

Plantain-mixed pasture collected in different climatic seasons produced less methane and ammonia than ryegrass–white clover pasture in vitro
Komahan Sivanandarajah A * , Daniel Donaghy A , German Molano B , David Horne A , Peter Kemp A , Soledad Navarrete A C , Thiagarajah Ramilan A and David Pacheco B
A * , Daniel Donaghy A , German Molano B , David Horne A , Peter Kemp A , Soledad Navarrete A C , Thiagarajah Ramilan A and David Pacheco B
A
B
C
Abstract
Plantain (PL) is recognised for reducing nitrate leaching and nitrous oxide emissions in pastoral systems. Evidence has shown that cows fed pure PL produced less methane (CH4) than cows fed ryegrass. However, it is unclear if the CH4 reduction can be achieved with PL in mixed pasture.
The study evaluated the in vitro rumen fermentation profiles of ryegrass–white clover (RWC) and medium-level PL (PLM, containing ~40% PL) pasture collected during different climatic seasons, to determine whether this inclusion level influences CH4 and rumen ammonia (NH3) production.
Substrates were selected from samples with various proportions of PL. Samples were categorised into three climatic seasons (i.e. spring, summer and autumn) and two pasture types (PLM and RWC). Representative samples for these scenarios were tested in an automated in vitro rumen batch culture system for gas, CH4 (mL/g DM) and NH3 (mM/g DM) production.
In summer samples, PLM produced approximately 8%, 14% and 19% less CH4 at 12 h, 24 h and potential CH4 production (PCH4), respectively. Although gas production (GP) was similar at 12 and 24 h, PLM had 13% lower potential GP than RWC (P < 0.05). In spring samples, PLM had approximately 11% greater GP and CH4 production at 12 h. For the autumn samples, GP and CH4 production were similar between PLM and RWC (P > 0.05). Net NH3 production from PLM substrates was significantly lower in spring (27%) and autumn (17%) samples, with no differences in summer, despite higher crude protein levels in the selected PLM.
Compared with RWC, PLM changed rumen fermentation parameters that could translate to potential environmental benefits: PLM produced less net NH3 in spring and autumn samples (27% and 17%, respectively), and up to 19% less CH4 production in summer samples.
Incorporating ~40% PL into RWC pasture showed a promising reduction of CH4 emissions and nitrogen losses in vitro. If the in vitro results translate to cows grazing pasture, this could offer greater environmental benefits with minimal input costs. In vitro results suggest that PLM’s potential to mitigate CH4 emissions can be influenced by seasonal variations in pasture quality compared with RWC. However, further animal studies are needed to fully comprehend the CH4 mitigation potential of this forage.
Keywords: forage quality, greenhouse gas, methane mitigation, mixed pasture, nitrogen losses, plant secondary metabolites, Plantago lanceolata, rumen fermentation.
Introduction
Based on grazed pasture systems, New Zealand’s dairy industry makes a significant contribution to the national economy by efficient milk production at comparatively lower costs than indoor feeding systems (Ozawa et al. 2005). However, this benefit comes with trade-offs of elevated nitrogen (N) losses (Pinxterhuis et al. 2024) and methane (CH4) emissions from enteric fermentation (Leahy et al. 2019). In 2022, the agriculture sector contributed half (53%) of total emissions, with CH4 from enteric fermentation accounting for 78.2% of sector’s emissions, followed by nitrous oxide (N2O) emissions from agricultural soils (MfE 2024).
Feeding pastures to cows can provide N in excess of the animals’ nutritional requirements (Pacheco and Waghorn 2008). High N intake may increase rumen ammonia (NH3) concentration, thereby potentially raising urinary N excretion (Beltran et al. 2019). This could lead to high N leaching and N2O emissions (Selbie et al. 2015) from the urine patch. Ruminants grazing on low-quality feeds produce around 75% of CH4 emissions globally (Leng 1993) and offering high-quality pasture can mitigate CH4 emission per unit of DM intake (DMI; Arndt et al. 2022). High-quality pasture is generally characterised by greater concentrations of non-structural carbohydrates (NSC), greater digestibility and crude protein (CP) and lower fibre, particularly neutral detergent fibre (NDF; Waghorn and Clark 2004).
In recent years, incorporating plantain (PL; Plantago lanceolata L.) into a permanent pasture mix (RWC) of perennial ryegrass (Lolium perenne) and white clover (Trifolium repens) has become increasingly popular in the New Zealand dairy system for its environmental benefits (DairyNZ 2023a). Plantain is a mineral-rich forage herb that is easy to establish, adaptable to various agricultural soils and drought tolerant (Stewart 1996). Numerous studies have reported the benefits of PL pasture in the diet of dairy cattle. These benefits include lowering urinary N concentration (Box et al. 2017; Minnee et al. 2020) and lowering CH4 emissions (Della Rosa et al. 2022).
According to the New Zealand’s greenhouse gas inventory, dairy cows produce 21–22 g of CH4 per kg of DMI (Pickering et al. 2024). Methane production from ruminants is influenced by various factors, such as pasture quality, digestibility, DMI, plant bioactive compounds and grazing management (Hristov et al. 2013; Danielsson et al. 2017). Among these, pasture quality, digestibility and availability of plant bioactive compounds can be influenced by climatic season (Roche et al. 2009; Navarrete et al. 2016; Lahart et al. 2024). When comparing PL and RWC pasture across seasons, PL mixed pasture is usually superior in nutritional value (i.e. greater digestibility, greater NSC:N content, greater mineral content and lower NDF) compared with RWC pasture (Nguyen et al. 2022; Herath et al. 2023), and PL pasture contains bioactive compounds, such as aucubin and acteoside (Tamura and Nishibe 2002). Both bioactive compounds influence the rumen fermentation profile by lowering net NH3 production, and modifying either the maximum gas production (acteoside) or the rate at which the gas is produced (aucubin; Navarrete et al. 2016).
When pure PL was tested both in vitro (Durmic et al. 2016) and in vivo (Della Rosa et al. 2022), it reduced CH4 emissions, but the effect of PL in a mixed pasture remains unclear. Hammond et al. (2014) observed a decrease in CH4 yield (g of CH4 per unit of DMI) from heifers grazing a mixture of ryegrass with five species, including wild PL. Wilson et al. (2020) reported a tendency for lower CH4 emissions from cows fed mixed pasture, which included PL, chicory and white clover, compared with those fed with a mixture of grass species and white clover. In contrast, Jonker et al. (2019) reported an increase in CH4 yield from animals fed diverse pastures including PL, but the total CH4 emissions were similar to those of RWC. In these studies, the reduction in CH4 emissions cannot be solely attributed to the PL, as other species were included in varying proportions. Additionally, in a more recent study, Koning et al. (2024) observed greater CH4 yield for pasture containing <25% PL compared with ryegrass pasture. As the responses reported vary widely, it is important to understand the sources of the variation of these results. This study examined the seasonal variability of PL mixed pasture in vitro to test its ability to mitigate CH4.
In the mixed pasture, maintaining higher levels of PL is challenging, as it declines naturally over time (Dodd et al. 2019; Nguyen et al. 2022). The results from the study site of the Massey University PL trial, which includes measurements in four production seasons, suggest that approximately 20–50% PL is achievable in practice using different sowing rates (MPI 2024). Thus, it is also crucial to assess how such proportions of PL in RWC mix affect CH4 emissions.
Accordingly, the present study aimed to assess the in vitro rumen fermentation profiles of RWC and PL mixed pasture, consisting of a medium-level of PL, ~40% PL (PLM), collected during different climatic seasons. The objectives of this study were to evaluate the chemical compositions of PLM and RWC substrates to determine their influence on CH4 emissions and NH3 production, with a particular focus on the potential of PL, at ~40% in the pasture, to reduce CH4 emissions, as it would be a more practicable PL percentage to have in a mixed pasture.
Methods
Experimental site description
Substrates assessed in the present study were selected from historical pasture samples collected from the Plantain Potency and Practice program’s (DairyNZ 2023b) Massey University trial site at Dairy 4, Palmerston North, New Zealand (40°23′27″S, 175°36′44″E) containing four pasture treatments with a targeted proportion of PL in a RWC pasture mix of 0% (PL0), 30% (PL30), 50% (PL50) and 70% (PL70). Pasture establishment and general management were reported previously by Nguyen et al. (2022). In brief, over four production seasons (September to May), 32 grazings were conducted with lactating dairy cows, and pasture quality was analysed for 23 events. Eighteen of those events were specifically analysed for the presence of bioactive compounds.
Botanical composition was measured by manually separating pasture components, oven drying at 75°C until they reached constant weight and were then reported as a percentage of DM. For pasture quality and bioactive compounds analysis, samples were oven-dried at 60°C until a constant weight was achieved. The dried samples were ground through a 1-mm sieve, and one subsample was taken and analysed for chemical composition of pasture quality at a commercial laboratory (RJ Hill Laboratories, Hamilton, New Zealand), whereas another subsample was taken to test the bioactive compounds, aucubin, acteoside and catalpol, using high-performance liquid chromatography, as described by Navarrete et al. (2016). Pasture samples were submitted for quality analysis after each grazing event, whereas bioactive compound concentrations were assessed collectively as one batch at the end of each production season. Once dried, the samples were stored in airtight, opaque containers at room temperature in a low-light environment until incubation.
Chemical composition analysis for pasture samples was conducted by using near-infrared spectroscopy (Hill-Labs 2022). The samples were tested for the following parameters: organic matter digestibility (OMD) in DM (determined using Australian Fodder Industry Association pepsin-cellulase procedure and derived as in vivo using a linear regression based on calibration samples from Lincoln University), NDF, acid detergent fibre (ADF), lignin (calibration based on acid detergent extraction followed by treatment with 72% sulfuric acid in the Ankom Daisy Incubator), ash (calibration based on weight loss after ashing at 600°C for 2 h), soluble sugars (calibration based on an 80:20 ethanol:water extraction and colorimetric determination), crude fat (calibration based on petroleum spirit extraction by Ankom auto analyser, AOCS official procedure AM-5-04), CP (N multiplied by 6.25, with N calibration based on total N by Dumas combustion), OMD in DM (DOMD; solubilised organic matter as a portion of the DM) and NSC = 100 – (CP + ash + crude fat + NDF).
Substrate selection for in vitro
A collated dataset was created using variables from the pasture quality and bioactive compound analysis to categorise the samples. These variables included PL percentage in the pasture, CP, NDF, ADF, lignin, (lignin/NDF), ash, DOMD, soluble sugar, NSC, catalpol, aucubin and acteoside.
The PL percentage varied across years, with the lowest annual production of 18% and the highest of 50% (MPI 2024). The targeted annual population for PL70 was not achieved in any year (Supplementary Table S1). Therefore, regardless of the pasture treatments (PL0, PL30, PL50 and PL70), 0–5% PL was defined as RWC, and 30–50% PL as PLM. Data were grouped by these categories (RWC, PLM) and by season (spring, summer, autumn), and based on the mean, a representative sample for each season and pasture type (n = 6) was selected for the in vitro incubation.
In vitro study experimental design and treatments
The in vitro fermentation was conducted in a fully automated batch culture system, adhering to the protocols outlined by Muetzel et al. (2014).
Each treatment (shown in Table 1) was incubated in two sets. The first set continuously measured CH4 and GP over 48 h, and the second set was used to obtain sub-samples at 3, 6, 9, 12, 24 and 48 h for NH3 plus one end-point sample for analysis of volatile fatty acids (VFA). At each time point, bottles from the second set were briefly removed from incubation one at a time and returned to the incubator after sampling. Using separate sets kept gas measurements and sample collection independent yet aligned to the same fermentation timeline. Each of these incubations was repeated three times (replicates) for statistical evaluation, and each replicate had two duplicate bottles (analytical replicates). Each replicate consisted of a mixture of rumen fluid from two cows.
Rumen sampling and in vitro medium preparation
A reduced carbonate-based buffer solution (6.0 mM Na2HPO4, 9.6 mM KH2PO4, 0.5 mM MgCl2, 64.5 mM NaHCO3 and 17.8 mM NH4HCO3) was prepared, as described by Mould et al. (2005). Rumen fluid was collected separately from two fistulated, non-lactating cows before the morning feeding to reduce variability between incubations. The donor cows grazed a RWC pasture year-round. The collection of rumen fluid from fistulated cows and the management of these animals were approved by the AgResearch Grasslands Animal Ethics Committee, Palmerston North, New Zealand (AE699), in accordance with the Animal Welfare Act of 1999 and its amendments in New Zealand. The collected rumen fluid was filled to the top of pre-warmed insulated flasks, maintaining both temperature and anaerobic conditions during transport to the laboratory. Equal volumes of rumen fluid from each donor cow were combined and added to the buffer solution, making up 20% of the total volume (v/v) of the in vitro rumen-buffer mixture (medium), which was continuously flushed with CO2 to help maintain anaerobic conditions.
Incubation preparation and sub-sample collection
The substrates were weighed to 500 ± 10 mg and added to 125-mL serum bottles, which were labelled and pre-warmed to 39°C in an incubator. A 50-mL aliquot of the medium was dispensed into each incubation bottle under CO2 flushing. The bottles were capped with butyl rubber stoppers, shaken manually and then randomly placed on a rack in a reciprocal shaker inside the incubator. The bottles were connected to the automated gas measurement system via a 23-gauge needle, shaken horizontally at 120 rpm. Gas pressure within each incubation bottle was automatically monitored at 1-min intervals throughout the incubation period, with gas released and sampled for gas chromatograph analysis whenever the internal pressure exceeded a threshold of 9 kPa (Muetzel et al. 2014).
A sample of the medium was collected for analysis of NH3 and VFA (0-h sample). Samples of the fermented material were collected using a 3-mL syringe connected to a manual valve via a needle. At each sampling point, the bottle was shaken manually and then a 1.8 mL of sample was pipetted into 2-mL Eppendorf tubes and centrifuged (21,000g for 10 min at 4°C). An aliquot of 900 μL of supernatant was mixed with 100 μL of internal standard (19 mM ethyl butyrate in 20% (v/v) phosphoric acid) in a 1.5-mL micro-tube, and stored at −20°C until further analysis of NH3 and VFA within the next 3 weeks.
Laboratory analysis for samples from in vitro incubation
The frozen samples were thawed and centrifuged at 21,000g for 10 min at 4°C. An aliquot of 800 μL supernatant was transferred into a 2-mL crimp cap vial for VFA analysis. Volatile fatty acids were analysed using gas chromatography, as described by Attwood et al. (1998). Approximately 100 μL of the remaining supernatant was collected for NH3 concentration analysis using the colorimetric method described by Weatherburn (1967) scaled down to run in 96-well microplate format.
Model fitting and data analysis
Gas and CH4 production from each bottle were separately fitted to a logistic model (France et al. 2000) as a function of time to estimate the in vitro gas and CH4 production parameters using the following formulae:
Where:
V: cumulative volume of gas or CH4 produced up to time t (mL/g DM)
a: potential gas or CH4 production, which is the maximum volume that can be produced per unit of DM (mL/g DM)
b: fermentation rate constant (/h)
c: constant determining curve steepness and lag phase
t: time (h)
a: half-time (h)
a: rate of GP at half-time (mL/h DM)
Statistical analysis
Ammonia values were corrected for the amount of incubated substrate and the time 0 NH3 concentration in the medium, and reported as net NH3 production (mM/g DM). All statistical analyses were performed using the statistical software R version 4.4.0 (R Core Team 2024). The chemical composition (NDF, ADF, lignin, % of lignin in NDF, DOMD, OMD, CP, NSC, acteoside, aucubin), gas, CH4 production parameters and endpoint VFA, were analysed using linear mixed models in the R package ‘lme4’ (Bates et al. 2015). Residuals were checked for normality using the Shapiro–Wilk test, and log transformations were performed where necessary (for acteoside and aucubin) to meet the assumptions of ANOVA. The R package ‘emmeans’ (Lenth 2023) was used for multiple comparisons using estimated marginal means, and P-values were adjusted using the Tukey post hoc test. Treatment effects were declared significant at adjusted P < 0.05. The ‘multcompView’ package (Graves et al. 2019) was used to convert the P-values into a character-based display in which characters identify groups that are significantly different/not different (P < 0.05). Back-transformed values were used to display the means in the respective log-transformed analyses.
The chemical composition was analysed using a two-way ANOVA, considering treatment and season of the year, as well as their interactions, as fixed effects. Replicates of the treatment and production year were included as random effects. Gas and CH4 production parameters, along with endpoint VFA concentrations, were analysed using a one-way ANOVA with treatment as a fixed effect, and biological replicates (n = 3; each consisting of a pooled rumen fluid mixture from two donor cows per run to capture natural biological variability) of the experiment as random effects. For the analysis of net NH3 production, a repeated measurement model including the fixed effects of treatment and time, the random effect of biological replicates, the incubation bottle as the subject of repeated measurement, and the interaction between treatment and time of sampling was used.
Results
Chemical composition of pasture types
Season is used, for brevity, to describe the results of samples collected at a particular time of the year, and does not mean that the experiments were conducted in different seasons.
The chemical composition of the pastures (PLM and RWC) varied seasonally and between pasture types (Table 2). A significant interaction between pasture type and season was found in NDF (P < 0.01), NSC (P < 0.05) and soluble sugars (P < 0.05). The NDF content of RWC pasture was significantly greater (~13%) than that of PLM in each season. For the PLM pasture, similar NDF content was observed across seasons, ranging from 386 to 398 g/kg DM. Additionally, the NDF content in autumn RWC was significantly greater than spring and/or summer RWC. A general trend of declining NSC content from spring to summer in both pasture types was observed. However, NSC content for PLM in each season was greater than that of RWC (P < 0.001), with differences of 18%, 22% and 53% in spring, summer and autumn, respectively. Soluble sugar content was greater in spring for both pasture types, with PLM significantly greater than RWC (P < 0.01); whereas in summer and autumn, both had similar soluble sugar content. On average, PLM pasture contained approximately 24% greater lignin content than RWC pasture across seasons, with seasonal effects also observed on lignin content (P < 0.001). Lignin content increased from spring to autumn for both pasture types, but the content in PLM was greater in each season compared with RWC. Pasture CP and DOMD were significantly affected by season (P < 0.001), and were similar between pasture types in each season. Digestibility (DOMD) was greater in spring compared with other seasons in both pasture types (P < 0.001).
| Season | Substrates | Chemical composition (g/kg DM) | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| NDF | ADF | NSC | Soluble sugar | CP | NSC/N (ratio) | DOMD | OMD in vivo | Lignin | Lignin in NDF (%) | Aucubin | Acteoside | ||||
| Average nutritive values across seasons | Spring | RWC | 430b | 244 abc | 220bc | 80b | 210b | 7.1ab | 701a | 786a | 54e | 12.7e | 0.1c | 1.4c | |
| PLM | 386c | 234c | 260a | 95a | 212b | 8.2a | 691a | 778a | 72cd | 18.9bc | 6.0a | 13.2a | |||
| Summer | RWC | 446b | 255a | 194c | 65c | 220ab | 6.1b | 660b | 741b | 70d | 15.7d | 0.8c | 1.1c | ||
| PLM | 398c | 244bc | 236ab | 65c | 226ab | 6.9b | 662b | 746b | 83b | 21.2b | 4.2b | 8.3b | |||
| Autumn | RWC | 477a | 251ab | 141d | 59c | 229ab | 4.5c | 657b | 742b | 79bc | 16.5cd | 1.3c | 0.3c | ||
| PLM | 393c | 236c | 216bc | 69bc | 237a | 6.2b | 664b | 757ab | 96a | 25.1a | 4.2ab | 10.1ab | |||
| P-values | s.e.m. | 19.1 | 10.9 | 11.8 | 7.2 | 22.3 | 10.9 | 18.0 | 19.6 | 4.3 | 1.2 | 0.7 | 1.1 | ||
| Pasture type (T) | <0.001 | <0.001 | <0.001 | <0.01 | n.s. | <0.001 | n.s. | n.s. | <0.001 | <0.001 | <0.001 | <0.001 | |||
| Season (S) | <0.001 | <0.01 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | n.s. | n.s. | |||
| T × S | <0.01 | n.s. | <0.05 | <0.05 | n.s. | n.s. | n.s. | n.s. | n.s. | n.s. | <0.001 | <0.001 | |||
| Samples selected for incubation | Spring | RWC | 451 | 242 | 179 | 64 | 240 | 4.7 | 688 | 761 | 49 | 10.9 | 0.2 | 0.6 | |
| PLM | 366 | 215 | 245 | 73 | 250 | 6.1 | 708 | 793 | 72 | 19. 7 | 5.0 | 11.1 | |||
| Summer | RWC | 458 | 261 | 213 | 94 | 201 | 6.7 | 669 | 738 | 64 | 13.6 | nd | nd | ||
| PLM | 377 | 226 | 241 | 84 | 234 | 6.4 | 694 | 785 | 74 | 19.6 | 2.6 | 9.5 | |||
| Autumn | RWC | 441 | 234 | 129 | 56 | 278 | 2.9 | 668 | 755 | 61 | 13.8 | 0.8 | 0.5 | ||
| PLM | 400 | 235 | 231 | 64 | 229 | 6.2 | 673 | 754 | 92 | 23 | 5 | 21 | |||
Values marked with the same letters within columns are not statistically different at the 5% significance level.
RWC, ryegrass–white clover pasture; PLM, medium-level plantain pasture (30–50%); nd, not detected; n.s., not significant; T × S, interaction between pasture type and season; NDF, neutral detergent fibre; ADF, acid detergent fibre; NSC, non-structural carbohydrate; CP, crude protein; N, nitrogen; DOMD, digestibility of organic matter in dry matter; OMD in vivo, organic matter digestibility in vivo; s.e.m., standard error of means of the pasture types.
The NSC:N ratio was significantly greater in PLM compared with RWC during autumn, and was statistically similar in spring and summer. A declining trend in NSC:N was observed from spring to autumn in both pastures; however, PLM had grater values in each season, with high absolute values in spring and summer, and greater values in autumn (P < 0.001). Both aucubin and acteoside concentrations were greater in PLM compared with RWC (P < 0.001), with acteoside levels being greater than aucubin. An interaction between seasons and pasture was observed for acteoside and aucubin, with PLM pasture having the greatest levels of these compounds in spring, the lowest levels in summer and levels in the autumn pasture being intermediate. Catalpol was detected at low levels in PLM (<1 g/kg DM in general, with a maximum value of 2.41 g/kg DM); therefore, it was excluded from the analysis (data not shown).
Numerical differences in the analysed mean values and absolute values of selected pasture samples for in vitro were observed (Table 2); however, these values remained within the typical range for the given season.
Gas and methane production parameters
Assessment of the methane effects requires measurement of both methane and gas production, to distinguish between results that indicate low fermentability of a substrate (i.e. low gas and methane production) and an inhibition of methanogenesis (i.e. lower methane proportion in the gas produced). Table 3 and Fig. 1 show total gas and CH4 production parameters measured in vitro over 48 h from selected pastures listed in Table 2.
| Seasons | Substrates | % of CH4/GP 12 h | % of CH4/GP 24 h | % of potential CH4/GP | GP at 12 h (mL/g DM) | GP at 24 h (mL/g DM) | PGP (mL/g DM) | CH4 at 12 h (mL/g DM) | CH4 at 24 h (mL/g DM) | PCH4 (mL/g DM) | T1/2 PGP (h) | T1/2 CH4 (h) | R1/2 PGP (mL/h DM) | R1/2 PCH4 (mL/h DM) | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Spring | RWC | 11.72ab | 13.82a | 14.38a | 179.36b | 253.03a | 285.22ab | 21.04bc | 35.00ab | 41.17a | 8.73a | 11.67a | 12.64b | 1.72a | |
| PLM | 11.83ab | 13.47a | 13.65ab | 198.94a | 255.03a | 282.29ab | 23.55a | 34.31ab | 38.53ab | 6.62bc | 9.34b | 14.25a | 1.75a | ||
| Summer | RWC | 12.12ab | 13.95a | 13.64ab | 191.25a | 252.66a | 298.70a | 23.18a | 35.25a | 40.89a | 7.89a | 10.25ab | 12.21bc | 1.70a | |
| PLM | 11.14c | 12.66b | 12.68b | 192.12a | 240.43ab | 261.32bc | 21.41b | 30.45c | 33.10b | 6.03c | 8.79b | 14.40a | 1.65a | ||
| Autumn | RWC | 12.19a | 13.96a | 13.93ab | 168.13c | 231.08b | 254.97c | 20.52bc | 32.26bc | 35.52ab | 8.13a | 10.37ab | 12.23bc | 1.74a | |
| PLM | 11.64bc | 13.38a | 13.28ab | 172.95bc | 227.99b | 268.42bc | 20.14c | 30.46c | 35.55ab | 7.70ab | 10.34ab | 11.19c | 1.44b | ||
| s.e.m. | 0.43 | 0.74 | 0.85 | 5.31 | 4.24 | 6.49 | 1.20 | 1.86 | 2.82 | 0.49 | 0.57 | 0.55 | 0.08 |
Values marked with the same lowercase letters within columns are not statistically different at the 5% significance level.
RWC, ryegrass-white clover pasture; PLM, medium-level plantain pasture (30–50%); GP, gas production; h, hours; PGP, potential gas production; CH4, methane production; PCH4, potential methane production; T1/2 PGP, half-time of potential gas production (h); T1/2 PCH4, half-time of potential CH4 production (h); R1/2 PGP, rate of PGP at T1/2; R1/2 PCH4, rate of PCH4 production at T1/2; s.e.m., standard error of the means.
Methane production (CH4, mL/g DM) for medium-level plantain pasture (PLM, Δ, dotted line) and ryegrass–white clover pasture (RWC, ●, solid line) collected in (a) spring, (b) summer and (c) autumn, over 48 h of in vitro batch culture incubation.
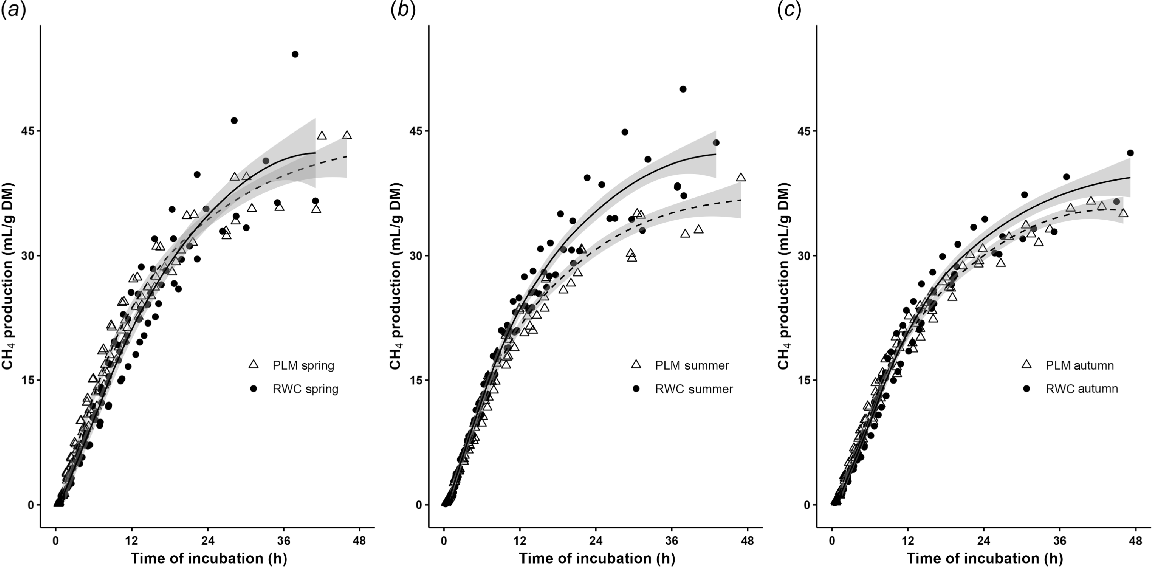
In summer pastures, PLM produced approximately 8%, 14% and 19% less CH4 at 12 h, 24 h and potential CH4 production (PCH4), respectively, compared with RWC. Gas production was similar at 12 h and 24 h, but a 13% lower PGP was observed in PLM. This led to reductions in CH4 proportion in GP (%CH4) of approximately 8% and 9% at 12 and 24 h; however, the %CH4 in PGP remained similar (Table 3).
In spring, PLM had approximately 11% greater GP and 12% greater CH4 production at 12 h. As time progressed beyond 12 h, both PLM and RWC produced similar GP and CH4 levels. Despite a general trend for a numerical reduction in the %CH4, no significant difference was found at any given time point for spring pastures (Table 3).
During autumn, PLM was not significantly different from RWC in GP and CH4 production at 12 h (P > 0.05). However, the PLM resulted in lower %CH4/GP at 12 h (P < 0.05), but was not different at 24 h (P > 0.05). Additionally, PGP and PCH4 remained similar between PLM and RWC.
The fermentation rate (R1/2) at half-time of PGP (T1/2 gas) was faster in PLM during spring and summer (13 and 18% greater, respectively). Similarly, in spring and summer, PLM reached its T1/2 gas approximately 24% faster than RWC. In autumn, both pastures reached their T1/2 gas and T1/2 CH4 at similar times. However, although the R1/2 gas was similar for both pastures in autumn, the R1/2 CH4 was significantly lower (17%) in PLM compared with RWC (Table 3).
Volatile fatty acids analysis
End-point VFA concentrations were similar between substrates collected in each season. A high acetate:propionate ratio was observed in PLM for the summer substrates (P < 0.05). Notably, the PLM substrate produced higher molar proportions of minor fatty acids compared with RWC in both summer and autumn. Additionally, a high molar proportion of butyrate was observed in the spring and autumn PLM pasture (Table 4).
| Seasons | Substrates | VFA (mM) | Acetate (%) | Propionate (%) | Butyrate (%) | Caproate (%) | Valerate (%) | Isobutyrate (%) | Isovalerate (%) | Minor (%) | (A:P) | (AcBu:PrVa) | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Spring | RWC | 83.61a | 65.89bc | 17.89ab | 9.89cd | 0.09c | 1.72b | 1.73b | 2.79b | 6.33b | 3.68bcd | 3.86c | |
| PLM | 83.74a | 65.95bc | 17.56b | 10.29b | 0.16a | 1.59c | 1.67c | 2.77bc | 6.20bc | 3.75bc | 3.98b | ||
| Summer | RWC | 83.91a | 66.02b | 17.47b | 10.86a | 0.08c | 1.55c | 1.49e | 2.53e | 5.65d | 3.78b | 4.04b | |
| PLM | 80.68ab | 67.08a | 16.75c | 10.10bc | 0.11b | 1.69b | 1.60d | 2.67d | 6.08c | 4.01a | 4.19a | ||
| Autumn | RWC | 81.87ab | 65.72bc | 18.02a | 9.55d | 0.09c | 1.91a | 1.83a | 2.88a | 6.71a | 3.65cd | 3.78c | |
| PLM | 76.65b | 65.51c | 18.08a | 10.27b | 0.12b | 1.66b | 1.66c | 2.70cd | 6.13c | 3.62d | 3.84c | ||
| s.e.m. | 2.71 | 0.35 | 0.10 | 0.10 | 0.01 | 0.07 | 0.10 | 0.19 | 0.36 | 0.03 | 0.04 |
Values marked with the same lowercase letters within columns are not statistically different at the 5% significance level.
RWC, ryegrass–white clover pasture; PLM, medium-level plantain pasture (30–50%); (%), denotes the molar proportions of respective VFA to total VFA production; (A:P), acetate to propionate ratio; (AcBu:PrVa), acetate + butyrate to propionate + valerate ratio; s.e.m., standard error of the means.
Net ammonia production
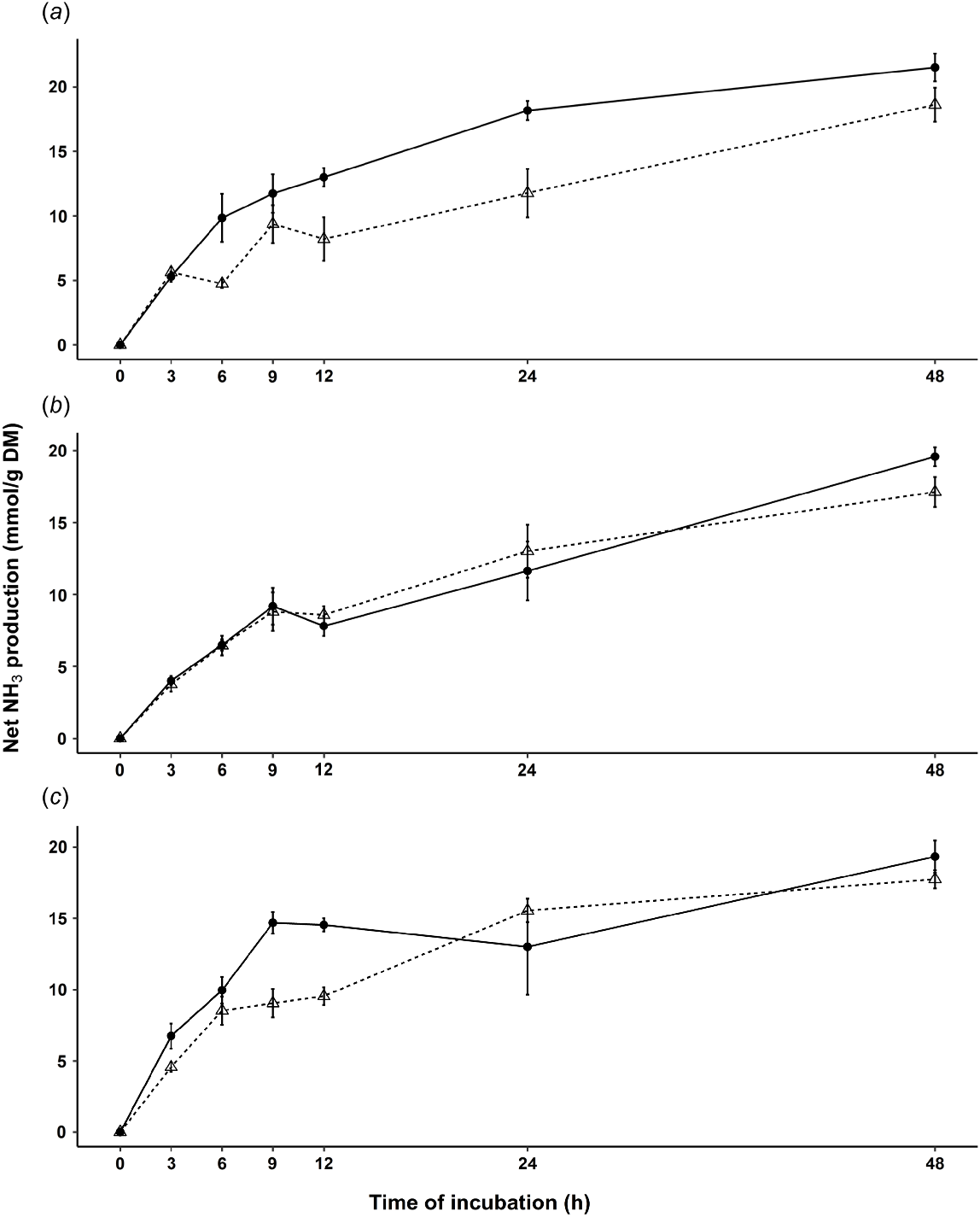
The net NH3 production over time was less for PLM compared with RWC from the spring and autumn samples. In spring, the net NH3 production was 27% lower for PLM pasture (Table 5). Specifically, PLM pasture collected during spring consistently showed lower net NH3 production at 6, 12 and 24 h, whereas during autumn, it was lower at 9 and 12 h (Fig. 2).
| Season | Substrates | Total ammonia (mM) | Net ammonia production (mM/g DM) | % difference in net ammonia compared with RWC | |
|---|---|---|---|---|---|
| Spring | RWC | 17.72a | 13.26a | – | |
| PLM | 16.12b | 9.73b | −26.63 | ||
| Summer | RWC | 16.13b | 9.79b | – | |
| PLM | 16.06b | 9.62b | −1.7 (n.s.) | ||
| Autumn | RWC | 17.61a | 13.05a | – | |
| PLM | 16.61b | 10.84b | −16.96 | ||
| s.e.m. | 1.54 | 0.61 | |||
| Source of variation | |||||
| Treatment | *** | *** | |||
| Time | *** | *** | |||
| Treatment × time | 0.06 | 0.06 | |||
Values marked with the same lowercase letters within columns are not statistically different at the 5% significance level.
RWC, ryegrass-white clover pasture; PLM, medium-level plantain pasture (30–50%).
***indicates significance at P < 0.001. Net ammonia = (total ammonia production – time 0 ammonia in the medium) per DM incubated.
In vitro net ammonia (NH3) production (mM/g DM) over time of medium-level plantain pasture (PLM, Δ, dotted line) and ryegrass–white clover pasture (RWC, ●, solid line) collected across seasons with comparison of (a) spring, (b) summer and (c) autumn, at each timepoint of in vitro batch culture incubation. Bars denoting standard error of the mean (s.e.m.) at each time point.
Discussion
A key finding of the present study was that PLM pasture has shown the potential to reduce CH4 production by up to 19% in vitro compared with RWC. This aligns with previous studies that reported lower CH4 production from pure PL pasture in vitro when compared with other forage species (Durmic et al. 2016), and in vivo when compared with ryegrass (Della Rosa et al. 2022). However, the measured reductions were not consistent across seasons for all parameters (e.g. CH4 production, GP and %CH4). This suggests that variations in the chemical composition of PLM, RWC or both, across different climatic seasons likely influenced the observed outcomes.
The effect of pasture quality on gas and methane emissions
Plantain generally has lower NDF, greater levels of NSC, lignin and improved or similar digestibility compared with ryegrass pasture (Stewart 1996; Minnee et al. 2019). In the present study, PLM substrates also had lower NDF, and greater NSC and lignin content compared with RWC, whereas digestibility, as measured by DOMD, remained similar between the two pasture types. The reported values of digestibility for PL mixed pastures were in a similar range of previous reports (Nkomboni et al. 2021; Herath et al. 2023). Others have reported that pure PL, particularly at its mature stage, has lower total tract DM digestibility (Della Rosa et al. 2022). In contrast to the latter study, the similar DOMD observed between pasture types in the present study could be due to the botanical and chemical composition of the pastures, as our PLM mix contained only approximately 40% PL.
Additionally, NDF levels remained relatively consistent across seasons for PL samples. However, for RWC, NDF content was greater in autumn compared with spring and summer (autumn > spring = summer). As the season progressed, NSC had a decreasing trend, whereas lignin increased in both pasture types. The DOMD was highest in spring, and declined in summer and autumn for both pastures. This aligned with the previous studies reporting PL pastures and RWC (Ulyatt et al. 2002; Navarrete et al. 2022; Herath et al. 2023).
Lignin serves as an indicator of potential NDF digestibility, and it correlates negatively with digestibility and CH4 emissions (Moore and Jung 2001; Hindrichsen et al. 2004), whereas NSC correlate positively with GP (Getachew et al. 2004). In the present study, lignin was greater in PLM than RWC across seasons. Despite the high lignin content, the total GP at 24 h was similar in both PLM and RWC regardless of the season. This suggests that the antagonistic effects of lignin and NSC on digestibility may have counterbalanced one another, and led to the forages to yield similar GP. Nevertheless, the limited sample size in the present study restricts the ability to construct a relationship between chemical composition and CH4 variables.
The PGP for PLM was lower than RWC in summer; however, in other seasons, PGP was similar between substrates. Notably, the GP at 24 h was similar between both substrates regardless of the season. This can be explained by the faster rate of degradation at half-time (R1/2), and the shorter time to reach half of the PGP (T1/2 PGP). These parameters suggest a more rapid utilisation of the fermentable substrate by microbes in the early part of the incubation. Box et al. (2018) reported a faster DM degradation rate in PL compared with RWC in sacco. These authors report that more PL DM had disappeared than RWC from hours 6–24. This suggests that with longer incubation times, once the fermentable carbohydrate pool is depleted, the degradation of substrate is slowed down, leading to the lower PGP in PLM.
In high-producing livestock, most digesta would rarely remain in the rumen for >24 h due to the rumen turnover (Keim et al. 2014). Consequently, the results from the first few hours of incubation in this study may be more relevant than PGP to explain what happens in vivo. Therefore, lower PGP – an indicator of lower total tract digestibility (Della Rosa et al. 2022) – may not be a significant concern for animal productivity. The lack of negative impacts on milk production from cows grazing the different PL pastures used in the current study (Nguyen 2023) suggests that the lower PGP observed in vitro may not be a direct indicator of the productivity effects of PL in vivo.
Although the spring PLM sample did not show potential for reducing CH4 emissions in vitro, a promising reduction, especially in summer, and a trend towards lower levels in autumn substrates suggest that animal trials are necessary to confirm whether CH4 reduction can be achieved at the farm level. Such trials would help to fully assess the potential of PLM as a CH4 mitigation tool.
The effect of pasture quality on net ammonia production
Another key finding of the present study was a reduction of up to 27% of net NH3 production from PL pasture collected in both spring and autumn. This finding is consistent with previous research by Durmic et al. (2016) and Navarrete et al. (2016), who reported similar reductions from pure PL pastures in in vitro studies. The present study demonstrated that such reductions can be achieved even with a medium-level of PL in the pasture. However, this effect was not seen in summer, indicating the effects on ammonia production may be influenced by seasonal variation in forage composition.
Protein degradation occurs in the rumen, where approximately 75–90% of the ingested CP in forage is degraded, with the remainder being rumen undegradable protein (Waghorn and Clark 2004). The protein degradation produces NH3, which is either utilised by rumen bacteria for their growth, forming microbial CP, or absorbed into the bloodstream. The absorbed NH3 can be recycled within the body or converted into urea and excreted primarily in the urine (Pacheco and Waghorn 2008). Hence, ruminal NH3 N not utilised for microbial protein synthesis will likely be excreted in urine. This represents a net loss to the animal and contributes to environmental pollution (Tamminga 1992).
A positive relationship between CP intake and urinary N losses has been well established (Box et al. 2018; Bougouin et al. 2022). Lower ruminal NH3 concentrations and greater microbial CP production have been shown to be typical responses to increased carbohydrate availability in rumen (Kolver et al. 1998), which indicates the importance of the ratio NSC:N in reducing NH3 in the rumen. The greater NSC:N ratios of PLM pasture compared with RWC in spring and autumn may have contributed to the significantly lower net NH3 production observed in PLM during these seasons (Table 4). In contrast, during summer, both RWC and PLM pastures had similar NSC:N ratios, resulting in similar net NH3 production.
The effect of bioactive compounds on methane and net ammonia production
Plantain contains various plant secondary compound groups, such as iridoid glycosides (aucubin) and phenylpropanoid glycoside (acteoside; Stewart 1996). The selected PLM pasture had approximately 11, 10 and 21 g/kg DM of acteoside, and 5, 3 and 5 g/kg DM of aucubin in spring, summer and autumn, respectively. The levels of acteoside and aucubin measured in this study were within the mid-range reported in previous studies (Navarrete Quijada 2015; Box and Judson 2018), likely due to the PLM containing only approximately 40% PL, leading to a proportional decrease in the concentrations of these bioactive compounds.
According to Navarrete et al. (2016) acteoside increases GP by serving as an additional energy source for microbes. Additionally, both acteoside and aucubin reduce net NH3 production, likely due to improved N utilisation by acteoside or the antimicrobial activity of aucubin (Navarrete et al. 2016). A study on Paulownia leaf extract, rich in acteoside with its derivatives, aucubin and other phenolic compounds, demonstrated a significant decrease in CH4 and NH3 production during in vitro rumen fermentation by altering microbial population, particularly methanogens and protozoa (Nowak et al. 2022).
In the present study, acteoside and aucubin found in PLM substrates may have influenced the NH3 and CH4 production to a certain extent. For example, the observed reductions in CH4 production in summer PLM and the lower R1/2 of PCH4 observed in autumn PLM may be due to the antimicrobial actions of bioactive compounds. In contrast, despite a numerically higher CP content in summer PLM samples (16% higher than RWC), the net NH3 production was similar between the forages, suggesting an effect of the bioactive compounds on this variable.
However, due to the confounding effects of chemical composition on net NH3 and CH4 production, and the lack of studies reporting the interaction between acteoside and aucubin in reducing these emissions, it is challenging to quantify the specific impact of these bioactive compounds on NH3 and CH4 production based on this study design.
Conclusions
The present study shows that in different seasons, PLM pasture exhibited evidence of potential environmental benefits compared with RWC pasture. Medium-level PL substrate (~40% PL) collected in summer produced up to 19% less CH4, suggesting a potential for PLM to reduce enteric emissions. Additionally, reductions in net NH3, up to 27% in spring and 17% in autumn, indicate the potential for lower urinary N output from cows, a critical source of nitrate leaching and N2O emissions. These findings are consistent with a broader body of in vivo research that shows that dietary inclusion of PL can reduce animal-level N losses to the environment. However, further animal studies at different seasons and inclusion levels of PL are required to confirm a CH4 mitigation potential for this forage.
Data availability
The data that support this study will be shared upon reasonable request to the corresponding author.
Conflicts of interest
David Pacheco is an Associate Editor for Animal Production Science but was not involved in the peer review or decision-making process for this paper.
Declaration of funding
This work was funded by the DairyNZ-led Sustainable Food and Fibre Futures Plantain Potency and Practice program funded by DairyNZ, the NZ Ministry for Primary Industries, PGG Wrightson Seeds and Fonterra. The primary author received postgraduate funding from Massey University, the TR Ellett Agricultural Research Trust and the Massey University Foundation.
Acknowledgements
The authors thank Massey University and AgResearch Grasslands for providing the facilities and support to conduct this research.
References
Arndt C, Hristov AN, Price WJ, McClelland SC, Pelaez AM, Cueva SF, Oh J, Dijkstra J, Bannink A, Bayat AR, Crompton LA, Eugène MA, Enahoro D, Kebreab E, Kreuzer M, McGee M, Martin C, Newbold CJ, Reynolds CK, Schwarm A, Shingfield KJ, Veneman JB, Yáñez-Ruiz DR, Yu Z (2022) Full adoption of the most effective strategies to mitigate methane emissions by ruminants can help meet the 1.5°C target by 2030 but not 2050. Proceedings of the National Academy of Sciences 119(20), e2111294119.
| Crossref | Google Scholar |
Attwood GT, Klieve AV, Ouwerkerk D, Patel BK (1998) Ammonia-hyperproducing bacteria from New Zealand ruminants. Applied and Environmental Microbiology 64(5), 1796-1804.
| Crossref | Google Scholar | PubMed |
Bates D, Mächler M, Bolker B, Walker S (2015) Fitting linear mixed-effects models using lme4. Journal of Statistical Software 67(1), 1-48.
| Crossref | Google Scholar |
Beltran IE, Gregorini P, Daza J, Balocchi OA, Morales A, Pulido RG (2019) Diurnal concentration of urinary nitrogen and rumen ammonia are modified by timing and mass of herbage allocation. Animals 9(11), 961.
| Crossref | Google Scholar | PubMed |
Bougouin A, Hristov A, Dijkstra J, Aguerre MJ, Ahvenjärvi S, Arndt C, Bannink A, Bayat AR, Benchaar C, Boland T, Brown WE, Crompton LA, Dehareng F, Dufrasne I, Eugène M, Froidmont E, Van Gastelen S, Garnsworthy PC, Halmemies-Beauchet-Filleau A, Herremans S, Huhtanen P, Johansen M, Kidane A, Kreuzer M, Kuhla B, Lessire F, Lund P, Minnée EMK, Muñoz C, Niu M, Nozière P, Pacheco D, Prestløkken E, Reynolds CK, Schwarm A, Spek JW, Terranova M, Vanhatalo A, Wattiaux MA, Weisbjerg MR, Yáñez-Ruiz DR, Yu Z, Kebreab E (2022) Prediction of nitrogen excretion from data on dairy cows fed a wide range of diets compiled in an intercontinental database: a meta-analysis. Journal of Dairy Science 105(9), 7462-7481.
| Crossref | Google Scholar | PubMed |
Box LA, Judson HG (2018) The concentration of bioactive compounds in Plantago lanceolata is genotype specific. Journal of New Zealand Grasslands 80, 113-118.
| Crossref | Google Scholar |
Box LA, Edwards GR, Bryant RH (2017) Milk production and urinary nitrogen excretion of dairy cows grazing plantain in early and late lactation. New Zealand Journal of Agricultural Research 60(4), 470-482.
| Crossref | Google Scholar |
Box LA, Edwards GR, Bryant RH (2018) In sacco digestion kinetics of plantain and ryegrass–white clover harvested in the morning and afternoon. New Zealand Journal of Animal Science and Production 78, 34-39.
| Google Scholar |
DairyNZ (2023a) Plantain. Available at https://www.dairynz.co.nz/feed/crops/environmental-benefits-of-plantain/
DairyNZ (2023b) Plantain potency and practice. Available at https://www.dairynz.co.nz/research/science-projects/plantain-potency-and-practice/
Danielsson R, Ramin M, Bertilsson J, Lund P, Huhtanen P (2017) Evaluation of a gas in vitro system for predicting methane production in vivo. Journal of Dairy Science 100(11), 8881-8894.
| Crossref | Google Scholar | PubMed |
Della Rosa M, Sandoval E, Luo D, Pacheco D, Jonker A (2022) Effect of feeding fresh forage plantain (Plantago lanceolata) or ryegrass-based pasture on methane emissions, total-tract digestibility, and rumen fermentation of nonlactating dairy cows. Journal of Dairy Science 105(8), 6628-6638.
| Crossref | Google Scholar |
Dodd MB, Moss RA, Pinxterhuis IJ (2019) A paddock survey of on-farm plantain use. Journal of New Zealand Grasslands 81, 125-130.
| Crossref | Google Scholar |
Durmic Z, Moate PJ, Jacobs JL, Vadhanabhuti J, Vercoe PE (2016) In vitro fermentability and methane production of some alternative forages in Australia. Animal Production Science 56(3), 641-645.
| Crossref | Google Scholar |
France J, Dijkstra J, Dhanoa MS, Lopez S, Bannink A (2000) Estimating the extent of degradation of ruminant feeds from a description of their gas production profiles observed in vitro: derivation of models and other mathematical considerations. British Journal of Nutrition 83(2), 143-150.
| Crossref | Google Scholar | PubMed |
Getachew G, Robinson PH, DePeters EJ, Taylor SJ (2004) Relationships between chemical composition, dry matter degradation and in vitro gas production of several ruminant feeds. Animal Feed Science and Technology 111(1–4), 57-71.
| Crossref | Google Scholar |
Hammond KJ, Humphries DJ, Westbury DB, Thompson A, Crompton LA, Kirton P, Green C, Reynolds CK (2014) The inclusion of forage mixtures in the diet of growing dairy heifers: impacts on digestion, energy utilisation, and methane emissions. Agriculture, Ecosystems & Environment 197, 88-95.
| Crossref | Google Scholar |
Herath G, Thomas C, McMillan N, Woods R, Bryant RH, Al-Marashdeh O (2023) Incorporating plantain into ryegrass-white clover mixed sward for an economically and environmentally sustainable dairy system: year one of a farm system study. Journal of New Zealand Grasslands 85, 321-331.
| Crossref | Google Scholar |
Hill-Labs (2022) Technical Note: Analysis of Feedstuffs Using Near Infra-Red Spectroscopy. Available at https://www.hill-labs.co.nz/media/qxvjxec3/8391v9_technical-note-pasture-feed-forage-quality.pdf
Hindrichsen IK, Wettstein H-R, Machmüller A, Soliva CR, Bach Knudsen KE, Madsen J, Kreuzer M (2004) Effects of feed carbohydrates with contrasting properties on rumen fermentation and methane release in vitro. Canadian Journal of Animal Science 84(2), 265-276.
| Crossref | Google Scholar |
Hristov AN, Oh J, Firkins JL, Dijkstra J, Kebreab E, Waghorn G, Makkar HPS, Adesogan AT, Yang W, Lee C, Gerber PJ, Henderson B, Tricarico JM (2013) Special topics – mitigation of methane and nitrous oxide emissions from animal operations: I. A review of enteric methane mitigation options. Journal of Animal Science 91(11), 5045-5069.
| Crossref | Google Scholar | PubMed |
Jonker A, Farrell L, Scobie D, Dynes R, Edwards G, Hague H, McAuliffe R, Taylor A, Knight T, Waghorn G (2019) Methane and carbon dioxide emissions from lactating dairy cows grazing mature ryegrass/white clover or a diverse pasture comprising ryegrass, legumes and herbs. Animal Production Science 59(6), 1063-1069.
| Crossref | Google Scholar |
Keim JP, López IF, Berthiaume R (2014) Nutritive value, in vitro fermentation and methane production of perennial pastures as affected by botanical composition over a growing season in the south of Chile. Animal Production Science 54(5), 598-607.
| Crossref | Google Scholar |
Kolver E, Muller LD, Varga G, Cassidy TJ (1998) Synchronization of ruminal degradation of supplemental carbohydrate with pasture nitrogen in lactating dairy cows. Journal of Dairy Science 81(7), 2017-2028.
| Crossref | Google Scholar | PubMed |
Koning L, Holshof G, Klop A, Klootwijk CW (2024) The effect of ribwort plantain (Plantago lanceolata) on enteric methane emission during grazing. In ‘Why grasslands? Proceedings of the 30th general meeting of the European Grassland Federation’, 9–13 June 24, Leeuwarden, Netherlands. pp. 214–216. (Brill)
Lahart B, Buckley F, Herron J, Fitzgerald R, Fitzpatrick E, Galvin N, Shalloo L (2024) Evaluating enteric methane emissions within a herd of genetically divergent grazing dairy cows. Journal of Dairy Science 107(1), 383-397.
| Crossref | Google Scholar | PubMed |
Leahy SC, Kearney L, Reisinger A, Clark H (2019) Mitigating greenhouse gas emissions from New Zealand pasture-based livestock farm systems. Journal of New Zealand Grasslands 81, 101-110.
| Crossref | Google Scholar |
Leng RA (1993) Quantitative ruminant nutrition – a green science. Australian Journal of Agricultural Research 44(3), 363-380.
| Crossref | Google Scholar |
Lenth R (2023) ‘Emmeans: estimated marginal means, aka least-squares means.’ (R package version 1.8.7) Available at https://CRAN.R-project.org/package=emmeans
MfE (2024) New Zealand’s greenhouse gas inventory 1990–2022. Ministry for the Environment, Wellington. Available at https://environment.govt.nz/assets/publications/GhG-Inventory/2024-GHG-inventory-2024/GHG-Inventory-2024-Vol-1.pdf
Minnee EMK, Kuhn-Sherlock B, Pinxterhuis IJB, Chapman DF (2019) Meta-analyses comparing the nutritional composition of perennial ryegrass (Lolium perenne) and plantain (Plantago lanceolata) pastures. Journal of New Zealand Grasslands 81, 117-124.
| Crossref | Google Scholar |
Minnee EMK, Leach CMT, Dalley DE (2020) Substituting a pasture-based diet with plantain (Plantago lanceolata) reduces nitrogen excreted in urine from dairy cows in late lactation. Livestock Science 239, 104093.
| Crossref | Google Scholar |
Moore KJ, Jung H-JG (2001) Lignin and fiber digestion. Journal of Range Management 54(4), 420-430.
| Crossref | Google Scholar |
Mould FL, Kliem KE, Morgan R, Mauricio RM (2005) In vitro microbial inoculum: a review of its function and properties. Animal Feed Science and Technology 123–124, 31-50.
| Crossref | Google Scholar |
MPI (2024) March to May 2024: plantain potency and practice progress report. MPI. Available at https://www.mpi.govt.nz/dmsdocument/64746-March-to-May-2024-Plantain-Potency-and-Practice-progress-report/sitemap
Muetzel S, Hunt C, Tavendale MH (2014) A fully automated incubation system for the measurement of gas production and gas composition. Animal Feed Science and Technology 196, 1-11.
| Crossref | Google Scholar |
Navarrete S, Kemp PD, Pain SJ, Back PJ (2016) Bioactive compounds, aucubin and acteoside, in plantain (Plantago lanceolata L.) and their effect on in vitro rumen fermentation. Animal Feed Science and Technology 222, 158-167.
| Crossref | Google Scholar |
Navarrete S, Rodriguez M, Horne D, Hanly J, Hedley M, Kemp P (2022) nitrogen excretion by dairy cows grazing plantain (Plantago lanceolata) based pastures during the lactating season. Animals 12(4), 469.
| Crossref | Google Scholar | PubMed |
Navarrete Quijada SDC (2015) Evaluation of herb pastures for New Zealand dairy systems. A thesis presented in partial fulfilment of the requirements for the degree of Doctor of Philosophy in Animal Sciences. Massey University, Manawatu, New Zealand. Available at http://hdl.handle.net/10179/10121
Nguyen TT (2023) Impact of plantain (Plantago lanceolata) based pasture on milk production of dairy cows and nitrate leaching from pastoral systems: a thesis presented in partial fulfilment of the requirements for the degree of Doctor of Philosophy in Animal Science at Massey University, Manawatu, New Zealand. Theses and Dissertations, Massey University.
Nguyen TT, Navarrete S, Horne DJ, Donaghy DJ, Kemp PD (2022) Incorporating plantain with perennial ryegrass-white clover in a dairy grazing system: dry matter yield, botanical composition, and nutritive value response to sowing rate, plantain content and season. Agronomy 12(11), 2789.
| Crossref | Google Scholar |
Nkomboni D, Bryant RH, Edwards GR (2021) Effect of increasing dietary proportion of plantain on milk production and nitrogen use of grazing dairy cows in late lactation. Animal Production Science 61(8), 770-779.
| Crossref | Google Scholar |
Nowak B, Moniuszko-Szajwaj B, Skorupka M, Puchalska J, Kozłowska M, Bocianowski J, Kołodziejski PA, Szumacher-Strabel M, Patra AK, Stochmal A, Cieslak A (2022) Effect of paulownia leaves extract levels on in vitro ruminal fermentation, microbial population, methane production, and fatty acid biohydrogenation. Molecules 27(13), 4288.
| Crossref | Google Scholar | PubMed |
Ozawa T, Lopez-villalobos N, Blair HT (2005) Dairy farming financial structures in Hokkaido, Japan and New Zealand. Animal Science Journal 76(4), 391-400.
| Crossref | Google Scholar |
Pacheco D, Waghorn GC (2008) Dietary nitrogen - definitions, digestion, excretion and consequences of excess for grazing ruminants. Proceedings of the New Zealand Grassland Association 70, 107-116.
| Crossref | Google Scholar |
Pickering AH, Gibbs JJA, Wear S, Fick J, Tomlin H (2024) Methodology for calculation of New Zealand’s agricultural greenhouse gas emissions. Ministry for Primary Industries, Manatū Ahu Matua: Wellington. Available at https://www.mpi.govt.nz/dmsdocument/13906-Detailed-methodologies-for-agricultural-greenhouse-gas-emission-calculation-
Pinxterhuis JB, Judson HG, Peterson ME, Navarrete S, Minnée E, Dodd MB, Davis SR (2024) Implementing plantain (Plantago lanceolata) to mitigate the impact of grazing ruminants on nitrogen losses to the environment: a review. Grass and Forage Science 79(2), 144-157.
| Crossref | Google Scholar |
R Core Team (2024) ‘R: a language and environment for statistical computing.’ (R Foundation for Statistical Computing: Vienna, Austria) Available at https://www.R-project.org/
Roche JR, Turner LR, Lee JM, Edmeades DC, Donaghy DJ, Macdonald KA, Penno JW, Berry DP (2009) Weather, herbage quality and milk production in pastoral systems. 4. Effects on dairy cattle production. Animal Production Science 49(3), 222-232.
| Crossref | Google Scholar |
Selbie DR, Buckthought LE, Shepherd MA (2015) Chapter four - the challenge of the urine patch for managing nitrogen in grazed pasture systems. In ‘Advances in agronomy, Vol. 129’. (Ed. DL Sparks) pp. 229–292. (Academic Press) 10.1016/bs.agron.2014.09.004
Stewart AV (1996) Plantain (Plantago lanceolata) – a potential pasture species. Proceedings of the New Zealand Grassland Association 58, 77-86.
| Crossref | Google Scholar |
Tamminga S (1992) Nutrition management of dairy cows as a contribution to pollution control. Journal of Dairy Science 75(1), 345-357.
| Crossref | Google Scholar |
Tamura Y, Nishibe S (2002) Changes in the concentrations of bioactive compounds in plantain leaves. Journal of Agricultural and Food Chemistry 50(9), 2514-2518.
| Crossref | Google Scholar | PubMed |
Ulyatt MJ, Lassey KR, Shelton ID, Walker CF (2002) Seasonal variation in methane emission from dairy cows and breeding ewes grazing ryegrass/white clover pasture in New Zealand. New Zealand Journal of Agricultural Research 45(4), 217-226.
| Crossref | Google Scholar |
Waghorn GC, Clark DA (2004) Feeding value of pastures for ruminants. New Zealand Veterinary Journal 52(6), 320-331.
| Crossref | Google Scholar | PubMed |
Weatherburn M (1967) Phenol-Hypochlorite reaction for determination of ammonia. Analytical Chemistry 39(8), 971-974.
| Crossref | Google Scholar |
Wilson RL, Bionaz M, Macadam JW, Beauchemin KA, Naumann HD, Ates S (2020) Milk production, nitrogen utilization, and methane emissions of dairy cows grazing grass, forb, and legume-based pastures. Journal of Animal Science 98(7), skaa220.
| Crossref | Google Scholar |