

Current issues in plant cryopreservation and importance for ex situ conservation of threatened Australian native species
Robyn Streczynski A F , Hamish Clark A F , Lily M. Whelehan A F , Sze-Tieng Ang A F , Lyndle K. Hardstaff A , Bryn Funnekotter A B , Eric Bunn A B C , Catherine A. Offord D , Karen D. Sommerville D and Ricardo L. Mancera A E
A B , Eric Bunn A B C , Catherine A. Offord D , Karen D. Sommerville D and Ricardo L. Mancera A E
A School of Pharmacy and Biomedical Sciences, Curtin Health Innovation Research Institute, Curtin University, GPO Box U1985, Perth, WA 6845, Australia.
B Kings Park Science, Department of Biodiversity, Conservation and Attractions, Kings Park, WA 6005, Australia.
C School of Biological Sciences, The University of Western Australia, Crawley, WA 6005, Australia.
D The Australian PlantBank, The Royal Botanic Gardens and Domain Trust, Mount Annan, NSW 2567, Australia.
E Corresponding author. Email: R.Mancera@curtin.edu.au
F Joint first authors.
Australian Journal of Botany 67(1) 1-15 https://doi.org/10.1071/BT18147
Submitted: 30 July 2018 Accepted: 6 February 2019 Published: 20 March 2019
Abstract
An alarming proportion of Australia’s unique plant biodiversity is under siege from a variety of environmental threats. Options for in situ conservation are becoming increasingly compromised as encroaching land use, climate change and introduced diseases are highly likely to erode sanctuaries regardless of best intentions. Ex situ conservation is currently limited to botanic garden living collections and seed banking, with in vitro and cryopreservation technologies still being developed to address ex situ conservation of species not amenable to conventional storage. Cryopreservation (storage in liquid nitrogen) has been used successfully for long-term biosecure storage of shoot tips of several species of threatened Australian plants. We present a case for building on this research and fostering further development and utilisation of cryopreservation as the best means of capturing critical germplasm collections of Australian species with special storage requirements (e.g. recalcitrant-seeded taxa and species with short-lived seeds) that currently cannot be preserved effectively by other means. This review highlights the major issues in cryopreservation that can limit survival including ice crystal damage and desiccation, toxicity of cryoprotective agents, membrane damage, oxidative stress and mitochondrial function. Progress in understanding and mitigating these stresses is vital for advancing cryopreservation for conservation purposes.
Additional keywords: cryobank, cryobiology, cryobiotechnology, cryostorage, freezing, in vitro conservation, plant tissue culture, vitrification.
Australian unique plant biodiversity under threat
Threats to Australian ecosystems and plant diversity include land clearing for agriculture and urban development, habitat fragmentation, changed water use, pollution (water, soil and air), natural disasters, illegal collection, loss of seed dispersal agents and the introduction of alien species (Engelmann 2004; Maunder et al. 2004; Li and Pritchard 2009; Williams et al. 2011; Berjak and Pammenter 2014; Merritt et al. 2014a; Millennium Seed Bank Partnership 2015; Broadhurst and Coates 2017; O’Donnell and Sharrock 2017; Sommerville et al. 2017). Introduced diseases, such as ‘root rot’ or ‘dieback’, caused by the soil-borne water mould Phytophthora cinnamomi, are linked to the decline of many rainforest and other species in the wet tropics (Williams et al. 2011; Sommerville et al. 2017) and south-west Western Australian eucalypt and banksia woodlands (Anderson et al. 2010; Duque-Lazo et al. 2016). A more recent invasive pathogen, myrtle rust (Austropuccinia psidii) (Beenken 2017), originating in South America and first observed in Australia in 2010, is of particular concern given the dominance of Myrtaceae in Australian ecosystems. A study by Carnegie et al. (2016) reported 232 potential host species for A. psidii in Australia, including a keystone wetland species (Melaleuca quinquenervia), with a further 115 species susceptible after artificial inoculation. Berthon et al. (2018) showed 1285 species to be at risk of exposure if the fungus continues its spread across Australia. Two major Australian plant biodiversity hotspots that contain a high percentage of rare or threatened plant (and animal) species most at risk include the Forests of East Australia, which also contain significant areas of rainforest that have persisted since Gondwanan times and sheltered species such as the endangered ‘living fossil’ Wollemia nobilis (Ashmore et al. 2011; Williams et al. 2011), and the south-west Australia hotspot with 5570 species of mainly endemic vascular plants including four endemic monospecific families: Ecdeiocoleaceae, Emblingiaceae, Eremosynaceae and Cephalotaceae (Conservation International 2018).
Conservation options for the Australian flora
Although in situ conservation of species and ecosystems is considered the simplest and most effective method of conservation, where such areas can be adequately protected (Prance 1997), in situ conservation alone cannot save all threatened species from the various threats outlined in the preceding paragraph. Ex situ conservation methods are necessary to provide support for in situ actions by preventing the extinction of species, providing a source of material for reintroduction or population enhancement of individual species, and providing a source of material for research to better understand the biological factors contributing to a species’ decline (Offord and Makinson 2009). A low cost, long-term method for ex situ conservation applicable to seed-bearing species is seedbanking (i.e. storage of seeds, after proper moisture equilibration, at –20°C). This option is applicable to a high proportion of species that possess orthodox (desiccation and low temperature tolerant) seed (Tweddle et al. 2003; Guerrant et al. 2014). Orthodox seeds generally have stable storage behaviour that allows drying, freezing, and storage at low temperatures over long periods of time (at least decades), after which germination and growth into healthy, reproductive adult plants is possible (Dickie and Pritchard 2002; Guerrant et al. 2014; O’Donnell and Sharrock 2017). However, not all seeds can be stored under conventional seed banking conditions. Seeds with special storage requirements are generally chilling sensitive, short lived, desiccation sensitive, or have a combination of those characteristics (Hay et al. 2010; Merritt et al. 2014b). Many of these seeds can be classified as recalcitrant, i.e. seeds that are unable to survive drying to low moisture contents (MC) and are sensitive to chilling (Ellis et al. 1990; Engelmann 2012). Most recalcitrant seeds studied to date are produced by evergreen rainforest tree or shrub species in moist tropical and subtropical zones. These seeds tend to be larger and have a higher water content than orthodox seeds, and are metabolically active and temperature sensitive; these characteristics make them unsuitable for storage by conventional seed banking methods (Tweddle et al. 2003; Li and Pritchard 2009; Hamilton et al. 2013; Funnekotter et al. 2017c). Many threatened species fall under the recalcitrant category and are thus much more suited to other ex situ storage methods such as in vitro (tissue) culture and/or cryopreservation (Ashmore et al. 2011).
Tissue culture is a technique used to grow plant cells, tissues and organs on synthetic media in an aseptic environment with controlled light, temperature and humidity conditions (Dagla 2012). Tissue culture allows a large amount of plant material to be produced from a small amount of starting material while maintaining key genotypic features of elite genotypes (or simply a limited number of existing plants in the case of threatened taxa), which may be lost in storage of genetically varying seed accessions (Ashmore et al. 2011; Bunn et al. 2011; Funnekotter et al. 2017c). However, tissue cultures require regular maintenance, may be accidentally contaminated via operator error, may accumulate somaclonal (or epigenetic) variation over many generations of continuous culture cycles, and may decline in viability over time (Larkin and Scowcroft 1981; Miguel and Marum 2011), and the presence of endophytes may hasten the death of the explants by availing more nutrients from either the plant culture medium or damaged tissues (Cassells 1991; Fouda et al. 2015). Hence other technologies are needed and cryo-storage or cryopreservation presents as a more attractive option for the long-term, biosecure storage of key germplasm collections (Benson 2008b).
Cryopreservation
Cryopreservation offers a superior long-term method of plant germplasm conservation that, in principle, overcomes the issues listed above for in vitro material, and also offers the only current alternative long-term storage option for recalcitrant-seeded species. Cryopreservation, in theory, allows indefinite storage of plant tissues in liquid nitrogen (LN), thereby maintaining the tissues at –196°C, with cellular metabolism effectively halted (Benson 2008b). Cryopreservation has been increasingly favoured as a plant conservation method in recent years as it requires very little storage space, minimal upkeep (i.e. periodic replenishing of LN), all but eliminates the risk of contamination, and is applicable to a wide range of plant tissues (Kaczmarczyk et al. 2012). It also mitigates the risk of any epigenetic variation in the stored tissues over time (Harding 2004). A wide range of plant tissues and organs can be cryopreserved including pollen, seeds, shoot tips, dormant buds, cell suspensions, embryonic cultures, somatic and zygotic embryos and callus tissue (Benelli et al. 2013; Engelmann 2012). The choice of propagule to use for cryo-storage is dependent on the conservation goal, with seeds and embryos capturing species diversity, whereas propagules like shoot tips and dormant buds capture specific genotypes (Reed 2008). Initial research on plant cryopreservation began in 1965, with a series of experiments investigating the use of the cryoprotective agent (CPA) dimethyl sulfoxide (DMSO) on mulberry twigs (Sakai 1965). Cryopreservation techniques have been steadily evolving and improving since these early beginnings.
Biophysics of cryo-storage
The development of cryopreservation protocols requires an understanding of water behaviour, cryo-injury and cryo-protection (Day et al. 2008). Temperature greatly influences the formation of the four most common states of water: liquid, glass, ice and vapour (Pukacki and Juszczyk 2015). The main goal that cryo-protocols aim to achieve for successful cryopreservation is the manipulation of water in its liquid, glassy and ice states, in order to avoid the formation of lethal intracellular ice crystals (Pukacki and Juszczyk 2015). Sufficient dehydration and cooling during cryo-storage ensures that the temperature drops rapidly below the glass transition temperature (Tg), resulting in the formation of an amorphous ‘glassy’ state (Shamblin et al. 1999; Walters 2015). The stability of this glassy state is related to the strength of the hydrogen-bonding network in water (Kreck and Mancera 2014). The advantages of the formation of this glassy state are that it (i) prevents cellular collapse; (ii) precludes chemical reactions requiring diffusion, ensuring stability during the storage period; and (iii) allows chaotropic solutes to be trapped by an amorphous glass which prevents them from becoming concentrated (Burke 1986; Moelbert et al. 2004).
Formation of ice crystals can occur during cryo-storage because the vitrified state is metastable. The behaviour of glasses in biological tissue is highly complex, and they can easily devitrify to form ice and/or revert back into a liquid (Benson 2008b; Day et al. 2008). When water molecules form intermolecular hydrogen bonds, the size of any ice crystals increases, so they become lethal to the cell (Walters 2015). Crystallisation can occur extracellularly or intracellularly depending on the rate of cooling. Rapid cooling results in intracellular ice formation, whereas slow cooling results in extracellular ice formation (Gonzalez-Arnao et al. 2007). Extracellular ice formation is responsible for osmotic shock injury (Benson 2008a). The formation of intracellular ice damages cell membranes directly as the growing ice crystals push apart and rupture membranes (Burke et al. 1976).
Desiccation of cells minimises the formation of ice crystals as the removal of water causes the cells to shrink and increases their solute concentration, in turn, increasing viscosity of the cytoplasm and improving the stability of the glassy state (Walters et al. 2010; Verhoeven et al. 2018). Successful cryopreservation protocols all centre on the controlled reduction of tissue water content to sufficiently low levels that the cell contents become concentrated and highly viscous, promoting the likelihood of vitrification taking place over ice crystal formation (Benson 2008a). Several measures can be incorporated into cryopreservation protocols to achieve this, including desiccation and pretreatment with CPAs (O’Donnell and Sharrock 2017).
Cryopreservation protocols
Cryopreservation is a multi-step process, involving (i) the initial excision of the germplasm such as seed embryos or shoot tips to obtain sufficiently small material; (ii) desiccation or preculture on osmotic media to reduce water content; (iii) cryoprotection through exposure to CPAs to promote vitrification; (iv) cryopreservation in LN; (v) re-warming; and finally (vi) the washing (unloading of CPA solutions) and recovery of the germplasm after cryopreservation (Fig. 1; Funnekotter et al. 2017b; Kaczmarczyk et al. 2012). There is wide variation in cryo-capability among species, and also among different clonal lines of the same species, therefore each species requires its own cryopreservation protocol to achieve the maximum possible post-cryogenic survival (Reed 2008). Different protocols need to be developed and optimised depending on the tissue and species to be cryopreserved, by experimentally altering factors such as desiccation conditions (preculture), mixtures of CPAs and recovery environments (Hamilton et al. 2009b).

|
CPAs can be classified into two main types that are often used in combination: non-penetrating, such as sucrose and polyethylene glycol; and penetrating (also known as ‘colligative’), such as DMSO and glycerol (Day et al. 2008; Berjak and Pammenter 2014). Non-penetrating CPAs do not cross cell membranes and remain external to the cells, causing potentially freezable water to be osmotically removed from the cells, thus increasing intracellular solute concentration (Pammenter and Berjak 2014). Penetrating CPAs cross cell membranes and act in the following various ways: (i) as agents that inhibit the formation and growth of intracellular ice by disrupting crystalline structure; (ii) as solutes that increase the number of particles dissolved in the intracellular water and, therefore, lower the freezing point; (iii) as diluents for intracellular electrolytes, which counteract the potentially toxic effects of increased concentrations of solute as water is removed; and (iv) as promoters of the glassy state (Berjak and Pammenter 2014). Molecular simulation studies have revealed the mechanism by which addition of CPAs to water promotes the formation of the glassy state of water (Kreck et al. 2011; Mandumpal et al. 2011).
The most commonly used combination of CPAs, Plant Vitrification Solution 2 (PVS2), was developed in 1990 by Sakai et al. (1990) and contains (in w/v) 30% glycerol, 15% DMSO and 15% ethylene glycol in Murashige-Tucker medium (however, Murashige-Skoog medium is now more commonly used) containing 0.15 M sucrose. One of the most widely used techniques incorporating CPAs today is droplet vitrification. This method was developed in 2004 by Panis et al. (2005) and involves treatment with CPAs before rapidly freezing each shoot tip in a droplet of vitrification solution and subsequently storing in LN. Using droplets rather than large volumes of CPAs contributes to higher cooling and re-warming rates, resulting in improved survival rates after cryopreservation (Sakai and Engelmann 2007). An alternative cryo-protocol, developed in the 1990s by Fabre and Dereuddre (1990) and known as encapsulation-dehydration, involves encapsulation of plant material within alginate beads which provide protection from mechanical damage. The beads are osmotically dehydrated in a concentrated sugar solution, then further dehydrated by silica gel or airflow to achieve sufficiently low water content for cellular vitrification, thereby avoiding the use of toxic CPAs (Fig. 1), before immersion in LN. A variation on this technique known as encapsulation-vitrification replaces the osmotic and air dehydration steps with cryoprotection using CPAs for more desiccation sensitive species (Sakai and Engelmann 2007).
Progress in the cryopreservation of Australian species
Most of the cryopreservation research for native species in Australia to date has focussed on Western Australian species due to the location of appropriate research facilities. There is, however, a range of herbaceous and woody species from different ecosystems represented in the literature (Table 1). Cryopreservation of other Australian species has previously been reported by Bunn et al. (2007), Kaczmarczyk et al. (2011) and Funnekotter et al. (2017b), while Merritt et al. (2014a) has reviewed cryopreservation success with seeds of various orchid species. Due to the high diversity of endemic species in Australia, little is known about the physiology of each species, resulting in slow progress in the development of new cryopreservation protocols. Further progress in understanding the fundamental requirements for successful cryopreservation and the development of novel protocols is vital to accelerate successful conservation of Australian species through cryopreservation.

|
Current issues in cryopreservation
Differential explant response
The materials most frequently used for cryopreservation of wild plants in Australia to date include tissue cultured shoot tips and whole seeds (Table 1). The use of excised seed embryos or embryonic axes (the root and shoot axis of the embryo with cotyledons removed) has, for the most part, been limited to commercially important recalcitrant and intermediate species such as almond, chestnut, coffee, hazelnut and rubber (Normah and Makeen 2008). Although both shoot tips and embryos are composed chiefly of meristematic cells, they can respond very differently to cryopreservation pretreatments and the processes of freezing and thawing. Shoot tips consist largely of apical meristem tissues surrounded by leaf primordia and subtended by partially differentiated cells forming the primary meristems, i.e. ground meristem (that will produce the stem pith), procambium (that will produce the vascular system) and protoderm (that will produce the epidermis) (Evert and Eichhorn 2013). Shoot tips – at least those produced in tissue culture – are generally intolerant of desiccation unless pre-treated with suitable cryoprotective agents (Volk and Walters 2006). Embryonic axes are composed largely of primary meristems with cell characteristics and tissue arrangement related to the mature tissues they will eventually form (Esau 1977). In addition to an apical meristem, the embryonic axis has a root meristem, which in some embryos may have developed further into a radicle (a primordial root), while the apical meristem may have developed further into a plumule (a stem-like axis with one or more leaves surrounding the apical meristem) (Evert and Eichhorn 2013). In some taxa, the embryonic axis has the capacity to survive rapid desiccation even though the whole seed is sensitive to drying (Normah and Makeen 2008). In other taxa, the axis may be just as sensitive to drying as a shoot tip (Pammenter and Berjak 2014; Sommerville et al. 2017).
There have not, as yet, been any published studies comparing desiccation-tolerant and desiccation-sensitive embryonic axes in terms of ultrastructure, membrane composition or other cellular components. Likewise, there have been no studies comparing the effect of cryopreservation on the ultrastructure of embryonic axes with shoot tips of the same species. There have, however, been several ultrastructural studies focussed on a single tissue type for individual species and these may provide some clues to the mechanisms at work. Wesley-Smith et al. (2015), for example, observed that cryopreservation of fresh, untreated, embryonic axes of Acer saccharinum resulted in far greater damage to the highly vacuolated cells of the plumule than to the cells of the radicle. The apical and root meristems were both lethally damaged and regrowth occurred only from the primary meristems – from ground meristem and procambium in the radicle, and from pockets of ground meristem and protoderm in the plumule. Similarly, in a study on the effects of cryopreservation on potato (Solanum tuberosum) shoot tips pre-treated with DMSO, Kaczmarczyk et al. (2008) observed extensive post-cryopreservation damage in the meristematic dome and part of the epidermis. Survival and regeneration occurred mainly in the leaf primordial tissues. These results suggest that material with a higher proportion of partially differentiated tissues may be more tolerant of cryopreservation than undifferentiated meristem. However, in a study of cryopreservation of yam (Dioscorea alata) shoot tips by encapsulation-dehydration, Barraco et al. (2014), found that only cells in the meristematic zone survived. Similarly, Volk and Caspersen (2007) found that recovery of cryopreserved Mentha × piperita shoot tips pre-treated with PVS2 occurred from surviving clumps of cells in the outer layers of the original meristem or in the axil of the shoot and leaf primordia, whereas Helliot et al. (2003) observed that recovery of banana meristems pre-treated with PVS2 occurred only from small areas of surviving cells in the leaf primordial tissues and in the tunica of the meristematic dome. These results may indicate that different cryoprotective treatments favour different tissues, or may be an indication of species-dependent differences in cell structure and composition.
Dormant buds are a possible alternative source of material for cryopreservation not yet tested in Australia. Dormant buds or scions holding dormant buds have been successfully used for preservation of genetic material in forest, fruit, nut and ornamental species (Towill and Ellis 2008; Benelli et al. 2013). The method used for cryopreservation is dependent on the species as well as on the level of cold-hardiness of the collected material (Towill and Ellis 2008). For successful cryopreservation of dormant buds, the most important factor identified to date is the extent of cold acclimation that the species can attain, and the extent of acclimation that the material possesses at the time of collection (Towill and Ellis 2008). Regrowth after cryopreservation can be achieved by direct grafting of the bud or micrografting of the excised shoot tip from the dormant bud (e.g. apple), by culture of the shoot tip on suitable medium (e.g. persimmon), or by direct rooting of the cryopreserved scion (e.g. willow) (Towill and Ellis 2008; Benelli et al. 2013). However, only 0.15% of the Australian continent experiences winter snow-cover (Pickering et al. 2008), and thus there are unlikely to be many species that are sufficiently cold tolerant to be suitable for this method.
Cryoprotective agent toxicity
The successful use of CPAs requires exposure to these agents in an adequate concentration and for a sufficiently long time to achieve dehydration and ultimately, vitrification. Volk and Walters (2006) showed that the extent of PVS2 penetration into garlic and mint shoot tips was directly proportional to the exposure time, and a significant decrease in water content was seen with increased exposure time to PVS2. The effect of CPAs on cellular ice formation can be measured through the use of thermal analysis via differential scanning calorimetry (DSC), with extended exposure times significantly reducing the formation of ice crystals (Benson 2008a; Menon et al. 2012). CPAs, however, can be toxic to cells, such that limiting the concentration of CPAs to reduce toxicity is a major barrier to cryopreservation by vitrification (Best 2015). CPAs are deemed toxic if they cause breaches or damage in the cell membrane, impair the function of enzymes, diminish cell development, and damage DNA, proteins or macromolecules (Best 2015). The high concentration of CPAs causes an exchange of intracellular water with the external environment, creating acute osmotic stress (Lai et al. 2015), with the resulting rapid efflux of water from cells causing them to collapse (Fuller 2004). Molecular simulation of model cell membranes has shown that addition of CPAs at high concentrations can damage membrane integrity, ultimately causing the formation of water pores (Fig. 2) and the subsequent destruction of the cell membrane (Hughes et al. 2012; Hughes et al. 2013; Hughes and Mancera 2013; Malajczuk et al. 2013). However, this damaging effect is reduced when CPAs are used in mixtures, such as in PVS2 (Hughes and Mancera 2014).

|
Various approaches have been developed to reduce CPA toxicity, including their sequential ‘loading’ and ‘unloading’ to prevent osmotic shock (Day et al. 2008). The temperature and timing of CPA treatments are critical in achieving sufficient cryoprotection, while avoiding toxicity and damage to cells (Kaczmarczyk et al. 2012; Pence 2014). In the development of cryopreservation protocols for Musacae (Panis et al. 2005) and Anigozanthos viridis (Turner et al. 2000), a 5 or 10 min difference in CPA incubation times was observed to translate to a change in survival rate of around 20%. The consequence of the damaging effects of CPAs for cryopreservation protocols is that optimising CPA incubation time is essential for post cryopreservation survival. If plant tissues are incubated insufficiently, ice crystals will damage cells, but just as much damage can occur with longer incubation times due to osmotic stress and CPA toxicity (Turner et al. 2000; Panis et al. 2005).
The role of cell membranes in cryopreservation
The fundamental purpose of the cell membrane is to form a stable but fluid barrier that is semi-permeable between two aqueous compartments: the intra- and extracellular environments. Cell membranes are considered one of the primary sites of cryo-injury. During cryopreservation, water stress damage occurs due to changes in intracellular and extracellular water concentrations caused by desiccation or freezing. This results in external osmotic pressure on cells, forcing the cells to shrink in volume (Moussa et al. 2008). The maintenance of membrane integrity and stability under water stress is considered a major factor in drought tolerance (Bajji et al. 2002). In cryopreservation protocols, plant material is desiccated on culture media containing a high concentration of sugars. This removal of water can mimic the effects of water stress on the cellular membrane. There are two widely accepted hypotheses around the effect of water stress on membranes: (i) that when electrolyte concentration reaches a critical level, the cell membrane is damaged, and the cells die from osmotic shock (Lovelock 1953); and (ii) that membrane damage occurs because cells are unable to shrink enough to maintain osmotic equilibrium (Meryman 1968). Additionally, the process of cryopreservation can result in physical membrane damages, such as (i) expansion-induced lysis, where overexpansion of cells occurs due to increased extracellular osmotic pressure during thawing/warming; (ii) loss of osmotic responsiveness, where due to a slow cooling rate there is no osmotic change during warming (i.e. cells stay dehydrated); (iii) altered osmotic behaviour, where water and solutes are released into the surroundings because cell membranes become ‘leaky’; and (iv) intracellular ice crystal formation due to rapid cooling that causes membrane disruption. (Steponkus 1984).
Minimising cryo-injury depends on membrane stabilisation, typically achieved through preconditioning, causing changes in lipid composition in cell membranes, accumulation of sugars and production of membrane-protecting polypeptides (Thomashow 1999). Preconditioning is the method of exposing plants to an abiotic stress before cryopreservation to alter its cellular properties (Kaczmarczyk et al. 2012). Temperature preconditioning is commonly used as a method to increase cell membrane stability at low temperatures through increased levels of unsaturated acyl chains (Menon et al. 2014). Studies on two Australian species (Grevillea scapigera and Loxocarya cinerea) exposed to 20/10°C alternating temperature preconditioning resulted in a decrease in common acyl chains (16 : 0, 18 : 0, 18 : 1 and 18 : 2) with an increase in α-linoleyl tails (18 : 3) (Funnekotter et al. 2013). Larger concentrations of the more unsaturated α-linoleyl tails induce a greater level of membrane molecular disorder, leading to increased membrane fluidity, stabilisation of the lamellar phase and maintenance of diffusion across membranes when subjected to reduced temperatures and desiccation. An increase in double bonds within phospholipids (PLs) was also seen in both species; however, these changes only correlated with increased cryopreservation survival in one (G. scapigera). Studies on cryopreservation of banana meristems have also seen positive correlation between unsaturation and cryopreservation survival (Zhu et al. 2006). Higher concentrations of double bonds are generally thought to be beneficial in dehydration and cooling by reducing the liquid crystal-to-gel transition temperature through increased membrane fluidity (Kong and von Aderkas 2011). However, too high a concentration of double bonds may not be beneficial as the membrane becomes more susceptible to free radicals acting on electron rich double bonds and causing lipid peroxidation (Benson 1990).
Oxidative stress damage in cryopreservation
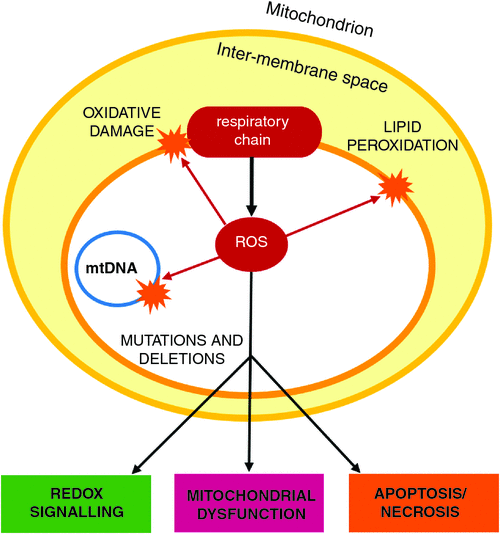
Plant tissues are susceptible to oxidative stress, caused primarily by reactive oxygen species (ROS) (Benson and Bremner 2004; Kaczmarczyk et al. 2012). ROS are oxygen-based molecules with an unstable number of electrons, making them highly reactive (Murphy 2009). Mitochondria are generally accepted as the major producers of ROS in the cell (Murphy 2009; Andreyev et al. 2015), although chloroplasts and peroxisomes also produce ROS in plants (Taylor et al. 2004; Suzuki et al. 2012). The accumulation of these ROS results in an imbalance of redox reactions within cells, resulting in oxidative stress and several other deleterious effects, potentially resulting in cell apoptosis (Fig. 3) (Berjak and Pammenter 2014).
|
Oxidative stress constitutes a major component of cryo-injury and is a major obstacle for successful cryopreservation (Benson and Bremner 2004; Chetverikova 2012; Funnekotter et al. 2017c). Each of the steps in the vitrification technique of cryopreservation (excision, pre-culture, loading solution, CPA treatment, cooling, rewarming, washing and regrowth) presents the possibility of oxidative stress, due to physical damage and osmotic stress involved in the process, and many studies have shown this (Uchendu et al. 2010). Hydrogen peroxide (H2O2) and superoxide production has been observed during excision, desiccation, exposure to LN, and rewarming and recovery (Johnston et al. 2007; Fang et al. 2008; Whitaker et al. 2010; Berjak et al. 2011; Skyba et al. 2012). During the desiccation stage, Johnston et al. (2007) found Ribes ciliatum produced more than three times the amount of ethylene (an indicator of oxidative membrane damage) as Ribes nigrum. Whitaker et al. (2010) showed that the tropical tree species Trichilia dregeana produced twice as much superoxide as Castanea sativa, and for a greater duration. One study found that H2O2 production in Hypericum perforatum persisted for several weeks after cryopreservation (Skyba et al. 2012). Roach et al. (2008) studied ROS formation during the first steps of the cryopreservation protocol, specifically wounding and desiccation, and showed that excision of embryonic axes from fresh seeds of recalcitrant sweet chestnut (C. sativa) induced a burst of extracellular superoxide in the first 5 min following excision. The isolated axes were subjected to varying levels of desiccation stress, and the combined effects of wounding and desiccation from 60 to 30% MC, were found to double the initial rates of extracellular superoxide production (Roach et al. 2008). These results indicated that there is a complex interaction between excision and subsequent drying of the isolated axes, which showed a reduction in viability and vigour and increase in an electrolyte leakage, which suggests impaired membrane integrity (Roach et al. 2008). Similar results were found by Whitaker et al. (2010) using recalcitrant seeds from the tropical tree species Trichilia dregeana.
Lipid peroxidation
ROS interact with cellular biomolecules, and can cause serious oxidative damage to proteins, nucleic acids and lipids, with lipid peroxidation in particular leading to disruption of membranes that are critical to cell survival (Sies and Cadenas 1985; Uchendu et al. 2010; Halliwell and Gutteridge 2015). Lipid peroxidation results from the oxidation of PLs and other unsaturated lipids when the production of ROS overwhelms the ability of scavenging antioxidant defence systems to protect cells (Rachmilevitch et al. 2006). Lipid peroxidation is concerning as it affects the integrity of membrane structure, and alters its functions, which leads to cell death (Uchendu et al. 2010).
Some of the by-products of lipid peroxidation, such as the aldehydes, are highly reactive and may be regarded as secondary toxic messengers, which spread and cause an increase in initial free radical events (Catalá 2006). The aldehydes that have been most intensively studied so far are malondialdehyde (MDA) and 4-hydroxy-2-nonenal (4-HNE), with the identification of their formation commonly used as an indicator of oxidative stress (Kaczmarczyk et al. 2012). Due to its carbonyl functional groups, MDA is chemically reactive; however, the toxicity of MDA is debatable, as its effects were previously overestimated and it has been found to be less reactive than HNE (Halliwell and Gutteridge 2015; Tsikas 2017). MDA can polymerise easily and undergoes various reactions with proteins (leading to loss of function) and DNA (leading to mutations) (Halliwell and Gutteridge 2015; Tsikas 2017). However, HNE formation has greater toxicity to cells than MDA, as it can cause mitochondrial damage, inhibit DNA and protein synthesis, and inhibit the action of chaperone repair proteins (Halliwell and Gutteridge 2015).
Ren et al. (2013) studied how peroxidation due to CPA treatment affects Arabidopsis thaliana seedling survival and found that MDA content increased after desiccation and cryoprotection (with PVS2) treatment in both 48 and 72 h (cultured) seedlings. Their data showed that high levels of MDA in seedlings reduced the capacity for regrowth following cryogenic treatment, with the 72 h seedlings (MDA increased almost 2-fold) suffering serious membrane damage from lipid peroxidation and subsequent cell death (Ren et al. 2013). Chen et al. (2015) also examined the survival tolerance of A. thaliana seedlings, which were germinated for 48 or 72 h, at five steps of cryopreservation (desiccation, cryoprotection, rapid warming, unloading and recovery), in order to determine the role of ROS (superoxide, H2O2 and OH•) in cryo-injury. The main cause of cryo-injury in the 48 h seedlings and 72 h seedlings (no survival), was that H2O2 (generated mainly in cotyledons, shoot tips and roots) induced oxidative stress at the cryoprotection step (with PVS2) and rapid rewarming steps (Chen et al. 2015). MDA content was higher throughout the steps in the 72 h seedlings but showed a similar trend to that seen in the 48 h seedlings (Chen et al. 2015). Other studies also found increases in MDA content, including a 3-fold increase in MDA detected during the desiccation and freezing of maize embryos (Wen et al. 2010), and substantially more MDA detected at each step of the cryopreservation of untreated blackberry shoot tips compared with shoot tips treated with antioxidants (Uchendu et al. 2010). The initial recovery period following cryopreservation is a critical stage: increased levels of oxidative stress have been linked to a decrease in post-cryogenic regrowth, with increased detection of superoxide burst during rehydration of cryopreserved Trichilia dregeana (Whitaker et al. 2010), increased MDA content in A. thaliana during 24 h recovery (Chen et al. 2015), and detection of 4-HNE and hydroperoxide in Coffea arabica (coffee), indicating lipid peroxidation during the first hour of seed rehydration following desiccation (Dussert et al. 2003). The above indicate lipid peroxidation can cause substantial damage to cryopreserved plant material unless controlled with remedial or mitigating treatments (e.g. antioxidants).
Impact of cryopreservation on mitochondrial function
Although there are few studies on plant mitochondria in cryopreservation, studies on how plant cells in general are physically affected by cryopreservation have been conducted. Vacuolisation, abnormalities and rupture of the nuclear envelope, cell lysis, and autophagic decomposition were all observed following exposure to LN (Helliot et al. 2003; Wesley-Smith et al. 2015). Helliot et al. (2003) determined that this damage occurred during either osmotic dehydration using PVS2 or freezing and thawing. The finding by the same study that surviving cells (1 week post-thaw) showed that the presence of numerous organelles, including mitochondria, is of particular significance as it suggests a high level of metabolic activity in the plant cells during the recovery period (Helliot et al. 2003). Despite the evidence that suggests cryopreservation often causes damage to mitochondria in animal cells, as well as to plant cells as a whole, Skyba et al. (2012) found that exposure to cryogenic temperatures in the plant species H. perforatum caused no significant observable structural damage to mitochondria, although they did not test for impacts on mitochondrial function. Ren et al. (2013) found that cryogenic treatment caused the downregulation of transporters related to oxidative phosphorylation in A. thaliana seedlings, which also suggests that the negative effect seen during cryopreservation on animal cell mitochondrial function may also apply to plant cells. However, the full extent (and duration) of this damage, and whether or not mitochondria can repair cryo-induced damage and resume normal function, is still unresolved.
Recovery medium
Recovery methods are an often overlooked but crucial aspect of cryopreservation protocols (Reed 2008). Optimal recovery methods vary greatly among species. Although most protocols employ a standard basal medium for recovery from cryopreservation (Gonzalez-Arnao and Engelmann 2006), several studies have found altering the contents of the medium to be beneficial. The addition of growth regulators, for example, stimulates shoot production in some species (Paulet et al. 1993). Uchendu et al. (2013) found that the addition of melatonin to the recovery medium improved regrowth of American elm when used in conjunction with a melatonin preculture stage. Conversely, some species benefit from the removal of growth hormones such as benzyl adenine (BA) from the recovery medium (Chang and Reed 1999; Gupta and Reed 2006), which can reduce callus formation, thus maintaining genetic integrity.
The addition of growth regulators to recovery media is common for cryopreservation of many Australian species (Touchell et al. 2002; Menon et al. 2012; Whiteley et al. 2016). Shoot tips of Grevillea scapigera, for example, fully recovered only in the presence of zeatin whereas, in the same study, Anigozanthos viridis showed greatest recovery levels with the addition of choline chloride (Touchell et al. 2002). Environmental conditions are also an important aspect of recovery protocols. Both A. viridis and G. scapigera showed increased recovery rates when stored in darkness for 7 and 14 days following thawing respectively (Touchell et al. 2002). Sugarcane and cassava protocols also include a period of darkness during recovery (Paulet et al. 1993; Escobar et al. 1997). However, both Carica papaya and Vasconcellea pubescens showed decreased recovery with the exclusion of light for any period of time after cryopreservation (Ashmore et al. 2007).
Progress in understanding and ameliorating cryo-injury
Role of antioxidants
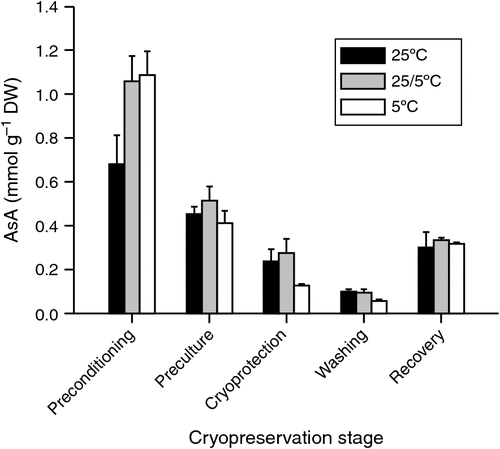
Plants have complex antioxidant systems, which are their natural defence against the accumulation of ROS and their by-products, and which enable them to resist external stress (Chen et al. 2015; Funnekotter et al. 2017b). Although antioxidants within the cells can balance the build-up of ROS (Mittler 2002), it has been shown that concentrations of antioxidants like glutathione (GSH) and ascorbic acid (AsA) decrease in plant tissues with each successive step of the cryopreservation process (Fig. 4) (Funnekotter et al. 2017b). Although Chen et al. (2015) also found a reduction in GSH and AsA antioxidants in thawing and recovery stages, levels of these antioxidants were found to increase in the desiccation period. However, the same study also showed a decrease in superoxide dismutase (SOD) over all stages of the cryopreservation process (Chen et al. 2015). Funnekotter et al. (2016) also examined the influence of abiotic stress preconditioning on antioxidant enzymes in shoot tips of an Australian species, Lomandra sonderi, before cryo-storage. A weak positive correlation was observed between SOD activity and post-cryogenic survival; however, the activities of glutathione reductase, glutathione peroxidase and catalase showed no correlation. Analysis of the antioxidants found that glutathione reductase activity decreased significantly after cryopreservation, but glutathione peroxidase and catalase did not change (Funnekotter et al. 2016).
|
The damaging effects of ROS formation can be modulated in two ways: through treatment to reduce ROS production, or by exogenous application of antioxidants which can exert a direct effect or may enhance endogenous antioxidant activity (Berjak et al. 2011). Several studies have shown that the application of exogenous antioxidants during the cryopreservation process can increase survival and regeneration post-cryopreservation, such as the use of vitamin C and E at critical steps during vitrification, which significantly improved regrowth of blackberry shoot tips following exposure to LN (Uchendu et al. 2010), and the use of low concentrations of glutathione in the preculture step of cryopreservation of citrus shoot tips, which was beneficial in post-thaw recovery (Wang and Deng 2004). Alternatives to antioxidants can also be used to mitigate oxidative stress damage to plant cells during cryopreservation. The use of desferrioxamine (a powerful Fe3+ chelator which interferes with OH• generation from H2O2 in the Fenton reaction), for example, promoted post-thaw recovery after cryopreservation of rice cell cultures (Benson et al. 1995), whereas cathodic water has been used to reduce oxidative stress and promote shoot development after cryopreservation of Strychnos gerrardii axes (Berjak et al. 2011).
Transcriptome profiling
A more recent area of study is that of transcriptome changes that occur due to the stresses imposed throughout cryopreservation. Studies by Ren et al. (2015) and Gross et al. (2017) explored the regulation of transcripts in response to exposure to CPAs and LN on A. thaliana. Ren et al. (2015) primarily looked at the effect of addition of vitamin C to PVS2, and its effect on genes related to ROS stresses. It was seen that, compared with controls, plant tissue incubated in PVS2 with vitamin C upregulated genes associated with antioxidant formation as well as genes associated with ROS-scavenging. During osmo-protection, receptors for drought, osmotic, cold and salt stresses were upregulated. Gross et al. (2017) characterised transcriptional regulation due to exposure to CPAs, PVS2 or PVS3 before cryopreservation, compared with a control of exposure to LN only. They found that 180 transcripts exhibited modified expression due to treatment with CPAs or cryopreservation. Of these, 67 were related to general stress responses, defence, wounding, lipid and carbohydrate production, abscisic acid regulation, oxidation, temperature stress and osmoregulation. It was observed that PVS2-regulated genes were involved in an oxidative response, whereas transcripts of PVS3-regulated genes were involved in a more general metabolic response. Transcriptional analysis allows for in-depth understanding of the stresses imposed on plant material during cryopreservation protocols and may help to identify changes in protocols that could result in greater survival.
Advancements in cryopreservation protocols
The vitrification protocol is currently the most commonly used cryopreservation technique, and hundreds of plant species have been successfully cryopreserved in this way (Sakai and Engelmann 2007). The vitrification method has continued to develop and improve over time, with approaches such as vacuum infiltration vitrification (VIV) (Nadarajan and Pritchard 2014; Funnekotter et al. 2015), cryo-plate protocols (Yamamoto et al. 2011, 2012), and a new cryo-mesh (Funnekotter et al. 2017a) protocol created to increase success in cryopreservation.
VIV as an alteration to cryopreservation protocols involves applying a vacuum to plant material during the incubation phases, whether that involves loading solutions, CPA solutions or a combination of both. The vacuum is thought to reduce trapped air bubbles on the surface of plant material, thus increasing the surface interactions between plant membranes and the surrounding solution and resulting in a more uniform infiltration and a potentially greater intake of CPAs (Nadarajan and Pritchard 2014). Using the VIV method, Nadarajan and Pritchard (2014) achieved greater post cryopreservation viability and regrowth in embryos of Carica papaya, Passiflora edulis and Laurus nobilis seeds, with a 10-fold reduction in PVS2 exposure times at both pretreatment temperatures of 0 and 25°C. Funnekotter et al. (2015) also investigated the use of VIV as an addition to cryopreservation protocols on shoot tips of species native to Western Australia. It was observed that the use of a vacuum during PVS2 incubation resulted in increased post-cryopreservation survival, along with a reduction in CPA incubation times and regeneration times. Of great interest were shoot tips of L. cinerea, which did not achieve shoot tip regeneration under a standard protocol but achieved up to 10% regeneration with the addition of vacuum conditions during PVS2 incubation.
The cryo-plate protocol developed by Yamamoto et al. (2011) is a modification of the encapsulation-vitrification protocol. The cryo-plate is an aluminium strip with wells of set diameter in which excised shoot tips can be placed. The excised shoot tips are embedded in wells with alginate to secure them to the well. Having multiple shoot tips attached to a cryo-plate helps to reduce mechanical damage through reduction of direct handling of shoot tips during the transition between solutions and LN (Yamamoto et al. 2011). The cryo-plate protocol has been further optimised, and in the case of Mentha spp. Yamamoto et al. (2012) achieved post-cryopreservation regrowth between 73–100% across 16 species. Funnekotter et al. (2017a) investigated an alternative to the cryo-plate protocol in which an aluminium mesh, deemed cryo-mesh, was used in place of the cryo-plate with fixed well sizes. Cryo-plates require specific and consistent production of wells or shoot tips may not be able to adhere. Cryo-mesh overcomes this by allowing the alginate to act as a glue between the mesh and the shoot tips (Fig. 5) (Funnekotter et al. 2017a). Cryo-mesh efficacy was tested in comparison to droplet-vitrification with cryopreservation of the West Australian native species A. viridis. No significant difference in post-cryogenic generation was observed between the two protocols: 78% for droplet-vitrification and 83% for cryo-mesh. Cryo-mesh, however, showed efficacy in reducing the manufacturing precision required over cryo-plates, as well as reducing the operator skill required in comparison to the droplet-vitrification protocol (Funnekotter et al. 2017a).

|
Meristem shoot tips from tissue cultured plants are commonly used in cryopreservation and are desiccated by osmotic means; however, excised zygotic embryos and embryonic axes are the preferred materials for cryopreservation as they allow for whole plants to be generated without complex tissue culture protocols (Ballesteros et al. 2014). It is known that the partial drying of material pre-cryopreservation is pivotal to survival. However, the loss of freezable water in recalcitrant species affects metabolism, with damage being dependent on the rate at which water is removed (Varghese et al. 2011). New desiccation methods, such as flash drying, followed by addition of CPAs, have been shown to reduce water contents further than slow drying without loss of survival (Wesley-Smith et al. 2001). The positive effect of flash drying has been shown in embryonic axes of jackfruit (Artocarpus heterophyllus) in which Wesley-Smith et al. (2001) found that axes that underwent rapid drying (<90 min) maintained 100% survival at WC of 0.4 g g–1 DW, in contrast to total mortality of embryonic axes brought down to this WC by 2–3 days of osmotic desiccation. Ballesteros et al. (2014) looked at the effect of flash drying on various species in relation to dehydration rates. Large differences between species were observed, with dehydration between 18–92% after 30 min at constant air flow. It was reported that, in general, the species with the highest rate of drying exhibited the highest rate of post-cryopreservation survival. Flash drying provides a novel technique that has the potential to be incorporated into cryopreservation protocols with Australian recalcitrant-seeded species, reducing desiccation times without having to risk additional stress and damage due to exposure to toxic CPAs or highly osmotic solutions.
Conclusions
The diversity and complex range of different physiologies exhibited by Australian plants, especially in regard to recalcitrant-seeded species (and other groups that have unusual seed characteristics), are major impediments to seed banking as the sole means of ex situ plant conservation. In addition, the time available to develop effective ex situ germplasm conservation for threatened species (and indeed the wider floras of threatened plant communities) before they become extinct is likely to shrink even further with the added threats of diseases such as myrtle rust. Cryopreservation is currently the only viable alternative for long-term biosecure germplasm storage of seeds from species that cannot be seed-banked effectively. Progress has been made in developing a more holistic understanding of the stresses that cryopreservation imposes on plant germplasm, and in developing novel cryopreservation protocols to counteract them. This has greatly improved the scope for conservation of rare, threatened and recalcitrant Australian species. However, this progress requires continued dedicated research to enable further development of viable and efficient cryopreservation protocols applicable to species in highly biodiverse but threatened ecosystems, such as Australian sub-tropical and temperate rainforest remnants, where a relatively high proportion of species possess recalcitrant seeds, and for species with special storage requirements such as the short-lived seeds of orchids. Failure to appreciate the urgency of the precarious future of many threatened Australian plant taxa will inevitably hasten their permanent demise from the unique palette of our biodiversity heritage.
Conflict of interest
The authors declare no conflicts of interest.
Acknowledgement
This work was supported by grant LP160101496 from the Australian Research Council (ARC).
References
Anderson P, Brundrett M, Grierson P, Robinson R (2010) Impact of severe forest dieback caused by Phytophthora cinnamomi on macrofungal diversity in the northern jarrah forest of Western Australia. Forest Ecology and Management 259, 1033–1040.| Impact of severe forest dieback caused by Phytophthora cinnamomi on macrofungal diversity in the northern jarrah forest of Western Australia.Crossref | GoogleScholarGoogle Scholar |
Andreyev A, Kushnareva Y, Murphy A, Starkov A (2015) Mitochondrial ROS metabolism: 10 years later. Biochemistry 80, 517–531.
| Mitochondrial ROS metabolism: 10 years later.Crossref | GoogleScholarGoogle Scholar | 26071769PubMed |
Ashmore SE, Drew RA, Azimi M (2007) Vitrification-based shoot tip cryopreservation of Carica papaya and a wild relative Vasconcellea pubescens Australian Journal of Botany 55, 541–547.
| Vitrification-based shoot tip cryopreservation of Carica papaya and a wild relative Vasconcellea pubescens Crossref | GoogleScholarGoogle Scholar |
Ashmore SE, Hamilton KN, Offord CA (2011) Conservation technologies for safeguarding and restoring threatened flora: case studies from Eastern Australia. In Vitro Cellular & Developmental Biology. Plant 47, 99–109.
| Conservation technologies for safeguarding and restoring threatened flora: case studies from Eastern Australia.Crossref | GoogleScholarGoogle Scholar |
Bajji M, Kinet J-M, Lutts S (2002) The use of the electrolyte leakage method for assessing cell membrane stability as a water stress tolerance test in durum wheat. Plant Growth Regulation 36, 61–70.
| The use of the electrolyte leakage method for assessing cell membrane stability as a water stress tolerance test in durum wheat.Crossref | GoogleScholarGoogle Scholar |
Ballesteros D, Sershen , Varghese B, Berjak P, Pammenter NW (2014) Uneven drying of zygotic embryos and embryonic axes of recalcitrant seeds: challenges and considerations for cryopreservation. Cryobiology 69, 100–109.
| Uneven drying of zygotic embryos and embryonic axes of recalcitrant seeds: challenges and considerations for cryopreservation.Crossref | GoogleScholarGoogle Scholar | 24909952PubMed |
Barraco G, Sylvestre I, Collin M, Escoute J, Lartaud M, Verdeil J-L, Engelmann F (2014) Histocytological analysis of yam (Dioscorea alata) shoot tips cryopreserved by encapsulation-dehydration. Protoplasma 251, 177–189.
| Histocytological analysis of yam (Dioscorea alata) shoot tips cryopreserved by encapsulation-dehydration.Crossref | GoogleScholarGoogle Scholar | 23926078PubMed |
Beenken L (2017) Austropuccinia: a new genus name for the myrtle rust Puccinia psidii placed within the redefined family Sphaerophragmiaceae (Pucciniales). Phytotaxa 297, 53–61.
| Austropuccinia: a new genus name for the myrtle rust Puccinia psidii placed within the redefined family Sphaerophragmiaceae (Pucciniales).Crossref | GoogleScholarGoogle Scholar |
Benelli C, De Carlo A, Engelmann F (2013) Recent advances in the cryopreservation of shoot-derived germplasm of economically important fruit trees of Actinidia, Diospyros, Malus, Olea, Prunus, Pyrus and Vitis. Biotechnology Advances 31, 175–185.
| Recent advances in the cryopreservation of shoot-derived germplasm of economically important fruit trees of Actinidia, Diospyros, Malus, Olea, Prunus, Pyrus and Vitis.Crossref | GoogleScholarGoogle Scholar | 23022736PubMed |
Benson EE (1990) ‘Free radical damage in stored plant germplasm.’ (International Board for Plant Genetic Resources: Rome)
Benson EE (2008a) Cryopreservation of phytodiversity: a critical appraisal of theory & practice. Critical Reviews in Plant Sciences 27, 141–219.
| Cryopreservation of phytodiversity: a critical appraisal of theory & practice.Crossref | GoogleScholarGoogle Scholar |
Benson EE (2008b) Cryopreservation theory. In ‘Plant cryopreservation: a practical guide’. (Ed. BM Reed) pp. 15–32. (Springer: New York)
Benson EE, Bremner D (2004) Oxidative stress in the frozen plant: a free radical point of view. In ‘Life in the frozen state’. (Eds BJ Fuller, N Lane, EE Benson) pp. 205–242. (CRC Press: Boca Raton, FL, USA)
Benson EE, Lynch PT, Jones J (1995) The use of the iron chelating agent desferrioxamine in rice cell cryopreservation: a novel approach for improving recovery. Plant Science 110, 249–258.
| The use of the iron chelating agent desferrioxamine in rice cell cryopreservation: a novel approach for improving recovery.Crossref | GoogleScholarGoogle Scholar |
Berjak P, Pammenter NW (2014) Cryostorage of germplasm of tropical recalcitrant-seeded species: approaches and problems. International Journal of Plant Sciences 175, 29–39.
| Cryostorage of germplasm of tropical recalcitrant-seeded species: approaches and problems.Crossref | GoogleScholarGoogle Scholar |
Berjak P, Varghese B, Pammenter NW (2011) Cathodic amelioration of the adverse effects of oxidative stress accompanying procedures necessary for cryopreservation of embryonic axes of recalcitrant-seeded species. Seed Science Research 21, 187–203.
| Cathodic amelioration of the adverse effects of oxidative stress accompanying procedures necessary for cryopreservation of embryonic axes of recalcitrant-seeded species.Crossref | GoogleScholarGoogle Scholar |
Berthon K, Esperon-Rodriguez M, Beaumont LJ, Carnegie AJ, Leishman MR (2018) Assessment and prioritisation of plant species at risk from myrtle rust (Austropuccinia psidii) under current and future climates in Australia. Biological Conservation 218, 154–162.
| Assessment and prioritisation of plant species at risk from myrtle rust (Austropuccinia psidii) under current and future climates in Australia.Crossref | GoogleScholarGoogle Scholar |
Best BP (2015) Cryoprotectant toxicity: facts, issues, and questions. Rejuvenation Research 18, 422–436.
| Cryoprotectant toxicity: facts, issues, and questions.Crossref | GoogleScholarGoogle Scholar | 25826677PubMed |
Broadhurst L, Coates D (2017) Plant conservation in Australia: current directions and future challenges. Plant Diversity 39, 348–356.
| Plant conservation in Australia: current directions and future challenges.Crossref | GoogleScholarGoogle Scholar | 30159528PubMed |
Bunn E, Turner SR, Panaia M, Dixon KW (2007) The contribution of in vitro technology and cryogenic storage to conservation of indigenous plants. Australian Journal of Botany 55, 345–355.
| The contribution of in vitro technology and cryogenic storage to conservation of indigenous plants.Crossref | GoogleScholarGoogle Scholar |
Bunn E, Turner SR, Dixon KW (2011) Biotechnology for saving rare and threatened flora in a biodiversity hotspot. In Vitro Cellular & Developmental Biology. Plant 47, 188–200.
| Biotechnology for saving rare and threatened flora in a biodiversity hotspot.Crossref | GoogleScholarGoogle Scholar |
Burke MJ (1986) The glassy state and survival of anhydrous biological systems. In ‘Membranes, metabolism and dry organisms’. (Ed. AC Leopold) pp. 358–364. (Cornell University Press: Ithaca, NY, USA)
Burke MJ, Gusta LV, Quamme HA, Weiser CJ, Li PH (1976) Freezing and injury in plants. Annual Review of Plant Physiology 27, 507–528.
| Freezing and injury in plants.Crossref | GoogleScholarGoogle Scholar |
Bustam BM, Dixon KW, Bunn E (2016a) Ex situ germplasm preservation and plant regeneration of a threatened terrestrial orchid, Caladenia huegelii, through micropropagation and cryopreservation. Australian Journal of Botany 64, 659–663.
| Ex situ germplasm preservation and plant regeneration of a threatened terrestrial orchid, Caladenia huegelii, through micropropagation and cryopreservation.Crossref | GoogleScholarGoogle Scholar |
Bustam BM, Dixon KW, Bunn E (2016b) A cryopreservation protocol for ex situ conservation of terrestrial orchids using asymbiotic primary and secondary (adventitious) protocorms. In Vitro Cellular & Developmental Biology. Plant 52, 185–195.
| A cryopreservation protocol for ex situ conservation of terrestrial orchids using asymbiotic primary and secondary (adventitious) protocorms.Crossref | GoogleScholarGoogle Scholar |
Carnegie AJ, Kathuria A, Pegg GS, Entwistle P, Nagel M, Giblin FR (2016) Impact of the invasive rust Puccinia psidii (myrtle rust) on native Myrtaceae in natural ecosystems in Australia. Biological Invasions 18, 127–144.
| Impact of the invasive rust Puccinia psidii (myrtle rust) on native Myrtaceae in natural ecosystems in Australia.Crossref | GoogleScholarGoogle Scholar |
Cassells AC (1991) Problems in tissue culture: culture contamination. In ‘Micropropagation’. (Eds PC Debergh, RH Zimmerman) pp. 31–44. (Springer: Dordrecht)
Catalá A (2006) An overview of lipid peroxidation with emphasis in outer segments of photoreceptors and the chemiluminescence assay. International Journal of Biochemistry & Cell Biology 38, 1482–1495.
| An overview of lipid peroxidation with emphasis in outer segments of photoreceptors and the chemiluminescence assay.Crossref | GoogleScholarGoogle Scholar |
Chang Y, Reed BM (1999) Extended cold acclimation and recovery medium alteration improve regrowth of rubus shoot tips following cryopreservation. Cryo Letters 20, 371–376.
Chen G-Q, Ren L, Zhang J, Reed BM, Zhang D, Shen X-H (2015) Cryopreservation affects ROS-induced oxidative stress and antioxidant response in Arabidopsis seedlings. Cryobiology 70, 38–47.
| Cryopreservation affects ROS-induced oxidative stress and antioxidant response in Arabidopsis seedlings.Crossref | GoogleScholarGoogle Scholar | 25489814PubMed |
Chetverikova EP (2012) DNA damage by reactive oxygen species in cryopreservation and the antioxidant properties of cryoprotectors. Biophysics 57, 263–269.
| DNA damage by reactive oxygen species in cryopreservation and the antioxidant properties of cryoprotectors.Crossref | GoogleScholarGoogle Scholar |
Conservation International (2018) Southwest Australia. Available at: https://www.cepf.net/our-work/biodiversity-hotspots/southwest-australia (accessed 28 May 2018)
Dagla HR (2012) Plant tissue culture: historical developments and applied aspects. Resonance 17, 759–767.
| Plant tissue culture: historical developments and applied aspects.Crossref | GoogleScholarGoogle Scholar |
Day JG, Harding KC, Nadarajan J, Benson EE (2008) Cryopreservation: conservation of bioresources at ultra-low temperatures. In ‘Molecular biomethods handbook’. (Eds J Walker, R Rapley) pp. 917–947. (Humana Press: Hertfordshire, UK)
Dickie JB, Pritchard HW (2002) Systematic and evolutionary aspects of desiccation tolerance in seeds. In ‘Desiccation and survival in plants: drying without dying’. (Eds M Black, HW Pritchard) pp. 239–259. (CAB International: Wallingford, UK)
Duque-Lazo J, van Gils H, Groen TA, Navarro-Cerrillo RM (2016) Transferability of species distribution models: the case of Phytophthora cinnamomi in southwest Spain and southwest Australia. Ecological Modelling 320, 62–70.
| Transferability of species distribution models: the case of Phytophthora cinnamomi in southwest Spain and southwest Australia.Crossref | GoogleScholarGoogle Scholar |
Dussert S, Chabrillange N, Montillet JL, Agnel JP, Engelmann F, Noirot M (2003) Basis of coffee seed sensitivity to liquid nitrogen exposure: oxidative stress or imbibitional damage? Physiologia Plantarum 119, 534–543.
| Basis of coffee seed sensitivity to liquid nitrogen exposure: oxidative stress or imbibitional damage?Crossref | GoogleScholarGoogle Scholar |
Ellis RH, Hong TD, Roberts EH (1990) An intermediate category of seed storage behaviour? I. Coffee. Journal of Experimental Botany 41, 1167–1174.
| An intermediate category of seed storage behaviour? I. Coffee.Crossref | GoogleScholarGoogle Scholar |
Engelmann F (2004) Plant cryopreservation: progress and prospects. In Vitro Cellular & Developmental Biology. Plant 40, 427–433.
| Plant cryopreservation: progress and prospects.Crossref | GoogleScholarGoogle Scholar |
Engelmann F (2012) Germplasm collection, storage, and conservation. In ‘Plant biotechnology and agriculture: prospects for the 21st century’. (Eds PM Hasegawa, A Altman) pp. 255–267. (Academic Press: San Diego, CA, USA)
Esau K (1977) ‘Anatomy of seed plants.’ (John Wiley & Sons: New York)
Escobar RH, Mafia G, Roca WM (1997) A methodology for recovering cassava plants from shoot tips maintained in liquid nitrogen. Plant Cell Reports 16, 474–478.
| A methodology for recovering cassava plants from shoot tips maintained in liquid nitrogen.Crossref | GoogleScholarGoogle Scholar | 30727635PubMed |
Evert RF, Eichhorn SE (2013) ‘Raven biology of plants.’ (W.H. Freeman & Co.: New York)
Fabre J, Dereuddre J (1990) Encapsulation-dehydration: a new approach to cryopreservation of Solanum shoot-tips. Cryo Letters 11, 413–426.
Fang J-Y, Wetten A, Johnston J (2008) Headspace volatile markers for sensitivity of cocoa (Theobroma cacao L.) somatic embryos to cryopreservation. Plant Cell Reports 27, 453–461.
| Headspace volatile markers for sensitivity of cocoa (Theobroma cacao L.) somatic embryos to cryopreservation.Crossref | GoogleScholarGoogle Scholar | 18043891PubMed |
Fouda AH, Hassan SE-D, Eid AM, Ewais EE-D (2015) Biotechnological applications of fungal endophytes associated with medicinal plant Asclepias sinaica (Bioss.). Annals of Agricultural Sciences 60, 95–104.
Fuller BJ (2004) Cryoprotectants: the essential antifreezes to protect life in the frozen state. Cryo Letters 25, 375–388.
Funnekotter B, Kaczmarczyk A, Turner SR, Bunn E, Zhou W, Smith S, Flematti G, Mancera RL (2013) Acclimation-induced changes in cell membrane composition and influence on cryotolerance of in vitro shoots of native plant species. Plant Cell, Tissue and Organ Culture 114, 83–96.
| Acclimation-induced changes in cell membrane composition and influence on cryotolerance of in vitro shoots of native plant species.Crossref | GoogleScholarGoogle Scholar |
Funnekotter B, Whiteley SE, Turner SR, Bunn E, Mancera RL (2015) Evaluation of the new vacuum infiltration vitrification (VIV) cryopreservation technique for native Australian plant shoot tips. Cryo Letters 36, 104–113.
Funnekotter B, Sortey A, Bunn E, Turner SR, Mancera RL (2016) Influence of abiotic stress preconditioning on antioxidant enzymes in shoot tips of Lomandra sonderi (Asparagaceae) prior to cryostorage. Australian Journal of Botany 64, 260–268.
| Influence of abiotic stress preconditioning on antioxidant enzymes in shoot tips of Lomandra sonderi (Asparagaceae) prior to cryostorage.Crossref | GoogleScholarGoogle Scholar |
Funnekotter B, Bunn E, Mancera RL (2017a) Cryo-mesh: a simple alternative cryopreservation protocol. Cryo Letters 38, 155–159.
Funnekotter B, Colville L, Kaczmarczyk A, Turner SR, Bunn E, Mancera RL (2017b) Monitoring of oxidative status in three native Australian species during cold acclimation and cryopreservation. Plant Cell Reports 36, 1903–1916.
| Monitoring of oxidative status in three native Australian species during cold acclimation and cryopreservation.Crossref | GoogleScholarGoogle Scholar | 28900717PubMed |
Funnekotter B, Mancera RL, Bunn E (2017c) Advances in understanding the fundamental aspects required for successful cryopreservation of Australian flora. In Vitro Cellular & Developmental Biology. Plant 53, 289–298.
| Advances in understanding the fundamental aspects required for successful cryopreservation of Australian flora.Crossref | GoogleScholarGoogle Scholar |
Gonzalez-Arnao MT, Engelmann F (2006) Cryopreservation of plant germplasm using the encapsulation-dehydration technique: review and case study on sugarcane. Cryo Letters 27, 155–168.
Gonzalez-Arnao MT, Panta A, Roca WM, Escobar RH, Engelmann F (2007) Development and large scale application of cryopreservation techniques for shoot and somatic embryo cultures of tropical crops. Plant Cell, Tissue and Organ Culture 92, 1–13.
| Development and large scale application of cryopreservation techniques for shoot and somatic embryo cultures of tropical crops.Crossref | GoogleScholarGoogle Scholar |
Gross B, Henk A, Bonnart R, Volk G (2017) Changes in transcript expression patterns as a result of cryoprotectant treatment and liquid nitrogen exposure in Arabidopsis shoot tips. Plant Cell Reports 36, 459–470.
| Changes in transcript expression patterns as a result of cryoprotectant treatment and liquid nitrogen exposure in Arabidopsis shoot tips.Crossref | GoogleScholarGoogle Scholar | 27999976PubMed |
Guerrant EO, Havens K, Vitt P (2014) Sampling for effective ex situ plant conservation. International Journal of Plant Sciences 175, 11–20.
| Sampling for effective ex situ plant conservation.Crossref | GoogleScholarGoogle Scholar |
Gupta S, Reed BM (2006) Cryopreservation of shoot tips of blackberry and raspberry by encapsulation-dehydration and vitrification Cryo Letters 27, 29–42.
Halliwell B, Gutteridge JM (2015) ‘Free radicals in biology and medicine.’ 5th edn (Oxford University Press: New York)
Hamilton K, Ashmore S (2008) Development of conservation technologies for Australia’s rainforest and tropical native fruits. Australasian Plant Conservation 17, 30–31.
Hamilton KN, Ashmore SE, Pritchard HW (2009a) Thermal analysis and cryopreservation of seeds of Australian wild Citrus species (Rutaceae): Citrus australasica, C. inodora and C. garrawayi. Cryo Letters 30, 268–279.
Hamilton KN, Turner SR, Ashmore SE (2009b) Cryopreservation. In ‘Plant germplasm conservation in Australia: strategies and guidelines for developing, managing and utilising ex situ collections’. (Eds CA Offord, PF Meagher) pp. 129–148. (Australian Network for Plant Conservation Inc.: Canberra, ACT)
Hamilton KN, Offord CA, Cuneo P, Deseo MA (2013) A comparative study of seed morphology in relation to desiccation tolerance and other physiological responses in 71 eastern Australian rainforest species. Plant Species Biology 28, 51–62.
| A comparative study of seed morphology in relation to desiccation tolerance and other physiological responses in 71 eastern Australian rainforest species.Crossref | GoogleScholarGoogle Scholar |
Harding K (2004) Genetic integrity of cryopreserved plant cells: a review. Cryo Letters 25, 3–22.
Hay FR, Merritt DJ, Soanes JA, Dixon KW (2010) Comparative longevity of Australian orchid (Orchidaceae) seeds under experimental and low temperature storage conditions. Botanical Journal of the Linnean Society 164, 26–41.
| Comparative longevity of Australian orchid (Orchidaceae) seeds under experimental and low temperature storage conditions.Crossref | GoogleScholarGoogle Scholar |
Helliot B, Swennen R, Poumay Y, Frison E, Lepoivre P, Panis B (2003) Ultrastructural changes associated with cryopreservation of banana (Musa spp.) highly proliferating meristems. Plant Cell Reports 21, 690–698.
Hughes ZE, Mancera RL (2013) Molecular dynamics simulations of mixed DOPC−β-sitosterol bilayers and their interactions with DMSO. Soft Matter 9, 2920–2935.
| Molecular dynamics simulations of mixed DOPC−β-sitosterol bilayers and their interactions with DMSO.Crossref | GoogleScholarGoogle Scholar |
Hughes ZE, Mancera RL (2014) Molecular mechanism of the synergistic effects of vitrification solutions on the stability of phospholipid bilayers. Biophysical Journal 106, 2617–2624.
| Molecular mechanism of the synergistic effects of vitrification solutions on the stability of phospholipid bilayers.Crossref | GoogleScholarGoogle Scholar | 24940779PubMed |
Hughes ZE, Mark AE, Mancera RL (2012) Molecular dynamics simulations of the interactions of DMSO with DPPC and DOPC phospholipid membranes. Journal of Physical Chemistry B 116, 11911–11923.
| Molecular dynamics simulations of the interactions of DMSO with DPPC and DOPC phospholipid membranes.Crossref | GoogleScholarGoogle Scholar |
Hughes ZE, Malajczuk CJ, Mancera RL (2013) The effects of cryosolvents on DOPC−β-sitosterol bilayers determined from molecular dynamics simulations. Journal of Physical Chemistry B 117, 3362–3375.
| The effects of cryosolvents on DOPC−β-sitosterol bilayers determined from molecular dynamics simulations.Crossref | GoogleScholarGoogle Scholar |
Johnston JW, Harding K, Benson EE (2007) Antioxidant status and genotypic tolerance of Ribesin vitro cultures to cryopreservation. Plant Science 172, 524–534.
| Antioxidant status and genotypic tolerance of Ribesin vitro cultures to cryopreservation.Crossref | GoogleScholarGoogle Scholar |
Kaczmarczyk A, Rutten T, Melzer M, Keller ERJ (2008) Ultrastructural changes associated with cryopreservation of potato (Solanum tuberosum L.) shoot tips. Cryo Letters 29, 145–156.
Kaczmarczyk A, Turner SR, Bunn E, Mancera RL, Dixon KW (2011) Cryopreservation of threatened native Australian species – what have we learned and where to from here? In Vitro Cellular & Developmental Biology. Plant 47, 17–25.
| Cryopreservation of threatened native Australian species – what have we learned and where to from here?Crossref | GoogleScholarGoogle Scholar |
Kaczmarczyk A, Funnekotter B, Menon A, Phang PY, Al-Hanbali A, Bunn E, Mancera RL (2012) Current issues in plant cryopreservation. In ‘Current frontiers in cryobiology’. (Ed. I Katkov) pp. 417–438. (IntechOpen Ltd: London)
Kaczmarczyk A, Funnekotter B, Turner SR, Bunn E, Bryant G, Hunt TE, Mancera RL (2013) Development of cryopreservation for Loxocarya cinerea – an endemic Australian plant species important for post-mining restoration. Cryo Letters 34, 508–519.
Kong L, von Aderkas P (2011) A novel method of cryopreservation without a cryoprotectant for immature somatic embryos of conifer. Plant Cell, Tissue and Organ Culture 106, 115–125.
| A novel method of cryopreservation without a cryoprotectant for immature somatic embryos of conifer.Crossref | GoogleScholarGoogle Scholar |
Kreck CA, Mancera RL (2014) Characterization of the glass transition of water predicted by molecular dynamics simulations using nonpolarizable intermolecular potentials. Journal of Physical Chemistry B 118, 1867–1880.
| Characterization of the glass transition of water predicted by molecular dynamics simulations using nonpolarizable intermolecular potentials.Crossref | GoogleScholarGoogle Scholar |
Kreck CA, Mandumpal JB, Mancera RL (2011) Prediction of the glass transition in aqueous solutions of simple amides by molecular dynamics simulations. Chemical Physics Letters 501, 273–277.
| Prediction of the glass transition in aqueous solutions of simple amides by molecular dynamics simulations.Crossref | GoogleScholarGoogle Scholar |
Lai D, Ding J, Smith GW, Smith GD, Takayama S (2015) Slow and steady cell shrinkage reduces osmotic stress in bovine and murine oocyte and zygote vitrification. Human Reproduction 30, 37–45.
| Slow and steady cell shrinkage reduces osmotic stress in bovine and murine oocyte and zygote vitrification.Crossref | GoogleScholarGoogle Scholar | 25355589PubMed |
Larkin PJ, Scowcroft WR (1981) Somaclonal variation – a novel source of variability from cell cultures for plant improvement. Theoretical and Applied Genetics 60, 197–214.
| Somaclonal variation – a novel source of variability from cell cultures for plant improvement.Crossref | GoogleScholarGoogle Scholar | 24276737PubMed |
Li D-Z, Pritchard HW (2009) The science and economics of ex situ plant conservation. Trends in Plant Science 14, 614–621.
| The science and economics of ex situ plant conservation.Crossref | GoogleScholarGoogle Scholar | 19818672PubMed |
Lovelock JE (1953) The haemolysis of human red blood-cells by freezing and thawing. Biochimica et Biophysica Acta 10, 414–426.
| The haemolysis of human red blood-cells by freezing and thawing.Crossref | GoogleScholarGoogle Scholar | 13058999PubMed |
Malajczuk CJ, Hughes ZE, Mancera RL (2013) Molecular dynamics simulations of the interactions of DMSO, mono- and polyhydroxylated cryosolvents with a hydrated phospholipid bilayer. Biochimica et Biophysica Acta - Biomembranes 1828, 2041–2055.
| Molecular dynamics simulations of the interactions of DMSO, mono- and polyhydroxylated cryosolvents with a hydrated phospholipid bilayer.Crossref | GoogleScholarGoogle Scholar |
Mandumpal JB, Kreck CA, Mancera RL (2011) A molecular mechanism of solvent cryoprotection in aqueous DMSO solutions. Physical Chemistry Chemical Physics 13, 3839–3842.
| A molecular mechanism of solvent cryoprotection in aqueous DMSO solutions.Crossref | GoogleScholarGoogle Scholar | 21206958PubMed |
Maunder M, Havens K, Guerrant EO, Jr, Falk DA (2004) Ex situ methods: a vital but underused set of conservation resources. In ‘Ex situ plant conservation: supporting species survival in the wild’. (Eds EO Guerrant Jr., K Havens, M Maunder) pp. 3–20. (Island Press: Covelo, CA, USA)
Menon A, Funnekotter B, Kaczmarczyk A, Bunn E, Turner S, Mancera RL (2012) Cryopreservation of Lomandra sonderi (Asparagaceae) shoot tips using droplet-vitrification. Cryo Letters 33, 259–270.
Menon A, Funnekotter B, Kaczmarczyk A, Bunn E, Turner SR, Mancera RL (2014) Cold-induced changes affect survival after exposure to vitrification solution during cryopreservation in the south-west Australian Mediterranean climate species Lomandra sonderi (Asparagaceae). Plant Cell, Tissue and Organ Culture 119, 347–358.
| Cold-induced changes affect survival after exposure to vitrification solution during cryopreservation in the south-west Australian Mediterranean climate species Lomandra sonderi (Asparagaceae).Crossref | GoogleScholarGoogle Scholar |
Merritt DJ, Touchell DH, Dixon KW, Plummer JA, Turner DW (2000) Moisture content influences survival of cryostored seed of Banksia ashbyi (Proteaceae). Australian Journal of Botany 48, 581–587.
| Moisture content influences survival of cryostored seed of Banksia ashbyi (Proteaceae).Crossref | GoogleScholarGoogle Scholar |
Merritt DJ, Hay FR, Swarts ND, Sommerville KD, Dixon KW (2014a) Ex situ conservation and cryopreservation of orchid germplasm. International Journal of Plant Sciences 175, 46–58.
| Ex situ conservation and cryopreservation of orchid germplasm.Crossref | GoogleScholarGoogle Scholar |
Merritt DJ, Martyn AJ, Ainsley P, Young RE, Seed LU, Thorpe M, Hay FR, Commander LE, Shackelford N, Offord CA, Dixon KW, Probert RJ (2014b) A continental-scale study of seed lifespan in experimental storage examining seed, plant, and environmental traits associated with longevity. Biodiversity and Conservation 23, 1081–1104.
| A continental-scale study of seed lifespan in experimental storage examining seed, plant, and environmental traits associated with longevity.Crossref | GoogleScholarGoogle Scholar |
Meryman H (1968) Modified model for the mechanism of freezing injury in erythrocytes. Nature 218, 333–336.
| Modified model for the mechanism of freezing injury in erythrocytes.Crossref | GoogleScholarGoogle Scholar | 5649669PubMed |
Miguel C, Marum L (2011) An epigenetic view of plant cells cultured in vitro: somaclonal variation and beyond. Journal of Experimental Botany 62, 3713–3725.
| An epigenetic view of plant cells cultured in vitro: somaclonal variation and beyond.Crossref | GoogleScholarGoogle Scholar | 21617249PubMed |
Mikuła A, Jata K, Rybczyński JJ (2009) Cryopreservation strategies for Cyathea australis (R.Br.) Domin. Cryo Letters 30, 429–439.
Millennium Seed Bank Partnership (2015) Seed conservation standards for ‘MSB partnership collections’. Available at: https://www.kew.org/sites/default/files/MSBP%20Seed%20Conservation%20Standards_Final%2005-02-15.pdf (accessed 20 February 2019).
Mittler R (2002) Oxidative stress, antioxidants and stress tolerance. Trends in Plant Science 7, 405–410.
| Oxidative stress, antioxidants and stress tolerance.Crossref | GoogleScholarGoogle Scholar | 12234732PubMed |
Moelbert S, Normand B, De Los Rios P (2004) Kosmotropes and chaotropes: modelling preferential exclusion, binding and aggregate stability. Biophysical Chemistry 112, 45–57.
| Kosmotropes and chaotropes: modelling preferential exclusion, binding and aggregate stability.Crossref | GoogleScholarGoogle Scholar | 15501575PubMed |
Moussa M, Dumont F, Perrier-Cornet J-M, Gervais P (2008) Cell inactivation and membrane damage after long-term treatments at sub-zero temperature in the supercooled and frozen states. Biotechnology and Bioengineering 101, 1245–1255.
| Cell inactivation and membrane damage after long-term treatments at sub-zero temperature in the supercooled and frozen states.Crossref | GoogleScholarGoogle Scholar | 18814283PubMed |
Murphy MP (2009) How mitochondria produce reactive oxygen species. The Biochemical Journal 417, 1–13.
| How mitochondria produce reactive oxygen species.Crossref | GoogleScholarGoogle Scholar | 19061483PubMed |
Nadarajan J, Pritchard HW (2014) Biophysical characteristics of successful oilseed embryo cryoprotection and cryopreservation using vacuum infiltration vitrification: an innovation in plant cell preservation. PLoS One 9, e96169
| Biophysical characteristics of successful oilseed embryo cryoprotection and cryopreservation using vacuum infiltration vitrification: an innovation in plant cell preservation.Crossref | GoogleScholarGoogle Scholar | 24788797PubMed |
Normah M, Makeen A (2008) Cryopreservation of excised embryos and embryonic axes. In ‘Plant Cryopreservation: A practical guide’. pp. 211–240. (Springer)
O’Donnell K, Sharrock S (2017) The contribution of botanic gardens to ex situ conservation through seed banking. Plant Diversity 39, 373–378.
| The contribution of botanic gardens to ex situ conservation through seed banking.Crossref | GoogleScholarGoogle Scholar | 30159531PubMed |
Offord CA, Makinson RO (2009) Options and major considerations for plant germplasm conservation. In ‘Plant germplasm conservation in Australia – strategies and guidelines for developing, managing and utilising ex situ collections’. (Eds CA Offord, PF Meagher) pp. 11–34. (Australian Network for Plant Conservation: Canberra, ACT)
Pammenter NW, Berjak P (2014) Physiology of desiccation-sensitive (recalcitrant) seeds and the implications for cryopreservation. International Journal of Plant Sciences 175, 21–28.
| Physiology of desiccation-sensitive (recalcitrant) seeds and the implications for cryopreservation.Crossref | GoogleScholarGoogle Scholar |
Panis B, Piette B, Swennen R (2005) Droplet vitrification of apical meristems: a cryopreservation protocol applicable to all Musaceae. Plant Science 168, 45–55.
| Droplet vitrification of apical meristems: a cryopreservation protocol applicable to all Musaceae.Crossref | GoogleScholarGoogle Scholar |
Paulet F, Engelmann F, Glaszmann J-C (1993) Cryopreservation of apices of in vitro plantlets of sugarcane (Saccharum sp. hybrids) using encapsulation/dehydration. Plant Cell Reports 12, 525–529.
| Cryopreservation of apices of in vitro plantlets of sugarcane (Saccharum sp. hybrids) using encapsulation/dehydration.Crossref | GoogleScholarGoogle Scholar | 24196115PubMed |
Pence VC (2014) Tissue cryopreservation for plant conservation: potential and challenges. International Journal of Plant Sciences 175, 40–45.
| Tissue cryopreservation for plant conservation: potential and challenges.Crossref | GoogleScholarGoogle Scholar |
Pickering C, Hill W, Green K (2008) Vascular plant diversity and climate change in the alpine zone of the Snowy Mountains, Australia. Biodiversity and Conservation 17, 1627–1644.
| Vascular plant diversity and climate change in the alpine zone of the Snowy Mountains, Australia.Crossref | GoogleScholarGoogle Scholar |
Prance G (1997) The conservation of botanical diversity. In ‘Plant genetic conservation: the in situ approach’. (Eds N Maxted, B Ford-Lloyd, J Hawkes) pp. 3–14. (Springer: Dordrecht, The Netherlands)
Pukacki PM, Juszczyk K (2015) Desiccation sensitivity and cryopreservation of the embryonic axes of the seeds of two Acer species. Trees 29, 385–396.
| Desiccation sensitivity and cryopreservation of the embryonic axes of the seeds of two Acer species.Crossref | GoogleScholarGoogle Scholar |
Rachmilevitch S, DaCosta M, Huang B (2006) Physiological and biochemical indicators for stress tolerance. In ‘Plant–environment interactions’. 3rd edn (Ed. B Huang) pp. 321–356. (CRC Press: Boca Raton, FL, USA)
Reed BM (2008) Cryopreservation - practical considerations. In ‘Plant cryopreservation: a practical guide’. (Ed. BM Reed) pp. 3–13. (Springer: New York)
Ren L, Zhang D, Jiang X-N, Gai Y, Wang W-M, Reed BM, Shen X-H (2013) Peroxidation due to cryoprotectant treatment is a vital factor for cell survival in Arabidopsis cryopreservation. Plant Science 212, 37–47.
| Peroxidation due to cryoprotectant treatment is a vital factor for cell survival in Arabidopsis cryopreservation.Crossref | GoogleScholarGoogle Scholar | 24094052PubMed |
Ren L, Zhang D, Chen G-q, Reed B, Shen X-h, Chen H-y (2015) Transcriptomic profiling revealed the regulatory mechanism of Arabidopsis seedlings’ response to oxidative stress from cryopreservation. Plant Cell Reports 34, 2161–2178.
| Transcriptomic profiling revealed the regulatory mechanism of Arabidopsis seedlings’ response to oxidative stress from cryopreservation.Crossref | GoogleScholarGoogle Scholar | 26373654PubMed |
Roach T, Ivanova M, Beckett RP, Minibayeva FV, Green I, Pritchard HW, Kranner I (2008) An oxidative burst of superoxide in embryonic axes of recalcitrant sweet chestnut seeds as induced by excision and desiccation. Physiologia Plantarum 133, 131–139.
| An oxidative burst of superoxide in embryonic axes of recalcitrant sweet chestnut seeds as induced by excision and desiccation.Crossref | GoogleScholarGoogle Scholar | 18452494PubMed |
Sakai A (1965) Survival of plant tissue at super-low temperature III. Relation between effective prefreezing temperatures and the degree of front hardiness. Plant Physiology 40, 882–887.
| Survival of plant tissue at super-low temperature III. Relation between effective prefreezing temperatures and the degree of front hardiness.Crossref | GoogleScholarGoogle Scholar | 16656170PubMed |
Sakai A, Engelmann F (2007) Vitrification, encapsulation-vitrification and droplet-vitrification: a review. Cryo Letters 28, 151–172.
Sakai A, Kobayashi S, Oiyama I (1990) Cryopreservation of nucellar cells of navel orange (Citrus sinensis Osb. var. brasiliensis Tanaka) by vitrification. Plant Cell Reports 9, 30–33.
| Cryopreservation of nucellar cells of navel orange (Citrus sinensis Osb. var. brasiliensis Tanaka) by vitrification.Crossref | GoogleScholarGoogle Scholar | 24226373PubMed |
Shamblin SL, Tang X, Chang L, Hancock BC, Pikal MJ (1999) Characterization of the time scales of molecular motion in pharmaceutically important glasses. Journal of Physical Chemistry B 103, 4113–4121.
| Characterization of the time scales of molecular motion in pharmaceutically important glasses.Crossref | GoogleScholarGoogle Scholar |
Shatnawi MA, Johnson KA (2004) Cryopreservation by encapsulation-dehydration of ‘Christmas bush’ (Ceratopetalum gummiferum) shoot tips. In Vitro Cellular & Developmental Biology. Plant 40, 239–244.
| Cryopreservation by encapsulation-dehydration of ‘Christmas bush’ (Ceratopetalum gummiferum) shoot tips.Crossref | GoogleScholarGoogle Scholar |
Shatnawi MA, Johnson KA, Torpy FR (2004) In vitro propagation and cryostorage of Syzygium francissi (Myrtaceae) by the encapsulation-dehydration method. In Vitro Cellular & Developmental Biology. Plant 40, 403–407.
| In vitro propagation and cryostorage of Syzygium francissi (Myrtaceae) by the encapsulation-dehydration method.Crossref | GoogleScholarGoogle Scholar |
Sies H, Cadenas E (1985) Oxidative stress: damage to intact cells and organs. Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences 311, 617–631.
| Oxidative stress: damage to intact cells and organs.Crossref | GoogleScholarGoogle Scholar | 2869521PubMed |
Skyba M, Petijová L, Košuth J, Koleva DP, Ganeva TG, Kapchina-Toteva VM, Čellárová E (2012) Oxidative stress and antioxidant response in Hypericum perforatum L. plants subjected to low temperature treatment. Journal of Plant Physiology 169, 955–964.
| Oxidative stress and antioxidant response in Hypericum perforatum L. plants subjected to low temperature treatment.Crossref | GoogleScholarGoogle Scholar | 22575055PubMed |
Sommerville KD, Clarke B, Keppel G, McGill C, Newby Z-J, Wyse SV, James SA, Offord CA (2017) Saving rainforests in the South Pacific: challenges in ex situ conservation. Australian Journal of Botany 65, 609–624.
| Saving rainforests in the South Pacific: challenges in ex situ conservation.Crossref | GoogleScholarGoogle Scholar |
Steponkus PL (1984) Role of the plasma membrane in freezing injury and cold acclimation. Annual Review of Plant Physiology 35, 543–584.
| Role of the plasma membrane in freezing injury and cold acclimation.Crossref | GoogleScholarGoogle Scholar |
Suzuki N, Koussevitzky S, Mittler R, Miller G (2012) ROS and redox signalling in the response of plants to abiotic stress. Plant, Cell & Environment 35, 259–270.
| ROS and redox signalling in the response of plants to abiotic stress.Crossref | GoogleScholarGoogle Scholar |
Taylor NL, Day DA, Millar AH (2004) Targets of stress-induced oxidative damage in plant mitochondria and their impact on cell carbon/nitrogen metabolism. Journal of Experimental Botany 55, 1–10.
| Targets of stress-induced oxidative damage in plant mitochondria and their impact on cell carbon/nitrogen metabolism.Crossref | GoogleScholarGoogle Scholar | 14645387PubMed |
Thomashow MF (1999) Plant cold acclimation: freezing tolerance genes and regulatory mechanisms. Annual Review of Plant Physiology and Plant Molecular Biology 50, 571–599.
| Plant cold acclimation: freezing tolerance genes and regulatory mechanisms.Crossref | GoogleScholarGoogle Scholar | 15012220PubMed |
Touchell DH (1995) Principles of cryobiology for conservation of threatened species. PhD Thesis, Department of Botany, University of Western Australia, Perth, WA, Australia.
Touchell DH, Dixon KW, Tan B (1992) Cryopreservation of shoot-tips of Grevillea scapigera (Proteaceae): a rare and endangered plant from Western Australia. Australian Journal of Botany 40, 305–310.
| Cryopreservation of shoot-tips of Grevillea scapigera (Proteaceae): a rare and endangered plant from Western Australia.Crossref | GoogleScholarGoogle Scholar |
Touchell DH, Turner SR, Bunn E, Dixon KW (2002) Cryostorage of somatic tissues of endangered Australian species. In ‘Cryopreservation of plant germplasm’. (Eds LE Towill, YPS Bajaj) pp. 357–372. (Springer-Verlag: Berlin)
Towill LE, Ellis DD (2008) Cryopreservation of dormant buds. In ‘Plant cryopreservation: A practical guide’. (Ed. BM Reed) pp. 421–442. (Springer: New York)
Tsikas D (2017) Assessment of lipid peroxidation by measuring malondialdehyde (MDA) and relatives in biological samples: analytical and biological challenges. Analytical Biochemistry 524, 13–30.
| Assessment of lipid peroxidation by measuring malondialdehyde (MDA) and relatives in biological samples: analytical and biological challenges.Crossref | GoogleScholarGoogle Scholar | 27789233PubMed |
Turner SR, Touchell DH, Dixon KW, Tan B (2000) Cryopreservation of Anigozanthos viridis ssp. viridis and related taxa from the south-west of Western Australia. Australian Journal of Botany 48, 739–744.
| Cryopreservation of Anigozanthos viridis ssp. viridis and related taxa from the south-west of Western Australia.Crossref | GoogleScholarGoogle Scholar |
Turner SR, Senaratna T, Bunn E, Tan B, Dixon KW, Touchell DH (2001) Cryopreservation of shoot tips from six endangered Australian species using a modified vitrification protocol. Annals of Botany 87, 371–378.
| Cryopreservation of shoot tips from six endangered Australian species using a modified vitrification protocol.Crossref | GoogleScholarGoogle Scholar |
Tweddle JC, Dickie JB, Baskin CC, Baskin JM (2003) Ecological aspects of seed desiccation sensitivity. Journal of Ecology 91, 294–304.
| Ecological aspects of seed desiccation sensitivity.Crossref | GoogleScholarGoogle Scholar |
Uchendu EE, Leonard SW, Traber MG, Reed BM (2010) Vitamins C and E improve regrowth and reduce lipid peroxidation of blackberry shoot tips following cryopreservation. Plant Cell Reports 29, 25–35.
| Vitamins C and E improve regrowth and reduce lipid peroxidation of blackberry shoot tips following cryopreservation.Crossref | GoogleScholarGoogle Scholar | 19898848PubMed |
Uchendu EE, Shukla MR, Reed BM, Saxena PK (2013) Melatonin enhances the recovery of cryopreserved shoot tips of American elm (Ulmus americana L.). Journal of Pineal Research 55, 435–442.
Varghese B, Sershen , Berjak P, Varghese D, Farrant J, Pammenter N (2011) A biochemical assessment of survival during slow and rapid drying of recalcitrant Trichilia dregeana embryonic axes. South African Journal of Botany 77, 563–564.
Verhoeven A, García-Plazaola JI, Fernández-Marín B (2018) Shared mechanisms of photoprotection in photosynthetic organisms tolerant to desiccation or to low temperature. Environmental and Experimental Botany 154, 66–79.
Volk GM, Caspersen AM (2007) Plasmolysis and recovery of different cell types in cryoprotected shoot tips of Mentha × piperita. Protoplasma 231, 215–226.
| Plasmolysis and recovery of different cell types in cryoprotected shoot tips of Mentha × piperita.Crossref | GoogleScholarGoogle Scholar | 17603746PubMed |
Volk GM, Walters C (2006) Plant vitrification solution 2 lowers water content and alters freezing behavior in shoot tips during cryoprotection. Cryobiology: International Journal of Low Temperature Biology and Medicine 52, 48–61.
| Plant vitrification solution 2 lowers water content and alters freezing behavior in shoot tips during cryoprotection.Crossref | GoogleScholarGoogle Scholar |
Walters C (2015) Orthodoxy, recalcitrance and in-between: describing variation in seed storage characteristics using threshold responses to water loss. Planta 242, 397–406.
| Orthodoxy, recalcitrance and in-between: describing variation in seed storage characteristics using threshold responses to water loss.Crossref | GoogleScholarGoogle Scholar | 25985842PubMed |
Walters C, Ballesteros D, Vertucci VA (2010) Structural mechanics of seed deterioration: standing the test of time. Plant Science 179, 565–573.
| Structural mechanics of seed deterioration: standing the test of time.Crossref | GoogleScholarGoogle Scholar |
Wang Z-C, Deng X-X (2004) Cryopreservation of shoot-tips of citrus using vitrification: effect of reduced form of glutathione. Cryo Letters 25, 43–50.
Wen B, Wang R, Cheng H, Song S (2010) Cytological and physiological changes in orthodox maize embryos during cryopreservation. Protoplasma 239, 57–67.
| Cytological and physiological changes in orthodox maize embryos during cryopreservation.Crossref | GoogleScholarGoogle Scholar | 19904484PubMed |
Wesley-Smith J, Pammenter NW, Berjak P, Walters C (2001) The effects of two drying rates on the desiccation tolerance of embryonic axes of recalcitrant jackfruit (Artocarpus heterophyllus Lamk.) seeds. Annals of Botany 88, 653–664.
| The effects of two drying rates on the desiccation tolerance of embryonic axes of recalcitrant jackfruit (Artocarpus heterophyllus Lamk.) seeds.Crossref | GoogleScholarGoogle Scholar |
Wesley-Smith J, Walters C, Pammenter NW, Berjak P (2015) Why is intracellular ice lethal? A microscopical study showing evidence of programmed cell death in cryo-exposed embryonic axes of recalcitrant seeds of Acer saccharinum. Annals of Botany 115, 991–1000.
| Why is intracellular ice lethal? A microscopical study showing evidence of programmed cell death in cryo-exposed embryonic axes of recalcitrant seeds of Acer saccharinum.Crossref | GoogleScholarGoogle Scholar | 25808653PubMed |
Whitaker C, Beckett RP, Minibayeva FV, Kranner I (2010) Production of reactive oxygen species in excised, desiccated and cryopreserved explants of Trichilia dregeana Sond. South African Journal of Botany 76, 112–118.
| Production of reactive oxygen species in excised, desiccated and cryopreserved explants of Trichilia dregeana Sond.Crossref | GoogleScholarGoogle Scholar |
Whiteley SE, Bunn E, Menon A, Mancera RL, Turner SR (2016) Ex situ conservation of the endangered species Androcalva perlaria (Malvaceae) by micropropagation and cryopreservation. Plant Cell, Tissue and Organ Culture 125, 341–352.
| Ex situ conservation of the endangered species Androcalva perlaria (Malvaceae) by micropropagation and cryopreservation.Crossref | GoogleScholarGoogle Scholar |
Williams KJ, Ford A, Rosauer DF, De Silva N, Mittermeier R, Bruce C, Larsen FW, Margules C (2011) Forests of east Australia: the 35th biodiversity hotspot. In ‘Biodiversity hotspots’. (Eds FE Zachos, JC Habel) (Springer-Verlag: Berlin)
Yamamoto S, Rafique T, Priyantha WS, Fukui K, Matsumoto T, Niino T (2011) Development of a cryopreservation procedure using aluminium cryo-plates. Cryo Letters 32, 256–265.
Yamamoto S, Rafique T, Fukui K, Sekizawa K, Niino T (2012) V-cryo-plate procedure as an effective protocol for cryobanks: case study of mint cryopreservation. Cryo Letters 33, 12–23.
Zhu G-Y, Geuns JMC, Dussert S, Swennen R, Panis B (2006) Change in sugar, sterol and fatty acid composition in banana meristems caused by sucrose-induced acclimation and its effects on cryopreservation. Physiologia Plantarum 128, 80–94.
| Change in sugar, sterol and fatty acid composition in banana meristems caused by sucrose-induced acclimation and its effects on cryopreservation.Crossref | GoogleScholarGoogle Scholar |