9 ± 3.1a | 41.4 ± 6.7ab | 4.4 ± 0.9abc | 3.5 ± 1.3c | |
| 5IWL-D | 5.4 ± 0.4bc | 83.4 ± 6.8bc | 13.3 ± 1.7bcd | 0.5 ± 0.1ab | 23.3 ± 3.3b | 5.2 ± 1.5ab | 3.1 ± 0.4cd | 22.2 ± 2.8bc | 22.5 ± 4.4ab | 3 ± 0.3ab | 0.7 ± 0.1c | |
| 5IWB | 7.8 ± 0.1a | 59.1 ± 2.7cd | 11.9 ± 0.4bc | 0.9 ± 0.1a | 38.9 ± 1.8a | 8.2 ± 0.4a | 9.2 ± 0.5a | 41.2 ± 3.2a | 35.9 ± 2.8a | 5 ± 0.5a | 8.2 ± 0.7ab | |
| 5IWB-B | 8 ± 2a | 47.9 ± 17.5de | 6.7 ± 3.3cd | 0.3 ± 0.4ab | 13.3 ± 7.2b | 3.9 ± 2ab | 2.4 ± 0.9cd | 14.8 ± 8.3bc | 13.5 ± 6.8ab | 2.7 ± 0.8abc | 0.7 ± 0.3c | |
| 5IWB-D | 6.8 ± 0.6ab | 39 ± 2.9e | 5.1 ± 0.4cd | 0.2 ± 0.2ab | 9.5 ± 0.4b | 2.9 ± 0.2b | 1.9 ± 0.3d | 10.3 ± 0.7c | 9.7 ± 0.8b | 2.2 ± 0.3bc | 0.6 ± 0.2c | |
| 5ICC | 3 ± 0.4d | 59 ± 6.5cde | 11.8 ± 2bcd | 0.7 ± 0.1ab | 20.7 ± 3.6b | 3.5 ± 0.6b | 4 ± 0.6c | 12.8 ± 1.6c | 13 ± 1.8b | 1.5 ± 0.3c | 25.7 ± 5.2a | |
| 5ICC-B | 3.1 ± 0.4d | 48 ± 6.8de | 8.3 ± 1.4cd | 0.4 ± 0.4ab | 12.6 ± 2.9b | 2.6 ± 0.3b | 2.4 ± 0.5cd | 11.5 ± 2.5c | 11.8 ± 3.3b | 1.8 ± 0.8bc | 7.1 ± 5.2abc | |
| 5ICC-D | 3.4 ± 0.8cd | 55.4 ± 14.8dce | 10.2 ± 3.7bcd | 0.7 ± 0.2ab | 14.6 ± 5.4b | 3.3 ± 1.1b | 2.4 ± 0.7cd | 18.3 ± 6.3bc | 18.6 ± 6.6ab | 2.8 ± 1abc | 0.8 ± 0.3c |
All values are reported as mean and standard deviation from three replicates. Values in a column followed by the same letter are not significantly different (at P = 0.05).
5IWB, 5IWB-B, 5IWB-D, fifth instar larvae from waste mixture subjected to homogenisation, blanching, and drying respectively; 5IWL, 5IWL-B, 5IWL-D, fifth instar larvae from supermarket waste subjected to homogenisation, blanching and drying respectively; 5ICC, fifth instar larvae from childcare centre waste subjected to homogenisation, blanching and drying respectively.
The unprocessed BSFL and blanched samples within the same group did not differ (P > 0.05) in terms of the composition of palmitic, stearic, and oleic acids, except for the blanched larvae from the WB groups. Blanched BSFL exhibited a lower concentration of these fatty acids than did the unprocessed BSFL from WB streams. As pertaining to lauric acid, no difference (P > 0.05) was observed between the unprocessed and blanched BSFL samples from all three waste streams. The concentration of fatty acids including lauric acid, palmitic acid, stearic acid, oleic acid, and nervonic acid in dried BSFL samples was found to be reduced and significantly (P < 0.05) different from that of the unprocessed samples from all three waste streams. It is noteworthy that no statistical difference (P > 0.05) was observed between blanched and dried BSFL samples for lauric acid and nervonic acid. Similar results were observed for palmitic, stearic, and oleic acids; however, the dried samples from WL were found to be lower (P < 0.05) in concentration than were the blanched samples obtained from the same waste stream.
It has been reported in previous studies that the SFA content in BSFL, except for lauric acid, does not vary greatly in BSFL on rearing them with different substrates and, similar results were obtained in our studies. However, the lauric acid content was also found to be similar. This may be due to the limited variability observed in the carbohydrate content of the three waste streams (Facility B) used for this study. The distribution of PUFA in BSFL is mainly influenced by the rearing substrate employed. The composition of PUFAs in unprocessed BSFL varied only for the samples from the CC waste stream because the constituents of WL and WB were similar.
The blanching and drying conditions implemented in our studies reduced the FA content of BSFL samples (on a DM basis). The extent of reduction caused by these processing methods was influenced mainly by the initial concentration of FAs in the unprocessed BSFL. It is noteworthy that the FA composition of the blanched and dried BSFL samples in this study is comparable to that of fishmeal and soybean meal, suggesting that the processing methods employed in this study do not compromise the use of BSFL as feed ingredients for poultry and aquaculture production (Zhou et al. 2018; Hossain et al. 2021).
Pearson correlation coefficients
Pearson correlation coefficients of the different variables in BSFL samples are depicted in Fig. 8. A moderate positive correlation was observed for the CF with most of the minerals and saturated fatty acids. The CF, ash content and the saturated fatty acid content in BSFL is primarily influenced by the substrate as observed in previous research and in this study (Gold et al. 2018; Siddiqui et al. 2022). These results show the prevalence and strong coexistence of these attributes in the food waste streams used for rearing BSFL. Similar results were observed for starch with CF, minerals, and FA content. This could be attributed to BSFL utilising carbohydrates to synthesise FAs and to increase the fat content (Ewald et al. 2020; Eggink et al. 2023). NDF was found to be moderately correlated with most of the AAs, but a very low positive correlation was observed with proteins. Minerals, including P, S and B, have moderative positive correlations with most of the AAs. Minerals such as Na were negatively correlated with the AA profile and most of the FAs. It is noteworthy that most of the minerals, except for Na and B, exhibited positive correlations with each other. This suggests the strong coexistence of these elements in the different substrates that were used for rearing BSFL (Alagappan et al. 2022a, 2022b). It has been reported in literature that food waste streams are observed to be characterised with a diverse set of minerals (Liu et al. 2023).
Pearson correlation matrix of different nutritional attributes of black soldier fly larvae reared with different food waste streams.

In case of the AAs, the LAAs such as Thr, Val, Ile, Leu, and Lys exhibited good correlations with the other AAs in BSFL. Moderate to poor correlations were observed for the other LAAs, including Met and His. BSFL in our studies were found to be higher in Met content than has been reported in previous studies. Higher concentrations of Met in several other species of insects belonging to the order of Diptera were found to affect their development and influence the balance of the overall AA profile (Weeks et al. 2019). This could have possibly resulted in the negative correlation observed for Met with several other AAs that were analysed. The non-LAAs such as Asp, Glu, Ser, Arg exhibited strong positive correlations with the other AAs. For Gly, Ala, Pro and Tyr, a moderate correlation with the other AAs was observed. The positive correlation observed for the AAs could be explained by their physiological importance at different stages in the life cycle of the larvae (Tomberlin et al. 2023). For instance, Lys, Arg and Glu were observed to play a major role in energy metabolism functions in several species of insects (Tomberlin et al. 2023). AAs including Thr and Ser are responsible for playing roles in development and reproduction in different species of insects (Tomberlin et al. 2023).
Minerals including Ca and Ba exhibited moderate positive correlations with FAs, whereas Se exhibited a good positive correlation with the FAs. Insects have been reported to maintain Ca ion homeostasis to ensure proper functioning of their lipid metabolism; this could explain the positive correlations observed for Ca with the different FAs (Kaczmarek and Boguś 2021). The positive correlation of Se with the FAs could probably be attributed to the role played of Se in influencing FA composition in plants and animals (Ferrari et al. 2023). The medium-chain saturated FAs were found to have a moderate correlation with the other SFAs in BSFL (Fig. 8). However, PUFAs were found to be strongly correlated with each other. This is because the concentration of PUFAs in BSFL is influenced by the substrate used (Ewald et al. 2020); these correlation coefficients suggest their prevalence and coexistence in the food waste streams used in this study. Also, SFAs are synthesised by BSFL, and their synthesis is influenced by the carbohydrate content of the substrate and the metabolic rigour of the larvae.
Conclusions
The nutritional profile of BSFL is influenced by several factors, including the rearing substrate, larval age and, post-harvest treatments, with substrate being the most crucial factor. The study investigated the effect of larval instar and post-harvest treatments on the nutritional profile of BSFL reared on different food waste streams in two different commercial production facilities. The post-harvest treatments conducted in these studies did not significantly alter the nutritional composition of the different nutritional parameters analysed. The larval instar was found to influence the crude fat and fatty acid content of the BSFL, but had only a partial effect on their amino acid composition. The mineral concentrations of the BSFL were also influenced by the substrate, with little to no effect established by the age of the BSFL. The outcomes of this study will assist commercial manufactures to select substrate(s) with desired nutritional properties to obtain BSFL with a desired nutritional quality. It will also assist producers to tailor their post-harvest treatments to obtain BSFL suitable to meet varying market requirements.
Data availability
The data that support this study will be shared upon reasonable request to the corresponding author.
Declaration of funding
The work has been supported by the End Food Waste Cooperative Research Centre (Project 2.4.1), whose activities are funded by the Australian Government’s Cooperative Research Centre Program.
Author contributions
Shanmugam Alagappan: data curation, formal analysis, data curation, investigation, writing original draft, review and edit, and visualisation; sampling, spectra collection, analysis, data interpretation, draft preparation; Hung Hong: methodology, data curation, validation and, original draft preparation; Deirdre Mikkelsen: writing – review and editing and, supervision; Sandra Olarte Mantilla: writing – review and editing and, supervision; Peter James: writing – review and editing and, supervision; Olympia Yarger: writing – review and editing and, supervision; Daniel Cozzolino: investigation, original draft preparation, review and editing and, supervision; Louwrens Hoffman: investigation, original draft preparation, review and editing, supervision, project administration, and funding acquisition.
References
Alagappan S, Hoffman LC, Mantilla SMO, Mikkelsen D, James P, Yarger O, Cozzolino D (2022a) Near Infrared spectroscopy as a traceability tool to monitor black soldier fly larvae (Hermetia illucens) intended as animal feed. Applied Sciences 12(16), 8168.
| Crossref | Google Scholar |
Alagappan S, Rowland D, Barwell R, Cozzolino D, Mikkelsen D, Olarte Mantilla SM, James P, Yarger O, Hoffman L (2022b) Organic side streams (bioproducts) as substrate for black soldier fly (Hermetia illucens) intended as animal feed: chemical safety issues. Animal Production Science 62(17), 1639-1651.
| Crossref | Google Scholar |
Alagappan S, Dong A, Mikkelsen D, Hoffman LC, Mantilla SMO, James P, Yarger O, Cozzolino D (2023) Near infrared spectroscopy for prediction of yeast and mould counts in black soldier fly larvae, feed and frass: a proof of concept. Sensors 23(15), 6946.
| Crossref | Google Scholar |
Amicarelli V, Lagioia G, Bux C (2021) Global warming potential of food waste through the life cycle assessment: an analytical review. Environmental Impact Assessment Review 91, 106677.
| Crossref | Google Scholar |
Barbi S, Macavei LI, Fuso A, Luparelli AV, Caligiani A, Ferrari AM, Maistrello L, Montorsi M (2020) Valorization of seasonal agri-food leftovers through insects. Science of The Total Environment 709, 136209.
| Crossref | Google Scholar |
Barragán-Fonseca K, Pineda-Mejia J, Dicke M, Van Loon JJ (2018) Performance of the black soldier fly (Diptera: Stratiomyidae) on vegetable residue-based diets formulated based on protein and carbohydrate contents. Journal of Economic Entomology 111(6), 2676-2683.
| Crossref | Google Scholar |
Barragan-Fonseca KB, Gort G, Dicke M, van Loon JJA (2019) Effects of dietary protein and carbohydrate on life-history traits and body protein and fat contents of the black soldier fly Hermetia illucens. Physiological Entomology 44(2), 148-159.
| Crossref | Google Scholar |
Bessa LW, Pieterse E, Marais J, Hoffman LC (2020) Why for feed and not for human consumption? The black soldier fly larvae. Comprehensive Reviews in Food Science and Food Safety 19(5), 2747-2763.
| Crossref | Google Scholar |
Beyers M, Coudron C, Ravi R, Meers E, Bruun S (2023) Black soldier fly larvae as an alternative feed source and agro-waste disposal route – a life cycle perspective. Resources, Conservation and Recycling 192, 106917.
| Crossref | Google Scholar |
Cammack JA, Tomberlin JK (2017) The Impact of diet protein and carbohydrate on select life-history traits of the black soldier fly Hermetia illucens (L.) (Diptera: Stratiomyidae). Insects 8(2), 56.
| Crossref | Google Scholar |
Cheng M, McCarl B, Fei C (2022) Climate change and livestock production: a literature review. Atmosphere 13(1), 140.
| Crossref | Google Scholar |
Chua ET, Brunner M, Atkin R, Eltanahy E, Thomas-Hall SR, Schenk PM (2019) The ionic liquid cholinium arginate is an efficient solvent for extracting high-value Nannochloropsis sp. lipids. ACS Sustainable Chemistry & Engineering 7(2), 2538-2544.
| Crossref | Google Scholar |
Danieli PP, Lussiana C, Gasco L, Amici A, Ronchi B (2019) The effects of diet formulation on the yield, proximate composition, and fatty acid profile of the black soldier fly (Hermetia illucens L.) prepupae intended for animal feed. Animals 9(4), 178.
| Crossref | Google Scholar |
Do S, Koutsos EA, Utterback PL, Parsons CM, de Godoy MRC, Swanson KS (2021) Amino acid digestibility and digestible indispensable amino acid score-like values of black soldier fly larvae fed different forms and concentrations of calcium using the precision-fed cecectomized rooster assay. Journal of Animal Science 99(6), skab124.
| Crossref | Google Scholar |
Dolan SP, Capar SG (2002) Multi-Element analysis of food by microwave digestion and inductively coupled plasma-atomic emission spectrometry. Journal of Food Composition and Analysis 15(5), 593-615.
| Crossref | Google Scholar |
Ebeneezar S, Linga Prabu D, Teipal CS, Jeena NS, Summaya R, Chandrasekar S, Sayooj P, Vijayagopa P (2021) Nutritional evaluation, bioconversion performance and phylogenetic assessment of black soldier fly (Hermetia illucens, Linn. 1758) larvae valorized from food waste. Environmental Technology & Innovation 23, 101783.
| Crossref | Google Scholar |
Eggink KM, Donoso IG, Dalsgaard J (2023) Optimal dietary protein to carbohydrate ratio for black soldier fly (Hermetia illucens) larvae. Journal of Insects as Food and Feed 9(6), 789-798.
| Crossref | Google Scholar |
Erhard AL, Águas Silva M, Damsbo-Svendsen M, Menadeva Karpantschof B-E, Sørensen H, Bom Frøst M (2023) Acceptance of insect foods among Danish children: effects of information provision, food neophobia, disgust sensitivity, and species on willingness to try. Food Quality and Preference 104, 104713.
| Crossref | Google Scholar |
Ewald N, Vidakovic A, Langeland M, Kiessling A, Sampels S, Lalander C (2020) Fatty acid composition of black soldier fly larvae (Hermetia illucens) – Possibilities and limitations for modification through diet. Waste Management 102, 40-47.
| Crossref | Google Scholar |
Ferrari L, Cattaneo DMIR, Abbate R, Manoni M, Ottoboni M, Luciano A, von Holst C, Pinotti L (2023) Advances in selenium supplementation: from selenium-enriched yeast to potential selenium-enriched insects, and selenium nanoparticles. Animal Nutrition 14, 193-203.
| Crossref | Google Scholar |
Fischer H, Romano N (2021) Fruit, vegetable, and starch mixtures on the nutritional quality of black soldier fly (Hermetia illucens) larvae and resulting frass. Journal of Insects as Food and Feed 7(3), 319-328.
| Crossref | Google Scholar |
Fitriana EL, Laconi EB, Astuti DA, Jayanegara A (2022) Effects of various organic substrates on growth performance and nutrient composition of black soldier fly larvae: a meta-analysis. Bioresource Technology Reports 18, 101061.
| Crossref | Google Scholar |
Gao Z, Wang W, Lu X, Zhu F, Liu W, Wang X, Lei C (2019) Bioconversion performance and life table of black soldier fly (Hermetia illucens) on fermented maize straw. Journal of Cleaner Production 230, 974-980.
| Crossref | Google Scholar |
Gold M, Tomberlin JK, Diener S, Zurbrügg C, Mathys A (2018) Decomposition of biowaste macronutrients, microbes, and chemicals in black soldier fly larval treatment: a review. Waste Management 82, 302-318.
| Crossref | Google Scholar |
Gold M, Cassar CM, Zurbrügg C, Kreuzer M, Boulos S, Diener S, Mathys A (2020) Biowaste treatment with black soldier fly larvae: increasing performance through the formulation of biowastes based on protein and carbohydrates. Waste Management 102, 319-329.
| Crossref | Google Scholar |
Hashmi MS, Akhtar S, Ismail T (2021) Biochemical characterization of vegetables wastes and development of functional bread: Vegetables wastes based functional bread. Journal of Microbiology, Biotechnology and Food Sciences 10(4), 691-696.
| Crossref | Google Scholar |
Heuel M, Kreuzer M, Sandrock C, Leiber F, Mathys A, Gold M, Zurbrügg C, Gangnat IDM, Terranova M (2021) Transfer of lauric and myristic acid from black soldier fly larval lipids to egg yolk lipids of hens is low. Lipids 56, 423-435.
| Crossref | Google Scholar | PubMed |
Hossain MS, Fawole FJ, Labh SN, Small BC, Overturf K, Kumar V (2021) Insect meal inclusion as a novel feed ingredient in soy-based diets improves performance of rainbow trout (Oncorhynchus mykiss). Aquaculture 544, 737096.
| Crossref | Google Scholar |
Isibika A, Simha P, Vinnerås B, Zurbrügg C, Kibazohi O, Lalander C (2023) Food industry waste – an opportunity for black soldier fly larvae protein production in Tanzania. Science of The Total Environment 858, 159985.
| Crossref | Google Scholar |
Kaczmarek A, Boguś M (2021) The metabolism and role of free fatty acids in key physiological processes in insects of medical, veterinary and forensic importance. PeerJ 9, e12563.
| Crossref | Google Scholar |
Kahsay BN, Moeller L, Imming P, Neubert RHH, Gebre-Mariam T (2022) Development and validation of a simple, selective, and accurate reversed-phase liquid chromatographic method with diode array detection (RP-HPLC/DAD) for the simultaneous analysis of 18 free amino acids in topical formulations. Chromatographia 85(7), 665-676.
| Crossref | Google Scholar |
Karkalas J (1985) An improved enzymic method for the determination of native and modified starch. Journal of the Science of Food and Agriculture 36(10), 1019-1027.
| Crossref | Google Scholar |
Kaya C, Generalovic TN, Ståhls G, Hauser M, Samayoa AC, Nunes-Silva CG, Roxburgh H, Wohlfahrt J, Ewusie EA, Kenis M, Hanboonsong Y, Orozco J, Carrejo N, Nakamura S, Gasco L, Rojo S, Tanga CM, Meier R, Rhode C, Picard CJ, Jiggins CD, Leiber F, Tomberlin JK, Hasselmann M, Blanckenhorn WU, Kapun M, Sandrock C (2021) Global population genetic structure and demographic trajectories of the black soldier fly, Hermetia illucens. BMC Biology 19(1), 94.
| Crossref | Google Scholar |
Lalander C, Diener S, Zurbrügg C, Vinnerås B (2019) Effects of feedstock on larval development and process efficiency in waste treatment with black soldier fly (Hermetia illucens). Journal of Cleaner Production 208, 211-219.
| Crossref | Google Scholar |
Lee JH, Kim T-K, Park S-Y, Kang M-C, Cha JY, Lim M-C, Choi Y-S (2023) Effects of blanching methods on nutritional properties and physicochemical characteristics of hot-air dried edible insect larvae. Food Science of Animal Resources 43(3), 428-440.
| Crossref | Google Scholar |
Li S, Zhu D, Li K, Yang Y, Lei Z, Zhang Z (2013) Soybean curd residue: composition, utilization, and related limiting factors. International Scholarly Research Notices 2013, 423590.
| Crossref | Google Scholar |
Liew CS, Yunus NM, Chidi BS, Lam MK, Goh PS, Mohamad M, Sin JC, Lam SM, Lim JW, Lam SS (2022) A review on recent disposal of hazardous sewage sludge via anaerobic digestion and novel composting. Journal of Hazardous Materials 423, 126995.
| Crossref | Google Scholar |
Liland NS, Biancarosa I, Araujo P, Biemans D, Bruckner CG, Waagbø R, Torstensen BE, Lock E-J (2017) Modulation of nutrient composition of black soldier fly (Hermetia illucens) larvae by feeding seaweed-enriched media. PLoS ONE 12(8), e0183188.
| Crossref | Google Scholar |
Liu X, Chen X, Wang H, Yang Q, ur Rehman K, Li W, Cai M, Li Q, Mazza L, Zhang J, Yu Z, Zheng L (2017) Dynamic changes of nutrient composition throughout the entire life cycle of black soldier fly. PLoS ONE 12(8), e0182601.
| Crossref | Google Scholar |
Liu Z, Minor M, Morel PCH, Najar-Rodriguez AJ (2018) Bioconversion of three organic wastes by black soldier fly (Diptera: Stratiomyidae) larvae. Environmental Entomology 47(6), 1609-1617.
| Crossref | Google Scholar |
Liu Z, de Souza TSP, Holland B, Dunshea F, Barrow C, Suleria HAR (2023) Valorization of food waste to produce value-added products based on its bioactive compounds. Processes 11(3), 840.
| Crossref | Google Scholar |
Liyanaarachchi GVV, Mahanama KRR, Somasiri HPPS, Punyasiri PAN, Kottawa-Arachchi JD (2020) Analysis of total amino acids in rice using a validated reversed-phase high performance liquid chromatographic method with diode array detection (RP-HPLC-DAD). International Journal of Analytical and Bioanalytical Methods 2, 008.
| Crossref | Google Scholar |
Lu S, Taethaisong N, Meethip W, Surakhunthod J, Sinpru B, Sroichak T, Archa P, Thongpea S, Paengkoum S, Purba RAP, Paengkoum P (2022) Nutritional composition of black soldier fly larvae (Hermetia illucens L.) and its potential uses as alternative protein sources in animal diets: a review. Insects 13(9), 831.
| Crossref | Google Scholar |
Makkar HPS, Tran G, Heuzé V, Ankers P (2014) State-of-the-art on use of insects as animal feed. Animal Feed Science and Technology 197, 1-33.
| Crossref | Google Scholar |
Maranga B, Kagali R, Mbogo K, Orina P, Munguti J, Ogello E (2022) Growth performance of african catfish (Clarias Gariepinus) fed on diets containing black soldier fly (Hermetia Illucens) larvae under aquaponic system. Aquaculture Studies 23(5), AQUAST910.
| Crossref | Google Scholar |
Meneguz M, Schiavone A, Gai F, Dama A, Lussiana C, Renna M, Gasco L (2018) Effect of rearing substrate on growth performance, waste reduction efficiency and chemical composition of black soldier fly (Hermetia illucens) larvae. Journal of the Science of Food and Agriculture 98(15), 5776-5784.
| Crossref | Google Scholar |
Montevecchi G, Macavei LI, Zanelli E, Benassi G, Pinotti G, D’Arco S, Buffagni S, Masino F, Maistrello L, Antonelli A (2023) Seasonal variability of the HO.RE.CA. food leftovers employed as a feeding substrate for black soldier fly (Hermetia illucens L.) larvae and effects on the rearing performance. Sustainable Chemistry and Pharmacy 33, 101061.
| Crossref | Google Scholar |
Nampijja Z, Kiggundu M, Kigozi A, Lugya A, Magala H, Ssepuuya G, Nakimbugwe D, Walusimbi SS, Mugerwa S (2023) Optimal substitution of black soldier fly larvae for fish in broiler chicken diets. Scientific African 20, e01636.
| Crossref | Google Scholar |
Nyakeri EM, Ogola HJO, Ayieko MA, Amimo FA (2017) Valorisation of organic waste material: growth performance of wild black soldier fly larvae (Hermetia illucens) reared on different organic wastes. Journal of Insects as Food and Feed 3(3), 193-202.
| Crossref | Google Scholar |
Opoku O, Hamidu JA, Adjei-Mensah B, Quaye B, Sasu P, Benante V, Amankrah MA, Donkoh A, Atuahene CC (2023) Alternative organic wastes as substrates and its impact on growth, proximate chemical and mineral composition of black soldier fly (Hermetia illucens) larvae for subsistent animal feed. Journal of Applied Animal Research 51(1), 564-572.
| Crossref | Google Scholar |
Ottoboni M, Spranghers T, Pinotti L, Baldi A, De Jaeghere W, Eeckhout M (2018) Inclusion of Hermetia Illucens larvae or prepupae in an experimental extruded feed: process optimisation and impact on in vitro digestibility. Italian Journal of Animal Science 17(2), 418-427.
| Crossref | Google Scholar |
Phaengphairee P, Boontiam W, Wealleans A, Hong J, Kim YY (2023) Dietary supplementation with full-fat Hermetia illucens larvae and multi-probiotics, as a substitute for antibiotics, improves the growth performance, gut health, and antioxidative capacity of weaned pigs. BMC Veterinary Research 19(1), 7.
| Crossref | Google Scholar |
Rahman MM, Mat K, Ishigaki G, Akashi R (2021) A review of okara (soybean curd residue) utilization as animal feed: nutritive value and animal performance aspects. Animal Science Journal 92(1), e13594.
| Crossref | Google Scholar |
Rampure SM, Velayudhannair K, Marimuthu N (2023) Characteristics of chitin extracted from different growth phases of black soldier fly, Hermetia illucens, fed with different organic wastes. International Journal of Tropical Insect Science 43(3), 979-987.
| Crossref | Google Scholar |
Ravi HK, Degrou A, Costil J, Trespeuch C, Chemat F, Vian MA (2020a) Effect of devitalization techniques on the lipid, protein, antioxidant, and chitin fractions of black soldier fly (Hermetia illucens) larvae. European Food Research and Technology 246(12), 2549-2568.
| Crossref | Google Scholar |
Ravi HK, Degrou A, Costil J, Trespeuch C, Chemat F, Vian MA (2020b) Larvae mediated valorization of industrial, agriculture and food wastes: biorefinery concept through bioconversion, processes, procedures, and products. Processes 8(7), 857.
| Crossref | Google Scholar |
Ribeiro N, Costa R, Ameixa OMCC (2022) The influence of non-optimal rearing conditions and substrates on the performance of the black soldier fly (Hermetia illucens). Insects 13(7), 639.
| Crossref | Google Scholar |
Romano N, Sinha A, Powell A, Fischer H (2023) Mineral composition in black soldier fly (Hermetia illucens) larvae and resulting frass from fruit and their peels. Journal of Insects as Food and Feed 9(1), 43-53.
| Crossref | Google Scholar |
Ryckebosch E, Muylaert K, Foubert I (2012) Optimization of an analytical procedure for extraction of lipids from microalgae. Journal of the American Oil Chemists’ Society 89(2), 189-198.
| Crossref | Google Scholar |
Scieuzo C, Franco A, Salvia R, Triunfo M, Addeo NF, Vozzo S, Piccolo G, Bovera F, Ritieni A, Francia AD, Laginestra A, Schmitt E, Falabella P (2023) Enhancement of fruit byproducts through bioconversion by Hermetia illucens (Diptera: Stratiomyidae). Insect Science 30(4), 991-1010.
| Crossref | Google Scholar |
Shah AA, Totakul P, Matra M, Cherdthong A, Hanboonsong Y, Wanapat M (2022) Nutritional composition of various insects and potential uses as alternative protein sources in animal diets. Animal Bioscience 35(2), 317-331.
| Crossref | Google Scholar | PubMed |
Shumo M, Khamis FM, Tanga CM, Fiaboe KKM, Subramanian S, Ekesi S, van Huis A, Borgemeister C (2019a) Influence of temperature on selected life-history traits of black soldier fly (Hermetia illucens) reared on two common urban organic waste streams in Kenya. Animals 9(3), 79.
| Crossref | Google Scholar |
Shumo M, Osuga IM, Khamis FM, Tanga CM, Fiaboe KKM, Subramanian S, Ekesi S, van Huis A, Borgemeister C (2019b) The nutritive value of black soldier fly larvae reared on common organic waste streams in Kenya. Scientific Reports 9(1), 10110.
| Crossref | Google Scholar |
Siddiqui SA, Ristow B, Rahayu T, Putra NS, Widya Yuwono N, Nisa K, Mategeko B, Smetana S, Saki M, Nawaz A, Nagdalian A (2022) Black soldier fly larvae (BSFL) and their affinity for organic waste processing. Waste Management 140, 1-13.
| Crossref | Google Scholar |
Siva Raman S, Stringer LC, Bruce NC, Chong CS (2022) Opportunities, challenges and solutions for black soldier fly larvae-based animal feed production. Journal of Cleaner Production 373, 133802.
| Crossref | Google Scholar |
Smets R, Verbinnen B, Van De Voorde I, Aerts G, Claes J, Van Der Borght M (2020) Sequential extraction and characterisation of lipids, proteins, and chitin from black soldier fly (Hermetia illucens) larvae, prepupae, and pupae. Waste and Biomass Valorization 11(12), 6455-6466.
| Crossref | Google Scholar |
Son J, Park SH, Jung HJ, You SJ, Kim BG (2023) Effects of drying methods and blanching on nutrient utilization in black soldier fly larva meals based on in vitro assays for pigs. Animals 13(5), 858.
| Crossref | Google Scholar |
Spranghers T, Ottoboni M, Klootwijk C, Ovyn A, Deboosere S, De Meulenaer B, Michiels J, Eeckhout M, De Clercq P, De Smet S (2017) Nutritional composition of black soldier fly (Hermetia illucens) prepupae reared on different organic waste substrates. Journal of the Science of Food and Agriculture 97(8), 2594-2600.
| Crossref | Google Scholar |
Tomberlin JK, Miranda C, Flint C, Harris E, Wu G (2023) Nutrients limit production of insects for food and feed: an emphasis on nutritionally essential amino acids. Animal Frontiers 13(4), 64-71.
| Crossref | Google Scholar |
Weeks EN, Baniszewski J, Gezan SA, Allan SA, Cuda JP, Stevens BR (2019) Methionine as a safe and effective novel biorational mosquito larvicide. Pest Management Science 75(2), 346-355.
| Crossref | Google Scholar |
Yakti W, Schulz S, Marten V, Mewis I, Padmanabha M, Hempel A-J, Kobelski A, Streif S, Ulrichs C (2022) The effect of rearing scale and density on the growth and nutrient composition of Hermetia illucens (L.) (Diptera: Stratiomyidae) Larvae. Sustainability 14(3), 1772.
| Crossref | Google Scholar |
Zhou JS, Liu SS, Ji H, Yu HB (2018) Effect of replacing dietary fish meal with black soldier fly larvae meal on growth and fatty acid composition of Jian carp (Cyprinus carpio var. Jian). Aquaculture Nutrition 24(1), 424-433.
| Crossref | Google Scholar |
Zulkifli NFNM, Seok-Kian AY, Seng LL, Mustafa S, Kim Y-S, Shapawi R (2022) Nutritional value of black soldier fly (Hermetia illucens) larvae processed by different methods. PLoS ONE 17(2), e0263924.
| Crossref | Google Scholar |