

Triple La Niña 2020–2022 revival: river red gum and coolibah recruitment in the Queensland Murray–Darling Basin floodplain, Australia
Michael R. Ngugi A * , Jaye Lobegeiger B , Janice L. Kerr C , Victor J. Neldner A , Jessica McCabe B , Glenn B. McGregor B and Luke Hogan AA
B
C
Abstract
Floodplain forests in the Queensland Murray–Darling Basin (QMDB) have experienced significant recruitment challenges following multiple droughts, including the Millennium Drought (1997–2009). This study investigated the impact of the rare triple-dip La Niña event (2020–2022) on floodplain tree recruitment patterns for river red gum (Eucalyptus camaldulensis Dehnh.) and coolibah (Eucalyptus coolabah Blakely & Jacobs). Field assessments across QMDB’s three water catchment areas showed recruitment at 34% of plots. Days since last flood (P = 0.04) was significantly associated with recruitment density, whereas bankfull exceedance floods showed a marginally positive effect on recruitment density (P = 0.080), and total days of exceedance had a marginally negative influence on recruitment density (P = 0.06). Open woodland and woodland areas with low canopy cover exhibited significantly higher recruitment rates (P = 0.001 and P = 0.021) compared to forests. Low native fauna grazing pressure showed strong association with increased recruitment (P = 0.007); during wet conditions, abundant grass cover diverted browsing pressure on seedlings. Demographic analysis demonstrated improved reverse J-curve distributions for both species compared with previous assessments, suggesting that this triple-dip event recruitment episode may help restore juvenile cohorts. Findings suggest that although flooding appears to contribute to recruitment through site preparation and seed dispersal, sustained rainfall appears crucial for seedling survival. These results have important implications for environmental water management, highlighting that infrequent but significant recruitment episodes may be sufficient for maintaining sustainable populations over the extended lifespans of these species, while demonstrating the limits of direct management interventions.
Keywords: climate variability, demographic recovery, environmental flows, episodic recruitment, floodplain forests, floodplain restoration, rangeland ecology, tree demographics.
Introduction
Murray–Darling Basin (MDB) floodplain and riparian ecosystems represent crucial biodiversity hotspots, supporting diverse native flora and fauna including many threatened taxa (MDBA 2024). This Basin, Australia’s largest river system (Fig. 1), encompasses over 1 million km2 across the states of Queensland, New South Wales, South Australia and Victoria, and the Australian Capital Territory (MDBA 2012). Coordinated water management throughout this vast Basin is provided by the Murray–Darling Basin Plan, which sets restoration targets for aquatic ecosystems, including floodplain trees (MDBA 2012). However, following European settlement, these ecosystems have experienced significant alteration and degradation from river regulation, water extraction, broadscale land clearing, loss of Aboriginal cultural practices, and shifting climate patterns (Kingsford 2000; Mac Nally et al. 2011; Capon et al. 2013).
Map of Australia showing the Murray–Darling Basin. The Basin is divided into the northern Basin, which includes the area shown in Queensland and parts of northern New South Wales (NSW), and southern Basin (southern NSW, Victoria, South Australia, and Australian Capital Territory).
In addition to these anthropogenic pressures, the Basin has also faced many droughts such as the World War II Drought (1937–1945) and the Millennium Drought (1997–2009), which were characterised by prolonged sequences of dry years that occurred consecutively, or interspersed with moderate or even wet years (BOM 2020a). From the mid-1970s, both the northern and southern Basins have experienced more dry than wet years, with multiple consecutive wet years typically required to recover from drought (BOM 2020a). The Millennium Drought from 1997 to 2009, which represented the longest consecutive period of below-median rainfall in south-eastern Australia since at least 1900 and which has been described as the worst drought on record for south-eastern Australia (Van Dijk et al. 2013), catalysed the development of the comprehensive Murray–Darling Basin Plan to restore river system health (MDBA 2012).
The natural structure and function of floodplain forest ecosystems depend intrinsically on flow regimes, with factors such as drought (extended low or no-flow periods), irregular precipitation patterns, and major flood events influencing the demographic composition of floodplain forest communities (Cooper et al. 2003). The Millennium Drought in the MDB triggered severe deterioration of riparian forests and woodlands (Doody et al. 2014), with extensive dieback of the dominant riparian species, river red gum (Eucalyptus camaldulensis Dehnh.), documented throughout the Victorian Murray River floodplain (Cunningham et al. 2011; Mac Nally et al. 2011). Such severe drought impacts on floodplain ecosystems are not specific to the MDB.
Owing to the direct connection between river flows and rainfall events, opportunities to better understand tree recruitment processes on floodplains have been restricted by multiple extended droughts in the MDB. These include periods preceding the Millennium Drought, the dry spell of 2012–2015, and the Tinderbox Drought of 2017–2019 (BOM 2020a, 2020b; Devanand et al. 2024), which have limited the occurrence of flooding events necessary for tree recruitment. The northern MDB experienced its lowest recorded rainfall in 2019 during the Tinderbox Drought, surpassing even the Federation Drought of 1900–1902 (BOM 2020b). Ngugi et al. (2022) highlighted the severe impacts of these droughts on floodplain tree recruitment through demographic assessments of river red gum, coolibah (Eucalyptus coolabah Blakely & Jacobs), and black box (Eucalyptus largiflorens F.Muell.) populations in the Queensland Murray–Darling Basin (QMDB). Their study reported significantly fewer juvenile trees than would be expected in a reverse J-shaped demographic curve. The expected pattern in healthy, self-sustaining tree populations is where there are abundant younger individuals and progressively fewer older ones creating a J-shape when graphed by age class.
Factors including water availability, seed predation, poor seedling survival and browsing have been reported to contribute to poor recruitment of floodplain trees species in the MDB (Moxham and Kenny 2024), potentially threatening long-term population viability as older trees die without sufficient replacement (George et al. 2005). Multiple studies and field observations have reported minimal recruitment across the MDB floodplains for over two decades (George et al. 2005; Woods et al. 2012). In addition to poor recruitment impairing the reverse J-shaped demographic structure of floodplain tree species, Ngugi et al. (2022) observed that floodplain tree stands frequently exhibited mosaics of evenly sized tree cohorts, suggesting episodic recruitment in response to specific rainfall events in the past. For example, Good et al. (2012) reported that following floods in the 1970s, dense regeneration of Eucalyptus coolabah occurred in New South Wales (NSW), Australia, reaching such prevalence that the species was classified as ‘invasive’ under state regulations.
Knowledge gaps remain regarding both the frequency of successful recruitment events for river red gum and coolibah floodplain species (Horner et al. 2016; Moxham et al. 2024) and whether the observed recruitment deficit represents a transient state linked to the sporadic occurrence of extended rainfall events within these long-lived species’ lifecycles, or a more concerning response to climate-change impacts. Opportunistic studies by Westbrooke et al. (2005) and Westbrooke and Florentine (2005) following an episodic high-rainfall and flood event in NSW in February 1997 documented that eucalypt germination occurred exclusively within flooded areas. In 2020, the QMDB experienced localised, high-density germination of floodplain tree seedlings, particularly river red gum and coolibah, following widespread rainfall and drought-breaking floods (BOM 2020b; Ngugi et al. 2022). This initial recruitment event was subsequently augmented by three consecutive La Niña events (2020–2022), creating prolonged wet conditions and multiple flood events across the region (Gillett and Taschetto 2022). Such ‘triple-dip’ La Niña conditions, characterised by three consecutive summers of La Niña influence, represent a rare climatic phenomenon that has occurred only twice since the 1950s (1973–1976 and 1998–2001) (Gillett and Taschetto 2022). These exceptional climatic circumstances presented a unique research opportunity to investigate the impact of sustained rainfall on floodplain tree species recruitment throughout the QMDB.
This study investigated the natural recruitment (seedling and sapling abundance) dynamics of river red gum and coolibah trees across the QMDB following three consecutive La Niña events (2020–2022). We hypothesised that (1) seed germination, recruitment, and early-stage survival of coolibah and river red gum depend critically on a major flood and multiple rainfall events, (2) recruitment is significantly inhibited by competition from ground vegetation and herbivory pressure, and (3) QMDB-wide recruitment episodes have the potential to replenish juvenile cohorts within population demographic structures, thereby enhancing long-term population viability. Field assessments were conducted within floodplains of major river systems in the QMDB: Border Rivers and Moonie; Condamine, Maranoa and Balonne; and Warrego, Paroo, and Nebine.
Methods
Study area and sampling
This study was conducted in the Queensland section of the northern MDB, encompassing the following three geographical regions that have their own water resource management plans (water plan areas, referred to here as catchments) under Queensland’s Water Act 2000: the Border Rivers and Moonie (Lobegeiger and Prior 2022a), the Condamine–Balonne (Lobegeiger and Prior 2022b), and the Paroo, Warrego and Nebine (RDMW 2022) (Fig. 2). The climate across the study area ranges from subtropical to semi-arid and is characterised by warm and wet summers and mild, dry winters. Average annual rainfall and mean monthly temperature ranged from 654 mm and 18.3°C in the Border Rivers catchment in the east, to 345 mm and 21.5°C in the Paroo River catchment in the west (BOM 2024).
Locations of plots surveyed in 2023 (total of 103 plots), showing plots where recruitment was recorded (green markers) and plots without recruitment in the Queensland Murray–Darling Basin. Catchment boundaries (black lines) represent Border Rivers and Moonie, Condamine and Balonne, and Warrego, Paroo, and Nebine.
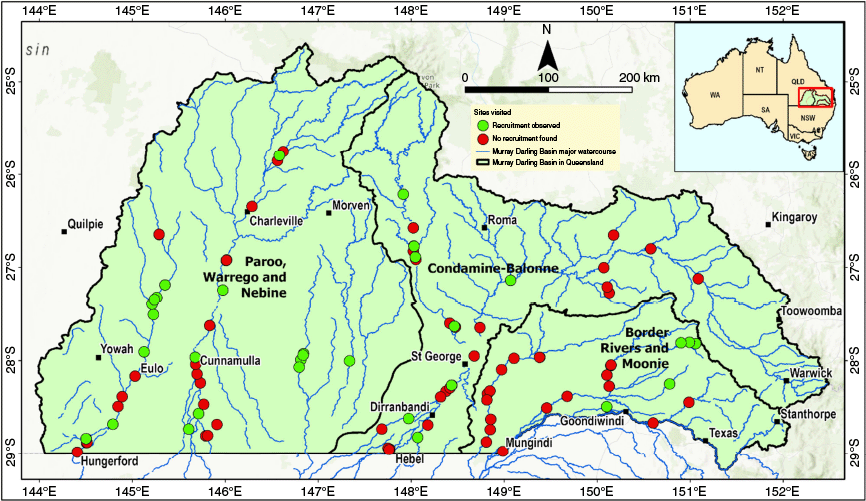
In 2023, the 131 plots previously used for vegetation condition and tree demographic assessments in 2017–2020 (Ngugi et al. 2022), were overlaid with regional ecosystem maps (Queensland Herbarium 2021), showing the spatial distribution of river red gum and coolibah within the Basin. During initial 2020 field assessments (Ngugi et al. 2022), new recruitment was observed at locations not represented in the original sampling design. Hence, to capture the full extent of recruitment patterns following the triple La Niña event, additional plots were strategically located using opportunistic sampling to target areas where active recruitment was evident. This adaptive sampling approach ensured comprehensive documentation of recruitment episodes across the study region, while acknowledging potential bias towards areas with visible recruitment activity. Owing to the linear nature of riparian vegetation stands (typically <50 m width), and to capture the diversity of riverine vegetation, a 25 m × 100 m (0.25 ha) plot was established and oriented parallel to the river channel. In locations where recruitment was confined to specific microenvironments (e.g. water ponds, creek edges, or channels), plot dimensions were reduced and scaled to best survey the recruitment, with the smallest plot measuring 50 m × 5 m. Within each plot, all seedlings (<0.3 m tall), saplings (0.3–2 m tall), and young trees (>2 m tall and <10 cm diameter at breast height [DBH]) were recorded.
Plot attributes potentially influencing recruitment were documented (Supplementary Table S1). Grazing pressure from native animals (kangaroos, wallabies, and other native herbivores) and non-native animals (cattle, sheep and goats) was assessed. Evidence of grazing by native and stocked animals was distinguished using exclusion fencing from adjacent properties, as well as other indicators such as footprints, tracks, and scats to determine which animals were consuming the recruits. Grazing pressure was assessed using visual indicators and classified into the following four categories: high (widespread evidence of trampling, bare ground, dung and damaged vegetation), medium (some evidence of trampling, dung and disturbance), low (minimal signs of trampling, dung or disturbance), or absent (no trampling, no dung or disturbance). Pig disturbance was also assessed as presence/absence. Additional variables recorded included soil texture (assessed by feel method by using the categories clay, loam, or sandy), inundation depth (assessed visually by bark discolouration on trees as indicators of previous flood height and classified as low [<0.5 m], medium [0.5–2 m], or high [>2 m]). The relative percentages of ground cover vegetation, plant litter, bare ground, rocks and stones, and coarse woody debris (fallen stems lying on the soil surface of >10 cm diameter) were assessed within 1 m × 1 m quadrats placed at 10 m intervals along the centreline of each plot, and relative percentages were averaged to provide site-level estimates. Quadrat spacing was maintained at 10 m intervals irrespective of the plot length. In plots with high seedling and sapling density, these quadrats were also used to estimate seedling and sapling abundance. The plots where seedlings and saplings were recorded were re-visited in September 2024 to monitor survival.
Rainfall and hydrology data
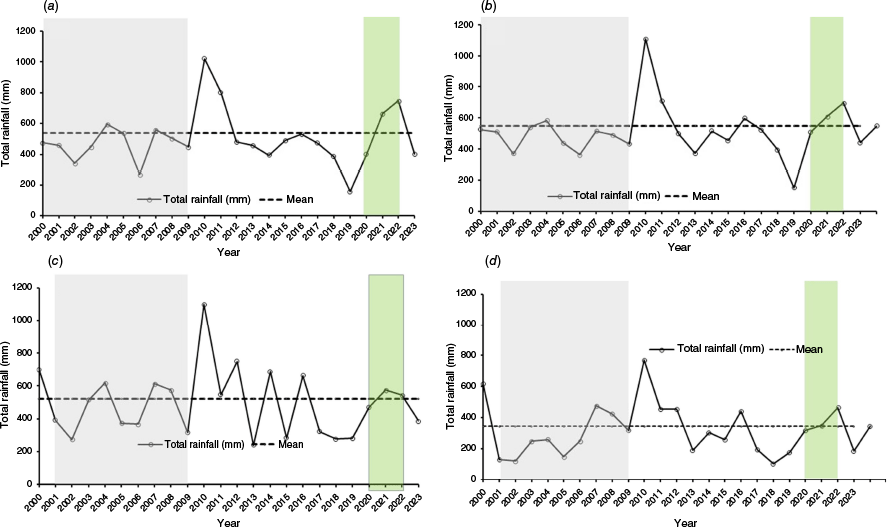
Rainfall data for a monitoring plot within each water plan area for the period between 2000 and 2023 were obtained from the SILO climate database (DESI 2024). Exceptional rainfall and floods in 2010–2011 across multiple catchments and the wet period during 2020–2022, corresponding to the triple-dip La Niña event, are shown in Fig. 3. These charts illustrate the temporal variability in rainfall and the wet periods that coincide with recruitment episodes observed in floodplain tree species.
Annual rainfall patterns (2000–2023) across four catchments in the Queensland Murray–Darling Basin: (a) Border Rivers and Moonie (Moonie4), (b) Condamine–Balonne (251), (c) Warrego (Augathella_2), and (d) Paroo (WP 11). Solid lines with circles represent annual total rainfall (mm), and dashed horizontal lines indicate long-term means for each catchment. Note the millennium drought 2000–2009 (grey shaded), exceptional rainfall in 2010–2011 (double year La Niña) across multiple catchments and the extended wet period during 2020–2022 (green shaded) corresponding to the triple-dip La Niña event.
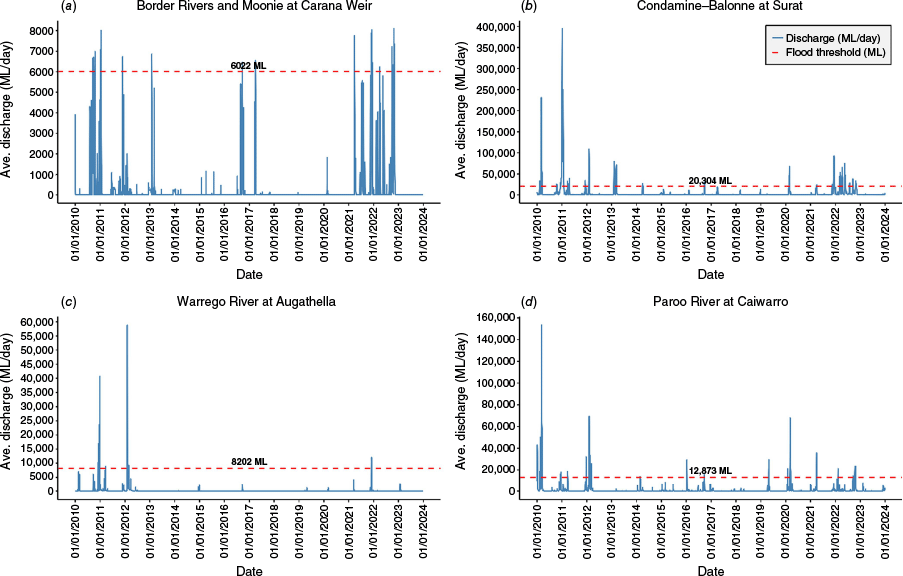
Hydrological daily stream discharge data were obtained from the Queensland Government surface water gauging station network through the Water Monitoring Information Portal (WMIP) (RDMW 2024). Field survey plots were associated with relevant gauging stations by intersecting plot locations with floodplain assessment reaches (FARs). These spatial units delineate river channels and floodplain areas where hydrological conditions are represented by flows at a specific gauge (DSITIA 2013; DES 2018a, 2018b, 2018c). Daily flow data from one gauging station within each water catchment area are presented in Fig. 4. To identify the effect of wet La Niña conditions on riverine flow, bankfull flow volume thresholds were determined by identifying the top of the bank from cross sections at the relevant gauge and converting the bank height to stream discharge by using a rating curve (RDMW 2024).
Daily flow data recorded across four water catchment areas in the Queensland Murray–Darling Basin. Bankfull flow threshold, indicating which flow events were associated with overbank flooding, is represented by the orange dashed line: (a) Border Rivers and Moonie at Carana Weir gauging station (416203A), (b) Condamine–Balonne at the Balonne River at Surat gauging station (422220A), (c) Warrego River at Augathella (gauging station 423204A), and (d) Paroo River at Caiwarro gauging station (424201A).
Drought
Drought was assessed using an evapotranspiration deficit index (Narasimhan and Srinivasan 2005). This index estimates climatic water balance and characterises drought severity by integrating rainfall, temperature, soil moisture and evaporation rates, providing a comprehensive assessment of drought conditions at each monitoring plot. The drought index (DI) was calculated as follows:
where DI is the drought index, P the total precipitation (mm) from first of January 2020, to the field sampling date in 2023, and E the total daily Morton’s areal actual evapotranspiration (mm) over the same period. Daily precipitation and Morton’s areal actual evapotranspiration data were obtained from the SILO climate database (DESI 2024).
The DI ranges from −1 to 1, with negative values indicating drier conditions and positive values indicating wetter conditions. DI = 1 occurs when evapotranspiration is zero (E = 0), indicating extremely wet conditions. Conversely, DI = −1 occurs when precipitation is zero (P = 0), indicating extremely dry conditions. A value of DI = 0 indicates neutral conditions, where total precipitation equals total evapotranspiration (P = E).
Statistical analysis
Analyses of daily stream discharge data (m3/s) obtained from WMIP between 2019 and 2023 and multiple linear regression analyses were performed using R (ver. 4.2.2; R Core Team 2022) and RStudio (ver. 2022.12.0; RStudio Team 2024), and the ‘tidyverse’ (ver. 2.0.0, https://tidyverse.tidyverse.org; Wickham et al. 2019) and ‘lubridate’ (ver. 1.9.4, https://lubridate.tidyverse.org; Grolemund and Wickham 2011) packages. Data for three hydrological attributes were derived from WMIP: the number of rainfall events (i.e. multiple consecutive days) that exceeded river bankfull thresholds; total count of days that the threshold was exceeded; and the count of days since the last flood occurred.
All seedling and sapling data collected at each plot were standardised to per hectare values. The relationships between seedling and sapling density (trees/ha) and plot factors, including species variables, hydrological features and variables, vegetation structure attributes, soil texture attributes, grazing pressure attributes and ground layer proportions (see list of attributes in Supplementary Table S2), were analysed using multiple linear regression to identify significant factors influencing tree recruitment. Categorical variables were incorporated into the models by creating binary proxy variables. For example, grazing pressure from exotic animals (categorised as high, medium, low, or absent) was represented by the following four subvariables: exotic-high, exotic-medium, exotic-low, and exotic-absent. For each plot, the observed category was assigned a value of 1, whereas unobserved categories were assigned 0. This approach allowed investigation of relationships between recruitment and grazing intensity or absence across the dataset.
Seedling and sapling density was log-transformed to address data skewness and improve normality of residuals. To assess linear regression model assumptions and validate our statistical inferences, we conducted regression diagnostics using the ‘lmtest’ package (ver. 0.9-40, https://cran.r-project.org/web/packages/lmtest; Zeileis and Hothorn 2002), the ‘car’ package (ver. 3.1-3, https://cran.r-project.org/web/packages/car) for variance inflation factor (VIF) analysis to detect potential issues with normality, influential observations (wide range of seedling and sapling densities observed), and multicollinearity among the plot factors (Fox and Weisberg 2019), ‘ggplot2’ for data visualisation (ver. 4.0.0, https://ggplot2.tidyverse.org; Wickham 2016), and ‘MASS’ package (ver. 7.3-65, https://cran.r-project.org/web/packages/MASS) for stepwise model selection (Venables and Ripley 2002). Multiple regression models were developed using all potential explanatory variables, allowing simultaneous evaluation of their relative contributions to recruitment variation, and the significance of individual predictors was determined on the basis of statistical significance (P = 0.05).
Fitting species demographic curves and temporal comparison
All tree diameter data for each species across the landscape from 2017 to 2020 assessments (Ngugi et al. 2022) were used as the baseline for demographic comparison. Because that dataset showed significantly fewer juvenile trees than expected in healthy populations, this study targeted assessment of juvenile trees of <10 cm DBH (0–10-cm-diameter class) in 2023. To compare demographic recovery between the two time periods, seedling and sapling data for each species were pooled and analysed by plotting diameter size classes against the number of individuals in each class for both the 2017–2020 (pre-La Niña) and 2023 (post-La Niña) datasets. A reversed-J shape, negative exponential curve was fitted for the relationship between tree density (N/ha) and the middle value of the diameter class value, allowing direct comparison of demographic structure between the drought period and post-La Niña period.
Results
Floodplain tree recruitment patterns
In total, 103 plots were surveyed for seedling and sapling abundance as an indicator of floodplain tree recruitment. The occurrence of seedlings and saplings was widespread but patchy, occurring at 34% of plots (Table 1), with distribution across catchments shown in Fig. 1. The frequency and duration of overbank flood events varied significantly across regions. The Border Rivers and Moonie catchment experienced an average of eight flood events, with a mean inundation duration of 63 days (Table 1). The Condamine–Balonne catchment had four flood events, on average, with a mean inundation duration of 50 days. The Warrego, Paroo, and Nebine catchment showed the least riverine flooding, averaging three flood events, with a mean inundation duration of 15 days (Fig. 2, Table 1). Although seedling and sapling heights varied, saplings within the same survey plot were generally of uniform size or cohort, suggesting that recruitment occurred following a flood event (Fig. 5). Mean seedling and sapling density was 277 trees/ha, excluding one exceptional plot. Including all the plots, density ranged from 4 to 298,180 stems/ha (Supplementary Table S3). The upper extreme reflects exceptional seedling and sapling abundance at one fenced plot adjacent to Callandoon Lake in the Border Rivers catchment, where river red gum density reached 298,180 stems per hectare within a 20 m × 500 m plot (Fig. 5a, b). Seedlings and saplings were also observed within channel sandy floors, but most of these were washed away by subsequent floods (Supplementary Fig. S1). All the plots where seedlings and saplings were observed in 2023 were revisited in 2024, and seedlings and saplings were found to be thriving and exceeding 2 m in height in 97% of the plots (Supplementary Table S3).
| Water catchment area | Number of plots | Plots with recruits | Proportion of sites with recruits (%) | Number of gauge stations | Average number of inundation days | Number of overbank flood events | |
|---|---|---|---|---|---|---|---|
| Border Rivers and Moonie | 25 | 5 | 20 | 9 | 63 | 8 | |
| Condamine–Balonne | 33 | 9 | 27 | 13 | 50 | 4 | |
| Warrego, Paroo, Nebine | 45 | 21 | 47 | 5 | 15 | 3 | |
| Total | 103 | 35 | 34 | 27 | 43 | 15 |
Recruitment of floodplain tree species in the Queensland Murray–Darling Basin following a rare triple-dip La Niña event (2020–2022): (a) Border Rivers at Callandoon Lake 2023 (river red gum), (b) Border Rivers at Callandoon Lake 2024 (river red gum), (c) Condamine–Balonne at the Maranoa River in 2024 (river red gum), (d) Warrego River at Wyandra in 2024 (coolibah), (e) Nebine Creek (river red gum), and (f) Paroo catchment in 2023 (coolibah).

Grazing evidence and pressure
Grazing by exotic animals (Fig. 6) was detected at 80% of plots, and by native animals at 97% of plots (Table 2). Grazing pressure was variable but generally low in intensity, primarily targeting saplings with abundant new leaves.
Saplings of coolibah (Eucalyptus coolabah) in the Weir River catchment (Border Rivers), showing (a) low grazing inside fenced paddock compared with (b) few recruits in adjacent heavily grazed stock route outside the fence in September 2023 and (c) recovery with no grazing of the same plot in September 2024.

| Water catchment area | Plots (n) | Animal type | Grazing pressure category (%) | |||||
|---|---|---|---|---|---|---|---|---|
| High | Medium | Low | Absent | Grazed | ||||
| Border Rivers and Moonie | 25 | Exotic | 16.0 | 24.0 | 52.0 | 8.0 | 92.0 | |
| Native | 4.0 | 44.0 | 48.0 | 4.0 | 96.0 | |||
| Condamine–Balonne | 31 | Exotic | 9.7 | 38.7 | 32.3 | 19.4 | 80.6 | |
| Native | 3.2 | 25.8 | 67.7 | 3.2 | 96.8 | |||
| Warrego, Paroo, Nebine | 47 | Exotic | 4.3 | 29.8 | 38.3 | 27.7 | 72.3 | |
| Native | 0.0 | 14.9 | 83.0 | 2.1 | 97.9 | |||
| Total sites | 103 | Exotic | 8.7 | 31.1 | 39.8 | 20.4 | 79.6 | |
| Native | 1.9 | 25.2 | 69.9 | 2.9 | 97.1 | |||
Factors influencing juvenile tree abundance
Vegetation structure was a strong predictor of floodplain tree recruitment (Table 3). Open woodland areas (crown cover of ≤20%) exhibited significantly higher recruitment rates than did other habitat types, followed by woodland (crown cover 20–50%) areas. River red gum plots showed marginally higher recruitment compared to coolibah plots, although this effect was less statistically robust. Hydrological features (creek permanence, ephemeral waterways, floodplain position) and soil texture variables showed no significant influence on recruitment patterns, despite their theoretical importance in seedling establishment.
| Attribute | Coefficient | Standard error | t-statistic | P-value | |
|---|---|---|---|---|---|
| Intercept | −0.717 | 1.270 | −0.564 | 0.574 | |
| Species | |||||
| River red gum | 1.132 | 0.652 | 1.736 | 0.086 | |
| Coolibah | 0.188 | 0.646 | 0.291 | 0.772 | |
| RRG and EC mixed | 0.516 | 0.681 | 0.758 | 0.450 | |
| Hydrological features | |||||
| Creek (Permanent) | 0.673 | 1.242 | 0.541 | 0.590 | |
| Ephemeral waterway | −0.021 | 0.934 | −0.022 | 0.982 | |
| Floodplain | 0.460 | 0.972 | 0.473 | 0.638 | |
| Vegetation structure | |||||
| Open woodland | 1.465 | 0.443 | 3.309 | 0.001** | |
| Woodland | 0.907 | 0.384 | 2.359 | 0.021* | |
| Soil texture | |||||
| Clay | −0.079 | 0.398 | −0.198 | 0.843 | |
| Loam | −0.061 | 0.438 | −0.140 | 0.889 | |
| Sandy | 0.115 | 0.621 | 0.185 | 0.854 | |
| Model statistics | |||||
| R-squared | 0.219 | ||||
| Adjusted R-squared | 0.115 | ||||
| F(12,90) = 2.099 | P = 0.025* | ||||
| Observations | 103 | ||||
The species are river red gum (RRG = Eucalyptus camaldulensis) and coolibah (EC = Eucalyptus coolabah and mixed stands of the two species. Asterisks denote significance levels: *P < 0.05; **P < 0.01; ***P < 0.001.
The drought index for the plots ranged from −0.267 to −0.022. This index range was notably less severe (less negative) than the 2018–2019 range (−0.587 to −0.401), the driest (most negative) period in a decade. Among hydrological variables (Table 4), drought index had a significant negative effect, indicating reduced seedling and sapling abundance during drought, whereas days since last flood showed a significant positive relationship, suggesting that germination and establishment increase after inundation (Table 4). The number of bankfull exceedance events showed a marginally positive effect, whereas total days of exceedance had a marginally negative influence, suggesting that the frequency of flooding events may be more important for seedling and sapling establishment than is flood duration. Although only 53 plots had a corresponding gauging station, results suggest that observed seedling and sapling abundance was linked to flooding events.
| Variable | Coefficient | Standard error | t-statistic | P-value | |
|---|---|---|---|---|---|
| Model: R2 = 0.299, Adjusted R2 = 0.116, F(11,42) = 1.632, P = 0.125 | |||||
| Intercept | −3.483 | 1.723 | −2.021 | 0.049* | |
| Hydrological variables | |||||
| Drought index | −16.280 | 6.562 | −2.482 | 0.017* | |
| Bankfull exceedance events | 0.468 | 0.2624 | 1.788 | 0.080 | |
| Total days of exceedance | −0.051 | 0.026 | −1.948 | 0.057 | |
| Days since last flood | 0.0006 | 0.0003 | 2.157 | 0.036* | |
| Grazing pressure | |||||
| Exotic grazing – absent | −0.109 | 1.910 | −0.057 | 0.955 | |
| Exotic grazing –low | −0.713 | 1.800 | −0.396 | 0.694 | |
| Exotic grazing – medium | −1.585 | 1.609 | −0.985 | 0.330 | |
| Native grazing – absent | 1.645 | 3.018 | 0.545 | 0.588 | |
| Native grazing –low | 2.354 | 0.838 | 2.810 | 0.007** | |
| Native grazing – high | 2.009 | 2.371 | 0.847 | 0.402 | |
| Pig – presence | −0.800 | 1.120 | −0.714 | 0.479 | |
| Ground layer attributes R2 = 0.166, Adjusted R2 = 0.047, F(5,35) = 1.392, P = 0.251, n = 41 | |||||
| Intercept | −296.600 | 316.200 | −0.938 | 0.355 | |
| Vegetation cover (%) | 2.968 | 3.161 | 0.939 | 0.354 | |
| Bare ground cover (%) | 3.020 | 3.161 | 0.955 | 0.346 | |
| Litter cover (%) | 3.026 | 3.158 | 0.958 | 0.345 | |
| Rock cover (%) | 3.033 | 3.162 | 0.959 | 0.344 | |
| Non-natives cover (%) | −0.008 | 0.065 | −0.123 | 0.903 | |
Significance levels: *P < 0.05; **P < 0.01; ***P < 0.001.
Low native grazing pressure emerged as a strong positive predictor of seedling and sapling abundance, with plots assessed as experiencing low native herbivory exhibiting substantially higher abundance (Table 4). Neither exotic grazing pressure nor pig presence significantly affected seedling and sapling density during prolonged wet conditions. Ground cover attributes (vegetation cover, bare ground cover, litter cover, and rock cover) were not significantly associated with seedling and sapling abundance (Table 4).
Species demographic structure
Field assessments in 2023/2024, following the 2020–2023 La Niña events, showed moderate seedling and sapling establishment in the <10-cm-diameter class (Fig. 7) for river red gum and coolibah. This was a substantial increase when compared with the abundance of trees in this size class prior to these events.
Tree size distribution of river red gum and coolibah recruits (<10-cm diameter at breast height, 1.3 m) recorded in 2023 across the Queensland Murray–Darling Basin. Error bars represent standard errors.
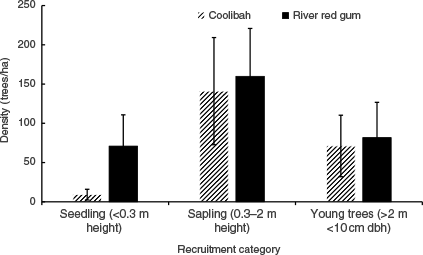
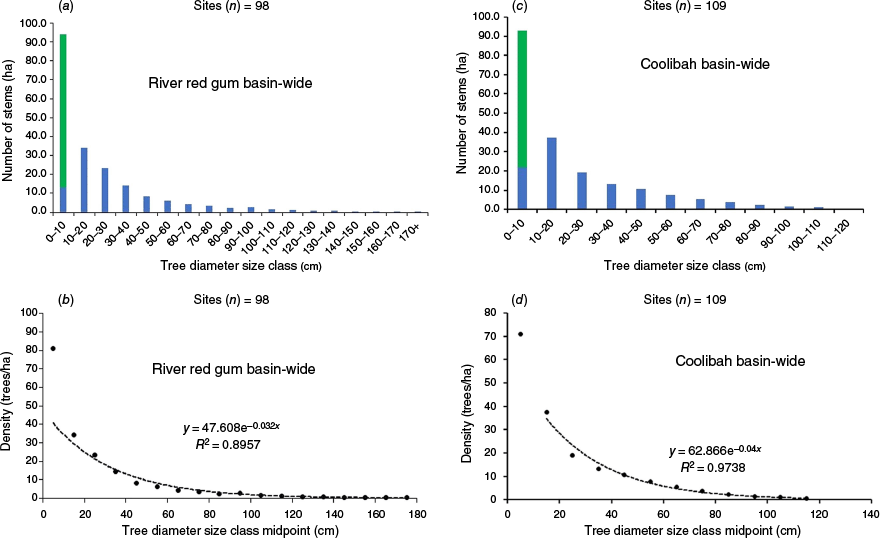
When species population data were categorised by demographic size, seedlings and saplings had the highest density, suggesting a strong likelihood of progressing to young trees under favourable growth conditions (Fig. 7). Using data from young trees (height of >2 m, DBH of <10 cm), reconstructed demographic structure curves for river red gum and coolibah showed an improved alignment to the reverse J-curve, indicating a more stable population structure if trees in the smallest size classes continue to thrive (Fig. 8).
Revised tree demographic structure, showing tree size-class distribution observed in 2017–2020 (blue bars) and seedlings and saplings observed in 2023 (green bars) following the triple-dip La Niña event (2020–2022). Reverse J-curves are fitted to size class mid-points for river red gum (a, b) and coolibah (c, d) across the Queensland Murray–Darling Basin.
Discussion
In arid and semi-arid regions, early signs of population decline often include shifts in species demographics, particularly reduced juvenile tree abundance (Ngugi et al. 2022). These dynamics were examined in the context of multiple droughts, followed by multiple consecutive rainfall events during a rare triple-dip La Niña (2020–2022). The duration of favourable weather conditions, rather than individual high-rainfall events, apparently played a crucial role in driving establishment of young trees.
We first hypothesised that landscape-scale recruitment of floodplain forests in the QMDB is dependent on major floods and extended wet periods. Results showed a more complex picture than initially proposed. Although seedling and sapling density were significantly (P < 0.05) associated with flooding events, observations were consistent with hydrochory (seed dispersal by water), because seeds deposited within flood-disturbed ecological niches showed higher rates of germination and subsequent survival to the seedling and sapling stage at open canopy plots than at plots with denser canopies. High-density recruitment occurred primarily on plots with minimal tree canopy cover, in bands along the banks of water channels or edges of waterbodies, or as randomly scattered patches within channels or on floodplain flats, where slow-moving floods and receding waters had deposited seeds on moist sandy or silty substrates. Hydrochory was also reported for coolibah in the Diamantina and Warburton River systems in north-eastern South Australia, where 70% of coolibah seeds floated for 5 days, after which most began to sink, the majority of which had commenced germination and displayed emergent radicles (Gillen 2017). The study also reported that coolibah seeds dispersed widely during uncommon extensive floods, both downstream and laterally across the adjacent floodplain. Our observations of hydrochory in river red gum populations are also consistent with those reported for major river red gum forests in Victoria and southern New South Wales, where seed dispersal has been reported to be aided by the movement of floodwaters and laboratory tests have indicated that seeds float for up to 36 h (Stefano 2002).
Tranmer et al. (2023) found that increased floodplain inundation led to greater potential for riparian recruitment during wet hydrologic years compared to average years in the South Fork Boise River, USA. However, they determined that inundation alone was a poor metric of seedling and sapling establishment, because establishment requires an appropriate sequence of geomorphic disturbance, seed delivery, and sustained water availability throughout the growing season. Our results align with these findings and the findings of Kerr et al. (2024), who demonstrated that sustained rainfall, rather than flooding alone, is crucial for sapling survival and growth. We observed direct evidence of this process, particularly where river red gum had established on in-channel sand bars along the Maranoa River. At these plots, initial flooding events deposited seeds, followed by sustained rainfall that supported seedling development, without disruption from subsequent major floods.
The spatial patchiness of observed seedling establishment (34% of plots) reflects the variable distribution of rainfall and hydrology across the QMDB, even during the triple La Niña event. This is consistent with the variability in recruitment observed after the 2010–2012 double-dip La Niña (BOM 2012). However, recruitment was extremely low (0–10 saplings) during the 2010–2012 event, except in a few locations of mass recruitment (Capon et al. 2012), whereas our study found more widespread recruitment (0–298,180 stems per hectare). Findings indicated that triple La Niña events (Gillett and Taschetto 2022) may provide opportunities for recruitment ‘reset’ at QMDB-wide scale, characterised by mass recruitment of evenly sized tree cohorts, whereas single La Niña events (BOM 2012) typically result in smaller, more localised recruitment events.
Second, we hypothesised that during long wet periods, seedling and sapling establishment is hampered by competition with ground vegetation and grazing pressure. Contrary to expectations, this relationship was shown to be complex. Ground vegetation cover, primarily grasses, did not significantly affect seedling and sapling density. However, during the dry winter months, the ground cover provided by dense drying grass may have enhanced the survival of coolibah seedlings, shading the soil and reducing evaporation (Good et al. 2014; Moxham et al. 2024), while providing alternative forage. Although grazing by both native and exotic animals can significantly affect seedling and sapling establishment (Stefano 2002; Dorrough and Moxham 2005), we observed minimal browsing on seedlings and saplings regardless of their height where grass was abundant. This suggests that abundant grass growth during the triple La Niña period provided alternative forage for herbivores, reducing browsing pressure on tree seedlings and saplings. However, rapid grass growth during the extended wet period of the triple La Niña event is likely to have created intense competition for water and nutrients, and increased the susceptibility of seedlings and saplings to potential bushfire events. This competition is likely to have slowed the growth of some seedlings and prevented them from reaching heights that would reduce their vulnerability to grazing. Both native and exotic animals can affect seedling and sapling abundance through browsing, grazing and trampling, particularly during short dry spells. We observed this impact at one plot where saplings growing at the edge of a drying waterbody were heavily browsed as animals came looking for water. This vulnerability to browsing and grazing, which can prevent seedlings from developing into saplings with foliage above browsing height (approximately 2 m), is consistent with other studies (Li et al. 2003; Reid et al. 2011; Horner et al. 2016).
Third, we hypothesised that a QMDB-wide seedling and sapling establishment episode has the potential to replenish the juvenile age classes of floodplain tree species that have been negatively affected by drought, increasing the proportion of young trees in the population and enhancing the viability of these populations. The observed QMDB-wide spatial distribution of seedling and sapling establishment, albeit patchy, supports this hypothesis. The better fit to reverse J-curve distributions for both river red gum and coolibah populations represents a substantial change from the findings of Ngugi et al. (2022), which reported fewer juvenile trees than expected in a typical reverse J-curve distribution. This change suggests that wide-spread establishment episodes, such as those triggered by triple-dip La Niña events, may be crucial for maintaining population viability over long time scales (Good et al. 2012). However, the demographic results must be interpreted cautiously because the observed juvenile tree size distribution is heavily skewed toward saplings (<2-cm diameter).
The extremely high seedling and sapling density observed near Callandoon Lake (298,180 saplings per hectare) demonstrates the remarkable potential for population regeneration under optimal germination conditions. This finding is consistent with dense germination of coolibah reported in NSW in the 1970s on cleared agricultural lands (Westbrooke and Florentine 2005; Good et al. 2012, 2014). However, the long-term persistence of these seedlings and saplings will depend on their capacity to develop drought tolerance mechanisms, particularly extensive root systems to access groundwater during low-rainfall periods (Balcombe et al. 2021). Management interventions such as prescribed thinning, along with natural self-thinning processes, will also be necessary to reduce competition associated with high stem density and achieve sustainable tree densities. Despite these challenges, the recent establishment associated with 3 years of sustained rainfall has shifted the demographics from a risky state depauperate of juvenile trees to a potentially more sustainable distribution.
In addition to the development of a sapling’s capacity to adapt to low moisture conditions and access groundwater resources during drought, bankfull flows are important for maintaining river red gum condition along the Murray River through lateral recharge of riparian soils (Doody et al. 2014). Although such flows could benefit established saplings, our findings suggest they may not substantially promote new seedling and sapling establishment. This understanding is particularly relevant given that mass recruitment was often found adjacent to temporary waterbodies and floodplain wetlands rather than permanent waterbodies. The establishment observed following the triple-dip La Niña event, only the third such occurrence since the 1950s, suggests that these climatic events could be crucial in maintaining population viability over the extended lifespans of these species. This challenges previous concerns that low establishment might represent a transient state, or a response to changing climate, and suggests that population maintenance may depend on these irregular but significant establishment episodes.
The significant association between seedling and sapling abundance and open woodland and woodland land systems, but not permanent waterbodies, adds important nuance to the understanding of regeneration dynamics, suggesting that these areas are particularly important for seedling and sapling establishment. This pattern is likely to reflect the different recruitment niches operating across the structural continuum from open woodland to forest systems. Open woodlands provide optimal conditions for seedling and sapling development through reduced competition for light and moisture, while allowing sufficient seed sources from mature trees. Conversely, denser forest systems may inhibit establishment through increased competition and reduced light penetration to the forest floor.
Although recruitment episodes, like those documented in this study, are clearly important, other demographic factors also influence long-term population viability. For example, tree condition affects reproductive output and seed production across these structural types, because healthier trees in open woodland systems may contribute greater seed fall and enhanced recruitment potential (Jensen 2008; Jensen et al. 2008; Kerr et al. 2024). Our adaptive sampling approach, although necessary to capture recruitment patterns comprehensively, introduced potential bias toward areas with visible recruitment activity, which may have overestimated recruitment rates across the broader landscape. Visual assessment methods for grazing pressure, soil texture, and inundation depth, although standardised through categorical classifications, introduced subjective interpretation that may have affected reproducibility between observers. Our study focused on a single post-La Niña assessment period, limiting the ability to assess longer-term recruitment persistence and survival rates, although the 2024 follow-up monitoring provides some insight into short-term survival. Consequently, the observational nature of this study limits the ability to establish definitive causal relationships between environmental variables and recruitment, because correlative analyses cannot account for all potential confounding factors.
Implications for water management
These findings have implications for environmental flow management and conservation strategies in the QMDB, contribute to refining concepts of ecological resilience in the Murray–Darling Basin Plan, and can inform future water management strategies. Both flooding, which aids seed dispersal and germination, and sustained rainfall, which supports seedling growth and establishment, are essential for large-scale floodplain tree recruitment. However, recruitment patterns remain patchy, influenced by multiple environmental factors including variable rainfall and water availability, but also potentially reflecting spatial variation in flowering, seed production, seed dispersal patterns, and differential seedling mortality from grazing or other stressors. Although the Murray–Darling Basin Plan and related strategies aim to improve floodplain vegetation through flow management, this study demonstrated that achieving these outcomes depends on a combination of conditions, including climatic factors that cannot be regulated through environmental water planning. Land-use factors such as managing grazing intensity and soil conservation following rainfall events that trigger recruitment events could complement water strategies (Dauwalter et al. 2018), but existing programs are often underfunded and lack coordination (Metcalfe and Costello 2021). Additionally, the requirement for sustained rainfall at key growth stages further underscores the limits of direct management interventions. Water management strategies should also consider that the germination phase and the longer-term survival and maintenance of tree health may require different water regimes, with early life stages benefiting from shorter-duration, widespread flooding events, whereas established trees may require longer-duration floods to recharge groundwater resources needed for maintaining physiological health.
The observed complex interactions among flow, climate, and habitat conditions suggest the need for realistic expectations about what flow management alone can achieve in supporting floodplain vegetation recruitment. Management strategies should consider multiple factors beyond flow management alone. The timing and duration of environmental flows should aim to mimic the sustained moisture conditions that support seedling and sapling establishment (noting that managing high flows onto floodplains while avoiding negative impacts is challenging). Management of grazing pressure must be coordinated with natural recruitment episodes. Protection of woodland and open woodland systems is crucial, because these systems showed stronger associations with seedling and sapling establishment than did forested riparian forests. Conservation strategies should recognise that infrequent but significant recruitment episodes may be sufficient for maintaining population viability. Ultimately, although water management plays a role in supporting floodplain ecosystems, broader climatic and environmental conditions shape long-term recruitment.
Tracking the survival and development of these recruitment cohorts is needed, particularly given the variable grazing pressures and potential for future drought. This should include both long-term monitoring and controlled experimental approaches to establish causal relationships rather than correlations. Specifically, manipulative experiments such as light attenuation treatments to simulate canopy competition effects, provision of constant water regimes to isolate hydrological factors, and exclusion fencing to quantify herbivory impacts could help disentangle the complex drivers of seedling establishment and persistence. Long-term monitoring coupled with these experimental interventions will be crucial for understanding whether these recruitment episodes are sufficient for maintaining population viability in the face of changing climatic conditions and increasing pressures on water resources in the Murray–Darling Basin.
Conclusions
This study has provided several key technical insights into floodplain forest regeneration following extreme climatic events. Triple La Niña events appear to create fundamentally different recruitment opportunities from those created by shorter La Niña cycles, with establishment occurring at 34% of plots and densities reaching up to 298,180 saplings per hectare under optimal conditions. The rarity of triple La Niña events (only three occurrences since the 1950s) suggests that floodplain tree populations may naturally depend on infrequent but massive establishment episodes rather than consistent annual recruitment.
Hydrochory (water-mediated seed dispersal) emerged as a critical mechanism, with seeds establishing preferentially in flood-disturbed areas with open canopies rather than dense forest understories. Importantly, sustained rainfall following initial flooding was shown to be more crucial for seedling survival than were flood events alone, highlighting the importance of extended favourable conditions rather than single precipitation events.
The study demonstrated that abundant grass growth during wet periods can simultaneously protect seedlings from herbivory (by providing alternative forage), while creating competition for resources. Open woodland systems consistently showed higher establishment rates than either dense forests or permanent waterbodies, suggesting that intermediate disturbance and light conditions optimise regeneration.
These findings challenge conventional assumptions about ‘recruitment failure’ in long-lived tree species, suggesting, instead, that episodic establishment during rare climatic events may be sufficient for population maintenance. For floodplain ecosystems globally, this implies that management strategies focused on facilitating consistent annual recruitment may be less effective than is protecting conditions that enable establishment during infrequent but significant climatic events.
Data availability
Access to the data used in this study are available from the corresponding author upon reasonable request.
Declaration of funding
This project was funded through the Murray–Darling Basin Authority’s Federation Funding Agreement (MDBA-FFA) administered by the Queensland Department of Local Government, Water and Volunteers (DLGWV), and executed by the Queensland Herbarium and Biodiversity Science, Department of Environment, Tourism, Science and Innovation (DETSI).
Acknowledgements
We acknowledge Aboriginal peoples of the Queensland Murray–Darling Basin as the First Nations peoples of the country on which this work took place. We recognise their enduring connection to land, water, sky, Country and community. We pay our respects to them, their cultures, and to their Elders past and present. We acknowledge and thank Peter Brownhalls (DLGWV) for assistance during the funding application process and project administration. We thank Jiaorong Li (DETSI) for assistance in GIS and preparation of maps. We also acknowledge advice provided by participants during consultation presentation and workshop, including Gabrielle Hunt and Cherie Campbell (MDBA), Peter Negus, Jonathan Marshall, and Alisha Steward (DETSI), Andrea Prior (DLGWV) and Andrew Biggs (Department of Resources). Thanks go to Keenan Williams (Industry placement student, Griffith University), Georgia Daley (Industry placement student, University of Sunshine Coast) and Enoch Chow (Volunteer) for data collation assistance. We gratefully acknowledge Jonathan Marshall, Peter Negus, Sara Clifford and Geoffrey Smith (DETSI) who provided review, comments and suggestions that have helped improve this paper. We are grateful to the three anonymous reviewers and Dr Paul Novelly, the Editor-in-Chief, for their detailed and insightful comments that greatly enhanced this work. Their constructive feedback was invaluable in strengthening the paper.
References
Balcombe SR, Biggs A, Prior A, Capon SJ (2021) Seedling root growth experiments for two Australian dryland riparian eucalypts provide new insights for environmental watering. River Research and Applications 37(10), 1463-1470.
| Crossref | Google Scholar |
BOM (2012) Record-breaking La Niña events. An analysis of the La Niña life cycle and the impacts and significance of the 2010–11 and 2011–12 La Niña events in Australia. Bureau of Meteorology. Available at http://www.bom.gov.au/climate/enso/history/La-Nina-2010-12.pdf [accessed 30 April 2025]
BOM (2020a) ‘Trends and historical conditions in the Murray–Darling Basin.’ (Bureau of Meteorology) Available at https://www.mdba.gov.au/sites/default/files/publications/bp-eval-2020-bom-trends-and-historical-conditions-report.pdf [accessed 30 April 2025]
BOM (2020b) ‘Special Climate Statement 70 update – drought conditions in Australia and impact on water resources in the Murray– Darling Basin.’ (Australian Bureau of Meteorology: Canberra, ACT, Australia) Available at http://www.bom.gov.au/climate/current/statements/scs70.pdf [accessed 30 April 2025]
BOM (2024) ‘Climate data online.’ Monthly rainfall and temperature data for Inglewood (station 041047) in the Border Rivers, and Kahmoo (station 044150) in the Paroo catchment (Australian Bureau of Meteorology) Available at http://www.bom.gov.au/climate/data/ [accessed 30 April 2025]
Capon SJ, Chambers LE, Mac Nally R, Naiman RJ, Davies P, Marshall N, Pittock J, Reid M, Capon T, Douglas M, Catford J, Baldwin DS, Stewardson M, Roberts J, Parsons M, Williams SE (2013) Riparian ecosystems in the 21st Century: hotspots for climate change adaptation? Ecosystems 16(3), 359-381.
| Crossref | Google Scholar |
Capon SJ, Rolls R, James C, Mackay S (2012) Regeneration of floodplain vegetation in response to large-scale flooding in the Condamine–Balonne and Border Rivers. Final report. Australian Rivers Institute, Griffith University, Qld. Available at https://www.researchgate.net/publication/267625845_Regeneration_of_Floodplain_Vegetation_in_Response_to_Large-scale_Flooding_in_the_Condamine-Balonne_and_Border_Rivers
Cooper DJ, Andersen DC, Chimner RA (2003) Multiple pathways for woody plant establishment on floodplains at local to regional scales. Journal of Ecology 91(2), 182-196.
| Crossref | Google Scholar |
Cunningham SC, Thomson JR, Mac Nally R, Read J, Baker PJ (2011) Groundwater change forecasts widespread forest dieback across an extensive floodplain system: groundwater change forecasts forest dieback. Freshwater Biology 56(8), 1494-1508.
| Crossref | Google Scholar |
Dauwalter DC, Fesenmyer KA, Miller SW, Porter T (2018) Response of riparian vegetation, instream habitat, and aquatic biota to riparian grazing ex closures. North American Journal of Fisheries Management 38(5), 1187-1200.
| Crossref | Google Scholar |
DES (2018a) Review of Water Plan (Border Rivers) 2003. Ecological Risk Assessment Report. Department of Environment and Science, Queensland Government. Available at https://www.mdba.gov.au/sites/default/files/publications/qld-border-rivers-ecological-risk-assessment-report-2018.pdf
DES (2018b) Review of Water Plan (Condamine and Balonne) 2004. Ecological Risk Assessment Report. Department of Environment and Science, Queensland Government. Available at https://www.mdba.gov.au/sites/default/files/publications/qld-condamine-balonne-ecological-risk-assessment-report-2018.pdf
DES (2018c) Review of Water Plan (Moonie) 2003. Ecological Risk Assessment Report. Department of Environment and Science, Queensland Government. Available at https://www.mdba.gov.au/sites/default/files/publications/qld-moonie-ecological-risk-assessment-report-2019.pdf
DESI (2024) Scientific Information for Landowners (SILO). Available at https://www.longpaddock.qld.gov.au/silo/ [accessed 30 April 2025]
Devanand A, Falster GM, Gillett ZE, Hobeichi S, Holgate CM, Jin C, Mu M, Parker T, Rifai SW, Rome KS, Stojanovic M, Vogel E, Abram NJ, Abramowitz G, Coats S, Evans JP, Gallant AJE, Pitman AJ, Power SB, Rauniyar SP, Taschetto AS, Ukkola AM (2024) Australia’s Tinderbox drought: an extreme natural event likely worsened by human-caused climate change. Science Advances 10(10), 3460.
| Crossref | Google Scholar | PubMed |
Doody TM, Benger SN, Pritchard JL, Overton IC (2014) Ecological response of Eucalyptus camaldulensis (river red gum) to extended drought and flooding along the River Murray, South Australia (1997–2011) and implications for environmental flow management. Marine and Freshwater Research 65(12), 1082.
| Crossref | Google Scholar |
Dorrough J, Moxham C (2005) Eucalypt establishment in agricultural landscapes and implications for landscape-scale restoration. Biological Conservation 123(1), 55-66.
| Crossref | Google Scholar |
DSITIA (2013) ‘Review of Water Resource (Warrego, Paroo, Bulloo and Nebine) Plan 2003 and Resource Operations Plan: environmental risk assessment for selected ecological assets.’ (Department of Science, Information Technology, Innovation and the Arts: Brisbane, Qld, Australia) Available at https://www.mdba.gov.au/sites/default/files/publications/16-DSITIA-2013a-Environmental-assessment-stage2-D16-40905.pdf [accessed 30 April 2025]
George AK, Walker KF, Lewis MM (2005) Population status of eucalypt trees on the River Murray floodplain, South Australia. River Research and Applications 21(2–3), 271-282.
| Crossref | Google Scholar |
Gillen JS (2017) ‘Coolibah (Eucalyptus coolabah Blakely & Jacobs) of the Diamantina and Warburton River systems in northeastern South Australia.’ (Fenner School of Environment and Society Australian National University, South Australian Arid Lands Natural Resources Management Board: Pt Augusta, SA, Australia) Available at https://cdn.environment.sa.gov.au/landscape/docs/saal/2_coolibah_final_hmeditfinal_1.pdf [accessed 30 April 2025]
Gillett Z, Taschetto AS (2022) ‘Multi-year La Niña events. ARC Centre of Excellence for Climate Extremes Briefing Note 20.’ (UNSW Sydney) Available at https://climateextremes.org.au/wp-content/uploads/Multi-year-La-Nina-Events-ARC-Centre-of-Excellence-for-Climate-Extremes.pdf
Good MK, Price JN, Clarke PJ, Reid N (2012) Dense regeneration of floodplain Eucalyptus coolabah: invasive scrub or passive restoration of an endangered woodland community? The Rangeland Journal 34(2), 219.
| Crossref | Google Scholar |
Good MK, Clarke PJ, Price JN, Reid N (2014) Seasonality and facilitation drive tree establishment in a semi-arid floodplain savanna. Oecologia 175(1), 261-271.
| Crossref | Google Scholar | PubMed |
Grolemund G, Wickham H (2011) Dates and Times Made Easy with Lubridate. Journal of Statistical Software 40(3), 1-25.
| Crossref | Google Scholar |
Horner GJ, Cunningham SC, Thomson JR, Baker PJ, Mac Nally R (2016) Recruitment of a keystone tree species must concurrently manage flooding and browsing. Journal of Applied Ecology 53(3), 944-952.
| Crossref | Google Scholar |
Jensen AE, Walker KF, Paton DC (2008) The role of seedbanks in restoration of floodplain woodlands. River Research and Applications 24(5), 632-649.
| Crossref | Google Scholar |
Kerr J, Harding D, Fawcett J, Prior A (2024) The phenology of Eucalyptus camaldulensis (Dehnh, 1832) and Eucalyptus coolabah (Blakely & Jacobs, 1934) in the northern Murray–Darling Basin and implications for recruitment on floodplains. Marine and Freshwater Research 75(12), MF24034.
| Crossref | Google Scholar |
Kingsford RT (2000) Ecological impacts of dams, water diversions and river management on floodplain wetlands in Australia. Austral Ecology 25(2), 109-127.
| Crossref | Google Scholar |
Li J, Duggin JA, Grant CD, Loneragan WA (2003) Germination and early survival of Eucalyptus blakelyi in grasslands of the New England Tablelands, NSW, Australia. Forest Ecology and Management 173(1–3), 319-334.
| Crossref | Google Scholar |
Lobegeiger J, Prior A (2022a) Border Rivers and Moonie long-term watering plan. Available at https://nla.gov.au/nla.obj-3139247754/view [accessed 30 April 2025]
Lobegeiger J, Prior A (2022b) Condamine and Balonne long-term watering plan. Available at https://nla.gov.au/nla.obj-3139248537/view [accessed 30 April 2025]
Mac Nally R, Cunningham SC, Baker PJ, Horner GJ, Thomson JR (2011) Dynamics of Murray–Darling floodplain forests under multiple stressors: the past, present, and future of an Australian icon. Water Resources Research 47(6), W00G05.
| Crossref | Google Scholar |
MDBA (2012) ‘Murray-Darling Basin Plan (Basin Plan).’ (Murray-Darling Basin Authority: Canberra, ACT, Australia) Available at https://www.mdba.gov.au/basin-plan-roll-out [accessed 30 April 2025]
MDBA (2024) ‘Plant and animals: The Basin supports a huge variety of birdlife, animals, fish and plants.’ (Murray–Darling Basin Authority: Canberra, ACT, Australia) Available at https://www.mdba.gov.au/basin/plants-and-wildlife [accessed 30 April 2025]
Metcalfe DJ, Costello O (2021) Australia state of the environment 2021: extreme events. Independent report to the Australian Government Minister for the Environment. Commonwealth of Australia: Canberra, ACT, Australia. doi:10.26194/316q-pv32
Moxham C, Kenny SA (2024) Browsing impacts on seedling survival of black box (Eucalyptus largiflorens). Ecological Management & Restoration 25(1), 32-36.
| Crossref | Google Scholar |
Moxham C, Kenny SA, Fanson B (2024) Environmental flows and microsite attributes influence floodplain eucalypt recruitment. Forest Ecology and Management 572, 122330.
| Crossref | Google Scholar |
Narasimhan B, Srinivasan R (2005) Development and evaluation of Soil Moisture Deficit Index (SMDI) and Evapotranspiration Deficit Index (ETDI) for agricultural drought monitoring. Agricultural and Forest Meteorology 133(1–4), 69-88.
| Crossref | Google Scholar |
Ngugi MR, Neldner VJ, Dowling RM, Li J (2022) Recruitment and demographic structure of floodplain tree species in the Queensland Murray–Darling basin, Australia. Ecological Management and Restoration 23(1), 64-73.
| Crossref | Google Scholar |
Queensland Herbarium (2021) Regional Ecosystem Description Database (REDD). Version 12 (March 2021). (Department of Environment and Science: Brisbane, Qld, Australia) Available at https://www.qld.gov.au/environment/plants-animals/plants/ecosystems/descriptions/download [accessed 30 April 2025]
R Core Team (2022) ‘R: a Language and Environment for Statistical Computing.’ (R Foundation for Statistical Computing: Vienna, Austria) Available at https://www.R-project.org/ [accessed 30 April 2025]
RDMW (2022) Warrego Paroo Bulloo and Nebine long-term watering plan. Available at https://nla.gov.au/nla.obj-3139249116/view [accessed 30 April 2025]
RDMW (2024) Water Monitoring Information Portal. Available at https://water-monitoring.information.qld.gov.au/ [accessed 30 April 2025]
Reid MA, Ogden R, Thoms MC (2011) The influence of flood frequency, geomorphic setting and grazing on plant communities and plant biomass on a large dryland floodplain. Journal of Arid Environments 75(9), 815-826.
| Crossref | Google Scholar |
RStudio Team (2024) ‘RStudio: Integrated Development for R.’ (RStudio, PBC: Boston, MA, USA) Available at http://www.rstudio.com [accessed 30 April 2025]
Stefano JD (2002) River red gum (Eucalyptus camaldulensis): a review of ecosystem processes, seedling regeneration and silvicultural practice. Australian Forestry 65(1), 14-22.
| Crossref | Google Scholar |
Tranmer AW, Benjankar R, Vidergar D, Tonina D (2023) Identifying failure mechanisms of native riparian forest regeneration in a variable-width floodplain using a spatially-distributed riparian forest recruitment model. Ecological Engineering 187, 106865.
| Crossref | Google Scholar |
Van Dijk AIJM, Beck HE, Crosbie RS, De Jeu RAM, Liu YY, Podger GM, Timbal B, Viney NR (2013) The Millennium Drought in southeast Australia (2001–2009): natural and human causes and implications for water resources, ecosystems, economy, and society. Water Resources Research 49(2), 1040-1057.
| Crossref | Google Scholar |
Westbrooke ME, Florentine SK (2005) Rainfall-driven Episodic Flood Events: are they a major factor in moulding New South Wales arid land vegetation patterns? Australian Geographer 36(2), 171-181.
| Crossref | Google Scholar |
Westbrooke ME, Florentine SK, Milberg P (2005) Arid land vegetation dynamics after a rare flooding event: influence of fire and grazing. Journal of Arid Environments 61(2), 249-260.
| Crossref | Google Scholar |
Wickham H, Averick M, Bryan J, Chang W, McGowan L, François R, Grolemund G, Hayes A, Henry L, Hester J, Kuhn M, Pedersen T, Miller E, Bache S, Müller K, Ooms J, Robinson D, Seidel D, Spinu V, Takahashi K, Vaughan D, Wilke C, Woo K, Yutani H (2019) Welcome to the Tidyverse. Journal of Open Source Software 4(43), 1686.
| Crossref | Google Scholar |
Zeileis A, Hothorn T (2002) Diagnostic checking in regression relationships. R News 2(3), 7-10.
| Google Scholar |