

The pros and cons of increasing soil organic matter in dryland cropping systems
Chelsea Janke A B * , John Kirkegaard C , James Hunt A D , Louise Barton E , Lindsay Bell F , Senani Karunaratne C , Lynne M. Macdonald A , Chiara Pasut A , Uta Stockmann C , Ehsan Tavakkoli G , Gupta Vadakattu A , Anton Wasson C and Mark Farrell A E *
A B * , John Kirkegaard C , James Hunt A D , Louise Barton E , Lindsay Bell F , Senani Karunaratne C , Lynne M. Macdonald A , Chiara Pasut A , Uta Stockmann C , Ehsan Tavakkoli G , Gupta Vadakattu A , Anton Wasson C and Mark Farrell A E *
A
B
C
D
E
F
G
Abstract
Soil organic matter (SOM) in drylands accounts for approximately 33% of global soil organic carbon (SOC) stocks and regulates many processes. Anthropogenic activities and climatic changes have influenced, and continue to significantly influence SOM contents. However, management practices that improve the soil carbon (C) and macronutrient balance can increase or maintain SOM. These include (1) maximising C inputs from grain crops, (2) integrating livestock and pasture phases, (3) using cover crops, (4) intercropping, (5) managing tillage and stubble, and (6) organic amendments. Estimated SOC increases achievable in drylands, ranging from 60 to 114 kg C ha year−1, fall short of the ambitious ‘4 per mille’ target, which is equivalent to a 0.4% annual increase in initial soil C, or at least 240 kg C ha−1 year−1 for drylands (assuming a global mean dryland SOC stock of 60 Mg C ha−1). In dryland systems, we propose a more rational approach, advocating for context-specific optima with a clear understanding of the benefits and costs to evaluate the suitability of management practices for improving SOM. The benefits include amelioration of soil constraints, improving nutrient and water availability, enhancing system resilience and sustainability, and potential participation in C markets. However, costs can be significant and are typically divided into the following two main categories: (1) economic (e.g. financial costs required for implementing management practices), and (2) environmental (e.g. the potential for increased nutrient loss via emissions or leaching as a result of enhanced nutrient cycling). The net benefit or cost is highly context-dependent, with the unique challenges of dryland environments being often overlooked in the literature. This review examines the primary strategies for maintaining or increasing SOM in dryland arable systems, the associated benefits and costs, methods for monitoring SOC stocks, and future challenges and opportunities.
Keywords: agronomy, carbon management, dry-land, organic carbon, rainfed, resilience, soil carbon sequestration, soil health.
Introduction
Drylands cover 41% of the Earth’s land surface, and arable systems within these regions produce 44% of the world’s crops (UNCCD 2017). These farming systems are found primarily in Mediterranean climates, where more than 65% of annual rainfall occurs in the winter, but they are also present in subtropical and subhumid regions where rainfall is sparse, irregular, and unevenly distributed (Koohafkan and Stewart 2008; Osman 2018; Fig. 1). Agriculture in dryland regions is often characterised by low soil fertility and inputs, which typically limit plant productivity (Novelli et al. 2023).
Global distribution of dryland agroecosystems in subhumid and semiarid regions. Note: minimal agricultural production occurs in the hyper-arid regions. Figure has been reproduced with permission by the Zoi Environment Network (2011).
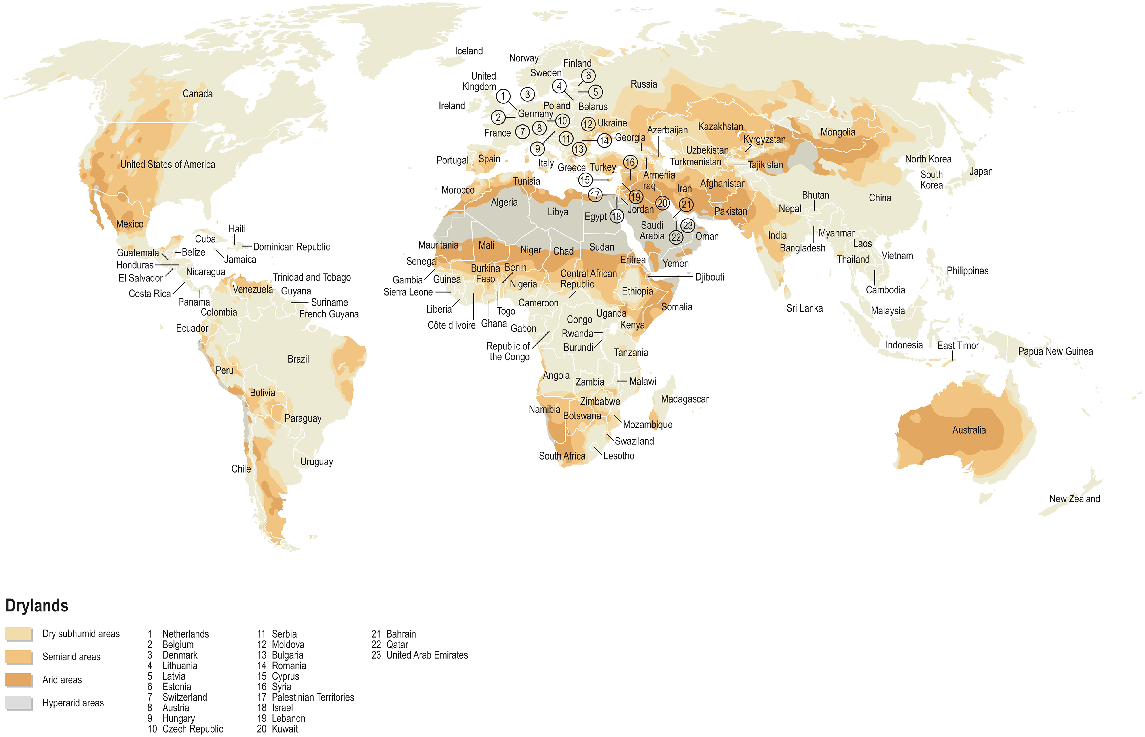
Despite their low fertility and soil organic matter (SOM) concentrations, drylands account for approximately 33% of global carbon (C) stocks (Safriel et al. 2005). Soil organic matter is a vital component of these soils, providing essential functions that support cost-effective primary production (Lal 2004). Key benefits of SOM include enhanced nutrient cycling, improved soil structure and water retention, and increased cation exchange and buffering capacities (Tiessen et al. 1994; Murphy 2014; Schjonning et al. 2018).
The term ‘soil health’ has been widely used within the context of agricultural sustainability (Kibblewhite et al. 2008), with its origins in the 1990s (Doran and Safley 1997). It has recently been defined by the USDA as ‘the continued capacity of soil to function as a vital living ecosystem that sustains plants, animals, and humans’ (p. 544; Lehmann et al. 2020). It is suggested that if soil organic C (SOC) falls below a threshold of ~2% (~3.4% SOM), then soil health may deteriorate significantly (Loveland and Webb 2003; Huber et al. 2008; Musinguzi et al. 2013, 2016). However, critical SOC concentrationss can vary depending on soil type and context. For instance, many Australian soils in the cropping belt contain less than 2% SOC (Baldock et al. 2013). The risks associated with SOM decline in drylands are increasingly recognised globally, leading to the development of technical briefs (Laban et al. 2018), frameworks for evaluating land-use impacts (FAO 2019; Fu et al. 2021; Sims et al. 2021), and governance policies (Stafford-Smith and Metternicht 2021). Carbon markets are one strategy implemented by several governing bodies (European Commission 2023), trading C credits to offset a large greenhouse-gas (GHG) footprint or for economic gain. However, it is unlikely that a C market approach will succeed in most dryland regions, where other socioeconomic and political factors take precedence (Plaza-Bonilla et al. 2015). Resilience of yield to drought also appears related to SOM (Kane et al. 2021).
Mitigating further SOM decline in drylands is crucial because increasing SOM is challenging, even under optimal conditions. Widely accepted practices to increase SOM include increasing C inputs through various methods (Schimel 2013; Machmuller et al. 2015; Pausch and Kuzyakov 2017), ensuring adequate co-nutrient application to enhance microbial C use efficiency (CUE; Kirkby et al. 2016; Kirkegaard et al. 2023), improving C stabilisation within soils (Janzen et al. 2022; Whalen et al. 2022; Tao et al. 2023), minimising soil disturbance (Dungait et al. 2019; Wu et al. 2023), and limiting soil erosion (Brooks et al. 2007).
Estimates of the C sequestration potential in drylands range from approximately 60–114 kg C ha−1 year−1 to a depth of 1 m, depending on various scenarios (Lal 2004). These values are even smaller, ~11–18 kg C ha−1 year−1, when considering only the top 0.3 m of soil, which is most affected by management techniques, in more common cropping/pasture contexts (Hoyle et al. 2013). This dryland C sequestration potential falls far short of the ambitious ‘quatre pour mille’ (4/1000) goal, which advocates for a 0.4% annual increase of the standing SOC stock (Minasny et al. 2017). The disparity suggests that context-specific targets for C sequestration in soils that consider climate, soil, and management factors are more appropriate than are broadly applied targets.
The upper limit of SOM accumulation and organic C storage in soils is a topic of ongoing debate. Some scientists argue that soils are an unlimited sink for C capture and storage (Vogel et al. 2014; Schweizer 2022; Begill et al. 2023), whereas others suggest that soils become C saturated, with certain soil types and environments having practical upper thresholds for C storage (Stewart et al. 2007; Cotrufo et al. 2019; Georgiou et al. 2022; Singh and Benbi 2023). Given the diverse and unique constraints on SOM accumulation in different soils, quantifying the upper limit potential of C sequestration is challenging, particularly as rates of accumulation change over time. This upper limit is likely to be dynamic rather than static, varying with C flows (i.e. inputs and losses) and changing constraints, such as microbial activity, nutrient deficiency, crop productivity, and soil structure (Karunaratne et al. 2014; Mattila and Vihanto 2024). Nevertheless, there is a consensus on the importance of maintaining or increasing SOM in soils.
This review examines current approaches to maintaining and increasing SOM in dryland arable systems, the associated benefits and costs of these strategies, methods for monitoring SOM, and future challenges and opportunities for SOM management. Although pasture phases and livestock inclusion are discussed as features of a rotation system, pasture dominated systems are not the focus of this review. The reader is directed to other reviews (e.g. Eyles et al. 2015; Whitehead 2020) that focus on management of livestock production systems to increase SOC.
Growing soil organic matter
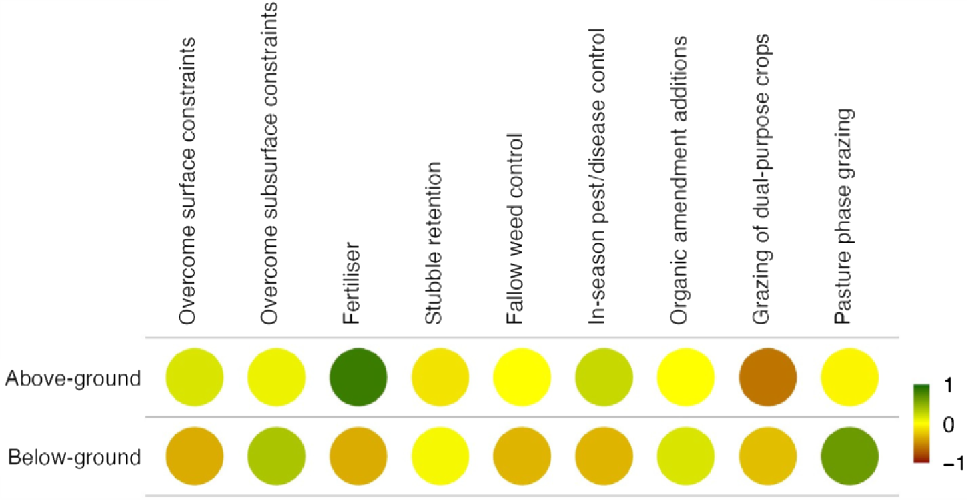
The primary source of C input to soils is C fixed through plant photosynthesis. Therefore, net primary productivity (NPP), i.e. the balance of C fixed in plant tissue by photosynthesis minus C lost by autotrophic respiration as C dioxide (CO2) in above- and below-ground components (Fig. 2), sets the upper limit for C inputs (Janzen et al. 2022). However, numerous factors influence NPP and, consequently, the availability of C for incorporation into the soil (Minasny et al. 2022). Additionally, over 90% of the added C from plant inputs can be lost through various pathways, including (1) respiration as CO2 (Angers et al. 2022), (2) leaching as dissolved organic C (DOC), and (3) erosion via aeolian processes or intense rainfall (Quinton et al. 2010; Chappell et al. 2016). The increasing prevalence of extreme weather events as a result of global heating is intensifying these C-loss pathways, particularly in topsoils, which are more exposed and typically contain higher SOC concentrations (Plaza et al. 2018; Lian et al. 2021; Wang et al. 2022).
Qualitative visualisation of the influence of various factors on above- and below-ground plant inputs of C. Green indicates a positive effect; and brown indicates a negative effect.
Mechanisms and properties of in situ C inputs and losses
The quality and degradability of above-ground inputs vary significantly (Fig. 2) depending on the plant part (e.g. stems, leaves, chaff), crop type (e.g. legumes vs monocotyledonous grains), and the development stage (e.g. young vs mature plants). Plant residues with high (>20) C:nitrogen (N) ratios tend to degrade more slowly because nutrient availability, particularly N, limits microbial activity (Heal et al. 1996), which immobilises available nutrient pools within decomposing residue and the surrounding soil, limiting crop nutrient availability. In contrast, plant residues with low (<20) C:N ratios (e.g. legumes, some crop leaves) tend to degrade rapidly, and the proportion of residue C respired by microbes may be lower because of more favourable C:nutrient ratios for microbial biomass growth. Faeces from grazing animals also typically add organic material with a lower C:nutrient ratio (Chen et al. 2003).
The amount of above-ground biomass C incorporated into the soil, particularly below the immediate surface layer (0–5 cm), varies significantly depending on management practices and soil characteristics. In dryland systems that use regular tillage, above-ground biomass is incorporated within the plough layer, typically 0–20 cm. However, over the past three decades, reduced (i.e. strategic, minimal) or zero-tillage systems have become more common and almost exclusively employed in some regions, such as Australian dryland cropping systems (Llewellyn et al. 2012). As a result, much above-ground biomass remains at the soil surface as a mulch layer following senescence. Although this mulch can protect the soil surface from erosion and improve water infiltration to depth, both being critical processes in dryland systems, only limited quantities (2–16%) of C readily enter the soil matrix through leaching (Soong et al. 2015). Incorporation of surface residue C into the soil matrix relies on a combination of macrofaunal activity (e.g. bioturbation by earthworms and termites), below-ground fungal degradation and transfer via hyphal networks, leaching of DOC following above-ground biotic or abiotic (e.g. UV) degradation, or pedoturbation in shrink-swell soils with vertic properties (López-Bellido et al. 2020). Innovations in machinery, such as stripper fronts, windrowers, and weed-seed destructors on harvesters, also alter the physical form of above-ground biomass remaining on the surface after harvest (Desbiolles et al. 2019). For instance, stripper fronts that remove only the heads of the crop leave a greater proportion of standing stubble, limiting the immediate interaction of much above-ground biomass with the soil. In contrast, low harvest heights and pulverisation of the chaff fraction by seed destructors significantly reduces the physical size of remaining above-ground biomass, potentially altering decomposition kinetics (Angers and Recous 1997). Additionally, removing crop residues (e.g. for sale as animal forage, hay or silage, bedding material, bioenergy production) reduces the amount of above-ground C inputs. In less developed dryland regions where subsistence farming is prevalent, the removal of crop residues, especially for animal feed, is common, and residue retention often does not improve soil fertility, particularly where other inputs (e.g. fertilisers) are scarce (Rusinamhodzi et al. 2015).
Below-ground C inputs in agricultural systems arise from various plant–soil interactions, including rhizodeposition, root decomposition, and relationships with soil microbial biomass (Fig. 2). The zone where below-ground C flux occurs, the rhizosphere, is broadly characterised as the soil immediately surrounding the plant roots, where complex biogeochemical interactions take place (Hinsinger et al. 2009). Due to the complexity of these processes and the difficulty in distinguishing below-ground C inputs from pre-existing SOC, the contribution of below-ground C inputs to total SOC is often overlooked. There remains considerable uncertainty about the extent to which below-ground C inputs contribute to total SOC. Several intrinsic factors of plants and soils, including microbes and fungi, influence the quantity and quality of below-ground C incorporation into SOC, with some being particularly relevant in dryland systems.
Rhizodeposition is the process of plant root material being lost or excreted from plant roots into the surrounding soil, such as sloughed root cells, senesced and lysed root cells, secreted mucilage, and exudation (e.g. sugars, amino compounds, and organic acids), including C flows to symbionts (e.g. mycorrhizas; Jones et al. 2009). These compounds are rich in C and nutrients, resulting in rapid utilisation by the microbial biomass through consumption or decomposition to CO2 (Paterson 2003). The remaining organic C inputs are defined as net rhizodeposition (Pausch and Kuzyakov 2017). Generally, net rhizodeposition processes contribute approximately half as much as root mass to total below-ground C (i.e. rhizodeposition/root ratio of 0.5; Pausch and Kuzyakov 2017). In dryland systems, which are typically infertile and have suboptimal moisture content, below-ground C from rhizodeposition processes may be somewhat reduced; not only is NPP typically lower in drylands, but drier conditions also reduce mass flow through soil profiles, lowering the potential for the diffusive release of many exudate compounds (Jones 1998, 1999; Holz et al. 2018). Grazing of pasture phases also alters rhizodeposition processes as plants reallocate resources in response to defoliation, with the intensity and length of rest periods being important drivers of SOC gains or losses (Sanderman et al. 2015; Bai and Cortrufo 2022).
Roots are a major contributor of C inputs in cropping soils (Balesdent and Balabane 1996), and their morphology and architecture significantly influence below-ground C inputs. Factors such as root type (primary or lateral), extent of branching (Henry et al. 2005; Engedal et al. 2023), and development stage (Personeni et al. 2007) determine rhizodeposition. In annual crops, root exudation is highest at the root apices and lowest in older, lignified primary roots (McCully and Canny 1985). In systems relying on annual crops or pastures, typical of many dryland farming enterprises, most root activity and potential for C deposition occur in the surface soil layers (<30 cm depth). Selection for maximal grain yield in annual crops has also significantly decreased the proportion of C allocated to root systems (Siddique et al. 1990; Aziz et al. 2017). In perennial crops, root systems often explore a greater soil volume and depth, potentially increasing organic C deposition deeper in the soil profile (Kirkby et al. 2016; Dietzel et al. 2017; Hirte et al. 2018; Thorup-Kristensen et al. 2020). However, this is not always the case (Rasse et al. 2006; Taylor et al. 2023a), and factors such as lignin and suberin content in roots can modify decomposition kinetics and the subsequent flow of root-derived C to SOC (Rasse et al. 2005). Moreover, there is a trade-off between decreasing rhizodeposition with rooting depth and increasing microbial stabilisation of those deposits (Peixoto et al. 2020). In dryland systems, agronomic factors affecting root morphology may be especially important for organic C contributions to the soil, as poor root development owing to water limitations, nutrient deficiencies, subsoil constraints, or root diseases can reduce the total volume of root material contributing to organic C inputs (Holz et al. 2018).
Carbon incorporation from organic material is primarily driven by decomposing microbial communities, including heterotrophic microorganisms (e.g. bacteria and fungi) and detritivores (e.g. earthworms, millipedes, enchytraeidae, and termites; Adl 2003). Some of the C is retained within the microbial biomass, whereas the remainder is excreted as metabolites or CO2 (Allison et al. 2010; Schimel 2013). This partitioning of C resources by soil microbes is known as the CUE of the microbial population, expressed as a proportion of new C retained for microbial growth rather than catabolised to CO2. Typically, CUE is higher where simple substrates, such as those found in rhizodeposits, are consumed, as they require less energy for their uptake and processing by microorganisms (Cotrufo et al. 2012). In contrast, more complex substrates demand substantial C expenditure for enzyme production necessary for their degradation, resulting in less C being available for microbial biomass growth. Nutrient availability relative to the amount of C processed also influences CUE, and microbial nutrient requirements are governed by tightly defined C:N:phosphorus (P):sulfur (S) ratios (Redfield 1958; Cleveland and Liptzin 2007).
In dryland agriculture, soil microbial activity is often limited (Gupta et al. 2011). Although the exact impacts of drought and aridity on microbial C metabolism remain unclear (Evans et al. 2022), reduced CO2 respiration and fewer metabolically active microbes are generally observed (Singh et al. 2021; Bian et al. 2022). Furthermore, bacterial and fungal diversity and abundance are typically lower in dryland than temperate systems (Maestre et al. 2015). Limited nutrient availability (e.g. N, P, S) in the generally infertile soils of drylands can further restrict microbial activity and C utilisation (Manzoni et al. 2012; Xu et al. 2015). However, certain microbial strains have shown promise in incorporating C inputs into SOM. Of particular interest are endophytic fungi, which convert simple C compounds (e.g. sugars exuded by plant roots) into complex, polymerised C compounds that resist microbial degradation (Manganyi and Ateba 2020; Liu et al. 2021). Fungal hyphae may also position these complex C forms within the anaerobic core of microaggregates (Killham and Foster 1994), providing further protection from oxidative processes and potentially enhancing long-term C stability (Wilson et al. 2009). Whereas C accumulation in soil by fungi varies among species and biomass (Ekblad et al. 2013; Fernandez and Koide 2014), fungal processes contribute more to C sequestration and stabilisation than do those of other microbial organisms, such as bacteria (Six et al. 2006; Li et al. 2015). Moreover, fungal prevalence increases under dry soil conditions (Haugwitz et al. 2013; De Vries et al. 2018; Singh et al. 2021) and fungi are effective at exploiting even poorly available nutrient resources through their extensive hyphal networks (Rashid et al. 2016; Adomako et al. 2022). Thus, fungi are crucial for effectively incorporating C into dryland agricultural soils (Hannula and Morrien 2022).
Once C enters the soil, it can follow one of the following three pathways: (1) remain unprocessed, (2) be microbially processed and stabilised, or (3) be microbially processed and lost as respired CO2 (Ernakovich et al. 2021), with the latter being the primary loss pathway of SOC in dryland systems. Excluding charcoal-like material (Lehmann et al. 2008), most C in soils is not chemically recalcitrant; instead, it is stabilised via organo-mineral associations or protected within aggregate structures (Dungait et al. 2012; Rassmussen et al. 2018).
The net flow and partitioning of metabolically consumed C is driven primarily by factors affecting microbial CUE, which then controls the amount of C available for SOC storage (Domeignoz-Horta et al. 2020; Tao et al. 2023). However, the relative importance of CUE for C storage efficiency, compared with factors such as C inputs, decomposition rates, and vertical transport, is currently debated (Basile-Doelsch et al. 2020; Button et al. 2022; Janzen et al. 2022; He et al. 2023). In dryland soils, microbial processing of C inputs is episodic, driven by large bursts of activity following wetting events (Fierer and Schimel 2003). Emerging evidence suggests that protozoa and viral diversity may play a key role in these processes (Frey et al. 2001; Geisen et al. 2021; Santos-Medellin et al. 2023). Coupled with the typically low nutrient availability of dryland soils, which tends to lower CUE (Manzoni et al. 2012), this suggests that CUE is typically limited in dryland agricultural soils and may pose a significant barrier to increased SOC stabilisation. Despite the recognised centrality of microbial processing for C stabilisation and turnover, recent findings have indicated that substantial quantities of plant inputs may be directly stabilised without microbial processing (Whalen et al. 2022). Overall, retention of C inputs in soils ranges from approximately 40% to less than 10%, depending on environmental and edaphic factors (Ladd et al. 1995; Berthelin et al. 2022).
Due to the microbially mediated nature of soil C cycling, feedback mechanisms can either result in the loss of existing SOC (Chowbury et al. 2014) or the retention of additional C following the addition of fresh inputs (Weng et al. 2017). This phenomenon, known as the priming effect (PE; Kuzyakov et al. 2000), can be influenced by various factors, including temperature, moisture availability, nutrient availability, soil type, quality of fresh C inputs, and microbial community structure (Coonan et al. 2020; Tang et al. 2023). Limited nutrient and moisture availability in dryland systems generally increases priming as microbes mine nutrients from SOM (Wu et al. 2020). Moreover, certain management practices, such as inversion tillage, deep ripping, or the introduction of deep-rooted crops, can mix microbial communities associated with old SOC and fresh inputs, further influencing PE (Álvaro-Fuentes et al. 2008; Taylor et al. 2023b; Unkovich et al. 2023). Depending on the specific situation, this may result in an increase or a decrease in PE.
Beyond microbial activity, other drivers of SOC losses include erosion and leaching. Although total rainfall in dryland systems is low, episodic events can be intense, resulting in the loss of colloidal and dissolved organic matter (DOM) through lateral erosion and leaching down the soil profile (Brooks et al. 2007).
Agronomic management practices to build soil C
Practices that increase SOM can be categorised into the following four mechanistic strategies: (1) increasing the total amount of fixed C; (2) enhancing the amount of fixed C introduced into the soil; (3) improving the proportion of fixed C converted to SOM (CUE); and (4) managing the influence of fresh inputs on existing SOM via the PE. Some practices operate through a single mechanism, whereas others may affect all four.
The fate of SOM in dryland agriculture has followed a similar trajectory in many regions globally in response to prevailing agronomic practices. Native perennial vegetation was often cleared as sedentary agriculture became the primary food source for human populations. This shift led to a decline in soil fertility because of nutrient removal, as early grazing and cropping practices preceded the widespread use of fertilisers. Additionally, cultivation and fallowing of land, later accelerated by mechanisation, were necessary for weed control and maintaining soil fertility for pastures and crops. However, these practices also resulted in erosional losses and accelerated microbial turnover of SOM. With increased awareness of the critical role of SOM in agricultural production, numerous studies have quantified the impact of various management practices on SOM. The response to most practices is notably variable, making meta-analyses (Guo and Gifford 2002; Alvarez 2005; Sun et al. 2020) valuable tools for understanding the variation and contextual factors influencing outcomes, while noting the potential for meta-analyses themselves to deliver unreliable conclusions (Krupnik et al. 2019).
In arable grain-producing systems, a significant proportion of C fixed by photosynthesis is removed in grain, but the majority remains in above- and below-ground crop residues. If this C is retained in the field, it becomes available for microbial degradation and incorporation into the soil. The effects of residue retention on SOC and crop yield vary with climate, typically providing benefits in dry environments (Sun et al. 2020). Interestingly, systems that remove above-ground residues through burning or physical removal do not always accumulate less SOM (Fettell and Gill 1995; Alvarez 2005), suggesting that factors other than C supply, such as CUE (Tao et al. 2023), limit SOM accumulation in some cases, or that C allocated below the soil surface plays a more important role.
The adoption of minimum or no-till (NT) practices is increasing globally, primarily to protect soil from erosion and reduce moisture loss (Kassam et al. 2022). In dryland systems, savings on fuel, time, labour, and machinery costs further encourage the adoption of these practices. Since most SOM is stabilised by its association with clay minerals (Wu et al. 2023) and through physical protection within aggregate structures (Dungait et al. 2019), the unnecessary disturbance of frequent tillage in agricultural systems exposes previously protected SOC to mineralisation. However, the capacity of NT practices to build appreciable amounts of SOC remains debated (Powlson et al. 2014; Kopittke et al. 2016). A recent global meta-analysis (Cai et al. 2022) indicated that SOC increases are confined to surface soil layers and primarily occur in the early years following NT adoption, whereas losses may continue in deeper soil layers. These findings are likely to stem from the reduced incorporation of surface residues deeper into the soil profile (Cai et al. 2022). Over time, nutrient stratification and compaction layers can limit root growth and exploration, reducing C inputs at depth.
Despite these challenges, the documented benefits of NT practices for soil health (e.g. improved soil structure and fertility, microbial diversity, biological functions) and water retention (Gupta et al. 2019; Mondal and Chakraborty 2022) suggest that strategic and infrequent tillage may help overcome some of the downsides of NT, while retaining the benefits of reduced tillage, which is especially important in dryland regions. For example, residue incorporation may distribute SOC more evenly throughout the soil profile (Helgason et al. 2014), with the potential for further success if supplementary nutrients are applied to assist in SOM formation, creating a nutrient ratio that enhances residue decomposition (Kirkegaard et al. 2023). Where subsoil constraints are present or subsoil horizons (e.g. clay in sandy texture contrast soils), deep ripping or other soil engineering practices may improve crop growth and increase C inputs while enhancing the ability of the soil to retain C (Feng et al. 2020).
Nitrogen plays a crucial role in increasing SOM in the mechanisms mentioned above. As the fourth-most abundant element in SOM after C, oxygen, and hydrogen, N is often the most limiting factor for plant growth in dryland systems (Cossani and Sadras 2018; Hochman and Horan 2018). When N limits plant growth, adding plant-available forms through synthetic fertilisers, organic wastes, or legumes can increase C fixation and thus above- and below-ground biomass. Residues with lower C:N ratios are converted to SOM with fewer respiratory losses, as they more closely match the stoichiometry of microbial biomass. However, arable systems that export large amounts of N in grain can develop a negative mass balance, leading to N-limited SOM formation and a decline in SOC despite a net positive C mass balance (Kirkby et al. 2016). Soil organic matter tends to increase (or decline more slowly) in systems with higher N inputs, whether from legumes (Armstrong et al. 2019), synthetic fertilisers (Alvarez 2005; Ladha et al. 2011; Guo et al. 2012), or organic sources.
In many agricultural systems, opportunities exist to increase C fixation without reducing crop yield or profitability. Among the three primary resources for photosynthesis, water is the most limiting in dryland environments. As a result, practices that increase crop yield in water-limited environments will generally also increase C fixation. Passioura (2006) proposed the following four mechanisms to increase crop yields:
Increase the supply of soil water to crops.
Increase the proportion of soil water supply that transpires while reducing unproductive losses to evaporation, drainage and runoff.
Increase the efficiency of soil water use for CO2 fixation (transpiration efficiency).
Increase the proportion of fixed C allocated to grain (harvest index).
These mechanisms also apply to C fixation in dryland environments, except for increasing the harvest index, which reduces C input efficiency.
The most obvious practice to increase soil water supply is irrigation, although it is relevant only to a small proportion of drylands globally. Well-managed NT systems with stubble retention can also improve the water balance for higher productivity, as discussed earlier. Other practices include ameliorating soil and subsoil constraints, such as acidity, sodicity, salinity, and compaction (Adcock et al. 2007; Sadras et al. 2019), allowing greater root exploration and associated water uptake. Other examples include controlling fallow weeds and maintaining soil cover with crop residues (Hunt et al. 2013). However, although fallowing increases water supply and C fixation during crop growth, the preceding period of lost production can negatively affect C balance. Additionally, crop uptake and export of N in grain that was mineralised during fallow periods generally causes SOC to decrease as fallow intensity increases (Cann et al. 2020a).
Systems with more than one crop per year (e.g. double- or triple-cropping) and a lower proportion of fallow have higher annual biomass production than do those with significant periods of fallow, because less water is lost to evaporation, runoff, and drainage. In dryland agriculture, the primary trade-off for additional C fixation from increased cropping intensity is with soil water stored during fallow periods. A recent meta-analysis (Liu et al. 2022) found that in regions with less than 600 mm of annual rainfall, increasing cropping intensity offers minimal benefits because of limited water availability. However, increased cropping intensity can significantly enhance SOC in regions of higher rainfall, and factors such as crop duration, type and species diversity become crucial.
Extending the life-cycle duration of annual crops by sowing them earlier or using cultivars with delayed development (e.g. increased vernalisation or photoperiod sensitivity) consistently increases biomass production (Gomez-Macpherson and Richards 1995; Cann et al. 2020b) through greater water use, reduced evaporative losses and increased transpiration efficiency (Kemanian et al. 2005; Flohr et al. 2020). Likewise, increasing plant population density or narrowing row spacing reduces evaporative losses while improving competitive ability against weeds (Lemerle et al. 2004; Borger et al. 2016).
Crop diversity helps reduce plant growth-limiting factors, such as root diseases and N deficiency when legumes are used (Dias et al. 2015; Peralta et al. 2018; Weisberger et al. 2019). Species and cultivars with functional traits (e.g. catch crops to reduce nitrate leaching, deep-rooting varieties to access water and nutrients at depth) can potentially balance higher long-term profitability with increased C inputs (Volsi et al. 2022). In addition to enhancing total biomass, crop species diversity modifies residue input quality, reducing SOC turnover (McDaniel et al. 2014; Chatterjee et al. 2016) and potentially enhancing microbial biomass and nutrient cycling (McDaniel et al. 2014; Tiemann et al. 2015).
Cover crops are grown as a single or mixed species to provide ground cover, capture unused resources (e.g. N), and improve soil health (Florence and McGuire 2020). Unlike cash crops, they are not harvested but are typically sown between or in place of cash crops, extending the duration of living plants in the soil, which can potentially boost system NPP and C inputs (Garland et al. 2021). Various cover crop types and mixtures have been tested, offering different benefits depending on agronomic needs, such as N fixation, weed suppression, disease management, and subsoil improvement. The major trade-off for soil C benefits often lies between selecting high-biomass species (e.g. cereals, C4 plants) and those that provide higher-quality residues or additional N inputs (e.g. legumes).
Longitudinal studies in the USA have shown that cover crops can increase permanganate-oxidisable C and SOC (Wood and Bowman 2021), with a meta-analysis across various environments consistently demonstrating positive effects on soil C (mean ~300 kg C ha−1 year−1; Bolinder et al. 2020). However, in dryland systems that rely on stored water for cash crop growth, using some moisture to support cover crops can reduce the growth, yield, and subsequent C input from the following cash crop (Garba et al. 2022). Therefore, it is crucial to consider the water requirements of the cash crop, the capacity to replenish the soil water during the fallow period, and the likelihood of excess water availability when considering cover crops in drier systems (Wunsch et al. 2017; Garba et al. 2022; Rose et al. 2022). Effective management of cover crop water use, including appropriate termination strategies, can mitigate some of these concerns (Whish et al. 2009; Cong et al. 2015; Wunsch et al. 2017; Rose et al. 2022). Few studies have examined the long-term effects of cover cropping on soil C accumulation in dryland arable systems, although short-term studies have shown an increase in biologically active C fractions and microbial activity (Williams et al. 2022). The potential benefits of below-ground biomass production for C accumulation in dryland systems also require further exploration. For example, selecting cover crop species on the basis of root architecture to maximise C inputs or using tuber-forming crops (e.g. tillage radish) to improve soil infiltration and conditions for subsequent cash crops could be beneficial. The advantages of cover cropping in dryland systems and the processes underlying these benefits warrant further investigation.
Unlike cover crops, which are grown separately from cash crops, intercropping involves planting two or more crops together in the same space and time (Stratton et al. 2022). Intercropping aims to increase crop yields per unit area by improving resource use efficiency (e.g. sunlight, water, nutrients), spreading production risks across different crops, and reducing disease pressure. While the biological processes of intercropping share similarities with those of cover cropping, the end goals differ, with intercropping focusing on maximising harvestable biomass rather than improving soil health.
Growing mixtures of crops with different functional traits can increase total system biomass production (Pappa et al. 2012; Isbell et al. 2017; Qui et al. 2023), with the extent of ‘overyielding’ (i.e. greater yield for a species when grown with others compared to monoculture) depending on plant species. Dryland regions may particularly benefit from intercropping (Li et al. 2014; Zhu et al. 2023) when one of the crops enhances nutrient availability, such as increased N inputs from legumes or greater P availability from species with P-solubilising traits. Indeed, a recent meta-analysis of intercropping systems in Africa and South Asia found that grain legume–cereal intercropping systems, on average, significantly increased SOC via increased biomass production, and that this effect was greatest in soils with an initial SOC of <1% and clay content of >32% (Kuyah et al. 2023).
However, positive effects of intercropping on SOC are not uniform. Parvin et al. (2025) demonstrated both increased available N and water in a wheat–vetch system, but this did not result in increased wheat yield or grain quality, possibly owing to sufficient in-season rainfall in a non-N limiting situation in the year of study. In a 4-year study of wheat–soybean and maize–soybean intercropping in Gansu Province, China, Wang et al. (2025) were not able to observe changes in surface soil SOC, but did observe an increase in partitioning of C by 5.6% to the more stable mineral-associated pool in intercropping treatments. Conversely, Wang et al. (2024) observed a decrease in subsoil C by 5.3% after 10 years of intercropping, despite increased yield and above-ground biomass production. In low-rainfall regions and especially in lower decile years, intercropping may offer limited benefits, and the different crop species may compete for scarce water resources (Duivenbooden et al. 2000).
Integrating livestock or phases of pastures into farming systems offers the following two key pathways to build or maintain SOM compared with grain-only systems: (1) increasing both above- and below-ground C inputs, and (2) improving the quality of C available to the microbial community, potentially enhancing the rate of C fixation (Richardson et al. 2019). Perennial grass-based pastures are particularly effective, with much higher above- and below-ground biomass production than with annual pasture species (Koteen et al. 2011). Numerous studies have shown that periods of perennial pastures in crop rotations can increase soil C by ~500 kg C ha year−1, although stocks tend to oscillate between accrual under pasture and then draw-down in the subsequent pasture phase (Díaz-Zorita et al. 2002), perhaps with a longer-term upward trend across multiple phases. Achieving high rates of C accumulation depends on the productivity of grass-based pastures, which are often fertilised or include legumes to provide an N source (Dalal et al. 2018). In contrast, low-productivity pastures offer smaller or negligible benefits to soil C. Incorporating legumes into pastures in appropriate ratios with grasses results in organic material with a lower C:N ratio, which can be used more effectively by microbes than grass residues alone, which typically immobilise available N and reduce C cycling.
Annual species typically have less developed root systems than do perennial species, owing to their short-lived growth, and are, therefore, less likely to build soil C than are perennial pastures. However, early sown annual dual-purpose crops (e.g. forages) can increase total above-ground biomass production by 2–3 Mg dry matter ha−1 compared with grain-only crops sown in traditional windows. Dual-purpose crops are common in dryland farming systems worldwide, where their resilience to drought risks is a motivating factor for adoption (e.g. low yield risks mitigated by the ability to graze livestock; Ground Cover 2023). These crops are valuable in both developed and developing countries (MacKown and Carver 2005; Arzadún et al. 2006; Kirkegaard et al. 2008; Tian et al. 2012; Giunta et al. 2014; Melesse et al. 2021).
Although grazing removes some above-ground biomass, including livestock in pasture phases does not necessarily reduce net C or N balances. In favourable conditions, grazed plants may increase root rhizodeposition, enhancing net C flow to the soil (Stanley et al. 2024). Root biomass often decreases in the short-term following grazing, but with sufficient rest, it can recover after defoliation (Dawson et al. 2000). Under optimal grazing intensities and frequencies, total biomass productivity can be maintained (Deng et al. 2023). However, overgrazing can reduce total biomass growth and the amount of organic material available for SOM incorporation. Conversely, very low grazing intensities may not stimulate as much biomass regrowth and/or root contributions to soil C. In dryland systems with variable rainfall, grazing intensities must be adjusted regularly to avoid over- or under-grazing.
Of the biomass consumed by livestock, approximately 40% is returned to the soil as manure, often in a form that is a more effective source of C for soil microbes. For example, some of the more resistant C forms in pasture residues (e.g. cellulose) are decomposed by cellulolytic microbes in the digestive tracts of grazing animals, lowering the C:N ratio and improving the availability of this C to soil microbes and for incorporation into SOM (Kristensen et al. 2022). The higher CUE of grazed and excreted organic material increases soil microbial biomass and the biologically active C fraction (Franzluebbers and Stuedemann 2015). In grazed systems, only small amounts of N are removed in animal products, with most consumed N being returned to the soil in manure and urine (Sprague et al. 2021).
Importing organic matter
In dryland regions where NPP and existing management practices are insufficient to increase SOM concentrations or where additional increases are desired, organic amendments (OAs) may be introduced into the farming system, although they are usually applied with the primary aim of exploiting their nutrient content. Because OA application directly adds organic C and nutrients, it typically improves SOC content and nutrient cycling (Dorado et al. 2003; Melero et al. 2007; Diacono and Montemurro 2010; Li et al. 2020; Soto et al. 2021). Various OAs have also improved dryland yields, likely via direct nutrition effects, although repeated or high-rate applications are often necessary to maintain efficacy (Tejada and Gonzalez 2007; Mlih et al. 2016; Calderon et al. 2018; Ma et al. 2022).
Types of organic amendment
A wide variety of OAs is being applied to agricultural soils, including (1) composted and uncomposted animal manures and litters, (2) processed urban organic resources (e.g. composts, biosolids, and biodigester residues), and (3) biochars, humates, and other novel amendments. The composition of these amendments can vary significantly among and within types (Fig. 3; Farrell et al. 2015a, 2015b; Abbott et al. 2018; Baldock et al. 2021). The availability of specific amendments for land application depends on the proximity to a suitable source and regulations governing their use. Although the C contained in OAs increases SOC on application, this increase does not necessarily represent long-term sequestration unless there is (1) a subsequent increase in SOC beyond the amount added because of improved crop growth and increased plant C inputs, or (2) additional stabilisation of the applied C. Given the considerable distances between major sources of OAs and many dryland farming regions (e.g. in large parts of Australia, Canada, and the USA), the costs and emissions associated with transport present significant barriers to widespread adoption.
Distribution of chemical attributes of 121 individual OAs sourced across Australia. Alkyl:O-alkyl and aromatic:aliphatic ratios are derived from 13C cross polarisation/magic angle spinning (CP/MAS) nuclear magnetic resonance (NMR) analysis and are metrics of decomposition and chemical stability respectively. Data from Baldock et al. (2021) and Farrell et al. (2015a, 2015b).
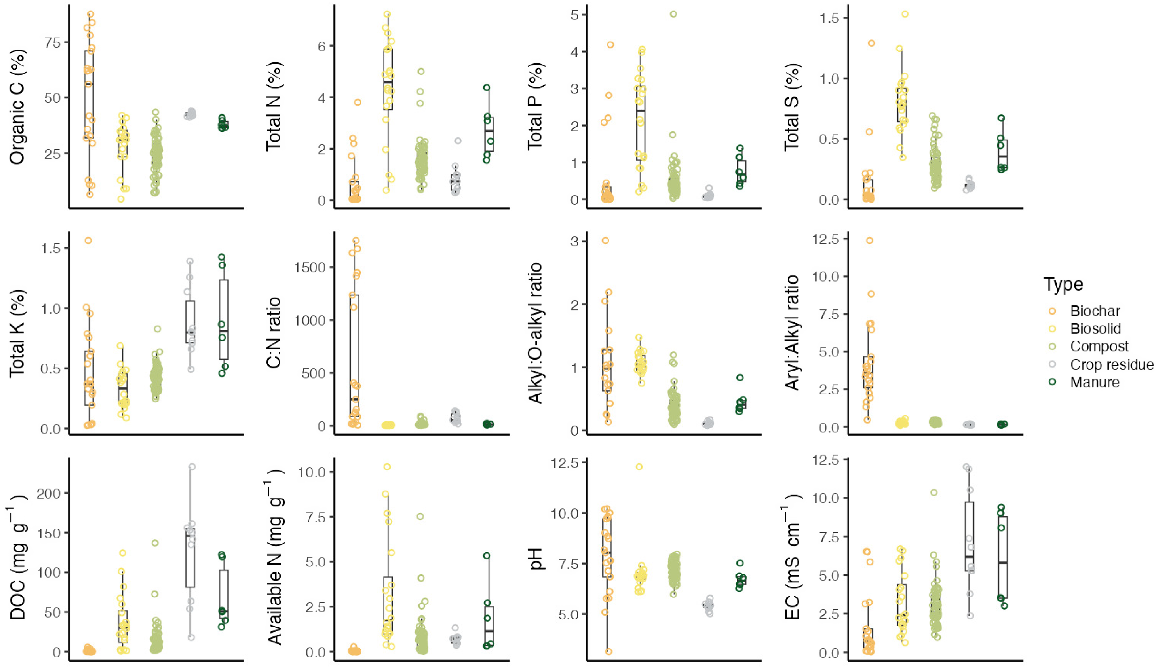
Manures and residues are among the most nutrient-rich OAs. Because of their typically low C:nutrient ratio (Fig. 3), manures may stabilise C more efficiently than do residues (Coonan et al. 2020). Similarly, biosolids may contain substantial nutrients, although their C and nutrient concentrations can vary significantly depending on their source and processing methods (Brown et al. 2020). Composts also exhibit a wide range of properties (Fig. 3) and may have high C:nutrient ratios that could initially immobilise nutrients after soil application. Depending on the source and season, composts often contain more woody lignin-rich debris, which may be particularly beneficial for improving soil porosity and structural properties (Agnew and Leonard 2003). However, repeated applications of composts, manures, and other more labile amendments are often required to achieve lasting changes in soil C and structural properties (Sodhi et al. 2009) The last major class of OAs is biochar, a recalcitrant, charcoal-like product created through the pyrolysis of organic feedstock in a low-oxygen environment (Jeffery et al. 2011). Biochars are typically low in nutrients and resistant to decomposition. Due to their high C density and lower application rate than other OAs, biochars may be transported further from production areas. However, biochars exhibit significant variability in their attributes are based on production conditions and feedstock composition, making it difficult to generalise their function, particularly given their interactions with soil characteristics (Macdonald et al. 2014).
Methods of application
Traditionally, OAs have been surface applied using bulk spreading machinery. Conventional tillage systems incorporate these surface applications into the soil through tillage. However, with the widespread shift to NT systems, which favour minimal soil disturbance and direct seeding, the incorporation of surface-spread OAs relies on leaching, bioturbation by soil macrofauna, and partial incorporation by seeder tines to move the material below the surface. Without incorporation, the translocation of OA-C from surface layers to moderate depths (e.g. 5–15 cm) can take significant periods of time (over 25 years), particularly in dryland systems where lower rainfall and reduced microbial activity limit incorporation (Nicoloso et al. 2018). Occasional strategic tillage may expedite OA-C addition to depth, with only minor and temporary adverse effects from soil disturbance (Conyers et al. 2019). As an alternative in NT systems, applying small amounts (<1 Mg ha−1) of suitable OAs (e.g. biochar; Farrell et al. 2012) via a second line of an air seeder directly into the seedbed places the amendment beneath the roots of the germinating seed. Although such methods do not add large quantities of C to the soil, they target areas where the effect is likely to be most immediate.
Deep placement of OAs such as manures or pelletised plant residues with and without inorganic amendments (e.g. nutrients, gypsum, lime) has been explored in some dryland cropping systems with mixed results (Celestina et al. 2019). Organic amendments may also be applied directly to the subsoil through intensive processes, including ripping through subsoil layers with deep tines to slot OAs down the rip-line or via surface application followed by deep or full-inversion tillage. These strategies are typically employed to address subsoil constraints to plant productivity (Unkovich et al. 2020; Sale et al. 2021), although their effects on crop yield are often confounded by simultaneous amelioration of constraints and nutrient addition (Celestina et al. 2019). Balanced comparisons of surface versus subsurface applications frequently show no significant impact of placement depth or interaction with amendment type (Celestina et al. 2018).
Potential benefits of increasing SOM
Nutrient and water availability
Increasing SOM generally enhances nutrient availability for plant growth (Fig. 4; Seiter and Horwath 2004; Sarker et al. 2018; Barton et al. 2022). The most significant yield benefits of higher SOM are often observed under conditions of low nutrient fertilisation, high nutrient demand, or when SOM can supply nutrients in soil profile positions that are difficult to reach with fertilisers (Johnston et al. 2009). Additionally, because SOM significantly contributes to cation exchange, increasing SOM improves the supply of positively charged nutrients (e.g. calcium, potassium; Tiessen et al. 1994).
Conceptual diagram illustrating the agronomic, economic and environmental benefits (left) and costs (right) of increasing soil organic matter (SOM). Cumulative benefits significantly outweigh cumulative costs when at low SOM contents but there are decreasing marginal benefits and increasing marginal costs as SOM contents increase beyond optimum range (x-axis). For example, the benefits of participating in carbon markets are highest when starting with low SOM content, but decrease as initial SOM content increases. Conversely, input costs of cash crops (e.g. fertiliser, water) rise with an increasing SOM content because of higher biomass production. Again, improvements in water availability as a result of either increasing infiltration in clayey soils with limited structure due to low SOM, or increasing retention in sandy soils with low SOM would be expected to tail off as SOM content increases along the x-axis. Note: what constitutes ‘low’ or ‘high’ SOM content varies among different soils and systems.
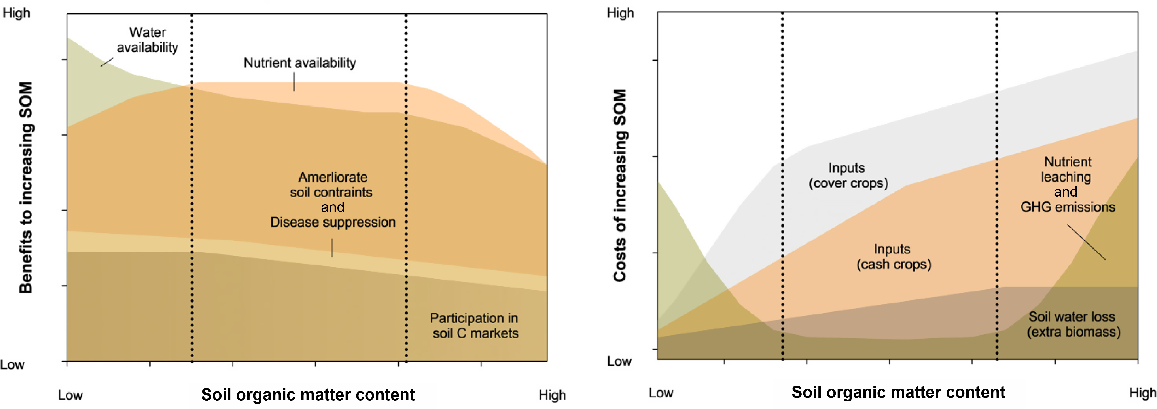
Soil organic matter also improves soil structure, aggregate stability, and bulk density (Hudson 1994; Krull et al. 2004), enhancing water-holding capacity. The extent to which SOM increases water retention depends on other soil characteristics. The greatest benefits occur in coarser-textured soils that do not retain significant amounts of water (Hudson 1994; Murphy 2015). Enhanced nutrient and water availability are the two most critical benefits of increasing SOM in dryland soils, because these factors typically limit primary production (Lu et al. 2018; Wang et al. 2022; Brown and Collins 2023). Studies attempting to partition the relative effects of enhanced nutrient cycling and water availability on crop yield responses to SOM have concluded that nutrient cycling is the dominant effect (Moore and Hunt 2010; Minasny and McBratney 2018; Oldfield et al. 2020).
Ameliorate soil constraints
Soil organic matter is especially beneficial for addressing soil structural deterioration and dispersive sodic soils, which reduce agricultural productivity, especially in dryland farming regions (Fig. 4; Rengasamy and Olsson 1991). Soil organic matter functions as a binding agent, promoting the formation of stable aggregates, which enhances both macro- and micro-porosity (Tisdall and Oades 1982), resulting in improved water infiltration, reduced surface runoff, and better conditions for root growth (Sale et al. 2021). Improved soil structure with increased SOM contributes to yield increases in some crops through more rapid root development, enhancing water and nutrient uptake, benefits not typically seen with increased nutrient applications alone (Johnston et al. 2009). Further, since soil constraints often appear in subsoil horizons, increasing SOM in these layers is necessary to alleviate constraints, meaning SOC increases must extend beyond topsoil layers; 0–10 cm). Fortunately, subsoil layers show potential for increased C storage due to their typically higher clay mineral content than that of topsoils (Hobley et al. 2015), which may provide the added benefit of more stable storage of C at depth (Feng et al. 2020).
Resilience and sustainability
The stability and sustainability of dryland cropping systems depends on resistance (the ability to withstand disturbance) and resilience (the ability to recover after disturbance). Emerging evidence links SOM with increased drought resistance in corn yields through direct and indirect mechanisms (Kane et al. 2021). Enhancing SOM in dryland agricultural soils improves soil resilience by enhancing soil structure and establishing a larger, more diverse, resilient microbial community. This community has a complex taxonomic and functional composition and stable microbial networks. It exhibits functional plasticity, modulating metabolic profiles to withstand stresses and recover efficiently (Shu et al. 2021; Dacal et al. 2022), and enables the soil system to maintain ecosystem functions (e.g. nutrient availability, C cycling and sequestration, plant health via biological suppression) despite disturbances (Gupta et al. 2011; Shade et al. 2012; Griffiths and Philippot 2013; Todman et al. 2016; Dynarski et al. 2020). Since dryland soils often face repeated disturbances (e.g. tillage, short-term fallows) and environmental (e.g. water-stress conditions) stresses that can lead to discrete soil environmental or habitat changes, SOM contents that support a large, diverse and functionally redundant microbial community and a diverse soil food web are beneficial for maintaining ecosystem services in arable soil (De Vries et al. 2013; Gupta et al. 2019; Garland et al. 2021; Steven et al. 2021; Shi et al. 2024).
Additionally, SOM influences soil macroporosity affecting the impact of wetting and drying events (Bonetti et al. 2017). In dryland systems exposed to typically dry but increasingly extreme weather events, buffering between large moisture fluctuations is crucial for soil resilience.
Participation in C markets
Carbon markets are increasingly becoming a primary tool for countries to meet their emissions reduction commitments (UNFCCC 2024). These markets set standards and governance requirements for C projects to create and trade C credits by reducing, storing, or avoiding C emissions. By building SOC, the land sector can supply C credits to the market or keep them and insert them into their enterprise C account (Ebersold et al. 2023).
Since C stocks in drylands are generally low on a per-area basis but cover large expanses (Hanan et al. 2021), even minor changes can result in appreciable SOC stock increases, generating C credits (Fig. 4). The growing emphasis on environmental, social and governance (ESG) commitments to net zero targets leads many hard-to-abate industries to seek C offsets in the land sector, presenting an opportunity for dryland farmers to improve their soil fertility and function while financially benefiting from SOC-increasing strategies. However, socio-political environments in many dryland regions may hinder the successful implementation of C markets. For example, crop residue removal for animal feed and excessive grazing, being practices often necessary for farmers’ livelihoods (Lal 2005; Lahmar et al. 2012), contribute to SOC depletion. There are also trade-offs between the need to retain stubble for its beneficial effects on wider soil health (and SOC stocks specifically) versus its value for the production of biofuels (Arsic et al. 2022), which are increasingly sought after to reduce reliance on fossil fuels in hard to abate sectors such as aviation. Nonetheless, modelling does suggest that in the southern and western cropping regions of Australia, it may be possible to remove up to 50–75% of residues without affecting SOC stocks (Zhao et al. 2015).
The global agri-food market, supply chain demands, and regulatory environments surrounding commodity emissions and broader sustainability credentials are rapidly evolving. As mandatory ESG reporting increases, farmers must decide between selling C offsets or in-setting them to improve their production footprint. A long-term low-emissions strategy that permits practices with high upfront emission costs (e.g. liming, amelioration) may make it more valuable for growers to retain their soil C credits to manage enterprise credentials and market access.
Potential costs of increasing SOM
The positive impacts of increasing SOM in arable dryland systems are widely recognised (Fig. 4). However, important economic and environmental considerations require careful discussion to highlight potential risks. Notably, if strategies to increase SOM are misapplied, they may incur economic or environmental costs. For instance, increasing N fertilisation to boost biomass production could result in excessive fertiliser use if underlying production constraints are not addressed, leading to fertiliser loss (Simpson et al. 2011; Vanlauwe et al. 2015; Abbott et al. 2018). Similarly, deep ripping to incorporate C inputs deeper in the soil profile (Davis et al. 2019) could lead to increased mineralisation of previously protected SOM in upper soil layers (Balesdent et al. 2000; Fiedler et al. 2016). This section discusses the potential risks of increasing SOM with correctly applied interventions.
Economic
The functions of SOM are crucial to agricultural productivity, making the concept of an optimum SOM range sensible from an economic perspective (Fig. 4). Especially in dryland systems, suboptimal SOM concentrations can negatively affect plant production, offering a strong economic incentive for improvement. However, the costs of increasing SOM may or may not always be balanced by the potential economic benefits from improved productivity or payments for sequestered SOC. At a global scale, the link between increased SOM and yield is variable, and the climate mitigation potential of increased SOM is modest (Moinet et al. 2022). Responses are highly context-dependent, influenced by specific pedo-climatic conditions, initial SOC stocks, and interactions and trade-offs between management practices and other yield-limiting processes. Consequently, the rationale and strategies to increase SOM must be evaluated against other economic, environmental, and practical factors influencing farm enterprise decision-making (Richardson et al. 2019).
In dryland agriculture, excessive biomass production can rapidly deplete stored water, causing significant stress during critical reproductive stages of grain crops. This phenomenon, known as ‘haying-off’, can result in economic losses because of reduced yield and grain quality (Fig. 4; Van Herwaarden et al. 1998). Breeding strategies and agronomic recommendations have focused on increasing the harvest index to minimise unproductive biomass production (Hunt et al. 2019). Where biotic or abiotic constraints are alleviated, allowing better use of available water and nutrients, higher-yielding crops may leave more biomass. However, optimal, rather than maximum biomass accumulation is advisable when water supply and timing are unpredictable.
Establishing pasture phases and adding or increasing livestock are long-term strategic business decisions. The profitability of mixed farming systems, including cropping and livestock, varies by context (e.g. crop selection, access to technology, water availability, farm size; Bowman and Zilberman 2013). Although systems that include pasture phases typically have higher long-term profitability in many regions (Sánchez et al. 2022), converting cropped land to pasture requires significant investment in infrastructure and upskilling (Krall and Schuman 1996), posing a substantial short-term opportunity cost. This cost must be weighed against long-term benefits such as improved soil fertility, weed control, enterprise diversity (e.g. high livestock prices), or C payments. For example, mixed farms may have similar gross margins to grain-only farms when the value of biologically fixed N from pasture phases is considered (Angus and Peoples 2012). However, the temporary rundown of stored SOM during crop phases may jeopardise C payment eligibility.
In regions with wet and mild winters and hot and dry summers (e.g. southern Australia, Mediterranean Europe), cover crops may have limited benefits and could compete for resources needed by subsequent cash crops (Fig. 4; Rose et al. 2022). Tactical opportunities for cover crops may arise in certain conditions, such as (1) after a failed winter crop or prolonged drought exposes soil to erosion, (2) when herbicide-resistant weeds require management, (3) when full soil profiles at harvest and wet seasonal outlooks allow cover crop growth and termination without pre-emptive competition, and (4) on sandy soils in high winter rainfall areas with minimal reliance on summer rainfall for cash crop yields. In contrast, temperate climates with more evenly distributed rainfall make cover crops more effective for providing ground cover, decreasing nitrate leaching and increasing SOM (Riviere et al. 2022).
No-till and stubble-retained management are primarily adopted for their economic benefits, such as timely crop sowing and reduced fuel, labour, and machinery costs (Llewellyn and Ouzman 2019). However, anticipated improvements in soil C have not been widely realised (Powlson et al. 2014; Kopittke et al. 2016). Long-term NT approaches may also face challenges such as herbicide-resistant weeds, disease, subsurface compaction and acidity, and non-wetting soils, leading to a more pragmatic approach that includes strategic tillage and stubble management (Conyers et al. 2019; Kirkegaard et al. 2020).
In systems where cereal stubble is retained, nutrient availability (e.g. N, P, S) is often a limiting factor for C sequestration and SOM increases (Lemke et al. 2010; Kirkby et al. 2016; Kirkegaard et al. 2023). Whereas supplementary nutrients may increase total SOM, the stability of stored C or aggregates may decrease (Fonte et al. 2009; Tang et al. 2023). Economic analyses suggest a positive return on investment in nutrient management strategies over time (Kirkegaard et al. 2023). Since N limitation remains the major cause of crop yield gaps, maintaining nutrient balance to optimise yield and improve SOM will come at little additional cost (Fig. 4; Meier et al. 2021). In developing countries, short- and long-term trade-offs between using crop residues for soil cover and other purposes (e.g. livestock feed) must be considered (Giller et al. 2015). Removing residues for other uses increases short-term income and/or capital and manages the risk of poor crop production and/or prices (Erenstein et al. 2015).
The primary economic cost of importing OAs to increase SOC is the expense of transporting bulky materials over long distances. In many dryland areas, the limited availability of OAs and their low nutrient concentrations relative to synthetic fertilisers make them expensive to ship. Variability in OAs, within and among categories (Fig. 3), complicates their precision management, and the potential for nutrient imbalances poses risks to crop and pasture performance and the environment (Möller 2018). Contamination of OAs with potentially toxic elements (PTEs), persistent organic pollutants, microplastics, pathogens, and antimicrobial resistance genes is a concern, particularly for biosolids from human waste streams (Petrie et al. 2015; Tiedje et al. 2019). Standards and regulations exist to manage PTEs, but emerging contaminants may pose long-term risks.
Perhaps perversely, importing C via OAs may also complicate eligibility for SOC payment programs. For example, the Australian soil carbon methodology (DCCEEW 2021) excludes C in external inputs from counting towards C credits for SOC, because it is already considered to be ‘removed’ from the atmosphere. Increased C inputs from improved crop and pasture performance as a result of management practice changes that include external C applications can be credited. However, detection of these inputs (which is already difficult; Stanley et al. 2023) against additional non-creditable C from OAs that may also have increased SOC heterogeneity, makes the task more challenging.
Environmental
The environmental costs of increasing SOM generally relate to enhanced microbial activity and nutrient cycling (Fig. 4), which are also essential for increased nutrient availability (Coonan et al. 2020). Key concerns include the loss of nutrients through gaseous emissions and leaching (Abbott et al. 2018; Qiu et al. 2020; Xenopoulos et al. 2021). Generally, the environmental risks of increasing SOM in dryland systems are considered low.
Large CO2 fluxes are primarily associated with the decomposition of ‘younger’ OM, whereas older OM reserves tend to resist mineralisation (Su et al. 2023). This resistance is particularly evident when newer C sources are incorporated into the DOM fraction (Boyer and Groffman 1996; Chantigny 2003; Qiu et al. 2015). Management practices that increase labile soil C also influence N2O emissions from cropped soils (e.g. Six et al. 2004; Helgason et al. 2005; Liu et al. 2014), likely because of the stimulating effect of bioavailable C on microbial SOM mineralisation and a concomitant increase in mineral N availability for nitrifiers and denitrifiers (Qiu et al. 2020). The highest N2O fluxes occur when increased SOM creates a temporal mismatch between mineral N availability and crop N demand, and when environmental conditions favour microbial processes responsible for soil N2O emissions (Barton et al. 2022). This is most notable when increased SOM decreases soil oxygen availability due to enhanced soil respiration or greater soil water-holding capacity (Groffman et al. 1988).
The rise in CO2 and N2O emissions following the addition of rapidly solubilised C inputs has implications for agricultural systems striving to increase SOM contents. For example, a substantial proportion of the recently applied C (e.g. crop residues) might be rapidly respired as CO2, potentially stimulating N2O emissions from subsequent N-fertiliser applications and/or SOM-derived N, especially if the residues are immature (i.e. N supply in residues exceeds immediate microbial needs; Olesen et al. 2023). Agricultural practices that enhance the stability of C inputs and/or slow crop residue decomposition may be necessary to reduce GHG (e.g. CO2, N2O) emission potential. These could include deeper incorporation of crop residues into the soil profile, where (1) DOM retention is higher and physical adsorption converts DOM to more recalcitrant forms (Angers and Eriksen-Hamel 2008; Kalbitz and Kaiser 2008), and (2) reduced microbial activity slows the decomposition rates of organic C inputs (Nicolardot et al. 2007; Naylor et al. 2022).
Dissolved organic matter is highly susceptible to leaching, where it is translocated down the soil profile and potentially into groundwater or nearby water bodies (Aitkenhead and McDowell 2000; Van der Berg et al. 2012; Xenopoulos et al. 2021). Excessive DOM contents in water systems pose a significant environmental risk, because high dissolved organic C concentrations can affect water potability, increase acidity, affect aquatic organisms, and potentially disrupt terrestrial C stores (Tipping et al. 1988; Freeman et al. 2001; Chapman et al. 2010). Similarly, nutrients within DOM (e.g. N and P) can threaten natural ecosystems when present at high concentrations, leading to issues such as eutrophication, hypoxic conditions, nutrient imbalances, and reduced light availability; Azevedo et al. 2015; Lu et al. 2023). In addition to DOM-associated losses, increasing SOM can increase direct N leaching losses (Murphy et al. 2000; Macdonald et al. 2016), and thus rate, timing, and placement of N inputs should still be considered carefully. Higher SOM content enhances N cycling in soils (Fisk et al. 2015), increasing the formation of negatively charged and highly mobile N species (i.e. nitrites, nitrates). These species have a high leaching risk, particularly in soils with structures that promote the preferential flow of dissolved N species, bypassing soil and plant processes that would otherwise retain N (Anderson et al. 1998; Barton et al. 2005).
In dryland systems, where rainfall is generally low and extended fallow periods exacerbate water loss from soils, the risk of leaching losses is likely limited to relatively rare intense precipitation events (Sigler et al. 2020), with the frequency of such events potentially increasing as a result of climate warming (Donat et al. 2016). Furthermore, most dryland agricultural soils are unlikely to have SOM contents that would pose a significant risk of excessive DOM loads reaching aquatic ecosystems (Smreczak and Ukalska-Jaruga 2021), even if substantial SOM increases were achieved.
Monitoring soil organic carbon stock change
Monitoring campaigns are essential for estimating changes in SOC stocks, assessing the effectiveness of soil C sequestration strategies, and guiding sustainable land management practices. Various methodological and technological approaches, such as direct analytical laboratory methods, proximally and remotely sensed techniques, and process-based models, are employed to measure and estimate SOC (Motavalli et al. 1994; Chatterjee et al. 2009). The accuracy of these methods decreases when moving from direct measurement to remote sensing, while the spatial representativeness increases (Manzoni and Porporato 2009). Accurate quantification requires precise measurement of bulk density (BD) and gravel content (>2 mm), as well as a representative sampling strategy capable of identifying expected changes (Stanley et al. 2023).
Determining soil organic carbon quantity and quality
Wet chemical oxidation, dry combustion, and ashing (e.g. loss on ignition) methods are used to measure SOC directly. Wet chemical oxidation approaches (Walkley and Black 1934) have been the most widely used, but may not fully quantify more recalcitrant C forms (Hardy and Dufey 2017). More recently, due to their reliability, dry combustion methods using elemental C analysers, such as the Dumas method, have become the standard for SOC measurements (Baldock et al. 2013). However, this approach measures total soil C and requires pre-treatment of samples to remove inorganic C, which is time-consuming and expensive, and can result in considerable error in soils with high inorganic C and low SOC concentrations (Schmidt et al. 2012).
The development of spectroscopic techniques has enabled rapid and reliable measurement of both total SOC and functionally distinct SOC fractions (Brown et al. 2006; Viscarra Rossel et al. 2006; Janik et al. 2007; Bellon-Maurel and McBratney 2011). Techniques include cross polarisation/magic angle spinning (CP/MAS) 13C nuclear magnetic resonance (NMR, Prietzel et al. 2018; Mustafa et al. 2022) for chemical characterisation, mid-infrared (MIR) and near-infrared (NIR, Jia et al. 2017; Jaconi et al. 2019; Vohland et al. 2022) for assessing functional groups and chemometric modelling, and synchrotron-based C near-edge X-ray adsorption fine structure (NEXAFS, Prietzel et al. 2018) for mostly qualitative investigations. In particular, infrared spectroscopy, such as MIR and NIR, are commonly used to quantify SOC through chemometric analysis (Viscarra Rossel et al. 2006; Bellon-Maurel and McBratney 2011), although they still require a calibration set of spectra that is representative of the soils studied.
Remote sensing uses earth observational (EO) datasets, such as satellite images and images taken by aeroplanes or drones, to derive proxy values of SOC stocks. Even though EO approaches can only estimate SOC stocks, they are time-efficient, non-destructive, and cover large areas, making them useful for monitoring changes over time. On-ground measurements of SOC stocks and remotely sensed EO datasets are used to derive empirical models across the landscape by using reflectance bands and other indices derived from a range of environmental covariates (Viscarra Rossel et al. 2014). With improved spatial (≤30 m), temporal (≤week revisit time), spectral (multi-spectral vs hyperspectral) and radiometric resolutions, next-generation sensor technologies are attracting growing interest in applications at the farm scale to monitor SOC stocks.
Calculating SOC stocks
Soil BD is required for a defined sampling depth, including correction for gravel content. However, its variability over time and space makes it challenging to detect differences in SOC stocks among sites (Kirkby et al. 2021). The BD measurement method depends on factors such as the purpose, time available, required accuracy and precision, the need for repeated measurements, cost, staff technical capabilities, and equipment availability (Coughlan et al. 2002). The intact core-based method is most commonly used for SOC stock calculations. However, there is increasing interest in indirect BD measurement methods (e.g. active gamma-ray attenuation sensors) for rapid and cost-effective estimation, including capturing spatial variability (Lobsey and Viscarra Rossel 2016). When detecting temporal changes in SOC stock, calculations require corrections for BD changes relative to baseline sampling using equivalent soil mass, which accounts for changes in BD that might otherwise exaggerate apparent changes in SOC stock (Don et al. 2010). This correction is especially relevant in agricultural systems where tillage or traffic can modify the BD of surface soil layers (Wendt and Hauser 2013).
Selecting a soil sampling strategy that appropriately captures spatiotemporal variation in SOC stocks is also critical. Most campaigns use one of two approaches, namely, design-based or model-based (De Gruijter et al. 2006). Model-based sampling schemes are more appropriate for local estimates (e.g. digital mapping of SOC stocks across the landscape at a defined grid interval), whereas design-based approaches are more suitable for global estimates of a given soil attribute (e.g. an estimate of SOC changes across a defined unit, such as soil-landscape units; Singh et al. 2012; Karunaratne et al. 2014). The extent, timeframe, and depth of the sampling campaign depend on the study aim. For example, Australia’s soil carbon research program collected 10 cores from a 25 × 25 m area (Baldock et al. 2013), whereas the current Australian soil carbon methodology mandate a minimum of three samples per stratum, with a minimum of three strata per area being tested (DCCEEW 2021).
Estimating and forecasting SOC stocks through space and time
Due to the expense of direct and even proximal or remote measurements across large areas, predictive modelling approaches provide valuable information that fills data gaps. Empirical regression models can estimate and forecast changes (e.g. annual) in SOC by using long-term ground measurements. This data-driven approach is cost- and time-effective, but requires a high volume of data for model training (Paustian et al. 2019). In contrast, process-based models such as RothC (Coleman and Jenkinson 1996), CENTURY (Parton et al. 1988), and APSIM (Verburg et al. 2025) use conceptual pools, with mass fluxes to and from different pools controlled by environmental conditions (e.g. soil type, temperature, water availability) and land management. Because of their cause–effect approach, these models can be applied across different spatial and time scales. However, process-based models require many parameters and long-term measurements for appropriate calibration and validation.
Merging process-based and empirical models, known as ‘surrogate’ models, can provide more information on soil C drivers and help assess areas of uncertainty (Luo et al. 2013, 2019). Surrogate models are data-driven models that use available variables to replicate the output of process-based models. This approach requires a limited amount of information, making it computation- and time-efficient compared with process-based models, and it can be used in regional- and national-scale applications (Luo et al. 2013, 2019).
Conclusions
Opportunities exist to limit SOC decline or maintain/increase stocks through improved management practices in farming systems across all dryland regions and contexts. Nonetheless, achieving ambitious targets such as ‘4/1000’ appears challenging. Most systems have an ‘optimum’ SOM content, where the benefits outweigh the costs of the management practice (Fig. 4). Beyond this point, continued investment in increasing SOM yields diminishes returns. This ‘optimum’ varies depending on soil type, region, and other factors, but is typically lower in dryland areas than in regions with higher rainfall and more fertile soil.
Identifying suitable management practices to increase SOM is particularly critical in drylands, where constraints and challenges vary widely and can significantly influence the success of implemented changes. Even if an end goal of increased SOM is achieved, it is possible that economic success may not be realised either through improved agronomic performance and/or resilience, or via monetisation of SOC via C credit schemes. This is because the magnitude of change in water-limited systems is likely to be relatively low, and the potential vulnerability of reversal owing to successive years of drought may overcome improved system resilience that is associated with higher SOM. Monitoring and verifying SOC changes can be complex, especially when seeking recognition for C credits. Pragmatic monitoring strategies will likely depend on a combination of conventional, proximal, and remote-sensing methods, which must be carefully considered to assess the effectiveness of management interventions. Despite these challenges, given the vast land area managed for cropping in dryland regions globally, small gains across wide areas may still amount to a significant increase on terrestrial C stocks at the planetary scale.
Of the interventions available to build SOM, the application of OAs clearly has the potential for most immediate impact, given it is a direct addition of exogenous OM. Even in no-till systems, repeated applications can lead to increased SOC down the profile, and coupled with an emerging recognition that occasional soil disturbance may also be required to overcome some physical soil constraints, this provides an opportunity for their direct incorporation. Nonetheless, availability of OAs and their cost of transport long distances from their source can be a barrier to adoption in many dryland regions, and despite the direct addition of C they provide, their diverse quality and interaction with different soil types does not guarantee production benefits.
All other interventions rely on increasing the amount of plant C inputs, increasing the amount of C stabilised (reducing losses), or both. In cropping systems that by design seek to optimise the amount of fixed C that is transferred to the harvested grain, this can be difficult to achieve through improved agronomy alone. Adoption of no-till or conservation tillage systems is not a guarantee of increased SOC, particularly when depths >30 cm are considered. This is despite improvements in water use efficiency, and thus crop biomass production, which are often observed. Other interventions including diversification of crop rotations, intercropping, or cover crops may have positive or negative impacts on SOC depending on specific circumstances, and it is important to understand changes in SOC throughout the soil profile.
Taking cover crops during the summer fallow periods that typify dryland cropping systems as an example of a more major intervention: if the cover crop is well managed, terminated effectively, and sufficient water remains available for the subsequent grain crop, then the greater C inputs from the cover crop and their likely deeper distribution should with time increase SOC. Although many studies in cool-temperate regions have shown improvements in broader soil health metrics as well as SOC, cover crops are typically grown in cooler wetter months in those systems where water availability is not a limiting factor to production. In the dryland scenario, if a cover crop uses water that would have been needed for the subsequent grain crop, especially at the critical early growth stages, its own productivity can be negatively affected, reducing both yields and C inputs. As with so many management decisions in dryland cropping systems, water availability and the ability of nutrients to maximise growth of the grain crop are the most important drivers of C inputs.
The other major intervention to increase C inputs is the incorporation of ley pasture phases into cropping systems. This reduces C exported from the system while increasing partitioning of C below ground, often to greater depths. Whereas the cycles of building or losing SOM under pasture and cropping phases of these mixed systems are expected, the length of these respective phases within mixed framing systems is driven by prevailing economic and environmental conditions.
In considering approaches to build SOM in dryland cropping systems, it is also important to focus on the end goal, the timeframe of intervention, and the downstream impacts of management changes to the farming system as a whole. Rather than focusing solely on building SOM for C sequestration, the benefits to system resilience of increased SOM are evident. However, as with most management interventions, economic costs may be difficult to offset or justify in both short- and long-term investment scenarios. Additionally, there may be unintended environmental impacts. Emerging issues such as the potential contest for biomass resources of stubble for biofuels versus their importance for soil surface protection and C inputs require resolution to ensure that well-intentioned actions do not affect other areas of the entire production system. Further research should focus on understanding these context-specific optima and the most effective strategies to achieve them while minimising risks in the whole-of-system context.
References
Abbott LK, Macdonald LM, Wong MTF, Webb MJ, Jenkins SN, Farrell M (2018) Potential roles of biological amendments for profitable grain production – a review. Agriculture, Ecosystems & Environment 256, 34-50.
| Crossref | Google Scholar |
Adcock D, McNeill AM, McDonald GK, Armstrong RD (2007) Subsoil constraints to crop production on neutral and alkaline soils in south-eastern Australia: a review of current knowledge and management strategies. Australian Journal of Experimental Agriculture 47(11), 1245-1261.
| Crossref | Google Scholar |
Adomako MO, Roiloa S, Yu F-H (2022) Potential roles of soil microorganisms in regulating the effect of soil nutrient heterogeneity on plant performance. Microorganisms 10(12), 2399.
| Crossref | Google Scholar |
Agnew JM, Leonard JJ (2003) The physical properties of compost. Compost Science & Utilization 11(3), 238-264.
| Crossref | Google Scholar |
Aitkenhead JA, McDowell WH (2000) Soil C:N ratio as a predictor of annual riverine DOC flux at local and global scales. Global Biogeochemical Cycles 14(1), 127-138.
| Crossref | Google Scholar |
Allison SD, Wallenstein MD, Bradford MA (2010) Soil-carbon response to warming dependent on microbial physiology. Nature Geoscience 3, 336-340.
| Crossref | Google Scholar |
Alvarez R (2005) A review of nitrogen fertilizer and conservation tillage effects on soil organic carbon storage. Soil Use and Management 21(1), 38-52.
| Crossref | Google Scholar |
Álvaro-Fuentes J, López MV, Cantero-Martinez C, Arrúe JL (2008) Tillage effects on soil organic carbon fractions in Mediterranean dryland agroecosystems. Soil Science Society of America Journal 72(2), 541-547.
| Crossref | Google Scholar |
Anderson GC, Fillery IRP, Dunin FX, Dolling PJ, Asseng S (1998) Nitrogen and water flows under pasture–wheat and lupin–wheat rotations in deep sands in Western Australia. 2. Drainage and nitrate leaching. Australian Journal of Agricultural Research 49(3), 345-362.
| Crossref | Google Scholar |
Angers DA, Recous S (1997) Decomposition of wheat straw and rye residues as affected by particle size. Plant and Soil 189, 197-203.
| Crossref | Google Scholar |
Angers DA, Eriksen-Hamel NS (2008) Full-inversion tillage and organic carbon distribution in soil profiles: a meta-analysis. Soil Science Society of America Journal 72(5), 1370-1374.
| Crossref | Google Scholar |
Angers D, Arrouays D, Cardinael R, Chenu C, Corbeels M, Demenois J, Farrell M, Martin M, Minasny B, Recous S, Six J (2022) A well-established fact: rapid mineralization of organic inputs is an important factor for soil carbon sequestration. European Journal of Soil Science 73(3), e13242.
| Crossref | Google Scholar |
Angus JF, Peoples MB (2012) Nitrogen from Australian dryland pastures. Crop & Pasture Science 63(9), 746-758.
| Crossref | Google Scholar |
Armstrong RD, Perris R, Munn M, Dunsford K, Robertson F, Hollaway GJ, O’Leary GJ (2019) Effects of long-term rotation and tillage practice on grain yield and protein of wheat and soil fertility on a Vertosol in a medium-rainfall temperate environment. Crop & Pasture Science 70, 1-15.
| Crossref | Google Scholar |
Arsic M, O’Sullivan CA, Wasson AP, Juliano P, MacMillan CP, Antille DL, Haling RE, Clarke WP (2022) Australia needs a national policy approach to successfully implement circular bioeconomy in agriculture and food systems. Farm Policy Journal 19, 46-60.
| Google Scholar |
Arzadún MJ, Arroquy JI, Laborde HE, Brevedan RE (2006) Effect of planting date, clipping height, and cultivar on forage and grain yield of winter wheat in Argentinean pampas. Agronomy Journal 98(5), 1274-1279.
| Crossref | Google Scholar |
Azevedo LB, Zelm RV, Leuven RSEW, Hendriks AJ, Huijbregts MAJ (2015) Combined ecological risks of nitrogen and phosphorus in European freshwaters. Environmental Pollution 200, 85-92.
| Crossref | Google Scholar |
Aziz MM, Palta JA, Siddique KHM, Sadras VO (2017) Five decades of selection for yield reduced root length density and increased nitrogen uptake per unit root length in Australian wheat varieties. Plant and Soil 413, 181-192.
| Crossref | Google Scholar |
Bai Y, Cortrufo MF (2022) Grassland soil carbon sequestration: current understanding, challenges, and solutions. Science 377(6606), 603-608.
| Crossref | Google Scholar |
Baldock JA, Hawke B, Sanderman J, Macdonald LM (2013) Predicting contents of carbon and its component fractions in Australian soils from diffuse reflectance mid-infrared spectra. Soil Research 51(8), 577-595.
| Crossref | Google Scholar |
Baldock JA, Creamer C, Szarvas S, McGowan J, Carter T, Farrell M (2021) Linking decomposition rates of soil organic amendments to their chemical composition. Soil Research 59, 630-643.
| Crossref | Google Scholar |
Balesdent J, Balabane M (1996) Major contribution of roots to soil carbon storage inferred from maize cultivated soils. Soil Biology and Biochemistry 28(9), 1261-1263.
| Crossref | Google Scholar |
Balesdent J, Chenu C, Balabane M (2000) Relationship of soil organic matter dynamics to physical protection and tillage. Soil and Tillage Research 53(3–4), 215-230.
| Crossref | Google Scholar |
Barton L, Schipper A, Barkle GF, McLeod M, Speir TW, Taylor MD, McGill AC, Van Schaik AP, Fitzgerald NB, Pandey SP (2005) Land application of domestic effluent onto four soil types: plant uptake and nutrient leaching. Journal of Environmental Quality 34, 635-643.
| Crossref | Google Scholar |
Barton L, Hoyle FC, Grace PR, Schwenke GD, Scanlan CA, Armstrong RD, Bell MJ (2022) Chapter one – soil nitrogen supply and N fertilizer losses from Australian dryland grain cropping systems. In ‘Advances in agronomy, Vol. 174’. (Ed. DL Sparks) pp. 1–52. (Academic Press) 10.1016/bs.agron.2022.03.001
Basile-Doelsch I, Balesdent J, Pellerin S (2020) Reviews and syntheses: the mechanisms underlying carbon storage in soil. Biogeosciences 17, 5223-5242.
| Crossref | Google Scholar |
Begill N, Don A, Poeplau C (2023) No detectable upper limit of mineral-associated organic carbon in temperate agricultural soils. Global Change Biology 29(16), 4662-4669.
| Crossref | Google Scholar |
Bellon-Maurel V, McBratney A (2011) Near-infrared (NIR) and mid-infrared (MIR) spectroscopic techniques for assessing the amount of carbon stock in soils – critical review and research perspectives. Soil Biology and Biochemistry 43(7), 1398-1410.
| Crossref | Google Scholar |
Berthelin J, Laba M, Lemaire G, Powlson D, Tessier D, Wander M, Baveye PC (2022) Soil carbon sequestration for climate change mitigation: mineralization kinetics of organic inputs as an overlooked limitation. European Journal of Soil Science 73(1), e13221.
| Crossref | Google Scholar |
Bian H, Li C, Zhu J, Xu L, Li M, Zheng S, He N (2022) Soil moisture affects the rapid response of microbes to labile organic C addition. Frontiers in Ecology and Evolution 10, 857185.
| Crossref | Google Scholar |
Bolinder MA, Crotty F, Elsen A, Frac M, Kismányoky T, Lipiec J, Tits M, Tóth Z, Kätterer T (2020) The effect of crop residues, cover crops, manures and nitrogen fertilization on soil organic carbon changes in agroecosystems: a synthesis of reviews. Mitigation and Adaptation Strategies for Global Change 25, 929-952.
| Crossref | Google Scholar |
Bonetti JdA, Anghinoni I, De Moraes MT, Fink JR (2017) Resilience of soils with different texture, mineralogy and organic matter under long-term conservation systems. Soil and Tillage Research 174, 104-112.
| Crossref | Google Scholar |
Borger CPD, Hashem A, Powles SB (2016) Manipulating crop row orientation and crop density to suppress Lolium rigidum. Weed Research 56(1), 22-30.
| Crossref | Google Scholar |
Bowman MS, Zilberman D (2013) Economic factors affecting diversified farming systems. Ecology and Society 18(1), 33.
| Crossref | Google Scholar |
Boyer JN, Groffman PM (1996) Bioavailability of water extractable organic carbon fractions in forest and agricultural soil profiles. Soil Biology and Biochemistry 28(6), 783-790.
| Crossref | Google Scholar |
Brooks PD, Haas PA, Huth AK (2007) Seasonal variability in the concentration and flux of organic matter and inorganic nitrogen in a semiarid catchment, San Pedro River, Arizona. JGR Biogeosciences 112, G03S04.
| Crossref | Google Scholar |
Brown RF, Collins SL (2023) As above, not so below: long-term dynamics of net primary production across a dryland transition zone. Global Change Biology 29(14), 3941-3953.
| Crossref | Google Scholar |
Brown DJ, Shepherd KD, Walsh MG, Dewayne Mays M, Reinsch TG (2006) Global soil characterization with VNIR diffuse reflectance spectroscopy. Geoderma 132(3–4), 273-290.
| Crossref | Google Scholar |
Brown S, Ippolito JA, Hundal LS, Basta NT (2020) Municipal biosolids – a resource for sustainable communities. Current Opinion in Environmental Science & Health 14, 56-62.
| Crossref | Google Scholar |
Button ES, Pett-Ridge J, Murphy DV, Kuzyakov Y, Chadwick DR, Jones DL (2022) Deep-C storage: biological, chemical and physical strategies to enhance carbon stocks in agricultural subsoils. Soil Biology and Biochemistry 170, 108697.
| Crossref | Google Scholar |
Cai A, Han T, Ren T, Sanderman J, Rui Y, Wang B, Smith P, Xu M, Li Y (2022) Declines in soil carbon storage under no tillage can be alleviated in the long run. Geoderma 425, 116028.
| Crossref | Google Scholar |
Calderon FJ, Vigil MF, Benjamin J (2018) Compost input effect on dryland wheat and forage yields and soil quality. Pedosphere 28(3), 451-462.
| Crossref | Google Scholar |
Cann DJ, Hunt JR, Malcolm B (2020a) Long fallows can maintain whole-farm profit and reduce risk in semi-arid south-eastern Australia. Agricultural Systems 178, 102721.
| Crossref | Google Scholar |
Cann DJ, Schillinger WF, Hunt JR, Porker KD, Harris FAJ (2020b) Agroecological advantages of early-sown winter wheat in semi-arid environments: a comparative case study from southern Australia and Pacific Northwest United States. Frontiers in Plant Science 11, 568.
| Crossref | Google Scholar |
Celestina C, Midwood J, Sherriff S, Trengove S, Hunt J, Tang C, Sale P, Franks A (2018) Crop yield responses to surface and subsoil applications of poultry litter and inorganic fertiliser in south-eastern Australia. Crop & Pasture Science 69(3), 303-316.
| Crossref | Google Scholar |
Celestina C, Hunt JR, Sale PWG, Franks AE (2019) Attribution of crop yield responses to application of organic amendments: a critical review. Soil and Tillage Research 186, 135-145.
| Crossref | Google Scholar |
Chantigny MH (2003) Dissolved and water-extractable organic matter in soils: a review on the influence of land use and management practices. Geoderma 113(3–4), 357-380.
| Crossref | Google Scholar |
Chapman PJ, McDonald AT, Tyson R, Palmer SM, Mitchell G, Irvine B (2010) Changes in water colour between 1986 and 2006 in the headwaters of the River Nidd, Yorkshire, UK. Biogeochemistry 101, 281-294.
| Crossref | Google Scholar |
Chappell A, Baldock J, Sanderman J (2016) The global significance of omitting soil erosion from soil organic carbon cycling schemes. Nature Climate Change 6, 187-191.
| Crossref | Google Scholar |
Chatterjee A, Lal R, Wielopolski L, Martin MZ, Ebinger MH (2009) Evaluation of different soil carbon determination methods. Critical Reviews in Plant Science 28(3), 164-178.
| Crossref | Google Scholar |
Chatterjee A, Cooper K, Klaustermeier A, Awale R, Cihacek LJ (2016) Does crop species diversity influence soil carbon and nitrogen pools? Agronomy Journal 108(1), 427-432.
| Crossref | Google Scholar |
Chen X, Cabrera ML, Zhang L, Shi Y, Shen SM (2003) Long-term decomposition of organic materials with different carbon/nitrogen ratios. Communications in Soil Science and Plant Analysis 34(1–2), 41-54.
| Crossref | Google Scholar |
Chowbury S, Farrell M, Bolan N (2014) Priming of soil organic carbon by malic acid addition is differentially affected by nutrient availability. Soil Biology and Biochemistry 77, 158-169.
| Crossref | Google Scholar |
Cleveland CC, Liptzin D (2007) C:N:P stoichiometry in soil: is there a ‘Redfield ratio’ for the microbial biomass? Biogeochemistry 85, 235-252.
| Crossref | Google Scholar |
Cong W-F, Hoffland E, Li L, Six J, Sun J-H, Bao X-G, Zhang F-S, Van Der Werf W (2015) Intercropping enhances soil carbon and nitrogen. Global Change Biology 21(4), 1715-1726.
| Crossref | Google Scholar |
Conyers M, Van der Rijt V, Oates A, Poile G, Kirkegaard J, Kirkby C (2019) The strategic use of minimum tillage within conservation agriculture in southern New South Wales, Australia. Soil and Tillage Research 193, 17-26.
| Crossref | Google Scholar |
Coonan EC, Kirkby CA, Kirkegaard JA, Amidy MR, Strong CL, Richardson AE (2020) Microorganisms and nutrient stoichiometry as mediators of soil organic matter dynamics. Nutrient Cycling in Agroecosystems 117, 273-298.
| Crossref | Google Scholar |
Cossani CM, Sadras VO (2018) Chapter six water–nitrogen colimitation in grain crops. In ‘Advances in agronomy, Vol. 150’. (Ed. DL Sparks) pp. 231–274. (Academic Press) 10.1016/bs.agron.2018.02.004
Cotrufo MF, Wallenstein MD, Boot CM, Denef K, Paul E (2012) The Microbial Efficiency-Matrix Stabilization (MEMS) framework integrates plant litter decomposition with soil organic matter stabilization: do labile plant inputs form stable soil organic matter? Global Change Biology 19(4), 988-995.
| Crossref | Google Scholar |
Cotrufo MF, Ranalli MG, Haddix ML, Six J, Lugato E (2019) Soil carbon storage informed by particulate and mineral-associated organic matter. Nature Geoscience 12, 989-994.
| Crossref | Google Scholar |
Dacal M, Garcia-Palacios P, Asensio S, Wang J, Singh BK, Maestre FT (2022) Climate change legacies contrastingly affect the resistance and resilience of soil microbial communities and multifunctionality to extreme drought. Functional Ecology 36(4), 908-920.
| Crossref | Google Scholar |
Dalal RC, Strong WM, Weston EJ, Cooper JE, Lehane KJ, King AJ (2018) Farming systems’ productivity and soil organic carbon stocks following fertilisers, no-tillage or legumes on a fertility-depleted soil in a semi-arid subtropical region. Soil Research 56(4), 429-440.
| Crossref | Google Scholar |
Davis S, Armstrong R, Macdonald L, Condon J, Petersen E (2019) Soil constraints: a role for stategic deep tillage. In ‘Australian agriculture in 2020: from conservation to automation’. (Eds J Pratley, J Kirkegaard) pp. 117–135. (Agronomy Australia & Charles Sturt University: Wagga Wagga, NSW, Australia)
DCCEEW (2021) ‘Carbon credits (carbon farming initiative—estimation of soil organic carbon sequestration using measurement and models) methodology determination 2021.’ (Department of Climate Change, the Environment and Water. Federal Register of Legislation) Available at https://www.legislation.gov.au/F2021L01696/latest/text
De Vries FT, Thebault E, Liiri M, Birkhofer K, Tsiafouli MA, Bjørnlund L, Bracht Jørgensen H, Brady MV, Christensen S, De Ruiter PC, d’Hertefeldt T, Frouz J, Hedlund K, Hemerik L, Hol WHG, Hotes S, Mortimer SR, Setälä H, Sgardelis SP, Uteseny K, Van der Putten WH, Wolters V, Bardgett RD (2013) Soil food web properties explain ecosystem services across European land use systems. Proceedings of the National Academy of Sciences 110, 14296-14301.
| Crossref | Google Scholar |
De Vries FT, Griffiths RI, Bailey M, Craig H, Girlanda M, Gweon HS, Hallin S, Kaisermann A, Keith AM, Kretzschmar M, Lemanceau P, Lumini E, Mason KE, Oliver A, Ostle N, Prosser JI, Thion C, Thomson B, Bardgett RD (2018) Soil bacterial networks are less stable under drought than fungal networks. Nature Communications 9, 3033.
| Crossref | Google Scholar |
Deng L, Shangguan Z, Bell SM, Soromotin AV, Peng C, An S, Wu X, Xu X, Wang K, Li J, Tang Z, Yan W, Zhang F, Li J, Wu J, Kuzyakov Y (2023) Carbon in Chinese grasslands: meta-analysis and theory of grazing effects. Carbon Research 2, 19.
| Crossref | Google Scholar |
Desbiolles J, Saunder C, Barr J, Riethmuller G, Northover G, Tullberg J, Antille D (2019) Machinery evolution for conservation agriculture. In ‘Australian agriculture in 2020: from conservation to automation’. (Eds J Pratley, J Kirkegaard) pp. 81–105. (Agronomy Australia and Charles Stuart University: Wagga Wagga, NSW, Australia)
Diacono M, Montemurro F (2010) Long-term effects of organic amendments on soil fertility. A review. Agronomy for Sustainable Development 30, 401-422.
| Crossref | Google Scholar |
Dias T, Dukes A, Antunes PM (2015) Accounting for soil biotic effects on soil health and crop productivity in the design of crop rotations. Journal of the Science of Food and Agriculture 95(3), 447-454.
| Crossref | Google Scholar |
Díaz-Zorita M, Duarte GA, Grove JH (2002) A review of no-till systems and soil management for sustainable crop production in the subhumid and semiarid Pampas of Argentina. Soil and Tillage Research 65(1), 1-18.
| Crossref | Google Scholar |
Dietzel R, Leibman M, Archontoulis S (2017) A deeper look at the relationship between root carbon pools and the vertical distribution of the soil carbon pool. SOIL 3, 139-152.
| Crossref | Google Scholar |
Domeignoz-Horta LA, Pold G, Liu X-JA, Frey SD, Melillo JM, DeAngelis KM (2020) Microbial diversity drives carbon use efficiency in a model soil. Nature Communications 11, 3684.
| Crossref | Google Scholar |
Don A, Schumacher J, Freibauer A (2010) Impact of tropical land-use change on soil organic carbon stocks – a meta-analysis. Global Change Biology 17(4), 1658-1670.
| Crossref | Google Scholar |
Donat MG, Lowry AL, Alexander LV, O’gorman PA, Maher N (2016) More extreme precipitation in the world’s dry and wet regions. Nature Climate Change 6, 508-513.
| Crossref | Google Scholar |
Dorado J, Zancada M-C, Almendros G, Lopez-Fando C (2003) Changes in soil properties and humic substances after long-term amendments with manure and crop residues in dryland farming systems. Journal of Plant Nutrition and Soil Science 166(1), 31-38.
| Crossref | Google Scholar |
Duivenbooden NV, Pala M, Studer C, Bielders CL, Beukes DJ (2000) Cropping systems and crop complementarity in dryland agriculture to increase soil water use efficiency: a review. NJAS: Wageningen Journal of Life Sciences 48(3), 213-236.
| Crossref | Google Scholar |
Dungait JAJ, Hopkins DW, Gregory AS, Whitmore AP (2012) Soil organic matter turnover is governed by accessibility not recalcitrance. Global Change Biology 18(6), 1781-1796.
| Crossref | Google Scholar |
Dynarski KA, Bossio DA, Scow KM (2020) Dynamic stability of soil carbon: reassessing the ‘permanence’ of soil carbon sequestration. Frontiers in Environmental Science 8, 514701.
| Crossref | Google Scholar |
Ebersold F, Hechelmann R-H, Holzapfel P, Meschede H (2023) Carbon insetting as a measure to raise supply chain energy efficiency potentials: opportunities and challenges. Energy Conversion and Management: X 20, 100504.
| Crossref | Google Scholar |
Ekblad A, Wallander H, Godbold DL, Cruz C, Johnson D, Baldrian P, Björk RG, Epron D, Kieliszewska-Rokicka B, Kjøller R, Kraigher H, Matzner E, Neumann J, Plassard C (2013) The production and turnover of extramatrical mycelium of ectomycorrhizal fungi in forest soils: role in carbon cycling. Plant and Soil 366, 1-27.
| Crossref | Google Scholar |
Engedal T, Magid J, Hansen V, Rasmussen J, Sorensen H, Jensen LS (2023) Cover crop root morphology rather than quality controls the fate of root and rhizodeposition C into distinct soil C pools. Global Change Biology 29(19), 5677-5690.
| Crossref | Google Scholar |
Erenstein O, Gerad B, Tittonell P (2015) Biomass use trade-offs in cereal cropping systems in the developing world: overview. Agricultural Systems 134, 1-5.
| Crossref | Google Scholar |
Ernakovich JG, Baldock J, Creamer C, Sanderman J, Kalbitz K, Farrell M (2021) A combined microbial and ecosystem metric of carbon retention efficiency explains land cover-dependent soil microbial biodiversity–ecosystem function relationships. Biogeochemistry 153, 1-15.
| Crossref | Google Scholar |
European Commission (2023) International carbon market. Available at https://climate.ec.europa.eu/eu-action/eu-emissions-trading-system-eu-ets/international-carbon-market_en#:~:text=Besides%20the%20EU%20emissions%20trading,Switzerland%20and%20the%20United%20States [accessed 1 Septemeber 2023]
Evans SE, Allison SD, Hawkes CV (2022) Microbes, memory and moisture: predicting microbial moisture responses and their impact on carbon cycling. Functional Ecology 36(6), 1430-1441.
| Crossref | Google Scholar |
Eyles A, Coghlan G, Hardie M, Hovenden M, Bridle K (2015) Soil carbon sequestration in cool-temperate dryland pastures: mechanisms and management options. Soil Research 53(4), 349-365.
| Crossref | Google Scholar |
Farrell M, Macdonald LM, Butler G, Chirino-Valle I, Condron LM (2012) Biochar and fertiliser applications influence phosphorus fractionation and wheat yield. Biology and Fertility of Soils 50, 169-178.
| Crossref | Google Scholar |
Feng Q, An C, Chen Z, Wang Z (2020) Can deep tillage enhance carbon sequestration in soils? A meta-analysis towards GHG mitigation and sustainable agricultural management. Renewable and Sustainable Energy Reviews 133, 110293.
| Crossref | Google Scholar |
Fernandez CW, Koide RT (2014) Initial melanin and nitrogen concentrations control the decomposition of ectomycorrhizal fungal litter. Soil Biology and Biochemistry 77, 150-157.
| Crossref | Google Scholar |
Fettell NA, Gill HS (1995) Long-term effects of tillage, stubble, and nitrogen management on properties of a red-brown earth. Australian Journal of Experimental Agriculture 35(7), 923-928.
| Crossref | Google Scholar |
Fiedler SR, Leinweber P, Jurasinski G, Eckhardt K-U, Glatzel S (2016) Tillage-induced short-term soil organic matter turnover and respiration. SOIL 2, 475-486.
| Crossref | Google Scholar |
Fierer N, Schimel JP (2003) A proposed mechanism for the pulse in carbon dioxide production commonly observed following the rapid rewetting of a dry soil. Soil Science Society of America Journal 67(3), 798-805.
| Crossref | Google Scholar |
Fisk LM, Barton L, Jones DL, Glanville HC, Murphy DV (2015) Root exudate carbon mitigates nitrogen loss in a semi-arid soil. Soil Biology and Biochemistry 88, 380-389.
| Crossref | Google Scholar |
Flohr BM, Hunt JR, Kirkegaard JA, Rheinheimer B, Swan T, Goward L, Evans JR, Bullock M (2020) Deep soil water-use determines the yield benefit of long-cycle wheat. Frontiers in Plant Science 11, 548.
| Crossref | Google Scholar |
Florence AM, McGuire AM (2020) Do diverse cover crop mixtures perform better than monocultures? A systematic review. Agronomy Journal 112(5), 3513-3534.
| Crossref | Google Scholar |
Fonte SJ, Yeboah E, Ofori P, Quansah GW, Vanlauwe B, Six J (2009) Fertilizer and residue quality effects on organic matter stabilization in soil aggregates. Soil Science Society of America Journal 73(3), 961-966.
| Crossref | Google Scholar |
Franzluebbers AJ, Stuedemann JA (2015) Does grazing of cover crops impact biologically active soil carbon and nitrogen fractions under inversion or no tillage management? Journal of Soil and Water Conservation 70(6), 365-373.
| Crossref | Google Scholar |
Freeman C, Evans CD, Monteith DT, Reynolds B, Fenner N (2001) Export of organic carbon from peat soils. Nature 412, 785.
| Crossref | Google Scholar |
Frey SD, Gupta VVSR, Elliott ET, Paustian K (2001) Protozoan grazing affects estimates of carbon utilization efficiency of the soil microbial community. Soil Biology and Biochemistry 33(12–13), 1759-1768.
| Crossref | Google Scholar |
Fu C, Chen Z, Wang G, Yu Z, Yu G (2021) A comprehensive framework for evaluating the impact of land use change and management on soil organic carbon stocks in global drylands. Current Opinion in Environmental Sustainability 48, 103-109.
| Crossref | Google Scholar |
Garba II, Bell LW, Williams A (2022) Cover crop legacy impacts on soil water and nitrogen dynamics, and on subsequent crop yields in drylands: a meta-analysis. Agronomy for Sustainable Development 42, 34.
| Crossref | Google Scholar |
Garland G, Edlinger A, Banerjee S, Degrune F, García-Palacios P, Pescador DS, Herzog C, Romdhane S, Saghai A, Spor A, Wagg C, Hallin S, Maestre FT, Philippot L, Rillig MC, Van der Heijden MGA (2021) Crop cover is more important than rotational diversity for soil multifunctionality and cereal yields in European cropping systems. Nature Food 2, 28-37.
| Crossref | Google Scholar |
Geisen S, Hu S, Dela Cruz TEE, Veen GF (2021) Protists as catalyzers of microbial litter breakdown and carbon cycling at different temperature regimes. The ISME Journal 15(2), 618-621.
| Crossref | Google Scholar |
Georgiou K, Jackson RJ, Vinduskova O, Abramoff RZ, Ahlström A, Feng W, Harden JW, Pellegrini AFA, Polley HW, Soong JL, Riley WJ, Torn MS (2022) Global stocks and capacity of mineral-associated soil organic carbon. Nature Communications 13, 3797.
| Crossref | Google Scholar |
Giller KE, Andersson JA, Corbeels M, Kirkegaard J, Mortensen D, Erenstein O, Vanlauwe B (2015) Beyond conservation agriculture. Frontiers in Plant Science 6, 870.
| Crossref | Google Scholar |
Giunta F, Motzo R, Fois G, Bacciu P (2014) Developmental ideotype in the context of the dual-purpose use of triticale, barley and durum wheat. Annals of Applied Biology 166(1), 118-128.
| Crossref | Google Scholar |
Gomez-Macpherson H, Richards RA (1995) Effect of sowing time on yield and agronomic characteristics of wheat in south-eastern Australia. Australian Journal of Agricultural Research 46(7), 1381-1399.
| Crossref | Google Scholar |
Griffiths BS, Philippot L (2013) Insights into the resistance and resilience of the soil microbial community. FEMS Microbiology Reviews 37(2), 112-129.
| Crossref | Google Scholar |
Guo LB, Gifford RM (2002) Soil carbon stocks and land use change: a meta analysis. Global Change Biology 8(4), 345-360.
| Crossref | Google Scholar |
Guo S, Wu J, Coleman K, Zhu H, Li Y, Liu W (2012) Soil organic carbon dynamics in a dryland cereal cropping system of the Loess Plateau under long-term nitrogen fertilizer applications. Plant and Soil 353, 321-332.
| Crossref | Google Scholar |
Gupta VVSR, Roper MM, Thompson J (2019) Harnessing the benefits of soil biology in conservation agriculture. In ‘Australian agriculture in 2020: from conservation to automation’. (Eds J Pratley, J Kirkegaard) pp. 237–253. (Agronomy Australia and Charles Sturt University: Wagga Wagga, NSW, Australia)
Hanan NP, Milne E, Aynekulu E, Yu Q, Anchang J (2021) A role for drylands in a carbon neutral world? Frontiers in Environmental Science 9, 786087.
| Crossref | Google Scholar |
Hannula SE, Morrien E (2022) Will fungi solve the carbon dilemma? Geoderma 413, 115767.
| Crossref | Google Scholar |
Hardy B, Dufey JE (2017) The resistance of centennial soil charcoal to the ‘Walkley-Black’ oxidation. Geoderma 303, 37-43.
| Crossref | Google Scholar |
Haugwitz MS, Bergmark L, Prieme A, Christensen S, Beier C, Michelsen A (2013) Soil microorganisms respond to five years of climate change manipulations and elevated atmospheric CO2 in a temperate heath ecosystem. Plant and Soil 374, 211-222.
| Crossref | Google Scholar |
He X, Abramoff R, Abs E, Georgiou K, Zhang H, Goll DS (2023) Contribution of carbon inputs to soil carbon accumulation cannot be neglected. bioRxiv 1-9.
| Crossref | Google Scholar |
Helgason BL, Janzen HH, Chantigny MH, Drury CF, Ellert BH, Gregorich EG, Lemke RL, Pattey E, Rochette P, Wagner-Riddle C (2005) Toward improved coefficients for predicting direct N2O emissions from soil in Canadian agroecosystems. Nutrient Cycling in Agroecosystems 72, 87-99.
| Crossref | Google Scholar |
Helgason BL, Gregorich EG, Janzen HH, Ellert BH, Lorenz N, Dick RP (2014) Long-term microbial retention of residue C is site-specific and depends on residue placement. Soil Biology and Biochemistry 68, 231-240.
| Crossref | Google Scholar |
Henry F, Nguyen C, Paterson E, Sim A, Robin C (2005) How does nitrogen availability alter rhizodeposition in Lolium multiflorum Lam. during vegetative growth? Plant and Soil 269, 181-191.
| Crossref | Google Scholar |
Hinsinger P, Bengough AG, Vetterlein D, Young IM (2009) Rhizosphere: biophysics, biogeochemistry and ecological relevance. Plant and Soil 321, 117-152.
| Crossref | Google Scholar |
Hirte J, Leifeld J, Abiven S, Oberholzer H-R, Mayer J (2018) Below ground carbon inputs to soil via root biomass and rhizodeposition of field-grown maize and wheat at harvest are independent of net primary productivity. Agriculture, Ecosystems & Environment 265, 556-566.
| Crossref | Google Scholar |
Hobley E, Wilson B, Wilkie A, Gray J, Koen T (2015) Drivers of soil organic carbon storage and vertical distribution in Eastern Australia. Plant and Soil 390, 111-127.
| Crossref | Google Scholar |
Hochman Z, Horan H (2018) Causes of wheat yield gaps and opportunities to advance the water-limited yield frontier in Australia. Field Crops Research 228, 20-30.
| Crossref | Google Scholar |
Holz M, Zarebanadkouki M, Kaestner A, Kuzyakov Y, Carminati A (2018) Rhizodeposition under drought is controlled by root growth rate and rhizosphere water content. Plant and Soil 423, 429-442.
| Crossref | Google Scholar |
Hoyle FC, Antuono MD, Overheu T, Murphy DV (2013) Capacity for increasing soil organic carbon stocks in dryland agricultural systems. Soil Research 51(8), 657-667.
| Crossref | Google Scholar |
Hudson BD (1994) Soil organic matter and available water capacity. Journal of Soil and Water Conservation 49(2), 189-194.
| Crossref | Google Scholar |
Hunt JR, Browne C, McBeath TM, Verburg K, Craig S, Whitbread AM (2013) Summer fallow weed control and residue management impacts on winter crop yield though soil water and N accumulation in a winter-dominant, low rainfall region of southern Australia. Crop & Pasture Science 64(9), 922-934.
| Crossref | Google Scholar |
Hunt JR, Lilley JM, Trevaskis B, Flohr BM, Peake A, Fletcher A, Zwart AB, Gobbett D, Kirkegaard JA (2019) Early sowing systems can boost Australian wheat yields despite recent climate change. Nature Climate Change 9, 244-247.
| Crossref | Google Scholar |
Isbell F, Adler PR, Eisenhauer N, Fornara D, Kimmel K, Kremen C, Letourneau DK, Liebman M, Polley HW, Quijas S, Scherer-Lorenzen M, Bardgett R (2017) Benefits of increasing plant diversity in sustainable agroecosystems. Journal of Ecology 105, 871-879.
| Crossref | Google Scholar |
Jaconi A, Poeplau C, Ramirez-Lopez L, Van Wesemael B, Don A (2019) Log-ratio transformation is the key to determining soil organic carbon fractions with near-infrared spectroscopy. European Journal of Soil Science 70(1), 127-139.
| Crossref | Google Scholar |
Janik LJ, Skjemstad JO, Shepherd KD, Spouncer LR (2007) The prediction of soil carbon fractions using mid-infrared-partial least square analysis. Australian Journal of Soil Research 45(2), 73-81.
| Crossref | Google Scholar |
Janzen HH, Van Groenigen KJ, Powlson DS, Schwinghamer T, Van Groenigen JW (2022) Photosynthetic limits on carbon sequestration in croplands. Geoderma 416, 115810.
| Crossref | Google Scholar |
Jeffery S, Verheijen FGA, Van derVelde M, Bastos AC (2011) A quantitative review of the effects of biochar application to soils on crop productivity using meta-analysis. Agriculture, Ecosystems & Environment 144(1), 175-187.
| Crossref | Google Scholar |
Jia X, Chen S, Yang Y, Zhou L, Yu W, Shi Z (2017) Organic carbon prediction in soil cores using VNIR and MIR techniques in an alpine landscape. Scientific Reports 7, 2144.
| Crossref | Google Scholar |
Johnston AE, Poulton PR, Coleman K (2009) Chapter 1. Soil organic matter: its importance in sustainable agriculture and carbon dioxide fluxes. In ‘Advances in agronomy’. (Ed. DL Sparks) pp. 1–57. (Academic Press) 10.1016/S0065-2113(08)00801-8
Jones DL (1998) Organic acids in the rhizosphere – a critical review. Plant and Soil 205, 25-44.
| Crossref | Google Scholar |
Jones DL (1999) Amino acid biodegradation and its potential effects on organic nitrogen capture by plants. Soil Biology and Biochemistry 31(4), 613-622.
| Crossref | Google Scholar |
Jones DL, Nguyen C, Finlay RD (2009) Carbon flow in the rhizosphere: carbon trading at the soil–root interface. Plant and Soil 321, 5-33.
| Crossref | Google Scholar |
Kalbitz K, Kaiser K (2008) Contribution of dissolved organic matter to carbon storage in forest mineral soils. Journal of Plant Nutrition and Soil Science 171(1), 52-60.
| Crossref | Google Scholar |
Kane DA, Bradford MA, Fuller E, Oldfield EE, Wood SA (2021) Soil organic matter protects US maize yields and lowers crop insurance payouts under drought. Environmental Research Letters 16(4), 044018.
| Crossref | Google Scholar |
Karunaratne SB, Bishop TFA, Odeh IOA, Baldock JA, Marchant BP (2014) Estimating change in soil organic carbon using legacy data as the baseline: issues, approaches and lessons to learn. Soil Research 52(4), 349-365.
| Crossref | Google Scholar |
Kassam A, Friedrich T, Derpsch R (2022) Successful experiences and lessons from conservation agriculture worldwide. Agronomy 12(4), 769.
| Crossref | Google Scholar |
Kemanian AR, Stockle CO, Huggins DR (2005) Transpiration-use efficiency of barley. Agricultural and Forest Meteorology 130(1–2), 1-11.
| Crossref | Google Scholar |
Kibblewhite MG, Ritz K, Swift MJ (2008) Soil health in agricultural systems. Philosophical Transactions of the Royal Society B: Biological Sciences 363, 685-701.
| Crossref | Google Scholar |
Kirkby CA, Richardson AE, Wade LJ, Conyers M, Kirkegaard JA (2016) Inorganic nutrients increase humification efficiency and C-sequestration in an annually cropped soil. PLoS ONE 11, e0153698.
| Crossref | Google Scholar |
Kirkby CA, Kirkegaard JA, Richardson AE (2021) Gravel-associated organic material is important to quantify soil carbon and nitrogen stocks to depth in an agricultural cropping soil. Soil Research 60(3), 224-233.
| Crossref | Google Scholar |
Kirkegaard JA, Sprague SJ, Dove H, Kelman WM, Marcroft SJ, Lieschke A, Howe GN, Graham JM (2008) Dual-purpose canola – a new opportunity in mixed farming systems. Australian Journal of Agricultural Research 59, 291-302.
| Crossref | Google Scholar |
Kirkegaard J, Kirkby C, Oates A, Van der Rijt V, Poile G, Conyers M (2020) Strategic tillage of a long-term, no-till soil has little impact on soil characteristics or crop growth over five years. Crop & Pasture Science 71(12), 945-958.
| Crossref | Google Scholar |
Kirkegaard JA, Richardson AE, Kirkby CA (2023) Fate and cost effectiveness of soil carbon sequestered using supplementary nutrients applied to crop residues under field conditions. Nutrient Cycling in Agroecosystems 126, 143-161.
| Crossref | Google Scholar |
Kopittke PM, Dalal RC, Finn D, Menzies NW (2016) Global changes in soil stocks of carbon, nitrogen, phosphorus, and sulphur as influenced by long-term agricultural production. Global Change Biology 23(6), 2509-2519.
| Crossref | Google Scholar |
Koteen LE, Baldocchi DD, Harte J (2011) Invasion of non-native grasses causes a drop in soil carbon storage in California grasslands. Environmental Research Letters 6(4), 044001.
| Crossref | Google Scholar |
Krall JM, Schuman GE (1996) Integrated dryland crop and livestock production systems on the great plains: extent and outlook. Journal of Production Agriculture 9(2), 187-191.
| Crossref | Google Scholar |
Kristensen JA, Svenning J-C, Georgiou K, Malhi Y (2022) Can large herbivores enhance ecosystem carbon persistence? Trends in Ecology & Evolution 37(2), 117-128.
| Crossref | Google Scholar |
Krupnik TJ, Andersson JA, Ruisnamhodzi L, Corbeels M, Shennan C, Gérard B (2019) Does size matter? A critical review of meta-analysis in agronomy. Experimental Agriculture 55(2), 200-229.
| Crossref | Google Scholar |
Kuyah S, Muoni T, Bayala J, Chopin P, Dahlin AS, Hughes K, Jonsson M, Kumar S, Sileshi GW, Dimobe K, Öborn I (2023) Grain legumes and dryland cereals contribute to carbon sequestration in the drylands of Africa and South Asia. Agriculture, Ecosystems & Environment 355, 108583.
| Crossref | Google Scholar |
Kuzyakov Y, Friedel JK, Stahr K (2000) Review of mechanisms and quantification of priming effects. Soil Biology and Biochemistry 32, 11-12 1485-1498.
| Crossref | Google Scholar |
Ladd JN, Amato M, Grace PR, Van Veen JA (1995) Simulation of 14C turnover through the microbial biomass in soils incubated with 14C-labelled plant residues. Soil Biology and Biochemistry 27(6), 777-783.
| Crossref | Google Scholar |
Ladha JK, Reddy CK, Padre AT, Van Kessel C (2011) Role of nitrogen fertilization in sustaining organic matter in cultivated soils. Journal of Environmental Quality 40(6), 1756-1766.
| Crossref | Google Scholar |
Lahmar R, Bationo BA, Lamso ND, Guero Y, Tittonell P (2012) Tailoring conservation agriculture technologies to West Africa semi-arid zones: building on traditional local practices for soil restoration. Field Crops Research 132, 158-167.
| Crossref | Google Scholar |
Lal R (2004) Carbon sequestration in dryland ecosystems. Environmental Management 33, 528-544.
| Crossref | Google Scholar |
Lal R (2005) Enhancing crop yields in the developing countries through restoration of the soil organic carbon pool in agricultural lands. Land Degradation & Development 17(2), 197-209.
| Crossref | Google Scholar |
Lehmann J, Skjemstad J, Sohi S, Carter J, Barson M, Falloon P, Coleman K, Woodbury P, Krull E (2008) Australian climate–carbon cycle feedback reduced by soil black carbon. Nature Geoscience 1, 832-835.
| Crossref | Google Scholar |
Lehmann J, Bossio DA, Kögel-Knabner I, Rillig MC (2020) The concept and future prospects of soil health. Nature Reviews Earth & Environment 1, 544-553.
| Crossref | Google Scholar |
Lemerle D, Cousens RD, Gill GS, Peltzer SJ, Moerkerk M, Murphy CE, Collins D, Cullis BR (2004) Reliability of higher seeding rates of wheat for increased competitiveness with weeds in low rainfall environments. The Journal of Agricultural Science 142(4), 395-409.
| Crossref | Google Scholar |
Lemke RL, Vandenbygaart AJ, Campbell CA, Lafond GP, Grant B (2010) Crop residue removal and fertilizer N: effects on soil organic carbon in a long-term crop rotation experiment on a Udic Boroll. Agriculture, Ecosystems & Environment 135(1–2), 42-51.
| Crossref | Google Scholar |
Li L, Tilman D, Lambers H, Zhang F-S (2014) Plant diversity and overyielding: insights from belowground facilitation of intercropping in agriculture. New Phytologist 203(1), 63-69.
| Crossref | Google Scholar |
Li N, Xu Y-Z, Han X-Z, He H-B, Zhang X-D, Zhang B (2015) Fungi contribute more than bacteria to soil organic matter through necromass accumulation under different agricultural practices during the early pedogenesis of a Mollisol. European Journal of Soil Biology 67, 51-58.
| Crossref | Google Scholar |
Li P, Wu M, Kang G, Zhu B, Li H, Hu F, Jiao J (2020) Soil quality response to organic amendments on dryland red soil in subtropical China. Geoderma 373, 114416.
| Crossref | Google Scholar |
Lian Z, Piao S, Chen A, Huntingford C, Fu B, Li LZX, Huang J, Sheffield J, Berg AM, Keenan TF, McVicar TR, Wada Y, Wang X, Wang T, Yang Y, Roderick ML (2021) Multifaceted characteristics of dryland aridity changes in a warming world. Nature Reviews Earth & Environment 2, 232-250.
| Crossref | Google Scholar |
Liu C, Lu M, Cui J, Li B, Fang C (2014) Effects of straw carbon input on carbon dynamics in agricultural soils: a meta-analysis. Global Change Biology 20(5), 1366-1381.
| Crossref | Google Scholar |
Liu X, Zhou Z-Y, Cui J-L, Wang M-L, Wang J-H (2021) Biotransformation ability of endophytic fungi: from species evolution to industrial applications. Applied Microbiology and Biotechnology 105, 7095-7113.
| Crossref | Google Scholar |
Liu X, Tan S, Song X, Wu X, Zhao G, Li S, Liang G (2022) Response of soil organic carbon content to crop rotation and its controls: a global synthesis. Agriculture, Ecosystems & Environment 335, 108017.
| Crossref | Google Scholar |
Llewellyn RS, D’emden FH, Kuehne G (2012) Extensive use of no-tillage in grain growing regions of Australia. Field Crops Research 132, 204-212.
| Crossref | Google Scholar |
Lobsey CR, Viscarra Rossel RA (2016) Sensing of soil bulk density for more accurate carbon accounting. European Journal of Soil Science 67(4), 504-513.
| Crossref | Google Scholar |
López-Bellido L, López-Bellido R, Fernández-García P, Muñoz-Romero V, Lopez-Bellido FJ (2020) Carbon storage in a rainfed Mediterranean vertisol: effects of tillage and crop rotation in a long-term experiment. European Journal of Soil Science 71(3), 472-483.
| Crossref | Google Scholar |
Loveland P, Webb J (2003) Is there a critical level of organic matter in the agricultural soils of temperate regions: a review. Soil and Tillage Research 70(1), 1-18.
| Crossref | Google Scholar |
Lu X-T, Liu Z-Y, Hu Y-Y, Zhang H-Y (2018) Testing nitrogen and water co-limitation of primary productivity in a temperate steppe. Plant and Soil 432, 119-127.
| Crossref | Google Scholar |
Lu Z, Che Y, Rejesus RM, Goodwin BK, Ghosh SK, Paudel J (2023) Unintended environmental benefits of crop insurance: nitrogen and phosphorus in water bodies. Ecological Economics 204, 107657.
| Crossref | Google Scholar |
Luo Z, Wang E, Bryan BA, Grant T, Liu DL, Cowie A, Farquharson R, Simmons A, Crawford D, Searle R, Moore A (2013) Meta-modeling soil organic carbon sequestration potential and its application at regional scale. Ecological Applications 23, 408-420.
| Crossref | Google Scholar |
Luo Z, Eady S, Sharma B, Grant T, Liu DL, Cowie A, Farquharson R, Simmons A, Crawford D, Searle R, Moore A (2019) Mapping future soil carbon change and its uncertainty in croplands using simple surrogates of a complex farming system model. Geoderma 337, 311-321.
| Crossref | Google Scholar |
Ma B, Ma B-L, McLaughlin NB, Li M, Liu J (2022) Improvement in dryland crop performance and soil properties with multiple annual applications of lignite-derived humic amendment. Soil and Tillage Research 218, 105306.
| Crossref | Google Scholar |
Macdonald LM, Farrell M, Zwieten LV, Krull ES (2014) Plant growth responses to biochar addition: an Australian soils perspective. Biology and Fertility of Soils 50, 1035-1045.
| Crossref | Google Scholar |
Macdonald BCT, Ringrose-Voase AJ, Nadelko AJ, Farrell M, Tuomi S, Nachimuthu G (2016) Dissolved organic nitrogen contributes significantly to leaching from furrow-irrigated cotton–wheat–maize rotations. Soil Research 55(1), 70-77.
| Crossref | Google Scholar |
Machmuller MB, Kramer MG, Cyle TK, Hill N, Hancock D, Thompson A (2015) Emerging land use practices rapidly increase soil organic matter. Nature Communications 6, 6995.
| Crossref | Google Scholar |
MacKown CT, Carver BF (2005) Fall forage biomass and nitrogen composition of winter wheat populations selected from grain-only and dual-purpose environments. Crop Science 45(1), 322-328.
| Crossref | Google Scholar |
Maestre FT, Delgado-Baquerizo M, Jeffries TC, Eldridge DJ, Ochoa V, Gozalo B, Quero JL, García-Gómez M, Gallardo A, Ulrich W, Bowker MA, Arredondo T, Barraza-Zepeda C, Bran D, Florentino A, Gaitán J, Gutiérrez JR, Huber-Sannwald E, Jankju M, Mau RL, Miriti M, Naseri K, Ospina A, Stavi I, Wang , D, Woods NN, Yuan X, Zaady E, Singh BK (2015) Increasing aridity reduces soil microbial diversity and abundance in global drylands. Proceedings of the National Academy of Sciences 112(51), 15684-15689.
| Crossref | Google Scholar |
Manganyi MC, Ateba CN (2020) Untapped potentials of endophytic fungi: a review of novel bioactive compounds with biological applications. Microorganisms 8(12), 1934.
| Crossref | Google Scholar |
Manzoni S, Porporato A (2009) Soil carbon and nitrogen mineralization: theory and models across scales. Soil Biology and Biochemistry 41(7), 1355-1379.
| Crossref | Google Scholar |
Manzoni S, Taylor P, Ritchter A, Porporato A, Agren GI (2012) Environmental and stoichiometric controls on microbial carbon-use efficiency in soils. New Phytologist 196(1), 79-91.
| Crossref | Google Scholar |
Mattila TJ, Vihanto N (2024) Agricultural limitations to soil carbon sequestration: plant growth, microbial activity, and carbon stabilization. Agriculture, Ecosystems & Environment 367, 108986.
| Crossref | Google Scholar |
McCully ME, Canny MJ (1985) Localisation of translocated 14C in roots and root exudates of field-grown maize. Physiologia Plantarum 65(4), 380-392.
| Crossref | Google Scholar |
McDaniel MD, Tiemann LK, Grandy AS (2014) Does agricultural crop diversity enhance soil microbial biomass and organic matter dynamics? A meta-analysis. Ecological Applications 24(3), 560-570.
| Crossref | Google Scholar |
Meier EA, Hunt JR, Hochman Z (2021) Evaluation of nitrogen bank, a soil nitrogen management strategy for sustainably closing wheat yield gaps. Field Crops Research 261, 108017.
| Crossref | Google Scholar |
Melero S, Madejon E, Ruiz JC, Herencia JF (2007) Chemical and biochemical properties of a clay soil under dryland agriculture system as affected by organic fertilization. European Journal of Agronomy 26(3), 327-334.
| Crossref | Google Scholar |
Melesse MB, Tirra AN, Ojiewo CO, Hauser M (2021) Understanding farmers’ trait preferences for dual-purpose crops to improve mixed crop–livestock systems in Zimbabwe. Sustainability 13(10), 5678.
| Crossref | Google Scholar |
Minasny B, McBratney AB (2018) Limited effect of organic matter on soil available water capacity. European Journal of Soil Science 69(1), 39-47.
| Crossref | Google Scholar |
Minasny B, Malone BP, McBratney AB, Angers DA, Arrouays D, Chambers A, Chaplot V, Chen Z-S, Cheng K, Das BS, Field DJ, Gimona A, Hedley CB, Hong SY, Mandal B, Marchant BP, Martin M, McConkey BG, Mulder VL, O’Rourke S, Richer-de-Forges AC, Odeh I, Padarian J, Paustian K, Pan G, Poggio L, Savin I, Stolbovoy V, Stockmann U, Sulaeman Y, Tsui C-C, Vågen T-G, Van Wesemael B, Winowiecki L (2017) Soil carbon 4 per mille. Geoderma 292, 59-86.
| Crossref | Google Scholar |
Minasny B, Arrouays D, Cardinael R, Chabbi A, Farrell M, Henry B, Koutika L-S, Ladha JK, McBratney AB, Padarian J, Román Dobarco M, Rumpel C, Smith P, Soussana J-F (2022) Current NPP cannot predict future soil organic carbon sequestration potential. Comment on ‘photosynthetic limits on carbon sequestration in croplands’. Geoderma 424, 115975.
| Crossref | Google Scholar |
Mlih R, Bol R, Amelung W, Brahim N (2016) Soil organic matter amendments in date palm groves of the Middle Eastern and North African region: a mini-review. Journal of Arid Land 8, 77-92.
| Crossref | Google Scholar |
Moinet GYK, Hijbeek R, Vuuren DPV, Giller KE (2022) Carbon for soils, not soils for carbon. Global Change Biology 29(9), 2384-2398.
| Crossref | Google Scholar |
Möller K (2018) Soil fertility status and nutrient input–output flows of specialised organic cropping systems: a review. Nutrient Cycling in Agroecosystems 112, 147-164.
| Crossref | Google Scholar |
Mondal S, Chakraborty D (2022) Global meta-analysis suggests that no-tillage favourably changes soil structure and porosity. Geoderma 405, 115443.
| Crossref | Google Scholar |
Moore A, Hunt J (2010) Managing to increase soil carbon in southern Australian cropping systems: what are the interactions with water-use efficiency? In ‘XIth European Society of Agronomy Congress’, 29 August–3 September 2010, Montpellier. pp. 687–688. (European Society of Agronomy) Available at http://hdl.handle.net/102.100.100/106945?index=1
Motavalli PP, Palm CA, Parton WJ, Elliott ET, Frey SD (1994) Comparison of laboratory and modeling simulation methods for estimating soil carbon pools in tropical forest soils. Soil Biology and Biochemistry 26(8), 935-944.
| Crossref | Google Scholar |
Murphy BW (2015) Impact of soil organic matter on soil properties—a review with emphasis on Australian soils. Soil Research 53(6), 605-635.
| Crossref | Google Scholar |
Murphy DV, Macdonald AJ, Stockdale EA, Goulding KWT, Fortune S, Gaunt JL, Poulton PR, Wakefield JA, Webster CP, Wilmer WS (2000) Soluble organic nitrogen in agricultural soils. Biology and Fertility of Soils 30, 374-387.
| Crossref | Google Scholar |
Musinguzi P, Tenywa JS, Ebanyat P, Tenywa MM, Mubiru DN, Basamba TA, Leip A (2013) Soil organic carbon thresholds and nitrogen management in tropical agroecosystems: concepts and prospects. Journal of Sustainable Development 6(12), 31-43.
| Crossref | Google Scholar |
Musinguzi P, Ebanyet P, Tenywa JS, Basamba TA, Tenywa MM, Mubiru DN (2016) Critical soil organic carbon range for optimal crop response to mineral fertiliser nitrogen on a Ferralsol. Experimental Agriculture 52(4), 635-653.
| Crossref | Google Scholar |
Mustafa A, Frouz J, Naveed M, Ping Z, Nan S, Minggang X, Núñez-Delgado A (2022) Stability of soil organic carbon under long-term fertilization: results from 13C NMR analysis and laboratory incubation. Environmental Research 205, 112476.
| Crossref | Google Scholar |
Naylor D, McClure R, Jansson J (2022) Trends in microbial community composition and function by soil depth. Microorganisms 10(3), 540.
| Crossref | Google Scholar |
Nicolardot N, Bouziri L, Bastian F, Ranjard L (2007) A microcosm experiment to evaluate the influence of location and quality of plant residues on residue decomposition and genetic structure of soil microbial communities. Soil Biology and Biochemistry 39(7), 1631-1644.
| Crossref | Google Scholar |
Nicoloso RS, Rice CW, Amado TJC, Costa CN, Akley EK (2018) Carbon saturation and translocation in a no-till soil under organic amendments. Agriculture, Ecosystems & Environment 264, 73-84.
| Crossref | Google Scholar |
Novelli LE, Caviglia OP, Jobbagy EG, Sadras VO (2023) Diversified crop sequences to reduce soil nitrogen mining in agroecosystems. Agriculture, Ecosystems & Environment 341, 108208.
| Crossref | Google Scholar |
Oldfield EE, Wood SA, Bradford MA (2020) Direct evidence using a controlled greenhouse study for threshold effects of soil organic matter on crop growth. Ecological Applications 30(4), e02073.
| Crossref | Google Scholar |
Olesen JE, Rees RM, Recous S, Bleken MA, Abalos D, Ahuja I, Butterbach-Bahl K, Carozzi M, De Notaris C, Ernfors M, Haas E, Hansen S, Janz B, Lashermes G, Massad RS, Petersen Søren O, Rittl TF, Scheer C, Smith KE, Thiébeau P, Taghizadeh-Toosi A, Thorman RE, Topp CFE (2023) Challenges of accounting nitrous oxide emissions from agricultural crop residues. Global Change Biology 29, 6846-6855.
| Crossref | Google Scholar |
Pappa VA, Rees RM, Walker RL, Baddeley JA, Watson CA (2012) Legumes intercropped with spring barley contribute to increased biomass production and carry-over effects. The Journal of Agricultural Science 150(5), 584-594.
| Crossref | Google Scholar |
Parton WJ, Stewart JWB, Cole CV (1988) Dynamics of C, N, P and S in grassland soils: a model. Biogeochemistry 5, 109-131.
| Crossref | Google Scholar |
Parvin S, Bajwa A, Uddin S, Sandral G, Rose MT, Van Zweiten L, Rose TJ (2025) Impact of wheat–vetch temporary intercropping on soil functions and grain yield in a dryland semi-arid environment. Plant and Soil 506, 457-470.
| Crossref | Google Scholar |
Passioura J (2006) Increasing crop productivity when water is scarce—from breeding to field management. Agricultural Water Management 80, 176-196.
| Crossref | Google Scholar |
Paterson E (2003) Importance of rhizodeposition in the coupling of plant and microbial productivity. European Journal of Soil Science 54(4), 741-750.
| Crossref | Google Scholar |
Pausch J, Kuzyakov Y (2017) Carbon input by roots into the soil: quantification of rhizodeposition from root to ecosystem scale. Global Change Biology 24(1), 1-12.
| Crossref | Google Scholar |
Paustian K, Collier S, Baldock J, Burgess R, Creque J, DeLonge M, Dungait J, Ellert B, Frank S, Goddard T, Govaerts B, Grundy M, Henning M, Izaurralde RC, Madaras M, McConkey B, Porzig E, Rice C, Searle R, Seavy N, Skalsky R, Mulhern W, Jahn M (2019) Quantifying carbon for agricultural soil management: from the current status toward a global soil information system. Carbon Management 10(6), 567-587.
| Crossref | Google Scholar |
Peixoto L, Elsgaard L, Rasmussen J, Kuzyakov Y, Banfield CC, Dippold MA, Olesen JE (2020) Decreased rhizodeposition, but increased microbial carbon stabilization with soil depth down to 3.6 m. Soil Biology and Biochemistry 150, 108008.
| Crossref | Google Scholar |
Peralta AL, Sun Y, Mcdaniel MD, Lennon JT (2018) Crop rotational diversity increases disease suppressive capacity of soil microbiomes. Ecosphere 9(5), e02235.
| Crossref | Google Scholar |
Personeni E, Nguyen C, Marchal P, Pages L (2007) Experimental evaluation of an efflux–influx model of C exudation by individual apical root segments. Journal of Experimental Botany 58(8), 2091-2099.
| Crossref | Google Scholar |
Petrie B, Barden R, Kasprzyk-Hordern B (2015) A review on emerging contaminants in wastewaters and the environment: current knowledge, understudied areas and recommendations for future monitoring. Water Research 72, 3-27.
| Crossref | Google Scholar |
Plaza-Bonilla D, Arrue JL, Cantero-Martinez C, Fanlo R, Iglesias A, Alvaro-Fuentes J (2015) Carbon management in dryland agricultural systems. A review. Agronomy For Sustainable Development 35, 1319-1334.
| Crossref | Google Scholar |
Plaza C, Zaccone C, Sawicka K, Méndez AM, Tarquis A, Gascó G, Heuvelink GBM, Schuur EAG, Maestre FT (2018) Soil resources and element stocks in drylands to face global issues. Scientific Reports 8, 13788.
| Crossref | Google Scholar |
Powlson DS, Stirling CM, Jat ML, Gerard BG, Palm CA, Sanchez PA, Cassman KG (2014) Limited potential of no-till agriculture for climate change mitigation. Nature Climate Change 4, 678-683.
| Crossref | Google Scholar |
Prietzel J, Müller S, Kogel-Knabner I, Thieme J, Jaye C, Fischer D (2018) Comparison of soil organic carbon speciation using C NEXAFS and CPMAS 13C NMR spectroscopy. Science of the Total Environment 628–629, 906-918.
| Crossref | Google Scholar |
Qiu Q, Wu L, Quyang Z, Li B, Xu Y, Wu S, Gregorich EG (2015) Effects of plant-derived dissolved organic matter (DOM) on soil CO2 and N2O emissions and soil carbon and nitrogen sequestrations. Applied Soil Ecology 96, 122-130.
| Crossref | Google Scholar |
Qiu W, Liu J, Li B, Wang Z (2020) N2O and CO2 emissions from a dryland wheat cropping system with long-term N fertilization and their relationships with soil C, N, and bacterial community. Environmental Science and Pollution Research 27, 8673-8683.
| Crossref | Google Scholar |
Qui Y, Li X, Tang Y, Xiong S, Han Y, Wang Z, Feng L, Wang G, Yang B, Lei Y, Du W, Zhi X, Xin M, Jiao Y, Zhang S, Li Y, Li X-F (2023) Directly linking plant N, P and K nutrition to biomass production in cotton-based intercropping systems. European Journal of Agronomy 151, 126960.
| Crossref | Google Scholar |
Quinton JN, Govers G, Oost KV, Bardgett RD (2010) The impact of agricultural soil erosion on biogeochemical cycling. Nature Geoscience 3, 311-314.
| Crossref | Google Scholar |
Rashid MI, Mujawar LH, Shahzad T, Almeelbi T, Ismail IMI, Oves M (2016) Bacteria and fungi can contribute to nutrients bioavailability and aggregate formation in degraded soils. Microbiological Research 183, 26-41.
| Crossref | Google Scholar |
Rasse DP, Rumpel C, Dignac M-F (2005) Is soil carbon mostly root carbon? Mechanisms for a specific stabilisation. Plant and Soil 269, 341-356.
| Crossref | Google Scholar |
Rasse DP, Mulder J, Moni C, Chenu C (2006) Carbon turnover kinetics with depth in a French loamy soil. Soil Science Society of America Journal 70(6), 2097-2105.
| Crossref | Google Scholar |
Rassmussen C, Heckman K, Wieder WR, Keiluweit M, Lawrence CR, Berhe AA, Blankinship JC, Crow SE, Druhan JL, Hicks Pries CE, Marin-Spiotta E, Plante AF, Schädel C, Schimel JP, Sierra CA, Thompson A, Wagai R (2018) Beyond clay: towards an improved set of variables for predicting soil organic matter content. Biogeochemistry Letters 137, 297-306.
| Crossref | Google Scholar |
Redfield AC (1958) The biological control of chemical factors in the environment. American Scientist 46, 3 205-221.
| Google Scholar |
Rengasamy P, Olsson KA (1991) Sodicity and soil structure. Australian Journal of Soil Research 29(6), 935-962.
| Crossref | Google Scholar |
Riviere C, Bethinger A, Bergez J-E (2022) The effects of cover crops on multiple environmental sustainability indicators – a review. Agronomy 12(9), 2011.
| Crossref | Google Scholar |
Rose TJ, Parvin S, Han E, Condon J, Flohr BM, Schefe C, Rose MT, Kirkegaard JA (2022) Prospects for summer cover crops in southern Australian semi-arid cropping systems. Agricultural Systems 200, 103415.
| Crossref | Google Scholar |
Rusinamhodzi L, Van Wijk MT, Corbeels M, Rufino MC, Giller KE (2015) Maize crop residue uses and trade-offs on smallholder crop–livestock farms in Zimbabwe: economic implications of intensification. Agriculture, Ecosystems & Environment 214, 31-45.
| Crossref | Google Scholar |
Safriel U, Adeel Z, Neimeijer D, Puigdefabregas J, White R, Lal R, Winslow M, Ziedler J, Prince S, Archer E, King C, Shapiro B, Wessels K, Nielsen T, Portnov B, Reshef I, Thonell J, Lachman E, McNab D (2005) Dryland systems. In ‘Ecosystems and human well-being: current state and trends, Vol. 1’. (Eds M El-Kassas, E Ezcurra) pp. 625–662. (Millennium Ecosystem Assessment: Washington, USA)
Sale P, Tavakkoli E, Armstrong R, Wilhelm N, Tang C, Desbiolles J, Malcolm B, O’Leary G, Dean G, Davenport D, Henty S, Hart M (2021) Chapter six – ameliorating dense clay subsoils to increase the yield of rain-fed crops. In ‘Advances in agronomy, Vol. 165’. (Ed. DL Sparks) pp. 249–300. (Academic Press) 10.1016/bs.agron.2020.08.003
Sánchez AC, Kamau HN, Grazioli F, Jones SK (2022) Financial profitability of diversified farming systems: a global meta-analysis. Ecological Economics 201, 107595.
| Crossref | Google Scholar |
Sanderman J, Reseigh J, Wurst M, Young M-A, Austrin J (2015) Impacts of rotational grazing on soil carbon in native grass-based pastures in southern Australia. PLoS ONE 10, e0136157.
| Crossref | Google Scholar |
Santos-Medellin C, Blazewicz SJ, Pett-Ridge J, Firestone MK, Emerson JB (2023) Viral but not bacterial community successional patterns reflect extreme turnover shortly after rewetting dry soils. Nature Ecology & Evolution 7, 1809-1822.
| Crossref | Google Scholar |
Sarker JR, Singh BP, Dougherty WJ, Fang Y, Badgery W, Hoyle FC, Dalal RC, Cowie AL (2018) Impact of agricultural management practices on the nutrient supply potential of soil organic matter under long-term farming systems. Soil and Tillage Research 175, 71-81.
| Crossref | Google Scholar |
Schimel J (2013) Microbes and global carbon. Nature Climate Change 3, 867-868.
| Crossref | Google Scholar |
Schmidt A, Smernik RJ, Mcbeath TM (2012) Measuring organic carbon in Calcarosols: understanding the pitfalls and complications. Soil Research 50(5), 397-405.
| Crossref | Google Scholar |
Schweizer SA (2022) Perspectives from the Fritz-Scheffer Awardee 2021: soil organic matter storage and functions determined by patchy and piled-up arrangements at the microscale. Journal of Plant Nutrition and Soil Science 185(6), 694-706.
| Crossref | Google Scholar |
Shade A, Peter H, Alllison SD, Baho DL, Berga M, Bürgmann H, Huber DH, Langenheder S, Lennon JT, Martiny JBH, Matulich KL, Schmidt TM, Handelsman J (2012) Fundamentals of microbial community resistance and resilience. Frontiers in Microbiology 3, 1-19.
| Crossref | Google Scholar |
Shi A, Cavagnaro TR, Sumby KM, McDonald G, Denton MD, Roy SJ, Schilling RK (2024). Chapter three – impact of agronomic management on the soil microbiome: a Southern Australian dryland broadacre perspective. In ‘Advances in agronomy, Vol. 186’. (Ed. DL Sparks) pp. 113–171. 10.1016/bs.agron.2024.02.008
Shu X, Daniell TJ, Hallett PD, Baggs EM, Mitchell S, Langarica-Fuentes A, Griffiths BS (2021) Role of microbial communities in conferring resistance and resilience of soil carbon and nitrogen cycling following contrasting stresses. European Journal of Soil Biology 104, 103308.
| Crossref | Google Scholar |
Siddique KHM, Belford RK, Tennant D (1990) Root:shoot ratios of old and modern, tall and semi-dwarf wheats in a Mediterranean environment. Plant and Soil 121, 89-98.
| Crossref | Google Scholar |
Sigler WA, Ewing SA, Jones CA, Payn RA, Miller P, Maneta M (2020) Water and nitrate loss from dryland agricultural soils is controlled by management, soils, and weather. Agriculture, Ecosystems & Environment 304, 107158.
| Crossref | Google Scholar |
Simpson RJ, Oberson A, Culvenor RA, Ryan MH, Veneklaas EJ, Lambers H, Lynch JP, Ryan PR, Delhaize E, Smith FA, Smith SE, Harvey PR, Richardson AE (2011) Strategies and agronomic interventions to improve the phosphorus-use efficiency of farming systems. Plant and Soil 349, 89-120.
| Crossref | Google Scholar |
Singh P, Benbi DK (2023) Organic carbon in soils’ fine fraction: thresholds in saturation capacity and its relationship with carbon stabilization. Tropical Ecology 64, 635-654.
| Crossref | Google Scholar |
Singh K, Murphy BW, Marchant BP (2012) Towards cost-effective estimation of soil carbon stocks at the field scale. Soil Research 50(8), 672-684.
| Crossref | Google Scholar |
Singh S, Mayes MA, Shekoofa A, Kivlin SN, Bansal S, Jagadamma S (2021) Soil organic carbon cycling in response to simulated soil moisture variation under field conditions. Scientific Reports 11, 10841.
| Crossref | Google Scholar |
Six J, Ogle SM, Breidt FJ, Conant RT, Mosier AR, Paustian K (2004) The potential to mitigate global warming with no-tillage management is only realized when practised in the long term. Global Change Biology 10(2), 155-160.
| Crossref | Google Scholar |
Six J, Frey SD, Thiet RK, Batten KM (2006) Bacterial and fungal contributions to carbon sequestration in agroecosystems. Soil Science Society of America Journal 70(2), 555-569.
| Crossref | Google Scholar |
Smreczak B, Ukalska-Jaruga A (2021) Dissolved organic matter in agricultural soils. Soil Science Annual 72(1), 132234.
| Crossref | Google Scholar |
Sodhi GPS, Beri V, Benbi DK (2009) Soil aggregation and distribution of carbon and nitrogen in different fractions under long-term application of compost in rice-wheat system. Soil and Tillage Research 103(2), 412-418.
| Crossref | Google Scholar |
Soong JL, Parton WJ, Calderon F, Campbell EE, Cotrufo MF (2015) A new conceptual model on the fate and controls of fresh and pyrolized plant litter decomposition. Biogeochemistry 124, 27-44.
| Crossref | Google Scholar |
Soto RL, Martinez-Mena M, Padilla MC, Vente JD (2021) Restoring soil quality of woody agroecosystems in Mediterranean drylands through regenerative agriculture. Agriculture, Ecosystems & Environment 306, 107191.
| Crossref | Google Scholar |
Sprague SJ, Lilley JM, Bullock MJ, Virgona JM, Kirkegaard JA, Hunt JR, Hopwood MDA, Faulkner MG, Angus JF (2021) Low nitrogen use efficiency of dual-purpose crops: causes and cures. Field Crops Research 267, 108129.
| Crossref | Google Scholar |
Stafford-Smith M, Metternicht G (2021) Governing drylands as global environmental commons. Current Opinion in Environmental Sustainability 48, 115-124.
| Crossref | Google Scholar |
Stanley P, Spertus J, Chiartas J, Stark PB, Bowles T (2023) Valid inferences about soil carbon in heterogeneous landscapes. Geoderma 430, 116323.
| Crossref | Google Scholar |
Stanley PL, Wilson C, Patterson E, Machmuller MB, Cotrufo MF (2024) Ruminating on soil carbon: applying current understanding to inform grazing management. Global Change Biology 30(3), e17223.
| Crossref | Google Scholar |
Steven B, Phillips ML, Belnap J, Gallegos-Graves LV, Kuske CR, Reed SC (2021) Resistance, resilience, and recovery of dryland soil bacterial communities across multiple disturbances. Frontiers in Microbiology 12, 648455.
| Crossref | Google Scholar |
Stewart CE, Paustian K, Conant RT, Plante AF, Six J (2007) Soil carbon saturation: concept, evidence and evaluation. Biogeochemistry 86, 19-31.
| Crossref | Google Scholar |
Stratton AE, Comin JJ, Siddique I, Zak DR, Dambroz Filipini L, Rodrigues Lucas R, Blesh J (2022) Assessing cover crop and intercrop performance along a farm management gradient. Agriculture, Ecosystems & Environment 332, 107925.
| Crossref | Google Scholar |
Su J, Zhang H, Han X, Lv R, Liu L, Jiang Y, Li H, Kuzyakov Y, Wei C (2023) 5300-year-old soil carbon is less primed than young soil organic matter. Global Change Biology 29(1), 260-275.
| Crossref | Google Scholar |
Sun W, Canadell JG, Yu L, Yu L, Zhang W, Smith P, Fischer T, Huang Y (2020) Climate drives global soil carbon sequestration and crop yield changes under conservation agriculture. Global Change Biology 26(6), 3325-3335.
| Crossref | Google Scholar |
Tang B, Rocci KS, Lehmann A, Rillig MC (2023) Nitrogen increases soil organic carbon accrual and alters its functionality. Global Change Biology 29(7), 1971-1983.
| Crossref | Google Scholar |
Tao F, Huang Y, Hungate BA, Manzoni S, Frey SD, Schmidt MWI, Reichstein M, Carvalhais N, Ciais P, Jiang L, Lehmann J, Wang Y-P, Houlton BZ, Ahrens B, Mishra U, Hugelius G, Hocking TD, Lu X, Shi Z, Viatkin K, Vargas R, Yigini Y, Omuto C, Malik AA, Peralta G, Cuevas-Corona R, Di Paolo LE, Luotto I, Liao C, Liang Y-S, Saynes VS, Huang X, Luo Y (2023) Microbial carbon use efficiency promotes global soil carbon storage. Nature 618, 981-985.
| Crossref | Google Scholar |
Taylor LS, Gonzalez A, Essington ME, Lenaghan SC, Stewart CN, Mundorff AZ, Steadman DW, DeBruyn JM (2023a) Soil elemental changes during human decomposition. PLoS ONE 18, e0287094.
| Crossref | Google Scholar |
Taylor K, Samaddar S, Schmidt R, Lundy M, Scow K (2023b) Soil carbon storage and compositional responses of soil microbial communities under perennial grain IWG vs. annual wheat. Soil Biology and Biochemistry 184, 109111.
| Crossref | Google Scholar |
Tejada M, Gonzalez LJ (2007) Influence of organic amendments on soil structure and soil loss under simulated rain. Soil and Tillage Research 93(1), 197-205.
| Crossref | Google Scholar |
Thorup-Kristensen K, Halberg N, Nicolaisen M, Olesen JE, Crews TE, Hinsinger P, Kirkegaard J, Pierret A, Dresbøll DB (2020) Digging deeper for agricultural resources, the value of deep rooting. Trends in Plant Science 25(4), 406-417.
| Crossref | Google Scholar |
Tian LH, Bell LW, Shen YY, Whish JPM (2012) Dual-purpose use of winter wheat in western China: cutting time and nitrogen application effects on phenology, forage production, and grain yield. Crop & Pasture Science 63(6), 520-528.
| Crossref | Google Scholar |
Tiedje JM, Wang F, Manaia CM, Virta M, Sheng H, Ma L, Zhang T, Topp E (2019) Antibiotic resistance genes in the human-impacted environment: a one health perspective. Pedosphere 29, 273-282.
| Crossref | Google Scholar |
Tiemann LK, Grandy AS, Atkinson EE, Marin-Spiotta E, Mcdaniel MD (2015) Crop rotational diversity enhances belowground communities and functions in an agroecosystem. Ecology Letters 18(8), 761-771.
| Crossref | Google Scholar |
Tiessen H, Cuevas E, Chacon P (1994) The role of soil organic matter in sustaining soil fertility. Nature 371, 783-785.
| Crossref | Google Scholar |
Tipping E, Hilton J, James B (1988) Dissolved organic matter in Cumbrian lakes and streams. Freshwater Biology 19(3), 371-378.
| Crossref | Google Scholar |
Tisdall JM, Oades JM (1982) Organic matter and water-stable aggregates in soils. European Journal of Soil Science 33(2), 141-163.
| Crossref | Google Scholar |
Todman LC, Fraser FC, Corstanje R, Deeks LK, Harris JA, Pawlett M, Ritz K, Whitmore AP (2016) Defining and quantifying the resilience of responses to disturbance: a conceptual and modelling approach from soil science. Scientific Reports 6, 28426.
| Crossref | Google Scholar |
UNFCCC (2024) The Paris Agreement. Available at https://unfccc.int/process-and-meetings/the-paris-agreement [accessed 23 January 2024]
Unkovich M, Mcbeath T, Llewellyn R, Hall J, Gupta VVSR, Macdonald LM (2020) Challenges and opportunities for grain farming on sandy soils of semi-arid south and south-eastern Australia. Soil Research 58(4), 323-334.
| Crossref | Google Scholar |
Unkovich M, Mcbeath T, Moodie M, Macdonald LM (2023) High soil strength and cereal crop responses to deeper tillage on sandy soils in a semi-arid environment. Field Crops Research 291, 108792.
| Crossref | Google Scholar |
Van der Berg LJL, Shotbolt L, Ashmore MR (2012) Dissolved organic carbon (DOC) concentrations in UK soils and the influence of soil, vegetation type and seasonality. Science of the Total Environment 427–428, 269-276.
| Crossref | Google Scholar |
Van Herwaarden AF, Farquhar GD, Angus JF, Richards RA, Howe GN (1998) ‘Haying-off’, the negative grain yield response of dryland wheat to nitrogen fertiliser. I. Biomass, grain yield, and water use. Australian Journal of Agricultural Research 49(7), 1067-1082.
| Crossref | Google Scholar |
Vanlauwe B, Descheemaeker K, Giller KE, Huising J, Merckx R, Nziguheba G, Wendt J, Zingore S (2015) Integrated soil fertility management in sub-Saharan Africa: unravelling local adaptation. SOIL 1, 491-508.
| Crossref | Google Scholar |
Verburg K, Pasley HR, Biggs JS, Vogeler I, Wang E, Mielenz H, Snow VO, Smith CJ, Pasut C, Basche AD, He D, Archontoulis SV, Gaydon DS, Huth NI, Holzworth DP, Sharp JM, Cichota R, Khaembah EN, Teixeira EI, Brown HE, Farrell M, Janke CK, Gupta VVSR, Thorburn PJ (2025) Review of APSIM’s soil nitrogen modelling capability for agricultural systems analyses. Agricultural Systems 224, 104213.
| Crossref | Google Scholar |
Viscarra Rossel RA, Walvoort DJJ, Mcbratney AB, Janik LJ, Skjemstad JO (2006) Visible, near infrared, mid infrared or combined diffuse reflectance spectroscopy for simultaneous assessment of various soil properties. Geoderma 131(1–2), 59-75.
| Crossref | Google Scholar |
Viscarra Rossel RA, Webster R, Bui EN, Baldock JA (2014) Baseline map of organic carbon in Australian soil to support national carbon accounting and monitoring under climate change. Global Change Biology 20(9), 2953-2970.
| Crossref | Google Scholar |
Vogel C, Mueller CW, Höschen C, Buegger F, Heister K, Schulz S, Schloter M, Kögel-Knabner I (2014) Submicron structures provide preferential spots for carbon and nitrogen sequestration in soils. Nature Communications 5, 2947.
| Crossref | Google Scholar |
Vohland M, Ludwig B, Seidel M, Hutengs C (2022) Quantification of soil organic carbon at regional scale: Benefits of fusing vis-NIR and MIR diffuse reflectance data are greater for in situ than for laboratory-based modelling approaches. Geoderma 405, 115426.
| Crossref | Google Scholar |
Volsi B, Higashi GE, Bordin I, Telles TS (2022) The diversification of species in crop rotation increases the profitability of grain production systems. Scientific Reports 12, 19849.
| Crossref | Google Scholar |
Walkley AJ, Black IA (1934) An examination of the degtjareff method for determining soil organic matter, and a proposed modification of the chromic acid titration method. Soil Science 37, 29-38.
| Crossref | Google Scholar |
Wang L, Jiao W, Macbean N, Rulli MC, Manzoni S, Vico G, D’Odorico P (2022) Dryland productivity under a changing climate. Nature Climate Change 12, 981-994.
| Crossref | Google Scholar |
Wang W, Chen G-R, Li M-Y, Chen Y, Wang Y, Tao H-Y, Hou H-Z, Rehman MMU, Ashraf M, Song Y, Kavagi L, Wang B-Z, Xiong Y-C (2024) Long-term cereal-legume intercropping accelerates soil organic carbon loss in subsoil of dryland. Resources, Conservation and Recycling 211, 107898.
| Crossref | Google Scholar |
Wang W, Li M-Y, Wen Q-H, Ma Y, Zhang A-M, Rehman MMU, Mo F, Tao H-Y, Ma B-L, Whalen JK, Xiong Y-C (2025) Cereal–legume intercropping stimulates straw decomposition and promotes soil organic carbon stability. Science China Life Sciences 68, 1498-1508.
| Crossref | Google Scholar |
Weisberger D, Nichols V, Liebman M (2019) Does diversifying crop rotations suppress weeds? A meta-analysis. PLoS ONE 14, e0219847.
| Crossref | Google Scholar |
Wendt JW, Hauser S (2013) An equivalent soil mass procedure for monitoring soil organic carbon in multiple soil layers. European Journal of Soil Science 64(1), 58-65.
| Crossref | Google Scholar |
Weng Z, Zwieten LV, Singh BP, Tavakkoli E, Joseph S, Macdonald LM, Rose TJ, Rose MT, Kimber SWL, Morris S, Cozzolino D, Araujo JR, Archanjo BS, Cowie A (2017) Biochar built soil carbon over a decade by stabilizing rhizodeposits. Nature Climate Change 7, 371-376.
| Crossref | Google Scholar |
Whalen ED, Grandy AS, Sokal NW, Keiluweit M, Ernakovich J, Smith RG, Frey SD (2022) Clarifying the evidence for microbial- and plant-derived soil organic matter, and the path toward a more quantitative understanding. Global Change Biology 28, 7167-7185.
| Crossref | Google Scholar |
Whish JPM, Price L, Castor PA (2009) Do spring cover crops rob water and so reduce wheat yields in the northern grain zone of eastern Australia? Crop & Pasture Science 60(6), 517-525.
| Crossref | Google Scholar |
Whitehead DE (2020) Management of grazed landscapes to increase soil carbon stocks in temperate, dryland grasslands. Frontiers in Sustainable Food Systems 4, 585913.
| Crossref | Google Scholar |
Williams A, Kay P, Stirling G, Weng X, Bell L (2022) Impacts of reducing fallow periods on indicators of soil function in subtropical dryland farming systems. Agriculture, Ecosystems & Environment 324, 107727.
| Crossref | Google Scholar |
Wilson GWT, Rice CW, Rillig MC, Springer A, Hartnett DC (2009) Soil aggregation and carbon sequestration are tightly correlated with the abundance of arbuscular mycorrhizal fungi: results from long-term field experiments. Ecology Letters 12(5), 452-461.
| Crossref | Google Scholar |
Wood SA, Bowman M (2021) Large-scale farmer-led experiment demonstrates positive impact of cover crops on multiple soil health indicators. Nature Food 2, 97-103.
| Crossref | Google Scholar |
Wu L, Xu H, Xiao Q, Huang Y, Suleman MM, Zhu P, Kuzyakov Y, Xu X, Xu M, Zhang W (2020) Soil carbon balance by priming differs with single versus repeated addition of glucose and soil fertility level. Soil Biology and Biochemistry 148, 107913.
| Crossref | Google Scholar |
Wu S, Konhauser KO, Chen B, Huang L (2023) ‘Reactive Mineral Sink’ drives soil organic matter dynamics and stabilization. npj Materials Sustainability 1, 3.
| Crossref | Google Scholar |
Wunsch EM, Bell LW, Bell MJ (2017) Can legumes provide greater benefits than millet as a spring cover crop in southern Queensland farming systems? Crop & Pasture Science 68(8), 746-759.
| Crossref | Google Scholar |
Xenopoulos M, Barnes RT, Boodoo KS, Butman D, Catalán N, D’Amario SC, Fasching C, Kothawala DN, Pisani O, Solomon CT, Spencer RGM, Williams CJ, Wilson HF (2021) How humans alter dissolved organic matter composition in freshwater: relevance for the Earth’s biogeochemistry. Biogeochemistry 154, 323-348.
| Crossref | Google Scholar |
Xu X, Hui D, King AW, Song X, Thornton PE, Zhang L (2015) Convergence of microbial assimilations of soil carbon, nitrogen, phosphorus and sulfur in terrestrial ecosystems. Scientific Reports 5, 17445.
| Crossref | Google Scholar |
Zhao G, Bryan BA, King D, Luo Z, Wang E, Yu Q (2015) Sustainable limits to crop residue harvest for bioenergy: maintaining soil carbon in Australia’s agricultural lands. GCB Bioenergy 7(3), 479-487.
| Crossref | Google Scholar |
Zhu S-G, Zhu H, Zhou R, Zhang W, Wang W, Zhou Y-N, Wang B-Z, Yang Y-M, Wang J, Tao H-Y, Xiong Y-C (2023) Intercrop overyielding weakened by high inputs: global meta-analysis with experimental validation. Agriculture, Ecosystems & Environment 342, 108239.
| Crossref | Google Scholar |