

Genetic approaches to enhancing phosphorus-use efficiency (PUE) in crops: challenges and directions
William D. Bovill A C D , Chun Y. Huang A B and Glenn K. McDonald AA Waite Research Institute and School of Agriculture, Food & Wine, The University of Adelaide, Waite Campus, Glen Osmond, SA 5064, Australia.
B Australian Centre for Plant Functional Genomics, The University of Adelaide, Waite Campus, Glen Osmond, SA 5064, Australia.
C Present Address: CSIRO Plant Industry, PO Box 1600, Canberra, ACT 2601, Australia.
D Corresponding author. Email: william.bovill@csiro.au
Crop and Pasture Science 64(3) 179-198 https://doi.org/10.1071/CP13135
Submitted: 19 April 2013 Accepted: 10 July 2013 Published: 6 August 2013
Journal Compilation © CSIRO Publishing 2013 Open Access CC BY-NC-ND
Abstract
Many soils have intrinsically low concentrations of available phosphorus (P), which is a major limitation to crop and pasture growth. Regular applications of P have underpinned agricultural productivity internationally, and fertiliser use now constitutes one of the largest variable input costs to farming. Globally, high-quality reserves of P are being depleted and price increases are likely in the future. In addition, the effects of P pollution on water quality are attracting legislative regulation. Hence, there is a need to improve P-use efficiency (PUE) in farming systems.
Progress in improving PUE has been limited for several reasons, including: inconsistent definitions of PUE, inappropriate phenotyping, incomplete understanding of the controls of P uptake, lack of field validation, and little consideration of genotype × environment interactions that affect the expression of PUE. With greater consideration of these limitations, the powerful array of molecular and genomic tools currently available promises considerable advances in developing more P-efficient crops. Stronger interaction between molecular science and the traditional disciplines of plant breeding, crop physiology, soil science, and agronomy will allow new opportunities to study genetic differences in PUE, bringing P-efficient crops closer to reality.
Introduction
Phosphorus (P) is an essential nutrient for plant growth and development. Due to the diverse functional and structural roles of P in plants, P-use efficiency (PUE) is a complex trait to dissect. In many soils, P deficiency is a major limitation to crop production. Although the total amount of P in soils can be high, plant-available P is often low. It is for this reason that regular applications of fertiliser P are applied to crops. However, only 10–20% of P is used in the year in which it is applied (McLaughlin et al. 1988), although Syers et al. (2008) argue that, when assessed over longer time-frames, use of residual P fertiliser is much higher. In crops to which P fertiliser has been applied, only a small proportion of the P taken up comes from the fertiliser in the year of application, and the majority comes from uptake of native soil P.
Phosphorus-use efficiency has become topical in recent times for several reasons. There were large price increases in 2007–08, and high prices are likely to continue in the future. Coincidentally, the concept of ‘peak P’ has gained some attention in the media, which has drawn attention to the environmental, economic, and social problems that might arise due to limited P reserves (Cordell et al. 2009; Lott et al. 2009). Unlike nitrogen (N), the amount of P available for use in agriculture is finite. Steen (1998) estimated that the depletion of current economically exploitable reserves would occur sometime in the next 60–130 years. Lower quality reserves will be mined when it becomes economical to do so, but the extra extraction and processing costs will result in an increase in the cost of P fertiliser in the future.
Improvements in the efficiency of P nutrition of crops will come from a variety of potential sources, including changes in fertiliser technology, improvements in exploiting soil biology, and better fertiliser management practices, as well as genetic improvement. The widespread realisation that improvements in P nutrition are crucial to the future need to raise global agricultural production has resulted in several recent reviews that have explored these different opportunities (Hinsinger 2001; McNeill and Penfold 2009; Richardson et al. 2009; Ryan et al. 2009; McLaughlin et al. 2011; Simpson et al. 2011). Overall improvement in PUE will ultimately come from integrating a range of different approaches to develop a more efficient farming system. However, one component of this will be varieties that are better able to take up and use soil P, and this aspect will be the focus of the current review.
Economics of phosphorus
Phosphorus fertiliser use in Australia
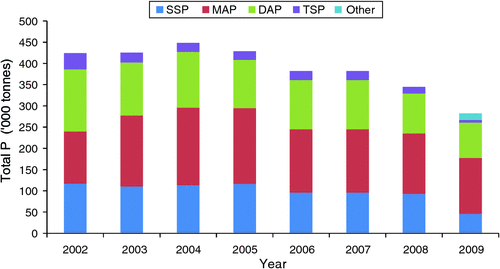
Australian soils are, by world standards, low in plant-available P, and this has been attributed to several factors including the small amount of P in parent rock and the slow rate of weathering (McLaughlin et al. 1992). Consequently, crop production in Australia relies on regular applications of P fertiliser to achieve profitable levels of production. On average ~390 000 tonnes of P per annum, supplied from domestic and imported sources, was used in Australia between 2002 and 2009 (FIFA 2003–2010; Fig. 1). The compound fertilisers, mono-ammonium phosphate (MAP) and di-ammonium phosphate (DAP), have been preferred over single and triple super phosphate. Recently, use of fertiliser P has declined in response to both the large price increases in fertiliser that spiked in 2007–08 and extended drought. The use of novel forms of P, particularly fluid P, increased in 2009, possibly as a response to the reported benefits of fluid P compared with granular P in some farming systems (Holloway et al. 2001; McBeath et al. 2005).
|
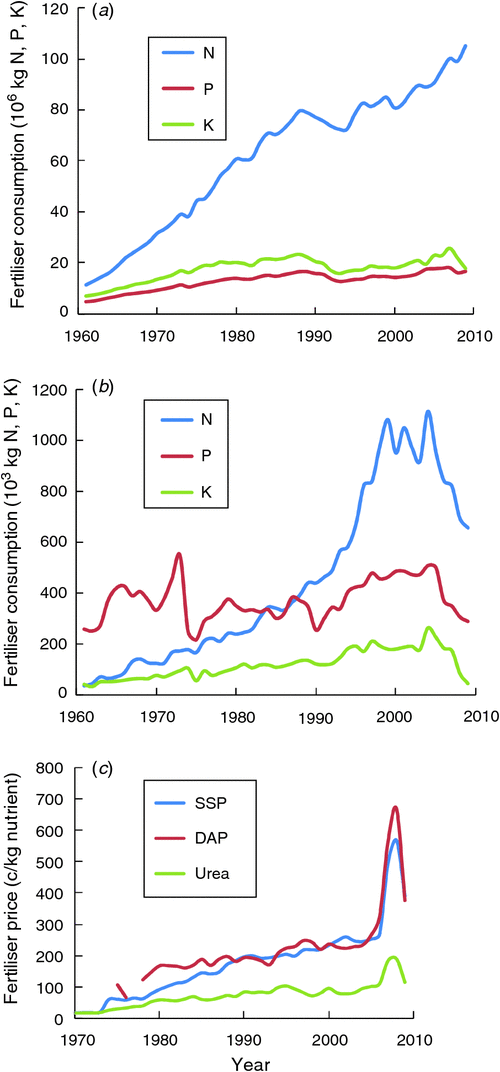
Compared with global consumption of fertiliser, Australian agriculture uses relatively large amounts of P fertiliser, and the P : N ratio is higher (Fig. 2a, b), which probably reflects the generally low P status of Australia’s agricultural land. However, unlike N fertiliser use, total consumption of P fertiliser in Australia has not shown a consistent change over the past 50 years, although there was a gradual increase in consumption in the 1990s and early 2000s. The variations in consumption since 1960 have reflected the costs of fertiliser and the profitability of agricultural production (Fig. 2c). For example, the recent decline in N and P use was associated with the large increase in the price of fertiliser and extended drought.
|
Phosphorus application rates
Phosphorus application rates per crop vary and depend on a range of factors including target yield, paddock fertiliser history and soil test results, soil type, and farm financial constraints. Management of P in cropping systems generally goes through several phases. In the majority of Australian soils where the levels of native soil P are low, soil P reserves are built up to levels that will not limit crop yields. Once this has been achieved, P rates can be reduced to levels that replace the amount of P removed in harvested grain or biomass (replacement levels).
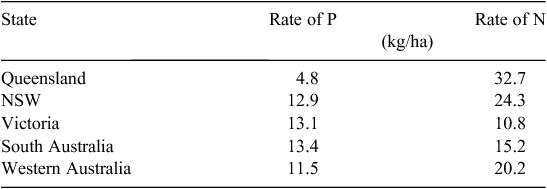
The average rate of P application in the wheat–sheep zone is ~11 kg P/ha, with 21 kg N/ha (ABARE 2010; Table 1). With the exception of Queensland, the average rate of P fertiliser used throughout the wheat–sheep zone does not vary greatly despite the large variation in rainfall and soils. The variability of P rates in the application fertiliser is less than the variability of N rates used. This perhaps suggests a relatively conservative approach to P fertiliser use among Australian grain farmers.
|
Grain P concentration differs among the various grains. Oilseeds have a higher percentage of P in seed than do pulses and cereals (Table 2) and require the highest replacement P levels per tonne of grain harvested. For example, at the paddock level, 26.7 and 14.8 kg P/ha would be exported from cottonseed and soybeans, respectively. Although the oilseeds have the highest concentration of P in harvested product and require higher replacement levels to maintain soil P levels, annual production is a fraction of that of the major cereals, and therefore, most P that is exported from Australian farms is in wheat and barley grain. Improving the PUE of the major winter cereals, and especially wheat, will therefore have the greatest impact on P removal in Australia.

|
Fertiliser as a percentage of farm costs
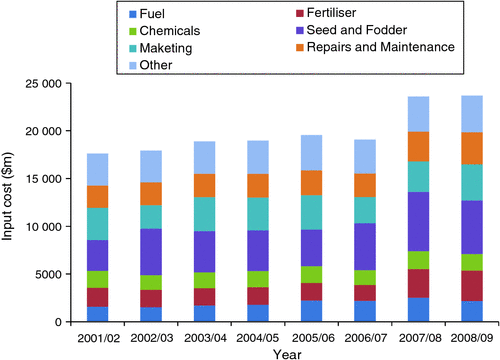
From 2001–02 to 2008–09, fertiliser costs represented an average of 10.6% of total farm input costs (excluding wages, interest paid, other overheads, and depreciation), with an average annual cost of AU$2.14 billon nationally (Fig. 3). Over this time period, the fertiliser input cost ranged from 8.7% in 2005–06 (total value of $1.66 billion) to 13.4% in 2008–09 ($3.17 billion). From 2001–02 to 2007–08, P represented an average of 46.5% of the fertiliser consumed in Australia (N represented 42.8% and potassium (K) 10.8%), when calculated on an elemental basis.
|
Phosphorus requirements of different crops
Improving the PUE of broadleaf crops has two potential advantages: it will allow P fertiliser to be used more efficiently on the individual crop, and it may also benefit the P nutrition of the following crop as there is evidence that cereal crops can benefit from increased levels of residual soil P following some grain legumes (Nuruzzaman et al. 2005a, 2005b). The ability to use soil P and responses to P fertiliser can vary among different crop species, although the extent of the difference is influenced by soil type (Bolland et al. 1999; Brennan and Bolland 2001; Bolland and Brennan 2008; Vu et al. 2010).
Canola (Brassica napus L.) generally shows large responses to P. In Western Australia, pot and field studies showed that the response to P in canola is greater than that of wheat (Triticum aestivum L.) and that it required up to 50% less P than wheat to reach 90% of maximum yield (Bolland et al. 1999; Brennan and Bolland 2001; Bolland and Brennan 2008). Canola also took up more P than wheat at a given amount a P fertiliser, suggesting that canola roots have a greater ability to utilise soil P, at least in the soils used in those studies. Vu et al. (2010) also found canola seedlings to be very responsive to P in a range of soils from Victoria. In northern New South Wales, canola often shows greater responses to P than wheat, and P nutrition is considered more critical in canola than in wheat (Serafin et al. 2005). The greater responsiveness of canola to P means that the general fertiliser recommendation is for similar or higher rates of P to be applied to canola compared with wheat despite it yielding less than wheat (Mullen and Gammie 2002; Duff et al. 2006).
Grain legumes show a range in responsiveness to P. White lupin (Lupinus albus L.) and chickpea (Cicer arietinum L.) have been reported to be less responsive to P fertiliser than wheat, which has been attributed to their ability to alter the pH of the rhizosphere (Bolland et al. 1999). Studies have shown that compared with wheat, chickpea, field pea (Pisum sativum L.), and lupin (Lupinus spp.) have the ability to acidify the rhizosphere by releasing organic acids, which has been associated with relatively better growth at low available P (Pearse et al. 2006). Faba bean (Vicia faba L.) has shown a variable response to P but it has been considered the grain legume that is most responsive to P when grown on neutral–acid soils of Western Australia (Bolland et al. 1999). Using three different soil types found in Victoria (Chromosol, Vertisol, and Calcarosol), Vu et al. (2010) also found that chickpea was more effective in using native soil P than wheat and canola but that wheat was quite efficient in using freshly applied P fertiliser. The various results of controlled environment and field studies suggest that faba bean and field pea are the most P-responsive legumes, while chickpea and white lupin are less responsive. These differences are consistent with the reported differences in seed P (Table 2). There is little local information on responses in lentil (Lens culinaris Medik.).
What is PUE?
Progress on genetic improvement in PUE is hampered because there is no generally agreed way of defining PUE. Numerous criteria for PUE are found in the literature (Table 3). To cloud the problem further, different terms are often used even though they are calculated in the same way. For example, Jones et al. (1989) and Ozturk et al. (2005) report on varietal differences in P efficiency ratio (as a criterion for determining PUE). Jones et al. (1989) calculate the ratio as grain yield per unit P uptake, whereas Ozturk et al. (2005) calculate the ratio as relative shoot growth. Manske et al. (2002) report varietal differences in P utilisation efficiency, using the same calculation that Jones et al. (1989) use for determining the P efficiency ratio. There are other discrepancies in the terminology (see Table 3), and future work on PUE would benefit from more consistent use of terminology and definitions.

|
An assessment of three different approaches
Ortiz-Monasterio et al. (2001) suggested that the definition and components of N-use efficiency (NUE) reported by Moll et al. (1982) should be adopted in studies of PUE; Manske et al. (2001) used this when classifying P-efficient wheat varieties. In this approach, PUE, which is defined as grain yield per unit of nutrient supplied (GY/Ns), is the product of uptake efficiency (Nt/Ns) and utilisation efficiency (GY/Nt) (Eqn 1):

where GY is grain yield, Ns is nutrient supplied, and Nt is total above-ground plant nutrient. A problem with using this definition of PUE to select for genotypes that are more efficient is that Ns will be the same across all genotypes, and hence genotypes with the highest PUE will be the highest yielding genotypes; i.e. one would simply be selecting for yield potential. If this definition were applied to a collection of land races as well as modern genotypes screened in the field, the land races (that may well be adapted to low P soils) would be classed as P-inefficient compared with modern genotypes, due to their lower yield potential. Clearly, yield potential confounds assessment of efficiency when this definition is used.
Agronomic PUE refers to the increase in yield of a variety following the addition of P fertiliser. Assessed as the difference in yield between fertilised and unfertilised treatments, divided by the difference in nutrient supplied in each of the treatments (Hammond et al. 2009), agronomic PUE is a measurement of the level of responsiveness to P. Thus, a variety with high agronomic efficiency will be one that has a high yield increase in response to increased P application. A variety with a high agronomic efficiency may result in higher yields at the levels of P currently recommended and may also show a large yield response with further additions of P. The question is whether this is the most appropriate variety if the goal of improving PUE is to reduce inputs of P fertiliser.
Graham (1984) defines the nutrient efficiency of a genotype as the ability to produce a high yield in soil that is limiting in that element for a standard genotype. This is essentially the same as the definition proposed by Moll et al. (1982), but importantly, it differs in that the relative yield of a genotype in limiting compared with non-limiting nutrient conditions is used to determine nutrient-use efficiency. The approach of Graham (1984) when screening for improved nutrient efficiency in the field has been to evaluate relative yield in paired plots (Eqn 2):

where GY– is yield of plots without nutrient, and GY+ is yield of plots with supplied nutrient. An efficient genotype is one that possesses a high relative yield.
Table 4 shows that the use of the various definitions can result in very different selection outcomes. The raw yield data come directly from the study of Moll et al. (1982) on NUE in maize (Zea mays L.), but are equally applicable to PUE. Using the Moll et al. (1982) definition (NUE = GY/Ns), hybrid 7 is the most efficient at low nutrient supply, whereas hybrid 6 is the most efficient at high nutrient supply. Hybrids 7 or 6 were classed as the most varieties with greatest NUE because they were the highest yielding hybrids at each of the two fertiliser levels. Hybrid 2 has the highest agronomic efficiency. Hybrid 7 has the highest relative yield and the lowest agronomic efficiency, and it would be the variety of choice when aiming to reduce nutrient inputs. Clearly, the definition used has a strong impact on selection outcomes.
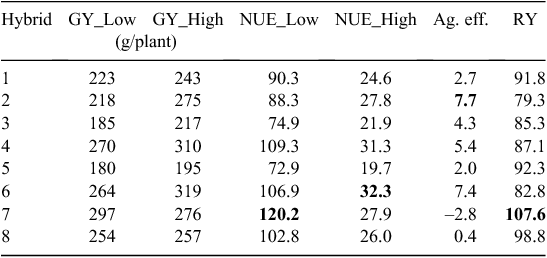
|
An economic perspective on PUE
The farm business is driven by profit, and so rather than focusing on the biological optimum P rate, the economic optimum P rate should also be considered (Fig. 4). In its simplest terms, the economic optimum fertiliser rate is when the marginal profit is zero, that is, the increase in return from adding fertiliser matches the additional cost of the fertiliser (Fig. 4a). The economic optimum P rate will be lower than the biological optimum rate. Unless there are changes in the responsiveness of varieties to P, the predicted increases in the price of fertiliser will result in the economic optimum P rate becoming progressively lower (Fig. 4b), which will eventually lead to lower yields as the economic optimum falls well below the biological optimum yield. There are several responses possible to the increased cost of fertiliser: (i) increase the responsiveness of a variety so that yield is not reduced at the lower level of P, and (ii) increase the ability to exploit native soil P (i.e. the yield at 0 kg P/ha; Fig. 4c). In the former case we are changing the agronomic efficiency, and in the latter the relative yield or the genetic PUE. Improving yield at 0 kg P/ha could result from plants accessing P from more pools, though over time, P removed by crops would need to be replaced. Improvements may also be achieved by increasing the yield potential of varieties without selecting directly for PUE-related traits (Fig. 4c). This simple analysis illustrates that there may not be a single ‘best’ way of increasing P efficiency of varieties and that several approaches need to be integrated.
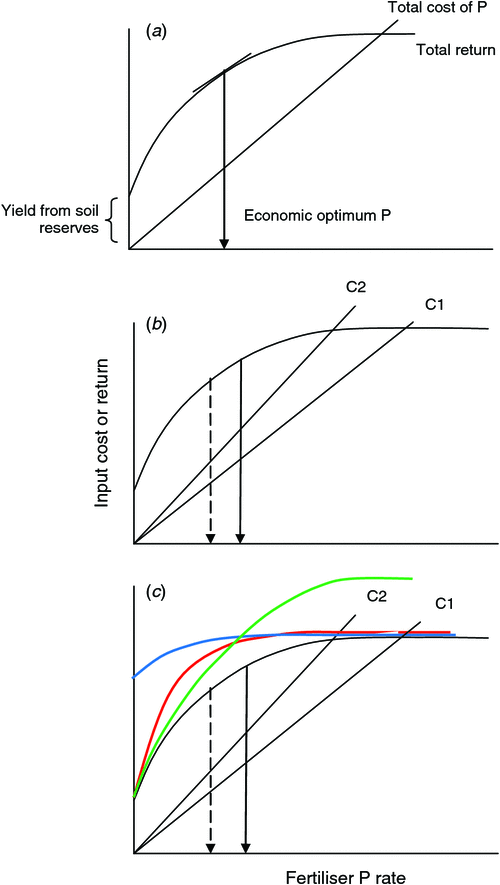
|
Defining the goal
In the face of increasing costs of P fertiliser inputs, genetic improvements in P efficiency can occur by developing varieties that either allow farmers to reduce fertiliser input costs without compromising yields, or by maintaining current rates of fertiliser but achieving equivalent or higher yields. The former case is indicated by relatively high yields at low or zero rates of fertiliser P, which may be achieved by increasing yield potential generally (in effect passively increasing P efficiency) or by selecting for a greater capacity to exploit native soil P. Achieving equivalent or higher yields with current rates of P will be achieved by an ability to respond to applications of P fertiliser. Again, this may occur passively by improving overall yield potential or by enhancing the ability of a variety to take up and use fertiliser P.
Plant breeders aim to produce high-yielding varieties that have broad adaptation, and genetic improvements in PUE should also show an ability to improve P efficiency across a range of conditions. The majority of grain farms in Australia have reasonable levels of soil P, and it is likely that farmers will aim to maintain this level of fertility into the future. Selecting for P uptake under severely P-limited conditions is perhaps unrealistic for these farms unless it can be demonstrated that traits that are suited to severely P-deficient soils would also be useful at higher levels of fertility. There are also situations where available soil P is low enough to limit productivity even with regular additions of P fertiliser. These include calcareous and acidic soils in which applied P is tightly bound in chemical forms that restrict its availability to plants. In this case, selecting for P uptake under low available soil P would be appropriate. A problem with much of the past work is that there has been no systematic evaluation of the usefulness of putative traits for PUE at different levels of P availability; much of the screening work has used severely P-deficient soil. Therefore, it is unclear whether characteristics that contribute to enhanced growth and P uptake under severe P stress will be useful at the lower levels of P stress likely to be encountered in Australian grain production. Examining genotypes across several sites and seasons where the level of P stress varies can provide some insight into this issue.
Ideally, a variety should show a high yield under low P as well as an ability to respond to P when P rates are increased or if there are high levels of soil P. Therefore, rather than a single criterion for PUE, it may be better to characterise genotypes for their relative response to P (i.e. Graham’s nutrient-use efficiency) as well as the yield of the genotype at low P. Using these two measures allows the yield potential of the variety and its responsiveness to be considered simultaneously.
Phosphorus uptake, utilisation, and signalling
Mechanisms of P uptake
The complexity of P nutrition of plants arises because the availability and uptake of P depends on the interaction between soil, plant, and microbial processes. The relative importance of different processes and the effectiveness of different plant characteristics are likely to vary according to soil type, cropping history, and climate and weather. Therefore, targeting one specific mechanism to improve P uptake by plants may provide limited success under commercial conditions. This is not to say that gains cannot be made, as there have been reported improvements in PUE in several crops in China (Yan et al. 2006).
Root system architecture
To maximise P acquisition in low P conditions, plants change root growth and development by promoting the formation of a shallow, highly branched root system through a reduction of primary root growth, and an increase in adventitious roots and lateral root density, as well as the development of more and longer root hairs. Architectural root traits associated with enhanced topsoil foraging include shallower growth angles of axial roots, a greater number of adventitious axial roots, and greater dispersion of lateral roots (Péret et al. 2009). Functional characterisation of the Pup1 quantitative trait locus (QTL) in rice (Oryza sativa L.) has led to the identification of a protein kinase gene (PSTOL1) which increases adventitious root development under low P conditions, conferring P-deficiency tolerance (Gamuyao et al. 2012). Several genes controlling lateral root development have been identified in Arabidopsis (Péret et al. 2009) and rice (Coudert et al. 2010). At least six root QTLs have been identified in maize, and are good candidates for further evaluation (Hund et al. 2011). Genetic variation in root hair length, and loci controlling barley (Hordeum vulgare L.) root hair formation, have also been identified (Gahoonia and Nielsen 1997; Szarejko et al. 2005).
Symbiosis with arbuscular mycorrhizal fungi
Arbuscular mycorrhizal (AM) associations are another avenue by which most plants can increase uptake of P. This symbiotic relationship between plant and fungus increases the ability of the plant to access P (as well as other nutrients such as zinc and copper; Marschener 1998), while in return the fungus receives carbohydrates from the host plant. In AM associations, two pathways for plant P uptake exist: the direct pathway (P uptake by roots) and the AM fungal pathway (Smith and Smith 2011). By using the AM pathway, the soil volume from which P can be accessed is increased. In addition, there is some evidence that the fungal hyphae can access pools of P that are unavailable to plant roots (Jayachandran et al. 1992; Koide and Kabir 2000). Genetic variation for AM associations has been demonstrated in wheat, barley, and other plant species (Baon et al. 1993; Zhu et al. 2001; Jakobsen et al. 2005; An et al. 2010). This offers the potential to select for AM responsiveness. Given that the AM pathway can be responsible for the majority of P that is taken up by plants, the ability to select for responsiveness warrants further investigation. However, the positive effect of mycorrhizal colonisation decreases as soil P levels increase. Further work would benefit from assessment of colonisation at P levels that are representative of agricultural soils, as results from some field studies suggest that high colonisation by AM is unimportant for crop productivity (Ryan et al. 2002; Ryan and Angus 2003). Nonetheless, understanding the physiological and genetic controls of the AM–plant interaction may enable root infection to occur even at high soil P concentrations, which may enhance P uptake over a wider range of soil P concentrations than occurs currently.
Exudates for mobilisation and scavenging inorganic P (Pi) from soil
One of the ways in which plants respond to low supplies in soil P is to release several root exudates that increase the availability of inorganic and organic forms of P into the rhizosphere (Rengel and Marschner 2005; George and Richardson 2008). The exudates include several organic acid anions, protons that increase availability of the sparingly soluble forms of P, and enzymes such as phosphatases that increase the P availability from organic P.
Several plant species, including some legumes and rapeseed/canola, can increase the exudation of organic acid anions (carboxylates) in response to low available P (Pearse et al. 2006; Kirkby and Johnston 2008) or aluminium toxicity (Ligaba et al. 2004). The carboxylates include citrate, malate, malonate, and oxalate. The anions chelate the metal ions that bind with P and displace the P from the soil matrix (Ca, Fe, and Al complexes). Several P-efficient wheat varieties from China, for example, were able to excrete more citric and malic acid than P-inefficient varieties (Yan et al. 2006).
While there are examples of species, such as white lupin, that have the capacity to release large quantities of carboxylates, there is still some debate as to whether the rates of exudation of the organic anions are large enough to substantially increase P uptake, and there is uncertainty as to the level of improvement that could be achieved (George and Richardson 2008; Kirkby and Johnston 2008). Overexpression of genes that are involved in organic anion synthesis in roots is a possible means of increasing the concentrations of exudates in the rhizosphere. The bacterial citrate synthase gene has been examined in tobacco (de la Fuente et al. 1997; Delhaize et al. 2001) with inconsistent results, and even with overexpression of the gene, there is still concern that the current levels of exudation are insufficient to substantially increase P uptake (George and Richardson 2008). Moreover, there is relatively little information on the level of genetic variation in exudate release among varieties of the major grain crops. A further complicating factor is that the organic anions provide a substrate for microbial growth and so there may only be transitory increases in their concentrations in the rhizosphere. In addition, the complex relationships between root exudates, soil microbes, and nematodes are not well understood (Bais et al. 2006).
Plants can also exploit organic P in the rhizosphere. Extracellular phosphatases can be released by plant roots in response to P starvation. Although there is genetic variation in phosphatase release in wheat, there is little association with growth and P nutrition when tested over a range of soil types (Rengel and Marschner 2005; George and Richardson 2008). The complexity of the rhizosphere environment may be a contributor to this lack of a clear link between phosphatase release and P uptake. Kirkby and Johnston (2008) point out that we still do not understand the interactions between the enzymes released into the rhizosphere and the rhizosphere microorganisms involved in the turnover of organic P.
The other form of organic P in soils is phytate (myo-inositol hexaphosphate), which is broken down by phytase. Plant roots release little phytase and its release depends on microbial activity. However, genetic variation in the use of phytate as the source of P has been demonstrated in wheat (Osborne and Rengel 2002b). Overexpression of microbial phytase genes in transgenic plants has shown improvements in growth and P uptake under controlled conditions, but these effects are not observed when tested in a range of soils (George and Richardson 2008). The variability in the importance of phytate as a source of organic P in soils, interactions with soil microorganisms, and the complex soil chemical environment may be important factors limiting the effect.
High-affinity Pi transporters and mycorrhizal-specific Pi transporters
The PHT1 gene family is the primary group of Pi transporters for Pi influx from soil solution to root cells. There are four closely related barley PHT1 genes (HvPHT1;1, HvPHT1;2, HvPHT1;9, and HvPHT1;10) identified so far in the barley genome, and closely related wheat homologues have also been identified (Huang et al. 2011). These four barley PHT1 paralogues are induced by P deficiency, and are similar in expression patterns in the root tip and elongation zone (Preuss et al. 2011; C. Y. Huang, N. Shirley, P. Langridge, unpubl. data). HvPHT1;1 is localised in the plasma membrane and is a high-affinity Pi transporter with a Km value of 1.9 µm (Preuss et al. 2011).
The establishment of symbiosis with AM fungi is a common adaptive P-starvation response, in which plants increase P acquisition assisted by the symbiotic fungi (Smith and Smith 2011). Mycorrhizal-specific Pi transporters (expressed in cells containing arbuscules) have been identified in several species. One mycorrhizal-specific barley Pi transporter (HvPHT1;8) and a homologue of wheat (TaPHT1;8) have been reported so far (Glassop et al. 2005), and two additional mycorrhizal-specific barley Pi transporters have also been identified recently (C. Y. Huang and E. J. Grace, unpubl. data). The functional roles of the two new mycorrhizal-specific barley Pi transporters in P acquisition remain to be revealed. Two mycorrhizal-specific Pi transporters have been identified in rice (Yang et al. 2012) and one in barrel medic (Medicago truncatula Gaertn.) (Javot et al. 2007). There is little information available for PHT1 or mycorrhizal-specific Pi transporters in canola.
Loading of Pi to the xylem
Transport of Pi from the root cortex into the xylem is crucial for Pi uptake. PHO1 plays a significant role in root-to-shoot translocation of Pi (Poirier et al. 1991). Recent studies show that PHO1 mediates Pi efflux from the cell and reduced Pi concentration in the vacuole (Rouached et al. 2011; Stefanovic et al. 2011). Export of Pi mediated by ectopic expression of PHO1 in plant cells is associated with Golgi and trans-Golgi networks (Arpat et al. 2012). The corresponding protein contains six potential trans-membrane spanning domains and a long N-terminal hydrophilic domain. PHO1 shows no homology to characterised solute transporters, including the family of plant PHT1 H+/Pi co-transporters, but contains two domains, named SPX and EXS, that have been identified in some Saccharomyces cerevisiae proteins that are involved in Pi transport or sensing, and in protein sorting to endomembranes (Secco et al. 2012).
Utilisation of P
Metabolic adaptation to P-deficient conditions
Under P starvation, plants increase P recycling from intracellular organic P compounds by increasing activities of acid phosphatases, purple acid phosphatases, and RNase, and by scavenging Pi by replacing membrane phospholipids with galacto- and sulfo-lipids. Plants also use metabolic bypasses in glycolysis, increased accumulation of starch deposition, and anthocyanins in shoots (Wasaki et al. 2003; Plaxton and Tran 2011).
Low-affinity transporters and organelle Pi transporters
Remobilisation of Pi within a plant is critical for sustaining growth and seed production under external Pi fluctuation. Some of the PHT1 transporters such as HvPHT1;6 showed a linear transport activity over a concentration range of 5–30 mm, suggestive of a low-affinity Pi transporter (Preuss et al. 2010). HvPHT1;6 is highly expressed in old leaves, and its transcript is abundant in phloem tissue (Rae et al. 2003). The expression of HvPHT1;6 in both shoots and roots is upregulated by P deficiency (Huang et al. 2008; Huang et al. 2011). These data suggest that HvPHT1;6 plays a major role in Pi remobilisation.
Functional AtPHT1;5 showed Pi-deficiency-induced expression, specifically in the phloem cells of older leaves and other sink tissues in Arabidopsis (Mudge et al. 2002). Loss-of-function mutants of AtPHT1;5 and transgenic lines overexpressing this gene in Arabidopsis were developed to demonstrate its role in Pi mobilisation between source and sink under different Pi conditions (Nagarajan et al. 2011). Little is known about the roles of PHT1 genes in P loading into grains in cereals such as wheat and barley. Grains contain ~50% or more of the P taken up by the plant, and understanding control of P loading into grain is important for future work on PUE. Characterisation of Pi-transporter genes responsible for P remobilisation from source tissues and loading into grains will be important in future studies.
The PHT2 family genes in Arabidopsis and potato have been functionally characterised (Daram et al. 1999; Rausch and Bucher 2002). For example, AtPHT2;1 is an H+/Pi symporter, and has a high apparent Km for Pi (0.4 mm), suggesting that the members of the PHT2 family are low-affinity Pi transporters (Daram et al. 1999). PHT2 gene expression is localised in plastids (Versaw and Harrison 2002; Rausch et al. 2004). PHT3 family genes are mitochondrial Pi transporters (Rausch and Bucher 2002). PHT4 family proteins are localised in either the plastid envelope or the Golgi apparatus (Guo et al. 2008), indicating that PHT2, PHT3, and PHT4 are involved in Pi flux in the organelles. There have been no reports on functional roles of these three gene families of cereals in PUE.
There were other gene families involved in the transport of small metabolites containing P such as the Glycerol-3-phosphate permease (G3Pp) family (Ramaiah et al. 2011). The G3Pp genes are not only involved in Pi homeostasis, but also alter root development (Ramaiah et al. 2011).
Micro RNAs (miRNAs) and targets
The miRNAs function as riboregulators for many downstream genes involved in P starvation responses (PSR) and also act as signal-transmitting molecules for systemic adaptive responses (Kuo and Chiou 2011). The MiR399 plays an important role in maintaining Pi homeostasis at the level of Pi acquisition, distribution, and remobilisation. Upon P deficiency, miR399 acts as a positive regulator to promote Pi uptake and root-to-shoot translocation of Pi (Kuo and Chiou 2011). The target gene of miR399 encodes a ubiquitin-conjugating E2 enzyme, PHO2/UBC24, which functions mainly in the roots to regulate Pi acquisition and root-to-shoot Pi translocation (Aung et al. 2006). The Arabidopsis pho2 mutant overaccumulates phosphate in shoots (Delhaize and Randall 1995). Not only does miR399 function as a riboregulator that regulates many downstream genes involved in PSR but also as a signal-transmitting molecule that coordinates systemic adaptive responses in roots. MiR399 is present in the phloem sap and transmitted to roots as a systemic signal for regulation of root responses to P deficiency. Overexpression of miR399 in Arabidopsis and rice leads to an excessive increase in P accumulation in the shoots when P is available (Aung et al. 2006; Hu et al. 2011).
Another miRNA, miR827, is also upregulated in shoots and roots by P starvation (Lin et al. 2010). One AtmiR827 target is the gene encoding a ubiquitin E3 ligase, which consists of an N-terminal SPX domain and a C-terminal RING domain ligase (Hsieh et al. 2009; Pant et al. 2009). This E3 ligase mutant displays an early senescence phenotype of nitrogen limitation adaptation (nla), and increases Pi uptake and Pi content, especially under low-nitrate and high-phosphate availability, relative to wild-type plants (Kant et al. 2011). It would be of interest to reveal the underlying mechanism of the antagonistic crosstalk between nitrogen and phosphate homeostasis via miR827.
It is noteworthy that overexpression of miR827 has been shown to increase drought tolerance in maize (Patent: US 2009/0165168), but the underlying mechanism for the improvement of drought tolerance by overexpression of miR827 has not been reported. Although miR827 has been identified in barley (Schreiber et al. 2011) little is known about the involvement of miR827 in P nutrition and drought tolerance in wheat and barley. Overexpression of OsmiR827 did not show any increase in Pi accumulation in rice (Lin et al. 2010).
Phosphorus starvation can also lead to downregulation of miRNAs such as miR169, miR395, and miR398 by. These miRNAs are also responsive to other nutrients and abiotic stresses such as N, copper, sulfur, and drought, suggesting stress that signal transduction pathways interconnect with each other (Kuo and Chiou 2011). Elucidation of their specific role in P-metabolism requires further work.
In addition, non-coding RNAs, such as IPS genes contain a conserved motif, which could form an RNA duplex with miR399. They can regulate miR399 cleavage activity (Franco-Zorrilla et al. 2007). Two HvIPS genes show a different binding affinity to HvmiR399 members, and correlate with expression levels of low-affinity HvPHT1 genes (Huang et al. 2011). Genetic variation in the expression of low-affinity Pi transporters has been found to be positively correlated with the expression of the HvIPS genes (Huang et al. 2011). Further studies are required on the relationship between the expression of low-affinity Pi transporters/IPS genes and Pi remobilisation.
Uptake v. utilisation
There is debate about the relative importance of P acquisition compared with P utilisation in improving PUE (Wang et al. 2010; Rose and Wissuwa 2012). This is perhaps surprising given that the contribution of each of these processes to PUE will depend upon soil type and P availability. It seems reasonable to expect that on a soil with low levels of plant-available P, acquisition of P is likely to be more important, but in a soil with high P availability where P acquisition is not limited, P utilisation is likely to dominate PUE. Experimental evidence for these observations has been provided by Manske et al.(2001) in wheat; in a calcareous soil at high P, utilisation of P was more important, whereas in an acid soil with high P-fixing ability, P acquisition was more important in both low- and high-P treatments.
Biomass production plays a large part in the calculation of both P acquisition and P utilisation. From screening a diverse Brassica spp. germplasm set, Hammond et al. (2009) suggested that there was a greater influence of shoot dry matter production on measures of PUE, in contrast to more efficient accumulation or internal use of P within the plant. Further, studies have shown that P acquisition and utilisation are negatively correlated (Wissuwa et al. 1998; Su et al. 2009), with those authors suggesting that improvement of a genotype for both P acquisition and utilisation will be difficult to achieve. Rose et al. (2011) provide further evidence for this phenomenon in rice, concluding that P acquisition and P utilisation are unavoidably linked in traditional soil- and field-based screening. Using a hydroponic-based method, Rose et al. (2011) reported that the linkage between P acquisition efficiency and P utilisation can be broken. It remains to be seen if this new approach will be of practical use given the reported poor relationship between hydroponic P experiments and pot- and field-based assays (Hayes et al. 2004).
Phosphorus signalling
There are several components involved in the global regulation of PSR. Transcription factors play a central role in regulation, whereas miRNAs act at the post-transcriptional level. Proteins involved in protein ubiquitination also play an important role in Pi signalling at the post-translational level. For example, PHO2 is an E2 conjugase, which is the target of miR399 as described above, whereas AtNLA encodes an E3 ligase (Kant et al. 2011). Many transcription factors have been identified, in which MYB transcription factors, such as AtPHR1 and OsPHR2, are primary regulators in PSR. The orthologues of AtPHR1 have not yet been reported for other crops. These MYB transcription factors regulate genes involved in the root system, Pi uptake and P remobilisation, and P mobilisation in soil (Rubio et al. 2001). Sugar signalling is an important regulatory pathway in plant growth and development as well as in metabolic activities (Rolland et al. 2006). Sugar signalling is also involved in plant responses to Pi starvation (Hammond and White 2008). Sucrose acts at the molecular level to influence multiple P starvation responses. Lei et al. (2011) provide genetic and genomic evidence to show that sucrose is a global regulator of plant responses to Pi starvation. In addition, plant hormones play an important role in PSR (Chiou and Lin 2011). The network that consists of these regulatory components and downstream genes coordinates sophisticated responses to Pi starvation.
Genetics of PUE
Relatively few studies have identified QTLs for PUE in the crop species that are the target of this review (Table 5). Generally, the process has been to measure traits of interest (e.g. biomass production and shoot P concentration) and report upon QTLs that are detected for these traits under both limiting and non-limiting P conditions. Phosphorus uptake or utilisation efficiency is then calculated from these component traits, and those QTLs reported. Given the central role of biomass and yield in most definitions of PUE, it is not surprising that the QTLs for biomass and yield often collocate with QTLs for P uptake and/or P utilisation efficiency.

|
For example, in wheat (Su et al. 2006, 2009), Brassica sp. (Hammond et al. 2009; Yang et al. 2010, 2011), soybean (Glycine max (L.) Merr.) (Zhang et al. 2009), and rice (Wissuwa et al. 1998), QTLs for P uptake efficiency co-located with QTLs for biomass production. This is because the correlation between biomass production and shoot P uptake is often extremely high (up to r = 0.96 in rice; Wissuwa et al. 1998), and shows that biomass production drives calculation of P uptake. Many studies show that QTLs detected for P uptake are co-located with QTLs for P-utilisation efficiency, although alleles at the QTL that lead to increased P uptake reduce P utilisation (Wissuwa et al. 1998; Su et al. 2006, 2009; Zhang et al. 2009). The authors suggest that it will be therefore be difficult to improve both traits simultaneously. It is not clear if this negative correlation is an artefact of the definitions that are used for P uptake and P-utilisation efficiency. If a genotype possesses high P-uptake efficiency (driven by biomass production), then P-utilisation efficiency (calculated as biomass production per unit P uptake) will be low. This further highlights the problem with commonly used definitions of PUE (see previous section: What is PUE?).
However, few authors have attempted to overcome this issue by assessing relative yield, and detecting QTLs for relative yield. Yang et al. (2010), in a study assessing the relationship between QTLs for root traits and P uptake in Brassica napus, found that QTLs for P uptake and biomass production were linked. In a later report on the same population, Yang et al. (2011) change their definition of PUE to relative yield and state that ‘these QTL were demonstrated to represent the true QTL for P efficiency’ (Yang et al. 2011). Unfortunately, differences and similarities in the location of the QTL between the two approaches cannot be assessed due to the format in which the maps have been provided in the papers (Yang et al. 2010, 2011). It would be worthwhile for previous studies, with data already available, to repeat the QTL analysis based upon relative yield, and compare the results with those previously published. In China, several P-efficient wheat varieties were identified by extensive screening of germplasm at high and low soil-P; the sole criterion for efficiency was relative yield, irrespective of the yield potential of the variety (Davies et al. 2002).
There have been several attempts to link QTLs for root development/architectural traits with QTLs for P uptake or utilisation efficiency (Liao et al. 2004; Beebe et al. 2006; Ochoa et al. 2006; Cichy et al. 2009; Hammond et al. 2009; Zhang et al. 2009; Liang et al. 2010; Yang et al. 2010, 2011). Common bean (Phaseolus vulgaris L.) has been the most widely studied species in this respect. In a population developed from a cross between Andean and Mesoamerican parents, Beebe et al. (2006) and Liao et al. (2004) analysed the same field dataset; Beebe et al. (2006) compared field and hydroponic results, whereas Liao et al. (2004) compared field and growth pouch results. Liao et al. (2004) found that three of the QTLs that contributed to P-acquisition efficiency (Pup4.1, Pup7.1, and Pup11.1) in the field were ‘closely linked’ (although the distance between QTLs appears to be ~30–40 cM in some cases) to some of the QTLs that were identified for root gravitropism in growth pouches, and concluded that root gravitropism contributes to P efficiency. Beebe et al. (2006) only reported two (Pup4.1 and Pup10.1) of the six QTLs for P-acquisition efficiency in the field found by Liao et al. (2004), using the same dataset. Nonetheless, some of the QTLs for root architectural traits identified in the field and in hydroponics were associated with P acquisition QTLs (Beebe et al. 2006). Ochoa et al. (2006) examined adventitious root formation in a related population (G2333/GI19839), but the QTLs for adventitious root formation were not located in the same region as the QTLs for P-acquisition efficiency as reported in Beebe et al. (2006) and Liao et al. (2004). Cichy et al. (2009) also could not find any relationship between root traits and P uptake in an Andean/Andean bean population. These reports show the contrasting results for the relationship between root architecture and PUE.
In some studies, developmental genes have been shown to be involved with PUE, although the reasons for this are unclear. In wheat, Su et al. (2006) identified a range of QTLs associated with P-deficiency tolerance, with three main clusters located on chromosomes 4B, 5A, and 5D. Interestingly, the 5A and 5D QTLs were associated with the major vernalisation genes, Vrn-A1 and Vrn-D1. In a later study on a different wheat population, the effect of Vrn-A1 and Vrn-D1 were not detected, but it is unclear whether this population was segregating at these loci (Su et al. 2006); it would be useful to genotype the Vrn alleles of these parents. In barley, George et al. (2011) attempted to identify QTLs for shoot P concentration in an association mapping panel of 120 barley genotypes comprising 56 winter and 64 spring types. No common QTLs could be detected between winter and spring types. However, the associations only just exceeded the threshold for detection, and George et al. (2011) attribute this to the relatively small population sizes that were used for the study and/or limited genetic variation. QTLs for PUE in common bean were linked with the fin gene, which regulates determinism in this species (Cichy et al. 2009). Whether or not these developmental associations are a result of the effect of P deficiency on development, or play a central role in PUE, deserves more attention.
There are not enough studies of different populations within a species to make a strong case for the likely effectiveness of any particular QTLs that have been detected. A further issue arises from the population sizes that are used in many studies. The effect of population size on the accuracy of QTL mapping is well known; in small populations, only QTLs with large effect are likely to be identified, and their effect can be over-inflated (Tanksley 1993). The small population sizes that are generally used probably arise from difficulty in phenotyping for PUE; the need to screen at both limiting and non-limiting conditions doubles the amount of phenotyping needed.
A review of the studies presented in Table 5 suggests clear directions that should be taken for future QTL mapping studies. A larger number of mapping studies within species will provide greater certainty of the value of QTL regions detected, and common QTLs could then be targeted for map-based cloning and potential transgenic approaches to improve PUE. Problems associated with definition of PUE need to be overcome—a comparison of results obtained using different definitions will help. Ideally, screening for PUE should be conducted on a range of different soil types and environments. Finally, the phenomenon of linkage of developmental genes with PUE should be investigated further, to determine whether certain combinations, particularly in wheat, can lead to improved PUE.
Transgenic approaches for improving PUE
Molecular engineering on a key component in the regulatory network of PSR represents a useful approach for molecular breeding of plants towards more efficient Pi uptake and use. This has been shown in Arabidopsis and rice using overexpression, gene knockdown, or knockout. Artificial target mimics of miRNAs can also be used for functional studies on PSR and this has potential to contribute to molecular breeding. Table 6 provides an overview of genes that have been assessed using transgenic approaches. Overexpression of OsPSTOL1 enhances P uptake in P-deficient soil (Gamuyao et al. 2012). The high P uptake by overexpression of OsPSTOL1 has also been validated in two different genetic background of rice (Gamuyao et al. 2012). Overexpressing AVP1 genes can enhance P uptake in different plant species and environments (Yang et al. 2007).

|
Transgenic rice lines overexpressing OsPHT1;8 increase the maximum influx by 3–5 fold, indicating that the transgenic approach can enhance Pi uptake from soil in this crop (Jia et al. 2011). By contrast, the overexpression of HvPHT1;1 in barley did not lead to an increase in Pi uptake (Rae et al. 2004). Overexpression of AtPHO1 leads to a 2–3-fold increase in shoot Pi content and a severe reduction in shoot growth (Stefanovic et al. 2011). The overexpression of AtPHO1 also largely increases Pi concentration in leaf xylem exudates as a result of a rapid reduction of the vacuolar Pi pool (Stefanovic et al. 2011). Overexpression of miR399 leads to a reduction in remobilisation of Pi in Arabidopsis (Aung et al. 2006) and in rice (Hu et al. 2011). The overexpression approach has also been used to modify secretion of acid phosphatase and protons in the roots of tomato, which facilitated the hydrolysis of soil organic P and dissolution of Pi (Gao et al. 2010).
Knockdown or knockout approaches have also been used for alteration of Pi homeostasis in plants. Knockdown of OsPHT1;8 reduces Pi uptake and translocation (Jia et al. 2011). Knockout of Osphf1 (Pi starvation-induced transcription factor 1) reduces Pi uptake and translocation from roots to shoots in rice, as well as arsenate (Wu et al. 2011). The knockout mutant of ltn1, an orthologue of AtPHO2, shows several typical Pi-starvation responses, such as stimulation of phosphatase and RNase activities, lipid composition alteration, and nitrogen assimilation repression (Hu et al. 2011). The elongation of primary and adventitious roots is also enhanced in the ltn1 mutant, suggesting that the modification of LTN1 expression may be able to enhance morphological, physiological, and biochemical responses to Pi starvation.
Transgene expression driven by a cell-specific promoter instead of a constitutive promoter will be preferred in some cases. Phosphatases that release phosphorus from organic compounds would be more useful if produced by shallow roots than by deep roots, since soil organic matter typically decreases with depth (Lynch 2011). By contrast, carboxylates capable of releasing phosphorus from Fe and Al oxides may be more useful when released into deeper soil horizons where these forms of phosphorus predominate (Lynch 2011).
Challenges to genetic improvement of PUE
There are several challenges to improving PUE, whether based upon conventional genetic approaches, or on GM approaches. These are summarised as follows:
-
Incomplete understanding of controls of P uptake: The growth and P uptake of a crop relies on a complex interaction between the plant, the chemical and physical characteristics of the soil, and the biological properties of the rhizosphere. While there have been significant advances in our understanding of specific components of this system and of their genetic basis, time and again the comment is made in reviews of P nutrition that our understanding of the operation of the system is incomplete. There are many examples of traits that result in substantial improvements in P nutrition under controlled conditions which fail to show similar advantages in field soil. This is one of the compelling reasons for rigorous testing in soils that are representative of commercial practice.
-
Appropriate phenotyping: Much of the reported work comes from glasshouse or hydroponic studies. This is out of convenience, as well as necessity in the case of genetic modification (GM) studies. Few studies have compared results from controlled-environment experiments with responses in field trials. Soil properties will influence the form and the availability of P, but our understanding of the importance of specific mechanisms of PUE in different soils is poor. High-throughput screening methods are desirable, but if they cannot be demonstrated to correlate with results obtained from commercial growing situations, then their application may be limited.
-
Limited long-term genotype field trials: Grain yields of rainfed crops show large environmental variation, which reflects differences in seasonal conditions. Variation in soil moisture and effects of companion stresses such as high Al and Fe as well as low pH are likely to influence P uptake and expression of PUE, but considerations such as these have not been adequately addressed as there are no long-term assessments of genetic diversity for PUE under commercial conditions. Identifying genotypes that show consistent PUE over a range of environments is key to identifying traits to select for improved PUE that will be commercially useful over large areas.
-
Limited population-based studies: A review of the studies presented in Table 5 suggests that there are limited numbers of population-based mapping studies. A larger number of mapping studies within species will provide greater certainty of the value of QTL regions detected, and common QTLs could then be targeted for map-based cloning and potential transgenic approaches to improve PUE.
-
Effect of developmental genes: A common response to P deficiency in crops is a delay in maturity. Studies in wheat and common bean have identified QTLs that are linked to developmental genes (Su et al. 2006; Cichy et al. 2009). Selection for PUE based upon relative development (i.e. development at limiting compared with non-limiting soil P) may provide a cost-effective means of screening for improved PUE, but the nature of the association between development and PUE needs to be examined further.
-
GM approaches: GM shows the potential to improve P uptake in acid soils (Delhaize et al. 2009). Increased P uptake has also been demonstrated by overexpression of AVP1 (Yang et al. 2007). Several other genes have been identified recently in Arabidopsis and rice (see Table 6) that also show promise for improvement of PUE in controlled environments. As discussed above, PUE is a complex trait in which the interactions between the plant and soil interface play a crucial role. Therefore, the potential benefits of GM approaches for improving PUE need to be verified under field conditions. The incomplete understanding of controls of P uptake (see point 1) is a bottleneck for both conventional and GM approaches. Compared with conventional approaches, there are additional hurdles such as consumer and export-market sentiment, regulatory control, and the costs of deregistration for GM approaches. Therefore, benefits for growers from GM approaches are not likely to be realised in the short term. However, GM approaches should be seen as necessary to provide functional validation of any genes identified from approaches such as QTL analysis.
Directions
Improvements in PUE in crop plants can contribute to the overall P efficiency of the cropping system. This may come from more effective and efficient use of fertiliser P or it may allow the more effective use of less-soluble forms of P such as rock phosphate or organic forms of P. In both cases, this may help to extend the life of current P reserves or allow cost savings to farmers. While the complexity of the system has been highlighted, the directions that follow will relate to plant-based traits. It is acknowledged that there are potential gains to be made in better understanding the microbiological properties of the rhizosphere and the interactions between soil microbes and plant roots. The complex soil reactions of P also mean that there would be benefits from closer collaboration between soil and plant scientists.
Based on the current state of knowledge of the P nutrition of crops, most of the proposed genetic solutions to improved P nutrition represent potential opportunities rather than achievable objectives. The complexity of P nutrition and the strong environmental influence on soil P availability and P uptake mean improvements will most likely be achieved from multiple traits being combined rather than relying on one or two traits as ‘best bets’.
The winter cereals, and especially wheat, are the most intensively studied crops. Much less work has been done in examining genetic opportunities in pulses and oilseed crops, although interspecific variation in PUE among these is well documented. The directions recommended this review are based upon need and likely achievement in short (5 years), medium (5–10 years), and longer (>10 years) time-frames (Table 7), focusing on improving PUE in wheat and barley.
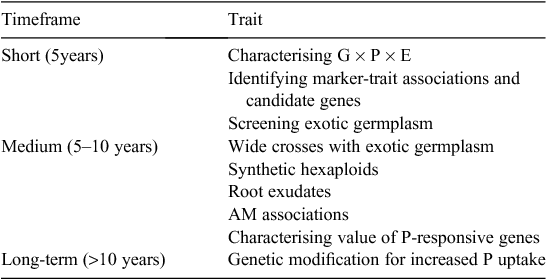
|
Characterising G × P × E
Although there is strong evidence of genetic diversity for PUE and component traits in a range of species, much of this comes from controlled environment experiments with plants grown in soils that are severely P-deficient. Evidence obtained under commercial field conditions is limited, and where obtained, results from the same varieties under both field and glasshouse conditions are often contrasting. Screening of wheat and barley varieties under field conditions (Bovill et al. 2011a, 2011b) has shown that the genotype × P × environment (G × P × E) interaction is large. A better understanding of these interactions is required, and this will only be achieved by obtaining data from a large number of site–season combinations. Therefore, multi-environment trial networks should be established to better understand G × P × E interactions.
Identifying marker-trait associations and candidate genes
QTL mapping
The QTL mapping of populations developed from biparental crosses offers a means to detect marker trait associations. Currently available populations are ideal, as the costs associated with population development are nil; however, further consideration of population structure in relation to segregation for developmental loci such as Ppd (photoperiod responsiveness), Vrn (vernalisation requirement), and Rht (reduced height) is required, as their effect on PUE may confound results.
Association mapping
The recent application of array-based methods for single nucleotide polymorphism (SNP) genotyping in wheat varieties (Akhunov 2011) is a valuable resource for genetic improvement in PUE in this crop. Phenotyping a large association mapping panel under field conditions may help to overcome some problems associated with more traditional biparental QTL mapping. In addition, the possibility exists for combining the association mapping approach with a candidate gene approach to identify genes that could be targeted for transgenic manipulation. The advantage of this approach is that the identified genes would have clear impact for breeding, as they would have been shown to be important under commercial growing conditions. Both the association and QTL mapping approaches should be focussed upon, as they are essential steps in improving the efficiency of selecting for improved PUE by breeding companies.
Screening exotic germplasm
Seed banks of landraces and wild relatives of wheat and barley, as well as material of promise identified internationally (for example, the Chinese germplasm mentioned previously), should be screened to assess for improved PUE. Notably, this approach has been successful in identifying superior salinity tolerance. Broad collections (such as the Watkins collection of wheat landraces from diverse geographical origins) or a more focussed approach (for example, by selecting germplasm adapted to regions with soils with high P-fixing capability) could be used. Ideally, screening would be conducted under field conditions, but the number of lines that are likely to be assessed in the field may limit the effectiveness of this approach. An alternative would be to screen a broad germplasm set under controlled-environment conditions in the first instance, and then advance the most promising lines to field assessment.
Wide crosses with exotic germplasm
Once PUE in landraces and wild relatives and other exotic germplasm has been assessed, wide crosses with promising lines should be instigated. Selection under field conditions will be essential for achieving the goal of improved PUE, and this is acknowledged by the longer time-frame that will be required to achieve this outcome.
Synthetic hexaploids
The value of synthetic hexaploids to PUE has not been examined. Some synthetic hexaploids have more vigorous root systems, which may improve uptake of P. The PUE of T. tauschii has not been systematically characterised but could be part of a program of screening exotic germplasm. A program of synthetic hexaploid development is being conducted in ICARDA and collaboration with this program to improve PUE should be encouraged.
Root exudates
Root exudation is a common strategy to improve P accessibility and uptake in a range of species. However, the suggested benefits of modifying root exudation are yet to be demonstrated under field conditions. Work of this nature is currently being conducted in wheat, using both traditional and transgenic approaches. Further research on genes such as ALMT1 and AVP1 should be supported.
AM associations
Few studies have assessed genotypic variation in AM responsiveness. Using P isotopes, recent data show that the contribution of AM to P uptake is significant even when biomass responses are small or not seen at all. Given that AM plants may be able to access forms of P unavailable for non-AM plants, exploitation of AM associations may provide significant improvements to PUE. However, one of the questions to be addressed is the contribution of mycorrhizae to P uptake in soils with P levels that are representative of current farming systems. Further research examining mycorrhizal responsiveness is warranted. Also a better understanding of the genetic controls of AM infection and P uptake and nutrition of the host plant is required to allow the synergistic relationship to be manipulated.
Characterising value of P-responsive genes
Recently, there has been rapid progress in the understanding of plant response to P starvation and the Pi signalling involved. The progress in Arabidopsis and rice is much more advanced than in winter cereals such as barley and wheat. However, the high conservation of the Pi signalling pathway in dicots and monocots suggests that the knowledge generated from model plant species such as rice and Arabidopsis is applicable to wheat, barley, and canola. We know relatively little about the genes involved in the Pi signalling pathway available in wheat and barley. Identification and characterisation of these key regulatory components will be the starting point to move forward in the improvement of PUE in winter cereals.
Genetic manipulation of key genes involved in the Pi signalling pathway could be used for enhanced P mobilisation in the rhizosphere via secretion of higher levels of metabolites, and enzymes such as phosphatases for degradation of organic P in soil, Pi acquisition, and P remobilisation (see Table 6). Genetic manipulation could also be used to alter root system architecture for higher PUE. Root system architecture is fundamental in the improvement of P efficiency because Pi diffuses slowly in soils. Root proliferation allows exploration of new soil patches containing Pi and delivers exudates to the rhizosphere with potential for increasing P mobility. An optimised root system becomes an integrated part of genetic manipulation in PUE. Genetic manipulation of OsPSTOL1 and AVP1 has provided an excellent example in optimising the root system for the improvement of tolerance to P deficiency (Yang et al. 2007; Gamuyao et al. 2012).
Signalling of Pi starvation is highly integrated with other signalling networks; changes in the Pi signalling pathway become part of dynamic responses to different stresses. Consequently, the genetic manipulation in the key genes involved in Pi signalling could have a pleiotropic effect. Precise manipulation may be necessary and could be achieved through cell-specific and stress-inducible expression. There is little information on genetic variation in the Pi signalling pathway, which could be explored using conventional breeding.
Genetic modification for increased P uptake
Genetic modification technologies promise major benefits for improvement of many traits. However, gene targets for improving PUE with GM approaches are not well defined due to our poor understanding of the physiological properties of PUE traits. Many factors could influence PUE via complex interactions among different processes. Whether the potential target traits could be improved, it is essential to verify the GM lines in both glasshouse and field conditions. The GM approach could be a relatively long-term strategy dependent on how much influence the target gene has on the phenotype. The manipulation of a complex trait such as PUE by any approach requires a consistent and long-term investment.
Concluding remarks
An adequate and balanced supply of essential nutrients is a cornerstone of improvements in crop productivity. Improved PUE will become increasingly important in the future as farmers strive to achieve higher levels of productivity and maintain profitable enterprises in the face of increasing fertiliser prices. In regions where crops are chronically malnourished, increases in soil P fertility through soil improvement and fertiliser use will underpin increases in productivity, while in areas where P has been applied in excess of the crop requirements, better use of the soil P bank and a more sustainable use of fertiliser P will be needed. In both cases, breeding for improved PUE can play an important role in increasing productivity.
There is considerable variability in how crop species and varieties exploit soil P and respond to fertiliser. The improvements in yield potential that have been achieved by breeding have resulted in a passive improvement in PUE, but there are few examples of commercial varieties being developed for their high PUE. The complexity of the soil and plant processes that influence the P status of crops, and incomplete understanding of the genetic control of PUE and its underlying physiological and molecular basis as well as a consistent conceptual understanding of PUE, have limited progress.
The rapid development of an array of molecular and genomic techniques provides an opportunity to overcome many of the hurdles that have hindered progress so far. Plant scientists are at the cusp of making considerable advances in understanding PUE and developing varieties that are more P-efficient. However, an important aspect of the use and implementation of this approach is that material needs to be tested under realistic field conditions. Marshalling new methods and technologies with the traditional disciplines of plant breeding, crop physiology, soil science, and agronomy provides expanded opportunities to study genetic differences in PUE and to link genotype to phenotype.
Acknowledgements
This work was funded by the Grains Research and Development Corporation.
References
ABARE (2009) ‘Australian Commodity Statistics.’ (Australian Bureau of Agricultural and Resource Economics, Canberra, ACT)ABARE (2010) ‘Australian Commodity Statistics.’ (Australian Bureau of Agricultural and Resource Economics: Canberra, ACT)
Akhunov E (2011) Next-generation tools for wheat genetics and breeding: high-throughput SNP genotyping assays and sequence-based genotyping. International Triticeae Mapping Initiative, CIMMYT, Mexico City.
An GH, Kobayashi S, Enoki H, Sonobe K, Muraki M, Karasawa T, Ezawa T (2010) How does arbuscular mycorrhizal colonization vary with host plant genotype? An example based on maize (Zea mays) germplasms. Plant and Soil 327, 441–453.
| How does arbuscular mycorrhizal colonization vary with host plant genotype? An example based on maize (Zea mays) germplasms.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3cXmslGgsg%3D%3D&md5=b15403037f58a8b4b92fe01c4d9d3b77CAS |
Arpat A, Magliano P, Wege S, Rouached H, Stefanovic A, Poirier Y (2012) Functional expression of PHO1 to the Golgi and trans-Golgi network and its role in export of inorganic phosphate. The Plant Journal 71, 479–491.
Aung K, Lin SI, Wu CC, Huang YT, Su CL, Chiou TJ (2006) pho2, a phosphate overaccumulator, is caused by a nonsense mutation in a MicroRNA399 target gene. Plant Physiology 141, 1000–1011.
| pho2, a phosphate overaccumulator, is caused by a nonsense mutation in a MicroRNA399 target gene.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD28Xnt1Ogtrw%3D&md5=c55b0140ec04e201679c1ce4ffc0c221CAS | 16679417PubMed |
Bais HP, Weir TL, Perry LG, Gilroy S, Vivanco JM (2006) The role of root exudates in rhizosphere interactions with plants and other organisms. Annual Review of Plant Biology 57, 233–266.
| The role of root exudates in rhizosphere interactions with plants and other organisms.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD28XosVKhtr8%3D&md5=46ab069eac082ed231465761d857989dCAS | 16669762PubMed |
Baon J, Smith S, Alston A (1993) Mycorrhizal responses of barley cultivars differing in P efficiency. Plant and Soil 157, 97–105.
Batten GD (1992) A review of phosphorus efficiency in wheat. Plant and Soil 146, 163–168.
| A review of phosphorus efficiency in wheat.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaK3sXitlaruro%3D&md5=a485e46a80edc7ca6978ad579eb3ccefCAS |
Beebe SE, Rojas-Pierce M, Yan X, Blair MW, Pedraza F, Munoz F, Tohme J, Lynch JP (2006) Quantitative trait loci for root architecture traits correlated with phosphorus acquisition in common bean. Crop Science 46, 413–423.
| Quantitative trait loci for root architecture traits correlated with phosphorus acquisition in common bean.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD28XhsFKjsbk%3D&md5=bff9540b57931ff47fe6f815fd21c45cCAS |
Bolland MDA, Brennan RF (2008) Comparing the phosphorus requirements of wheat, lupin, and canola. Australian Journal of Agricultural Research 59, 983–998.
| Comparing the phosphorus requirements of wheat, lupin, and canola.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD1cXht1equrnP&md5=f6fb027f5d2e9bf8ee6bea36e9cec7caCAS |
Bolland MDA, Siddique KHM, Loss SP, Baker MJ (1999) Comparing responses of grain legumes, wheat and canola to applications of superphosphate. Nutrient Cycling in Agroecosystems 53, 157–175.
| Comparing responses of grain legumes, wheat and canola to applications of superphosphate.Crossref | GoogleScholarGoogle Scholar |
Bovill WD, Huang CY, Mather DE, McKay A, McNeill A, Porker K, Shoobridge W, Stangoulis JCR, Wheeler R, McDonald GK (2011a) Improving phosphorus use efficiency in barley. In ‘15th Australian Barley Technical Symposium’. Adelaide, S. Aust.
Bovill WD, Huang CY, Mather DE, McKay A, McNeill A, Porker K, Shoobridge W, Stangoulis JCR, Wheeler R, McDonald GK (2011b) Improving phosphorus use efficiency in wheat. In ‘Wheat Breeding Assembly’. Perth, W. Aust.
Brennan R, Bolland M (2001) Comparing fertilizer phosphorus requirements of canola, lupin, and wheat. Journal of Plant Nutrition 24, 1885–1900.
| Comparing fertilizer phosphorus requirements of canola, lupin, and wheat.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD3MXovVCksLw%3D&md5=b907ccbce01ff3ac7e0b0f01b88e0288CAS |
Chiou T-J, Lin S-I (2011) Signaling network in sensing phosphate availability in plants. Annual Review of Plant Biology 62, 185–206.
| Signaling network in sensing phosphate availability in plants.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3MXnslans78%3D&md5=3759fb98c7cfdabbf40f2d92be893c06CAS | 21370979PubMed |
Cichy KA, Blair MW, Galeano Mendoza CH, Snapp SS, Kelly JD (2009) QTL analysis for root architecture traits and low phosphorus tolerance in an Andean bean population. Crop Science 49, 49–68.
Cordell D, Drangert J-O, White S (2009) The story of phosphorus: Global food security and food for thought. Global Environmental Change 19, 292–305.
| The story of phosphorus: Global food security and food for thought.Crossref | GoogleScholarGoogle Scholar |
Coudert Y, Périn C, Courtois B, Khong NG, Gantet P (2010) Genetic control of root development in rice, the model cereal. Trends in Plant Science 15, 219–226.
| Genetic control of root development in rice, the model cereal.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3cXks1WksLw%3D&md5=e332cc98f2b127981eba7abffa873ab4CAS | 20153971PubMed |
Daram P, Brunner S, Rausch C, Steiner C, Amrhein N, Bucher M (1999) Pht2;1 encodes a low-affinity phosphate transporter from Arabidopsis. The Plant Cell 11, 2153–2166.
Davies TGE, Ying J, Xu Q, Li ZS, Li J, Gordon-Weeks R (2002) Expression analysis of putative high-affinity phosphate transporters in Chinese winter wheats. Plant, Cell & Environment 25, 1325–1339.
| Expression analysis of putative high-affinity phosphate transporters in Chinese winter wheats.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD38XotVCqsrg%3D&md5=d589acf53ee7b18952ad043d39d4b3f4CAS |
de la Fuente JM, Ramírez-Rodríguez V, Cabrera-Ponce JL, Herrera-Estrella L (1997) Aluminum tolerance in transgenic plants by alteration of citrate synthesis. Science 276, 1566–1568.
| Aluminum tolerance in transgenic plants by alteration of citrate synthesis.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaK2sXjslGku7Y%3D&md5=c6690f0debe16e3a0d67ef060daf99d8CAS | 9171061PubMed |
Delhaize E, Randall P (1995) Characterization of a phosphate-accumulator mutant of Arabidopsis thaliana. Plant Physiology 107, 207–213.
Delhaize E, Hebb DM, Ryan PR (2001) Expression of a Pseudomonas aeruginosa citrate aynthase gene in tobacco is not associated with either enhanced citrate accumulation or efflux. Plant Physiology 125, 2059–2067.
| Expression of a Pseudomonas aeruginosa citrate aynthase gene in tobacco is not associated with either enhanced citrate accumulation or efflux.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD3MXjtFKqu78%3D&md5=0d2d7463bacc51b500a485f17e9d63deCAS | 11299385PubMed |
Delhaize E, Taylor P, Hocking PJ, Simpson RJ, Ryan PR, Richardson AE (2009) Transgenic barley (Hordeum vulgare L.) expressing the wheat aluminium resistance gene (TaALMT1) shows enhanced phosphorus nutrition and grain production when grown on an acid soil. Plant Biotechnology Journal 7, 391–400.
| Transgenic barley (Hordeum vulgare L.) expressing the wheat aluminium resistance gene (TaALMT1) shows enhanced phosphorus nutrition and grain production when grown on an acid soil.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD1MXmvFejtrY%3D&md5=8c46e780dd1ad01dbc6c3cd5ddc397e1CAS | 19490502PubMed |
Duff J, Sermon D, Walton G, Mangano P, Newman C, Waldon K, Barbetti M, Addison B, Ecksteen D, Leach B (2006) ‘Growing western canola.’ (Oilseeds Industry Association of Western Australia: Belmont, W. Aust.)
FIFA (2003–2010) ‘Fertilizer Industry Public Environment Report.’ (Fertilizer Australia: Canberra, ACT)
Franco-Zorrilla JM, Valli A, Todesco M, Mateos I, Puga MI, Rubio-Somoza I, Leyva A, Weigel D, Garcia JA, Paz-Ares J (2007) Target mimicry provides a new mechanism for regulation of microRNA activity. Nature Genetics 39, 1033–1037.
| Target mimicry provides a new mechanism for regulation of microRNA activity.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2sXot1CmsLY%3D&md5=9cc5327dd399bbf1b86c67a64711733eCAS | 17643101PubMed |
Gahoonia TS, Nielsen NE (1997) Variation in root hairs of barley cultivars doubled soil phosphorus uptake. Euphytica 98, 177–182.
| Variation in root hairs of barley cultivars doubled soil phosphorus uptake.Crossref | GoogleScholarGoogle Scholar |
Gamuyao R, Chin JH, Pariasca-Tanaka J, Pesaresi P, Catausan S, Dalid C, Slamet-Loedin I, Tecson-Mendoza EM, Wissuwa M, Heuer S (2012) The protein kinase Pstol1 from traditional rice confers tolerance of phosphorus deficiency. Nature 488, 535–539.
| The protein kinase Pstol1 from traditional rice confers tolerance of phosphorus deficiency.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC38Xht1emsLzE&md5=995c6726a6b420df423f131e2bc2bc73CAS | 22914168PubMed |
Gao N, Su Y, Min J, Shen W, Shi W (2010) Transgenic tomato overexpressing ath-miR399d has enhanced phosphorus accumulation through increased acid phosphatase and proton secretion as well as phosphate transporters. Plant and Soil 334, 123–136.
| Transgenic tomato overexpressing ath-miR399d has enhanced phosphorus accumulation through increased acid phosphatase and proton secretion as well as phosphate transporters.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3cXhtVaqtbnE&md5=8442e557f8218d9b1e6ba62920020c17CAS |
George TS, Richardson AE (2008) Potential and limitations to improving crops for enhanced phosphorus utilization. In ‘The ecophysiology of plant-phosphorus interactions’. Vol. 7. (Eds PJ White, JP Hammond) pp. 247–270. (Springer: Dordrecht, The Netherlands)
George TS, Brown LK, Newton AC, Hallett PD, Sun BH, Thomas WTB, White PJ (2011) Impact of soil tillage on the robustness of the genetic component of variation in phosphorus (P) use efficiency in barley (Hordeum vulgare L.). Plant and Soil 339, 113–123.
| Impact of soil tillage on the robustness of the genetic component of variation in phosphorus (P) use efficiency in barley (Hordeum vulgare L.).Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3MXmsVCiug%3D%3D&md5=db66add909b5c1da3f8bd4ff1bef194aCAS |
Glassop D, Smith S, Smith F (2005) Cereal phosphate transporters associated with the mycorrhizal pathway of phosphate uptake into roots. Planta 222, 688–698.
| Cereal phosphate transporters associated with the mycorrhizal pathway of phosphate uptake into roots.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2MXhtFyntr%2FF&md5=5e67296eb74ec6533bdce54791e56006CAS | 16133217PubMed |
Graham RD (1984) Breeding for nutritional characteristics in cereals. In ‘Advances in plant nutrition’. Vol. 1. (Eds PB Tinker, A Lauchli) pp. 57–102. (Praeger Publishers: New York)
Guo B, Jin Y, Wussler C, Blancaflor EB, Motes CM, Versaw WK (2008) Functional analysis of the Arabidopsis PHT4 family of intracellular phosphate transporters. New Phytologist 177, 889–898.
| Functional analysis of the Arabidopsis PHT4 family of intracellular phosphate transporters.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD1cXktVSnsLw%3D&md5=54c2fe96e68906735481a61fa4969d72CAS | 18086223PubMed |
Hamburger D, Rezzonico E, MacDonald-Comber Petétot J, Somerville C, Poirier Y (2002) Identification and characterization of the Arabidopsis PHO1 gene involved in phosphate loading to the xylem. The Plant Cell 14, 889–902.
| Identification and characterization of the Arabidopsis PHO1 gene involved in phosphate loading to the xylem.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD38XjsFWktro%3D&md5=291b97183c9013f5714a7de003c5cb6bCAS | 11971143PubMed |
Hammond JP, White PJ (2008) Sucrose transport in the phloem: integrating root responses to phosphorus starvation. Journal of Experimental Botany 59, 93–109.
| Sucrose transport in the phloem: integrating root responses to phosphorus starvation.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD1cXht1SitbY%3D&md5=358c04a55fc894ea2a9a3992ff28f986CAS | 18212031PubMed |
Hammond JP, Broadley MR, White PJ, King GJ, Bowen HC, Hayden R, Meacham MC, Mead A, Overs T, Spracklen WP, Greenwood DJ (2009) Shoot yield drives phosphorus use efficiency in Brassica oleracea and correlates with root architecture traits. Journal of Experimental Botany 60, 1953–1968.
| Shoot yield drives phosphorus use efficiency in Brassica oleracea and correlates with root architecture traits.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD1MXmtFSjtbY%3D&md5=034df31fb900a6fe0f991a7a7522543dCAS | 19346243PubMed |
Hayes JE, Zhu Y-G, Mimura T, Reid RJ (2004) An assessment of the usefulness of solution culture in screening for phosphorus efficiency in wheat. Plant and Soil 261, 91–97.
| An assessment of the usefulness of solution culture in screening for phosphorus efficiency in wheat.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2cXlvVehtLk%3D&md5=63fce0ee73b407a5ea67eb9eb54e426eCAS |
Hinsinger P (2001) Bioavailability of soil inorganic P in the rhizosphere as affected by root-induced chemical changes: a review. Plant and Soil 237, 173–195.
| Bioavailability of soil inorganic P in the rhizosphere as affected by root-induced chemical changes: a review.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD38XovVWlsQ%3D%3D&md5=8991bca6a5f81d192cc3ee94d2ed0671CAS |
Holloway RE, Bertrand I, Frischke AJ, Brace DM, McLaughlin MJ, Shepperd W (2001) Improving fertiliser efficiency on calcareous and alkaline soils with fluid sources of P, N and Zn. Plant and Soil 236, 209–219.
| Improving fertiliser efficiency on calcareous and alkaline soils with fluid sources of P, N and Zn.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD3MXptlKhurc%3D&md5=6814c318b26e40135af9c61034039c8fCAS |
Hsieh L-C, Lin S-I, Shih AC-C, Chen J-W, Lin W-Y, Tseng C-Y, Li W-H, Chiou T-J (2009) Uncovering small RNA-mediated responses to phosphate deficiency in Arabidopsis by deep sequencing. Plant Physiology 151, 2120–2132.
| Uncovering small RNA-mediated responses to phosphate deficiency in Arabidopsis by deep sequencing.Crossref | GoogleScholarGoogle Scholar | 19854858PubMed |
Hu B, Zhu C, Li F, Tang J, Wang Y, Lin A, Liu L, Che R, Chu C (2011) LEAF TIP NECROSIS1 plays a pivotal role in the regulation of multiple phosphate starvation responses in rice. Plant Physiology 156, 1101–1115.
| LEAF TIP NECROSIS1 plays a pivotal role in the regulation of multiple phosphate starvation responses in rice.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3MXptFWlu7o%3D&md5=e380aebb3b6b32b15c007105778227d2CAS | 21317339PubMed |
Huang CY, Roessner U, Eickmeier I, Genc Y, Callahan DL, Shirley N, Langridge P, Bacic A (2008) Metabolite profiling reveals distinct changes in carbon and nitrogen metabolism in phosphate-deficient barley plants (Hordeum vulgare L.). Plant & Cell Physiology 49, 691–703.
| Metabolite profiling reveals distinct changes in carbon and nitrogen metabolism in phosphate-deficient barley plants (Hordeum vulgare L.).Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD1cXnvFWns78%3D&md5=aaedc45e340365d22c27c1074dbc8716CAS |
Huang CY, Shirley N, Genc Y, Shi B, Langridge P (2011) Phosphate utilization efficiency correlates with expression of low-affinity phosphate transporters and non-coding RNA, IPS1 in barley. Plant Physiology 156, 1217–1229.
| Phosphate utilization efficiency correlates with expression of low-affinity phosphate transporters and non-coding RNA, IPS1 in barley.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3MXptFWksrw%3D&md5=707e060c16e5d23e15fd154c4821c767CAS | 21606317PubMed |
Hund A, Reimer R, Messmer R (2011) A consensus map of QTLs controlling the root length of maize. Plant and Soil 344, 143–158.
| A consensus map of QTLs controlling the root length of maize.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3MXnsVCjtbs%3D&md5=bf419110db5b8b1afca1a502ad159aadCAS | 1:CAS:528:DC%2BC3MXnsVCjtbs%3D&md5=bf419110db5b8b1afca1a502ad159aadCAS |
Jakobsen I, Chen B, Munkvold L, Lundsgaard T, Zhu Y-G (2005) Contrasting phosphate acquisition of mycorrhizal fungi with that of root hairs using the root hairless barley mutant. Plant, Cell & Environment 28, 928–938.
| Contrasting phosphate acquisition of mycorrhizal fungi with that of root hairs using the root hairless barley mutant.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2MXms12gsb4%3D&md5=ead322d6083ce7adf3a3ff8201475ca8CAS | 1:CAS:528:DC%2BD2MXms12gsb4%3D&md5=ead322d6083ce7adf3a3ff8201475ca8CAS |
Javot H, Penmetsa RV, Terzaghi N, Cook DR, Harrison MJ (2007) A Medicago truncatula phosphate transporter indispensable for the arbuscular mycorrhizal symbiosis. Proceedings of the National Academy of Sciences of the United States of America 104, 1720–1725.
| A Medicago truncatula phosphate transporter indispensable for the arbuscular mycorrhizal symbiosis.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2sXhslGisr0%3D&md5=6c9870d39d1e87bcacb038f39a605e99CAS | 1:CAS:528:DC%2BD2sXhslGisr0%3D&md5=6c9870d39d1e87bcacb038f39a605e99CAS | 17242358PubMed |
Jayachandran K, Schwab AP, Hetricic BAD (1992) Mineralization of organic phosphorus by vesicular-arbuscular mycorrhizal fungi. Soil Biology & Biochemistry 24, 897–903.
| Mineralization of organic phosphorus by vesicular-arbuscular mycorrhizal fungi.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaK3sXhvFemsw%3D%3D&md5=9f0d59e1655ed0f0f694deb57ce25ba3CAS | 1:CAS:528:DyaK3sXhvFemsw%3D%3D&md5=9f0d59e1655ed0f0f694deb57ce25ba3CAS |
Jia H, Ren H, Gu M, Zhao J, Sun S, Zhang X, Chen J, Wu P, Xu G (2011) The phosphate transporter gene OsPht1;8 is involved in phosphate homeostasis in rice. Plant Physiology 156, 1164–1175.
| The phosphate transporter gene OsPht1;8 is involved in phosphate homeostasis in rice.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3MXptFWlu7Y%3D&md5=2ea628e3a7f0e127a013aac0260fe365CAS | 1:CAS:528:DC%2BC3MXptFWlu7Y%3D&md5=2ea628e3a7f0e127a013aac0260fe365CAS | 21502185PubMed |
Jones GPD, Blair GJ, Jessop RS (1989) Phosphorus efficiency in wheat—A useful selection criterion? Field Crops Research 21, 257–264.
| Phosphorus efficiency in wheat—A useful selection criterion?Crossref | GoogleScholarGoogle Scholar |
Jones GPD, Jessop RS, Blair GJ (1992) Alternative methods for the selection of phosphorus efficiency in wheat. Field Crops Research 30, 29–40.
| Alternative methods for the selection of phosphorus efficiency in wheat.Crossref | GoogleScholarGoogle Scholar |
Kant S, Peng M, Rothstein SJ (2011) Genetic regulation by NLA and microRNA827 for maintaining nitrate-dependent phosphate homeostasis in Arabidopsis. PLOS Genetics 7, e1002021
| Genetic regulation by NLA and microRNA827 for maintaining nitrate-dependent phosphate homeostasis in Arabidopsis.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3MXktl2ks7c%3D&md5=f3b4aa04e76ca23f518bb41b630ae2b9CAS | 1:CAS:528:DC%2BC3MXktl2ks7c%3D&md5=f3b4aa04e76ca23f518bb41b630ae2b9CAS | 21455488PubMed |
Kirkby EA, Johnston AE (2008) Soil and fertilizer phosphorus in relation to crop nutrition. In ‘The ecophysiology of plant–phosphorus interactions’. Vol. 7. (Eds PJ White, JP Hammond) pp. 177–223. (Springer: Dordrecht, The Netherlands)
Koide RT, Kabir Z (2000) Extraradical hyphae of the mycorrhizal fungus Glomus intraradices can hydrolyse organic phosphate. New Phytologist 148, 511–517.
| Extraradical hyphae of the mycorrhizal fungus Glomus intraradices can hydrolyse organic phosphate.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD3MXpvVaktQ%3D%3D&md5=db3cfbdee678b6036bf2efef4b14edb8CAS | 1:CAS:528:DC%2BD3MXpvVaktQ%3D%3D&md5=db3cfbdee678b6036bf2efef4b14edb8CAS |
Kuo H-F, Chiou T-J (2011) The role of microRNAs in phosphorus deficiency signaling. Plant Physiology 156, 1016–1024.
| The role of microRNAs in phosphorus deficiency signaling.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3MXptFWlurs%3D&md5=e5d3820e2097e39aee926bc10ab18582CAS | 1:CAS:528:DC%2BC3MXptFWlurs%3D&md5=e5d3820e2097e39aee926bc10ab18582CAS | 21562333PubMed |
Lei M, Liu Y, Zhang B, Zhao Y, Wang X, Zhou Y, Raghothama KG, Liu D (2011) Genetic and genomic evidence that sucrose is a global regulator of plant responses to phosphate starvation in Arabidopsis. Plant Physiology 156, 1116–1130.
| Genetic and genomic evidence that sucrose is a global regulator of plant responses to phosphate starvation in Arabidopsis.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3MXptFWlu7s%3D&md5=dff687a70f589ecb76c1220e1f07f9a3CAS | 1:CAS:528:DC%2BC3MXptFWlu7s%3D&md5=dff687a70f589ecb76c1220e1f07f9a3CAS | 21346170PubMed |
Liang Q, Cheng X, Mei M, Yan X, Liao H (2010) QTL analysis of root traits as related to phosphorus efficiency in soybean. Annals of Botany 106, 223–234.
| QTL analysis of root traits as related to phosphorus efficiency in soybean.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3cXotVWjsb0%3D&md5=ec892ad823d6d4bbc327787d55c41cc1CAS | 1:CAS:528:DC%2BC3cXotVWjsb0%3D&md5=ec892ad823d6d4bbc327787d55c41cc1CAS | 20472699PubMed |
Liao H, Yan X, Rubio G, Beebe SE, Blair MW, Lynch JP (2004) Genetic mapping of basal root gravitropism and phosphorus acquisition efficiency in common bean. Functional Plant Biology 31, 959–970.
| Genetic mapping of basal root gravitropism and phosphorus acquisition efficiency in common bean.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2cXosFSqu7s%3D&md5=c95d4da702fb8a625c33e98d0606a95bCAS | 1:CAS:528:DC%2BD2cXosFSqu7s%3D&md5=c95d4da702fb8a625c33e98d0606a95bCAS |
Liao M, Hocking PJ, Dong B, Delhaize E, Richardson AE, Ryan PR (2008) Variation in early phosphorus-uptake efficiency among wheat genotypes grown on two contrasting Australian soils. Australian Journal of Agricultural Research 59, 157–166.
| Variation in early phosphorus-uptake efficiency among wheat genotypes grown on two contrasting Australian soils.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD1cXitVGhsbg%3D&md5=3460fcf9deb03964c6d2bb9188e73a88CAS | 1:CAS:528:DC%2BD1cXitVGhsbg%3D&md5=3460fcf9deb03964c6d2bb9188e73a88CAS |
Ligaba A, Shen H, Shibata K, Yamamoto Y, Tanakamaru S, Matsumoto H (2004) The role of phosphorus in aluminium-induced citrate and malate exudation from rape (Brassica napus). Physiologia Plantarum 120, 575–584.
| The role of phosphorus in aluminium-induced citrate and malate exudation from rape (Brassica napus).Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2cXjtFWmu78%3D&md5=c62416b9af93fda18d94ea43108a11a3CAS | 1:CAS:528:DC%2BD2cXjtFWmu78%3D&md5=c62416b9af93fda18d94ea43108a11a3CAS | 15032819PubMed |
Lin S-I, Santi C, Jobet E, Lacut E, El Kholti N, Karlowski WM, Verdeil J-L, Breitler JC, Périn C, Ko S-S, Guiderdoni E, Chiou T-J, Echeverria M (2010) Complex regulation of two target genes encoding SPX-MFS proteins by rice miR827 in response to phosphate starvation. Plant & Cell Physiology 51, 2119–2131.
| Complex regulation of two target genes encoding SPX-MFS proteins by rice miR827 in response to phosphate starvation.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3cXhsFGrs73J&md5=51dbcbdb77fa276502da285e03b9c99cCAS | 1:CAS:528:DC%2BC3cXhsFGrs73J&md5=51dbcbdb77fa276502da285e03b9c99cCAS |
Liu F, Wang ZY, Ren HY, Shen CJ, Li Y, Ling HQ, Wu CY, Lian XM, Wu P (2010) OsSPX1 suppresses the function of OsPHR2 in the regulation of expression of OsPT2 and phosphate homeostasis in shoots of rice. The Plant Journal 62, 508–517.
| OsSPX1 suppresses the function of OsPHR2 in the regulation of expression of OsPT2 and phosphate homeostasis in shoots of rice.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3cXmtFygsLc%3D&md5=490bd559ceaec56dc001df7bad0478daCAS | 1:CAS:528:DC%2BC3cXmtFygsLc%3D&md5=490bd559ceaec56dc001df7bad0478daCAS | 20149131PubMed |
Lott JNA, Ockenden I, Raboy V, Batten GD (2000) Phytic acid and phosphorus in crop seeds and fruits: a global estimate. Seed Science Research 10, 11–33.
Lott JNA, Bojarski M, Kolasa J, Batten GD, Campbell LC (2009) A review of the phosphorus content of dry cereal and legume crops of the world. International Journal of Resources. Governance and Ecology 8, 351–370.
Lynch JP (2011) Root phenes for enhanced soil exploration and phosphorus acquisition: Tools for future crops. Plant Physiology 156, 1041–1049.
| Root phenes for enhanced soil exploration and phosphorus acquisition: Tools for future crops.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3MXptFWlurY%3D&md5=dede40b9ed15ca42f4f80923eada4c4aCAS | 1:CAS:528:DC%2BC3MXptFWlurY%3D&md5=dede40b9ed15ca42f4f80923eada4c4aCAS | 21610180PubMed |
Manske GGB, Ortiz-Monasterio JI, van Ginkel M, González RM, Fischer RA, Rajaram S, Vlek PLG (2001) Importance of P uptake efficiency versus P utilization for wheat yield in acid and calcareous soils in Mexico. European Journal of Agronomy 14, 261–274.
| Importance of P uptake efficiency versus P utilization for wheat yield in acid and calcareous soils in Mexico.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD3MXkvVeksbw%3D&md5=d1b1316d7c7eb1b824792810f7d82251CAS | 1:CAS:528:DC%2BD3MXkvVeksbw%3D&md5=d1b1316d7c7eb1b824792810f7d82251CAS |
Manske GGB, Ortiz-Monasterio JI, van Ginkel RM, Rajaram S, Vlek PLG (2002) Phosphorus use efficiency in tall, semi-dwarf and dwarf near-isogenic lines of spring wheat. Euphytica 125, 113–119.
| Phosphorus use efficiency in tall, semi-dwarf and dwarf near-isogenic lines of spring wheat.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD38XksFeiur0%3D&md5=029f7b3adc39e523d4e9207b1354cfb0CAS | 1:CAS:528:DC%2BD38XksFeiur0%3D&md5=029f7b3adc39e523d4e9207b1354cfb0CAS |
Marschener H (1998) Role of root growth, arbuscular mycorrhiza, and root exudates for the efficiency in nutrient acquisition. Field Crops Research 56, 203–207.
| Role of root growth, arbuscular mycorrhiza, and root exudates for the efficiency in nutrient acquisition.Crossref | GoogleScholarGoogle Scholar |
McBeath TM, Armstrong RA, Lombi E, McLaughlin MJ, Holloway RE (2005) Responsiveness of wheat (Triticum aestivum) to liquid and granular phosphorus fertilisers in southern Australian soils. Soil Research 43, 203–212.
| Responsiveness of wheat (Triticum aestivum) to liquid and granular phosphorus fertilisers in southern Australian soils.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2MXivVOjurc%3D&md5=91e9bf04afdcea3f7287eac89a50bfdcCAS | 1:CAS:528:DC%2BD2MXivVOjurc%3D&md5=91e9bf04afdcea3f7287eac89a50bfdcCAS |
McLaughlin MJ, Alston AM, Martin JK (1988) Phosphorus cycling in wheat-pasture rotations. I. The source of phosphorus taken up by wheat. Australian Journal of Soil Research 26, 323–331.
| Phosphorus cycling in wheat-pasture rotations. I. The source of phosphorus taken up by wheat.Crossref | GoogleScholarGoogle Scholar |
McLaughlin MJ, Fillery IR, Till AR (1992) Operation of the phosphorus, sulphur and nitrogen cycles. In ‘Australia’s renewable resources: Sustainability and global change’. (Eds RM Gifford, MM Barson) pp. 67–116. (Bureau of Rural Resources: Canberra, ACT)
McLaughlin MJ, McBeath TM, Smernik R, Stacey SP, Ajiboye S, Guppy C (2011) The chemical nature of P-accumulation in agricultural soils—implications for fertiliser management and design: an Australian perspective. Plant and Soil 349, 69–87.
| The chemical nature of P-accumulation in agricultural soils—implications for fertiliser management and design: an Australian perspective.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3MXhsFKntbzP&md5=f885e17b7ee646f300d17e711f261b0eCAS | 1:CAS:528:DC%2BC3MXhsFKntbzP&md5=f885e17b7ee646f300d17e711f261b0eCAS |
McNeill AM, Penfold CM (2009) Agronomic management options for phosphorus in Australian dryland organic and low-input cropping systems. Crop & Pasture Science 60, 163–182.
| Agronomic management options for phosphorus in Australian dryland organic and low-input cropping systems.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD1MXitlyrs74%3D&md5=d5e47e507ab2b834c288cbb28372ce92CAS | 1:CAS:528:DC%2BD1MXitlyrs74%3D&md5=d5e47e507ab2b834c288cbb28372ce92CAS |
Moll RH, Kamprath EJ, Jackson WA (1982) Analysis and interpretation of factors which contribute to efficiency of nitrogen utilization. Agronomy Journal 74, 562–564.
| Analysis and interpretation of factors which contribute to efficiency of nitrogen utilization.Crossref | GoogleScholarGoogle Scholar |
Mudge SR, Rae AL, Diatloff E, Smith FW (2002) Expression analysis suggests novel roles for members of the Pht1 family of phosphate transporters in Arabidopsis. The Plant Journal 31, 341–353.
| Expression analysis suggests novel roles for members of the Pht1 family of phosphate transporters in Arabidopsis.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD38XntFelsb8%3D&md5=8e3f02872af17dd13c27ab77392f267bCAS | 1:CAS:528:DC%2BD38XntFelsb8%3D&md5=8e3f02872af17dd13c27ab77392f267bCAS | 12164813PubMed |
Mullen CL, Gammie RL (2002) Phosphorus nutrition for winter crops. Agfact P1.4.5, NSW Agriculture, Orange, NSW.
Nagarajan VK, Jain A, Poling MD, Lewis AJ, Raghothama KG, Smith AP (2011) Arabidopsis Pht1;5 mobilizes phosphate between source and sink organs and influences the interaction between phosphate homeostasis and ethylene signaling. Plant Physiology 156, 1149–1163.
| Arabidopsis Pht1;5 mobilizes phosphate between source and sink organs and influences the interaction between phosphate homeostasis and ethylene signaling.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3MXptFWlu7k%3D&md5=090f0cbeea2115bdb93fa8f7c9276059CAS | 1:CAS:528:DC%2BC3MXptFWlu7k%3D&md5=090f0cbeea2115bdb93fa8f7c9276059CAS | 21628630PubMed |
Nilsson L, Muller R, Nielsen TH (2007) Increased expression of the MYB-related transcription factor, PHR1, leads to enhanced phosphate uptake in Arabidopsis thaliana. Plant, Cell & Environment 30, 1499–1512.
| Increased expression of the MYB-related transcription factor, PHR1, leads to enhanced phosphate uptake in Arabidopsis thaliana.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2sXhsVyjsbbJ&md5=59a4de19e1fb98affb3a69763e024474CAS | 1:CAS:528:DC%2BD2sXhsVyjsbbJ&md5=59a4de19e1fb98affb3a69763e024474CAS |
Nuruzzaman M, Lambers H, Bolland MDA, Veneklaas EJ (2005a) Phosphorus benefits of different legume crops to subsequent wheat grown in different soils of Western Australia. Plant and Soil 271, 175–187.
| Phosphorus benefits of different legume crops to subsequent wheat grown in different soils of Western Australia.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2MXks1Ojt7g%3D&md5=1a2b90571ab9d01ebabd52d15629bdd8CAS | 1:CAS:528:DC%2BD2MXks1Ojt7g%3D&md5=1a2b90571ab9d01ebabd52d15629bdd8CAS |
Nuruzzaman M, Lambers H, Bolland MDA, Veneklaas EJ (2005b) Phosphorus uptake by grain legumes and subsequently grown wheat at different levels of residual phosphorus fertiliser. Australian Journal of Agricultural Research 56, 1041–1047.
| Phosphorus uptake by grain legumes and subsequently grown wheat at different levels of residual phosphorus fertiliser.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2MXhtFChsLbE&md5=6761d987d961fab6217bbafcc59f73e6CAS | 1:CAS:528:DC%2BD2MXhtFChsLbE&md5=6761d987d961fab6217bbafcc59f73e6CAS |
Ochoa IE, Blair MW, Lynch JP (2006) QTL analysis of adventitious root formation in common bean under contrasting phosphorus availability. Crop Science 46, 1609–1621.
| QTL analysis of adventitious root formation in common bean under contrasting phosphorus availability.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD28XotV2jurg%3D&md5=e228fa64bc3a7237fc378d65079e58feCAS | 1:CAS:528:DC%2BD28XotV2jurg%3D&md5=e228fa64bc3a7237fc378d65079e58feCAS |
Ortiz-Monasterio JI, Manske GGB, van Ginkel M (2001) Nitrogen and phosphorus use efficiency. In ‘Application of physiology in wheat breeding’. (Eds MP Reynolds, JI Ortiz-Monasterio, A McNab) pp. 200–207. (CIMMYT: Mexico City)
Osborne LD, Rengel Z (2002a) Genotypic differences in wheat for uptake and utilisation of P from iron phosphate. Australian Journal of Agricultural Research 53, 837–844.
| Genotypic differences in wheat for uptake and utilisation of P from iron phosphate.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD38Xmt1GmtLg%3D&md5=4ff33f50fd6acee4165b04d1555d60c2CAS | 1:CAS:528:DC%2BD38Xmt1GmtLg%3D&md5=4ff33f50fd6acee4165b04d1555d60c2CAS |
Osborne LD, Rengel Z (2002b) Growth and P uptake by wheat genotypes supplied with phytate as the only P source. Australian Journal of Agricultural Research 53, 845–850.
| Growth and P uptake by wheat genotypes supplied with phytate as the only P source.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD38Xmt1GmtLk%3D&md5=e89bca83f83ab7d475b233bb5a75bda1CAS | 1:CAS:528:DC%2BD38Xmt1GmtLk%3D&md5=e89bca83f83ab7d475b233bb5a75bda1CAS |
Ozturk L, Eker S, Torun B, Cakmak I (2005) Variation in phosphorus efficiency among 73 bread and durum wheat genotypes grown in a phosphorus-deficient calcareous soil. Plant and Soil 269, 69–80.
| Variation in phosphorus efficiency among 73 bread and durum wheat genotypes grown in a phosphorus-deficient calcareous soil.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2MXks1Ojur8%3D&md5=b7aec6bab8d961882cbc48733360e2c2CAS | 1:CAS:528:DC%2BD2MXks1Ojur8%3D&md5=b7aec6bab8d961882cbc48733360e2c2CAS |
Pant BD, Musialak-Lange M, Nuc P, May P, Buhtz A, Kehr J, Walther D, Scheible W-R (2009) Identification of nutrient-responsive Arabidopsis and rapeseed microRNAs by comprehensive real-time polymerase chain reaction profiling and small RNA sequencing. Plant Physiology 150, 1541–1555.
| Identification of nutrient-responsive Arabidopsis and rapeseed microRNAs by comprehensive real-time polymerase chain reaction profiling and small RNA sequencing.Crossref | GoogleScholarGoogle Scholar | 19465578PubMed |
Pearse S, Veneklaas E, Cawthray G, Bolland M, Lambers H (2006) Carboxylate release of wheat, canola and 11 grain legume species as affected by phosphorus status. Plant and Soil 288, 127–139.
| Carboxylate release of wheat, canola and 11 grain legume species as affected by phosphorus status.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD28XhtFaksL7F&md5=5d36e8e5910ea3ec15d2488a3f11d454CAS |
Péret B, De Rybel B, Casimiro I, Benková E, Swarup R, Laplaze L, Beeckman T, Bennett MJ (2009) Arabidopsis lateral root development: an emerging story. Trends in Plant Science 14, 399–408.
| Arabidopsis lateral root development: an emerging story.Crossref | GoogleScholarGoogle Scholar | 19559642PubMed |
Plaxton WC, Tran HT (2011) Metabolic adaptations of phosphate-starved plants. Plant Physiology 156, 1006–1015.
| Metabolic adaptations of phosphate-starved plants.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3MXptFWluro%3D&md5=41d4cf9e6120c6bb39a28b8b71e6dd4bCAS | 1:CAS:528:DC%2BC3MXptFWluro%3D&md5=41d4cf9e6120c6bb39a28b8b71e6dd4bCAS | 21562330PubMed |
Poirier Y, Thoma S, Somerville C, Schiefelbein J (1991) A mutant of Arabidopsis deficient in xylem loading of phosphate. Plant Physiology 97, 1087–1093.
| A mutant of Arabidopsis deficient in xylem loading of phosphate.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaK38XotVGluw%3D%3D&md5=9dca9285118b48d9eaf83e42a045b6e8CAS | 1:CAS:528:DyaK38XotVGluw%3D%3D&md5=9dca9285118b48d9eaf83e42a045b6e8CAS | 16668493PubMed |
Preuss CP, Huang CY, Gilliham M, Tyerman SD (2010) Channel-like characteristics of the low-affinity barley phosphate transporter PHT1;6 when expressed in Xenopus oocytes. Plant Physiology 152, 1431–1441.
| Channel-like characteristics of the low-affinity barley phosphate transporter PHT1;6 when expressed in Xenopus oocytes.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3cXmsF2lsbo%3D&md5=1dc5712a0aaae5f0e69ca7fb6356a275CAS | 1:CAS:528:DC%2BC3cXmsF2lsbo%3D&md5=1dc5712a0aaae5f0e69ca7fb6356a275CAS | 20053709PubMed |
Preuss CP, Huang CY, Tyerman SD (2011) Proton-coupled high-affinity phosphate transport revealed from heterologous characterization in Xenopus of barley-root plasma membrane transporter, HvPHT1;1. Plant, Cell & Environment 34, 681–689.
| Proton-coupled high-affinity phosphate transport revealed from heterologous characterization in Xenopus of barley-root plasma membrane transporter, HvPHT1;1.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3MXltFaksro%3D&md5=000ecde952daef465fa2d7e3df6de504CAS | 1:CAS:528:DC%2BC3MXltFaksro%3D&md5=000ecde952daef465fa2d7e3df6de504CAS |
Rae AL, Cybinski DH, Jarmey JM, Smith FW (2003) Characterization of two phosphate transporters from barley; evidence for diverse function and kinetic properties among members of the Pht1 family. Plant Molecular Biology 53, 27–36.
| Characterization of two phosphate transporters from barley; evidence for diverse function and kinetic properties among members of the Pht1 family.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD3sXpvVGgtLo%3D&md5=a44813c002c4e56d19980493b81f4f81CAS | 1:CAS:528:DC%2BD3sXpvVGgtLo%3D&md5=a44813c002c4e56d19980493b81f4f81CAS | 14756304PubMed |
Rae AL, Jarmey JM, Mudge SR, Smith FW (2004) Over-expression of a high-affinity phosphate transporter in transgenic barley plants does not enhance phosphate uptake rates. Functional Plant Biology 31, 141–148.
| Over-expression of a high-affinity phosphate transporter in transgenic barley plants does not enhance phosphate uptake rates.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2cXhsl2gtro%3D&md5=ff0d0399aca3856018838d709d6f4cbcCAS | 1:CAS:528:DC%2BD2cXhsl2gtro%3D&md5=ff0d0399aca3856018838d709d6f4cbcCAS |
Ramaiah M, Jain A, Baldwin JC, Karthikeyan A, Raghothama KG (2011) Characterization of the phosphate-starvation-induced Glycerol-3-phosphate permease gene family in Arabidopsis. Plant Physiology 157, 279–291.
| Characterization of the phosphate-starvation-induced Glycerol-3-phosphate permease gene family in Arabidopsis.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3MXht1Sit7zO&md5=ed20be0d61d70e66250cd74c8d78e229CAS | 1:CAS:528:DC%2BC3MXht1Sit7zO&md5=ed20be0d61d70e66250cd74c8d78e229CAS | 21788361PubMed |
Rausch C, Bucher M (2002) Molecular mechanisms of phosphate transport in plants. Planta 216, 23–37.
| Molecular mechanisms of phosphate transport in plants.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD38Xps1eht70%3D&md5=8ad6d05986eed950ee862c8b8dcefc41CAS | 1:CAS:528:DC%2BD38Xps1eht70%3D&md5=8ad6d05986eed950ee862c8b8dcefc41CAS | 12430011PubMed |
Rausch C, Zimmermann P, Amrhein N, Bucher M (2004) Expression analysis suggests novel roles for the plastidic phosphate transporter Pht2;1 in auto- and heterotrophic tissues in potato and Arabidopsis. The Plant Journal 39, 13–28.
| Expression analysis suggests novel roles for the plastidic phosphate transporter Pht2;1 in auto- and heterotrophic tissues in potato and Arabidopsis.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2cXms1yktbg%3D&md5=fa92e05a4a731c2d80b649ebb36c8d14CAS | 1:CAS:528:DC%2BD2cXms1yktbg%3D&md5=fa92e05a4a731c2d80b649ebb36c8d14CAS | 15200639PubMed |
Rengel Z, Marschner P (2005) Nutrient availability and management in the rhizosphere: exploiting genotypic differences. New Phytologist 168, 305–312.
| Nutrient availability and management in the rhizosphere: exploiting genotypic differences.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2MXht1Smu7bK&md5=fad8f0f659d78a53f79bae78e58bc457CAS | 1:CAS:528:DC%2BD2MXht1Smu7bK&md5=fad8f0f659d78a53f79bae78e58bc457CAS | 16219070PubMed |
Richardson AE, Hocking PJ, Simpson RJ, George TS (2009) Plant mechanisms to optimise access to soil phosphorus. Crop & Pasture Science 60, 124–143.
| Plant mechanisms to optimise access to soil phosphorus.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD1MXitlyrs7s%3D&md5=b89d3512cf5612f00b3e6f953863de94CAS | 1:CAS:528:DC%2BD1MXitlyrs7s%3D&md5=b89d3512cf5612f00b3e6f953863de94CAS |
Rolland F, Baena-Gonzalez E, Sheen J (2006) Sugar sensing and signaling in plants: Conserved and novel mechanisms. In ‘Annual review of plant biology’. Vol. 57. pp. 675–709. (Annual Reviews: Palo Alto, CA)
Rose TJ, Wissuwa M (2012) Rethinking internal phosphorus utilization efficiency: a new approach is needed to improve PUE in grain crops. Advances in Agronomy 116, 185–217.
| Rethinking internal phosphorus utilization efficiency: a new approach is needed to improve PUE in grain crops.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC38XhslSlur%2FP&md5=bed7199080f5d32166906c5226ef6e47CAS | 1:CAS:528:DC%2BC38XhslSlur%2FP&md5=bed7199080f5d32166906c5226ef6e47CAS |
Rose TJ, Rose MT, Pariasca-Tanaka J, Heuer S, Wissuwa M (2011) The frustration with utilization: why have improvements in internal phosphorus utilization efficiency in crops remained so elusive? Frontiers in Plant Science
| The frustration with utilization: why have improvements in internal phosphorus utilization efficiency in crops remained so elusive?Crossref | GoogleScholarGoogle Scholar | 22639608PubMed | 22639608PubMed |
Rouached H, Stefanovic A, Secco D, Bulak Arpat A, Gout E, Bligny R, Poirier Y (2011) Uncoupling phosphate deficiency from its major effects on growth and transcriptome via PHO1 expression in Arabidopsis. The Plant Journal 65, 557–570.
| Uncoupling phosphate deficiency from its major effects on growth and transcriptome via PHO1 expression in Arabidopsis.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3MXjtVaitrw%3D&md5=077d565e070182ae355a1885ba01d879CAS | 1:CAS:528:DC%2BC3MXjtVaitrw%3D&md5=077d565e070182ae355a1885ba01d879CAS | 21288266PubMed | 21288266PubMed |
Rubio V, Linhares F, Solano R, Martín AC, Iglesias J, Leyva A, Paz-Ares J (2001) A conserved MYB transcription factor involved in phosphate starvation signaling both in vascular plants and in unicellular algae. Genes & Development 15, 2122–2133.
| A conserved MYB transcription factor involved in phosphate starvation signaling both in vascular plants and in unicellular algae.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD3MXmt1ajsb8%3D&md5=3ad01576868ec81ea883248cfbf8a679CAS | 1:CAS:528:DC%2BD3MXmt1ajsb8%3D&md5=3ad01576868ec81ea883248cfbf8a679CAS |
Ryan MH, Angus JF (2003) Arbuscular mycorrhizae in wheat and field pea crops on a low P soil: increased Zn-uptake but no increase in P-uptake or yield. Plant and Soil 250, 225–239.
| Arbuscular mycorrhizae in wheat and field pea crops on a low P soil: increased Zn-uptake but no increase in P-uptake or yield.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD3sXit1CisLY%3D&md5=e672228d3a1ce61761a83af0c76c5d27CAS | 1:CAS:528:DC%2BD3sXit1CisLY%3D&md5=e672228d3a1ce61761a83af0c76c5d27CAS |
Ryan MH, Norton RM, Kirkegaard JA, McCormick KM, Knights SE, Angus JF (2002) Increasing mycorrhizal colonisation does not improve growth and nutrition of wheat on Vertosols in south-eastern Australia. Australian Journal of Agricultural Research 53, 1173–1181.
| Increasing mycorrhizal colonisation does not improve growth and nutrition of wheat on Vertosols in south-eastern Australia.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD38Xos1ektLw%3D&md5=3d72f81a0c5f883a2d4b0908a071d982CAS |
Ryan PR, Dessaux Y, Thomashow LS, Weller DM (2009) Rhizosphere engineering and management for sustainable agriculture. Plant and Soil 321, 363–383.
| Rhizosphere engineering and management for sustainable agriculture.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD1MXos1emsr8%3D&md5=e9dc3f65e2ca1af70a4ab681ea350e4eCAS | 1:CAS:528:DC%2BD1MXos1emsr8%3D&md5=e9dc3f65e2ca1af70a4ab681ea350e4eCAS |
Schreiber A, Shi B-J, Huang C-Y, Langridge P, Baumann U (2011) Discovery of barley miRNAs through deep sequencing of short reads. BMC Genomics 12, 129
| Discovery of barley miRNAs through deep sequencing of short reads.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3MXjtVylt70%3D&md5=8ba1066c934d78ad9084ab3cb6c039e0CAS | 1:CAS:528:DC%2BC3MXjtVylt70%3D&md5=8ba1066c934d78ad9084ab3cb6c039e0CAS | 21352554PubMed | 21352554PubMed |
Secco D, Wang C, Shou H, Whelan J (2012) Phosphate homeostasis in the yeast Saccharomyces cerevisiae, the key role of the SPX domain-containing proteins. FEBS Letters 586, 289–295.
| Phosphate homeostasis in the yeast Saccharomyces cerevisiae, the key role of the SPX domain-containing proteins.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC38XhvVOqsL0%3D&md5=62a7a1d4bfd6405718e2e5155e8735fbCAS | 1:CAS:528:DC%2BC38XhvVOqsL0%3D&md5=62a7a1d4bfd6405718e2e5155e8735fbCAS | 22285489PubMed | 22285489PubMed |
Serafin L, Holland J, Bambach R, McCaffery G (2005) Canola: northern NSW planting guide. New South Wales Department of Primary Industries, April 2005. Available at: www.nvtonline.com.au/wp-content/uploads/2013/03/Crop-Guide-Canola-Northern-NSW-Planting-Guide.pdf.
Simpson R, Oberson A, Culvenor R, Ryan M, Veneklaas E, Lambers H, Lynch J, Ryan P, Delhaize E, Smith F, Smith S, Harvey P, Richardson A (2011) Strategies and agronomic interventions to improve the phosphorus-use efficiency of farming systems. Plant and Soil 349, 89–120.
| Strategies and agronomic interventions to improve the phosphorus-use efficiency of farming systems.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3MXhsFKntb7M&md5=58a92eb9456fb3c552e063bf8f46d977CAS | 1:CAS:528:DC%2BC3MXhsFKntb7M&md5=58a92eb9456fb3c552e063bf8f46d977CAS |
Smith FA, Smith SE (2011) What is the significance of the arbuscular mycorrhizal colonisation of many economically important crop plants? Plant and Soil 348, 63–79.
| What is the significance of the arbuscular mycorrhizal colonisation of many economically important crop plants?Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3MXht1Gnt73J&md5=18b2f9d46f2120581b23d74c00665c36CAS |
Steen I (1998) Phosphorus availability in the 21st century: Management of a non-renewable resource. Potassium and Phosphorus 217, 25–31.
Stefanovic A, Arpat AB, Bligny R, Gout E, Vidoudez C, Bensimon M, Poirier Y (2011) Over-expression of PHO1 in Arabidopsis leaves reveals its role in mediating phosphate efflux. The Plant Journal 66, 689–699.
| Over-expression of PHO1 in Arabidopsis leaves reveals its role in mediating phosphate efflux.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3MXmvFerurw%3D&md5=e9be0414361b1447516918c613f68067CAS | 21309867PubMed | 21309867PubMed |
Su J, Xiao Y, Li M, Liu Q, Li B, Tong Y, Jia J, Li Z (2006) Mapping QTLs for phosphorus-deficiency tolerance at wheat seedling stage. Plant and Soil 281, 25–36.
| Mapping QTLs for phosphorus-deficiency tolerance at wheat seedling stage.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD28XktlyhsLY%3D&md5=35a880b44640f640674dcd933c5c483bCAS |
Su JY, Zheng Q, Li HW, Li B, Jing RL, Tong YP, Li ZS (2009) Detection of QTLs for phosphorus use efficiency in relation to agronomic performance of wheat grown under phosphorus sufficient and limited conditions. Plant Science 176, 824–836.
| Detection of QTLs for phosphorus use efficiency in relation to agronomic performance of wheat grown under phosphorus sufficient and limited conditions.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD1MXksVWisb0%3D&md5=e8b73d825f8198798abb2860eca59ff0CAS | 1:CAS:528:DC%2BD1MXksVWisb0%3D&md5=e8b73d825f8198798abb2860eca59ff0CAS |
Syers JK, Johnston AE, Curtin D (2008) ‘Efficiency of soil and fertilizer phosphorus use.’ FAO Fertilizer and Plant Nutrition Bulletin No. 18. (FAO: Rome)
Szarejko I, Janiak A, Chmilelwska B, Nawrot M (2005) Genetic analysis of several root hair mutants of barley. Barley Genetics Newsletter 35, 36–38.
Tanksley SD (1993) Mapping polygenes. Annual Review of Genetics 27, 205–233.
| Mapping polygenes.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaK2cXht1KitLc%3D&md5=cfab77380756c2e9d7a2bf4d7b1dad08CAS | 1:CAS:528:DyaK2cXht1KitLc%3D&md5=cfab77380756c2e9d7a2bf4d7b1dad08CAS | 8122902PubMed | 8122902PubMed |
Versaw WK, Harrison MJ (2002) A chloroplast phosphate transporter, PHT2;1, influences allocation of phosphate within the plant and phosphate-starvation responses. The Plant Cell 14, 1751–1766.
| A chloroplast phosphate transporter, PHT2;1, influences allocation of phosphate within the plant and phosphate-starvation responses.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD38XmslChu7w%3D&md5=41f2eadb16c160424cc364c3d65fd3f1CAS | 12172020PubMed | 12172020PubMed |
Vu DT, Armstrong RD, Sale PWG, Tang CX (2010) Phosphorus availability for three crop species as a function of soil type and fertilizer history. Plant and Soil 337, 497–510.
| Phosphorus availability for three crop species as a function of soil type and fertilizer history.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3cXhsVWmtbrP&md5=6203fbd3fa773d76d49bb1cdbe385b7dCAS | 1:CAS:528:DC%2BC3cXhsVWmtbrP&md5=6203fbd3fa773d76d49bb1cdbe385b7dCAS |
Wang X, Shen J, Liao H (2010) Aqquisition of utilization, which is more critical for enhancing phosphorus efficiency in modern crops? Plant Science 179, 302–306.
| Aqquisition of utilization, which is more critical for enhancing phosphorus efficiency in modern crops?Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3cXpvFKkur0%3D&md5=10ca6886e930cf7037ecfff78b4b3083CAS | 1:CAS:528:DC%2BC3cXpvFKkur0%3D&md5=10ca6886e930cf7037ecfff78b4b3083CAS |
Wasaki J, Yonetani R, Kuroda S, Shinano T, Yazaki J, Fujii F, Shimbo K, Yamamoto K, Sakata K, Sasaki T, Kishimoto N, Kikuchi S, Yamagishi M, Osaki M (2003) Transcriptomic analysis of metabolic changes by phosphorus stress in rice plant roots. Plant, Cell & Environment 26, 1515–1523.
| Transcriptomic analysis of metabolic changes by phosphorus stress in rice plant roots.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD3sXotFyqsr8%3D&md5=e1fd8a36491608b6c6aa2211e14f3ddeCAS | 1:CAS:528:DC%2BD3sXotFyqsr8%3D&md5=e1fd8a36491608b6c6aa2211e14f3ddeCAS |
Wissuwa M, Yano M, Ae N (1998) Mapping of QTLs for phosphorus-deficiency tolerance in rice (Oryza sativa L.). Theoretical and Applied Genetics 97, 777–783.
| Mapping of QTLs for phosphorus-deficiency tolerance in rice (Oryza sativa L.).Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DyaK1cXotVahtrY%3D&md5=a3dc3cb1f6374113e2022f1e0e8e4b06CAS | 1:CAS:528:DyaK1cXotVahtrY%3D&md5=a3dc3cb1f6374113e2022f1e0e8e4b06CAS |
Wu Z, Ren H, McGrath SP, Wu P, Zhao FJ (2011) Investigating the contribution of the phosphate transport pathway to arsenic accumulation in rice. Plant Physiology 157, 498–508.
| Investigating the contribution of the phosphate transport pathway to arsenic accumulation in rice.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3MXht1Sit73E&md5=169d513d939c8f6d634acc38729538cbCAS | 1:CAS:528:DC%2BC3MXht1Sit73E&md5=169d513d939c8f6d634acc38729538cbCAS | 21715673PubMed | 21715673PubMed |
Yan X, Wu P, Ling H, Xu G, Xu F, Zhang Q (2006) Plant nutriomics in China: An overview. Annals of Botany 98, 473–482.
| Plant nutriomics in China: An overview.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD28XhtVequrnE&md5=d71a595743bbdbcc2f4d1a41ae0d76b2CAS | 1:CAS:528:DC%2BD28XhtVequrnE&md5=d71a595743bbdbcc2f4d1a41ae0d76b2CAS | 16735410PubMed | 16735410PubMed |
Yang H, Knapp J, Koirala P, Rajagopal D, Peer WA, Silbart LK, Murphy A, Gaxiola RA (2007) Enhanced phosphorus nutrition in monocots and dicots over-expressing a phosphorus-responsive type IH+-pyrophosphatase. Plant Biotechnology Journal 5, 735–745.
| Enhanced phosphorus nutrition in monocots and dicots over-expressing a phosphorus-responsive type IH+-pyrophosphatase.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2sXhtlWjsrfL&md5=9e7fddcefc957088f5c43dca694571b4CAS | 1:CAS:528:DC%2BD2sXhtlWjsrfL&md5=9e7fddcefc957088f5c43dca694571b4CAS | 17711412PubMed | 17711412PubMed |
Yang M, Ding G, Shi L, Feng J, Xu F, Meng J (2010) Quantitative trait loci for root morphology in response to low phosphorus stress in Brassica napus. Theoretical and Applied Genetics 121, 181–193.
| Quantitative trait loci for root morphology in response to low phosphorus stress in Brassica napus.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3cXmtFagu7Y%3D&md5=2e873ccd9da47e5cf1ad5891f641d76fCAS | 20217384PubMed | 20217384PubMed |
Yang M, Ding G, Shi L, XU F, Meng J (2011) Detection of QTL for phosphorus efficiency at vegetative stage in Brassica napus. Plant and Soil 339, 97–111.
| Detection of QTL for phosphorus efficiency at vegetative stage in Brassica napus.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC3MXmsVCguw%3D%3D&md5=9a8584339bc44f3309ae9a9a01f0e4d3CAS |
Yang S-Y, Grønlund M, Jakobsen I, Grotemeyer MS, Rentsch D, Miyao A, Hirochika H, Kumar CS, Sundaresan V, Salamin N, Catausan S, Mattes N, Heuer S, Paszkowski U (2012) Nonredundant regulation of rice arbuscular mycorrhizal symbiosis by two members of the PHOSPHATE TRANSPORTER1 gene family. The Plant Cell 24, 4236–4251.
| Nonredundant regulation of rice arbuscular mycorrhizal symbiosis by two members of the PHOSPHATE TRANSPORTER1 gene family.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BC38XhvVKls7rE&md5=f48d35b9eba3eb6611cfc30dc0d4ad5cCAS | 1:CAS:528:DC%2BC38XhvVKls7rE&md5=f48d35b9eba3eb6611cfc30dc0d4ad5cCAS | 23073651PubMed | 23073651PubMed |
Yi K, Wu Z, Zhou J, Du L, Guo L, Wu Y, Wu P (2005) OsPTF1, a novel transcription factor involved in tolerance to phosphate starvation in rice. Plant Physiology 138, 2087–2096.
| OsPTF1, a novel transcription factor involved in tolerance to phosphate starvation in rice.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD2MXps12lsL8%3D&md5=47e0797b85a18646ac6ff23d98bfdd73CAS | 1:CAS:528:DC%2BD2MXps12lsL8%3D&md5=47e0797b85a18646ac6ff23d98bfdd73CAS | 16006597PubMed | 16006597PubMed |
Zhang D, Cheng H, Geng L, Kan G, Cui S, Meng Q, Gai J, Yu D (2009) Detection of quantitative trait loci for phosphorus deficiency tolerance at soybean seedling stage. Euphytica 167, 313–322.
| Detection of quantitative trait loci for phosphorus deficiency tolerance at soybean seedling stage.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD1MXltl2gur8%3D&md5=527da543ff41e9bb1de576e0a8c8b5b9CAS | 1:CAS:528:DC%2BD1MXltl2gur8%3D&md5=527da543ff41e9bb1de576e0a8c8b5b9CAS |
Zhou J, Jiao FC, Wu ZC, Li YY, Wang XM, He XW, Zhong WQ, Wu P (2008) OsPHR2 is involved in phosphate-starvation signaling and excessive phosphate accumulation in shoots of plants. Plant Physiology 146, 1673–1686.
| OsPHR2 is involved in phosphate-starvation signaling and excessive phosphate accumulation in shoots of plants.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD1cXkvVWisr4%3D&md5=9f5863d2a349f38cf4dfc13d1bcf2896CAS | 1:CAS:528:DC%2BD1cXkvVWisr4%3D&md5=9f5863d2a349f38cf4dfc13d1bcf2896CAS | 18263782PubMed | 18263782PubMed |
Zhu YG, Smith SE, Barritt AR, Smith FA (2001) Phosphorus (P) efficiencies and mycorrhizal responsiveness of old and modern wheat cultivars. Plant and Soil 237, 249–255.
| Phosphorus (P) efficiencies and mycorrhizal responsiveness of old and modern wheat cultivars.Crossref | GoogleScholarGoogle Scholar | 1:CAS:528:DC%2BD38XovVWlug%3D%3D&md5=ed5831721d7e8b9c24cf788cbaa83d72CAS | 1:CAS:528:DC%2BD38XovVWlug%3D%3D&md5=ed5831721d7e8b9c24cf788cbaa83d72CAS |