

Carbon and nitrogen metabolism in rice cultivars affected by salt-alkaline stress
Liying Guo A , Yuya Lu A , Shuying Bao A , Qiang Zhang A , Yanqiu Geng A B and Xiwen Shao A B
A , Yuya Lu A , Shuying Bao A , Qiang Zhang A , Yanqiu Geng A B and Xiwen Shao A B
A Jilin Agricultural University, Changchun 130118, China.
B Corresponding authors. Email: ccgyq@163.com; shaoxiwen@126.com
Crop and Pasture Science 72(5) 372-382 https://doi.org/10.1071/CP20445
Submitted: 5 November 2020 Accepted: 10 March 2021 Published: 26 May 2021
Journal Compilation © CSIRO 2021 Open Access CC BY-NC-ND
Abstract
Salt-alkaline stress generally leads to soil compaction and fertility decline. It also restricts rice growth and nutrient acquisition, so reduces rice yield. However, little is known about the effect of salt-alkaline stress on the carbon and nitrogen metabolism of rice. In this study, two relatively salt-alkaline tolerant (Changbai 9 and Dongdao 12) and sensitive (Jinongda 138 and Tongyu 315) rice cultivars were grown in a field experiment conducted with two soil types including black soil and salt-alkaline soil to evaluate the characteristics of yield, carbon and nitrogen metabolism in rice. Results showed that yield and nitrogen use efficiency (NUE) in rice greatly decreased under salt-alkaline stress. Changbai 9 (CB9) and Dongdao 12 (DD12) showed higher NUE than Jinongda 138 (JND138) and Tongyu 315 (TY315) under the salt-alkaline stress. Additionally, carbon and nitrogen metabolism enzyme activity and compound content were significantly affected by salt-alkali stress; different performances were observed between cultivars under salt-alkaline stress. In conclusion, our results indicate that salt-alkaline tolerant rice cultivars may have more stable carbon and nitrogen metabolism than sensitive genotypes under salt-alkaline conditions.
Keywords: agriculture, carbon metabolism, nitrogen metabolism, rice, salt-alkaline soil, yield, sustainable development, genotypes, sensitive, tolerant, global food security.
Introduction
Globally, more than 900 million ha of land is harmed by salt-alkaline conditions, accounting for 20% of the world’s total cultivated land (Flowers and Yeo 1995; Li et al. 2020). Due to global climate change, population growth, increased industrial pollution, the development of irrigated agriculture, and improper use of chemical fertilisers and other factors, soil salinisation is becoming an increasingly serious problem (Tian et al. 2016). It is estimated that 30% of cultivated land will be salinised in the next 25 years, and it will reach 50% by 2050 (Cheng et al. 2007). In China, more than 6.67 million ha of land is damaged by salt-alkaline conditions, and nearly one-fifth of the cultivated land has been salinised (Eraslan et al. 2007; Qadir et al. 2014). Rice is one of the most important food crops worldwide, and sustainable development of rice agriculture is an essential part to ensure global food security (Qadir et al. 2014). Rice cultivation in salt-alkaline land is an effective soil improvement and utilisation measure, as well as an important way to increase food production, as shown by many years of agricultural production practices. Therefore, research on the morphological and physiological characteristics of salt-alkaline tolerant rice is necessary for sustainable development.
Exploring the physiological and biochemical metabolic activities and tolerance mechanisms of salt-alkaline tolerant rice under salt-alkaline stress is the basis for the breeding of salt-alkaline tolerant rice. In 1939, the strong saline-alkali-tolerant rice variety ‘Pokkali’ was bred and planted over large areas in Sri Lanka. Studies have found that under salt (NaCl) stress conditions, the salt-alkaline tolerant variety ‘Pokkali’ can reduce the ratio of ascorbate/dehydroascorbic acid, increase the activity of antioxidant enzymes and increase its salt-alkaline tolerance, by maintaining high activity of key enzymes (glyoxalase I and II) in the methyldiacetaldehyde detoxification system (El-Shabrawi et al. 2010; Kabir et al. 2016). The salt-alkaline tolerant rice variety ‘FL478’ was exposed to salt stress by inhibiting the metabolism of organic acid (OA) to inhibit its growth to adapt to salt stress the early stage of salt stress, in the later stage of growth, it accumulates a large amount of soluble sugars, amino acids, and other small molecules that act as osmotic adjustment substances to relieve osmotic stress and improve its own stress resistance (Shobbar et al. 2012; Zhao et al. 2014). Furthermore, plants can enhance the activities of antioxidase (superoxide dismutase, ferredoxin, ascorbate peroxidase) to ease the damage of reactive oxygen species (Zhang and Mu 2009). Past studies have focussed on the structural stability and metabolic balance of plants. However, salt-alkaline stress also inhibits plant growth through nutrient absorption and utilisation, the mechanism of the effect of salt-alkali stress on nutrient absorption and utilization needs to be further studied. Carbon and nitrogen metabolism are the two basic metabolism pathways in plants, and their stability under salt-alkaline stress is essential for the stable and high yield of salt-alkaline tolerant rice cultivation.
Salt-alkaline stress seriously affects crop yield (Abdullah et al. 2001; Nemati et al. 2011); doing so through two important mechanisms. On the one hand, high salt ions cause osmotic stress and ion poisoning, especially the excessive accumulation of Na+ in the leaves that destroy the cell membrane structure, affect the production of photosynthetic substances, and cause a decline in yield. On the other hand, high pH values cause ion precipitation and nutrient stress, resulting in low nutrient absorption and utilisation of crops in salt-alkaline soils, resulting in reduced yield. Studies have also shown that under salt-alkaline conditions, the absorption and utilisation of nutrients in rice is significantly inhibited (Dluzniewska et al. 2007; Tian et al. 2016). Nutrient absorption and utilisation-related gene expression and enzyme activity are also regulated by salt-alkaline stress (Surabhi et al. 2008; Abouelsaad et al. 2016). Therefore, research on rice photosynthetic characteristics, material production capacity, and nutrient absorption and utilisation under salt-alkaline stress is important. In this study, two relatively salt-alkaline tolerant rice cultivars (Changbai (CB) 9 and Dongdao (DD) 12) and two relatively salt-alkaline sensitive rice cultivars (Jinongda (JND) 138 and Tongyu (TY) 315) were planted into black and salt-alkaline soils. We investigated the effects of salt-alkaline stress on yield, and carbon and nitrogen metabolism the four rice cultivars. Different changes among the salt-alkaline tolerant and sensitive cultivars are also discussed.
Materials and methods
Plant materials and site description
Field experiments were conducted at a farm belonging to Jilin Agricultural University (43°05′N, 125°38′E) during the rice growing season of 2018, and repeated in 2019. Four japonica rice cultivars with similar growth periods, among them, Changbai 9 (CB9) and Dongdao 12 (DD12) are relatively salt-alkaline tolerant cultivars, whereas Jinongda 138 (JND138) and Tongyu 315 (TY315) are relatively salt-alkaline sensitive cultivars. Prior to the experiments, two types of wild soils (Table 1), black soil (BS) and salt-alkaline soil (SAS), soil samples from the upper 20-cm layer were collected to analyse the soil chemical properties.

|
Experimental design
The experiment was arranged in a completely randomised block design with three replications. The seeds were sown on 12 April 2018 and 9 April 2019. At the four-leaf heart stage, seedlings with the same growth were selected and transplanted on 25 May 2018 and 2019, at a hill spacing of 13.3 × 30.0 cm with two seedlings per hill and a plot size of 16 m2 in 2018 and 2019, plots were separated at their perimeters by a cement ridge to prohibit the exchange of irrigation water and fertiliser between plots. Nitrogen (180 kg N ha−1) fertiliser were applied as basal fertiliser, tillering fertiliser, and panicle fertiliser. The proportion of N split was 48, 32 and 20, respectively, at these three stages. Phosphorous (90 kg P2O5 ha−1) was used in the basal fertiliser. Potassium (45 kg K2O ha−1) was used in the basal and panicle fertilizers.
Sampling and measurements
Leaf photosynthesis and physiological parameters
At filling stage, the leaf area of the three plants growing in the three hills selected for each treatment was measured with a leaf area meter (CI-203, CID, USA). The photosynthetic rate (Pn) of latest full-grown leaves on the main plant stems were measured from 09:00 to 11:00 hours using a CIRAS-3 (PP Systems, USA) photosynthetic instrument. Leaf temperature during measurements was maintained at 27°C, leaf chamber humidity during measurements was maintained at 48–50%, with a photosynthetic photon flux density of 1200 μmol m–2 s–1. Photosynthetic nitrogen use efficiency (PNUE, μmol g–1 s–1) was calculated as follows (Ju et al. 2015):
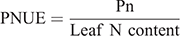
At filling stage, the latest full-grown leaves on the main plant stems collected from six representative hills in each plot were sampled to examine the differences in the compound content and the activities of key enzymes. The soluble sugar content was determined according to Maness (2010). The sucrose content was determined according to Zhang and Qu (2003). The leaf nitrogen content was determined by an elemental analyser (Elementar Vario Macro cube, Hanau, Germany). The soluble protein content was determined according to Zhang and Qu (2003). The free amino acids and activity of Rubisco was determined according to Wang et al. (2000). The activity of sucrose phosphate synthase (SPS) was determined according to Nakamura et al. (1989). The activity of nitrate reductase (NR) was determined according to Gibon et al. (2004). The activity of glutamine synthetase (GS) was determined according to Sun et al. (2014).
Grain yield and NUE
At maturity, grain yield was determined for except border plants within 1 m2 area (three replicates) in each plot and adjusted to a moisture content of 0.14 g H2O g−1 fresh weight. The plants in the nine representative hills of each plot were sampled to determine the yield components. Plant samples from each hill were separated into straw and panicle. The panicle number of each hill was recorded to determine the panicle number per hectare. Filled and unfilled grains of the panicles were manually separated to measure the grain number per panicle and seed setting rate. Dry weights of rachis, filled and unfilled grains, and straw were determined after oven drying at 80°C to a constant weight. Randomly selected filled grains from each hill were used for 1000-grain weight measurements. Tissue N content was determined by an elemental analyser (Elementar Vario Macro cube) to calculate aboveground N uptake. Total N accumulation (TNA, g m–2) was considered the total amount of N accumulated in a plant at maturity. The N utilisation efficiency for grain production (NUEg, g g–1), The N utilisation efficiency for biomass production (NUEb, g g–1), and N harvest index (NHI, g g–1) were calculated as follows:
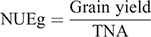

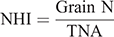
Statistical analysis
Data were statistically analysed using Microsoft Excel 2010 and SPSS 19.0 software (Softonic International, Barcelona, Spain). Means were compared using least significant difference at P < 0.05 (l.s.d. 0.05). Graphs were drawn with GraphPad Prism 8 software (GraphPad, San Diego, CA, USA).
Results
Grain yield and yield components
As shown in Table 2, the grain yield, effective panicles, grain per panicle and seed setting rate were significantly lower in the salt-alkaline soil than in the black soil. Compared with the black soil treatment, the effective panicles of CB9 (21.59% 2018 same below, 19.20% 2019 same below) and DD12 (23.67%, 20.68%) less relative decrease1 than JND138 (26.75%, 27.14%) and TY315 (28.24%, 28.11%) under the salt-alkaline soil treatment. The grain per panicle of CB9 (11.74%, 9.66%), DD12 (13.50%, 13.41%) and JND138 (13.50%, 13.41%) less relative decrease than TY315 (23.06%, 23.03%) under the salt-alkaline soil treatment. The seed setting rate of CB9 (6.60%, 6.10%) and DD12 (7.06%, 7.02%) less relative decrease than JND138 (13.97%, 14.43%) and TY315 (15.35%, 15.81%) under the salt-alkaline soil treatment. The yield of CB9 (31.57%, 31.19%) and DD12 (35.03%, 34.81%) less relative decrease than JND138 (49.18%, 49.07%) and TY315 (50.54%, 51.13%) under the salt-alkaline soil treatment.

|
N use efficiency and the N harvest index
As shown in Table 3, the TNA, NUEb, NUEg and NHI were significantly lower in the salt-alkaline soil than in the black soil. Compared with the black soil treatment, the TNA of CB9 (27.17%, 24.23%) and DD12 (24.62%, 23.83%) less relative decrease than JND138 (34.29%, 35.56%) and TY315 (34.77%, 36.60%) under the salt-alkaline soil treatment. The NUEb of CB9 (1.88%, 1.79%) and DD12 (10.75%, 9.46%) less relative decrease than JND138 (18.80%, 18.30%) and TY315 (19.49%, 21.13%) under the salt-alkaline soil treatment. The NUEg of CB9 (6.00%, 6.87%) and DD12 (12.18%, 13.87%) less relative decrease than JND138 (22.62%, 22.03%) and TY315 (24.07%, 25.43%) under the salt-alkaline soil treatment. The HNI of CB9 (9.68%, 6.67%) and DD12 (21.21%, 20.00%) less relative decrease than JND138 (29.23%, 26.98%) and TY315 (22.22%, 26.56%) under the salt-alkaline soil treatment.

|
Photosynthetic characteristics and leaf area index
As shown in Fig. 1, the leaf area index (LAI), Pn and PNUE were significantly lower in plants grown in salt-alkaline soil than in the black soil. Compared with the black soil treatment, LAI of CB9 (17.65%, 18.25%) and DD12 (21.72%, 26.11%) less relative decrease than JND138 (36.11%, 32.68%) and TY315 (29.79%, 31.07%) under the salt-alkaline soil treatment. The Pn of CB9 (23.88%, 23.88%) and DD12 (27.63%, 31.56%) less relative decrease than JND138 (42.34%, 39.98%) and TY315 (45.42%, 37.89%) under the salt-alkaline soil treatment. The PNUE of CB9 (3.16%, 3.25%) and DD12 (13.93%, 13.97%) less relative decrease than JND138 (25.28%, 24.72%) and TY315 (20.30%, 20.57%) under the salt-alkaline soil treatment.

|
Content of carbon and nitrogen compounds
As shown in Fig. 2, the soluble protein content was significantly lower, whereas the free amino acid, soluble sugar, and sucrose contents were significantly higher in plants grown in the salt-alkaline soil than in the black soil. Compared with the black soil treatment, the soluble protein content of CB9 (10.76%, 10.60%) and DD12 (8.55%, 8.74%) less relative decrease than JND138 (24.27%, 23.78%) and TY315 (26.96%, 27.02%) under the salt-alkaline soil treatment. The free amino acid content of CB9 (31.57%, 31.12%) and DD12 (28.74%, 28.72%) more relative increase than JND138 (14.07%, 14.21%) and TY315 (16.99%, 17.07%) under the salt-alkaline soil treatment. The soluble sugar content of CB9 (175.47%, 178.87%) and DD12 (136.51%, 138.90%) more relative increase than JND138 (28.47%, 28.22%) and TY315 (71.49%, 68.57%) under the salt-alkaline soil treatment. The sucrose content of CB9 (173.36%, 178.87%) and DD12 (135.61%, 132.48%) more relative increase than JND138 (27.48%, 29.70%) and TY315 (79.31%, 71.82%) under the salt-alkaline soil treatment.

|
Enzymatic activities related to carbon and nitrogen metabolism
As shown in Fig. 3, the Rubisco activity and NR activity were significantly lower, whereas the SPS activity was significantly higher in plants grown in the salt-alkaline soil than in the black soil. A significant difference among genotypes was observed in the GS activity in the salt-alkaline soil treatment. Compared with the black soil treatment, the Rubisco activity of CB9 (5.65%, 5.41%) and DD12 (9.25%, 8.96%) less relative decrease than JND138 (21.93%, 21.96%) and TY315 (16.67%, 16.82%) under the salt-alkaline soil treatment. The SPS activity of CB9 (112.67%, 121.52%) and DD12 (108.98%, 104.48%) more relative increase than JND138 (96.98%, 85.01%) and TY315 (101.33%, 83.66%) under the salt-alkaline soil treatment. The NR activity of JND138 (14.93%, 14.97%) less relative decrease than CB9 (17.63%, 17.87%), DD12 (16.75%, 16.70%) and TY315 (16.47%, 16.44%) under the salt-alkaline soil treatment. The GS activity of CB9 (12.53%, 12.75%), DD12 (7.67%, 8.98%) and TY315 (1.33%, 1.33%) were increased, and JND138 (4.97%, 3.02%) was decreased under the salt-alkaline soil treatment.

|
Correlation analysis
The relative effect of the salt-alkaline treatment on Pn, LAI, soluble protein, free amino acid, soluble sugar, sucrose contents, and RuBisco, SPS, and GS activities were significantly and positively correlated with the relative effect on TNA, NUEb, NUEg, PNUE and yield for rice under the salt-alkaline soil conditions. The relative effect of the salt-alkaline treatment on NR activity was significantly and negatively correlated with the relative effect on TNA, NUEb, NUEg, PNUE and yield for rice under the salt-alkaline soil conditions (Table 4).

|
Discussion
Rice is the preferred food crops for the development and utilisation of beaches and salt-alkaline land, but it is particularly sensitive to soil salinity (Krishnamurthy et al. 2009). The salt-alkaline tolerance of rice is a complex physiological and biochemical process involving the coordination of many genes, proteins, and various salt-alkaline tolerance mechanisms. Different salt tolerance varieties of rice also have differences in saline-alkali tolerance mechanisms (Tian et al. 2016). It is vital to explore the physiological and biochemical mechanism of salt-alkaline tolerant rice and to breed salt-alkaline tolerant rice varieties by analysing the effects of salt-alkaline stress on the yield, nitrogen use efficiency, carbon and nitrogen metabolism of different genotype rice varieties.
Yield and NUE
The growth and development of plants in high-salinity environments is restricted, and the leaves are often yellow and curled, which affects cell division and elongation, slowing growth of crops and reducing yield (Han et al. 2016). In this study, under the salt-alkaline stress, the yield and components of the four rice cultivars were all affected, and the effective panicles, grain per panicle, seed setting rate, and yield decreased. Among these measurements, effective panicles were most seriously affected. The decrease in the effective panicles is mainly due to the inhibition of tillering and reduction of productive tiller percentage by salt-alkaline stress (Zeng and Shannon 2000a). This indicates that the decrease in the effective panicles is the main reason for the reduced rice yield under the salt-alkaline stress. Compared with salt-alkaline sensitive cultivars, the yield and component factors of salt-alkaline tolerant cultivars was less affected under salt-alkaline stress. The LAI is closely related to the material production capacity of crops, and is an important parameter of whether the crop population structure is reasonable (Croft et al. 2013). Therefore, a reasonable LAI is one of the important conditions to ensure high and stable rice yields. In this study, the LAI and Pn of rice are significantly reduced under salt-alkaline stress, similar to previous studies (Nieman and Clark 1976; Yang et al. 2007). It may due to the higher pH under salt-alkaline stress, which damages the plasma membrane system and photosynthetic structure of the cell, leading to aerobic respiration and a decline in photosynthetic function. Additionally, compared with salt-alkaline sensitive cultivars, the LAI and Pn of salt-alkaline tolerant cultivars was less affected under salt-alkali stress. This shows that the salt-alkaline-tolerant cultivars CB9 and DD12 have a greater capacity for growth and material production under the salt-alkaline conditions than the less tolerant cultivars tested.
Salt-alkaline soil affects the normal metabolism and various physiological activities of crops (Woodrow et al. 2017). Accumulation of a large amount of Na+ will destroy the structure and function of the enzyme, leading to hindered free radical scavenging, accelerating the damage of plasma membrane and membrane protein, and increasing the permeability of protoplasm (He and Zhu 2008). At the same time, excessive absorption of Na+ leads to limited absorption of K+ and other metal elements necessary to maintain the normal physiological activities of crops. In the protoplast, Na+ is replaced by K+, K+ is extravagated, and the value of Na+/K+ increases, and the ions and nutrients of the cell fluid are out of balance, resulting in insufficient or lack of nutrients (Stevens et al. 2006). In current study, the TNA, NUEb, NUEg, and NHI reductions in CB9 and DD12 were significantly lower than JND138 and TY315 under the salt-alkaline conditions. Among them, the TNA has the largest decrease, indicating that nutrient absorption is the main factor limiting the increase in rice yield under the salt-alkaline conditions. Under the black soil conditions, the TNA, NUEb, NUEg, and NHI of CB9 were lower than DD12, JND138 and TY315, whereas under the salt-alkaline soil conditions, these indexes of CB9 decrease less than DD12, JND138, and TY315. These results show that under the salt-alkaline stress, nitrogen absorption and utilisation efficiency can be used as an index to evaluate the salt-alkaline tolerance of rice. Additionally, in order to meet human demand for food, rice cultivars with high NUE under normal cultivation conditions and stable NUE under salt-alkali stress should be selected.
Carbon and nitrogen metabolism
Carbon and nitrogen metabolism are the two most basic metabolic pathways of rice, so their stability under salt-alkali stress is an important guarantee for stable yield and survival of rice (Ball and Anderson 1986; Smirnoff and Colombé 1988; Wang et al. 2012; Baghel et al. 2016). In this study, salt-alkaline stress significantly affected the four cultivars’ carbon and nitrogen metabolisms. There were also significant differences between salt-alkaline tolerant rice cultivars and salt-alkaline sensitive rice cultivars under salt-alkaline stress. The results of correlation analysis shown that carbon and nitrogen metabolism traits are closely related to yield and NUE.
Rice yield is the result of the process of production, accumulation, and distribution of photosynthetic products, which are the material basis for the formation of rice yield. Rice leaves are the main source of photosynthesis, whereas rice panicles are organic matter storage and the ultimate expression of rice economic output (Kato et al. 2004). In the current study, Pn and PNUE reductions in CB9 and DD12 were significantly lower than JND138 and TY315 under the salt-alkaline conditions. It may be that the salt-alkaline tolerant cultivars have higher Rubisco activity under the salt-alkaline conditions. Rice grain yield comes from the transportation of storage substances in vegetative organs before heading and the direct transportation of leaf photosynthetic products to the grain after heading (Masoni et al. 2007). Under stress conditions, the soluble sugar and sucrose in rice acts as an osmotic regulator and a nutrient that protects the biofilm and its protein (Yildiztugay et al. 2014; Guo et al. 2018). In this study, the soluble sugar and sucrose contents were increased in CB9 and DD12, and were significantly higher in these cultivars than in JND138 and TY315 under the salt-alkaline conditions: a result that is consistent with findings of previous studies (Ghoulam et al. 2002; Li et al. 2017). Sucrose is the main product of plant photosynthesis, and is main form of carbohydrate used in transportation. The SPS enzyme is a key regulatory enzyme for sucrose metabolism, and its activity is closely related to the dry matter accumulation and yield formation of rice. In this study, SPS activity significantly increased under salt-alkali stress conditions. Plaut et al. (2004) showed that there may be competition between the development of grain and need assimilates of leaves to adapt to stress under water or salt stress. These observations indicate that part of the total assimilate from rice leaves is transported to the panicle under saline-alkali stress, whereas some assimilates need to be retained for osmotic adjustment.
Soluble protein content is an important osmotic adjustment substance in plants, and plays an key role in relieving the damage caused by stress. The level of soluble protein content is closely related to plant nitrogen metabolism and senescence (Xin et al. 2019). In this study, the soluble protein content of rice leaves decreased significantly under salt-alkaline stress. Additionally, it may be due to the salt-alkaline stress that the rice is in a state of nitrogen deficiency for a long time, and the plant accumulates free amino acids through protein hydrolysis, which may be an adaptive mechanism of the plant. NR is the rate-limiting enzyme of nitrogen assimilation that is regulated by nitrate, light, and stress. In this study, salt-alkaline stress significantly reduced NR activity in rice, consistent with results of previous studies (Wang et al. 2007; Shao et al. 2015). This change may be induced by a decrease in nitrate content of leaves caused by salt-alkaline stress. Zeng and Shannon (2000b) showed that salt-alkaline stress significantly increased the NR activity in salt-tolerant rice buds, but significantly reduced the NR activity in salt-sensitive rice buds at the bud stage. In this study, the decrease in NR activity of salt-alkaline tolerant cultivars leaves at the filling stage under saline-alkali stress was less than that of salt-alkaline-sensitive cultivars, showing a certain degree of adaptability to salt-alkaline stress. This indicates that the plant’s ability to tolerate salt-alkaline conditions may also be related to its ability to quickly adapt to the environment of nitrogen deficiency. Cao et al. (2008) reported that higher GS activities were beneficial for N absorption and accumulation, which are key enzymes involved in plants N metabolism. In this study, compared with the black soil conditions, the GS activity in CB9, DD12, and TY315 increased under the salt-alkaline soil conditions, whereas GS activity of JND138 decreased. These results indicate that higher GS activity is important for maintaining the stability of NUE under the salt-alkaline stress. Salt-alkaline stress produced a significant effect on the grain yield differences between rice cultivars with contrasting salt-alkaline resistance. Overall, compared with salt-alkaline sensitive cultivars, salt-alkaline tolerant cultivars obtained more N from the soil under salt-alkaline conditions. At the same time, salt-alkaline tolerant cultivars NUE and PNUE were significantly higher than salt-alkaline sensitive cultivars, indicating that under salt-alkaline stress, the salt-alkaline tolerant cultivars can obtain more N, provide more basic substances and energy for carbon metabolism, and thus obtain higher yields.
Conclusions
Most previous studies on salt-alkaline effects on plant growth have used chemical agents to simulate salt-alkaline stress. In this study, black soil and saline-alkaline soil were tested in the field, so results can directly guide the selection and breeding of salt-alkaline tolerant rice varieties and the improvement of cultivation measures on these soils. In our study, four cultivars with similar growth periods showed different yields, NUE, and carbon and nitrogen metabolisms under the two soil types. Our results show that the limitation of rice yield was mainly due to changes in effective panicles and N acquisition capacity under salt-alkaline conditions. The results also demonstrated that the salt-alkaline tolerant cultivars CB9 and DD12 have higher carbon and nitrogen compound contents and enzyme activity to maintain a more stable osmotic balance, and maintain carbon and nitrogen metabolisms. This shows that under salt-alkaline stress, higher carbon and nitrogen metabolism capacity is an important guarantee for stable rice yield. The nitrogen use efficiency (NUEb and NUEg) of CB9 under black soil conditions was lower than that of TY315 and JND138, whereas the opposite was true under salt-alkaline soil conditions, indicating that nitrogen use efficiency can be used as an important indicator for evaluating the salt-alkaline of rice under salt-alkaline stress. This study provides a basis for breeding salt-alkaline tolerant rice cultivars and the improvement of cultivation measures, which are important for the sustainable development of rice production.
Conflicts of interest
The authors declare no conflicts of interest.
Author contributions
All authors conceived and designed the experiments; Liying Guo and Yanqiu Geng performed the experiments; Liying Guo analysed the data and wrote the paper. All authors have read and approved the final manuscript.
Acknowledgements
This research was supported by the National Key Research and Development Program of China (2018YFD0300207-3); Key Scientific and Technological Research Projects of Jilin Province, China (20200402004NC); Key laboratory of Straw Biology and Utilisation (Jilin Agricultural University), Ministry of Education, Jilin Province Education Department ‘the 13th Five-Year Plan’ Science and Technology Research Program (JJKH20190933KJ).
References
Abdullah Z, Khan MA, Flowers T (2001) Causes of sterility in seed set of rice under salinity stress. Journal of Agronomy & Crop Science 187, 25–32.| Causes of sterility in seed set of rice under salinity stress.Crossref | GoogleScholarGoogle Scholar |
Abouelsaad I, Weihrauch D, Renault S (2016) Effects of salt stress on the expression of key genes related to nitrogen assimilation and transport in the roots of the cultivated tomato and its wild salt-tolerant relative. Scientia Horticulturae 211, 70–78.
| Effects of salt stress on the expression of key genes related to nitrogen assimilation and transport in the roots of the cultivated tomato and its wild salt-tolerant relative.Crossref | GoogleScholarGoogle Scholar |
Baghel L, Kataria S, Guruprasad KA (2016) Static magnetic field treatment of seeds improves carbon and nitrogen metabolism under salinity stress in soybean. Bioelectromagnetics 37, 455–470.
| Static magnetic field treatment of seeds improves carbon and nitrogen metabolism under salinity stress in soybean.Crossref | GoogleScholarGoogle Scholar |
Ball MC, Anderson JM (1986) Sensitivity of photosystems ii to NaCl in relation to salinity tolerance. comparative studies with thylakoids of the salt tolerant mangrove, Avicennia marina, and the salt-sensitive pea, Pisum sativum. Functional Plant Biology 13, 689–698.
| Sensitivity of photosystems ii to NaCl in relation to salinity tolerance. comparative studies with thylakoids of the salt tolerant mangrove, Avicennia marina, and the salt-sensitive pea, Pisum sativum.Crossref | GoogleScholarGoogle Scholar |
Cao Y, Fan XR, Sun SB, Xu GH, Hu J, Shen QR (2008) Effect of nitrate on activities and transcript levels of nitrate reductase and glutamine synthetase in rice. Pedosphere 18, 664–673.
| Effect of nitrate on activities and transcript levels of nitrate reductase and glutamine synthetase in rice.Crossref | GoogleScholarGoogle Scholar |
Cheng S, Cao L, Zhuang J, Chen S, Zhan X, Fan Y, Zhu D, Min S (2007) Super hybrid rice breeding in China: achievements and prospects. Journal of Integrative Plant Biology 49, 805–810.
| Super hybrid rice breeding in China: achievements and prospects.Crossref | GoogleScholarGoogle Scholar |
Croft H, Chen JM, Zhang Y, Simic A (2013) Modelling leaf chlorophyll content in broadleaf and needle leaf canopies from ground, CASI, Landsat TM 5 and MERIS reflectance data. Remote Sensing of Environment 133, 128–140.
| Modelling leaf chlorophyll content in broadleaf and needle leaf canopies from ground, CASI, Landsat TM 5 and MERIS reflectance data.Crossref | GoogleScholarGoogle Scholar |
Dluzniewska P, Gessler A, Dietrich H, Schnitzler JP, Rennenberg H (2007) Nitrogen uptake and metabolism in Populus × canescens as affected by salinity. New Phytologist 173, 279–293.
| Nitrogen uptake and metabolism in Populus × canescens as affected by salinity.Crossref | GoogleScholarGoogle Scholar |
El-Shabrawi H, Kumar B, Kaul T, Reddy MK, Singla-Pareek SL, Sopory SK (2010) Redox homeostasis, antioxidant defense, and methylglyoxal detoxification as markers for salt tolerance in Pokkali rice. Protoplasma 245, 85
| Redox homeostasis, antioxidant defense, and methylglyoxal detoxification as markers for salt tolerance in Pokkali rice.Crossref | GoogleScholarGoogle Scholar |
Eraslan F, Inal A, Gunes A, Alpaslan M (2007) Impact of exogenous salicylic acid on the growth, antioxidant activity and physiology of carrot plants subjected to combined salinity and boron toxicity. Scientia Horticulturae 113, 120–128.
| Impact of exogenous salicylic acid on the growth, antioxidant activity and physiology of carrot plants subjected to combined salinity and boron toxicity.Crossref | GoogleScholarGoogle Scholar |
Flowers TJ, Yeo AR (1995) Breeding for salinity resistance in crop plants: where next? Australian Journal of Plant Physiology 22, 875–884.
Ghoulam C, Foursy A, Fares K (2002) Effects of salt stress on growth, inorganic ions and proline accumulation in relation to osmotic adjustment in five sugar beet cultivars. Environmental and Experimental Botany 47, 39–50.
| Effects of salt stress on growth, inorganic ions and proline accumulation in relation to osmotic adjustment in five sugar beet cultivars.Crossref | GoogleScholarGoogle Scholar |
Gibon Y, Blaesing OE, Hannemann J, Carillo P, Höhne M, Hendriks JHM, Palacios N, Cross J, Selbig J, Stitt M (2004) A robot-based platform to measure multiple enzyme activities in Arabidopsis using a set of cycling assays: comparison of changes of enzyme activities and transcript levels during diurnal cycles and in prolonged darkness. The Plant Cell 16, 3304–3325.
| A robot-based platform to measure multiple enzyme activities in Arabidopsis using a set of cycling assays: comparison of changes of enzyme activities and transcript levels during diurnal cycles and in prolonged darkness.Crossref | GoogleScholarGoogle Scholar |
Guo YY, Yu HY, Yang MM, Kong DS, Zhang YJ (2018) Effect of drought stress on lipid peroxidation, osmotic adjustment and antioxidant enzyme activity of leaves and roots of Lycium ruthenicum Murr. seedling. Russian Journal of Plant Physiology 65, 244–250.
| Effect of drought stress on lipid peroxidation, osmotic adjustment and antioxidant enzyme activity of leaves and roots of Lycium ruthenicum Murr. seedling.Crossref | GoogleScholarGoogle Scholar |
Han XF, Wang L, Yang JM (2016) An extension of two-Higgs-doublet model and the excesses of 750 GeV diphoton, muon g-2 and h → μτ. Physics Letters. [Part B] 757, 537–547.
| An extension of two-Higgs-doublet model and the excesses of 750 GeV diphoton, muon g-2 and h → μτ.Crossref | GoogleScholarGoogle Scholar |
He Y, Zhu ZJ (2008) Exogenous salicylic acid alleviates NaCl toxicity and increases antioxidative enzyme activity in Lycopersicon esculentum. Biologia Plantarum 52, 792–795.
| Exogenous salicylic acid alleviates NaCl toxicity and increases antioxidative enzyme activity in Lycopersicon esculentum.Crossref | GoogleScholarGoogle Scholar |
Ju C, Buresh RJ, Wang Z, Zhang H, Liu L, Yang J, Zhang J (2015) Root and shoot traits for rice varieties with higher grain yield and higher nitrogen use efficiency at lower nitrogen rates application. Field Crops Research 175, 47–55.
| Root and shoot traits for rice varieties with higher grain yield and higher nitrogen use efficiency at lower nitrogen rates application.Crossref | GoogleScholarGoogle Scholar |
Kabir AH, Zaman R, Begum MC, Haque A, Haider SA (2016) Upregulation of, OsNAS1, OsPCS1, and DREB1A transcripts along with anti- oxidative defense confers salt tolerance in rice (Oryza sativa L. cv Pokkali). Archives of Agronomy and Soil Science 62, 1381–1395.
| Upregulation of, OsNAS1, OsPCS1, and DREB1A transcripts along with anti- oxidative defense confers salt tolerance in rice (Oryza sativa L. cv Pokkali).Crossref | GoogleScholarGoogle Scholar |
Kato M, Kobayashi K, Ogiso E, Yokoo M (2004) Photosynthesis and dry-matter production during ripening stage in a female-sterile line of rice. Plant Production Science 7, 184–188.
| Photosynthesis and dry-matter production during ripening stage in a female-sterile line of rice.Crossref | GoogleScholarGoogle Scholar |
Krishnamurthy P, Ranathunge K, Franke R, Prakash HS, Schreiber L, Mathe MK (2009) The role of root apoplastic transport barriers in salt tolerance of rice (Oryza sativa L.). Planta 230, 119–134.
| The role of root apoplastic transport barriers in salt tolerance of rice (Oryza sativa L.).Crossref | GoogleScholarGoogle Scholar |
Li Q, Yang A, Zhang WH (2017) Comparative studies on tolerance of rice genotypes differing in their tolerance to moderate salt stress. BMC Plant Biology 17, 141
| Comparative studies on tolerance of rice genotypes differing in their tolerance to moderate salt stress.Crossref | GoogleScholarGoogle Scholar |
Li X, Zheng H, Wu W, Liu H, Wang J, Jia Y, Li J, Yang L, Lei L, Zou D, Zhao H (2020) QTL mapping and candidate gene analysis for alkali tolerance in Japonica rice at the bud stage based on linkage mapping and genome-wide association study. Rice 13, 48
| QTL mapping and candidate gene analysis for alkali tolerance in Japonica rice at the bud stage based on linkage mapping and genome-wide association study.Crossref | GoogleScholarGoogle Scholar |
Maness N (2010) Extraction and analysis of soluble carbohydrates. Methods in Molecular Biology 639, 341–370.
| Extraction and analysis of soluble carbohydrates.Crossref | GoogleScholarGoogle Scholar |
Masoni A, Ercoli L, Mariott M, Arduini I (2007) Post-anthesis accumulation and remobilization of dry matter, nitrogen and phosphorus in durum wheat as affected by soil type. European Journal of Agronomy 26, 179–186.
| Post-anthesis accumulation and remobilization of dry matter, nitrogen and phosphorus in durum wheat as affected by soil type.Crossref | GoogleScholarGoogle Scholar |
Nakamura Y, Yuki K, Park SY, Toshihide O (1989) Carbohydrate metabolism in the developing endosperm of rice grains. Plant & Cell Physiology 30, 833–839.
| Carbohydrate metabolism in the developing endosperm of rice grains.Crossref | GoogleScholarGoogle Scholar |
Nemati I, Moradi F, Gholizadeh S, Esmaeili M, Bihamta M (2011) The effect of salinity stress on ions and soluble sugars distribution in leaves, leaf sheaths and roots of rice (Oryza sativa L.) seedlings. Plant, Soil and Environment 57, 26–33.
| The effect of salinity stress on ions and soluble sugars distribution in leaves, leaf sheaths and roots of rice (Oryza sativa L.) seedlings.Crossref | GoogleScholarGoogle Scholar |
Nieman RH, Clark RA (1976) Interactive effects of salinity and phosphorus nutrition on concentrations of phosphate and phosphate esters in mature photosynthesizing corn leaves. Plant Physiology 57, 157–161.
| Interactive effects of salinity and phosphorus nutrition on concentrations of phosphate and phosphate esters in mature photosynthesizing corn leaves.Crossref | GoogleScholarGoogle Scholar |
Plaut Z, Butow BJ, Blumenthal CS, Wrigley CW (2004) Transport of dry matter into developing wheat kernels and its contribution to grain yield under post-anthesis water deficit and elevated temperature. Field Crops Research 86, 185–198.
| Transport of dry matter into developing wheat kernels and its contribution to grain yield under post-anthesis water deficit and elevated temperature.Crossref | GoogleScholarGoogle Scholar |
Qadir M, Quillerou E, Nangia V, Murtaza G, Singh M, Thomas R, Drechsel P, Noble A (2014) Economics of salt-induced land degradation and restoration. Natural Research Forum 38, 282–295.
| Economics of salt-induced land degradation and restoration.Crossref | GoogleScholarGoogle Scholar |
Shao QS, Shu S, Du J, Xing WW, Guo SR, Sun J (2015) Effects of NaCl stress on nitrogen metabolism of cucumber seedlings. Russian Journal of Plant Physiology 62, 595–603.
| Effects of NaCl stress on nitrogen metabolism of cucumber seedlings.Crossref | GoogleScholarGoogle Scholar |
Shobbar MS, Azhari O, Shobbar ZS, Niknam V, Askari H, Pessarakli M, Ebrahimzadeh H (2012) Comparative analysis of some physiological responses of rice seedlings to cold, salt, and drought stress. Journal of Plant Nutrition 35, 1037–1052.
| Comparative analysis of some physiological responses of rice seedlings to cold, salt, and drought stress.Crossref | GoogleScholarGoogle Scholar |
Smirnoff N, Colombé SV (1988) Drought influences the activity of enzymes of the chloroplast hydrogen peroxide scavenging system. Journal of Experimental Botany 39, 1097–1108.
| Drought influences the activity of enzymes of the chloroplast hydrogen peroxide scavenging system.Crossref | GoogleScholarGoogle Scholar |
Stevens J, Senaratna T, Sivasithamparam K (2006) Salicylic acid induces salinity tolerance in tomato (Lycopersicon esculentum cv. Roma): associated changes in gas exchange, water relations and membrane stabilisation. Plant Growth Regulation 49, 77–83.
Sun H, Qian Q, Wu K, Luo J, Wang S, Zhang C, Ma Y, Liu Q, Huang X, Yuan Q, Han R, Zhao M, Dong G, Guo L, Zhu X, Gou Z, Wang W, Wu Y, Lin H, Fu X (2014) Heterotrimeric G proteins regulate nitrogen-use efficiency in rice. Nature Genetics 46, 652–656.
| Heterotrimeric G proteins regulate nitrogen-use efficiency in rice.Crossref | GoogleScholarGoogle Scholar |
Surabhi GK, Reddy AM, Kumari GJ, Sudhakar C (2008) Modulations in key enzymes of nitrogen metabolism in two high yielding genotypes of mulberry (Morus alba L.) with differential sensitivity to salt stress. Environmental and Experimental Botany 64, 171–179.
| Modulations in key enzymes of nitrogen metabolism in two high yielding genotypes of mulberry (Morus alba L.) with differential sensitivity to salt stress.Crossref | GoogleScholarGoogle Scholar |
Tian Z, Li J, Jia X, Yang F, Wang Z (2016) Assimilation and translocation of dry matter and phosphorus in rice genotypes affected by salt-alkaline stress. Sustainability 8, 568
| Assimilation and translocation of dry matter and phosphorus in rice genotypes affected by salt-alkaline stress.Crossref | GoogleScholarGoogle Scholar |
Wang H, Lee P, Chen W, Huang D, Su J (2000) Osmotic stress induced changes of sucrose metabolism in cultured sweet potato cells. Journal of Experimental Botany 51, 1991–1999.
| Osmotic stress induced changes of sucrose metabolism in cultured sweet potato cells.Crossref | GoogleScholarGoogle Scholar |
Wang ZQ, Yuan YZ, Ou JQ, Lin QH, Zhang CF (2007) Glutamine synthetase and glutamate dehydrogenase contribute differentially to proline accumulation in leaves of wheat (Triticum aestivum) seedlings exposed to different salinity. Journal of Plant Physiology 164, 695–701.
| Glutamine synthetase and glutamate dehydrogenase contribute differentially to proline accumulation in leaves of wheat (Triticum aestivum) seedlings exposed to different salinity.Crossref | GoogleScholarGoogle Scholar |
Wang H, Zhang M, Guo R, Shi D, Liu B, Lin X, Yang C (2012) Effects of salt stress on ion balance and nitrogen metabolism of old and young leaves in rice (Oryza sativa L.). BMC Plant Biology 12, 194
| Effects of salt stress on ion balance and nitrogen metabolism of old and young leaves in rice (Oryza sativa L.).Crossref | GoogleScholarGoogle Scholar |
Woodrow P, Ciarmiello LF, Annunziata MG, Pacifico S, Iannuzzi F, Mirto A, D’Amelia L, Dell’Aversana E, Piccolella S, Fuggi A, Carillo P (2017) Durum wheat seedling responses to simultaneous high light and salinity involve a fine reconfiguration of amino acids and carbohydrate metabolism. Physiologia Plantarum 159, 290–312.
| Durum wheat seedling responses to simultaneous high light and salinity involve a fine reconfiguration of amino acids and carbohydrate metabolism.Crossref | GoogleScholarGoogle Scholar |
Xin W, Zhang L, Zhang W, Gao J, Yi J, Zhen X, Li Z, Zhao Y, Peng C, Zhao C (2019) An integrated analysis of the rice transcriptome and metabolome reveals differential regulation of carbon and nitrogen metabolism in response to nitrogen availability. International Journal of Molecular Sciences 20, 2349
| An integrated analysis of the rice transcriptome and metabolome reveals differential regulation of carbon and nitrogen metabolism in response to nitrogen availability.Crossref | GoogleScholarGoogle Scholar |
Yang C, Chong J, Li C, Ki C, Shi D, Wang D (2007) Osmotic adjustment and ion balance traits of an alkali resistant halophyte Kochia sieversiana during adaptation to salt and alkali conditions. Plant and Soil 294, 263–276.
| Osmotic adjustment and ion balance traits of an alkali resistant halophyte Kochia sieversiana during adaptation to salt and alkali conditions.Crossref | GoogleScholarGoogle Scholar |
Yildiztugay E, Ozfidan-Konakci C, Kucukoduk M, Duran Y (2014) Variations in osmotic adjustment and water relations of Sphaerophysa kotschyana: glycine, betaine, proline and choline accumulation in response to salinity. Botanical Studies 55, 6
| Variations in osmotic adjustment and water relations of Sphaerophysa kotschyana: glycine, betaine, proline and choline accumulation in response to salinity.Crossref | GoogleScholarGoogle Scholar |
Zeng L, Shannon MC (2000a) Salinity effects on seedling growth and yield components of rice. Crop Science 40, 996–1003.
| Salinity effects on seedling growth and yield components of rice.Crossref | GoogleScholarGoogle Scholar |
Zeng L, Shannon MC (2000b) Effects of salinity on grain yield and yield components of rice at different seeding densities. Agronomy Journal 92, 418–423.
| Effects of salinity on grain yield and yield components of rice at different seeding densities.Crossref | GoogleScholarGoogle Scholar |
Zhang JT, Mu CS (2009) Effects of saline and alkaline stresses on the germination, growth, photosynthesis, ionic balance and anti-oxidant system in an alkali-tolerant leguminous forage Lathyrus quinquenervius. Soil Science and Plant Nutrition 55, 685–697.
| Effects of saline and alkaline stresses on the germination, growth, photosynthesis, ionic balance and anti-oxidant system in an alkali-tolerant leguminous forage Lathyrus quinquenervius.Crossref | GoogleScholarGoogle Scholar |
Zhang ZL, Qu WJ (2003) ‘Experimental guidance in plant physiology.’ 3rd edn. (Higher Education Press: Beijing) [in Chinese]
Zhao X, Wang W, Zhang F, Deng J, Li Z, Fu B (2014) Comparative metabolite profiling of two rice genotypes with contrasting salt stress tolerance at the seedling stage. PLoS One 9, e108020
| Comparative metabolite profiling of two rice genotypes with contrasting salt stress tolerance at the seedling stage.Crossref | GoogleScholarGoogle Scholar |
1 Under salt-alkaline conditions, most of the indicators of rice will decrease; the relative decrease of different genotypes is different. The ‘less relative decrease’ indicates that rice of this genotype is more stable and has a certain tolerance under salt-alkaline conditions.