

The composition, richness, and evenness of seedlings from the soil seed bank of a semi-arid steppe in northern China are affected by long-term stocking rates of sheep and rainfall variation
A. Hu A , J. Zhang A , X. J. Chen A , J. P. Millner B , S. H. Chang A , S. Bowatte A and F. J. Hou A CA State Key Laboratory of Grassland Agro-Ecosystems, Key Laboratory of Grassland Livestock Industry Innovation, Ministry of Agriculture, China, College of Pastoral Agriculture Science and Technology, Lanzhou University, Lanzhou, 730000, Gansu, China.
B Massey University, Palmerston North, New Zealand.
C Corresponding author. Email: cyhoufj@lzu.edu.cn
The Rangeland Journal 41(1) 23-32 https://doi.org/10.1071/RJ18025
Submitted: 6 March 2018 Accepted: 14 December 2018 Published: 22 January 2019
Journal compilation © Australian Rangeland Society 2019 Open Access CC BY-NC-ND
Abstract
The soil seed bank has a large influence on the potential for grassland restoration. This study aimed to characterise the composition, density, richness, and evenness of seedlings emerging from the soil seed bank under different sheep stocking rates, in a summer grazing system, in semi-arid China. Soil was sampled in 2015, a year with extreme drought conditions and in 2016, a normal rainfall year. The soil seed bank was assessed by measuring seedling emergence under laboratory conditions. Comprising 16 species, 85.4% of the seedlings were concentrated within a depth of 0–5 cm. Drought significantly reduced the density and richness of the seedlings. Grazing increased the richness of seedlings by increasing the richness of aboveground species, and grazing significantly reduced the evenness of the seedlings by reducing the evenness of aboveground species. Drought significantly reduced the similarities between the seedlings and the aboveground species, whereas grazing increased similarities in both years. This study revealed that the density and richness of seedlings were higher in higher stocking rate in drought year. We conclude that negative effects on density, richness and evenness of the seedlings caused by drought can be overcome by rotational grazing especially at higher stocking rate.
Additional keywords: biodiversity, drought, grasslands, grazing intensity, seed banks.
Introduction
The soil seed bank comprises seeds on the soil surface and within the soil; it is a repository for the production of subsequent generations of plants, enabling their survival. The soil seed bank of grasslands is an important component of the grassland ecosystem as it is the basis of population settlement, survival, reproduction, and expansion (Milberg and Hansson 1994). It is an important seed source for restoring communities and an important constraint of vegetation dynamics (Gioria et al. 2014; Tessema et al. 2016). The period during which seeds are stored in the soil seed bank (called the latent stage) reflects the history and current situation of a community (Thompson 2000; Kalamees et al. 2012). The size and the composition of the seed bank can be affected by any factors contributing to grassland degradation, reduction in biodiversity or vegetation productivity (Pugnaire and Lázaro 2000; Dreber and Esler 2011; Báldi et al. 2013). Seed production and release are both affected by the distribution of rainfall (Gutterman 2012). The magnitude of the overall heterogeneity in the soil seed bank is strongly influenced by selective grazing and disturbance-related environmental changes (Solomon et al. 2006; Kassahun et al. 2009). Kinloch and Friedel (2005) reported that heavy grazing over several decades affected both the size and composition of the germinable seed bank, but lighter grazing had no detectable effect on the seed bank over a short period. High livestock pressure favours annual species, which are generally more tolerant of intense grazing and treading because of adaptations including prostrate growth and the production of small easily buried seeds (Navie and Rogers 1997). Under moderate grazing on the Loess Plateau, grassland is composed of perennial species, with few annuals (Hu et al. 2015).
The Loess Plateau is in the arid and semi-arid area of northern China, ~640 000 km2 in area and is vulnerable to soil erosion (Hu et al. 2015). Forty percent of the area is grassland and is grazed by livestock (Gao et al. 2011; Chen et al. 2017). Rainfall on the Loess Plateau has decreased in the last 50 years (Sun et al. 2016). Long-term heavy grazing and climate change are considered to be the two main factors contributing to soil erosion, grassland degradation, reduction of biodiversity and vegetation productivity on the Loess Plateau (Liu et al. 2011; Hu et al. 2015; Chen et al. 2017). This experiment was conducted using a long-term summer rotational grazing system and an ungrazed enclosure. The experiment was conducted in 2015, a very dry year and in 2016, a year with normal rainfall, allowing comparison of seedling characteristics after consecutive years of contrasting rainfall. This study characterises and compares the composition, density, species richness, and evenness of seedlings emerging from the seed bank under different stocking rates in a semi-arid grassland over consecutive years. In particular, we investigate whether (1) the density, species richness, and evenness of seedlings emerging from the soil bank after 15 years of rotation grazing are less disturbed by low rainfall; (2) the similarity between the seedlings and aboveground species in years with contrasting rainfall after long-term grazing.
Materials and methods
Site description
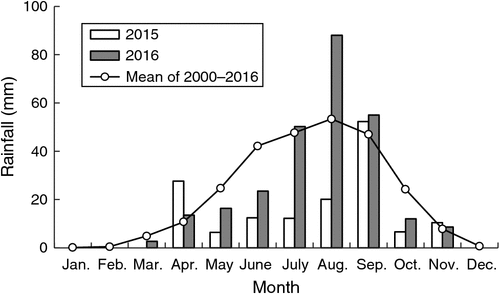
This study was conducted in a long-term sheep grazing system experimental site begun in 2001 (Chen et al. 2010, 2015) in Huanxian County, eastern Gansu Province, North-west China (37°07ʹN, 106°48ʹE, 1700 m a.s.l.). The main land use is pastoral farming basin area. The mean daily air temperature is 7.5°C; winters are long and cold (the frost-free period is 125 days), and summers are hot (mean of annually accumulated temperatures 3633 degree-days above 0°C). The average annual rainfall is 265 mm (2000–2016), but ranges from 148 to 433 mm, (coefficient of variation = 25.6%). Annual rainfall in 2015 was 148 mm; rainfall in 2016 was normal – 270 mm (Fig. 1). More than 70% of rainfall occurs from July to September, typical of a continental monsoon climate. Mean annual evaporation is 1993 mm. The soil is classified as sandy free-draining loess, and the rangeland is typical temperate steppe (Gong et al. 2007; Ren et al. 2008). The dominant species at the study site are perennial low shrubs (Artemisia capillaries Thunb. and Lespedeza davurica (Laxm.) Schindl) and perennial grass (Stipa bungeana Trin.) (Table 1). The biomass of the three dominant species accounts for 70–90% of the total biomass.
|

|
Experimental design
The long-term sheep grazing system experimental site had been rotationally grazed since 2001 by 4-month-old Tan sheep (wethers), a traditional breed in the region. The current experiment was conducted in the summer-grazing only area and included nine paddocks of equal size (50 m × 100 m) for grazing with three separate enclosed areas (50 m × 50 m) utilised as ungrazed controls. Three grazing treatments comprising 4, 8 and 13 sheep corresponding to stocking rates of 2.7, 5.3, and 8.7 sheep ha–1 respectively with three replicates were randomly assigned. Sheep grazed each paddock for 10 days so that 30 days were required to complete a full rotation of all replicates. Three 30-day rotations occurred from June to September each year. The grazing sequence in the three replicate paddocks within a stocking rate treatment was the same each year but the start of the grazing season varied from 1 to 15 June to prevent any rotation effects on plant growth stage.
Sampling
Aboveground vegetation
To compare the species emerging from the soil seed bank with the established plant species, the latter were assessed during the peak growing season (mid-August) in both 2015 and 2016. Four quadrats (1 m2) were randomly established within each paddock, totaling 48 (9 grazing plots × 4 quadrats + 3 ungrazed plots × 4 quadrats) samples per year. In each quadrat, plant species and density were identified (Cheng and Zhu 2012) and recorded, and samples of the tillers or branches and the fertile stems of each species were collected and subsequently dried at 65°C to a constant weight to determine dry matter.
Soil sampling
Soil sampling was carried out after seed production in mid-November of 2015 and 2016. Twelve soil samples per paddock were collected from grazed paddocks. In each paddock soil samples were collected from 10-cm × 10-cm areas within three transact lines spaced 10 m apart. Four soil samples spaced 20 m apart were collected from each line. From the enclosed (ungrazed) plots, four soil samples were collected from two transact lines spaced 20 m apart and two soil samples spaced 10 m were collected within each line. The soil samples were collected from 0–5-cm and 5–10-cm depths (Liu et al. 2011; Tessema et al. 2012). The soil samples collected at the same depth within a paddock were pooled and mixed. Each soil sample was then divided into two equal parts for germination testing and soil moisture analysis. The gravimetric soil moisture was determined by oven drying a known moist soil weight for 48 h at 105°C. The soil samples selected for germination testing were further divided into two equal parts as replicates resulting in 48 samples for soil seed bank estimation (9 grazing paddocks × 2 soil depths × 2 replications + 3 control samples × 2 soil depths × 2 replications). All soil samples were incubated outdoors for 2 months to ensure that the seeds were subjected to freezing temperatures, after which they were transferred to a glasshouse (Brock and Rogers 1998).
Germination tests
The number of seedlings of different species that emerged from the soil samples served as a measure of both the number of viable seeds and the composition of the soil seed bank (Espeland et al. 2010); this approach determines the relative abundance of viable seeds and excludes nonviable seeds (Poiani and Johnson 1988). The soil samples were dried at room temperature, after which any large stones and grass roots were removed. The soil aggregates were broken apart to release any seeds. Each soil sample (2 cm thick) was put on top of a 2-cm layer of (seed-free) sand within a germination tray (20 cm diameter). Germination trays were placed under natural indoor light conditions at 20 ± 5°C, and the soil in the trays was kept moist. Seedlings started to emerge after 1 week. During the entire growing period, the emerging seedlings were identified as soon as possible, recorded, and removed. Those seedlings that were difficult to identify were counted but were otherwise maintained regularly until they could be identified. The seeds in the samples were allowed to germinate for 3 months (Thompson and Grime 1979). No attempt was made to assess the number of ungerminated seeds potentially remaining in the soil samples.
Data analyses
The density of seeds (number of emerged seedlings per plot, seedlings m−2), species richness (the number of species per paddock, species 0.06 m−2), and species composition were identified according to Cheng and Zhu (2012) and recorded. Species evenness (Evar), which is calculated as:
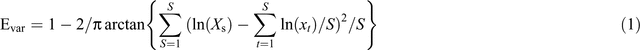
where Xs is the abundance of the Sth species (Smith and Wilson 1996).
The Jaccard coefficient of similarity (Kalacska et al. 2004) was used to test for similarities between the composition of seedlings and aboveground plant species in response to different stocking rates. The Jaccard similarity (J) is calculated as:

where a is the number of common species present in the two treatments (the seedlings and aboveground vegetation under the same stocking rate), and b is the total number of species in the two treatments. The species compositions of the seedlings and aboveground vegetation were compared between years by a nonmetric multidimensional scaling (NMDS) analysis using the Bray–Curtis dissimilarity matrix; the analysis was based on the relative density of the species in the 12 sample units (9 grazing paddocks and 3 control sites). The NMDS analysis was performed using R statistical package version R.342.
A goodness-of-fit (Shapiro-Wilk) test indicated that the data collected during this study were normally distributed. To test for statistical differences in the density, richness, and evenness of the seedlings, a general linear model was used; rainfall, stocking rate, and soil depth as well as their interactions were considered independent factors. Differences in the density, richness, and evenness of the seedlings between different stocking rates, soil depths, and years were verified by Tukey a posteriori tests. In addition, to test for differences in Jaccard similarity between the seedlings and aboveground species, a general linear model that included stocking rate and year and their interaction was utilised. These analyses were carried out using the software SAS 9.3 (SAS Institute Inc., Cary, NC, USA).
We used structural equation modelling to estimate the contributions of stocking rates and annual rainfall through the aboveground vegetation to seedling assessments (density, richness, and evenness). The fertile stem numbers of A. capillaries were used in the structural equation modelling to explain the density of the seedlings because it was the dominant plant species in aboveground vegetation and number of seedlings (21.4–74.6% of seedlings and 68–94.8% fertile stems in both years). The primary advantage of structural equation modelling is its ability to evaluate complex causality between variables; structural equation modelling translates hypothesised causal relationships into patterns of expected statistical relationships among the data (Grace 2006). In our model, we assumed that the stocking rate and rainfall had the potential to alter the density, richness and evenness of the seedlings directly and indirectly through their influence on the fertile stems of A. capillaries, richness, and evenness of the aboveground species, the stocking rate and rainfall had the potential to alter those same parameters indirectly. We used the chi-square test to evaluate the fit of the model indicated by 0 ≤ χ2 / d.f. ≤ 2 and 0.05 < P ≤ 1. Here, a large P-value (> 0.05) indicates that the covariance structure of the data and the expected model did not differ significantly (Grace 2006). All variables were tested for normality, and structural equation modelling analyses were performed using AMOS 21 (Arbuckle 2010).
Results and analyses
Composition of seedlings
A total of 16 plant species germinated from the seeds in the soil; these species belonged to nine families (Asteraceae, Gramineae, Leguminosae, Rosaceae, Linaceae, Plantaginaceae, Chenopodiaceae, Brassicaceae, Boraginaceae) comprising 12 perennial and 4 annual herbaceous species (Table 2). No annual species were observed in the aboveground vegetation (Table 1). Fifteen and sixteen species germinated in 2015 and 2016 respectively. Cleistogenes squarrosa (Trin.) Keng was only found in 2016, and Cleistogenes songorica (Roshev.) Ohwi, Agriophyllum squarrosum (Linn.) Moq. and Torularia humilis (C. A. Meyer) O. E. Schulz were only found in the grazed plots in both years; the dominant species A. capillaries was found in all of the sample plots in both years. A. squarrosum, an annual plant, was only found as seedlings in the 2.7 and 5.3 sheep ha−1 stocking rates. The seedlings were mainly concentrated within the 0−5-cm soil layer, and were rarely found in the 5−10-cm samples (Table 2).

|
Seedling density
Rainfall (indicated by year data) and soil sampling depth significantly affected seedling density (P < 0.0001), and stocking rate had no main effect (P > 0.05). All interactions between rainfall, stocking rate, and soil depth significantly affected seedling density (P < 0.05) (Table 3). Seedling density initially increased and then decreased with increased stocking rate in 2015. Maximum seedling numbers occurred at 5.3 sheep ha−1 (1206 seedlings m–2) in 2015; in 2016, seedling densities were 2831, 2255, 2254 and 1914 seedlings m−2 at 0, 2.7, 5.3, and 8.7 sheep ha−1 (Table 2). The density of seedlings in the 0−5-cm soil layer was significantly higher than the 5−10-cm layer in both years and for all stocking rates (Tables 2 and 3).
|
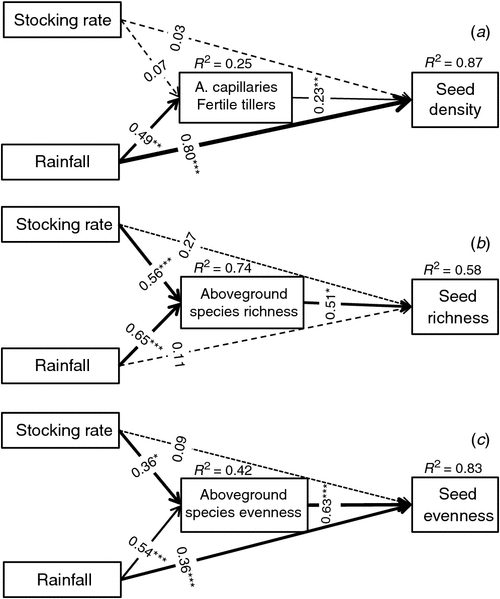
The structural equation modelling analysis explained 87% of the variation in the density of seedlings (Fig. 5a). Stocking rate influenced seed density and fertile tillers of A. capillaries (standardised path coefficients of −0.03 and 0.07; both P > 0.05; Fig. 5a). The direct effects of rainfall on seedling density and fertile tillers of A. capillaries were positive (0.80 and 0.49; P < 0.001 and P < 0.01 respectively; Fig. 5a); the effect of fertile tillers of A. capillaries on the seed density was significantly positive (0.23; P < 0.01; Fig. 5a); the indirect effects of stocking rate and rainfall (via fertile tillers of A. capillaries) were also positive (0.02 and 0.11).
Species richness
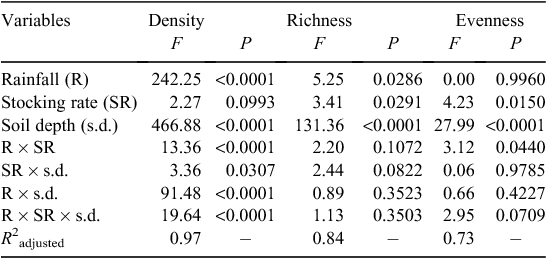
Rainfall, stocking rate, and soil depth all significantly affected richness of seedlings (P < 0.05), but their interactions did not (Table 3). The number of species observed in 2015 (2.2 species 0.06 m−2) was, significantly lower than in 2016 (2.9 species 0.06 m−2). The 0−5-cm soil layer (4.3 species 0.06 m−2) produced significantly more species than the 5−10-cm soil layer (0.8 species 0.06 m−2). Species richness was significantly higher in the 5.3 and 8.7 sheep ha−1 stocking rates (3.1 and 3.0 species 0.06 m−2, respectively) than in the ungrazed plots (1.9 species 0.06 m−2) (P < 0.05). Species richness was intermediate at 2.7 sheep ha−1 (2.3 species 0.06 m−2) (Fig. 2).

|
The structural equation modelling analysis (Fig. 5b) explained 58% of the variation in seedling species richness. The direct effects of stocking rate and rainfall on seedling species richness were not significant (standardised path coefficients 0.27 and 0.11, respectively; P > 0.05), but they did influence aboveground species richness (standardised path coefficients 0.56 and 0.65; P < 0.001). The aboveground species richness had significantly positive effect on the seedling richness (0.51; P < 0.05). Both stocking rate and rainfall indirectly affected seedling species richness through the aboveground species richness (0.29 and 0.33).
Species evenness
Stocking rate and soil depth both significantly affected the species evenness of seedlings (P < 0.05 and P < 0.0001, respectively), rainfall alone had no significant effect, but there was a significant interaction between rainfall and stocking rate on species evenness (P < 0.05) (Table 3). Species evenness of seedlings was similar (0.59 and 0.66) in 2015 and 2016 respectively, and significantly higher in the 5–10-cm soil layer (0.82) than in the 0–5-cm soil layer (0.50). With respect to the stocking rate, species evenness of seedlings in plots under a stocking rate of 8.7 sheep ha−1 (0.56) was significantly lower than in the ungrazed plots (0.78); species evenness was 0.61 and 0.59 in 2.7 and 5.3 sheep ha−1 stocking rates, respectively (Fig. 2).
The structural equation modelling analysis (Fig. 5c) explained 83% of the variation in species evenness. Stocking rate did not directly influence seedling evenness (standardised path coefficient − 0.09; P > 0.05), but did significantly affect the species evenness of aboveground vegetation (standardised path coefficient − 0.36; P < 0.05). Rainfall significantly influenced species evenness of both seedlings and aboveground vegetation (standardised path coefficients − 0.36 and − 0.54; P < 0.001). The aboveground species evenness had significantly positive effect on the seedling evenness (0.63; P < 0.001). Both stocking rate and rainfall indirectly affected species evenness of seedlings through aboveground species evenness (−0.23 and −0.34).
Similarity analyses of seedlings and aboveground vegetation
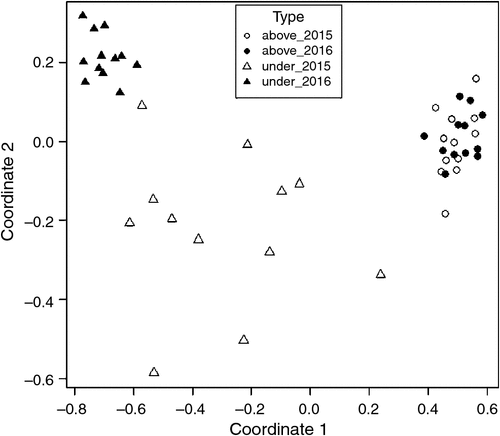
The nonmetric multidimensional scaling analysis showed that species composition significantly differed between seedlings and aboveground vegetation. Aboveground vegetation was clustered over time; seedlings were more heterogeneous, clustered in 2016 but separated at the two coordinates in 2015 (Fig. 3), indicating that drought strongly influenced composition of seedlings but minimally influenced composition of the aboveground vegetation.
|
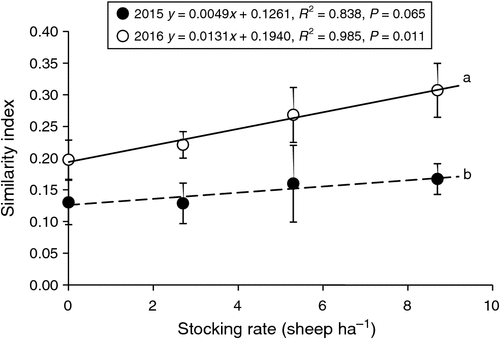
The Jaccard coefficients of similarity were relatively low (< 0.33) (Mendes et al. 2015). The similarity of aboveground vegetation and seedlings increased as stocking rate increased in both 2015 and 2016, but this effect was only significant in 2016. The similarities were significantly higher in 2016 than in 2015 for each stocking rate (P < 0.05; Fig. 4).
|
|
Discussion
Density of seedlings
Previous studies have shown that grazing can affect the density and composition of seedlings emerging from the soil seed bank (Tessema et al. 2012; van Langevelde et al. 2016). This study shows that stocking rate significantly affected seedling density and was rainfall dependent (P < 0.001 for stocking rate × year) (Table 3). In the drought year of 2015, seedling density first increased but then decreased with increasing stocking rate. Densities within the 0−10-cm depth were 271, 524, 1442, and 1022 seedlings m−2 for the stocking rates 0, 2.7, 5.3, and 8.7 sheep ha−1, respectively (Table 2). The density of seedlings was highest under a stocking rate of 5.3 sheep ha–1; moderate grazing can result in higher seedling density emerging from the soil seed bank (Zhao et al. 2001; Dreber and Esler 2011). This may be due to sheep treading in previous years which may facilitate the storage of seeds in the soil (Willms and Quinton 1995). In contrast, in the normal rainfall year 2016, density decreased with increasing stocking rate (Table 2). Grazing enclosures have been shown to increase the density and species richness of seedlings emerging from the soil seed bank on the Loess Plateau because livestock, especially high stocking rates, consume plant shoots, reducing seed production (Zhao et al. 2008).
Similarly, rainfall can affect the influence of stocking rate on soil seed bank density (Yan et al. 2012; Pol et al. 2014). Very little rainfall occurred in 2015, which would have reduced seed production irrespective of stocking rate.
E. pilosa and C. glaucum (both annual cropland weeds plants (Hu et al. 2015)) were only found as seedlings (Tables 1 and 2). A. squarrosum and L. deserticola were found only in aboveground vegetation in the normal rainfall year (2016), but both species were recorded as seedlings for both years. A. squarrosum, an annual plant, was only found as seedlings in the 2.7 and 5.3 sheep ha−1 stocking rates. Gates et al. (2017) reported that annuals generally emerge first during the recovery of degraded grasslands, indicating that annuals in the soil seed bank are pioneer species. The perennial and annual species in the soil seed bank indicate that the grassland of the study area is not in a highly degenerative state.
The effect of stocking rate on density of the most dominant species, A. capillaries, was similar to stocking rate effects for community density in both years (Table 2). However, seedling densities of the next most dominant species, (S. bungeana and L. bicolor) were lower at the highest stocking rate (8.7 sheep ha−1). P. lanceolata is a good forage species but was only found as seedlings. Future efforts to restore grassland in this area could endeavour to enhance the germination of P. lanceolata from the soil seed bank. More legume seedlings were recorded at 5.3 sheep ha−1. Legumes may not be the most preferred forage of sheep, but they improve soil fertility through biological nitrogen fixation, increasing the productivity of the grassland.
The structural equation modelling analysis examined the contributions of the direct and indirect effects of stocking rate and annual rainfall on the density of seedlings (Fig. 5a). Rainfall had positive effects on A. capillaries growth, producing higher fertile tiller numbers, resulting in higher seed production and seed fall which resulted in greater seedling numbers. Rainfall was the main factor influencing the density of seedlings in the study area.
Seedling species richness
Both rainfall and stocking rate significantly influenced the species richness of seedlings emerging from the soil seed bank (Table 3), but the structural equation modelling analysis revealed that stocking rate and rainfall had no direct effect on the it, the model suggesting that stocking rate and rainfall both significantly influenced the species richness of the grassland, and the species richness of the grassland had significant effect on the seedlings from the seed bank (Fig. 5b). Grazing can influence the species richness of seedlings from the soil seed bank (Harrison et al. 2003) through its influence on grassland vegetation species richness (Edwards et al. 2007; Báldi et al. 2013). Limitations on seed dispersal can also influence the species richness of the seedlings from soil seed bank (López-Mariño et al. 2000). Differences in grazing intensity have less influence on species richness in soil seed banks (Tessema et al. 2016).
The coefficients in the structural equation modelling are based on linear responses, whereas curvilinear regression, for example, richness versus stocking rates are generally present. In this study they were all linear responses.
Species evenness
The species evenness of seedlings was significantly influenced by the interaction of stocking rate and the rainfall (P = 0.0440 for stocking rate × year) (Table 3), which means that any effects of stocking rate on seedling evenness is dependent on rainfall (stocking rate × year, P = 0.0440). The structural equation modelling analysis can be used to describe the interaction effect. Both stocking rate and rainfall have direct negative effect on the evenness of aboveground species, and the aboveground species evenness have a significant direct effect on seedling evenness, leading to both rainfall and stocking rate indirectly affecting the seedling evenness through the evenness of the aboveground species. Hence, any effects on seedling evenness by stocking rate were dependent on rainfall effects on aboveground species evenness (Fig. 5c). Rainfall controls species emergence (Pugnaire and Lázaro 2000). Grazing and rainfall can influence the species evenness of seedlings from the soil seed bank by affecting grassland vegetation (Sudebilige et al. 2000; Baggio et al. 2018). In addition, dominance of a single species, for example, A. capillaries in our study, can reduce the coverage of other aboveground species (Flombaum and Sala 2008), which may negatively influence the species evenness of seedlings (Pugnaire and Lázaro 2000). The shrub canopy strongly affects which species appear in the understory, and seed dispersal did not limit species abundance (Pugnaire and Lázaro 2000).
Similarity between seedlings and aboveground vegetation
In this study, the similarity between species in the aboveground vegetation and species of the seedlings was low (Fig. 4). The similarity was significantly influenced by rainfall, being higher in the year of normal rainfall (2016) than in the dry year of 2015. Aboveground plant growth in 2016 was higher than in 2015. Consequently plants almost certainly produced more seeds, increasing the soil seed bank. Similarity increased with an increasing stocking rate in the normal rainfall year. Previous studies by Fenner (1985) and Leck and Simpson (1994) showed that the similarity between the soil seed bank and aboveground vegetation is relatively high in grazed grasslands. Many grassland studies have investigated the spatial patterns of seeds and found them to be clustered around parent plants (Shaukat and Siddiqui 2004), meaning that similarity tends to increase over time (Hopfensperger 2007). This is especially true with short dispersal distances (Bossuyt and Hermy 2004). However, Sanderson et al. (2014) reported low similarity between the soil seed bank and aboveground vegetation. Milberg and Hansson (1994) reported that similarity is strongly correlated with seed germination characteristics, and species with high turnover in a community are more prevalent in the soil than species with low turnover. The number of factors influencing the similarity between the seedlings and aboveground species is high and their interactions are not well understood; this warrants further studies.
Conclusion
Both rainfall and stocking rate were important factors regulating the composition, richness, and evenness of seedlings, and the similarity between aboveground vegetation and seedlings emerging from the soil seed bank in a semi-arid rangeland in North-west China. The density, richness, and the similarity of the seedlings significantly decreased during the dry year (2015), and the richness and similarity of the seedlings increased as the stocking rate increased. This study showed long-term grazing (15 years) enhanced the stability of the soil seed bank as the density and species richness of seedlings from the soil seed bank were higher in grazed plots than in ungrazed plots under the drought. We propose that adopting more judicious long-term grazing strategies has the potential of offsetting the negative effects of environmental changes anticipated in the future on aboveground vegetation and the soil seed banks of semi-arid grasslands in China.
Conflicts of interest
The authors declare no conflicts of interest.
Acknowledgements
We gratefully thank Xiaowen Hu for the experimental design and acknowledge the support of the Strategic Priority Research Program of Chinese Academy of Sciences (XDA2010010203), the Program for Changjiang Scholars and Innovative Research Team in University (IRT-17R50), and the National Natural Science Foundation of China (31672472). The comments of the anonymous reviewers led to substantial improvements in the paper.
References
Arbuckle, J. (2010). ‘IBM SPSS Amos 19 User’s Guide.’ (Amos Development Corporation: Crawfordville, FL.)Baggio, R., Medeiros, R. B., Focht, T., Boavista, L. R., Pillar, V. D., and Müller, S. C. (2018). Effects of initial disturbances and grazing regime on native grassland invasion by Eragrostis plana in southern Brazil. Perspectives in Ecology and Conservation 16, 158–165.
| Effects of initial disturbances and grazing regime on native grassland invasion by Eragrostis plana in southern Brazil.Crossref | GoogleScholarGoogle Scholar |
Báldi, A., Batáry, P., and Kleijn, D. (2013). Effects of grazing and biogeographic regions on grassland biodiversity in Hungary–analysing assemblages of 1200 species. Agriculture, Ecosystems & Environment 166, 28–34.
| Effects of grazing and biogeographic regions on grassland biodiversity in Hungary–analysing assemblages of 1200 species.Crossref | GoogleScholarGoogle Scholar |
Bossuyt, B., and Hermy, M. (2004). Seed bank assembly follows vegetation succession in dune slacks. Journal of Vegetation Science 15, 449–456.
| Seed bank assembly follows vegetation succession in dune slacks.Crossref | GoogleScholarGoogle Scholar |
Brock, M. A., and Rogers, K. H. (1998). The regeneration potential of the seed bank of an ephemeral floodplain in South Africa. Aquatic Botany 61, 123–135.
| The regeneration potential of the seed bank of an ephemeral floodplain in South Africa.Crossref | GoogleScholarGoogle Scholar |
Chen, X. J., Hou, F. J., Matthew, C., and He, X. Z. (2010). Stocking rate effects on metabolizable energy intake and grazing behaviour of Tan sheep in steppe grassland on the Loess Plateau of Northwest China. The Journal of Agricultural Science 148, 709–721.
| Stocking rate effects on metabolizable energy intake and grazing behaviour of Tan sheep in steppe grassland on the Loess Plateau of Northwest China.Crossref | GoogleScholarGoogle Scholar |
Chen, J. B., Hou, F. J., Chen, X. J., Wan, X. L., and Millner, J. (2015). Stocking rate and grazing season modify soil respiration on the Loess Plateau, China. Rangeland Ecology and Management 68, 48–53.
| Stocking rate and grazing season modify soil respiration on the Loess Plateau, China.Crossref | GoogleScholarGoogle Scholar |
Chen, X. J., Hou, F. J., Matthew, C., and He, X. Z. (2017). Soil C, N, and P stocks evaluation under major land uses on China’s Loess Plateau. Rangeland Ecology and Management 70, 341–347.
| Soil C, N, and P stocks evaluation under major land uses on China’s Loess Plateau.Crossref | GoogleScholarGoogle Scholar |
Cheng, J. M., and Zhu, R. B. (2012). ‘Illustrated Handbook of Common Plants in the Loess Plateau of China.’ (Science Press: Beijing.)
Dreber, N., and Esler, K. J. (2011). Spatio-temporal variation in soil seed banks under contrasting grazing regimes following low and high seasonal rainfall in arid Namibia. Journal of Arid Environments 75, 174–184.
| Spatio-temporal variation in soil seed banks under contrasting grazing regimes following low and high seasonal rainfall in arid Namibia.Crossref | GoogleScholarGoogle Scholar |
Edwards, A. R., Mortimer, S. R., Lawson, C. S., Westbury, D. B., Harris, S. J., Woodcock, B. A., and Brown, V. K. (2007). Hay strewing, brush harvesting of seed and soil disturbance as tools for the enhancement of botanical diversity in grasslands. Biological Conservation 134, 372–382.
| Hay strewing, brush harvesting of seed and soil disturbance as tools for the enhancement of botanical diversity in grasslands.Crossref | GoogleScholarGoogle Scholar |
Espeland, E. K., Perkins, L. B., and Leger, E. A. (2010). Comparison of seed bank estimation techniques using six weed species in two soil types. Rangeland Ecology and Management 63, 243–247.
| Comparison of seed bank estimation techniques using six weed species in two soil types.Crossref | GoogleScholarGoogle Scholar |
Fenner, M. (1985). ‘Seed Ecology.’ (Springer: Dordrecht, The Netherlands.)
Flombaum, P., and Sala, O. E. (2008). Higher effect of plant species diversity on productivity in natural than artificial ecosystems. Proceedings of the National Academy of Sciences of the United States of America 105, 6087–6090.
| Higher effect of plant species diversity on productivity in natural than artificial ecosystems.Crossref | GoogleScholarGoogle Scholar | 18427124PubMed |
Gao, Y., Cheng, J. M., and Liu, W. (2011). Community characteristics of different types of grassland in the Loess Plateau. Pratacultural Science 28, 1066–1069.
Gates, E. A., Vermeire, L. T., Marlow, C. B., and Waterman, R. C. (2017). Fire and season of postfire defoliation effects on biomass, composition, and cover in mixed-grass prairie. Rangeland Ecology and Management 70, 430–436.
| Fire and season of postfire defoliation effects on biomass, composition, and cover in mixed-grass prairie.Crossref | GoogleScholarGoogle Scholar |
Gioria, M., Jarošík, V., and Pyšek, P. (2014). Impact of invasions by alien plants on soil seed bank communities: emerging patterns. Perspectives in Plant Ecology, Evolution and Systematics 16, 132–142.
| Impact of invasions by alien plants on soil seed bank communities: emerging patterns.Crossref | GoogleScholarGoogle Scholar |
Gong, Z., Lei, W., Chen, Z., Gao, Y., Zeng, S., Zhang, G., Xiao, D., and Li, S. (2007). Chinese Soil Taxonomy. Bulletin of the Chinese Academy of Sciences 21, 36–38.
Grace, J. B. (2006). ‘Structural Equation Modeling and Natural Systems.’ (Cambridge University Press: Cambridge, UK.)
Gutterman, Y. (2012). ‘Seed Germination in Desert Plants.’ (Springer Science & Business Media: Berlin.)
Harrison, S., Inouye, B. D., and Safford, H. D. (2003). Ecological heterogeneity in the effects of grazing and fire on grassland diversity. Conservation Biology 17, 837–845.
| Ecological heterogeneity in the effects of grazing and fire on grassland diversity.Crossref | GoogleScholarGoogle Scholar |
Hopfensperger, K. N. (2007). A review of similarity between seed bank and standing vegetation across ecosystems. Oikos 116, 1438–1448.
| A review of similarity between seed bank and standing vegetation across ecosystems.Crossref | GoogleScholarGoogle Scholar |
Hu, A., Chen, H., Chen, X. J., and Hou, F. J. (2015). Soil seed banks of cropland and rangeland on the Loess Plateau. Pratacultural Science 32, 1035–1040.
Kalacska, M., Sanchez-Azofeifa, G. A., Calvo-Alvarado, J. C., Quesada, M., Rivard, B., and Janzen, D. H. (2004). Species composition, similarity and diversity in three successional stages of a seasonally dry tropical forest. Forest Ecology and Management 200, 227–247.
| Species composition, similarity and diversity in three successional stages of a seasonally dry tropical forest.Crossref | GoogleScholarGoogle Scholar |
Kalamees, R., Püssa, K., Zobel, K., and Zobel, M. (2012). Restoration potential of the persistent soil seed bank in successional calcareous (alvar) grasslands in Estonia. Applied Vegetation Science 15, 208–218.
| Restoration potential of the persistent soil seed bank in successional calcareous (alvar) grasslands in Estonia.Crossref | GoogleScholarGoogle Scholar |
Kassahun, A., Snyman, H. A., and Smit, G. N. (2009). Soil seed bank evaluation along a degradation gradient in arid rangelands of the Somali region, eastern Ethiopia. Agriculture, Ecosystems & Environment 129, 428–436.
| Soil seed bank evaluation along a degradation gradient in arid rangelands of the Somali region, eastern Ethiopia.Crossref | GoogleScholarGoogle Scholar |
Kinloch, J. E., and Friedel, M. H. (2005). Soil seed reserves in arid grazing lands of central Australia. Part 1: seed bank and vegetation dynamics. Journal of Arid Environments 60, 133–161.
| Soil seed reserves in arid grazing lands of central Australia. Part 1: seed bank and vegetation dynamics.Crossref | GoogleScholarGoogle Scholar |
Leck, M. A., and Simpson, R. L. (1994). Tidal freshwater wetland zonation: seed and seedling dynamics. Aquatic Botany 47, 61–75.
| Tidal freshwater wetland zonation: seed and seedling dynamics.Crossref | GoogleScholarGoogle Scholar |
Liu, H., Jiang, Q., Wang, Z. J., and Pan, Z. B. (2011). A study on soil seed bank in desert steppe of different enclosing years in Ningxia. Research of Soil and Water Conservation 18, 96–98.
López-Mariño, A., Luis-Calabuig, E., Fillat, F., and Bermúdez, F. F. (2000). Floristic composition of established vegetation and the soil seed bank in pasture communities under different traditional management regimes. Agriculture, Ecosystems & Environment 78, 273–282.
| Floristic composition of established vegetation and the soil seed bank in pasture communities under different traditional management regimes.Crossref | GoogleScholarGoogle Scholar |
Mendes, L. B., Silva, K. D. A., Santos, D. D. M., Santos, J. M. D. F. F., Albuquerque, U. P. D., and Araujo, E. D. L. (2015). What happens to the soil seed bank 17 years after clear cutting of vegetations? Revista de Biología Tropical 63, 321–332.
Milberg, P., and Hansson, M. L. (1994). Soil seed bank and species turnover in a limestone grassland. Journal of Vegetation Science 5, 35–42.
| Soil seed bank and species turnover in a limestone grassland.Crossref | GoogleScholarGoogle Scholar |
Navie, S. C., and Rogers, R. W. (1997). The relationship between attributes of plants represented in the germinable seed bank and stocking pressure in a semi-arid subtropical rangeland. Australian Journal of Botany 45, 1055–1071.
| The relationship between attributes of plants represented in the germinable seed bank and stocking pressure in a semi-arid subtropical rangeland.Crossref | GoogleScholarGoogle Scholar |
Poiani, K. A., and Johnson, W. C. (1988). Evaluation of the emergence method in estimating seed bank composition of prairie wetlands. Aquatic Botany 32, 91–97.
| Evaluation of the emergence method in estimating seed bank composition of prairie wetlands.Crossref | GoogleScholarGoogle Scholar |
Pol, R. G., Sagario, M. C., and Marone, L. (2014). Grazing impact on desert plants and soil seed banks: Implications for seed-eating animals. Acta Oecologica 55, 58–65.
| Grazing impact on desert plants and soil seed banks: Implications for seed-eating animals.Crossref | GoogleScholarGoogle Scholar |
Pugnaire, F. I., and Lázaro, R. (2000). Seed bank and understorey species composition in a semi-arid environment: the effect of shrub age and rainfall. Annals of Botany 86, 807–813.
| Seed bank and understorey species composition in a semi-arid environment: the effect of shrub age and rainfall.Crossref | GoogleScholarGoogle Scholar |
Ren, J. Z., Hu, Z. Z., Zhao, J., Zhang, D. G., Hou, F. J., Lin, H. L., and Mu, X. D. (2008). A grassland classification system and its application in China. The Rangeland Journal 30, 199–209.
| A grassland classification system and its application in China.Crossref | GoogleScholarGoogle Scholar |
Sanderson, M. A., Stout, R., Goslee, S., Gonet, J., and Smith, R. G. (2014). Soil seed bank community structure of pastures and hayfields on an organic farm. Canadian Journal of Plant Science 94, 621–631.
| Soil seed bank community structure of pastures and hayfields on an organic farm.Crossref | GoogleScholarGoogle Scholar |
Shaukat, S. S., and Siddiqui, I. A. (2004). Spatial pattern analysis of seeds of an arable soil seed bank and its relationship with above-ground vegetation in an arid region. Journal of Arid Environments 57, 311–327.
| Spatial pattern analysis of seeds of an arable soil seed bank and its relationship with above-ground vegetation in an arid region.Crossref | GoogleScholarGoogle Scholar |
Smith, B., and Wilson, J. B. (1996). A consumer’s guide to evenness indices. Oikos 76, 70–82.
| A consumer’s guide to evenness indices.Crossref | GoogleScholarGoogle Scholar |
Solomon, T. B., Snyman, H. A., and Smit, G. N. (2006). Soil seed bank characteristics in relation to land use systems and distance from water in a semi-arid rangeland of southern Ethiopia. South African Journal of Botany 72, 263–271.
| Soil seed bank characteristics in relation to land use systems and distance from water in a semi-arid rangeland of southern Ethiopia.Crossref | GoogleScholarGoogle Scholar |
Sudebilige, H., Li, Y., Yong, S., and Ren, S. (2000). Germinable soil seed bank of Artemisia frigida grassland and its response to grazing. Acta Ecologica Sinica 20, 43–48.
Sun, W. Y., Mu, X. M., Song, X. Y., Wu, D., Cheng, A. F., and Qiu, B. (2016). Changes in extreme temperature and precipitation events in the Loess Plateau (China) during 1960–2013 under global warming. Atmospheric Research 168, 33–48.
| Changes in extreme temperature and precipitation events in the Loess Plateau (China) during 1960–2013 under global warming.Crossref | GoogleScholarGoogle Scholar |
Tessema, Z. K., Boer, W. F. B., Baars, R. M. T., and Prins, H. H. T. (2012). Influence of grazing on soil seed banks determines the restoration potential of aboveground vegetation in a semi-arid savanna of Ethiopia. Biotropica 44, 211–219.
| Influence of grazing on soil seed banks determines the restoration potential of aboveground vegetation in a semi-arid savanna of Ethiopia.Crossref | GoogleScholarGoogle Scholar |
Tessema, Z. K., Boer, W. F. B., and Prins, H. H. T. (2016). Changes in grass plant populations and temporal soil seed bank dynamics in a semi-arid African savanna: Implications for restoration. Journal of Environmental Management 182, 166–175.
| Changes in grass plant populations and temporal soil seed bank dynamics in a semi-arid African savanna: Implications for restoration.Crossref | GoogleScholarGoogle Scholar | 27472053PubMed |
Thompson, K. (2000). The functional ecology of soil seed banks. In: ‘Seeds: the Ecology of Regeneration in Plant Communities’. 2nd edn. (Ed. M. Fenner.) pp. 215–235. (CABI Publishing: Wallingford, UK.)
Thompson, K., and Grime, J. P. (1979). Seasonal variation in the seed banks of herbaceous species in ten contrasting habitats. Journal of Ecology 67, 893–921.
| Seasonal variation in the seed banks of herbaceous species in ten contrasting habitats.Crossref | GoogleScholarGoogle Scholar |
van Langevelde, F., Tessema, Z. K., de Boer, W. F., and Prins, H. H. T. (2016). Soil seed bank dynamics under the influence of grazing as alternative explanation for herbaceous vegetation transitions in semi-arid rangelands. Ecological Modelling 337, 253–261.
| Soil seed bank dynamics under the influence of grazing as alternative explanation for herbaceous vegetation transitions in semi-arid rangelands.Crossref | GoogleScholarGoogle Scholar |
Willms, W. D., and Quinton, D. A. (1995). Grazing effects on germinable seeds on the fescue prairie. Journal of Range Management 48, 423–430.
| Grazing effects on germinable seeds on the fescue prairie.Crossref | GoogleScholarGoogle Scholar |
Yan, R. R., Wei, Z. J., Xin, X. P., Liu, H. M., Yang, J., and Wuren, Q. Q. G. (2012). Effects of the grazing systems on germinable soil seed bank of desert steppe. Sciences in Cold and Arid Regions 4, 40–45.
| Effects of the grazing systems on germinable soil seed bank of desert steppe.Crossref | GoogleScholarGoogle Scholar |
Zhao, W. Z., Liu, Z. M., and Chang, X. L. (2001). Influence of grazing intensity on seed bank of a sandy grassland in Horqin Steppe of China. Annals of Arid Zone 40, 397–404.
Zhao, L. P., Cheng, J. M., Wan, H. E., Shi, H. L., and Tan, S. T. (2008). Characteristics of soil seed banks in grassland under fencing and grazing in Loess Plateau. Pratacultural Science 25, 78–83.